intracellular sodium homeostasis in rat hippocampal astrocytes
TRANSCRIPT

Journal of Physiology (1996), 491.2, pp.291-305
Intracellular sodium homeostasis in rat hippocampalastrocytes
Christine R. Rose and Bruce R. Ransom
Department of Neurology, Yale University School of Medicine, New Haven,CT 06520, USA
1. We determined the intracellular Na+ concentration ([Na+]1) and mechanisms of itsregulation in cultured rat hippocampal astrocytes using fluorescence ratio imaging of theNae indicator SBFI-AM (acetoxymethylester of sodium-binding benzofuran isophthalate,10 /SM). Dye signal calibration within the astrocytes showed that the ratiometric dye signalchanged monotonically with changes in [Nae]i from 0 to 140 mm. The K+ sensitivity of thedye was negligible; intracellular pH changes, however, slightly affected the 'Nae' signal.
2. Baseline [Nae]i was 14-6 + 4-9 mm (mean + S.D.) in C02/HC03--containing saline with3 mM K+. Removal of extracellular Na+ decreased [Na+]i in two phases: a rapid phase of[Nae], reduction (0-58 + 0-32 mm min-) followed by a slower phase (0-15 + 0 09 mm min-).
3. Changing from C02/HC03--free to C02/HCO3--buffered saline resulted in a transientincrease in [Na+]i of -5 mm, suggesting activation of inward Na+-HCO3- cotransport byCO2/HCO3. During furosemide (frusemide, 1 mM) or bumetanide (50 /SM) application, aslow decrease in [Na+]i of -2 mm was observed, indicating a steady inward transport ofNae via Na+-K+-2C- cotransport under control conditions. Tetrodotoxin (100 /SM) did notinfluence [Nae]i in the majority of cells (85%), suggesting that influx of Nae throughvoltage-gated Nae channels contributed to baseline [Nae], in only a small subpopulation ofhippocampal astrocytes.
4. Blocking Na+,K+-ATPase activity with cardiac glycosides (ouabain or strophanthidin,1 mM) or removal of extracellular K+ led to an increase in [Nae] of about 2 and4 mM min-', respectively. This indicated that Na+,K+-ATPase activity was critical inmaintaining low [Na+]i in the face of a steep electrochemical gradient, which would favoura much higher [Nae]i.
5. Elevation of extracellular K+ concentration ([K+]O) by as little as 1 mm (from 3 to 4 mM)resulted in a rapid and reversible decrease in [Nae]1. Both the slope and the amplitude of the[K+]O-induced reductions in [Na+]i were sensitive to bumetanide. A reduction of [K+]o by1 mm increased [Nae], by 3 0 + 2-3 mm. In contrast, changing extracellular Naeconcentration by 20 mm resulted in changes in [Na+]i of less than 3 mM.
6. These results implied that in hippocampal astrocytes low baseline [Nae]i is determined bythe action of Na+-HCO3- cotransport, Na+-K+-2CF cotransport and Na+,K+-ATPase,and that both Na+,K+-ATPase and inward Na+-K+-2CF- cotransport are activated bysmall, physiologically relevant increases in [K+]O. These mechanisms are well suited to helpbuffer increases in [K+]O associated with neural activity.
Glial cells, like most other cell types, possess a variety of three Nae out of the cell in exchange for the uptake of twotransport systems that regulate the exchange of ions or K+. Glial Na+,K+-ATPase, in addition to regulation bylarger molecules with the extracellular space. The vast intracellular Nae concentration ([Na+]i), is believed to bemajority of these processes are energized by the inwardly activated by elevations in extracellular K+ concentrationdirected electrochemical gradient for Nae (e.g. Nae-coupled ([K+]O) (for review see Sweadner, 1995), which makes thisuptake of neurotransmitters). The Na+ gradient, in turn, is transporter suitable for a role in extracellular K+ clearancedue to the activity of Na+,K+-ATPase, which transports (Walz, 1989; Ransom & Sontheimer, 1992). Astrocytes are
4695 291
) by guest on July 13, 2011jp.physoc.orgDownloaded from J Physiol (

C. R. Rose and B. R. Ransom
believed to regulate many extracellular ions and molecules,and appear to do so in most instances by Na+ gradient-dependent mechanisms (Walz, 1989).
Regulation of astrocyte [Nae]i, which is central to Nae-dependent transport, has not been well studied. Someevidence suggests that Na+ influx, necessary to provide a
steady supply of Nae for Na+,K+-ATPase, occurs via inwardNa+-K+-2Cl- cotransport, leading to a so-called 'trans-membrane sodium cycle' in astrocytes (Walz & Hinks, 1986).Another mechanism which could maintain high [Nae]i isinflux of Nae through voltage-gated Nae channels, as
described for cultured spinal cord astrocytes (Sontheimer,Fernandez-Marques, Ullrich, Pappas & Waxman, 1994).These models, however, rely mainly on indirect measure-
ments of [Na+]i, such as radioisotope methods, becauseion-sensitive microelectrodes are not ideal for measurementof [Nae]i in small cells like astrocytes.
In the present study we have measured [Nae], in culturedrat hippocampal astrocytes using fluorescence ratio imagingwith the Nae indicator dye SBFI, which has been employedsuccessfully in a variety of other cell types (Harootunian,Kao, Eckert & Tsien, 1989; Negulescu, Harootunian, Tsien& Machen, 1990; Levi, Lee & Brooksby, 1994). Dye signalcalibrations within astrocytes after each experimentallowed us to determine absolute values of [Na+]i and toinvestigate the mechanisms that regulate baseline [Nae]1 inthese cells. Some of the results have been published inabstract form (Rose & Ransom, 1995).
METHODSAstrocyte culturesCell cultures were prepared as previously described (Sontheimer,Ransom, Cornell-Bell, Black & Waxman, 1991). Briefly, rat pups
(Sprague-Dawley rats, postnatal day 0) were anaesthetized byCO2 narcosis and decapitated. Hippocampal segments were
removed and dissociated by 30 min of enzyme exposure
(20 U ml-' papain, Worthington) and trituration. Cells were
plated onto polylysine-coated coverslips at a density of about80 cells mm-2 and grown at 37 °C in a 5% C02-95% humid airatmosphere. They were fed every 4 days with DMEM-F12(Dulbecco's modified Eagle's medium, No. 56499, JRH Biosciences,Lenexa, KS, USA; 1 :1) supplemented with 10% bovine serum
(JRH Biosciences). This protocol yielded cultures containing> 95% astrocytes, as judged by immunostaining for glial fibrillaryacidic protein. The absence of low density lipoprotein stainingindicated the virtual absence of microglia (Giulian & Baker, 1985).The cells grew to confluency in about 10 days and cells from10-17 days in vitro were used for experiments.
SolutionsStandard saline used for the experiments had the followingcomposition (mM): 115-75 NaCl, 3 KCl, 2 MgSO4, 2 CaCl2, 1-25NaH2PO4, 23 NaHCO3, 10 glucose, and was continously bubbledwith 5% C02-95% 02 resulting in a pH of 7.38. C02/HC03--freesaline had the same amount of KCl, MgSO4, CaCl2, NaH2PO4 andglucose, but 126-25 mm NaCl and 25 mm Hepes and was titrated toapH of 7-4 with NaOH or HCl.
Drugs and chemicals were obtained from Sigma except for thefluorescent dye SBFI-AM (acetoxymethylester of sodium-bindingbenzofuran isophthalate) which was purchased from Teflabs(Austin, TX, USA). The drugs, which were (except fortetrodotoxin) prepared as 1 M stock solutions in DMSO (dimethylsulphoxide) and stored frozen, were added to the saline shortlybefore use. Addition of DMSO alone (0 2%) to C02/HC03--buffered saline did not affect the 345 nm/385 nm ratio of SBFI (3coverslips, n = 28, data not shown).
The solutions for intracellular calibration of the [Na+]1 sensitivityof SBFI contained (mM): 150 (K+ + Na+), 30 Cl-, 120 gluconicacid. Unless otherwise specified, all calibration solutions contained10 Hepes, 2 MgSO4, and 2 CaCl2, and were adjusted to pH 7-15with KOH (see below). Gramicidin (3 FM), monensin (10 FM) andouabain (1 mM) were added for equilibration of extra- and intra-cellular Nae.To test the K+ sensitivity of intracellular SBFI, cells wereperfused with calibration solutions containing (mM): 140, 120, 100or 60 KCl and 10 NaCl. In saline solutions with reduced KCl, KClwas replaced by equivalent amounts of CsCl (K+ + Nae +Cs+ = 150 mM). Calibration solutions used for testing the pHsensitivity were titrated to pH 6-2, 6-6, 7 0 and 7-4 with KOH.
Loading and intracellular location of the dyeFor measurement of [Nae],, cells were loaded with the fluorescentdye SBFI-AM (10 /M) for 90 min in C02/HCOO3-free saline atroom temperature (20-22 00). Pluronic acid (0-1 %) was added toimprove the dye uptake. Following dye loading the cells weretransferred to an experimental chamber (volume t 500 ul) andperfused with C02/HC03--buffered solution (standard flow rate,6 ml min-), warmed to 370C. Due to a dead space between theswitching site and the chamber, complete solution exchangerequired about 60 s from the time of switching at standard flowrate. Experiments were begun 15-30 min after placing cells in thechamber to allow equilibration of the cells with the saline.
