atf3 expression in cardiomyocytes preserves homeostasis in
TRANSCRIPT

ATF3 expression in cardiomyocytes preserves
homeostasis in the heart and controls peripheral
glucose tolerance
Roy Kalfon1 Lilach Koren1 Sharon Aviram1 Ortal Schwartz2 Tsonwin Hai3 and
Ami Aronheim1
1Department of Cell Biology and Cancer Science B Rappaport Faculty of Medicine Technion-Israel Institute of Technology 7th Efron St Bat-Galim Haifa 31096 Israel 2BiomedicalCore Facility B Rappaport Faculty of Medicine Technion-Israel Institute of Technology Haifa Israel and 3Departments of Biological Chemistry and Pharmacology Ohio State University1060 Carmack road Columbus OH 43210 USA
Received 4 April 2016 revised 1 September 2016 editorial decision 16 October 2016 accepted 28 October 2016 online publish-ahead-of-print 7 November 2016
Time of primary review 34 days
Aims Obesity and type 2 diabetes (T2D) trigger a harmful stress-induced cardiac remodeling process known as cardiomy-opathy These diseases represent a serious and widespread health problem in the Western world however theunderlying molecular basis is not clear ATF3 is an lsquoimmediate earlyrsquo gene whose expression is highly and transientlyinduced in response to multiple stressors such as metabolic oxidative endoplasmic reticulum and inflammationstressors that are involved in T2D cardiomyopathy The role of ATF3 in diabetic cardiomyopathy is currentlyunknown Our research has aimed to study the effect of ATF3 expression on cardiomyocytes heart function andglucose homeostasis in an obesity-induced T2D mouse model
Methods andresults
We used wild type mice (WT) as well as mutant mice with a cardiac-specific ATF3 deficiency (ATF3-cKO) Mice werefed a high-fat diet (HFD) for 15 weeks HFD induced high ATF3 expression in cardiomyocytes Mice were examined forcardiac remodeling processes and the diabetic state was assessed HFD-fed ATF3-cKO mice exhibited severe cardiac fib-rosis higher levels of heart hypertrophic markers increased inflammation and worse cardiac function as compared toWT mice Interestingly HFD-fed ATF3-cKO mice display increased hyperglycemia and reduced glucose tolerance despitehigher blood insulin levels as compared to HFD-fed WT mice Elevated levels of the cardiac inflammatory cytokines IL-6and TNFa leading to impaired insulin signalling may partially explain the peripheral glucose intolerance
Conclusions Cardiac ATF3 has a protective role in dampening the HFD-induced cardiac remodeling processes ATF3 exerts
both local and systemic effects related to T2D-induced cardiomyopathy This study provides a strong relationshipbetween heart remodeling processes and blood glucose homeostasis
Keywords Type 2 diabetes bull Cardiac remodeling bull Cytokines bull High fat diet bull ATF3
1 Introduction
Current research has estimated that approximately 371 million peopleworldwide suffer from diabetes some 90 of these cases are type 2 dia-betes (T2D) The number of diagnosed diabetes cases has doubled since1980 due to the aging population and the rising incidence of risk factorssuch as obesity1 Cardiovascular diseases which include heart failure arethe leading cause of morbidity and mortality in T2D patients Of theseT2D patients about two-thirds die from heart failure23 Diabetesincreases the risk of developing diabetic cardiomyopathy3ndash5 Diabeticcardiomyopathy is a disease that damages the structure and function of
the heart resulting in cardiac metabolic changes hypertrophic growthmyocardial fibrosis and arrhythmias which eventually cause heart fail-ure67 The molecular mechanism of this linkage between obesity-diabetes-cardiomyopathy is not well understood In light of the growingand wide-spread incidence of T2D and obesity in modern society this isa major research challenge that can help to improve public health
The heart meets its energy requirements via the oxidation of fattyacids glucose lactate and other oxidizable substrates The heart func-tions best when both fatty acids (70) and glucose (30) are simultane-ously used as substrates for ATP production6 Increased fatty acid levelsin the blood promote the production of hormones and inflammatory
Corresponding author Tel thorn972 4 8295454 fax thorn972 4 8295225 E-mail aronheimtxtechnionacil
Published on behalf of the European Society of Cardiology All rights reserved VC The Author 2016 For Permissions please email journalspermissionsoupcom
Cardiovascular Research (2017) 113 134ndash146doi101093cvrcvw228
Dow
nloaded from httpsacadem
icoupcomcardiovascresarticle11321342687691 by guest on 29 June 2022
cytokines by adipocytes resulting in whole-body insulin resistance8
Under these conditions the heart rapidly adapts by consuming primarilyfatty acids as a source of ATP generation Despite an increase in fattyacid metabolism it is likely that fatty acid uptake exceeds oxidation ratesthereby resulting in cellular lipid accumulation-induced lipotoxicity5910
At the molecular level insulin signalling activates multiple pathways tar-geting various transcription factors resulting in re-programing of adaptiveor maladaptive cardiac remodeling processes11
In this report we identify an important transcription factor that playsa major role in this process Activating Transcription Factor 3 (ATF3)ATF3 is a member of the basic leucine zipper (bZIP) superfamily of tran-scription factors and is encoded by an immediate early gene whoseexpression is low under normal condition but is rapidly and highlyinduced by multiple stressors12
ATF3 binds to the cyclic AMP response element (CRE) and typicallydampens the expression of multiple target genes Depending on the con-text ATF3-mediated response can be adaptive or maladaptive1213 Forexample the role of ATF3 in cardiac diseases varies depending on thestress paradigms Neuroendocrine hormones stimuli induce ATF3 tran-scription both in vitro14 and in vivo15 Mice with ATF3 deficiency displayreduced cardiac remodeling and hypertrophy in a phenylephrine-induced pressure overload model1617 Consistently ectopic ATF3expression in cardiomyocytes is sufficient to promote heart growth andcardiac dysfunction in transgenic mice1618 Together this suggests a del-eterious role of ATF3 in cardiac diseases In contrast studies using ATF3deficient mice in the context of transverse aortic constriction suggest acardiac protective role for ATF31920 Thus the importance of ATF3 incardiac remodeling models is well documented but its effects varydepending on the stress paradigms21ndash24
In addition to cardiac diseases ATF3 has been shown to play a role inother diseases including diabetes Previously we showed that mice defi-cient in ATF3 had reduced glucose tolerance and more severe T2D thanthe wild type mice under a high-fat diet (HFD)25 We investigated ATF3in this context since ATF3 is known to be induced by stresses that play arole in obesity-induced T2D cardiomyopathy including metabolic stressoxidative stress endoplasmic reticulum (ER) stress and inflamma-tion1226 However the role of ATF3 in diabetes-induced cardiomyop-athy has never been reported
In this article we set out to test whether cardiomyocytes-selectivedeletion of ATF3 alters HFD-induced T2D cardiomyopathy We presentevidence that following a HFD cardiac ATF3 deficiency exacerbates car-diac remodeling inflammation maladaptive metabolic response and car-diac dysfunction Surprisingly it also worsens blood glucose intoleranceincluding a reduced ability of the peripheral tissues to respond to insulinSince ATF3 deletion is selectively in cardiomyocytesmdashwhich is not a celltype known to play a role in diabetesmdashthis result is novel and intriguing
2 Methods
21 MiceThis study was carried out in strict accordance with the Guide for theCare and Use of Laboratory Animals of the National Institute of HealthIn addition our protocol was approved by the Committee of the Ethicsof Animal Experiments of the Technion All mice strains used in thisstudy were backcrossed to C57Bl6J for at least six generations Micestrains used in this study whole-body ATF3 KO (ATF3-KO)27 ATF3-flox (ATF3ff)28 aMHC-Cre29 and loxP-XFP transgenic (confetti) mice30
To generate mice with a cardiomyocyte-specific deletion of ATF3
(ATF3-cKO) we crossed mice carrying ATF3-flox alleles with transgenicmice harbouring Cre recombinase driven by a aMHC promoter Matingcages were set up to produce progeny with only one copy of aMHC-Cre to avoid variability in Cre expression31 ATF3ff and ATF3-cKO malemice littermates were housed in the same cage on a 12-h lightdark cycleand given ad libitum access to food and water All experimental groupswere fed with normal chow (NC 44 fat) for 10 weeks Subsequentlymice were either left on the NC diet or switched to high-fat diet (HFD60 fat TD06414 Harlan Inc) for additional 15 weeks
22 Glucose tolerance test fasting glucosetest and blood biochemistryThe mice fasted for 6 h and were then intraperitoneally injected with2 gkg of glucose Blood samples were obtained from their snipped tailand analysed using a portable glucometer Blood from non-fasting micewas collected from submandibular bleeds and serum insulin levels weremeasured by ELISA (Merck Millipore) Total cholesterol and high densitylipoprotein (HDL) cholesterol were measured at the Rambam ClinicalBiochemistry Laboratory
23 Insulin signallingFor insulin signalling analysis HFD-fed ATF3ff and ATF3-cKO micefasted for 6 h and were then intraperitoneally injected with insulin (10 Ukg Lantus SoloStar Sanofi-Aventis NJ) Mice were euthanized by aninjection of 01 mL of 12 mgmL Xylazine 60 mgmL Ketamine mixture
Heart liver and skeletal muscle (gastrocnemius) were harvested15 min following insulin stimulation Tissue lysates were then prepared
24 EchocardiographyMice were anesthetized with 1 of isoflurane and kept on a 37 Cheated plate throughout the analysis An echocardiography was per-formed using a Vevo2100 micro-ultrasound imaging system(VisualSonics Fujifilm) which was equipped with 13ndash38 MHz (MS 400)and 22ndash55 MHz (MS550D) linear array transducers The person per-forming echocardiography and the data analysis was blinded to the micegenotype Cardiac size shape and function were analysed by conven-tional two-dimensional imaging and M-Mode recordings Maximal leftventricles end-diastolic (LVDd) and end-systolic (LVDs) dimensionswere measured in short-axis M-mode images Fractional shortening (FS)was calculated as FS () = [(LVDd-LVDs)LVDd]100 All values werebased on the average of at least five measurements
