membrane lipid homeostasis in bacteria
TRANSCRIPT

Bacterial survival depends on membrane lipid home-ostasis and on an ability to adjust lipid composition to acclimatize the bacterial cell to different environ-ments. Bacterial membranes consist of proteins that are embedded in a lipid matrix that closely approximates a phospholipid bilayer. Although there is a considerable diversity of phospholipid structures in the bacterial world, most membrane phospholipids are glycerolipids that contain two fatty acid chains. These phospholipid acyl chains determine the viscosity of the membrane, which in turn influences many crucial membrane-associated functions, such as the passive permeability of hydrophobic molecules, active solute transport and protein–protein interactions. The adjustments in fatty acid composition that maintain the biophysical proper-ties of membranes are referred to as homoviscous adap-tation, and are interpreted as a mechanism that modifies the permeability of the phospholipid bilayer to minimize energy expenditure and optimize growth.
The ability of bacteria to modify their membrane composition in response to environmental changes, such as in temperature, osmolarity, salinity and pH, was determined early in the study of bacterial lipid metabo-lism1. This Review discusses the primary genetic and biochemical processes that underlie these adjustments and are responsible for membrane phospholipid home-ostasis in bacteria. The biophysical properties of the membrane are defined in large part by the fatty acid structures that are incorporated into the component phospholipids. Phospholipid fatty acids are present in various chain lengths of between 14 and 20 carbons, and many contain a single cis double bond, or iso or anteiso methyl branches (TABLE 1). Bacteria have evolved
mechanisms to control the formation of new fatty acids and modify the structure of existing fatty acids, and these allow bacteria to adjust membrane viscosity to match environmental requirements. The de novo fatty acid biosynthetic pathway is a major focal point for the regulatory events that control membrane homeostasis. The control points that regulate total phospholipid content and fatty acid composition are discussed here. However, bacteria are often subjected to abrupt changes in their extracellular environment that neces-sitate the modification of existing fatty acids to rapidly adapt to the new conditions, and this Review will also cover these homeostatic mechanisms. These membrane alterations are important not only for free-living bacte-ria, but also for pathogens that must also adapt to the host environment. For example, many pathogens alter their membrane phospholipid structures to increase their resistance to antimicrobial peptides that are pro-duced by the innate immune system. Phospholipids are also used as intermediates in the formation of other important membrane and cell-wall constituents, and the lipid by-products of these pathways destabilize the membrane if they are allowed to accumulate.
Here, a phospholipid is defined as a lipid that is derived from the acylation of sn-glycerol-3-phosphate (glycerol-3-P), and the focus is the plasma membrane, as opposed to cell-wall or outer-membrane lipopoly-saccharides. Therefore, diglucosyl-diacylglycerols are grouped with phospholipids in this Review, because they are derived from glycerol-3-P, whereas the lipid A constituents of the outer membrane of Gram-negative organisms and the lipoteichoic acids that contribute to the cell wall of Gram-positive bacteria are not. Readers
Department of Infectious Diseases, St. Jude Children’s Research Hospital, 332 N. Lauderdale, Memphis, Tennessee 38105, USA. e-mails: [email protected]; [email protected]:10.1038/nrmicro1839Published online 11 February 2008
Membrane lipid homeostasis in bacteriaYong-Mei Zhang and Charles O. Rock
Abstract | The ability of bacteria to control the biophysical properties of their membrane phospholipids allows them to thrive in a wide range of physical environments. Bacteria precisely adjust their membrane lipid composition by modifying the types of fatty acids that are produced by the biosynthetic pathway and altering the structures of pre-existing phospholipids. The recycling of phospholipids that are used as intermediates in the biosynthesis of other major membrane components is also crucial to bilayer stability in dividing cells. Here, the principal genetic and biochemical processes that are responsible for membrane lipid homeostasis in bacteria are reviewed.
R E V I E W S
222 | MARCH 2008 | VOLUME 6 www.nature.com/reviews/micro
© 2008 Nature Publishing Group
Type II fatty acid biosynthetic pathwayThe pathway by which fatty acids are synthesized in bacteria, chloroplasts and mitochondria. Distinct enzymes, each of which are encoded by an individual gene, catalyse the different steps of the pathway.
are referred to recent reviews on lipid A2 and lipotei-choic acid3 modifications in response to environmental factors. Here, we will discuss the bacterial fatty acid biosynthetic machinery and its role in controlling the biophysical properties of the membrane, the modifica-tion of existing membrane phospholipids and how cells deal with phospholipid turnover.
The fatty acid biosynthetic machineryFatty acid synthesis in bacteria is achieved by a highly conserved set of genes in which each gene encodes an individual step in the type II fatty acid biosynthetic pathway (FIG. 1). All of the proteins in this pathway are located in the cytosol, and each has been purified and biochemically characterized. Furthermore, the struc-ture and catalytic mechanism of each enzyme is known in considerable detail4. Similar dissociated fatty acid synthase II (FASII) pathways are found in chloroplasts, mitochondria and apicoplasts.
The acyl carrier protein as a carrier. The central player in the FASII pathway is the acyl carrier protein (ACP), a low-molecular-mass protein that carries all of the FASII- pathway intermediates. The fatty acid chains are attached as thioesters to the terminus of the 4 -phosphopantetheine prosthetic group of ACP, which is covalently attached to a serine that is located on an extended loop between the first and second helices of the 4-helix bundle structure of ACP5,6. An acyl chain is accommodated by a hydrophobic binding cavity that is formed by the interior surfaces of the four helices of the ACP and varies in size depending
on the nature of the acyl chain7. Protein–ACP interac-tions occur between the conserved electronegative and hydrophobic structure of helix-2 of the ACP, also known as the recognition helix, and a corresponding electroposi-tive and hydrophobic docking surface that is adjacent to the entrance to the active site on the target proteins8. The conserved nature of the protein–ACP interface enables ACPs from diverse organisms to be used interchange-ably in the Escherichia coli FASII system8. Protein–protein interactions in the FASII system have considerable plas-ticity, because mutations that compromise protein–ACP interactions in vitro do not always lead to deficiencies in fatty acid synthesis in vivo9.
Initiation of fatty acid biosynthesis. The initiation phase of fatty acid synthesis is shown in FIG. 1a. The first step in fatty acid synthesis is the conversion of acetyl-CoA to mal-onyl-CoA by acetyl-CoA carboxylase (ACC). ACC is con-structed from four separate proteins and requires biotin as a covalently attached cofactor10. Next, the malonyl group is transferred to ACP by the malonyl transacylase FabD. FabH — a condensing enzyme that initiates the formation of a new acyl chain11 — is primed by acyl-CoA, and the acyl-enzyme intermediate is condensed with malonyl-ACP to form the first -ketoacyl-ACP intermediate in the FASII pathway.
The FASII pathway shown in FIG. 1 shows the for-mation of straight-chain fatty acids, but there can be a major variation on this theme. Branched-chain fatty acids are produced in most Gram-positive bacteria, and exist in two forms that are characterized by the
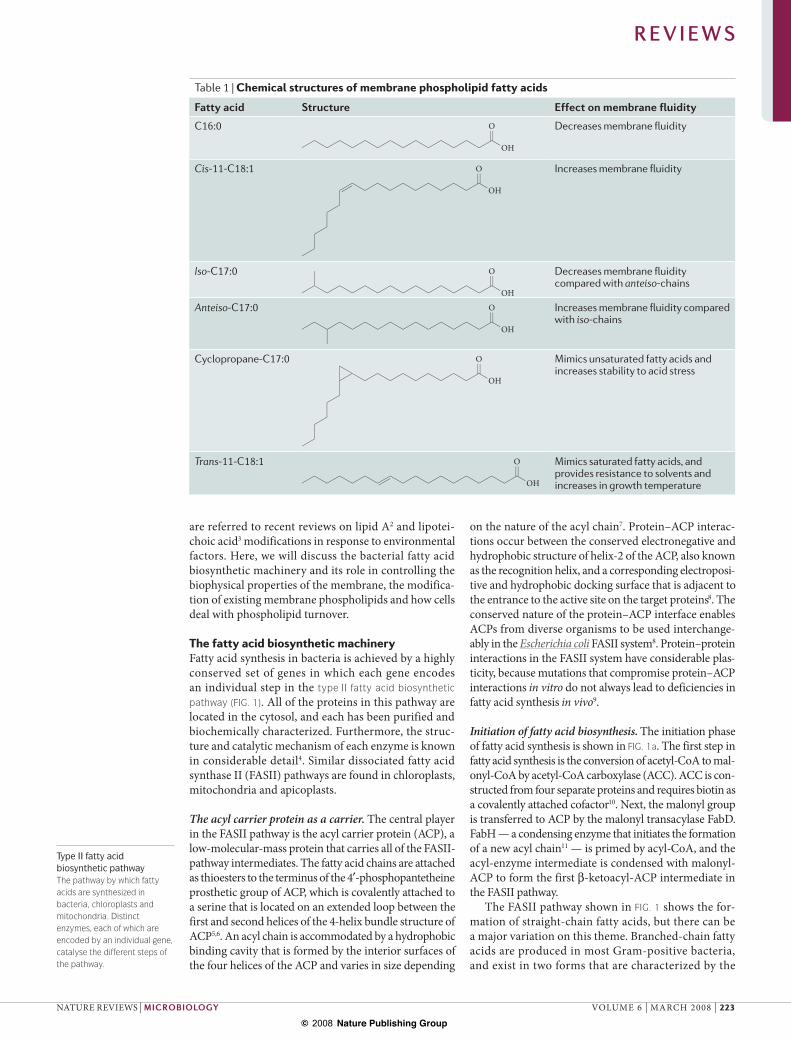
Table 1 | Chemical structures of membrane phospholipid fatty acids
Fatty acid Structure Effect on membrane fluidity
C16:0
OH
O Decreases membrane fluidity
Cis-11-C18:1
OH
O Increases membrane fluidity
Iso-C17:0
OH
O Decreases membrane fluidity compared with anteiso-chains
Anteiso-C17:0
OH
O Increases membrane fluidity compared with iso-chains
Cyclopropane-C17:0
OH
O Mimics unsaturated fatty acids and increases stability to acid stress
Trans-11-C18:1
OH
O Mimics saturated fatty acids, and provides resistance to solvents and increases in growth temperature
R E V I E W S
NATURE REVIEWS | MICROBIOLOGY VOLUME 6 | MARCH 2008 | 223
© 2008 Nature Publishing Group

Nature Reviews | Microbiology
SACP
O
HO
O
O– SCoA
O
–O SCoA
OO
–O SACP
OOR S
CoAO
R SACP
OO
R SACP
O
R SACP
O
R SACP
OOH
+
ATP
ACP
Malonyl-ACP
H2O
NADPH NADP+
ADPACC
FabD
FabH
NADHNAD+
FabG
FabZ or FabA
FabI or FabK
FabF or FabB
PlsX
PlsYPlsC
PlsC
PlsBPALPA
LPA
P
PALPALPA
a
c
b
Cytoplasm
Lysophosphatidic acid1-acyl-sn-glycero-3-phosphate; the first intermediate in membrane phospholipid formation.