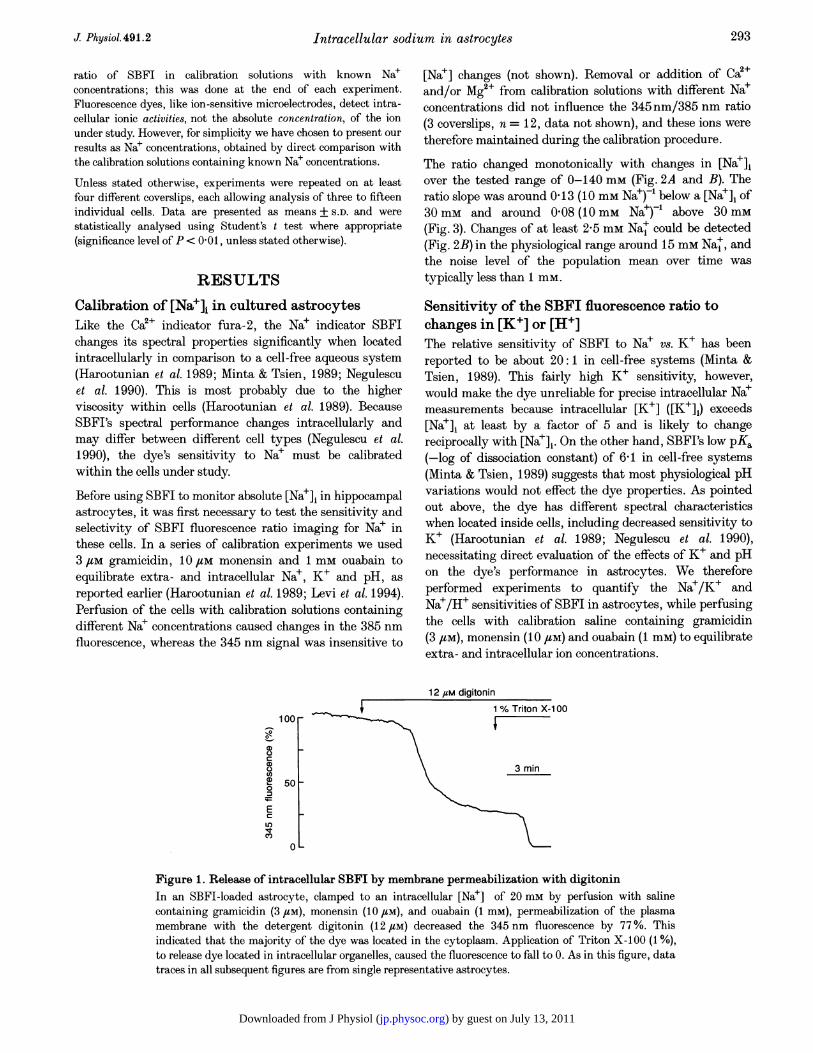
Addition of digitonin to permeabilize the plasma membraneindicated that the majority of the dye was located in thecytoplasm after this loading procedure. In a calibration solution inwhich [Nae]i was clamped to 20 mm, digitonin (12 FM) resulted ina decrease of the 345 nm SBFI fluorescence signal (Na+-insensitivefluorescence) of 77 + 12% (3 coverslips, n= 10; Fig. 1).Subsequent addition of the detergent Triton X-100 (1 %) releasedthe rest of the dye presumably from intracellular organelles. Theratioed dye signal, therefore, primarily reflected cytoplasmic[Nae]1.Intracellular [Na+] measurementThe experimental chamber was mounted on the stage of a Nikon-Diaphot-TMD inverted epifluorescence microscope (Nikon, Japan)equipped with a x 40 epifluorescence oil-immersion objective lens.Cells were excited every 5 s with dual digikrom 120 mono-chromators (CVI Laser Corporation, Albuquerque, NM, USA) at345 and 385 nm. Autofluorescence (< 1 %) and dye bleaching werenegligible during the experiments. Emission fluorescence wascollected above 510 nm by a GenIISys image intensifier systemconnected to a videocamera (MTI CCD72; Dage-MTI, MichiganCity, IN, USA) and quantified by the use of an image acquisitionprogram from Georgia Instruments (Roswell, GA, USA).Data were collected and analysed by a personal computer. Toimprove the signal-to-noise ratio, two temporally adjacent datapoints were averaged. [Na+]i was determined for each individualcell by a two- or three-point calibration of the 345 nm/385 nm
292 J Physiol.491.2
) by guest on July 13, 2011jp.physoc.orgDownloaded from J Physiol (
Intracellular sodium in astrocytes
ratio of SBFI in calibration solutions with known Na+concentrations; this was done at the end of each experiment.Fluorescence dyes, like ion-sensitive microelectrodes, detect intra-cellular ionic activities, not the absolute concentration, of the ionunder study. However, for simplicity we have chosen to present ourresults as Nae concentrations, obtained by direct comparison withthe calibration solutions containing known Nae concentrations.
Unless stated otherwise, experiments were repeated on at leastfour different coverslips, each allowing analysis of three to fifteenindividual cells. Data are presented as means + S.D. and werestatistically analysed using Student's t test where appropriate(significance level of P < 0 01, unless stated otherwise).
RESULTS
Calibration of [Nat]i in cultured astrocytesLike the Ca2+ indicator fura-2, the Na+ indicator SBFIchanges its spectral properties significantly when locatedintracellularly in comparison to a cell-free aqueous system(Harootunian et al. 1989; Minta & Tsien, 1989; Negulescuet al. 1990). This is most probably due to the higherviscosity within cells (Harootunian et al. 1989). BecauseSBFI's spectral performance changes intracellularly andmay differ between different cell types (Negulescu et al.1990), the dye's sensitivity to Na+ must be calibratedwithin the cells under study.
Before using SBFI to monitor absolute [Na+]i in hippocampalastrocytes, it was first necessary to test the sensitivity andselectivity of SBFI fluorescence ratio imaging for Na+ inthese cells. In a series of calibration experiments we used3 4um gramicidin, 10 /LM monensin and 1 mm ouabain toequilibrate extra- and intracellular Na+, K+ and pH, asreported earlier (Harootunian et al. 1989; Levi et al. 1994).Perfusion of the cells with calibration solutions containingdifferent Na+ concentrations caused changes in the 385 nmfluorescence, whereas the 345 nm signal was insensitive to
[Na+] changes (not shown). Removal or addition of Ca2+and/or Mg2+ from calibration solutions with different Na+concentrations did not influence the 345nm/385 nm ratio(3 coverslips, n = 12, data not shown), and these ions weretherefore maintained during the calibration procedure.
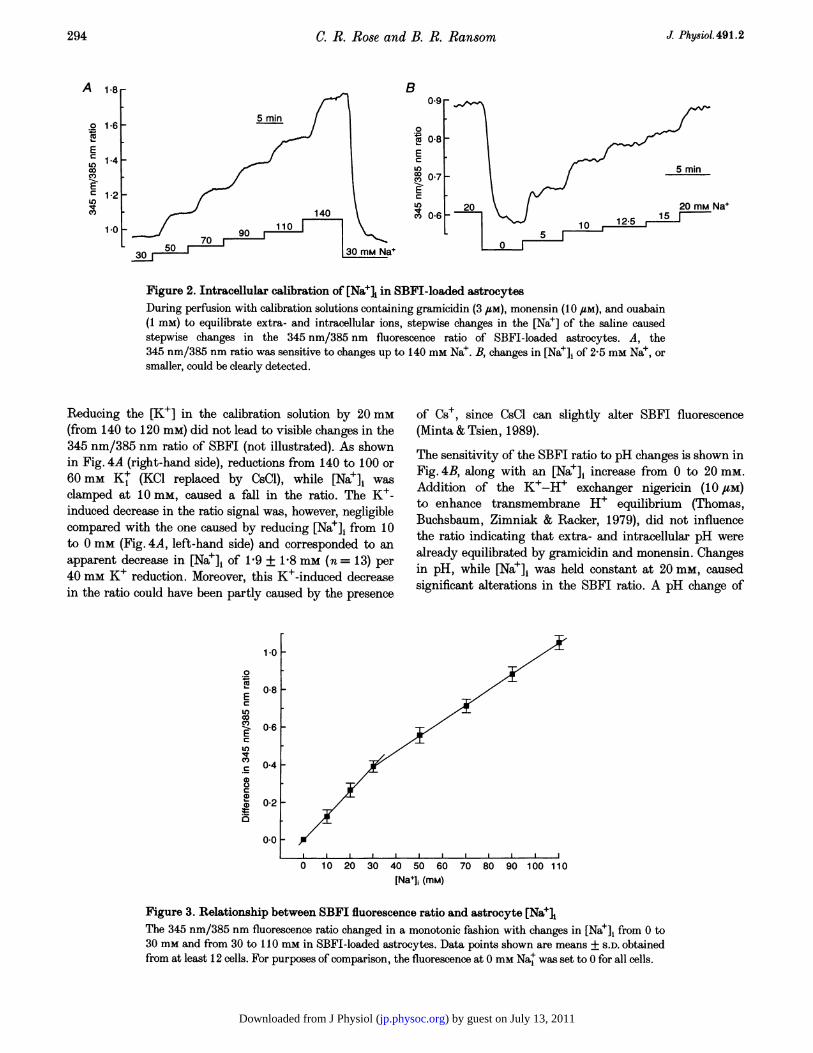
The ratio changed monotonically with changes in [Na+]1over the tested range of 0-140 mm (Fig. 2A and B). Theratio slope was around 0-13 (10 mM Na+)-1 below a [Na+], of30 mM and around 0-08 (10 mM Na+)-1 above 30 mM(Fig. 3). Changes of at least 2-5 mm Nat could be detected(Fig. 2B) in the physiological range around 15 mm Nat, andthe noise level of the population mean over time wastypically less than 1 mm.
Sensitivity of the SBFI fluorescence ratio tochanges in [K+] or [H+]The relative sensitivity of SBFI to Na+ vs. K+ has beenreported to be about 20: 1 in cell-free systems (Minta &Tsien, 1989). This fairly high K+ sensitivity, however,would make the dye unreliable for precise intracellular Na+measurements because intracellular [K+] ([K+]i) exceeds[Na+]i at least by a factor of 5 and is likely to changereciprocally with [Na+]1. On the other hand, SBFI's low pKa(-log of dissociation constant) of 6-1 in cell-free systems(Minta & Tsien, 1989) suggests that most physiological pHvariations would not effect the dye properties. As pointedout above, the dye has different spectral characteristicswhen located inside cells, including decreased sensitivity toK+ (Harootunian et al. 1989; Negulescu et al. 1990),necessitating direct evaluation of the effects of K+ and pHon the dye's performance in astrocytes. We thereforeperformed experiments to quantify the Na+/K+ andNa+/H+ sensitivities of SBFI in astrocytes, while perfusingthe cells with calibration saline containing gramicidin(3 /SM), monensin (10 /,M) and ouabain (1 mM) to equilibrateextra- and intracellular ion concentrations.
12 ,uM digitonin
100r1 % Triton X-1 00
3 min50
0'
Figure 1. Release of intracellular SBFI by membrane permeabilization with digitoninIn an SBFI-loaded astrocyte, clamped to an intracellular [Na+] of 20 mm by perfusion with salinecontaining gramicidin (3 /sM), monensin (10 /SM), and ouabain (1 mM), permeabilization of the plasmamembrane with the detergent digitonin (12 /sM) decreased the 345 nm fluorescence by 77%. Thisindicated that the majority of the dye was located in the cytoplasm. Application of Triton X-100 (1 %),to release dye located in intracellular organelles, caused the fluorescence to fall to 0. As in this figure, datatraces in all subsequent figures are from single representative astrocytes.