25 MRI hardware and animal monitoringMRI was performed on a 94T bore scanner (Bruker Biospec EttlingenGermany) using a cylindrical volume coil (86 mm inner diameter) for sig-nal excitation and a single channel surface coil (20 mm diameter) for sig-nal reception The animal was put under anaesthesia with aconcentration of 05ndash15 isoflurane and supplemented with oxygen(05 Lmin) Respiration was monitored during imaging (Small AnimalInstruments Stony Brook New York NY) and body temperature wasmaintained using thermostat-regulated circulating hot water
26 Cardiac magnetic resonance (CMR)sequencesThe mice were scanned using cine FLASH sequence acquired from theself-gating technique IntraGate (Bruker BioSpin MRI EttlingenGermany) The performed CMR protocol comprised of three sets ofscans (i) Single slice Long Axis view for general evaluation of the heart
ATF3 expression in cardiomyocytes preserves homeostasis 135D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
structure and accurate planning of the subsequent scans (ii) Single slicetwo chamber Short Axis view perpendicular to LV walls at the level ofthe papillary muscles and (iii) Contiguous multi slices two chamber ShortAxis view for entire LV coverage
Cardiac MR sequences parameters
i Single slice Long Axis view 1 mm thickness FOV = 3 cm3 cm matrixdimension = 400400 spatial resolution = 7575lm2 repetitionechotime (TRTE) = 6532 ms number of repetitions = 100 10 cardiacmovie frames (cardiac phases)
ii Single slice two chamber Short Axis view 07 mm thicknessFOV=3 cm3 cm matrix dimension=400400 spatial reso-lution=7575lm2 repetitionecho time (TRTE) = 6532 ms numberof repetitions = 150 15 cardiac movie frames (cardiac phases)
iii Multi slices two chamber Short Axis view Contiguous 9ndash12 slicesdepending on heart size 07 mm thickness FOV = 3 cm3 cm matrixdimension = 400400 spatial resolution=7575lm2 repetitionechotime (TRTE) = 81-12032 ms (depending on slices number) number ofrepetitions = 120 15 cardiac movie frames (cardiac phases)
The total acquisition time for each animal was40 min
27 MRI image analysisImage analysis was performed using the freely available Segment soft-ware (Medviso AB Lund Sweden)32 Manual segmentation of the endo-cardium and epicardium of the LV was performed to measure LV massand LV functional parametersmdashend-systolic and end-diastolic (ESV andEDV respectively) volumes Ejection fraction (EF) was calculated as EF() = [(EDV-ESV)EDV]100 The person performing the MRI and thedata analysis was blinded to the mice genotype
28 Heart harvestingAt endpoint mice were anesthetized weighed and sacrificed Heartswere excised and ventricles were then divided into three pieces thatwere used for protein extraction RNA purification and histologicalanalysis
29 mRNA extractionmRNA was purified from ventricles using an Aurum total RNA fatty orfibrous tissue kit (732-6830 Bio-Rad) or from isolated cardiomyocytesusing a High Pure RNA Isolation kit (11828665001 Roche Diagnostics)according to the manufacturerrsquos instructions
210 Quantitative real-time PCR (RT-qPCR)cDNA was synthesized from 800 or 50 ng of purified mRNA derivedfrom the ventricles or isolated from the cardiomyocytes respectivelyPurified mRNA was added to a total reaction mix of high-capacity cDNAreverse transcription kit (4368814 Applied Biosystems) in a final vol-ume of 20 mL Real-time PCR was performed using Rotor-Gene6000TM (Corbett) equipment with absolute blue SYBER green ROXmix (Thermo Scientific AB-4162B) Serial dilutions of a standard samplewere included for each gene to generate a standard curve Values werenormalized to b2 microglobulin expression levels The primer sequencesare shown in Table S1 (see Supplementary material online)
211 Western-blot analysisHarvested tissues were homogenized in RIPA buffer (PBS containing 1NP-40 5 mgmL Na-deoxycholate 01 SDS) and supplemented withprotease inhibitor cocktail (P-8340 SigmandashAldrich) Homogenization
was performed at 4 C using the Bullet Blender homogenizer (BBX24Next advance) according to the manufacturerrsquos instructions as previ-ously described17
212 ImmunohistochemistryTissues were fixed in 4 formaldehyde overnight then embedded inparaffin serially sectioned at 10 lm intervals and then mounted onslides Sections were processed for deparaffinization (xylene 20 min)dehydration (isopropanol) rehydration (H2O) and antigen unmasking(10 mM sodium citrate pH-6 90 C 12 min) Immunostaining was per-formed using Histostain kit (959643 Invitrogen) according to the man-ufacturerrsquos instructions Anti-ATF3 antibodies were diluted 150Following immunostaining nuclei were stained with hematoxylin
213 AntibodiesThe primary antibodies used anti-SMA (Cat A2547) anti-a-tubulin(Cat T-9026) and anti-a-actinin (Cat A7811) which were all pur-chased from SigmandashAldrich Anti-GFP (SC-8334) was purchased fromSanta-Cruz Anti-p38 (Cat 9212) and anti-phospho-p38 (Cat 9211)were purchased from Cell signalling Anti-pAkt (Cat AF887) and anti-Akt (Cat MAB 2055) was purchased from RampD Systems anti-Foxo3(Cat 07-1719) was purchased from Merck Millipore and anti-Glut4(Cat ab654) was purchased from Abcam Lastly anti-ATF3 antibodywas generated in Aronheimrsquos lab15
214 Cell size analysisSections were stained following deparaffinization with Wheat-germagglutinin TRITC-conjugated (SigmandashAldrich Cat L5266) and diluted toa 1100 phosphate-buffered saline (PBS) Sections were washed 3 timeswith PBS and mounted in Fluoromount-G (Southern BiotechnologyBirmingham AL 0100-01) Sections were then viewed using a ZeissLSM700 confocal microscope (Thornwood NY) equipped with a 40oil objective and solid state 555nm laser
Quantification of the cell size was performed with Image Pro Plus soft-ware Five fields in each slide were photographed Unstained areas werethen identified and segmented using Image Pro Plus software In eachstained area the mean cell perimeter and area was calculated and thenumber of cells was measured
215 Fibrosis stainingHeart tissue was fixed in 4 formaldehyde overnight embedded in par-affin serially sectioned at 10 mm intervals and then mounted on slidesMassonrsquos trichrome staining was performed according to the standardprotocol Images were acquired by using Virtual Microscopy (Olympus)Stained areas were segmented and quantified following calibration usingImage Pro Plus software Five fields were quantified for each slide
216 In vitro cardiomyocyte cultureAdult cardiomyocytes were isolated from 12- to 25-week-old NC- orHFD-fed mice by a modified Langendorff perfusion apparatus asdescribed33 Isolated cells were plated in culture media (MEMmdashEagleHanksrsquo salts) supplemented with penicillin 100 UmL glutamine 2 mMNaHCO3 4 mM HEPES 10 mM bovine serum albumin (BSA) 0225 mM blebbistatin (cat B0560 SigmandashAldrich) and 5 foetal bovineserum Cardiomyocytes were then plated on 35-mm dishes coated withmouse laminin (L2020 SigmandashAldrich) at a density of 105 cellsmL inserum free medium
136 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
217 Palmitate-treated cardiomyocytesForty-five minutes after plating the medium was replaced with a culturemedium containing either 04 mM palmitate or BSA (control) Palmitatestock solution (20 mM) was diluted in 001 M NaOH by heating at 70Cfor 30 min as described10 Following overnight culture conditionedmedium was collected and cardiomyocytes were harvested for RNAextraction In addition overnight palmitate- or BSA-treated cardiomyo-cytes were incubated with 100 nM insulin (I-5500 SigmandashAldrich) for30 min and were then harvested for protein extraction
218 Enzyme-linked immuno-absorbentassayInterleukin 6 (IL-6) and tumour necrosis factor a (TNFa) levels in condi-tioned medium from an overnight culture of primary cardiomyocyteswere measured by ELISA using the DuoSet ELISA kit (RampD Systems)according to the manufacturerrsquos instructions Blood from non-fastingHFD-fed mice was collected from submandibular bleeds and serum IL-6and TNFa levels were measured by ELISA (RampD Systems)
219 StatisticsOur data is expressed as means plusmn SD The statistical test for the GTTwas performed by repeated-measures analysis of variance (ANOVA)The comparison of two means was analysed by the Studentrsquos t-test Thecomparison between several means was analysed by one-way ANOVAfollowed by Tukeyrsquos post hoc analysis All statistical analyses were per-formed using GraphPad Prism 5 software (La Jolla CA) A P value lt_ 005was accepted as statistically significant
3 Results
31 aMHC promoter specificityFirst we verified the tissue specific expression of the Cre recombinaseunder the control of the aMHC promoter by crossing aMHC-Cremouse with the loxP-XFP transgenic (confetti) mouse30 In the confettimouse lox-P specific recombination resulted in the random expressionof one out of four fluorescent proteins (BFP GFP RFP or YFP) Celllysates were prepared from various tissues of either the confettiff mouse(without Cre -) or the double transgenic mouse (containing Cre 1) fol-lowed by western-blot analysis using anti-GFP antibodies As expectedGFP-related proteins were strongly detected in the heart lysate (seeSupplementary material online Figure S1) The lysate derived from lungtissue displayed a lower level of GFP-related proteins as compared tothe heart lysate probably due to low transcription activity of the aMHCpromoter in the lungs34 GFP-related protein expression was undetect-able in all of the other tissues tested (see Supplementary material onlineFigure S1)
32 In vitro analysis of cardiomyocytestreated with free fatty acidsTo reveal the possible role of ATF3 in cardiomyopathy induced by freefatty acids (FFA) (palmitate) we analysed adult cardiomyocyte primarycultures derived from ATF3ff and ATF3-cKO mice (ATF3-cKO geno-type ATF3ffaMHC-Cre) Cardiomyocytes were isolated by theLangendorff perfusion apparatus followed by overnight treatment with04 mM palmitate10 qRT-PCR analysis showed a 4- to 5-fold increase inATF3 induction by palmitate in the ATF3ff cardiomyocytes (Figure 1A)Immuno-fluorescence analysis of cardiomyocytes treated with either 1 h
or overnight palmitate displayed a robust ATF3 nuclear staining (Figure1B) Consistent with a previous study35 western-blot analysis of the car-diomyocytes lysate following overnight palmitate treatment demon-strated a significant induction of the ATF3 protein (Figure 1C) SinceATF3 induction is dependent on the activation of the p38-MAPK signal-ling pathway1026 we measured the p38-MAPK (p38) activation stateusing anti-phospho-p38 (p-p38) While p-p38 levels were increased fol-lowing palmitate treatment p38 remained unchanged independent ofthe treatment (Figure 1C) As expected neither ATF3 transcript (Figure1A) nor ATF3 protein (Figure 1C) was detected in the palmitate-treatedATF3-cKO cardiomyocytes Importantly palmitate treatment inducedthe expression of cardiac hypertrophic markers (ANP and BNP) inflam-mation markers (IL-6 and TNFa) a metabolic marker (PDK4) and anextracellular matrix (ECM) remodeling protein (TIMP1) (Figure 1A)Strikingly significantly higher level of induction of these genes wasobserved in the ATF3-cKO cardiomyocytes as compared with ATF3ff