Phosphatidic acid1,2-diacyl-sn-glycero-3-phosphate; the key intermediate in the formation of most bacterial membrane phospholipids.
presence of either iso or anteiso branches at their ter-mini (TABLE 1). The branched chains are introduced into the pathway in the initiation phase (FIG. 1a) by FabH enzymes of the appropriate substrate specifi-city. FabH of E. coli, which produces straight-chain fatty acids, is selective for acetyl-CoA, whereas the FabH enzymes of Gram-positive bacteria prefer larger branched-chain substrates12,13. Gram-positive bacte-ria also have a branched-chain ketoacid dehydro-genase that produces the branched-chain acyl-CoA precursors from leucine, valine and isoleucine. The
FabH from Mycobacterium tuberculosis has the largest acyl-chain-binding pocket, and can accommodate acyl-CoAs of 14 carbons and longer14,15.
Chain elongation. Four enzymatic reactions consti-tute the core elongation machine, which advances the growing acyl chain by two carbons (FIG. 1b). Each new round is initiated by an elongation condensing enzyme: FabF or FabB. The FabF isoform is universally expressed in bacteria, but some bacteria also express the FabB isoform, which has a specific role in unsatu-rated fatty acid (UFA) formation (discussed below). FabG is the -ketoacyl-ACP reductase, and only a single isoform of this enzyme has been identified so far in bacteria. The next step in the elongation cycle is the dehydration of the -hydroxyacyl-ACP to trans-2-enoyl-ACP. Two isoforms, FabA and FabZ, catalyse the dehydration of -hydroxyacyl-ACPs, albeit with different substrate specificities16. FabZ is universally expressed in bacteria, whereas FabA is expressed only in organisms that use the FabA–FabB system to introduce double bonds into the growing acyl chain (discussed below). Importantly, dehydratases are readily reversible, and cycles of elongation must be pulled to completion by an enoyl-ACP reductase (FabI)17. FabI is a widely expressed reductase, but some bacteria use alternative enoyl-ACP reductases — the flavoprotein FabK18,19 or FabV20 — to complete cycles of elongation.
Transfer to the membrane. The first step in the forma-tion of a membrane phospholipid is the acylation of glycerol-3-P (FIG. 1c). The most ubiquitous system is the PlsX–PlsY pathway, which is found in all but one family of proteobacteria21. PlsX, a peripheral mem-brane protein22, transfers the acyl group from the long-chain acyl-ACP end products of the elongation pathway to inorganic phosphate to form a reactive acylphosphate intermediate. This activated fatty acid is then used by PlsY, an integral plasma-membrane protein23, to acylate the 1-position of glycerol-3-P, thereby forming lysophosphatidic acid (LPA). A second route to the formation of LPA is catalysed by PlsB, an inner-membrane acyltranferase that uses either acyl-ACP or acyl-CoA as the acyl donor24. Following acyla-tion by PlsX–PlsY or PlsB, the integral membrane protein PlsC21,25, which is expressed by all bacteria, acylates the 2-position of LPA to form phosphatidic acid (PA), the central intermediate in the formation of membrane glycerolipids.
Phospholipid head-group diversity. There are a huge range of different types of polar head groups that can be attached to the basic PA core structure in bacteria. Although the E. coli phospholipid composition is not typical, E. coli does produce three of the most widely distributed phospholipids in bacteria, and its biosyn-thetic pathways have been studied in detail (FIG. 2a). Another major phospholipid in bacteria is cardiolipin, which is formed by the condensation of two phosphati-dylglycerol (PtdGro) molecules. A common variation
Figure 1 | Conserved pathway for the formation of phosphatidic acid in bacteria. a | The initiation phase of type II fatty acid synthesis. Acetyl-CoA carboxylase (ACC) is composed of four subunits and provides malonyl-CoA elongation units to the elongation cycle. The malonyl groups are transferred to the acyl carrier protein (ACP) by malonyl-CoA–ACP transacylase (FadD). -ketoacyl-ACP synthase III (FabH) is the initiating condensing enzyme that is primed by an acyl-CoA to condense with malonyl-ACP to start a new fatty acid. b | The elongation of fatty acid chains is an iterative process that involves 2-carbon building blocks. The -ketoacyl-ACP intermediate is reduced by the FabG -ketoreductase, and the -hydroxyacyl-ACP is dehydrated to enoyl-ACP by FabA or FabZ. The cycle is pulled to completion by the enoyl-ACP reductase (FabI or FabK). New rounds of elongation are initiated by the elongation condensing enzymes. FabF is a universally expressed isoform, but some bacteria also express FabB, a condensing enzyme that has a specific function in unsaturated fatty acid formation. c | Transfer of fatty acids to glycerol-3-phosphate. The long-chain acyl-ACP end products are recognized as substrates by the acyltransferases. The peripheral membrane protein PlsX catalyses the formation of an acylphosphate (Acyl-P) from acyl-ACP, and PlsY uses this unique intermediate to acylate glycerol-3-phosphate to 1-acyl-glycerol-3-phosphate (also known as lysophosphatidic acid (LPA)). PlsB acylates glycerol-3-phosphate using either acyl-ACP or acyl-CoA as the acyl donor. PlsC transfers a fatty acid to the 2-position of LPA to form phosphatidic acid (PA), the key intermediate in the synthesis of all membrane glycerolipids.
R E V I E W S
224 | MARCH 2008 | VOLUME 6 www.nature.com/reviews/micro
© 2008 Nature Publishing Group

Nature Reviews | Microbiology
CL
PtdGro
PtdGro-P
PtdEtn
PA
CdsA
CDP-DAG
PtdSer
O O R1
O
O
O
R2
PHO O
–O
OP
O
O
O–
N
NH2
ON
O
OH OH
O O R1
O
O
O
R2
PO
O–
O O R1
O
O
O
R2
PO O
–O
–O
O
NH3+
O O R1
O
O
O
R2
PO O
–O
+H3N
O O R1
O
O
O
R2
PO O
–O
OP
OHHO O
–O
O O R1
O
O
O
R2
PO O
–O
HOOH
O O R1
O
O
O
R2
P–O O
O
O O R3
O
O
O
R4
PO O
–O
HO
PgsAGro-3-P
PssAL-Ser
CTP
ClsPtdGro
Gro
PgpPPi
PsdCO2
a
b
PtdEtnPtdEtn PtdGroPtdGroPsd
PssA
PssA
PgsA PgpP
Cytoplasm
–– – – – –
CDP-DAGCDP-DAG
CDP
on the basic phospholipid biosynthetic theme is the formation of glycolipids by the reaction of a urid-ine diphosphate sugar with diacylglycerol (DAG)26. Glucosyl- and diglycosyl-diacylglycerols are abundant lipids in many Gram-positive bacteria, and these lipids function as membrane anchors for the lipoteichoic acid constituents of the cell wall3. In addition, ornithine-derived lipids are produced in phosphate-limiting envi-ronments so that available phosphate can be diverted to macromolecular synthesis27,28. Ornithine lipids are also crucial structural components of the membranes of photosynthetic bacteria29.
Controlling membrane homeostasisControl at the level of fatty acid biosynthesis is crucial for membrane homeostasis, because the biophysical properties of membranes are determined in large part by the composition of the fatty acids that are produced by de novo biosynthesis. There is no noteworthy alternative fate for fatty acids in most bacteria, and all of the long-chain acyl-ACPs that are produced by the biosynthetic pathway are incorporated into the membrane. Almost all of the metabolic energy that is used to produce membrane lipids is expended in the formation of fatty acids, and therefore their production must be precisely controlled to support membrane biogenesis and prevent the wasteful expenditure of ATP. Although the biophysi-cal properties of membranes can be changed by altering the ratio of the polar head groups in membrane phos-pholipids, bacteria seem to use biochemical and genetic mechanisms to modify the composition of the fatty acids that are produced by FASII.
Control of total membrane lipid content. The long-chain acyl-ACP end products of fatty acid biosynthesis have emerged as the key regulators of the biosynthetic pathway (FIG. 1). The importance of long-chain acyl-ACP regulation was determined through the demonstra-tion that acyl-ACP hydrolysis by thioesterases leads to deregulated fatty acid biosynthesis and to the release of fatty acids into the medium30–32. There are three enzymes that are regulated by acyl-ACP: ACC, FabH and FabI. The ACC reaction (FIG. 1a) is inhibited by long-chain acyl-ACP33, and lower ACC activity limits the supply of malonate groups for chain initiation and elongation. The first step in fatty acid elongation, which is catalysed by FabH (FIG. 1a), is potently inhibited by mature acyl-ACP chain lengths34,35. Regulation of FabH prevents the initiation of new acyl chains and limits the total number of fatty acids that are produced35. FabI, which produces acyl-ACP (FIG. 1b), is inhibited by acyl-ACP34 through product inhibition. Because FabI has a determinant role in the completion of rounds of fatty acid elongation17, a reduction in FabI activity slows the rate of fatty acid elongation.