0-
(D0cJCoa)0
ECLO
C')
J Physiol. 491.2 293
) by guest on July 13, 2011jp.physoc.orgDownloaded from J Physiol (
C. R. Rose and B. R. Ransom J Physiol.491.2
0-9r
IF
-
LNa+
Figure 2. Intracellular calibration of [Nael in SBFI-loaded astrocytesDuring perfusion with calibration solutions containing gramicidin (3 FM), monensin (10 uM), and ouabain(1 mM) to equilibrate extra- and intracellular ions, stepwise changes in the [Na+] of the saline causedstepwise changes in the 345 nm/385 nm fluorescence ratio of SBFI-loaded astrocytes. A, the345 nm/385 nm ratio was sensitive to changes up to 140 mm Nae. B, changes in [Nae]1 of 2-5 mm Na+, orsmaller, could be clearly detected.
Reducing the [K+] in the calibration solution by 20 mm(from 140 to 120 mM) did not lead to visible changes in the345 nm/385 nm ratio of SBFI (not illustrated). As shownin Fig. 4A (right-hand side), reductions from 140 to 100 or60 mm K+ (KCl replaced by CsCl), while [Nae]i wasclamped at 10 mm, caused a fall in the ratio. The K+-induced decrease in the ratio signal was, however, negligiblecompared with the one caused by reducing [Nae]i from 10to 0 mM (Fig. 4A, left-hand side) and corresponded to anapparent decrease in [Nae]i of 1P9 + 1P8 mm (n = 13) per40 mm K+ reduction. Moreover, this K+-induced decreasein the ratio could have been partly caused by the presence
of Cs+, since CsCl can slightly alter SBFI fluorescence(Minta & Tsien, 1989).
The sensitivity of the SBFI ratio to pH changes is shown inFig. 4B, along with an [Nae]i increase from 0 to 20 mm.Addition of the K+-H+ exchanger nigericin (10 /M)to enhance transmembrane H+ equilibrium (Thomas,Buchsbaum, Zimniak & Racker, 1979), did not influencethe ratio indicating that extra- and intracellular pH werealready equilibrated by gramicidin and monensin. Changesin pH, while [Nae]i was held constant at 20 mm, causedsignificant alterations in the SBFI ratio. A pH change of
1-0 I
0
co
Ec)
LO
coU)C')C
LO
0C
0
08 F
06 -
0-4 F
0-2 F
0-0
90 100 110
Figure 3. Relationship between SBFI fluorescence ratio and astrocyte [Na+iThe 345 nm/385 nm fluorescence ratio changed in a monotonic fashion with changes in [Nae]i from 0 to30 mm and from 30 to I 10 mm in SBFI-loaded astrocytes. Data points shown are means + S.D. obtainedfrom at least 12 cells. For purposes of comparison, the fluorescence at 0 mm Nae was set to 0 for all cells.
I-
294
A 18
o 1-6
E' 1-4C')2c 1-2c)
1*0
B
5 min0co 0-8Ec
X 0-7
C') 06
0 10 20 30 40 50 60 70 80[Na+]J (mM)
) by guest on July 13, 2011jp.physoc.orgDownloaded from J Physiol (

J Physiol.491.2 Intracellular sodium in astrocytes
A 10 mM 10 mMNa+I Na
Na+
0
EC
CV)
It
CV)
1*0
0-9
60 mM 1 00 mMK+ K+
I
0-8 _
B
r .~~0EC
CV)
10 uM nigericin
pH1 6-6 pHi 6-2 pH, 7-4
5 min
Figure 4. Sensitivity of intracellular SBFI ratio to [K+; and pHiA, experiments performed while perfusing astrocytes with calibration solution containing gramicidin(3/uM), monensin (10 ,M), and ouabain (1 mM), showed that the K+-sensitivity of intracellular SBFI was
negligible. A decrease in [Nae], by 10 mm (left-hand side) caused a 2- to 3-fold larger decrease in the SBFIratio than decreasing the [K+], from 140 to 60 mm (right-hand side). B, with astrocytes in calibrationsolution, addition of nigericin (10 FM) did not alter the SBFI ratio, indicating that H+ ions were alreadyequilibrated due to gramicidin and monensin. Compared with an increase in [Nae]i of 20 mm, pH, changesof 0 4 units significantly influenced the 345 nm/385 nm ratio. Acidification from pH 7 0 to 6-6 and then6-2 caused decreases in the SBFI ratio, whereas alkalinization to pH 7-4 led to an increase in the dye ratio.On average a 0 4 pH unit shift caused a change in SBFI ratio that suggested a -6 mm change in [Nae]1.Upward-pointing arrows in this and the following figures indicate return to the control saline.
0 4 units mimicked a change in [Na+]1 of 6-1 + 1-5 mM(n = 14), with acidification leading to a decrease andalkalinization to an increase in the ratio (Fig. 4B).
Because of this pH sensitivity, all calibration solutions were
titrated to a pH of 7-15, the apparent baseline intracellularpH (pHi) of cultured rat hippocampal astrocytes inC02/HC037-buffered solutions (Pappas & Ransom, 1994).
Effect of extracellular [Na+] on baseline [Na']iCalibrations performed after each experiment revealed a
baseline [Nae], of 14-6 + 4.9 mm (n = 270) in culturedhippocampal astrocytes in standard C02/HCO3 -bufferedsaline containing 3 mm K+. This corresponds to a Naeequilibrium potential of +57 mV. For most experiments,two-point calibrations (0 and 30 mm Na+) were performed(Fig. 5). When [Nae]i increased to more than 30 mm during
O Na+-cal 30 Na+-cal
30r
25
i
z
0 Na+
20 _-
15
10
5
0
5 min
Figure 5. Effect of [Nae]. on [Nae]iRemoval of Nao (0 Na+) was followed by a decrease in [Nae]1. This decrease consisted of a rapid declinefollowed by a slower, linear decline that persisted (compare dashed lines). At the end of this experiment, a
two-point calibration was performed with saline solutions containing gramicidin (3juM), monensin(10 FM), and ouabain (1 mM), and 0 mm Nae (0 Na-cal) or 30 mM Nae (30 Na-cal), respectively. Note thetransient increase in [Nae], following perfusion with the 0 Nae-calibration solution (arrow; see text).
295
) by guest on July 13, 2011jp.physoc.orgDownloaded from J Physiol (
C. R. Rose and B. R. Ransom
an experiment, a three-point calibration using 0, 30 and50 mm Nae was performed.
Changing from standard saline to a calibration solutioncaused a rapid and transient increase in [Na+]i in about30% of the cells (Fig. 5, arrow) which was not investigatedfurther. It might indicate the presence of voltage-activatedNa+ channels opening during the mixing of standard salinewith the high-K+ calibration solution. Alternatively, Naeionophores might mediate the increase in [Nae]i byincreasing the Na+ permeablility before high extracellularNae ([Nae]o) has been completely replaced by the low [Nae].in the calibration saline (i.e. '0 Na-cal', Fig. 5).
Removal of [Nae]o caused a decrease in the 345 nm/385 nmratio consisting of a rapid phase during the first 1-2 min,followed by a slower phase (compare dashed lines in Fig. 5).The maximal decline in [Nae]i was 0-58 + 0-32 mM min-'(n = 22). Considering that these cells rapidly acidify byabout 0 4 pH units with Nae removal (Pappas & Ransom,1993, 1994), the first drop in the SBFI ratio in 0 Nae salinecould be partly due to the pH sensitivity of the dye. As theacidification fully develops in the first 2 min, the followingslower phase of the ratio decrease (0-15 + 0 09 mM min-';n = 22; Fig. 5) is likely to reflect the true loss of Nai. Asjudged by the subsequent calibration, [Nae]i approachedvalues near 0 after a 20 min exposure to 0 Na+ saline.
When Nao was reintroduced, [Nae]i rapidly increased tovalues higher than the initial baseline and subsequently fellby 4-6 + 3-2 mm (n = 22) to reach a new steady-state[Nae]i, which was significantly lower than the initialbaseline (Fig. 5). Since the intracellular pH returns tobaseline pH after a few minutes (Pappas & Ransom, 1993,1994), this lower steady-state ratio is not likely to beinfluenced by the pH sensitivity of SBFI.
Influence of COJHCO3- on [Nae]iHippocampal astrocytes appear to express an electrogenicNae-HCO3- cotransporter (O'Connor, Sontheimer &Ransom, 1994; Pappas & Ransom, 1994). This Nae-dependent transporter might influence [Na+]i, and weinvestigated this possibility. To activate Na+-HCOjcotransport, cells were perfused with C02/HCO,--free,Hepes-buffered saline for 30 min following dye loading andthen exposed to C02/HCO3--buffered saline. For this seriesof experiments, the pH of the calibration solutions wasadjusted to 6 9 to compensate for the more acidic intra-cellular pH of hippocampal astrocytes in the absence ofCO2/HCO3- (Pappas & Ransom, 1993).