cardiomyocytes (Figure 1A) In addition while the expression level of theinsulin receptor substrate IRS1 was induced 2-fold by palmitate treat-ment in ATF3ff IRS1 expression was suppressed in ATF3-cKO cardio-myocytes (Figure 1A)
Next we quantified the level of secreted inflammatory cytokines byanalysing the conditioned medium derived from palmitate-treated cardi-omyocytes The levels of IL-6 and TNFa were significantly higher in theconditioned medium derived from palmitate-treated ATF3-cKO cardio-myocytes as compared to their conditioned media levels derived frompalmitate-treated ATF3ff cardiomyocytes (Figure 1D)
Collectively the above-mentioned results support an increase of ATF3expression in cardiomyocytes in response to acute and chronic exposureto FFA The analysis of cardiomyocytes derived from wild type and ATF3-cKO genotypes in vitro is consistent with a possible protective role ofATF3 by dampening the cardiomyocytes remodeling response to FFA
33 The effect of high-fat diet on wild typeand ATF3-cKO miceTo determine whether cardiac ATF3 expression plays a role in responseto a high-fat diet (HFD)-regimen we used mice harbouring acardiomyocyte-selective deletion ATF3-cKO and their wild type (ATF3f
f) counterparts Mice were fed either a normal chow (NC 44 fat) or aHFD (60 fat) The HFD was initiated at 10 weeks of age and continuedfor 15 weeks Both ATF3ff and ATF3-cKO mice fed with a HFD gainedweight at significantly higher rates than NC-fed mice with no significantdifferences between the genotypes (see Supplementary material onlineFigure S2) In addition the levels of circulating total cholesterol and highdensity lipoprotein (HDL) were significantly higher in HFD-fed mice incomparison to NC-fed mice with no significant difference between thegenotypes (see Supplementary material online Figure S3A and B)
34 Cardiomyocyte-selective ATF3deficiency induces cardiac remodelingSince ATF3 is highly induced in cardiomyocytes that are treated withFFA (04 mM palmitate) we studied the expression of ATF3 in heart tis-sue following 15 weeks of a HFD regimen by qRT-PCR ATF3 expres-sion was induced 6-fold in the hearts of HFD-fed ATF3ff mice whencompared to either the NC-fed mice or to HFD-fed ATF3-cKO (Figure2A) We next performed immuno-histochemistry staining of heart slicesderived from NC- and HFD-fed mice with anti-ATF3 antibodies Whileno ATF3 staining was observed in the hearts derived from NC-fed mice
ATF3 expression in cardiomyocytes preserves homeostasis 137D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
or HFD-fed ATF3-cKO mice efficient ATF3 nuclear staining wasobserved in HFD-fed ATF3ff mice (Figure 2B)
In addition western-blot analysis using anti-ATF3 antibodies with car-diomyocyte cell lysate confirmed a high level of ATF3 protein expressionin HFD-fed as compared with NC-fed mice (Figure 2C and D) We nextexamined the cardiac expression levels of various gene specific markersas an indication for cardiac remodeling processes of a stressed heartHFD-fed ATF3-cKO but not ATF3ff mice exhibited significantly higherlevels of the foetal gene programming markers (ANP BNP and bMHC)when compared to the NC-fed wild type counterparts (Figure 2A)Concomitantly a-myosin heavy chain (aMHC) expression was signifi-cantly reduced in HFD-fed mice as compared to NC-fed mice independ-ent of their genotype (Figure 2A) The switch from aMHC to bMHC is anindication of the cardiac remodeling process as well as a cardiac disease
state Heart sections stained with wheat-germ hemagglutinin revealed asignificant increase in the cross sectional area in HFD-fed mice withlarger increased area in ATF3-cKO mice as compared with ATF3ff mice(Figure 2E and F) Taken together cardiac remodeling induced by a HFDis more pronounced in the ATF3-cKO mice than that in the controlATF3ff mice
We next examined fibrosis markers and found elevated expression ofcollagen 1a cTGF and a-SMA in HFD-fed mice as compared to the cor-responding NC-fed mice (Figure 3A) In all cases the ATF3-cKO miceshowed a more pronounced phenotype (Figure 3A) Western-blot analysiswith cell lysates derived from both NC-fed and HFD-fed mice confirmedincreased levels of a-SMA in HFD-fed mice but these high levels weremore noticeable in the lysate derived from HFD-fed ATF3-cKO mice(Figure 3B and C) Another fibrosis marker TGFb3 is significantly elevated
A C
B D
Figure 1 Free fatty acids affect the expression of hypertrophic inflammatory metabolic and fibrosis markers in primary cardiomyocytes Cardiomyocyteswere isolated from adult ATF3ff and ATF3-cKO mice and primary cultures were generated (A) Cardiomyocytes were exposed to 04 mM palmitate orvehicle control for overnight mRNA was extracted and the expression levels of the indicated genes were studied by qRT-PCR The mean ratio of palmitatetreated vs untreated cells for each genotype is shown The results represent the mean ratio plusmn SD (n = 5 independent preparations) (B) ATF3ff cardiomyo-cytes were exposed to 04 mM palmitate for either 1 h or overnight and then fixed and stained with anti-ATF3 (red) and anti-a-actinin (green) antibodiesNuclei were stained with DAPI (blue) (C) Western-blot analysis of cardiomyocytes lysate either palmitate treated overnight or left untreated probed withanti-ATF3 anti-p38 and anti-phospho-p38 antibodies The expression level of a-tubulin was used as loading control Each lane represents a lysate preparedfrom a pool derived from three independent cardiomyocytes culture (D) Isolated cardiomyocytes were treated as described in A Conditioned mediumwas collected and the levels of the indicated cytokines were assessed by ELISA The results represent the mean ratio between palmitate treated tountreatedplusmn SD (n = 5ndash6 independent preparations) P lt_ 005 comparing treated to untreated cells of the same genotype daggerP lt_ 005 comparing palmitate-treated cells between genotypes
138 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
in ATF3-cKO mice but not in the control ATF3ff mice (Figure 3A) againindicating that HFD has a more profound effect on ATF3-cKO mice
We next examined fibrosis histologically by Massonrsquos trichrome stain-ing This analysis confirmed a significant increased fibrosis in the hearts ofHFD-fed mice with ATF3-cKO mice exhibiting the highest fibrosis level(Figure 3D and E) Fibrosis is a result of the deposition of collagen in theECM The net effect of ECM degrading enzymes such as metalloprotei-nases (MMPs) and their inhibitors (TIMPs) determines the level of colla-gen deposition36 The levels of MMP9 were significantly decreased inHFD-fed mice as compared to NC-fed mice independent of the micegenotype (Figure 3A) In contrast TIMP1 levels were significantlyincreased in HFD-fed mice with considerably higher levels in the ATF3-cKO mice consistent with the finding that ATF3-cKO mice displayhigher level of fibrosis (Figure 3A)
Cardiac remodeling processes and diabetes are typically associatedwith increased inflammation We therefore measured several inflamma-tory markers in the hearts of NC- and HFD-fed ATF3ff and ATF3-cKOmice IL-6 expression was highly induced in the hearts of NC- and HFD-fed ATF3-cKO mice as compared with ATF3ff mice Significantly HFD-fed ATF3-cKO mice displayed the highest level (Figure 4A) SimilarlyTNFa was expressed at significantly higher levels in HFD-fed ATF3-cKOmice (Figure 4A) To identify the source of IL-6 and TNFa expression wehave isolated cardiomyocytes from mice fed with HFD for 15 weeks andexamined the cytokinersquos transcription and secretion levels by both qRT-PCR and ELISA respectively Indeed the transcription (Figure 4B) and
secretion (Figure 4C) levels of IL-6 and TNFa were significantly higher incardiomyocytes derived from HFD-fed ATF3-cKO mice as compared totheir ATF3ff cardiomyocytes counterparts To reveal whether theinflammatory response in the heart following a HFD affects circulatingIL-6 and TNFa levels we measured the cytokines in the serum of HFD-fed mice using ELISA Indeed a significant increase in IL-6 and TNFa lev-els were detected in the serum of ATF3-cKO mice (Figure 4D and Erespectively) Collectively we conclude that cardiomyocytes are thecells responsible at least in part for the expression and secretion of IL-6and TNFa in HFD-fed mice with significantly higher circulating levels inATF3 deficient mice
Lastly since inflammation is often associated with a maladaptive meta-bolic response we analysed the expression levels of various metabolicgenes in the heart The level of IRS1 a gene involved in insulin signallingwas significantly reduced in HFD-fed mice The reduction was more pro-nounced in the ATF3-cKO than in control mice (Figure 5A) Similarly thelevel of fatty acid translocase (CD36) was significantly higher in HFD-fedATF3ff mice IRS2 was much higher in the NC-fed ATF3-cKO mice thanin the control counterparts (NC-fed ATF3ff mice) consistent with ourprevious data that its gene expression is suppressed by ATF337
Interestingly IRS2 is induced in HFD-fed ATF3ff mice to comparable levelas HFD-fed ATF3-cKO mice The biological implication of this finding isnot clear One possibility is that induction of IRS2 a pro-survival gene bya HFD is an attempt by the cardiac cells to cope with the HFD stress thisadaptation is not necessary in the ATF3-cKO heart since IRS2 levels are
A
B
C
D
E
F
Figure 2 HFD-fed ATF3-cKO mice display increased expression of cardiac remodeling markers ATF3ff and ATF3-cKO mice were fed with either normalchow (NC) for 25 weeks or switched to a 15-week HFD regimen at 10 weeks of age At endpoint mice were sacrificed and hearts were excised (A) mRNAwas extracted from ventricles and the expression level of ATF3 and the indicated remodeling markers were measured by qRT-PCR Expression levels arepresented as relative values (compared to NC-fed ATF3ff mice determined as 1 for each gene) The results represent the meansplusmn SD (n = 6ndash8group) (B)Immuno-histochemistry analysis of paraformaldehyde fixed heart slices as described in A Representative nuclear staining is indicated by yellow arrowsScale bar = 50lm (C) Western-blot analysis of heart cell lysate probed with anti-ATF3 antibodies The expression level of a-tubulin was used as loadingcontrol (D) Quantification of ATF3 expression from C The results represent the meansplusmn SD (n = 3ndash5group) ratio of ATF3 expression of the indicatedgenotypes relative to NC ATF3ff determined as 1 (E) Ventricles sections were stained with TRITC-labelled wheat germ agglutinin and the cell size was ana-lysed Scale bar = 20lm (F) Quantification of cell size from E The results represent the meansplusmn SD (n = 6ndash8group) P lt_ 005 difference between dietsdaggerP lt_ 005 difference between genotypes