Cellular acyl-ACP levels are controlled by regulation at the glycerol-3-P acyltransferase step (FIG. 1c); obstruct-ing PlsB activity using conditional mutants leads to the accumulation of long-chain acyl-ACP and the cessation of fatty acid biosynthesis36. A physiological regulator of PlsB activity is guanosine 3 ,5 -bispyrophosphate (ppGpp),
Figure 2 | Diversification of polar head groups. The enzymes in phospholipid synthesis are either intrinsic membrane proteins or are associated with the bilayer. a | A key intermediate in phospholipid synthesis is cytosine diphosphate (CDP)-diacylglycerol (CDP-DAG), which is formed by CdsA from phosphatidic acid and cytosine triphosphate (CTP). Phosphatidylethanolamine (PtdEtn), a major phospholipid in many bacteria, is produced from CDP-DAG in two steps. Serine is exchanged with cytosine monophosphate (CMP) by phosphatidylserine (PtdSer) synthase (PssA), and the resulting PtdSer is rapidly decarboxylated by PtdSer decarboxylase (Psd) to generate PtdEtn. Phosphatidylglycerol (PtdGro) is also synthesized from CDP-DAG in two steps. The first step exchanges glycerol-3-phosphate (Gro-3-P) for CMP by PtdGro-phosphate synthase (PgsA); this is followed by the rapid dephosphorylation of PtdGro-phosphate by phosphatidylglycerol phosphate phosphatase (PgpP), which is encoded by either the pgpA or pgpB genes. Cardiolipin is synthesized through the condensation of two PtdGro molecules by cardiolipin synthase (Cls). b | In the biochemical regulation of phospholipid composition, the zwitterionic (PtdEtn) and acidic (PtdGro and cardiolipin) branches of phospholipid synthesis compete for a common pool of CDP-DAG. PtdGro is constitutively produced by the action of PgsA and PgpP. PtdEtn formation is regulated by the mole fraction of PtdGro and cardiolipin in the membrane, which controls the association and activation of PssA.
R E V I E W S
NATURE REVIEWS | MICROBIOLOGY VOLUME 6 | MARCH 2008 | 225
© 2008 Nature Publishing Group

ZwitterionicA compound that is electrically neutral, but that carries formal positive and negative charges on different atoms.
a global regulator of gene expression and stable RNA synthesis in bacteria37. Elevating ppGpp levels in vivo inhibits PlsB activity, which, in turn, triggers an increase in acyl-ACP and a reduction in fatty acid synthesis36. Conversely, the inhibition of fatty acid elongation using the antibiotic cerulenin stimulates the accumulation of ppGpp38. Recent evidence suggests that an interaction between ACP and SpoT (also known as (p)ppGpp syn-thase II) couples fatty acid synthesis to ppGpp levels39. In Gram-positive bacteria, the inactivation of PlsX leads to the cessation of fatty acid synthesis22, which indicates that acyl-ACP also functions as a feedback regulator in these organisms. However, the mechanism of PlsX inhibition is unknown. Gram-positive bacteria also produce ppGpp in response to protein-synthesis arrest, but the role of this ppGpp production in the regulation of lipid metabolism is not clear. Many Gram-positive bacteria have a global transcriptional regulator, called FapR40, that is absent in Gram-negative bacteria. FapR coordinates the expression of all the genes that function in fatty acid and phospholipid synthesis, and malonyl-CoA is the ligand that controls FapR binding to DNA41.
Control of membrane phospholipid composition. The proportion of phospholipid polar head groups in the membrane is tightly controlled to balance the compo-sition of zwitterionic or neutral phospholipids (such as phosphatidylethanolamine (PtdEtn) and glucosyldia-cylglycerol) and acidic phospholipids (such as PtdGro and cardiolipin). The importance of maintaining this balance in bacterial membranes is clearly illustrated by the phenotypes of mutants that are deficient in various enzymes (FIG. 2a). Phospholipids are determinants of membrane-protein topology, and the failure of mem-brane proteins to correctly fold in mutants that have altered phospholipid compositions gives rise to defects in solute transport, electron transport, the initiation of DNA replication and cell division42–44. Phospholipid compositional homeostasis is achieved by the bio-chemical regulation of enzymes45,46 (FIG. 2b). The artifi-cial amplification of either phosphatidylserine (PtdSer) synthase (PssA)47 or PtdSer decarboxylase (Psd)48 has no substantive effect on the phospholipid composition of E. coli. PssA is an extrinsic membrane protein that associates with, and is activated by, bilayers that contain the anionic phospholipids PtdGro and cardiolipin49,50. Thus, as the proportion of PtdEtn increases, the mem-brane association and activity of PssA decreases, thereby slowing the rate of PtdEtn formation. Support for this model comes from experiments in which Bacillus subtilis PssA, a molecularly distinct integral membrane protein, was used to replace the transiently membrane-associated PssA in E. coli. Amplification of B. subtilis PssA increases the relative and absolute amounts of PtdEtn and impairs growth51,52. Therefore, the balance of zwitterionic and acidic phospholipids in E. coli arises from the continual formation of PtdGro by the integral membrane PtdGro-phosphate synthase–phosphatase (PgsA–PgpA/B) sys-tem, which is coupled with the tight regulation of PtdEtn synthesis through the PssA–Psd pathway (FIG. 2b).
Cardiolipin is a less abundant acidic membrane phos-pholipid than PtdGro, and it is synthesized by a reversible phosphatidyl group transfer from PtdGro (FIG. 2a). Cells that are deficient in cardiolipin have longer doubling times, are less viable in the stationary phase and have altered antibiotic sensitivity and transport functions53,54. Cardiolipin is an important determinant of antibiotic sensitivity, and the cls gene, which encodes cardiolipin synthase, was first recognized as a locus that increases the sensitivity of cells to novobiocin55. Alterations in car-diolipin content represent an important adaptive change in response to pH, salinity and organic solvents45,56. Cardiolipin content increases significantly as the salinity of the growth medium increases — cls mutants cannot grow in a medium that has a sodium chloride concentra-tion of more than 1 molar57. Regulation of cardiolipin content is determined, in part, by the induction of cls expression in high salt environments57 or as the cells enter the stationary phase58; however, cardiolipin synthase is also biochemically regulated by product inhibition59.
Regulation of membrane biophysical properties. The most intensively investigated and unifying theme in membrane phospholipid homeostasis research is adjustments in the structure of the fatty acyl chains that are attached to glycerol backbones. The structures of these fatty acids determine the biophysical proper-ties of the membrane bilayer, and the biological relevance of regulating fatty acid structure was recognized early in the investigation of bacterial lipid metabolism1. A simplified way to think about the problem is to con-sider the shapes of the fatty acid chains. The straight-chain saturated fatty acids (SFAs), such as palmitic acid (16:0), are linear and pack together efficiently to produce a bilayer that has a high phase transition and low permeability properties (TABLE 1). The cis-UFA introduces a pronounced kink in the chain (TABLE 1), which disrupts the order of the bilayer and results in lower transition temperatures and higher per-meability. The composition of the branched-chain fatty acids affects membrane fluidity owing to the disruptive effect of the methyl group on acyl-chain packing. The anteiso fatty acids promote a more fluid membrane structure than the iso fatty acids, because the methyl branch is further from the end of the fatty acid. Bacteria that produce these fatty acids modify their iso:anteiso ratio in response to temperature and pH stress. These membrane alterations have been well studied in Listeria monocytogenes, a pathogen that can survive at low temperatures60,61.
Bacteria that can introduce double bonds into growing acyl chains can adjust membrane fluidity by regulating the production of cis-UFAs1,24. The proportion of cis-UFA is determined by competition for intermediates at a spe-cific branch point in the pathway which, in E. coli, occurs at the FabA step (FIG. 1b). FabA defines a distinct protein family of dehydratases that converts the -hydroxy to the trans-2-decenoyl-ACP intermediate and functions as an isomerase that establishes a rapid equilibrium between trans-2- and cis-3-decenoyl-ACP at the 10-carbon stage. The SFA:UFA ratio depends on the ratio between the use
R E V I E W S
226 | MARCH 2008 | VOLUME 6 www.nature.com/reviews/micro
© 2008 Nature Publishing Group

of cis-3-decenoyl-ACP by FabB and the use of trans-2-decenoyl-ACP by FabI. The introduction of the double bond at the 10-carbon intermediate and the subsequent elongation steps give rise to the two major UFAs that are found in bacteria: 9-16:1 and 11-18:1 (also known as cis-vaccenate). The FabB condensing enzyme has the unique catalytic property of elongating cis-3-decenoyl-ACP, whereas FabF does not. Thus, both FabA and FabB are required for UFA synthesis.
In E. coli, the fabA and fabB genes are regulated by the transcription factors FadR and FabR. FadR, a bifunctional transcription factor, coordinately regulates fatty acid synthesis and oxidation62. FadR binds at a site downstream of the fad gene promoters and represses transcription of the entire regulon. Exogenous fatty acids are converted to CoA thioesters, which bind to FadR and thereby induce a conformational change that releases FadR from DNA63. The FadR cognate sequence also exists in the –40 regions of the fabA and fabB fatty acid biosynthetic genes64,65, and FadR functions as an activator of the fabA and fabB genes. FabR is a repressor that binds downstream of the FadR sites on the fabA and fabB promoters66. Thus, these two key biosynthetic genes are controlled by a combinatorial lock that involves a transcriptional activator and a repressor. The ligand that controls the binding of FabR to DNA is unknown.
A variation on the FabA–FabB theme for UFA synthe-sis was discovered in Enterococcus faecalis. This organism possesses a special variant of FabZ (called FabN) that has FabA-like activity and a unique FabF (called FabO) that replaces FabB67. The catalytic acid is always an aspartate in FabA and a glutamate in FabZ; however, swapping these residues does not interconvert their activities. The domain swapping between FabZ and FabN sug-gests that the dehydratases can catalyse the trans-2- and cis-3-decenoyl-ACP isomerization reactions because of the shape of the active-site tunnels, which permits the acyl chain to kink at the 3-position to accommodate the cis double bond68. Another variation of the FabA–FabB theme occurs in Streptococcus pneumoniae, which uses a different branch point in fatty acid biosynthesis to achieve UFA formation. In S. pneumoniae, the competi-tive factors are two unique enzymes — FabK competes with the cis–trans isomerase FabM. Again, the introduc-tion of a double bond occurs primarily at the 10-carbon intermediate, which suggests that FabM, like FabA, is a molecular ruler. The proportion of UFA is controlled by FabT, a transcriptional repressor that regulates the expression of fabK, but not fabM69. FabT belongs to the MarR family of transcriptional regulators, but the ligand that regulates its DNA-binding properties is unknown.