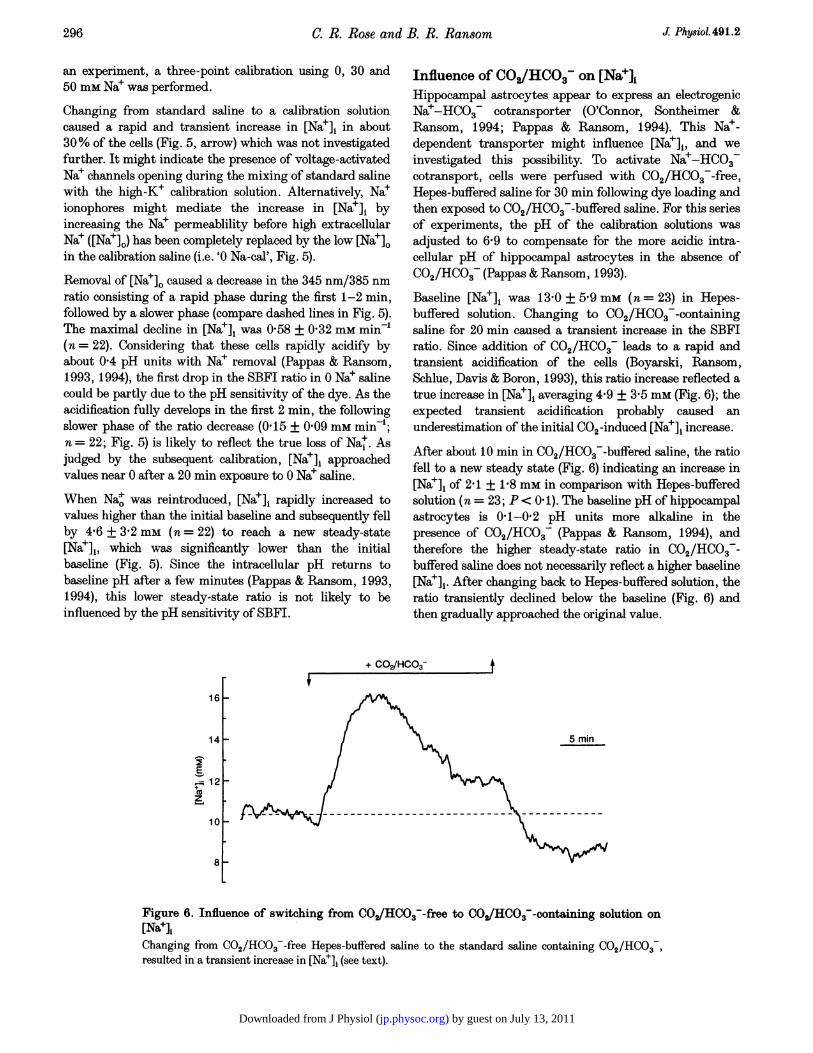
Baseline [Nae]i was 13 0 + 5.9 mm (n = 23) in Hepes-buffered solution. Changing to C02/HC03--containingsaline for 20 min caused a transient increase in the SBFIratio. Since addition of C02/HCO3- leads to a rapid andtransient acidification of the cells (Boyarski, Ransom,Schlue, Davis & Boron, 1993), this ratio increase reflected atrue increase in [Nae]i averaging 4-9 + 3.5 mm (Fig. 6); theexpected transient acidification probably caused anunderestimation of the initial CO2 -induced [Nae]i increase.
After about tO min in CO2/HCO3--buffered saline, the ratiofell to a new steady state (Fig. 6) indicating an increase in[Nae]i of 2-1 + 1-8 mm in comparison with Hepes-bufferedsolution (n = 23; P< 0f 1). The baseline pH of hippocampalastrocytes is 0f1-0f2 pH units more alkaline in thepresence of CO2/HC03 (Pappas & Ransom, 1994), andtherefore the higher steady-state ratio in CO2/HCO3-buffered saline does not necessarily reflect a higher baseline[Na+]1. After changing back to Hepes-buffered solution, theratio transiently declined below the baseline (Fig. 6) andthen gradually approached the original value.
+ C02/HC03-
161
14 -
12z
f
5 min
lO1
8
Figure 6. Influence of switching from COHCO3 -free to COJHCO3-containing solution on[NaelChanging from C02/HC03--free Hepes-buffered saline to the standard saline containing C02/HC03,resulted in a transient increase in [Na+]i (see text).
296 J Physiol.491.2
) by guest on July 13, 2011jp.physoc.orgDownloaded from J Physiol (
Intracellular sodium in astrocytes
12
E
Caz
5 min
10 -
8
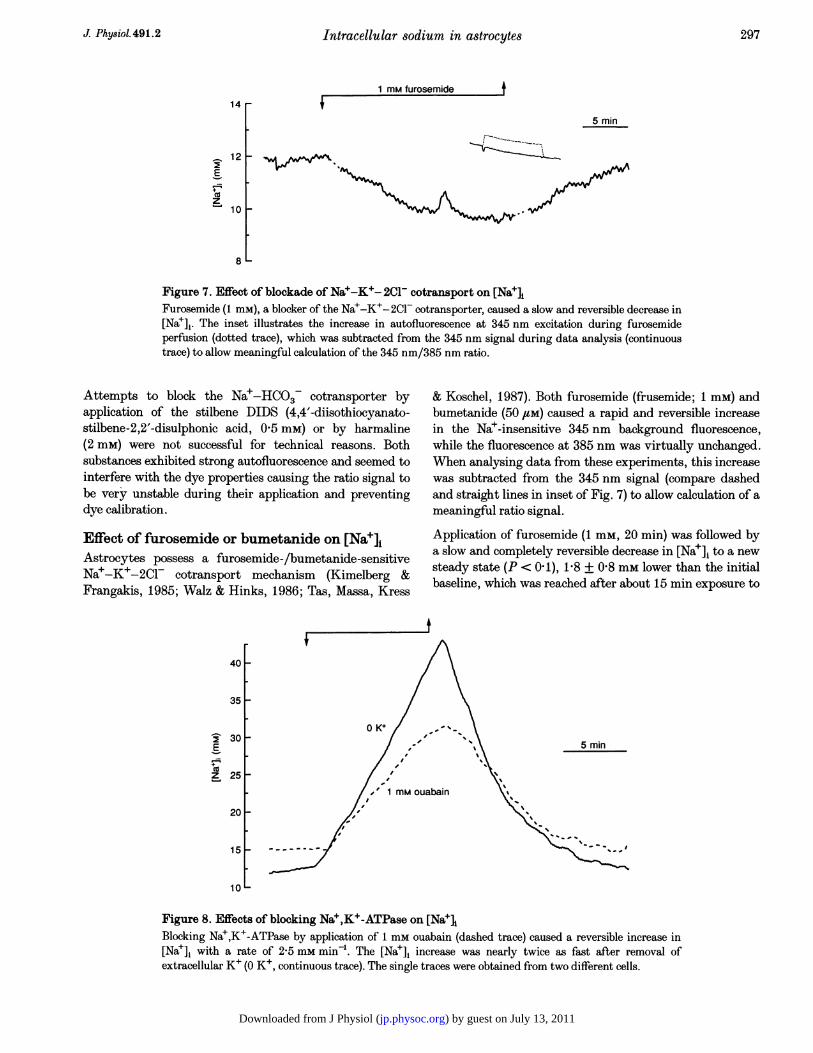
Figure 7. Effect of blockade of Na+-K+- 2C1- cotransport on [NajlFurosemide (1 mM), a blocker of the Na+-K+-2Cl- cotransporter, caused a slow and reversible decrease in[Nae],. The inset illustrates the increase in autofluorescence at 345 nm excitation during furosemideperfusion (dotted trace), which was subtracted from the 345 nm signal during data analysis (continuoustrace) to allow meaningful calculation of the 345 nm/385 nm ratio.
Attempts to block the Na+-HCO3- cotransporter byapplication of the stilbene DIDS (4,4'-diisothiocyanato-stilbene-2,2'-disulphonic acid, 0 5 mm) or by harmaline(2 mM) were not successful for technical reasons. Bothsubstances exhibited strong autofluorescence and seemed tointerfere with the dye properties causing the ratio signal tobe very unstable during their application and preventingdye calibration.
Effect of furosemide or bumetanide on [Naf]iAstrocytes possess a furosemide-/bumetanide-sensitiveNa+-K+-2Cl- cotransport mechanism (Kimelberg &Frangakis, 1985; Walz & Hinks, 1986; Tas, Massa, Kress
& Koschel, 1987). Both furosemide (frusemide; 1 mM) andbumetanide (50 /M) caused a rapid and reversible increasein the Na+-insensitive 345 nm background fluorescence,while the fluorescence at 385 nm was virtually unchanged.When analysing data from these experiments, this increasewas subtracted from the 345 nm signal (compare dashedand straight lines in inset of Fig. 7) to allow calculation of ameaningful ratio signal.
Application of furosemide (1 mm, 20 min) was followed bya slow and completely reversible decrease in [Na+]i to a newsteady state (P < 0 1), 1-8 + 0-8 mm lower than the initialbaseline, which was reached after about 15 min exposure to
401-
35
30 _E
z 25
0 K+
5 min
20 _
15 _
10 L_
Figure 8. Effects of blocking Nae,K+-ATPase on [NaeIBlocking Na+,K+-ATPase by application of 1 mm ouabain (dashed trace) caused a reversible increase in[Nae]i with a rate of 2-5 mm min-. The [Na+]i increase was nearly twice as fast after removal ofextracellular K+ (O K+, continuous trace). The single traces were obtained from two different cells.
liim iurubtfilllufd
J Physiol.491.2 297
14 r-
) by guest on July 13, 2011jp.physoc.orgDownloaded from J Physiol (

C. R. Rose and B. R. Ransom
the drug (n = 15; Fig. 7). Bumetanide (50 /SM) had virtuallythe same effect on [Na']i ([Na+]j decreased by 1P8 ± 0 9 mM,n = 18), which was also reversible (not shown). Theseexperiments indicated that the Na+-K+- 2Cl- cotransportercaused a steady Nae influx and contributed to the baseline[Nae], in these cells.
Effect of tetrodotoxin on [Nae]iThe presence of voltage-gated Na+ channels has beendemonstrated in cultured hippocampal astrocytes(Sontheimer et al. 1991). We determined whether voltage-gated Na+ channels influenced [Nae], by applying theblocker tetrodotoxin (TTX, 100,uM) for 15 min. Duringthese experiments the flow rate was reduced to about0 7 ml min-' to minimize the amount ofTTX necessary foreach experiment. The [Nae], was unchanged in most cells,but in six of forty cells investigated (15%) a change in[Nae]i was detected. In three cells, TTX reduced [Nae]i by0 37 mm min-', an effect which was not fully reversibleduring the course of the experiment (washout time about20 min) (data not shown). In three other cells, 5TXreversibly stopped a steady [Nae]i increase by-0-27 mm min-', observed under control conditions (datanot shown).
Effects of blocking Na+,K+-ATPase on [Nae]iOne of the most important functions of the Na+,K+-ATPase is the maintenance of a low [Na+]i against a steepinwardly directed electrochemical gradient for Nae. We
A
E
z
13
12
1 1
109
8
7
studied the influence of Na+,K+-ATPase inhibition on[Na+]1 in astrocytes using the cardiac glycoside blockers ofthe Na+,K+-ATPase, ouabain or strophanthidin, or byremoval of K+, which also blocks this transporter.
Ouabain (1 mM) caused [Na+]i to increase at a rate of2-5 + 0 9 mm min-' (determined during the first 3 min ofdrug exposure, n = 24). During a 10 min exposure to thedrug, [Nae], increased to 41P9 + 8-7 mm. After its removal,[Na+]j rapidly recovered in all cells observed (n = 24;Fig. 8). Strophanthidin application (1 mM) caused [Nae], toincrease at a rate of 1-9 + 1-6 mm min-', an effect whichwas fully reversible in seven of eighteen cells investigated(washout time 20 min; data not shown).