ATF3 expression in cardiomyocytes preserves homeostasis 139D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
already high The expression levels of other lipid and glycolytic regulatorssuch as peroxisome proliferator-activated receptor alpha (PPARa) carni-tine palmitoyltransferase 1 and 2 (CPT12) and pyruvate dehydrogenasekinase 4 (PDK4) were all elevated in HFD-fed mice with no significant dif-ference between the genotypes (Figure 5A) Lastly the level of the glucosetransporter 4 (Glut4) were unaffected by both diet and genotype at themRNA level (Figure 5A) However Glut4 was significantly elevated at theprotein level in HFD-fed mice with no significant difference between thegenotypes (see Supplementary material online Figure S4A and B)Collectively the expression of key metabolic genes was unalteredbetween the various genotypes in response to HFD In view of thechanges observed in metabolic genes in cardiomyocytes treated with pal-mitate (Figure 1A) this result was unexpected Thus we hypothesizedthat ATF3-dependent alterations in metabolic gene expression in cardio-myocytes may be masked by non-cardiomyocytes cells in the heartTherefore we studied the metabolic gene expression changes in isolatedcardiomyocytes derived from HFD- and NCndashfed mice from both ATF3ff
and ATF3-cKO mice (Figure 5B) Indeed the expression of all metabolicgenes tested was augmented in cardiomyocytes derived from HFD-fed ascompared with NC-fed mice Importantly the expression level of CD36CPT1 and CPT2 were significantly upregulated in HFD-fed ATF3-cKOmice as compared with their ATF3ff counterparts (Figure 5B) In contrastthe induction of both IRS1 and IRS2 were significantly lower in cardio-myocytes derived from HFD-fed ATF3-cKO mice (Figure 5B)Collectively the deleterious effects of HFD on cardiac remodeling fibro-sis and inflammation are more pronounced in ATF3-cKO mice suggest-ing that cardiac ATF3 plays a protective role in the HFD paradigm
35 HFD-induced cardiac dysfunctionNext we characterized the effect of HFD and cardiac remodeling oncardiac structure and function in ATF3ff and ATF3-cKO mice by
echocardiography Mice fed with NC had normal heart dimensions andfractional shortening (FS) (Figure 6A and B) HFD significantly reduced FSin both genotypes of HFD-fed mice as compared to corresponding NC-fed mice (Figure 6B) However the reduction was significantly more pro-nounced in the ATF3-cKO mice suggesting deteriorated systolic func-tion in these mice (Figure 6B) The reduced FS in the HFD-fed ATF3-cKO mice seems to be largely driven by an increase in LV internal end-systolic (LVIDs) dimension (Figure 6A) Heart rate was slightly increasedin the HFD-fed mice however this increase cannot explain the cardiacdysfunction observed in ATF3-cKO mice (Figure 6B) In addition weused cardiac MRI the gold standard for LV function and mass asses-sment38 LV mass was significantly higher in the ATF3-cKO mice follow-ing a 15-week HFD regimen (Figure 6C) In addition end-diastolic andend-systolic volumes were significantly larger in ATF3-cKO mice as com-pared to ATF3ff mice Moreover ATF3-cKO mice displayed a signifi-cantly lower ejection fraction as compared to ATF3ff mice (Figure 6D)These results are consistent with the reduced cardiac function found inthe echocardiographic analysis Images of mid-ventricular short-axis(Figure 6E) and long-axis (Figure 6E) slices are demonstrating the cardiaceccentric pattern observed in HFD-fed ATF3-cKO mice Collectivelythe results from the MRI show a greater increase in LV mass and volumein the ATF3-cKO mice as compared to the ATF3ff mice following aHFD suggesting fundamental structural differences in the heart betweenthe ATF3-cKO and ATF3ff mice
36 Impaired glucose tolerance in HFDATF3-cKO miceLastly we measured fasting glucose levels to assess the diabetic state ofthe mice As expected higher glucose levels were observed in HFD-fedmice as compared to NC-fed counterparts Interestingly the
A B
C
D
E
Figure 3 HFD-fed ATF3-cKO mice have higher level of fibrosis in the heart ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) mRNA wasextracted from ventricles and the expression levels of the indicated fibrosis markers were measured by qRT-PCR The expression levels are presented asrelative values (compared to NC-fed ATF3ff mice determined as 1 for each gene n = 6ndash8group) (B) Western-blot analysis of heart cell lysate probed withanti-a-SMA antibody The expression level of a-tubulin was used as loading control (C) Quantification of aSMA expression from B The results representthe meansplusmn SD (n= 3ndash5group) ratio of aSMA expression of the indicated genotypes relative to NC ATF3ff determined as 1 (D) Paraffin-embedded heartsections were stained with Massonrsquos trichrome to visualize fibrosis Scale bar = 100 lm (E) The level of fibrosis () stained by Massonrsquos trichrome was quan-tified and the results represent the meansplusmn SD (n = 6ndash8group) P lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes
140 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
hyperglycemia state of the ATF3-cKO mice was significantly higher thanin the ATF3ff mice following 15 weeks of HFD (Figure 7A) Consistentlythe glucose tolerance test (GTT) revealed that HFD-fed ATF3-cKOmice displayed higher blood glucose levels and longer glucose clearancetime when compared to HFD-fed ATF3ff whereas NC-fed ATF3ff andATF3-cKO mice had normal glucose clearance (Figure 7B)
Interestingly whole body ATF3-KO mice fed with HFD also displayedimpaired glucose clearance (see Supplementary material onlineFigure S5A)39 Previously we reported that decreased glucose tolerancein whole body ATF3-KO mice is at least in part due to reduced insulingene expression in b-cells (see Supplementary material online FigureS5B)39 To determine insulin level in HFD-fed ATF3-cKO mice weanalysed blood insulin levels by ELISA In both HFD-fed ATF3ff andATF3-cKO mice the serum insulin levels were significantly higher ascompared to their NC-fed counterpart mice with the ATF3-cKO micedisplaying the highest insulin levels (Figure 7C) Together these resultssuggest that peripheral tissues of ATF3-cKO mice have reduced ability
to uptake glucose despite their high insulin level in the blood Next wesought to measure the protein levels of components along the insulinpathway Akt and Foxo340 Both Akt and Foxo3 protein levels were sim-ilar in HFD-fed mice independent of the genotype (see Supplementarymaterial online Figure S4A and C) Interestingly phosphorylation of Akt(Ser473) which indicates the activity of the insulin pathway was slightlyelevated in HFD-fed mice however this did not reach statistical signifi-cance and was similar in both of the genotypes (see Supplementary material online Figure S4D) To measure the activity of the insulin signallingpathway following insulin stimulation we used the ratio between phos-phorylated Akt (Ser473) and total Akt expression HFD-fed mice wereinjected with insulin (10 Ukg) and the heart liver and skeletal musclewere harvested 15 min after the injection (Figure 7D and E and seeSupplementary material online Figure S6AndashD) Insulin injection resultedin a significant increase of pAktAkt ratio in the hearts of ATF3ff micewhile no significant change was observed in Akt phosphorylation inATF3-cKO mice (Figure 7D and E) Similarly low Akt phosphorylation
A
D E
B C
Figure 4 HFD-fed ATF3-cKO mice display inflammatory response in the heart (A) ATF3ff and ATF3-cKO mice were fed as described in Figure 2 mRNAwas extracted from ventricles and the expression levels of the indicated inflammatory markers were measured by qRT-PCR Expression levels are presentedas relative values (compared to NC-fed ATF3ff mice determined as 1 for each gene n= 6ndash8group) (B) Mice were fed as described in Figure 2Cardiomyocytes were isolated and primary cultures were generated mRNA was extracted from an overnight culture and the expression levels of IL-6 andTNFa were studied by qRT-PCR The mRNA level obtained in cardiomyocytes derived from NC-fed ATF3ff mice was determined as 1 and the relativeexpression of all other treatment is shown (n = 5ndash6 independent cardiomyocyte preparations) (C) Conditioned media was collected from cardiomyocytesdescribed in B and ELISA assay for the indicated cytokines is shown Cytokines level is presented as relative values to NC-fed ATF3ff mice determined as 1for each cytokine (n = 5ndash6 independent preparations) (D and E) Blood was collected from HFD-fed mice and IL-6 and TNFa serum levels were quantifiedby ELISA (n = 8ndash12group) The results represent the meansplusmn SD P lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes
ATF3 expression in cardiomyocytes preserves homeostasis 141D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
levels were also observed in both the liver and skeletal muscle of theATF3-cKO mice however this did not reach statistically significance (seeSupplementary material online Figure S6AndashD) Consistently palmitate-treated cardiomyocytes derived from ATF3-cKO mice displayedreduced phosphorylation of Akt following 30-min insulin stimulation(see Supplementary material online Figure S6E)
Collectively our findings revealed a surprising phenomenon that car-diac ATF3 deficiency results in increased cardiac remodeling leading topotentiation of local and peripheral insulin resistance
4 Discussion
A HFD is a commonly used regimen to induce T2D The majority of theliterature focuses on diabetic symptoms without addressing the issue ofT2D-induced cardiomyopathy Even when it is addressed there is noclear consensus regarding the extent of cardiac effects imposed byHFD41 Diet-induced obesity in mouse models is associated with globalinsulin resistance and increased myocardial fatty acid utilization how-ever the reported phenotypes on cardiac function vary from extensivehypertrophy and heart failure4 to lack of cardiac dysfunction4243 Thesevariations have been attributed to diet regimens andor sensitivity ofmouse strains to a HFD In this report we present evidence that cardiacATF3 deficiency exacerbates the deleterious effect of a HFD on theheart such as maladaptive cardiac remodeling fibrosis inflammation andcardiac dysfunction Our work may provide a potential molecular insightto variations in the literature regarding the impact of HFD on the heartAs ATF3 is a stress-inducible gene we propose that its proper inductionin the cardiomyocytes is protective and helps the heart to cope withHFD stress It is possible that the induction of ATF3mdashits expressionlevel duration and kineticsmdashvaries in different HFD regimens andormouse strains contributing to the variations observed in the literature