There is a biochemical mechanism that allows the elongation cycle to respond to the environmental temperature. This mechanism was carefully studied in E. coli, and FabF was identified as a key player in the alteration of membrane composition in response to temperature change70. At low temperatures, E. coli FabF promotes the production of 11-18:1, whereas at higher temperatures, FabF is inactivated and the formation of
11-18:1 is reduced71.
Modifying existing phospholipid componentsDe novo biosynthesis is finely tuned by environmental signals to produce acyl chains with the properties that are required to optimize membrane function. However, bacteria are often subjected to abrupt changes, for exam-ple in temperature and organic-solvent concentration, that necessitate the immediate modification of existing membrane phospholipid acyl chains to optimize growth under the new conditions. This section describes three biochemical processes that alter the structure of the fatty acids and the biophysical properties of membranes (FIG. 3). Although the enzyme systems that are involved in these modifications are restricted to specific cellular compartments that provide access to only one aspect of the bilayer, virtually all of the phospholipids are modi-fied, which shows that phospholipid flipping across the bilayer and movement between membranes is rapid. The study of these pathways using gene knockouts is possible because these pathways are not essential for growth in the laboratory.
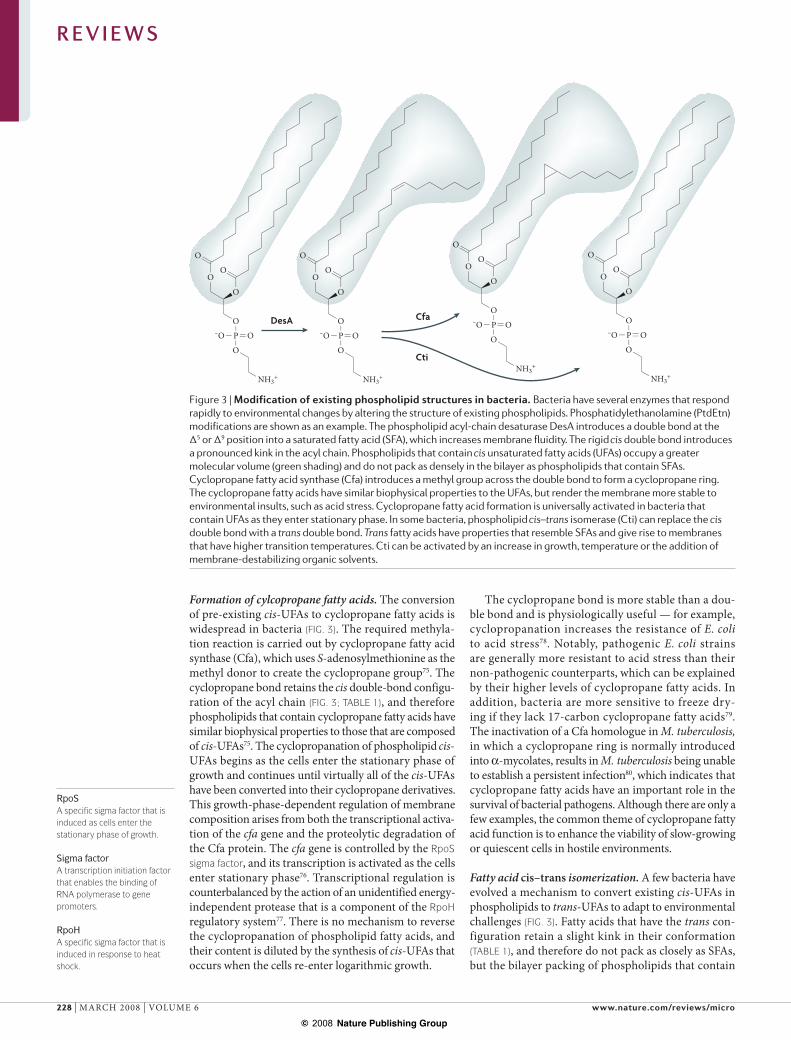
Phospholipid acyl desaturases. Desaturases introduce a cis double bond into pre-existing fatty acids (FIG. 3). An interesting system that has been uncovered in B. subtilis is the regulation of membrane fluidity by a molecular thermosensor72. Specifically, expression of a membrane-bound, oxygen-dependent phos-pholipid acyl-chain desaturase (DesA; also known as delta-5-desaturase) introduces a cis double bond at the 5-position of acyl chains that are attached to existing phospholipids. Transcription of the desA gene is controlled by a two-component regulator that senses changes in membrane properties which are due to abrupt temperature change. It will be interesting to determine exactly how the membrane component of this transcriptional regulatory system senses membrane biophysical properties and transmits the signal to the transcriptional apparatus.
Pseudomonas species have two fatty acyl desaturase systems that play a part in membrane homeostasis. The first system is similar to the one in Bacillus spp. (FIG. 3) and consists of the membrane phospholipid desaturase DesA. However, Pseudomonas spp. DesA introduces the double bond at the 9-position, and the regulator that controls desA expression is unknown. DesA permits the growth of P. aeruginosa in the absence of cis-UFA pro-duction by FASII in fabA mutants73. The second system, DesB, is responsible for introducing double bonds at the 9-position of acyl-CoAs that are derived from exogenous fatty acids73. This desaturase system is inducible and is regulated by DesT, a transcriptional regulator that has the unusual property of being able to sense the fatty acid composition of the acyl-CoA pool to adjust the expres-sion of the desaturase. DesT binds tightly to the desCB promoter to repress transcription in the presence of unsaturated acyl-CoA, but is released from the promoter to induce the synthesis of DesB if bound to a saturated acyl-CoA74. The DesB system provides a mechanism that allows cells to substitute environmental fatty acids for energy-intensive synthesized fatty acids and at the same time maintain membrane homeostasis.
R E V I E W S
NATURE REVIEWS | MICROBIOLOGY VOLUME 6 | MARCH 2008 | 227
© 2008 Nature Publishing Group
Nature Reviews | Microbiology
O
O
O
O
OPO
–O O
NH3+
O
O
O
O
OPO
–O O
NH3+
O
O
O
O
OPO
–O O
NH3+
O
O
O
O
OPO
–O O
NH3+
DesA Cfa
Cti
RpoSA specific sigma factor that is induced as cells enter the stationary phase of growth.
Sigma factorA transcription initiation factor that enables the binding of RNA polymerase to gene promoters.
RpoHA specific sigma factor that is induced in response to heat shock.
Formation of cylcopropane fatty acids. The conversion of pre-existing cis-UFAs to cyclopropane fatty acids is widespread in bacteria (FIG. 3). The required methyla-tion reaction is carried out by cyclopropane fatty acid synthase (Cfa), which uses S-adenosylmethionine as the methyl donor to create the cyclopropane group75. The cyclopropane bond retains the cis double-bond configu-ration of the acyl chain (FIG. 3; TABLE 1), and therefore phospholipids that contain cyclopropane fatty acids have similar biophysical properties to those that are composed of cis-UFAs75. The cyclopropanation of phospholipid cis-UFAs begins as the cells enter the stationary phase of growth and continues until virtually all of the cis-UFAs have been converted into their cyclopropane derivatives. This growth-phase-dependent regulation of membrane composition arises from both the transcriptional activa-tion of the cfa gene and the proteolytic degradation of the Cfa protein. The cfa gene is controlled by the RpoS sigma factor, and its transcription is activated as the cells enter stationary phase76. Transcriptional regulation is counterbalanced by the action of an unidentified energy-independent protease that is a component of the RpoH regulatory system77. There is no mechanism to reverse the cyclopropanation of phospholipid fatty acids, and their content is diluted by the synthesis of cis-UFAs that occurs when the cells re-enter logarithmic growth.
The cyclopropane bond is more stable than a dou-ble bond and is physiologically useful — for example, cyclopropanation increases the resistance of E. coli to acid stress78. Notably, pathogenic E. coli strains are generally more resistant to acid stress than their non-pathogenic counterparts, which can be explained by their higher levels of cyclopropane fatty acids. In addition, bacteria are more sensitive to freeze dry-ing if they lack 17-carbon cyclopropane fatty acids79. The inactivation of a Cfa homologue in M. tuberculosis, in which a cyclopropane ring is normally introduced into -mycolates, results in M. tuberculosis being unable to establish a persistent infection80, which indicates that cyclopropane fatty acids have an important role in the survival of bacterial pathogens. Although there are only a few examples, the common theme of cyclopropane fatty acid function is to enhance the viability of slow-growing or quiescent cells in hostile environments.
Fatty acid cis–trans isomerization. A few bacteria have evolved a mechanism to convert existing cis-UFAs in phospholipids to trans-UFAs to adapt to environmental challenges (FIG. 3). Fatty acids that have the trans con-figuration retain a slight kink in their conformation (TABLE 1), and therefore do not pack as closely as SFAs, but the bilayer packing of phospholipids that contain
Figure 3 | Modification of existing phospholipid structures in bacteria. Bacteria have several enzymes that respond rapidly to environmental changes by altering the structure of existing phospholipids. Phosphatidylethanolamine (PtdEtn) modifications are shown as an example. The phospholipid acyl-chain desaturase DesA introduces a double bond at the !5 or !9 position into a saturated fatty acid (SFA), which increases membrane fluidity. The rigid cis double bond introduces a pronounced kink in the acyl chain. Phospholipids that contain cis unsaturated fatty acids (UFAs) occupy a greater molecular volume (green shading) and do not pack as densely in the bilayer as phospholipids that contain SFAs. Cyclopropane fatty acid synthase (Cfa) introduces a methyl group across the double bond to form a cyclopropane ring. The cyclopropane fatty acids have similar biophysical properties to the UFAs, but render the membrane more stable to environmental insults, such as acid stress. Cyclopropane fatty acid formation is universally activated in bacteria that contain UFAs as they enter stationary phase. In some bacteria, phospholipid cis–trans isomerase (Cti) can replace the cis double bond with a trans double bond. Trans fatty acids have properties that resemble SFAs and give rise to membranes that have higher transition temperatures. Cti can be activated by an increase in growth, temperature or the addition of membrane-destabilizing organic solvents.