Removal of K+ caused [Na+]i to increase at a rate of4-2 + 1-8 mm min-1, faster than with ouabain or strophan-thidin. After a 10 min exposure to the K+-free saline,[Nae]i had increased to 60-0 + 21-0 mM (n = 44, Fig. 8).This increase was fully reversible in only 32% of the cells(washout time 20 min; Fig. 8).
Effects of small changes in [K+]O or [Nae] on[Nae]iIntense neural activity can cause brain [K+]O to increasefrom a baseline level of 3 mm up to a so-called 'ceiling-level'of 12-13 mM (Heinemann & Lux, 1977), whereas [Nae]odecreases by about 1 mm (Dietzel, Heinemann, Hofmeier &Lux, 1982). Glial cells play a key role in the regulationof [K+]o by uptake of excess extracellular K+. The
8 mM K+ 13 mM K+
B 20[
E 15_
zCv 5 min10j_7
Figure 9. Effects of small increases in [K+]. on [Nae1A, increasing [K+]o from 3 mm to 4, 5, 6, 7 or 8 mm for 3 min immediately decreased [Na+]j. Note,however, that the decreases in [Nae], did not increase in a linear fashion with increases in [K+]O, and thatthe exposure to elevated [K+]O in this set of experiments was too short to make statements about a newsteady-state [Nae]1. The decrease in [Nae], was greater for switches to 5 mm K+ than for switches to 7 mMK+ (see also Fig. 10). B, the effects of 10 min elevations of [K+]O from 3 to 8 mm and subsequently to13 mm on steady-state [Nae], are shown.
J Phy8iol. 491.2298
4 mm 5 mm 6 mm 7 mm 8 mm
fn, iK+ t T.+j
5 min
) by guest on July 13, 2011jp.physoc.orgDownloaded from J Physiol (

Intracellular sodium in astrocytes
8 mMK+
14
E
z
12
10
8
6
5 mMg8 mM
K-
50 /M bumetanide5 mM
5 min
Figure 10. Effects of bumetanide on KE-induced decreases in [Nae1Application of bumetanide (50 uM) to block the Na+-K+-2CF- cotransporter doubled both the slope andthe amplitude of the [Nae], decrease caused by a 3 min elevation of [K+]. from 3 to 8 mm (compare dottedlines). In contrast, the slope of the [Na+]i decrease caused by switching to 5 mm K+ was reduced (comparedashed lines), and its amplitude was only slighty increased (see text).
mechanisms of glial K+ uptake are not fully understood,but it has been proposed that it is mediated by activation ofNa+,K+-ATPase and/or Na+-K+-2C- cotransport, as wellas by passive influx of KCl through Ba2+-sensitive ionchannels (Walz & Hertz, 1982; Ballanyi, Grafe & TenBruggencate, 1987).
We investigated the influence of small changes in [K+]O and[Nae]o on astrocyte [Nae]1. Increasing [K+]O by as little as1 mM caused a decrease in [Na+]j. Neither the amplitude,nor the time course of the K+-induced [Na+]i decreases,however, changed in a linear fashion with increases in[K+]O. These results were consistent regardless of the orderin which changes in [K+]o were made.
Typical [Nae], responses to a series of small, 3 minduration changes in [K+]O are illustrated in Fig. 9A. Theseapplication periods were too brief to result in a new steady-state [Nae]1, defined as a stable [Nae]i for at least 5 minduring a given manipulation. A 3 min elevation of [K+]Ofrom 3 to 4 mm, was followed by a rapid and reversibledecrease in [Nae], to a level, which was 2-5 + 1-5 mm lowerthan the control [Nae]i (n = 42; Fig. 9A). Stepping from 3
A 2 mM K+ 1 mM K+
50 - j.\ 5 minj40 -
E30 -
Z 20 -
10
to 5 mM Ko for 3 min caused [Na+]i to decrease transientlyby 3 0 + 1P9 mm (n = 42), followed by an increase towardsthe initial [Nae]1, even when [K+]O was still elevated.Increasing [K+]o to 6 or 7 mm, however, led to only smalland much slower decreases in [Nae], (12 + 0-8 and1-4 + 0-8 mm, respectively). During a 3 min exposure to8 mM K+, [Nae], fell by 2-3 + 0-6 mM (n = 42; Fig. 9A).
During longer exposures to 8 mm K+ solution (> 10 min),[Na+] fell by 5-8 + 2-2 mM (n = 22; Fig. 9B). This newsteady-state [Nae]i was reached after 5-8 min and wassignificantly lower than the control value in 3 mm K+. Insaline containing 13 mm K+ (applied for > 10 min), steady-state [Nae]i decreased by 7-1 + 2'9 mm, again significantlylower than in control saline with 3 mm K+ (n = 22;Fig. 9B).
We investigated the influence of Na+-K+-2C- cotransporton the [K+]O-induced decreases in [Na+]i, using the selectiveblocker bumetanide (50 /sm; see above). Bumetanide loweredbaseline [Nae]i and significantly increased both the slope ofthe [Na+]i reduction caused by a 3 min exposure to 8 mMK+ (from 0-8 + 0-6 to 1P6 + 0-6 mm min-) and its
B
E
z
120 mM Na+ 160 mM Na+ tT
5 min
Figure 11. Effects of decreases in [KfL or changes in [NaeL on [NaelA, reduction in [K+]O from 3 to 2 mm increased [Nae]i by about 3 mm, whereas reduction to 1 mm K+ ledto a steady increase in [Nae], that persisted until standard saline was returned. B, changing [Nae]o by20 mm resulted in small changes in [Nae]1 of 2-3 mM.
299J Phy8iol.491.2
) by guest on July 13, 2011jp.physoc.orgDownloaded from J Physiol (
C. R. Rose and B. R. Ransom
amplitude (from 1-6 + 0-8 to 3-6 + 1P4 mm min-', n = 35;Fig. 10). Surprisingly, bumetanide significantly slowed,rather than accelerated, the [Na+]i decrease caused bya 3 min exposure 5 mm K+ (from 4-7 + 3-7 to1P4 + 1'2 mM min-'; n = 35). Since [Nae]i fell during theentire exposure to 5 mM K+, the amplitude of the [Na+],decrease was, however, slightly larger than in the control(it increased from 2'7 + 1P0 to 3-1 + 1 1 mm; n = 35;P< 0 1; Fig. 10).
[Na+]i increased when [K+]. was decreased. As shown inFig. 1 1A, perfusion with saline containing 2 mm K+ for15 min, increased steady-state [Nae]i by 3 0 + 2-3 mM(n = 23; P< 0 05) compared with baseline. Reduction of[K+]0 to 1 mm caused a steep increase in [Na+]i thatcontinued during the entire application period (15 min).The slope of this [Nae], increase was 1P9 + 0-8 mM min-'(n = 23) which was comparable with the [Na+], increasecaused by inhibition of Na+,K+-ATPase by 1 mmstrophanthidin.
Changes in [Nae]. by 10 or 20 mm only slightly alteredsteady-state [Nae]j. A 15 min reduction of [Nae]o from 140to 130 mm (Nae replaced with N-methyl-D-glucamine(NMDG) to maintain isosmolarity) resulted in a reductionof steady-state [Nae]i by 0 9 + 1-6 mM (n = 16; P< 0-25;not shown). A reduction of [Nae]o by 20 mM, significantlydecreased [Nae], by 2-7 + 2-3 mM (n = 43; Fig. liB).Addition of 20 mm Nae was followed by an increase in[Nae]i by 1-6 + 1-7 mm (n= 27; P< 0 1; Fig. llB). Thisincrease could have been, at least partly, caused by anincrease in osmolarity, because addition of 20 mm NMDGalso slightly increased [Nae]i by 0-9 +10 mm (n = 16;P< 0 4; data not shown).
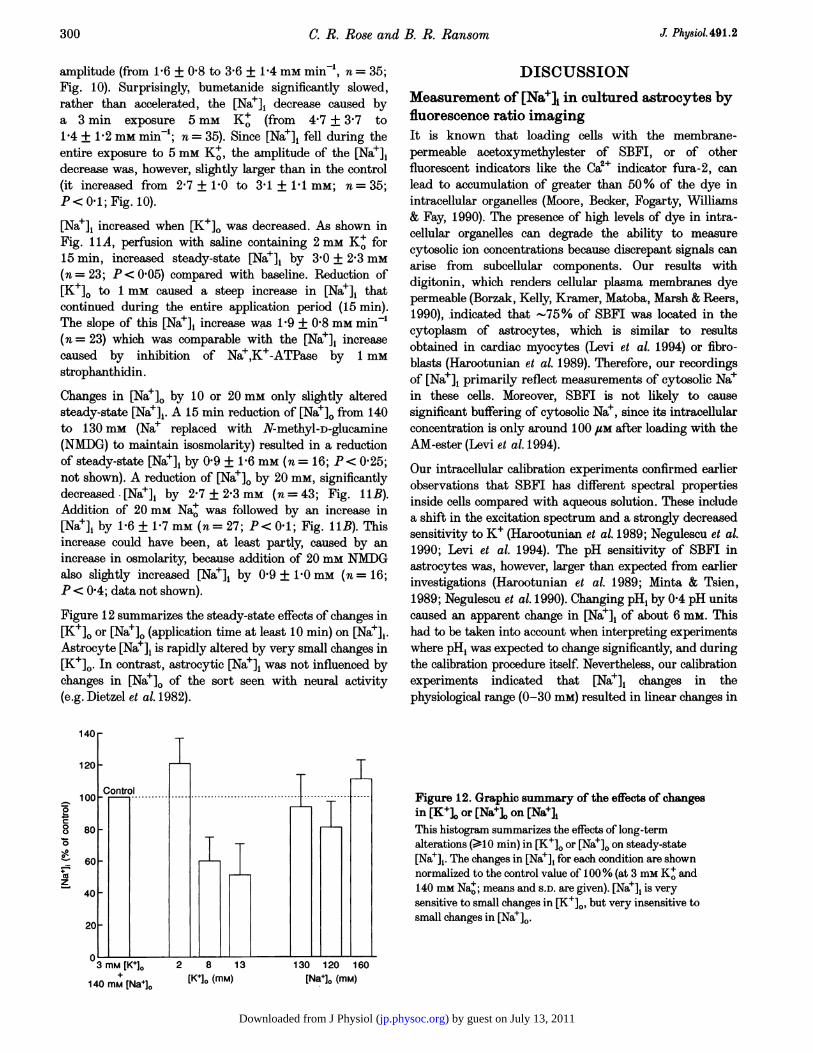
Figure 12 summarizes the steady-state effects of changes in[K+]. or [Nae]o (application time at least 10 min) on [Nae]j.Astrocyte [Nae], is rapidly altered by very small changes in[K+]O. In contrast, astrocytic [Na+], was not influenced bychanges in [Na+]o of the sort seen with neural activity(e.g. Dietzel et al. 1982).