HFD regimen induced a significant higher level of fibrosis in the heartof ATF3-cKO mice We have identified TIMP1 as a possible mechanismto explain the increase in fibrosis in ATF3-cKO cardiomyocytes Severallines of evidence suggest that TIMP1 is possibly an ATF3 target gene
First we observed an increase in TIMP1 transcription in the whole heartand in isolated cardiomyocytes (Figures 1A and 3A) Second TIMP 1 3and 4 were identified as significantly highly expressed genes in the heartof ATF3-KO mice as compared with wild type mice following aorticbanding20 Third a functional AP-1 regulatory element within the TIMP1promoter was described44 Indeed preliminary results in our lab suggestthat ATF3 downregulates the TIMP1 promoter (data not shown) Thusit is possible that TIMP1 is suppressed by ATF3 and therefore results in anet increase in MMPs activity that helps to maintain ECM formationdeg-radation homeostasis
Our findings are consistent with previous studies demonstrating that adeficiency in ATF3 can promote cardiac maladaptive changes which leadto cardiac dilatation and hypertrophy in an aortic banding model1920 Incontrast a different study using a pressure overload model reportedthat ATF3 induces a maladaptive cardiac response17 These seeminglyconflicting data may be explained by the different patterns of ATF3expression induced by the different models In the pressure overloadmodel the acute and transient expression of ATF3 in cardiomyocytesinitiates a cascade of maladaptive changes17 whereas in the aortic band-ing model and the HFD model described here ATF3 expression is longlasting
In this context this is the first report using mice with cardiac-selectivedeficiency of ATF3 (ATF3-cKO) Previous reports on the roles of ATF3in cardiac function were carried out using whole body knockout ofATF325 or transgenic mice ectopically expressing ATF3 in cardiomyo-cytes16 A key advancement of ATF3-cKO mice over the whole bodyknockout mice is that they provide insights into the function of ATF3selectively in cardiomyocytes without the complication of ATF3 defi-ciency in other cells Thus the ATF3-cKO mouse is a useful tool to bet-ter understand the function of cardiac ATF3
Consistent with our previous report25 here we showed that cardiacATF3 deficiency has a more severe HFD-induced diabetic phenotypethan the control wild type mice However while reduced insulin genetranscription in ATF3-deficient b-cells was observed25 our current dataindicates that a lack of insulin is not the explanation for the glucose intol-erance found in the HFD-fed ATF3-cKO mice In contrast these mice
A B
Figure 5 HFD-fed ATF3-cKO mice show metabolic response in the heart ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) mRNA wasextracted from ventricles and the expression levels of the indicated metabolic markers were measured by qRT-PCR Expression levels are presented as rela-tive values (compared to NC-fed ATF3ff mice determined as 1 for each gene n= 6ndash8group) (B) Cardiomyocytes were isolated and plated on laminin for1 h followed by mRNA preparation (n = 4ndash5 independent preparations) The expression levels of the indicated metabolic markers were measured by qRT-PCR Expression levels are presented as relative values (to NC-fed ATF3ff mice determined as 1 for each gene) The results represent the meansplusmn SDP lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes
142 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
had higher blood insulin level (Figure 7C) Therefore upon HFD chal-lenge cardiac-deficiency of ATF3 exacerbates HFD-induced T2D Thislsquoperipheral effectrsquo is surprising since ATF3 is selectively deleted in thecardiomyocytes
The question then is how can we explain the effect of cardiacATF3 deficiency on peripheral glucose intolerance One possibility isthe reduced IRS1 expression in ATF3-deficient cardiomyocytes bothin vivo and in vitro IRS1 suppression may lead to impaired insulin sig-nalling in the heart thereby exacerbating glucose intolerance Indeedwe observed that HFD-fed ATF3-cKO mice displayed insulin resist-ance upon insulin challenge when compared to HFD-fed ATF3ff
mice In addition palmitate-treated cardiomyocytes lacking ATF3 dis-played lower insulin signalling as compared to their wild type coun-terparts Since the heart is only a minor glucose consuming organ incomparison to skeletal muscle cells or the brain the peripheral sys-temic effect cannot be solely based on this observation A case inpoint transgenic mice with insulin receptor (IR) deficiency in theheart do not display a diabetic phenotype4546 Another attractive
possibility for the peripheral effect of cardiac ATF3-deletion is itsimpact on inflammation It is well established that low-grade inflam-mation years in advance of the disease onset is involved in the patho-genic processes causing T2D8 Mediators of inflammation such asTNFa interleukin (IL)-1b the IL-6 family of cytokines IL-18 and cer-tain chemokines have been proposed to be involved in the eventscausing both forms of diabetes (T1D and T2D)8 Interestingly wehave shown that IL-6 a well-known ATF3 target gene47 is known toenhance its transcription and secretion of cytokines such as IL-6 andTNFa in palmitate-treated primary cultures of ATF3-cKO cardio-myocytes as well as in cardiomyocytes derived from HFD-fed ATF3-cKO mice IL-6 has been proposed to play a role in glucose homeo-stasis and metabolism directly and indirectly via its action on skeletalmuscle cells adipocytes hepatocytes pancreatic b-cells and neuro-endocrine cells48
Therefore we hypothesize that elevated levels of inflammatory cyto-kines in ATF3-cKO mice mediate a cross-talk in peripheral tissues leadingto impaired glucose tolerance Although circulating IL-6 and TNFa
A
B
C
D
E
F
Figure 6 HFD-fed ATF3-cKO mice present deteriorated heart function ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) At endpointmice hearts were examined by micro ultrasound The following parameters were measured interventricular septal end diastole (IVSd) left ventricular pos-terior wall end diastole (LVPWd) maximal left ventricular internal end-diastole (LVIDd) end-systole (LVIDs) and heart rate The results represent themeansplusmn SD (n = 6ndash8 in each group as shown in panel B) (B) Heart function was assessed by calculating percentage of fractional shortening (FS) accordingto FS () = [(LVDd-LVDs)LVDd]100 The results represent the meansplusmn SD (n = 6ndash8 in each group) (C and D) Left ventricular cardiac volumes massand function were examined by a cardiac MRI The following parameters were measured left ventricular end-diastolic (LVEDV) and left ventricular end-sys-tolic volume (LVESV) left ventricular (LV) mass and ejection fraction (EF) The results represent the meansplusmn SD (n = 7 in each group) (E and F)Representative images of mid-ventricular short-axis (E) and long-axis (F) slice at peak diastole and systole P lt_ 005 difference between diets daggerP lt_ 005 dif-ference between genotypes
ATF3 expression in cardiomyocytes preserves homeostasis 143D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
cytokines may be derived from different tissues we provide ample evi-dence that ATF3-defficient cardiomyocytes produce and secrete higherlevels of these cytokines as compared to their wild type counterpartsThis indicates a lsquocardiac-to-peripheralrsquo effect in T2D-induced cardiomyop-athy This is a reverse direction from the lsquoperipheral-to-cardiacrsquo effect dis-cussed in the introduction where inflammation in the peripheral tissuesleads to cardiac dysfunction Since inflammatory cytokines are soluble
factors they can exert a systemic effect This is why it makes sense thattheir effect can go both ways Thus diabetes and cardiomyopathy are apart of a vicious cycle Previously it was shown that cardiac specific dele-tion of the thyroid hormone complex mediator MED13 results in aperipheral effect upon HFD Cardiac MED13 deficiency suppresses meta-bolic genes in the heart which leads to a decrease in energy expenditureand an obese phenotype most likely by the secretion of cardiokines that
A
B
C
D
E
Figure 7 HFD-fed ATF3-cKO mice exhibit impaired glucose tolerance ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) Fasting (6 h)blood glucose concentrations (mgdl) The results represent the meansplusmn SD (n = 20ndash30group) P lt_ 005 difference between diets daggerP lt_ 005 differencebetween genotypes (B) Glucose tolerance test (GTT) Values are presented as relative to initial valueplusmn SD (n = 20ndash25group) P lt_ 005 difference betweendiets daggerP lt_ 005 difference between genotypes (C) Serum insulin concentrations were quantified by ELISA Values are presented as relative values (com-pared to NC-fed ATF3ff mice determined as 1 n = 15ndash20group) P lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes (D and E)Effect of HFD on phosphorylated Akt (Ser473 pAkt) to Akt ratio levels in heart lysate following either saline or insulin injection (10 U1 kg) is shownRepresentative blots (D) Densitometric analysis is presented as means ratio (n = 3ndash6group) pAktAkt ratio is presented as relative values (compared toHFD-fed saline injected ATF3ff mice determined as 1) P lt_ 005 insulin vs saline treatment daggerP lt_ 005 difference between genotypes (E)
144 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
mediates the cross talk between the heart and peripheral tissues49ndash51
Here we also observed increased levels of natriuretic peptides (ANP andBNP) which together with IL-6 and TNFa may mediate the cross-talk withthe peripheral tissues to induce glucose intolerance
Does this finding have any relevance to cardiac disease in humansSeveral lines of evidence suggests that congestive heart failure in patientsis associated with an increase in blood glucose levels52 In addition highblood glucose levels in non-diabetic patients with acute myocardialinfarction serves as a prognostic predictor of long-term mortality5354
Thus here we provide evidence that high blood glucose levels are a partof the symptoms of cardiac remodeling process
Collectively our findings suggest that cardiac ATF3 expression isresponsible for an adaptive cardiac response to the stressful environmentunder a HFD (Figure 8) ATF3 dampens the inflammatory response andsuppresses the expression of ECM remodeling genes and thus amelioratescardiac function In HFD-fed mice with ATF3 deficiency or in mice withcardiac remodeling the inflamed heart affects also peripheral tissuesthereby worsening T2D in peripheral tissues resulting in elevated bloodglucose level and turning on a vicious cycle to worsen cardiomyopathy