R E V I E W S
228 | MARCH 2008 | VOLUME 6 www.nature.com/reviews/micro
© 2008 Nature Publishing Group

Nature Reviews | Microbiology
a
b
c
PtdEtn
DgkAMDOMdoB
MsbA
Cytoplasm
–
PtdGro DAGPtdGro PAPADAG
ATP ADP
LTALtaS
Cytoplasm
Cytoplasm
–
PtdGro DAGPtdGro PAPADAG
ATP ADP
DgkB
PtdEtn 2-acyl-GPE
2-acyl-GPE
LpIT
Lnt
Lpp Acyl-Lpp
FA + ATPAMP + PPi
PlsC
ACP
Aas
ACP
P
P
DefensinA cationic peptide that is produced by the innate immune system and that kills bacteria by disrupting the phospholipid bilayer.
trans-UFAs mimics the physical properties of phos-pholipids that contain SFAs. The cis–trans isomeri-zation reaction leads to membranes that have higher phase-transition temperatures, increased rigidity and decreased permeability to solutes1. An increase in tem-perature is one environmental change that triggers a significant increase in phospholipid trans-UFAs81,82, but cis–trans isomerization has been most extensively stud-ied in the response of Pseudomonas putida to organic solvents that intercalate into, and destabilize, the mem-brane83. The isomerization reaction is carried out by a periplasmic, haem-containing cis–trans isomerase (Cti)83,84. Cti is constitutively expressed and increases trans-UFA formation in response to solvents or tem-perature change85,86. It is, therefore, unclear how Cti activity is regulated by the properties of the membrane bilayer. As the trans-UFAs cannot be returned to the cis conformation by Cti when the environmental stress is removed, cis fatty acids must be replenished by de novo biosynthesis or an as-yet-unidentified process.
Modification of surface charge. Bacterial membranes have an overall high density of negative charge, a prop-erty that is exploited by the innate immune system to disrupt microbial membranes by using various cationic peptides. The formation of lysine adducts of the acidic membrane phospholipids is an important modifica-tion that protects pathogens from attack by the innate immune system87. These lipids are formed by the reaction of a charged lysyl-tRNA with PtdGro to form lysyl-PtdGro88. This modification decreases the nega-tive charge of the membrane bilayer, which prevents the intercalation of defensins into the bilayer and increases resistance to neutrophil killing and other cationic antimicrobial agents (such as vancomycin)89–92.
Membrane phospholipid turnoverPortions of phospholipids are used as precursors in the biosynthesis of other major structural molecules. The accumulation of phospholipid-degradation products that arises from these biosynthetic pathways would disrupt the architecture of the cell membrane and these products must be either eliminated or recycled into phospholipids to maintain membrane homeostasis. In light of the high energy expenditure that is incurred in the production of phospholipid acyl chains, recycling components is the preferred, and sometimes the only, method of dealing with these membrane perturbants. This section uses well-studied phospholipid-turnover events to illustrate how membrane homeostasis is maintained in phospholipid turnover (FIG. 4).
Phosphatidylglycerol turnover. PtdGro is a metaboli-cally active phospholipid and its glycerol-1-P head group undergoes rapid turnover. In Gram-negative bacteria, the glycerol-1-P of PtdGro is transferred to membrane-derived oligosaccharides (MDOs)93 (FIG. 4a), which function in osmotic homeostasis and use PtdGro as a substrate for their biosynthesis in the periplasmic space94. The resulting product, DAG, is converted to PA for the re-synthesis of PtdGro by
an inner membrane lipid kinase called DgkA95. dgkA mutants are viable under growth conditions that do not osmotically stress the bacteria and when MDO production is minimal; however, dgkA is an essential
Figure 4 | Membrane phospholipid turnover. Phospholipids are intermediates in the biosynthesis of other cellular structures, and the lipid by-products must be recycled to maintain membrane homeostasis. a | In Gram-negative bacteria, MdoB catalyses the decoration of membrane-derived oligosaccharides (MDOs) with glycerol-1-phosphate groups in the periplasmic space. The diacylglycerol (DAG) flips to the inner leaflet of the membrane to access the active site of the integral membrane DAG kinase, DgkA. b | In Gram-positive bacteria, the glycerol-1-phosphate head group of phosphatidylglycerol (PtdGro) is used by LtaS, the enzyme that is responsible for the polymerization of glycerol-1-phosphate chains on lipoteichoic acid (LTA). The resulting DAG spontaneously flips from the outer to the inner leaflet, where it is converted to phosphatidic acid (PA) by a soluble DAG kinase, DgkB. c | 2-acylglycerolphosphoeth-anolamine (2-acyl-GPE) is generated by transfer of the 1-position fatty acid of phosphatidylethanolamine (PtdEtn) to a membrane lipoprotein (Lpp), a process that is catalysed by the phospholipid–protein acyltransferase (Lnt). 2-acyl-GPE is transported to the cytosolic side of the membrane by the lysophospholipid flippase LplT, where it is re-acylated by acyl carrier protein (ACP) synthetase (Aas) using acyl-ACP. Aas either acquires acyl-ACP from the biosynthetic pathway or activates a free fatty acid with ATP–Mg2+ and transfers it to ACP to form acyl-ACP. The ATP-binding cassette (ABC) transporter MsbA is involved in the efflux of lipid A and PtdEtn.
R E V I E W S
NATURE REVIEWS | MICROBIOLOGY VOLUME 6 | MARCH 2008 | 229
© 2008 Nature Publishing Group

FabF or FabB
FabI
FabK
FabGFabZ or
FabA
Nature Reviews | Microbiology
Polyphenols
CeruleninThiolactomycinPlatensimycinPlatencin
TriclosanAminopyridinesCephalochrominIsoniazid
3-decynoyl-NACNAS-91 NAS-21
AtromentinLeucomeloneAG205
UndecaprenolA 55-carbon isoprenoid alcohol that functions as a carrier of sugars in the synthesis of peptidoglycan.
Major facilitator superfamilyA large family of transporters that has varied substrate specificity; members of this family possess 12–14 transmembrane segments.
gene in cells that are growing in osmotically chal-lenging environments96,97. Mutational inactivation of DgkA interrupts the DAG cycle, which leads to the accumulation of DAG98,99. DgkA defines a large family of prokaryotic DAG kinases (Pfam01219). However, not all of the members of this family use DAG as a sub-strate; some phosphorylate other lipid substrates, such as undecaprenol, instead100.
The sn-glycerol-1-P polymer that decorates Gram-positive lipoteichoic acids arises from PtdGro (FIG. 4b). Lipoteichoic acids are polydisperse amphiphiles that contain poly(sn-glycerol-1-P) attached to a glycolipid anchor and that form a continuum of anionic charge that protects the bacterium3. The length of the poly-glycerol-P chain varies from 14 to 33 repeating units, and in the prototypical Gram-positive bacterium B. subtilis the repeating units are attached to diglucosyl-DAG. PtdGro is the source of the glycerol-1-P groups101, which means that the biosynthesis of a single lipotei-choic acid molecule requires, on average, 25 PtdGro molecules. Accordingly, PtdGro turnover is rapid102, and the large amount of DAG that is formed is metabo-lized by DgkB, another DAG kinase100 (FIG. 4b). DgkB is
a soluble lipid kinase that closely resembles mammalian DAG kinases. The dgkB gene is essential, probably owing to the accumulation of DAG that occurs when the kinase is inactivated100. PtdGro turnover to DAG occurs on the outer leaflet of the membrane, and the topological problem of returning DAG to the inner leaflet to access the active sites of either DgkA or DgkB is probably solved by the spontaneous flipping of DAG across the bilayer.
Fatty acid turnover in phosphatidylethanolamine. The fatty acid at the 1-position of PtdEtn is used for the post-translational acylation of lipoproteins, and the resulting membrane-disruptive 2-acylgly-cerolphosphoethanolamine (2-acyl-GPE) must be either recycled or destroyed (FIG. 4c). 2-acyl-GPE is re-acylated by the bifunctional enzyme 2-acyl-GPE acyltransferase and acyl-ACP synthetase (Aas)103. Like DAG, 2-acyl-GPE is generated outside the cell and, therefore, must be flipped inside before it can be recycled; however, 2-acyl-GPE does not rapidly dif-fuse across the membrane. This topological problem is solved by a lysophospholipid flippase (LplT), which translocates 2-acyl-GPE to the cytoplasmic face of the inner membrane104. LplT is an integral membrane protein that comprises 12 predicted membrane-spanning segments and belongs to the major facilitator superfamily of membrane transporters104. The 2-acyl-GPE recy-cling pathway is not essential, because 2-acyl-GPE does not accumulate in aas mutants, which is due to the existence of phospholipases that degrade this membrane-disruptive lysophospholipid detergent105. By contrast, DAG continues to accumulate in the absence of DgkA and DgkB, owing to the absence of suitable degradative enzymes.
PerspectivesThe future promises to be productive for investiga-tors of membrane homeostasis in bacteria. A par-ticularly interesting outstanding question is how do the sensors of membrane viscosity operate at the molecular level? These sensors are either coupled to transcription factors or are enzymes that have activi-ties that are intrinsically regulated by the biophysical properties of the membrane. The functions of minor phospholipids in bacterial membranes remain to be elucidated, and not all membrane phospholipid turnover is associated with the metabolic cycles that are described in this Review. An ever-expanding database of bacterial-genome sequences allows the prediction of lipid metabolic pathways, recognition of organisms that have unique properties and identi-fication and knockout of genes that are proposed to be involved in membrane homeostasis. In addition, application of the emerging technologies in liquid chromatography and mass spectrometry will stream-line the detection and structural characterization of new lipid structures. More importantly, these technologies will facilitate the study of minor mem-brane lipid constituents that may have important roles in bacterial physiology.