140rr
-
0c
m-0
oz
140 mM [Na+]o2 8
[K+]o (mm)130 120 160
[Na+]o (mM)
DISCUSSION
Measurement of [Na]i in cultured astrocytes byfluorescence ratio imagingIt is known that loading cells with the membrane-permeable acetoxymethylester of SBFI, or of otherfluorescent indicators like the Ca2+ indicator fura-2, canlead to accumulation of greater than 50% of the dye inintracellular organelles (Moore, Becker, Fogarty, Williams& Fay, 1990). The presence of high levels of dye in intra-cellular organelles can degrade the ability to measurecytosolic ion concentrations because discrepant signals canarise from subcellular components. Our results withdigitonin, which renders cellular plasma membranes dyepermeable (Borzak, Kelly, Kramer, Matoba, Marsh & Reers,1990), .indicated that -75% of SBFI was located in thecytoplasm of astrocytes, which is similar to resultsobtained in cardiac myocytes (Levi et al. 1994) or fibro-blasts (Harootunian et al. 1989). Therefore, our recordingsof [Na+]i primarily reflect measurements of cytosolic Na+in these cells. Moreover, SBFI is not likely to causesignificant buffering of cytosolic Nae, since its intracellularconcentration is only around 100 ,uM after loading with theAM-ester (Levi et al. 1994).
Our intracellular calibration experiments confirmed earlierobservations that SBFI has different spectral propertiesinside cells compared with aqueous solution. These includea shift in the excitation spectrum and a strongly decreasedsensitivity to K+ (Harootunian et al. 1989; Negulescu et al.1990; Levi et al. 1994). The pH sensitivity of SBFI inastrocytes was, however, larger than expected from earlierinvestigations (Harootunian et al. 1989; Minta & Tsien,1989; Negulescu et al. 1990). Changing pH, by 0 4 pH unitscaused an apparent change in [Nae]i of about 6 mm. Thishad to be taken into account when interpreting experimentswhere pH, was expected to change significantly, and duringthe calibration procedure itself. Nevertheless, our calibrationexperiments indicated that [Na+], changes in thephysiological range (0-30 mM) resulted in linear changes in
Figure 12. Graphic summary of the effects of changesin [K+]o or [Na]o on [Nae],This histogram summarizes the effects of long-termalterations (>10 min) in [K+]. or [Nae]. on steady-state[Nae],. The changes in [Na']i for each condition are shownnormalized to the control value of 100% (at 3 mm K+ and140 mm Nao; means and S.D. are given). [Na], is verysensitive to small changes in [K+]0, but very insensitive tosmall changes in [Nae]o.
J. Physiol. 491.2300
1 1
) by guest on July 13, 2011jp.physoc.orgDownloaded from J Physiol (

Intracellular sodium in astrocytes
the 345 nm/385 nm ratio of SBFI. Therefore, absolute[Na+]i could be extrapolated from two- or three-pointcalibrations at the end of each experiment.
Baseline [Na]i in astrocytesIn a physiological solution at 37 °C, we found that [Na+],was about 15 mm in cultured rat hippocampal astrocytes,corresponding to an intracellular Nae activity (aNaj) ofabout 11 mm. Using Nae-sensitive microelectrodes, Buehrle& Sonnhof (1983) reported an aNa, around 40 mm in glialcells of the frog spinal cord. The aNai in glial cells, probablyastrocytes, of guinea pig olfactory cortex slices was 25 mM(Ballanyi et al. 1987). Cultured mouse oligodendrocyteswere reported to have an aNa, of 15 mm in physiologicalsolutions with 5-4 mm K+ (Ballanyi & Kettenmann, 1990).[Nae], of C6 glioma cells was about 6 mm (Erecinska,Dagani, Nelson, Deas & Silver, 1991). Using radioisotopetechniques, an [Na+]1 of 28-1 mm was calculated in culturedmouse astrocytes (Walz & Hertz, 1983). Glial [Nae], has alsobeen measured in invertebrate preparations: aNa1 of glialcells in drone retina was 37 mm (Coles & Orkand, 1985),whereas aNa1 was 5-11 mm in leech neuropile glial cells(Doerner, Ballanyi & Schlue, 1990; Deitmer, 1991).
The higher values of [Nae], measured previously invertebrate astrocytes might reflect special properties of theparticular cells under study, or these differences could betechnique dependent. For example, ion-sensitive electrodesmight damage small cells during impalement and produceartificially high [Nae]1. In this regard, the use of intra-cellular dyes for measuring ion concentration offers theadvantage of being very non-invasive.
Interestingly, in studies in which [Nae]i of both neuronsand glial cells were determined for the same preparation(and with the same technique), glial [Nae], was alwayshigher than neuronal [Nae]i (Buehrle & Sonnhof, 1983;Coles & Orkand, 1985; Doerner et al. 1990). The meaningof this is not clear, but perhaps the higher glial [Na]iassists in K+ removal by insuring that [Nae]i is not therate-limiting step for glial Na+,K+-ATPase activity, whichcontributes to K+ uptake (Sweadner, 1995).
Transmembrane Nae movementNae effluxRemoval of Nao led to a substantial decrease of [Na+]i incultured astrocytes, as also reported by Sontheimer et al.(1994). Na+ efflux is favoured thermodynamically by thissituation and Nae movement could occur by passive move-ment through channels or via Na+-dependent transporterslike the Na+-H+ exchanger, Na+-Ca2+ exchanger, or theNa+-HCO3- cotransporter.
The decrease in the 345 nm/385 nm ratio after removal ofNa+ consisted of a rapid phase in the first minutes followedby a slow, linear decline in [Nae]1. As pointed out inResults, the first rapid phase could have been amplified byan artifact due to the pH sensitivity of SBFI. However,
very similar biphasic time courses of [Na+]i decrease uponremoval of Nat+ were observed in gastric cells (Negulescu etal. 1990) and in leech neuropile glial cells (Deitmer, 1992).Since the latter study was performed using Na+-sensitivemicroelectrodes, the pH-induced artifact in our experimentswas presumably absent. The maximal rate of Nae loss inthe leech glial cells amounted to 1P35 mm min-', which isabout twice as fast as observed in the present study. Itsslope was reduced in C02/HC03--buffered solution,indicating that a significant amount of the rapid Nae losswas due to outwardly directed Nae-HCO3- cotransport inthe leech (Deitmer, 1992). Although not tested in our cells,this is a plausible mechanism for the reduction in [Nae]seen here with removal of external Na+.
Nae-dependent transportNae-HCO3- cotransport. Electrogenic Nae-HCO3cotransport, operating with a stoichiometry of2 HC03-: 1 Nae, has been described in several glial celltypes including cultured astrocytes from hippocampus(O'Connor et al. 1994; Pappas & Ransom, 1994). Thetransient increase in [Na+]i by about 5 mm uponintroduction of C02/HC03- in the present study could bedue, at least partly, to the activation of inwardly directedNa+-HCO3- cotransport, which is favoured under theseconditions because of the steep inwardly directed electro-chemical gradient for HC03- and Nae.The steady-state contribution of the Nae-HCO3- co-transporter to baseline [Nae]i of hippocampal astrocytesseems relatively small. If one neglects that part of thesteady-state SBFI ratio increase in C02/HC03--bufferedsolution might be due to an artifact caused by the pHsensitivity of SBFI (see Results), the presence ofC02HC03- caused only a 2 mm, or 10%, increase inbaseline [Na+]j. In contrast, baseline [Na+]i of leechneuropile glial cells increased by as much as 40%, whenCO2/HCO3- was added to cells bathed in C02/HC03--freesolution due to activation of Na+-HCO3- cotransport(Deitmer, 1991, 1992).
Na+-K+-2ClP cotransport. Several investigations havedescribed an electroneutral, inward transport of Nae, K+,and Cl- into cultured astrocytes, which can be blocked byfurosemide and bumetanide (Kimelberg & Frangakis,1985). This Na+-K+-2Cl- cotransport appears to play arole in regulatory volume increase (Kimelberg & Frangakis,1985), and in the clearance of elevated extracellular K+ byastrocytes (Walz & Hertz, 1982). Its relatively low value forhalf-maximal K+ activation, which is 2-7 mm (Tas et al.1987), suggests, however, that the transporter is constantlyactive, even at a baseline [K+]0 of about 3 mm.
Application of furosemide or bumetanide to hippocampalastrocytes resulted in a slow, reversible decrease in [Na+]iby about 2 mm. This indicated that the Na+-K+-2Clcotransporter was responsible for a steady inward movementof Na+ under control conditions, and contributed to baseline
J Phy8iol. 491.2 301
) by guest on July 13, 2011jp.physoc.orgDownloaded from J Physiol (

C. R. Rose and B. R. Ransom
[Na+]i in these cells. These results support a modelproposed by Walz & Hinks (1986) that inward transport ofNae via the Na+-K+-2Cl cotransporter fuels Na+,K+-ATPase, thereby completing a so-called 'transmembraneNae cycle' in astrocytes. But Na+-K+-2CG- cotransport isnot the only way that Nae can enter hippocampalastrocytes as noted above (and see below).
Voltage-gated Nae channelsHippocampal astrocytes in culture express TTX-sensitive,neuron-like Na+ currents from day 1 to about day 5 inculture, whereas less ITX-sensitive Na currents with glial-type characteristics are observed after that period. Thepercentage of astrocytes that express Na currents decreasesfrom 75 to 30% with increasing time in culture (Sontheimeret al. 1991). This means that in the present study aboutone-third of the cells investigated should have voltage-gated Nae channels.