Supplementary material
Supplementary material is available at Cardiovascular Research online
AcknowledgementsThe authors wish to thank Drs Keren Bitton-Worms and Ran Rostokerand Prof Derek LeRoith from the Rambam Medical Center for their
assistance in GTT and ITT Dr Edith Suss Toby Maya Holdengreber andLior Liba from the BCF Bioimaging Center (Faculty of MedicineTechnion) for their assistance with imaging and image analysis Prof DEMichelle from University of Michigan for cardiomyocytes culture techni-que and Prof Yehiel Zick from the Weizmann institute of Sciences forcritical comments
Conflict of interest none declared
FundingThis work was partially supported by the United States-Israel BinationalGrant 2009179 to AA and TH In addition we wish to thank the MarsPittsburgh Fund for Medical Research to AA the Department of Defensegrant W81XWH-14-1-0179 to TH and to D Dan and Betty Kahn travelgrant support to LK
References
1 Roger VL Go AS Lloyd-Jones DM Benjamin EJ Berry JD Borden WB Bravata DMDai S Ford ES Fox CS Fullerton HJ Gillespie C Hailpern SM Heit JA Howard VJKissela BM Kittner SJ Lackland DT Lichtman JH Lisabeth LD Makuc DM MarcusGM Marelli A Matchar DB Moy CS Mozaffarian D Mussolino ME Nichol GPaynter NP Soliman EZ Sorlie PD Sotoodehnia N Turan TN Virani SS Wong NDWoo D Turner MB American Heart Association Statistics C Stroke Statistics SHeart disease and stroke statisticsndash2012 update a report from the American HeartAssociation Circulation 2012125e2ndashe220
2 Wilson C Diabetes ACCORD 5-year outcomes of intensive glycemic control NatRev Endocrinol 20117314
3 Ernande L Derumeaux G Diabetic cardiomyopathy myth or reality Arch CardiovascDis 2012105218ndash225
4 Battiprolu PK Hojayev B Jiang N Wang ZV Luo X Iglewski M Shelton JM GerardRD Rothermel BA Gillette TG Lavandero S Hill JA Metabolic stress-induced
InsulinGF
Cytokines
FFA
GlucoseATF3
Cardiac
Remodeling
Factors
Paracrine action
Collagens
and ECM
proteins
Fibrosis
Cardiac
Dysfunction
Systemic Effects
Glucose Tolerance
TNFα
IL-6
Cardiomyocyte
ATF3
Homeostasis
Homeostasis
TNFα
IL-6
TIMP1
BNP
ANP
Figure 8 lsquoPeripheral-to-cardiacrsquo and lsquoCardiac-to-peripheralrsquo model scheme Schematic diagram showing the ATF3 adaptive role in maintaining hearthomeostasis following HFD-induced T2D In contrast stressful environment under HFD leads to maladaptive cardiac response in the absence of ATF3Cardiac remodeling caused by ATF3 deficiency induces an increase in cardiac ECM remodeling and inflammatory genes and mediates the communication ofcardiomyocytes with heart resident cells potentially mediated by IL-6 TNFa ANP BNP and TIMP1 In this situation increased cardiac fibrosis and inflamma-tory response worsen cardiac function The inflamed failing heart mediates the communication of cardiomyocytes with distant organs as well thereby affect-ing diabetic outcome and glucose homeostasis
ATF3 expression in cardiomyocytes preserves homeostasis 145D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
activation of FoxO1 triggers diabetic cardiomyopathy in mice J Clin Invest20121221109ndash1118
5 Mandavia CH Aroor AR Demarco VG Sowers JR Molecular and metabolic mecha-nisms of cardiac dysfunction in diabetes Life Sci 201392601ndash608
6 Ventura-Clapier R Garnier A Veksler V Energy metabolism in heart failure J Physiol20045551ndash13
7 Guo S Insulin signaling resistance and the metabolic syndrome insights from mousemodels into disease mechanisms J Endocrinol 2014220T1ndashT23
8 Montane J Cadavez L Novials A Stress and the inflammatory process a major causeof pancreatic cell death in type 2 diabetes Diabetes Metab Syndr Obes 2014725ndash34
9 Ashrafian H Frenneaux MP Opie LH Metabolic mechanisms in heart failureCirculation 2007116434ndash448
10 Tanabe K Liu Y Hasan SD Martinez SC Cras-Meneur C Welling CM Bernal-MizrachiE Tanizawa Y Rhodes CJ Zmuda E Hai T Abumrad NA Permutt MA Glucose andfatty acids synergize to promote B-cell apoptosis through activation of glycogen syn-thase kinase 3beta independent of JNK activation PLoS One 20116e18146
11 Heineke J Molkentin JD Regulation of cardiac hypertrophy by intracellular signallingpathways Nat Rev Mol Cell Biol 20067589ndash600
12 Hai T Wolford CC Chang YS ATF3 a hub of the cellular adaptive-response net-work in the pathogenesis of diseases is modulation of inflammation a unifying com-ponent Gene Expr 2010151ndash11
13 Takii R Inouye S Fujimoto M Nakamura T Shinkawa T Prakasam R Tan K HayashidaN Ichikawa H Hai T Nakai A Heat shock transcription factor 1 inhibits expression ofIL-6 through activating transcription factor 3 J Immunol 20101841041ndash1048
14 Giraldo A Barrett OP Tindall MJ Fuller SJ Amirak E Bhattacharya BS Sugden PHClerk A Feedback regulation by Atf3 in the endothelin-1-responsive transcriptomeof cardiomyocytes Egr1 is a principal Atf3 target Biochem J 2012444343ndash355
15 Hasin T Elhanani O Abassi Z Hai T Aronheim A Angiotensin II signaling up-regulates the immediate early transcription factor ATF3 in the left but not the rightatrium Basic Res Cardiol 2011106175ndash187
16 Koren L Elhanani O Kehat I Hai T Aronheim A Adult cardiac expression of theactivating transcription factor 3 ATF3 promotes ventricular hypertrophy PLoS One20138e68396
17 Koren L Alishekevitz D Elhanani O Nevelsky A Hai T Kehat I Shaked Y AronheimA ATF3-dependent cross-talk between cardiomyocytes and macrophages promotescardiac maladaptive remodeling Int J Cardiol 2015198232ndash240
18 Okamoto Y Chaves A Chen J Kelley R Jones K Weed HG Gardner KL Gangi LYamaguchi M Klomkleaw W Nakayama T Hamlin RL Carnes C Altschuld R BauerJ Hai T Transgenic mice with cardiac-specific expression of activating transcriptionfactor 3 a stress-inducible gene have conduction abnormalities and contractile dys-function Am J Pathol 2001159639ndash650
19 Zhou H Shen DF Bian ZY Zong J Deng W Zhang Y Guo YY Li H Tang QZActivating transcription factor 3 deficiency promotes cardiac hypertrophy dysfunc-tion and fibrosis induced by pressure overload PLoS One 20116e26744
20 Zhou H Guo H Zong J Dai J Yuan Y Bian ZY Tang QZ ATF3 regulates multipletargets and may play a dual role in cardiac hypertrophy and injury Int J Cardiol2014174838ndash839
21 Yang J Yang CJ Yang J Fan ZX ATF3 a promotion effect or a inhibition effect in car-diac maladaptive remodeling Int J Cardiol 2015201245ndash246
22 Koren L Shaked Y Aronheim A Response letter ldquoATF3 a promoter or inhibitor ofcardiac maladaptive remodelingrdquo Int J Cardiol 2015201692
23 Ma ZG Wei WY Xu SC Zhang WB Dai J Tang QZ ATF3 a potential target forcardiac maladaptive remodeling Int J Cardiol 201620250ndash51
24 Zhou H Yuan Y Ni J Guo H Deng W Bian ZY Tang QZ Pleiotropic and puzzlingeffects of ATF3 in maladaptive cardiac remodeling Int J Cardiol 201620687ndash88
25 Zmuda EJ Qi L Zhu MX Mirmira RG Montminy MR Hai T The roles of ATF3 anadaptive-response gene in high-fat-diet-induced diabetes and pancreatic beta-celldysfunction Mol Endocrinol 2010241423ndash1433
26 Lu D Chen J Hai T The regulation of ATF3 gene expression by mitogen-activatedprotein kinases Biochem J 2007401559ndash567
27 Hartman MG Lu D Kim ML Kociba GJ Shukri T Buteau J Wang X Frankel WLGuttridge D Prentki M Grey ST Ron D Hai T Role for activating transcription fac-tor 3 in stress-induced beta-cell apoptosis Mol Cell Biol 2004245721ndash5732
28 Wolford CC McConoughey SJ Jalgaonkar SP Leon M Merchant AS Dominick JLYin X Chang Y Zmuda EJ OrsquoToole SA Millar EK Roller SL Shapiro CL OstrowskiMC Sutherland RL Hai T Transcription factor ATF3 links host adaptive response tobreast cancer metastasis J Clin Iinvest 20131232893ndash2906
29 Agah R Frenkel PA French BA Michael LH Overbeek PA Schneider MD Generecombination in postmitotic cells Targeted expression of Cre recombinase pro-vokes cardiac-restricted site-specific rearrangement in adult ventricular musclein vivo J Clin Invest 1997100169ndash179
30 Livet J Weissman TA Kang H Draft RW Lu J Bennis RA Sanes JR Lichtman JWTransgenic strategies for combinatorial expression of fluorescent proteins in thenervous system Nature 200745056ndash62
31 Davis J Maillet M Miano JM Molkentin JD Lost in transgenesis a userrsquos guidefor genetically manipulating the mouse in cardiac research Circ Res2012111761ndash777
32 Heiberg E Sjogren J Ugander M Carlsson M Engblom H Arheden H Design andvalidation of segment ndash freely available software for cardiovascular image analysisBMC Med Imaging 2010101
33 Kabaeva Z Zhao M Michele DE Blebbistatin extends culture life of adult mouse car-diac myocytes and allows efficient and stable transgene expression Am J Physiol HeartC 2008294H1667ndash1674
34 Subramaniam A Jones WK Gulick J Wert S Neumann J Robbins J Tissue-specificregulation of the alpha-myosin heavy chain gene promoter in transgenic mice J BiolChem 199126624613ndash24620
35 Cunha DA Hekerman P Ladriere L Bazarra-Castro A Ortis F Wakeham MCMoore F Rasschaert J Cardozo AK Bellomo E Overbergh L Mathieu C Lupi R HaiT Herchuelz A Marchetti P Rutter GA Eizirik DL Cnop M Initiation and executionof lipotoxic ER stress in pancreatic beta-cells J Cell Sci 20081212308ndash2318
36 Wynn TA Ramalingam TR Mechanisms of fibrosis therapeutic translation forfibrotic disease Nat Med 2012181028ndash1040
37 Li D Yin X Zmuda EJ Wolford CC Dong X White MF Hai T The repression ofIRS2 gene by ATF3 a stress-inducible gene contributes to pancreatic beta-cell apop-tosis Diabetes 200857635ndash644
38 Myerson SG Bellenger NG Pennell DJ Assessment of left ventricular mass by cardi-ovascular magnetic resonance Hypertension 200239750ndash755
39 Zmuda EJ Viapiano M Grey ST Hadley G Garcia-Ocana A Hai T Deficiency ofAtf3 an adaptive-response gene protects islets and ameliorates inflammation in asyngeneic mouse transplantation model Diabetologia 2010531438ndash1450
40 Riehle C Abel ED Insulin signaling and heart failure Circ Res 20161181151ndash116941 Schreyer SA Wilson DL LeBoeuf RC C57BL6 mice fed high fat diets as models for
diabetes-accelerated atherosclerosis Atherosclerosis 199813617ndash2442 Brainard RE Watson LJ Demartino AM Brittian KR Readnower RD Boakye AA
Zhang D Hoetker JD Bhatnagar A Baba SP Jones SP High fat feeding in mice isinsufficient to induce cardiac dysfunction and does not exacerbate heart failure PLoSOne 20138e83174
43 Gupte AA Minze LJ Reyes M Ren Y Wang X Brunner G Ghosn M Cordero-Reyes AM Ding K Pratico D Morrisett J Shi ZZ Hamilton DJ Lyon CJ Hsueh WAHigh-fat feeding-induced hyperinsulinemia increases cardiac glucose uptake and mito-chondrial function despite peripheral insulin resistance Endocrinology20131542650ndash2662