Box 1 | Antibacterial agents that target fatty acid biosynthesis
Fatty acid biosynthesis is essential for bacterial growth, and the fatty acid synthase II (FASII) pathway has attracted considerable attention because it presents multiple potential targets for antibiotic development106. The elucidation of FASII enzyme structures has greatly facilitated the structure-based design of new inhibitors and improved existing antibacterial agents4. The principle that is followed is that each FASII enzyme is essential and a potential target. Among the FASII enzymes, the most exploited are the elongation condensing enzymes (FabF and FabB) and enoyl-acyl carrier protein reductase (FabI), all of which catalyse the key regulatory reactions. There are many examples of natural products (see the figure; green), such as thiolactomycin, cerulenin and platensimycin, that have evolved to target the FabF and FabB condensing enzymes. FabI is targeted by the antitubercular drug isoniazid107, the natural product cephalochromin108 and triclosan109,110, an antibacterial agent that is widely used in consumer products. However, a drawback to the development of FabI inhibitors as broad-spectrum antimicrobials is the existence of a non-homologous isozyme, FabK, in clinically important pathogens, such as Streptococcus pneumoniae18. Both natural-product111 and chemical112 (see the figure; grey) FabK inhibitors were recently discovered that provide leads to specific drugs that could treat S. pneumoniae infection. Other steps in the pathway have received limited experimental attention106. There is only a single isoform of FabG in bacteria, which suggests that FabG could be a target for broad-spectrum antibiotics. FabH and acetyl-CoA carboxylase enzymes in the initiation phase of FASII are involved in pathway regulation (FIG. 1) and present reasonable objectives for drug discovery. The rising tide of multidrug-resistant pathogens is energizing the search for new antimicrobials, and FASII inhibitors will probably have their most immediate effect on meticillin-resistant Staphylococcus aureus infections because of the sensitivity of this organism to both FabF and FabI inhibitors.
R E V I E W S
230 | MARCH 2008 | VOLUME 6 www.nature.com/reviews/micro
© 2008 Nature Publishing Group

So far, most of the research on bacterial lipid metabolism has focused on the E. coli model system. However, in light of the diversity of lipid structures and ecological niches that are occupied by bacteria, the E. coli paradigm cannot be considered to be representative of bacteria in general. Importantly, advances in genomics and lipo-domics will facilitate the future biomedical investigation of membrane homeostasis in major human pathogens. Many effec-tive antibiotics function by disrupting the membrane, and lipid synthesis is a validated target for new drugs against the expanding repertoire of bacteria that are resistant to current therapies106 (BOX 1). Also, under-standing membrane homeostasis is crucial for the
construction of ‘designer’ organisms that are engi-neered to either produce complex organic molecules or degrade selected environmental pollutants.
Finally, discoveries in bacterial lipid metabolism will impact other fields. For example, FASII is not confined to bacteria, but also exists in chloroplasts and mito-chondria. Thus, understanding the regulation of fatty acid compositional control is crucial for the genetic engineering of plants that can produce oils with desired physical properties. The role (or roles) of mitochondrial fatty acid synthesis remains to be fully appreciated, but it is clear from genetic experiments in the yeast system that the pathway is essential for the proper functioning of these organelles.
1. Cronan, J. E. Jr & Gelmann, E. P. Physical properties of membrane lipids: biological relevance and regulation. Bacteriol. Rev. 39, 232–256 (1975).
2. Raetz, C. R., Reynolds, C. M., Trent, M. S. & Bishop, R. E. Lipid A modification systems in Gram-negative bacteria. Annu. Rev. Biochem. 76, 295–329 (2007).
3. Neuhaus, F. C. & Baddiley, J. A continuum of anionic charge: structures and functions of D-alanyl-teichoic acids in Gram-positive bacteria. Microbiol. Mol. Biol. Rev. 67, 686–723 (2003).
4. White, S. W., Zheng, J., Zhang, Y. M. & Rock, C. O. The structural biology of type II fatty acid biosynthesis. Annu. Rev. Biochem. 74, 791–831 (2005).
5. Kim, Y. M. & Prestegard, J. H. A dynamic model for the structure of acyl carrier protein in solution. Biochemistry 28, 8792–8797 (1989).
6. Xu, G. Y. et al. Solution structure of B. subtilis acyl carrier protein. Structure 9, 277–287 (2001).
7. Roujeinikova, A. et al. Structural studies of fatty acyl-(acyl carrier protein) thioesters reveal a hydrophobic binding cavity that can expand to fit longer substrates. J. Mol. Biol. 365, 135–145 (2007).
8. Zhang, Y. M., Marrakchi, H., White, S. W. & Rock, C. O. The application of computational methods to explore the diversity and structure of bacterial fatty acid synthase. J. Lipid Res. 44, 1–10 (2003).
9. De Lay, N. R. & Cronan, J. E. In vivo functional analyses of the type II acyl carrier proteins of fatty acid biosynthesis. J. Biol. Chem. 282, 20319–20328 (2007).
10. Cronan, J. E. Jr & Waldrop, G. L. Multi-subunit acetyl-CoA carboxylases. Prog. Lipid Res. 41, 407–435 (2002).
11. Jackowski, S. & Rock, C. O. Acetoacetyl-acyl carrier protein synthase, a potential regulator of fatty acid biosynthesis in bacteria. J. Biol. Chem. 262, 7927–7931 (1987).Reports the discovery of FabH, the condensing enzyme that initiates fatty acid elongation.
12. Choi, K. H., Heath, R. J. & Rock, C. O. -ketoacyl-acyl carrier protein synthase III (FabH) is a determining factor in branched-chain fatty acid biosynthesis. J. Bacteriol. 182, 365–370 (2000).
13. Qiu, X. et al. Crystal structure and substrate specificity of the -ketoacyl-acyl carrier protein synthase III (FabH) from Staphylococcus aureus. Protein Sci. 14, 2087–2094 (2005).
14. Choi, K. H., Kremer, L., Besra, G. S. & Rock, C. O. Identification and substrate specificity of -ketoacyl (acyl carrier protein) synthase III (mtFabH) from Mycobacterium turberculosis. J. Biol. Chem. 275, 28201–28207 (2000).
15. Musayev, F., Sachdeva, S., Neel Scarsdale, J., Reynolds, K. A. & Wright, H. T. Crystal structure of a substrate complex of Mycobacterium tuberculosis -ketoacyl-acyl carrier protein synthase III (FabH) with
lauroyl-coenzyme A. J. Mol. Biol. 346, 1313–1321 (2005).
16. Heath, R. J. & Rock, C. O. Roles of the FabA and FabZ -hydroxyacyl-acyl carrier protein dehydratases in
Escherichia coli fatty acid biosynthesis. J. Biol. Chem. 271, 27795–27801 (1996).
17. Heath, R. J. & Rock, C. O. Enoyl-acyl carrier protein reductase (fabI) plays a determinant role in completing cycles of fatty acid elongation in Escherichia coli. J. Biol. Chem. 270, 26538–26542 (1995).
18. Heath, R. J. & Rock, C. O. A triclosan-resistant bacterial enzyme. Nature 406, 145–146 (2000).Reports the discovery of FabK, a flavoprotein enoyl-ACP reductase that substitutes for FabI.
19. Marrakchi, H. et al. Characterization of Streptococcus pneumoniae enoyl-(acyl carrier protein) reductase (FabK). Biochem. J. 370, 1055–1062 (2003).
20. Massengo-Tiasse, R. P. & Cronan, J. E. Vibrio cholerae FabV defines a new class of enoyl acyl-carrier-protein reductase. J. Biol. Chem. 283, 1308–1316 (2008).
21. Lu, Y. J. et al. Acyl-phosphates initiate membrane phospholipid synthesis in Gram-positive pathogens. Mol. Cell 23, 765–772 (2006).Reports the discovery of the PlsX–PlsY pathway for glycerol-phosphate acylation, which involves an acyl-phosphate intermediate. This is the principal pathway that initiates membrane phospholipid synthesis in bacteria.
22. Paoletti, L., Lu, Y. J., Schujman, G. E., de Mendoza, D. & Rock, C. O. Coupling of fatty acid and phospholipid synthesis in Bacillus subtilis. J. Bacteriol. 189, 5816–5824 (2007).
23. Lu, Y. J., Zhang, F., Grimes, K. D., Lee, R. E. & Rock, C. O. Topology and active site of PlsY: the bacterial acylphosphate:glycerol-3-phosphate acyltransferase. J. Biol. Chem. 282, 11339–11346 (2007).
24. Cronan, J. E. Jr & Rock, C. O. Biosynthesis of Membrane Lipids in Escherichia coli and Salmonella typhimurium: Cellular and Molecular Biology (eds Neidhardt, F. C. et al.) 612–636 (American Society of Microbiology, Washington DC, 1996).
25. Coleman, J. Characterization of the Escherichia coli gene for 1-acyl-sn-glycerol-3-phosphate acyltransferase (plsC). Mol. Gen. Genet. 232, 295–303 (1992).
26. Jorasch, P., Wolter, F. P., Zahringer, U. & Heinz, E. A UDP glucosyltransferase from Bacillus subtilis successively transfers up to four glucose residues to 1,2-diacylglycerol: expression of ypfP in Escherichia coli and structural analysis of its reaction products. Mol. Microbiol. 29, 419–430 (1998).
27. Weissenmayer, B., Gao, J. L., Lopez-Lara, I. M. & Geiger, O. Identification of a gene required for the biosynthesis of ornithine-derived lipids. Mol. Microbiol. 45, 721–733 (2002).
28. Gao, J. L. et al. Identification of a gene required for the formation of lyso-ornithine lipid, an intermediate in the biosynthesis of ornithine-containing lipids. Mol. Microbiol. 53, 1757–1770 (2004).
29. Agun-Sunar, S. et al. Ornithine lipid is required for optimal steady-state amounts of c-type cytochromes in Rhodobacter capsulatus. Mol. Microbiol. 61, 418–435 (2006).
30. Voelker, T. A. & Davies, H. M. Alteration of the specificity and regulation of fatty acid synthesis of Escherichia coli by expression of a plant medium-chain acyl–acyl carrier protein thioesterase. J. Bacteriol. 176, 7320–7327 (1994).
31. Jiang, P. & Cronan, J. E. Jr. Inhibition of fatty acid synthesis in Escherichia coli in the absence of phospholipid synthesis and release of inhibition by thioesterase action. J. Bacteriol. 176, 2814–2821 (1994).
32. Cho, H. & Cronan, J. E. Jr. Defective export of a periplasmic enzyme disrupts regulation of bacterial fatty acid synthesis. J. Biol. Chem. 270, 4216–4219 (1995).
33. Davis, M. S. & Cronan, J. E. Jr. Inhibition of Eschericia coli acetyl coenzyme A carboxylase by acyl–acyl carrier protein. J. Bacteriol. 183, 1499–1503 (2001).
34. Heath, R. J. & Rock, C. O. Regulation of fatty acid elongation and initiation by acyl–acyl carrier protein in Escherichia coli. J. Biol. Chem. 271, 1833–1836 (1996).