The function of these and other voltage-activated ionchannels in glial cells is still under discussion. In culturedspinal cord astrocytes, which have a very high density ofNae channels, application of high concentrations of TTXled to a decrease in [Na+]i as measured by ratio imaging ofSBFI (Sontheimer et al. 1994). These and other results ledto the suggestion that Nae entered these cells through asmall population of Na+ channels open at resting membranepotential. It was argued that this Nae influx was essentialfor cell survival because it provided a source of intracellularNae for maintenance of Na+,K+-ATPase activity(Sontheimer et al. 1994).
In our study, most astrocytes showed no change in [Nae]iwith application of 100 FM TTX. In about 15% ofhippocampal astrocytes, TTX slowly decreased [Nae]j, orreversibly stopped an increase in [Na+]i which was observedin normal saline before TTX application. Under theassumption that these effects were restricted to the 30% ofcells which possess voltage-gated Nae channels, it can beconcluded that baseline [Nae]1 might be elevated in a smallnumber of cells by Na+ currents present at resting membranepotential. It seems doubtful that Nae influx through Naechannels was critical for normal cell function, even in thosefew cells that exhibited TTX sensitivity, because otherpathways for Nae influx exist (cf. Sontheimer et al. 1994).
Nae,K+-ATPaseNa+,K+-ATPase activity establishes and maintains theelectrochemical gradients for Na+ and K+ across the cellmembrane. Its activity is, therefore, essential for a multitudeof other transport processes which use the inwardly directedNa+ gradient as an energy source, like the Na+-Ca2+exchanger, or the Na+-H+ exchanger. The glial isoform ofthis enzyme is relatively insensitive to cardiac glycosides,requiring concentrations of up to 1 mm for inhibition (Walz& Hertz, 1982; Sweadner, 1995). Its inhibition leads to aslow membrane depolarization of cultured astrocytes byabout 1 mV min-' (Sontheimer et al. 1994).
After inhibition of the Na+,K+-ATPase with ouabain orstrophanthidin, [Nae]i of hippocampal astrocytes increasedvery rapidly by 2-5 and 1-9 mm min'1, respectively. Thesevalues for [Nae]i with Na+,K+-ATPase inhibition are similarto those observed in other cell types, like smooth musclecells (Moore & Fay, 1993), or gastric cells (Negulescu et al.1990). They reflect high Nae influx across astrocytic cellmembranes driven by the steep electrochemical gradient forthis ion. In contrast to astrocytes, cultured mouse oligo-dendrocytes exhibit less Nae influx with Na+,K+-ATPaseinhibition. In 40% of the cells investigated, ouabainapplication increased [Nae], only by about 1P3 mm min-',and it had an even smaller, or barely detectable, effect onthe rest of the cells (Ballanyi & Kettenmann, 1990). Thesurface/volume ratio of oligodendrocytes is probablysmaller than with the cells studied here and this couldcontribute to the slower [Na+]i increase in oligodendrocytes.
Following removal of extracellular K+, the rate of [Na+]iincrease was twice that seen with cardiac glycosides. Thiscould indicate that K+ removal produces a more completeblock of Na+,K+-ATPase activity than these drugs, partlybecause extracellular K+, present in the standard saline,might have reduced the binding of the cardiac glycosides(Sweadner, 1995). Application of K+-free saline, however,not only inhibits Na+,K+-ATPase activity, but may alsoforce the Na+-K+-2Cl- cotransporter to operate in theoutward mode. At least transiently, astrocytes will bestrongly hyperpolarized by the removal of K+, and thiswill also increase the driving force for Na+ influx. Both ofthe latter could explain the steeper [Nae]i increase in 0 K+as compared with that seen with pharmacologicalinhibition of Na+,K+-ATPase.
Effects of small changes in [K+]O on [Nat]j:implications for extracellular K+ homeostasisNeural activity causes increases in [K+]O that would reachlevels capable of disrupting normal brain function if K+removal was purely on the basis of passive diffusion(Ransom & Sontheimer, 1992; Newman, 1995). Astrocytesaugment removal of excess K+ by direct uptake, and do soby several mechanisms. These include K+ influx throughBa2+-sensitive K+ channels, and activation of Na+,K+-ATPase and Na+-K+-2Cl- cotransport by elevated [K+]O(Walz & Hertz, 1982; Kimelberg & Frangakis, 1985; Grafe& Ballanyi, 1986; Walz, 1989). Both of the latter processescould directly influence [Nae], in these cells. Indeed, the[Na+]i of cultured mouse oligodendrocytes, as measuredwith ion-sensitive microelectrodes, fell by 5 mm whenincreasing [K+]O by 10 mm (Ballanyi & Kettenmann, 1990).Changes in intraglial [Nae]i following increases in [K+]0were also observed under more physiological conditions.During electrical stimulation, the [Nae], of glial cells inguinea-pig olfactory cortex slices decreased by about 8 mmin parallel with an increase in extracellular and intraglialK+ (Ballanyi et al. 1987). This is in sharp contrast toneurons, where the [K+]i decreases and the [Na+]i increases
J Phy8iol. 491.2302
) by guest on July 13, 2011jp.physoc.orgDownloaded from J Physiol (

Intracellular sodium in astrocytes
during activity (Grafe, Rimpel, Reddy & Ten Bruggencate,1982; Coles & Orkand, 1985; Lasser-Ross & Ross, 1992).
Small increases in [K+]. powerfully decreased [Na+]i inhippocampal astrocytes. One millimolar increases in [K+]O,from 3 to 4 mm, decreased [Na+]1 by about 2 5 mM. This[Nae], decrease was probably due to the activation ofNa+,K+-ATPase by increased [K+]0, because the ouabain-sensitive uptake of K+ into cultured astrocytes has beenshown to saturate only at an [K+]0 of 10-15 mm (Grisar,Frere & Franck, 1979; Walz & Hinks, 1986). On the otherhand, the stimulus-induced increases of glial [K+], in theguinea-pig olfactory cortex, as well as the decrease in[Nae]i following high-K+ solution in cultured mouseoligodendrocytes, was only partially blocked by ouabain(Ballanyi et al. 1987; Ballanyi & Kettenmann, 1990). Theseresults indicate a contribution of cell swelling to the decreasein intraglial [Nae]i (Walz & Hinks, 1986). The outwardtransport of Na+ due to enhanced Na+,K+-ATPase activityis apparently superimposed on the inward Na+ transport bythe K+ activation of Na+-K+-2C- cotransport (Walz &Hinks, 1986), as well as by the Nae-HCO3- cotransporter,which is activated by the membrane depolarization accom-panying the increase in [K+]0 (Pappas & Ransom, 1994).Ion and water movements associated especially withNa+-K+-2Cl- cotransport cause cell swelling (Walz &Hinks, 1986).
In fact, the decrease in [Nae]1 with increase in [K+]0 wasnon-linear, suggesting that the [K+]. increase recruiteddifferent mechanisms, some tending to decrease [Nae]1, andsome tending to increase it. The effects of blocking
A[K+]o = 3 mM
Na+-K+-2Cl- cotransport on K+-induced changes in [Na+]iwere complex, but this transporter appeared to transportNae into astrocytes when [K+]. was increased from 3 to8 mm. Because very little change in [Nae]i occurred when[K+]. was increased from 3 to 6 mm (see Fig. 9), it isplausible that at this [K+]O, Nae removal by Na+,K+-ATPase balances Na+ influx by Na+-K+-2ClF cotransport,and perhaps other transporters, with minimal net changein [Nae]1.Whereas the reduction of [K+]0 to 2 mm caused only asmall elevation in [Nae],, we observed a steep increase in[Nae]1 following reduction of [K+]0 to 1 mm. These effectscan also be explained by K+ actions on Na+,K+-ATPaseactivity; lowering [K+]0 would remove baseline stimulationof this enzyme and also increase the driving force for Naeinflux by causing hyperpolarization.
ConclusionsThe present investigation shows that fluorescence ratioimaging with the dye SBFI is a valuable tool for themeasurement of absolute values of [Na+], in culturedastrocytes. As in other cell types, maintenance of low[Nae]i against a steep electrochemical gradient is mainlyachieved by the Na+,K+-ATPase (Fig. 13A). Our findingsindicated that this enzyme is also well suited to take upexcess K+, because it was quickly activated by smallincreases in [K+]O. The Na+,K+-ATPase slowed withsubnormal [K+]0 and this would allow K+ accumulatedwithin astrocytes to be slowly returned to the extracellularspace during the period of decreased [K+]0 that followsneural activity (Ransom & Sontheimer, 1992).
B
Na+K+2
2CI-
Figure 13. Model of [Nael homeostasis in rat hippocampal astrocytes under normal conditionsand in the presence of elevated [K1]OA, under baseline conditions where [K+]o is about 3 mm, astrocyte [Nae]i is about 15 mm. Three transportmechanisms probably involved in determining the resting value of [Nae]i are shown. Voltage-gated Naechannels contribute to [Nae]i homeostasis under these conditions in only a minority of astrocytes (impliedby the dashed line). B, when neural activity increases [K+]O, as shown here from 3 to 5 mm, the externalcation site of the Na+,K+-ATPase is stimulated, leading to a lowering of [Nae]1. Increases in [K+]O of only1 mm are sufficient to produce a decrease in [Nae]1. Increases in [K+]O may stimulate the Na+-K+-Cl-cotransport which could then act to replenish the Na+ removed by the Na+ pump. The Na+-HCO3-cotransporter would also participate in the resupply of Nat+ because this cotransporter is stimulated bymembrane depolarization caused by the increase in [K+]O The complex pattern of the change in [Nae]1seen with small increases in [K+]O is probably a consequence of the dynamic interplay between these threetransport mechanisms (see text).
J Physiol.491.2 303
) by guest on July 13, 2011jp.physoc.orgDownloaded from J Physiol (

C. R. Rose and B. R. Ransom
Because glial cells do not experience Na+ influx with actionpotentials or synaptic potentials, it has been debated howNae enters these cells to supply the Na+,K+-ATPase. Ourexperiments indicated that influx of Nae through voltage-gated Nae channels only contributed to baseline [Nae]i ina minority of cultured hippocampal astrocytes. TheNa+-K+-2Cl- cotransporter, however, provided a constantNae influx under resting conditions. The Nae-HCO3cotransporter normally acts as a pH-regulating transportsystem, moving Nae and HC03- into astrocytes, and wouldalso, therefore, provide intracellular Na+ for the Na+,K+-ATPase (Fig. 13A). Both the Na+-K+-2C- cotransportand Nae-HCO3- cotransport are activated by increases in[K+]o and would be able to accommodate the increaseddemand for intracellular Na+ as Na+,K+-ATPase activityincreased.