44 Logan SK Garabedian MJ Campbell CE Werb Z Synergistic transcriptionalactivation of the tissue inhibitor of metalloproteinases-1 promoter via functionalinteraction of AP-1 and Ets-1 transcription factors J Biol Chem1996271774ndash782
45 Belke DD Betuing S Tuttle MJ Graveleau C Young ME Pham M Zhang DCooksey RC McClain DA Litwin SE Taegtmeyer H Severson D Kahn CR AbelED Insulin signaling coordinately regulates cardiac size metabolism and contractileprotein isoform expression J Clin Invest 2002109629ndash639
46 Fernandez AM Kim JK Yakar S Dupont J Hernandez-Sanchez C Castle AL FilmoreJ Shulman GI Le Roith D Functional inactivation of the IGF-I and insulin receptors inskeletal muscle causes type 2 diabetes Genes Dev 2001151926ndash1934
47 Gilchrist M Thorsson V Li B Rust AG Korb M Roach JC Kennedy K Hai TBolouri H Aderem A Systems biology approaches identify ATF3 as a negative regu-lator of Toll-like receptor 4 Nature 2006441173ndash178
48 Kristiansen OP Mandrup-Poulsen T Interleukin-6 and diabetes the good the bad orthe indifferent Diabetes 200554(Suppl 2)S114ndashS124
49 Grueter CE van Rooij E Johnson BA DeLeon SM Sutherland LB Qi X Gautron LElmquist JK Bassel-Duby R Olson EN A cardiac microRNA governs systemic energyhomeostasis by regulation of MED13 Cell 2012149671ndash683
50 Baskin KK Grueter CE Kusminski CM Holland WL Bookout AL Satapati S KongYM Burgess SC Malloy CR Scherer PE Newgard CB Bassel-Duby R Olson ENMED13-dependent signaling from the heart confers leanness by enhancing metabo-lism in adipose tissue and liver EMBO Mol Med 201461610ndash1621
51 Nakamura M Sadoshima J Heart over mind metabolic control of white adipose tis-sue and liver EMBO Mol Med 201461521ndash1524
52 Tenenbaum A Fisman EZ Impaired glucose metabolism in patients with heart failurepathophysiology and possible treatment strategies Am J Cardiovasc Drugs20044269ndash280
53 Suskin N McKelvie RS Burns RJ Latini R Pericak D Probstfield J Rouleau JL SigouinC Solymoss CB Tsuyuki R White M Yusuf S Glucose and insulin abnormalitiesrelate to functional capacity in patients with congestive heart failure Eur Heart J2000211368ndash1375
54 Aronson D Hammerman H Kapeliovich MR Suleiman A Agmon Y Beyar RMarkiewicz W Suleiman M Fasting glucose in acute myocardial infarction incremen-tal value for long-term mortality and relationship with left ventricular systolic func-tion Diabetes Care 200730960ndash966
146 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022

cytokines by adipocytes resulting in whole-body insulin resistance8
Under these conditions the heart rapidly adapts by consuming primarilyfatty acids as a source of ATP generation Despite an increase in fattyacid metabolism it is likely that fatty acid uptake exceeds oxidation ratesthereby resulting in cellular lipid accumulation-induced lipotoxicity5910
At the molecular level insulin signalling activates multiple pathways tar-geting various transcription factors resulting in re-programing of adaptiveor maladaptive cardiac remodeling processes11
In this report we identify an important transcription factor that playsa major role in this process Activating Transcription Factor 3 (ATF3)ATF3 is a member of the basic leucine zipper (bZIP) superfamily of tran-scription factors and is encoded by an immediate early gene whoseexpression is low under normal condition but is rapidly and highlyinduced by multiple stressors12
ATF3 binds to the cyclic AMP response element (CRE) and typicallydampens the expression of multiple target genes Depending on the con-text ATF3-mediated response can be adaptive or maladaptive1213 Forexample the role of ATF3 in cardiac diseases varies depending on thestress paradigms Neuroendocrine hormones stimuli induce ATF3 tran-scription both in vitro14 and in vivo15 Mice with ATF3 deficiency displayreduced cardiac remodeling and hypertrophy in a phenylephrine-induced pressure overload model1617 Consistently ectopic ATF3expression in cardiomyocytes is sufficient to promote heart growth andcardiac dysfunction in transgenic mice1618 Together this suggests a del-eterious role of ATF3 in cardiac diseases In contrast studies using ATF3deficient mice in the context of transverse aortic constriction suggest acardiac protective role for ATF31920 Thus the importance of ATF3 incardiac remodeling models is well documented but its effects varydepending on the stress paradigms21ndash24
In addition to cardiac diseases ATF3 has been shown to play a role inother diseases including diabetes Previously we showed that mice defi-cient in ATF3 had reduced glucose tolerance and more severe T2D thanthe wild type mice under a high-fat diet (HFD)25 We investigated ATF3in this context since ATF3 is known to be induced by stresses that play arole in obesity-induced T2D cardiomyopathy including metabolic stressoxidative stress endoplasmic reticulum (ER) stress and inflamma-tion1226 However the role of ATF3 in diabetes-induced cardiomyop-athy has never been reported
In this article we set out to test whether cardiomyocytes-selectivedeletion of ATF3 alters HFD-induced T2D cardiomyopathy We presentevidence that following a HFD cardiac ATF3 deficiency exacerbates car-diac remodeling inflammation maladaptive metabolic response and car-diac dysfunction Surprisingly it also worsens blood glucose intoleranceincluding a reduced ability of the peripheral tissues to respond to insulinSince ATF3 deletion is selectively in cardiomyocytesmdashwhich is not a celltype known to play a role in diabetesmdashthis result is novel and intriguing
2 Methods
21 MiceThis study was carried out in strict accordance with the Guide for theCare and Use of Laboratory Animals of the National Institute of HealthIn addition our protocol was approved by the Committee of the Ethicsof Animal Experiments of the Technion All mice strains used in thisstudy were backcrossed to C57Bl6J for at least six generations Micestrains used in this study whole-body ATF3 KO (ATF3-KO)27 ATF3-flox (ATF3ff)28 aMHC-Cre29 and loxP-XFP transgenic (confetti) mice30
To generate mice with a cardiomyocyte-specific deletion of ATF3
(ATF3-cKO) we crossed mice carrying ATF3-flox alleles with transgenicmice harbouring Cre recombinase driven by a aMHC promoter Matingcages were set up to produce progeny with only one copy of aMHC-Cre to avoid variability in Cre expression31 ATF3ff and ATF3-cKO malemice littermates were housed in the same cage on a 12-h lightdark cycleand given ad libitum access to food and water All experimental groupswere fed with normal chow (NC 44 fat) for 10 weeks Subsequentlymice were either left on the NC diet or switched to high-fat diet (HFD60 fat TD06414 Harlan Inc) for additional 15 weeks
22 Glucose tolerance test fasting glucosetest and blood biochemistryThe mice fasted for 6 h and were then intraperitoneally injected with2 gkg of glucose Blood samples were obtained from their snipped tailand analysed using a portable glucometer Blood from non-fasting micewas collected from submandibular bleeds and serum insulin levels weremeasured by ELISA (Merck Millipore) Total cholesterol and high densitylipoprotein (HDL) cholesterol were measured at the Rambam ClinicalBiochemistry Laboratory
23 Insulin signallingFor insulin signalling analysis HFD-fed ATF3ff and ATF3-cKO micefasted for 6 h and were then intraperitoneally injected with insulin (10 Ukg Lantus SoloStar Sanofi-Aventis NJ) Mice were euthanized by aninjection of 01 mL of 12 mgmL Xylazine 60 mgmL Ketamine mixture
Heart liver and skeletal muscle (gastrocnemius) were harvested15 min following insulin stimulation Tissue lysates were then prepared
24 EchocardiographyMice were anesthetized with 1 of isoflurane and kept on a 37 Cheated plate throughout the analysis An echocardiography was per-formed using a Vevo2100 micro-ultrasound imaging system(VisualSonics Fujifilm) which was equipped with 13ndash38 MHz (MS 400)and 22ndash55 MHz (MS550D) linear array transducers The person per-forming echocardiography and the data analysis was blinded to the micegenotype Cardiac size shape and function were analysed by conven-tional two-dimensional imaging and M-Mode recordings Maximal leftventricles end-diastolic (LVDd) and end-systolic (LVDs) dimensionswere measured in short-axis M-mode images Fractional shortening (FS)was calculated as FS () = [(LVDd-LVDs)LVDd]100 All values werebased on the average of at least five measurements
25 MRI hardware and animal monitoringMRI was performed on a 94T bore scanner (Bruker Biospec EttlingenGermany) using a cylindrical volume coil (86 mm inner diameter) for sig-nal excitation and a single channel surface coil (20 mm diameter) for sig-nal reception The animal was put under anaesthesia with aconcentration of 05ndash15 isoflurane and supplemented with oxygen(05 Lmin) Respiration was monitored during imaging (Small AnimalInstruments Stony Brook New York NY) and body temperature wasmaintained using thermostat-regulated circulating hot water
26 Cardiac magnetic resonance (CMR)sequencesThe mice were scanned using cine FLASH sequence acquired from theself-gating technique IntraGate (Bruker BioSpin MRI EttlingenGermany) The performed CMR protocol comprised of three sets ofscans (i) Single slice Long Axis view for general evaluation of the heart
ATF3 expression in cardiomyocytes preserves homeostasis 135D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
structure and accurate planning of the subsequent scans (ii) Single slicetwo chamber Short Axis view perpendicular to LV walls at the level ofthe papillary muscles and (iii) Contiguous multi slices two chamber ShortAxis view for entire LV coverage
Cardiac MR sequences parameters
i Single slice Long Axis view 1 mm thickness FOV = 3 cm3 cm matrixdimension = 400400 spatial resolution = 7575lm2 repetitionechotime (TRTE) = 6532 ms number of repetitions = 100 10 cardiacmovie frames (cardiac phases)
ii Single slice two chamber Short Axis view 07 mm thicknessFOV=3 cm3 cm matrix dimension=400400 spatial reso-lution=7575lm2 repetitionecho time (TRTE) = 6532 ms numberof repetitions = 150 15 cardiac movie frames (cardiac phases)
iii Multi slices two chamber Short Axis view Contiguous 9ndash12 slicesdepending on heart size 07 mm thickness FOV = 3 cm3 cm matrixdimension = 400400 spatial resolution=7575lm2 repetitionechotime (TRTE) = 81-12032 ms (depending on slices number) number ofrepetitions = 120 15 cardiac movie frames (cardiac phases)
The total acquisition time for each animal was40 min