35. Heath, R. J. & Rock, C. O. Inhibition of -ketoacyl-acyl carrier protein synthase III (FabH) by acyl–acyl carrier protein in Escherichia coli. J. Biol. Chem. 271, 10996–11000 (1996).
36. Heath, R. J., Jackowski, S. & Rock, C. O. Guanosine tetraphosphate inhibition of fatty acid and phospholipid synthesis in Escherichia coli is relieved by overexpression of glycerol-3-phosphate acyltransferase (plsB). J. Biol. Chem. 269, 26584–26590 (1994).
37. Magnusson, L. U., Farewell, A. & Nystrom, T. ppGpp: a global regulator in Escherichia coli. Trends Microbiol. 13, 236–242 (2005).
38. Seyfzadeh, M., Keener, J. & Nomura, M. spoT-dependent accumulation of guanosine tetraphosphate in response to fatty acid starvation in Escherichia coli. Proc. Natl Acad. Sci. USA 90, 11004–11008 (1993).
39. Battesti, A. & Bouveret, E. Acyl carrier protein/SpoT interaction, the switch linking SpoT-dependent stress response to fatty acid metabolism. Mol. Microbiol. 62, 1048–1063 (2006).
40. Schujman, G. E., Paoletti, L., Grossman, A. D. & de Mendoza, D. FapR, a bacterial transcription factor involved in global regulation of membrane lipid biosynthesis. Dev. Cell 4, 663–672 (2003).
41. Schujman, G. E. et al. Structural basis of lipid biosynthesis regulation in Gram-positive bacteria. EMBO J. 25, 4074–4083 (2006).Identifies malonyl-CoA as the ligand for FapR, a global transcriptional regulator of fatty acid biosynthetic genes in Gram-positive bacteria.
42. Xie, J., Bogdanov, M., Heacock, P. & Dowhan, W. Phosphatidylethanolamine and monoglucosyldiacylglycerol are interchangeable in supporting topogenesis and function of the polytopic membrane protein lactose permease. J. Biol. Chem. 281, 19172–19178 (2006).
43. Zhang, W., Campbell, H. A., King, S. C. & Dowhan, W. Phospholipids as determinants of membrane protein topology. Phosphatidylethanolamine is required for the proper topological organization of the -aminobutyric acid permease (GabP) of Escherichia
coli. J. Biol. Chem. 280, 26032–26038 (2005).44. Tsatskis, Y. et al. The osmotic activation of transporter
ProP is tuned by both its C-terminal coiled-coil and osmotically induced changes in phospholipid composition. J. Biol. Chem. 280, 41387–41394 (2005).
45. Dowhan, W. Molecular basis for membrane phospholipid diversity: why are there so many lipids? Annu. Rev. Biochem. 66, 199–232 (1997).
46. Rilfors, L. et al. Reconstituted phosphatidylserine synthase from Escherichia coli is activated by anionic phospholipids and micelle-forming amphiphiles. Biochim. Biophys. Acta 1438, 281–294 (1999).
47. Raetz, C. R. H., Larson, T. J. & Dowhan, W. Gene cloning for the isolation of enzymes of membrane lipid synthesis: phosphatidylserine synthase overproduction in Escherichia coli. Proc. Natl Acad. Sci. USA 74, 1412–1416 (1977).
R E V I E W S
NATURE REVIEWS | MICROBIOLOGY VOLUME 6 | MARCH 2008 | 231
© 2008 Nature Publishing Group

48. Tyhach, R. J., Hawrot, E., Satre, M. & Kennedy, E. P. Increased synthesis of phosphatidylserine decarboxylase in a strain of Escherichia coli bearing a hybrid plasmid. Altered association of enzyme with the membrane. J. Biol. Chem. 254, 627–633 (1979).
49. Salamon, Z., Lindblom, G., Rilfors, L., Linde, K. & Tollin, G. Interaction of phosphatidylserine synthase from E. coli with lipid bilayers: coupled plasmon–waveguide resonance spectroscopy studies. Biophys. J. 78, 1400–1412 (2000).
50. Linde, K., Grobner, G. & Rilfors, L. Lipid dependence and activity control of phosphatidylserine synthase from Escherichia coli. FEBS Lett. 575, 77–80 (2004).
51. Saha, S. K., Nishijima, S., Matsuzaki, H., Shibuya, I. & Matsumoto, K. A regulatory mechanism for the balanced synthesis of membrane phospholipid species in Escherichia coli. Biosci. Biotechnol. Biochem. 60, 111–116 (1996).
52. Saha, S. K., Furukawa, Y., Matsuzaki, H., Shibuya, I. & Matsumoto, K. Directed mutagenesis, Ser-56 to Pro, of Bacillus subtilis phosphatidylserine synthase drastically lowers enzymatic activity and relieves amplification toxicity in Escherichia coli. Biosci. Biotechnol. Biochem. 60, 630–633 (1996).
53. Tropp, B. E. Cardiolipin synthase from Escherichia coli. Biochim. Biophys. Acta 1348, 192–200 (1997).
54. Bernal, P., Munoz-Rojas, J., Hurtado, A., Ramos, J. L. & Segura, A. A Pseudomonas putida cardiolipin synthesis mutant exhibits increased sensitivity to drugs related to transport functionality. Environ. Microbiol. 9, 1135–1145 (2007).
55. Tropp, B. E. et al. Identity of the Escherichia coli cls and nov genes. J. Bacteriol. 177, 5155–5157 (1995).
56. Shibuya, I., Miyazaki, C. & Ohta, A. Alteration of phospholipid composition by combined defects in phosphatidylserine and cardiolipin synthases and physiological consequences in Escherichia coli. J. Bacteriol. 161, 1086–1092 (1985).
57. Lopez, C. S., Alice, A. F., Heras, H., Rivas, E. A. & Sanchez-Rivas, C. Role of anionic phospholipids in the adaptation of Bacillus subtilis to high salinity. Microbiology 152, 605–616 (2006).
58. Hiraoka, S., Matsuzaki, H. & Shibuya, I. Active increase in cardiolipin synthesis in the stationary growth phase and its physiological significance in Escherichia coli. FEBS Lett. 336, 221–224 (1993).
59. Ragolia, L. & Tropp, B. E. The effects of phosphoglycerides on Escherichia coli cardiolipin synthase. Biochim. Biophys. Acta 1214, 323–332 (1994).
60. Zhu, K., Bayles, D. O., Xiong, A., Jayaswal, R. K. & Wilkinson, B. J. Precursor and temperature modulation of fatty acid composition and growth of Listeria monocytogenes cold-sensitive mutants with transposon-interrupted branched-chain -ketoacid dehydrogenase. Microbiology 151, 615–623 (2005).
61. Giotis, E. S., McDowell, D. A., Blair, I. S. & Wilkinson, B. J. Role of branched-chain fatty acids in pH stress tolerance in Listeria monocytogenes. Appl. Environ. Microbiol. 73, 997–1001 (2007).
62. Cronan, J. E. Jr & Subrahmanyam, S. FadR, transcriptional co-ordination of metabolic expediency. Mol. Microbiol. 29, 937–943 (1998).
63. DiRusso, C. C., Heimert, T. L. & Metzger, A. K. Characterization of FadR, a global transcriptional regulator of fatty acid metabolism in Escherichia coli. Interaction with the fadB promoter is prevented by long chain fatty acyl coenzyme A. J. Biol. Chem. 267, 8685–8691 (1992).
64. Henry, M. F. & Cronan, J. E. Jr. A new mechanism of transcriptional regulation: release of an activator triggered by small molecule binding. Cell 70, 671–679 (1992).Reveals that FadR is an activator of fabA gene transcription.
65. Campbell, J. W. & Cronan, J. E. Jr. Escherichia coli FadR positively regulates transcription of the fabB fatty acid biosynthetic gene. J. Bacteriol. 183, 5982–5990 (2001).
66. Zhang, Y. M., Marrakchi, H. & Rock, C. O. The FabR (YijC) transcription factor regulates unsaturated fatty acid biosynthesis in Escherichia coli. J. Biol. Chem. 277, 15558–15565 (2002).Identifies FabR as a transcription factor that regulates UFA biosynthesis as a repressor of fabA and fabB gene expression.
67. Wang, H. & Cronan, J. E. Functional replacement of the FabA and FabB proteins of Escherichia coli fatty acid synthesis by Enterococcus faecalis FabZ and FabF homologues. J. Biol. Chem. 279, 34489–34495 (2004).
68. Lu, Y. J., White, S. W. & Rock, C. O. Domain swapping between Enterococcus faecalis FabN and FabZ proteins localizes the structural determinants for isomerase activity. J. Biol. Chem. 280, 30342–30348 (2005).
69. Lu, Y. J. & Rock, C. O. Transcriptional regulation of fatty acid biosynthesis in Streptococcus pneumoniae. Mol. Microbiol. 59, 551–566 (2006).
70. Garwin, J. L., Klages, A. L. & Cronan, J. E. Jr. -ketoacyl-acyl carrier protein synthase II of
Escherichia coli. Evidence for function in the thermal regulation of fatty acid synthesis. J. Biol. Chem. 255, 3263–3265 (1980).Identifies FabF as the condensing enzyme that is responsible for the temperature-dependent change in cis-vaccenate formation.
71. Ulrich, A. K., de Mendoza, D., Garwin, J. L. & Cronan, J. E. Jr. Genetic and biochemical analyses of Escherichia coli mutants altered in the temperature-dependent regulation of membrane lipid composition. J. Bacteriol. 154, 221–230 (1983).
72. Aquilar, P. S., Hernandez-Arriaga, A. M., Cybulski, L. E., Erazo, A. C. & de Mendoza, D. Molecular basis of thermosensing: a two component signal transduction thermometer in Bacillus subtilis. EMBO J. 20, 1681–1691 (2001).Describes DesR as a molecular sensor of the biophysical properties of membrane phospholipids.
73. Zhu, K., Choi, K. H., Schweizer, H. P., Rock, C. O. & Zhang, Y. M. Two aerobic pathways for the formation of unsaturated fatty acids in Pseudomonas aeruginosa. Mol. Microbiol. 60, 260–273 (2006).
74. Zhang, Y. M., Zhu, K., Frank, M. W. & Rock, C. O. A Pseudomonas aeruginosa transcription factor that senses fatty acid structure. Mol. Microbiol. 66, 622–632 (2007).