According to this scheme, baseline [Nae]i is a value set bythe dynamic equilibrium of at least three transportmechanisms: Na+,K+-ATPase, Nae-HCO3- cotransport,and Na+-K+-2C- cotransport (Fig. 13A). It is likely thatNa+-H+ exchange and perhaps other Na+-dependent carrierssuch as the amino acid uptake systems, also participate indetermining [Nae]1. This formulation suggests that essentialmembrane transport processes interact in a highly com-plementary manner. The idea that astrocyte Na+ influx ismediated by a channel that simply degrades the steep Naegradient without any immediate benefit for the cell is lessattractive, and perhaps is a strategy that is only employedtransiently during development (Sontheimer et al. 1994).
This model of astrocyte [Na+]i homeostasis represents asystem that can react effectively to increases in [K+]o(Fig. 13B). As [K+]o increases and activates the externalcation site on the Na+,K+-ATPase, causing greater enzymeactivity, [Nae]i would fall and could eventually limit K+removal by this mechanism. The increased [K+]0, however,would also increase Nae influx by Na+-K+-2CF-cotransport (Tas et at. 1987) and Nae-HCO3- cotransport(Pappas & Ransom, 1994), keeping pace in a dynamic waywith the astrocyte's enhanced need for intracellular Nae.
BALLANYI, K., GRAFE, P. & TEN BRUGGENCATE, G. (1987). Ionactivities and potassium uptake mechanisms of glial cells in guinea-pig olfactory cortex slices. Journal of Physiology 382, 159-174.
BALLANYI, K. & KETTENMANN, H. (1990). Intracellular Na activityin cultured mouse oligodendrocytes. Journal of NeuroscienceResearch 26, 455-460.
BORZAK, S., KELLY, R. A., KRAMER, B. K., MATOBA, Y., MARSH,J. D. & REERS, M. (1990). In situ calibration of fura-2 and BCECFfluorescence in adult rat ventricular myocytes. American Journal ofPhysiology 259, H973-981.
BOYARSKI, G., RANSOM, B. R., SCHLUE, W.-R., DAvIs, M. B. E. &BORON, W. F. (1993). Intracellular pH regulation in single culturedastrocytes from rat forebrain. Glia 8, 241-248.
BUEHRLE, C. P. & SONNHOF, U. (1983). Intracellular ion activities andequilibrium potential in motoneurons and glial cells of the frogspinal cord. Pfluigers Archiv 396, 144-153.
COLES, J. A. & ORKAND, R. K. (1985). Changes in sodium activityduring light stimulation in photoreceptors, glia and extracellularspace in drone retina. Journal of Physiology 362, 415-435.
DEITMER, J. W. (1991). Electrogenic sodium-dependent bicarbonatesecretion by glial cells of the leech central nervous system. Journalof General Physiology 98, 637-655.
DEITMER, J. W. (1992). Bicarbonate-dependent changes of intra-cellular sodium and pH in identified leech glial cells. Pfligers Archiv420, 584-589.
DIETZEL, I., HEINEMANN, U., HOFMEIER, G. & Lux, H. D. (1982).Stimulus-induced changes in extracellular Na+ and Cl-concentration in relation to changes in extracellular space.Experimental Brain Research 46, 73-84.
DOERNER, R., BALLANYI, K. & SCHLUE, W.-R. (1990). Glutaminergicresponses of neuropile glial cells and Retzius neurones in the leechcentral nervous system. Brain Research 523, 111-116.
ERECINSKA, M., DAGANI, F., NELSON, D., DEAS, J. & SILVER, J. A.(1991). Relations between intracellular ions and energy metabolism:A study with monensin in synaptosomes, neurons, and C6 gliomacells. Journal of Neuroscience 11, 2410-2421.
GIULIAN, D. & BAKER, T. J. (1985). Peptides released by ameboidmicroglia regulate astroglia proliferation. Journal of Cell Biology101, 2411-2415.
GRAFE, P. & BALLANYI, K. (1987). Cellular mechanisms of potassiumhomeostasis in the mammalian nervous system. Canadian Journalof Physiology and Pharmacology 65, 1038-1042.
GRAFE, P., RIMPEL, J., REDDY, M. M. & TEN BRUGGENCATE, G. (1982).Changes of intracellular sodium and potassium ion concentrations infrog spinal motoneurons induced by repetitive synaptic stimulation.Neuroscience 7, 3213-3220.
GRISAR, T., FRERE, J. M. & FRANCK, G. (1979). Effects of K+ ions onkinetic properties of the (Na+,K+)-ATPase (EC 3.6.1.3) of bulkisolated glia cells, perikarya and synaptosomes from rabbit braincortex. Brain Research 165, 87-103.
HAROOTUNIAN, A. T., KAO, J. P. Y., ECKERT, B. K. & TsIEN, R. Y.(1989). Fluorescence ratio imaging of cytosolic free Nae inindividual fibroblasts and lymphocytes. Journal of BiologicalChemistry 264, 19458-19467.
HEINEMANN, U. & Lux, H. D. (1977). Stimulus-induced rises inextracellular K+ in cerebral cortex of cat. Brain Research 120,231-249.
KIMELBERG, H. K. & FRANGAKIS, M. V. (1985). Furosemide andbumetanide-sensitive ion transport and volume control in primarycultures from rat brain. Brain Research 361, 125-134.
LASSER-ROSS, N. & Ross, W. N. (1992). Imaging voltage andsynaptically activated sodium transients in cerebellar Purkinje cells.Proceedings of the Royal Society B 247, 35-39.
LEVI, A. J., LEE, C. 0. & BROOKSBY, P. (1994). Properties of thefluorescent sodium indicator 'SBFI' in rat and rabbit cardiacmyocytes. Journal of Cardiovascular Electrophysiology 5, 241-257.
MINTA, A. & TsIEN, R. Y. (1989). Fluorescent indicators for cytosolicsodium. Journal of Biological Chemistry 264, 19449-19457.
MOORE, E. D. W., BECKER, P. L., FOGARTY, K. E., WILLIAMS, D. A. &FAY, F. S. (1990). Ca2+ imaging in single living cells: Theoretical andpractical issues. Cell Calcium 11, 157-179.
MOORE, E. D. W. & FAY, F. S. (1993). Isoproterenol stimulatesextrusion of sodium from isolated smooth muscle cells. Proceedingsof the National Academy of Sciences of the USA 90, 8058-8062.
304 J. Physiol.491.2
) by guest on July 13, 2011jp.physoc.orgDownloaded from J Physiol (

Intracellular sodium in astrocytes
NEGULESCU, P. A., HAROOTUNIAN, A., TsIEN, R. Y. & MACHEN, T. E.(1990). Fluorescence measurements of cytosolic free Naconcentration, influx and efflux in gastric cells. Cell Regulation 1,259-268.
NEWMAN, E. A. (1995). Glial cell regulation of extracellularpotassium. In Neuroglia, ed. KETTENMANN, H. & RANSOM, B. R.,pp. 717-731. Oxford University Press, New York.
O'CONNOR, E. R., SONTHEIMER, H. & RANSOM, B. R. (1994). Rathippocampal astrocytes exhibit electrogenic sodium-bicarbonate co-
transport. Journal of Neurophysiology 72, 2580-2589.PAPPAS, C. A. & RANSOM, B. R. (1993). A depolarization-stimulated,
bafilomycin-inhibitable H+ pump in hippocampal astrocytes. Glia 9,280-291.
PAPPAS, C. A. & RANSOM, B. R. (1994). Depolarization-inducedalkalinization (DIA) in rat hippocampal astrocytes. Journal ofNeurophysiology 72, 2816-2826.
RANSOM, B. R. & SONTHEIMER, H. (1992). The neurophysiology ofglial cells. Journal of Clinical Neurophysiology 9, 224-251.
ROSE, C. R. & RANSOM, B. R. (1995). Fluorescence imaging of intra-cellular sodium in rat hippocampal astrocytes. Society forNeuroscience Abstracts 21, 783.1.
SONTHEIMER, H., FERNANDEZ-MARQUES, E., ULLRICH, N., PAPPAS,C. A. & WAXMAN, S. G. (1994). Astrocyte Nae channels are requiredfor maintenance of Na`/K+-ATPase activity. Journal ofNeuroscience 14, 2464-2475.
SONTHEIMER, H., RANSOM, B. R., CORNELL-BELL, A. H., BLACK, J. A.& WAXMAN, S. G. (1991). Nae-current expression in rat hippocampalastrocytes in vitro: Alterations during development. Journal ofNeurophysiology 65, 3-19.
SWEADNER, K. J. (1995). Sodium, potassium-adenosine triphosphataseand its isoforms. In Neuroglia, ed. KETTENMANN, H. & RANSOM, B.R., pp. 259-272. Oxford University Press, New York.
TAs, P. W. L., MASSA, P. T., KRESS, H. G. & KOSCHEL, K. (1987).Characterization of an Na/K+/CI- cotransport in primary culturesof rat astrocytes. Biochimica et Biophysica Acta 903, 411-416.
THOMAS, J. A., BUCHSBAUM, R. N., ZIMNIAK, A. & RACKER, E. (1979).Intracellular pH measurements in Ehrlich ascites tumor cellsutilizing spectroscopic probes generated in situ. Biochemistry 18,2210-2218.
WALZ, W. (1989). Role of glial cells in the regulation of the brain ionmicroenvironment. Progress in Neurobiology 33, 309-333.
WALZ, W. & HERTZ, L. (1982). Ouabain-sensitive and ouabain-resistant uptake of potassium into astrocytes and neurons inprimary cultures. Journal of Neurochemistry 39, 70-77.
WALZ, W. & HERTZ, L. (1983). Intracellular ion changes of astrocytesin response to extracellular potassium. Journal of NeuroscienceResearch 10, 411-423.
WALZ, W. & HINKS, E. C. (1986). A transmembrane sodium cycle inastrocytes. Brain Research 368, 226-232.
AcknowledgementsWe thank Dr Kai Kaila for helpful comments on an earlier versionof the manuscript. This study was supported by a fellowship fromthe Deutsche Forschungsgemeinschaft to C.R.R. (Ro 1130/1-1)and by NIH grants (NS-15589 and NS-06208) to B.R.R.
Received 12 June 1995; accepted 26 September 1995.
305J Physiol.491.2
) by guest on July 13, 2011jp.physoc.orgDownloaded from J Physiol (