75. Grogan, D. W. & Cronan, J. E. Jr. Cyclopropane ring formation in membrane lipids of bacteria. Microbiol. Mol. Biol. Rev. 61, 429–441 (1997).
76. Wang, A. Y. & Cronan, J. E. Jr. The growth phase dependent synthesis of cyclopropane fatty acids in Escherichia coli is due to an RpoS (KatF) dependent promoter plus enzyme instability. Mol. Microbiol. 11, 1009–1017 (1994).
77. Chang, Y. Y., Eichel, J. & Cronan, J. E. Jr. Metabolic instability of Escherichia coli cyclopropane fatty acid synthase is due to RpoH-dependent proteolysis. J. Bacteriol. 182, 4288–4294 (2000).Together with reference 76, describes the dual regulation of Cfa by RpoH- and RpoS-dependent mechanisms.
78. Chang, Y. Y. & Cronan, J. E. Jr. Membrane cyclopropane fatty acid content is a major factor in acid resistance of Escherichia coli. Mol. Microbiol. 33, 249–259 (1999).
79. Munoz-Rojas, J. et al. Involvement of cyclopropane fatty acids in the response of Pseudomonas putida KT2440 to freeze-drying. Appl. Environ. Microbiol. 72, 472–477 (2006).
80. Glickman, M. S., Cox, J. S. & Jacobs, W. R. Jr. A novel mycolic acid cyclopropane synthetase is required for cording, persistence, and virulence of Mycobacterium tuberculosis. Mol. Cell 5, 717–727 (2000).Shows that cyclopropane fatty acid formation is required for the persistence and virulence of M. tuberculosis.
81. Morita, N. et al. Evidence for cis–trans isomerization of a double bond in the fatty acids of the psychrophilic bacterium Vibrio sp. strain ABE-1. J. Bacteriol. 175, 916–918 (1993).
82. Heipieper, H. J., Meulenbeld, G., van Oirschot, Q. & de Bont, J. A. M. Effect of environmental factors on the trans/cis ratio of unsaturated fatty acids in Pseudomonas putida S12. Appl. Environ. Microbiol. 62, 2773–2777 (1996).
83. Junker, F. & Ramos, J. L. Involvement of the cis/trans isomerase Cti in solvent resistance of Pseudomonas putida DOT-T1E. J. Bacteriol. 181, 5693–5700 (1999).
84. Holtwick, R., Meinhardt, F. & Keweloh, H. cis–trans isomerization of unsaturated fatty acids: cloning and sequencing of the cti gene from Pseudomonas putida P8. Appl. Environ. Microbiol. 63, 4292–4297 (1997).
85. Kiran, M. D., Annapoorni, S., Suzuki, I., Murata, N. & Shivaji, S. cis–trans isomerase gene in psychrophilic
Pseudomonas syringae is constitutively expressed during growth and under conditions of temperature and solvent stress. Extremophiles 9, 117–125 (2005).
86. Halverson, L. J. & Firestone, M. K. Differential effects of permeating and nonpermeating solutes on the fatty acid composition of Pseudomonas putida. Appl. Environ. Microbiol. 66, 2414–2421 (2000).
87. Peschel, A. How do bacteria resist human antimicrobial peptides? Trends Microbiol. 10, 179–186 (2002).
88. Oku, Y., Kurokawa, K., Ichihashi, N. & Sekimizu, K. Characterization of the Staphylococcus aureus mprF gene, involved in lysinylation of phosphatidylglycerol. Microbiology 150, 45–51 (2004).
89. Peschel, A. et al. Staphylococcus aureus resistance to human defensins and evasion of neutrophil killing via the novel virulence factor MprF is based on modification of membrane lipids with L-lysine. J. Exp. Med. 193, 1067–1076 (2001).Connects the formation of lysyl-phosphatidylglycerol with the resistance of bacteria to defensins.
90. Staubitz, P. & Peschel, A. MprF-mediated lysinylation of phospholipids in Bacillus subtilis — protection against bacteriocins in terrestrial habitats? Microbiology 148, 3331–3332 (2002).
91. Thedieck, K. et al. The MprF protein is required for lysinylation of phospholipids in listerial membranes and confers resistance to cationic antimicrobial peptides (CAMPs) on Listeria monocytogenes. Mol. Microbiol. 62, 1325–1339 (2006).
92. Nishi, H., Komatsuzawa, H., Fujiwara, T., McCallum, N. & Sugai, M. Reduced content of lysyl-phosphatidylglycerol in the cytoplasmic membrane affects susceptibility to moenomycin, as well as vancomycin, gentamicin, and antimicrobial peptides, in Staphylococcus aureus. Antimicrob. Agents Chemother. 48, 4800–4807 (2004).
93. Goldberg, D. E., Rumley, M. K. & Kennedy, E. P. Biosynthesis of membrane-derived oligosaccharides: a periplasmic phosphoglyceroltransferase. Proc. Natl Acad. Sci. USA 78, 5513–5517 (1981).
94. Kennedy, E. P. Osmotic regulation and biosynthesis of membrane-derived oligosaccharides in Escherichia coli. Proc. Natl Acad. Sci. USA 79, 1092–1095 (1982).
95. Raetz, C. R. H. & Newman, K. F. Diglyceride kinase mutants of Escherichia coli: inner membrane association of 1,2-diglyceride and its relation to synthesis of membrane-derived oligosaccharides. J. Bacteriol. 137, 860–868 (1979).
96. Rumley, M. K., Therisod, H., Weissborn, A. C. & Kennedy, E. P. Mechanisms of regulation of the biosynthesis of membrane-derived oligosaccharides in Escherichia coli. J. Biol. Chem. 267, 11806–11810 (1992).
97. Lacroix, J. M., Loubens, I., Tempete, M., Menichi, B. & Bohin, J. P. The mdoA locus of Escherichia coli consists of an operon under osmotic control. Mol. Microbiol. 5, 1745–1753 (1991).
98. Raetz, C. R. H. & Newman, K. F. Neutral lipid accumulation in the membranes of Escherichia coli mutants lacking diglyceride kinase. J. Biol. Chem. 253, 3882–3887 (1978).
99. Bielawska, A., Perry, D. K. & Hannun, Y. A. Determination of ceramides and diglycerides by the diglyceride kinase assay. Anal. Biochem. 298, 141–150 (2001).
100. Jerga, A., Lu, Y. J., Schujman, G. E., de Mendoza, D. & Rock, C. O. Identification of a soluble diacylglycerol kinase required for lipoteichoic acid production in Bacillus subtilis. J. Biol. Chem. 282, 21738–21745 (2007).
101. Koch, H. U., Haas, R. & Fischer, W. The role of lipoteichoic acid biosynthesis in membrane lipid metabolism of growing Staphylococcus aureus. Eur. J. Biochem. 138, 357–363 (1984).
102. Taron, D. J., Childs, W. C. & Neuhaus, F. C. Biosynthesis of D-alanyl-lipoteichoic acid: role of diglyceride kinase in the synthesis of phosphatidylglycerol for chain elongation. J. Bacteriol. 154, 1110–1116 (1983).
103. Jackowski, S., Jackson, P. D. & Rock, C. O. Sequence and function of the aas gene in Escherichia coli. J. Biol. Chem. 269, 2921–2928 (1994).
104. Harvat, E. M. et al. Lysophospholipid flipping across the Escherichia coli inner membrane catalyzed by a transporter (LplT) belonging to the major facilitator superfamily. J. Biol. Chem. 280, 12028–12034 (2005).
105. Hsu, L., Jackowski, S. & Rock, C. O. Isolation and characterization of Escherichia coli K-12 mutants
R E V I E W S
232 | MARCH 2008 | VOLUME 6 www.nature.com/reviews/micro
© 2008 Nature Publishing Group

lacking both 2-acyl-glycerophosphoethanolamine acyltransferase and acyl–acyl carrier protein synthetase activity. J. Biol. Chem. 266, 13783–13788 (1991).
106. Zhang, Y. M., White, S. W. & Rock, C. O. Inhibiting bacterial fatty acid synthesis. J. Biol. Chem. 281, 17541–17544 (2006).
107. Revill, W. P., Bibb, M. J. & Hopwood, D. A. Purification of a malonyltransferase from Streptomyces coelicolor A3(2) and analysis of its genetic determinant. J. Bacteriol. 177, 3946–3952 (1995).
108. Zheng, C. J. et al. Cephalochromin, a FabI-directed antibacterial of microbial origin. Biochem. Biophys. Res. Commun. 362, 1107–1112 (2007).
109. McMurray, L. M., Oethinger, M. & Levy, S. Triclosan targets lipid synthesis. Nature 394, 531–532 (1998).
110. Heath, R. J. et al. Mechanism of triclosan inhibition of bacterial fatty acid synthesis. J. Biol. Chem. 274, 11110–11114 (1999).
111. Zheng, C. J., Sohn, M. J. & Kim, W. G. Atromentin and leucomelone, the first inhibitors specific to enoyl-ACP reductase (FabK) of Streptococcus pneumoniae. J. Antibiot. 59, 808–812 (2006).
112. Takahata, S. et al. AG205, a novel agent directed against FabK of Streptococcus pneumoniae. Antimicrob. Agents Chemother. 50, 2869–2871 (2006).
AcknowledgementsResearch in the authors’ laboratory is supported by National Institute of General Medical Sciences grant GM34496, Cancer Center (CORE) Support grant CA21765 and the American Lebanese Syrian Associated Charities.
DATABASESEntrez Genome Project: http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=genomeprjBacillus subtilis | Enterococcus faecalis | Escherichia coli | Listeria monocytogenes | Mycobacterium tuberculosis | Pseudomonas putida | Streptococcus pneumoniae
FURTHER INFORMATIONCharles O. Rock’s homepage: http://www.stjude.org/stjude/v/index.jsp?vgnextoid=3a6e10e88ce70110VgnVCM1000001e0215acRCRD&vgnextchannel=0dd413c016118010VgnVCM1000000e2015acRCRD
ALL LINKS ARE ACTIVE IN THE ONLINE PDF
R E V I E W S
NATURE REVIEWS | MICROBIOLOGY VOLUME 6 | MARCH 2008 | 233
© 2008 Nature Publishing Group