27 MRI image analysisImage analysis was performed using the freely available Segment soft-ware (Medviso AB Lund Sweden)32 Manual segmentation of the endo-cardium and epicardium of the LV was performed to measure LV massand LV functional parametersmdashend-systolic and end-diastolic (ESV andEDV respectively) volumes Ejection fraction (EF) was calculated as EF() = [(EDV-ESV)EDV]100 The person performing the MRI and thedata analysis was blinded to the mice genotype
28 Heart harvestingAt endpoint mice were anesthetized weighed and sacrificed Heartswere excised and ventricles were then divided into three pieces thatwere used for protein extraction RNA purification and histologicalanalysis
29 mRNA extractionmRNA was purified from ventricles using an Aurum total RNA fatty orfibrous tissue kit (732-6830 Bio-Rad) or from isolated cardiomyocytesusing a High Pure RNA Isolation kit (11828665001 Roche Diagnostics)according to the manufacturerrsquos instructions
210 Quantitative real-time PCR (RT-qPCR)cDNA was synthesized from 800 or 50 ng of purified mRNA derivedfrom the ventricles or isolated from the cardiomyocytes respectivelyPurified mRNA was added to a total reaction mix of high-capacity cDNAreverse transcription kit (4368814 Applied Biosystems) in a final vol-ume of 20 mL Real-time PCR was performed using Rotor-Gene6000TM (Corbett) equipment with absolute blue SYBER green ROXmix (Thermo Scientific AB-4162B) Serial dilutions of a standard samplewere included for each gene to generate a standard curve Values werenormalized to b2 microglobulin expression levels The primer sequencesare shown in Table S1 (see Supplementary material online)
211 Western-blot analysisHarvested tissues were homogenized in RIPA buffer (PBS containing 1NP-40 5 mgmL Na-deoxycholate 01 SDS) and supplemented withprotease inhibitor cocktail (P-8340 SigmandashAldrich) Homogenization
was performed at 4 C using the Bullet Blender homogenizer (BBX24Next advance) according to the manufacturerrsquos instructions as previ-ously described17
212 ImmunohistochemistryTissues were fixed in 4 formaldehyde overnight then embedded inparaffin serially sectioned at 10 lm intervals and then mounted onslides Sections were processed for deparaffinization (xylene 20 min)dehydration (isopropanol) rehydration (H2O) and antigen unmasking(10 mM sodium citrate pH-6 90 C 12 min) Immunostaining was per-formed using Histostain kit (959643 Invitrogen) according to the man-ufacturerrsquos instructions Anti-ATF3 antibodies were diluted 150Following immunostaining nuclei were stained with hematoxylin
213 AntibodiesThe primary antibodies used anti-SMA (Cat A2547) anti-a-tubulin(Cat T-9026) and anti-a-actinin (Cat A7811) which were all pur-chased from SigmandashAldrich Anti-GFP (SC-8334) was purchased fromSanta-Cruz Anti-p38 (Cat 9212) and anti-phospho-p38 (Cat 9211)were purchased from Cell signalling Anti-pAkt (Cat AF887) and anti-Akt (Cat MAB 2055) was purchased from RampD Systems anti-Foxo3(Cat 07-1719) was purchased from Merck Millipore and anti-Glut4(Cat ab654) was purchased from Abcam Lastly anti-ATF3 antibodywas generated in Aronheimrsquos lab15
214 Cell size analysisSections were stained following deparaffinization with Wheat-germagglutinin TRITC-conjugated (SigmandashAldrich Cat L5266) and diluted toa 1100 phosphate-buffered saline (PBS) Sections were washed 3 timeswith PBS and mounted in Fluoromount-G (Southern BiotechnologyBirmingham AL 0100-01) Sections were then viewed using a ZeissLSM700 confocal microscope (Thornwood NY) equipped with a 40oil objective and solid state 555nm laser
Quantification of the cell size was performed with Image Pro Plus soft-ware Five fields in each slide were photographed Unstained areas werethen identified and segmented using Image Pro Plus software In eachstained area the mean cell perimeter and area was calculated and thenumber of cells was measured
215 Fibrosis stainingHeart tissue was fixed in 4 formaldehyde overnight embedded in par-affin serially sectioned at 10 mm intervals and then mounted on slidesMassonrsquos trichrome staining was performed according to the standardprotocol Images were acquired by using Virtual Microscopy (Olympus)Stained areas were segmented and quantified following calibration usingImage Pro Plus software Five fields were quantified for each slide
216 In vitro cardiomyocyte cultureAdult cardiomyocytes were isolated from 12- to 25-week-old NC- orHFD-fed mice by a modified Langendorff perfusion apparatus asdescribed33 Isolated cells were plated in culture media (MEMmdashEagleHanksrsquo salts) supplemented with penicillin 100 UmL glutamine 2 mMNaHCO3 4 mM HEPES 10 mM bovine serum albumin (BSA) 0225 mM blebbistatin (cat B0560 SigmandashAldrich) and 5 foetal bovineserum Cardiomyocytes were then plated on 35-mm dishes coated withmouse laminin (L2020 SigmandashAldrich) at a density of 105 cellsmL inserum free medium
136 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
217 Palmitate-treated cardiomyocytesForty-five minutes after plating the medium was replaced with a culturemedium containing either 04 mM palmitate or BSA (control) Palmitatestock solution (20 mM) was diluted in 001 M NaOH by heating at 70Cfor 30 min as described10 Following overnight culture conditionedmedium was collected and cardiomyocytes were harvested for RNAextraction In addition overnight palmitate- or BSA-treated cardiomyo-cytes were incubated with 100 nM insulin (I-5500 SigmandashAldrich) for30 min and were then harvested for protein extraction
218 Enzyme-linked immuno-absorbentassayInterleukin 6 (IL-6) and tumour necrosis factor a (TNFa) levels in condi-tioned medium from an overnight culture of primary cardiomyocyteswere measured by ELISA using the DuoSet ELISA kit (RampD Systems)according to the manufacturerrsquos instructions Blood from non-fastingHFD-fed mice was collected from submandibular bleeds and serum IL-6and TNFa levels were measured by ELISA (RampD Systems)
219 StatisticsOur data is expressed as means plusmn SD The statistical test for the GTTwas performed by repeated-measures analysis of variance (ANOVA)The comparison of two means was analysed by the Studentrsquos t-test Thecomparison between several means was analysed by one-way ANOVAfollowed by Tukeyrsquos post hoc analysis All statistical analyses were per-formed using GraphPad Prism 5 software (La Jolla CA) A P value lt_ 005was accepted as statistically significant
3 Results
31 aMHC promoter specificityFirst we verified the tissue specific expression of the Cre recombinaseunder the control of the aMHC promoter by crossing aMHC-Cremouse with the loxP-XFP transgenic (confetti) mouse30 In the confettimouse lox-P specific recombination resulted in the random expressionof one out of four fluorescent proteins (BFP GFP RFP or YFP) Celllysates were prepared from various tissues of either the confettiff mouse(without Cre -) or the double transgenic mouse (containing Cre 1) fol-lowed by western-blot analysis using anti-GFP antibodies As expectedGFP-related proteins were strongly detected in the heart lysate (seeSupplementary material online Figure S1) The lysate derived from lungtissue displayed a lower level of GFP-related proteins as compared tothe heart lysate probably due to low transcription activity of the aMHCpromoter in the lungs34 GFP-related protein expression was undetect-able in all of the other tissues tested (see Supplementary material onlineFigure S1)
32 In vitro analysis of cardiomyocytestreated with free fatty acidsTo reveal the possible role of ATF3 in cardiomyopathy induced by freefatty acids (FFA) (palmitate) we analysed adult cardiomyocyte primarycultures derived from ATF3ff and ATF3-cKO mice (ATF3-cKO geno-type ATF3ffaMHC-Cre) Cardiomyocytes were isolated by theLangendorff perfusion apparatus followed by overnight treatment with04 mM palmitate10 qRT-PCR analysis showed a 4- to 5-fold increase inATF3 induction by palmitate in the ATF3ff cardiomyocytes (Figure 1A)Immuno-fluorescence analysis of cardiomyocytes treated with either 1 h
or overnight palmitate displayed a robust ATF3 nuclear staining (Figure1B) Consistent with a previous study35 western-blot analysis of the car-diomyocytes lysate following overnight palmitate treatment demon-strated a significant induction of the ATF3 protein (Figure 1C) SinceATF3 induction is dependent on the activation of the p38-MAPK signal-ling pathway1026 we measured the p38-MAPK (p38) activation stateusing anti-phospho-p38 (p-p38) While p-p38 levels were increased fol-lowing palmitate treatment p38 remained unchanged independent ofthe treatment (Figure 1C) As expected neither ATF3 transcript (Figure1A) nor ATF3 protein (Figure 1C) was detected in the palmitate-treatedATF3-cKO cardiomyocytes Importantly palmitate treatment inducedthe expression of cardiac hypertrophic markers (ANP and BNP) inflam-mation markers (IL-6 and TNFa) a metabolic marker (PDK4) and anextracellular matrix (ECM) remodeling protein (TIMP1) (Figure 1A)Strikingly significantly higher level of induction of these genes wasobserved in the ATF3-cKO cardiomyocytes as compared with ATF3ff
cardiomyocytes (Figure 1A) In addition while the expression level of theinsulin receptor substrate IRS1 was induced 2-fold by palmitate treat-ment in ATF3ff IRS1 expression was suppressed in ATF3-cKO cardio-myocytes (Figure 1A)
Next we quantified the level of secreted inflammatory cytokines byanalysing the conditioned medium derived from palmitate-treated cardi-omyocytes The levels of IL-6 and TNFa were significantly higher in theconditioned medium derived from palmitate-treated ATF3-cKO cardio-myocytes as compared to their conditioned media levels derived frompalmitate-treated ATF3ff cardiomyocytes (Figure 1D)
Collectively the above-mentioned results support an increase of ATF3expression in cardiomyocytes in response to acute and chronic exposureto FFA The analysis of cardiomyocytes derived from wild type and ATF3-cKO genotypes in vitro is consistent with a possible protective role ofATF3 by dampening the cardiomyocytes remodeling response to FFA
33 The effect of high-fat diet on wild typeand ATF3-cKO miceTo determine whether cardiac ATF3 expression plays a role in responseto a high-fat diet (HFD)-regimen we used mice harbouring acardiomyocyte-selective deletion ATF3-cKO and their wild type (ATF3f
f) counterparts Mice were fed either a normal chow (NC 44 fat) or aHFD (60 fat) The HFD was initiated at 10 weeks of age and continuedfor 15 weeks Both ATF3ff and ATF3-cKO mice fed with a HFD gainedweight at significantly higher rates than NC-fed mice with no significantdifferences between the genotypes (see Supplementary material onlineFigure S2) In addition the levels of circulating total cholesterol and highdensity lipoprotein (HDL) were significantly higher in HFD-fed mice incomparison to NC-fed mice with no significant difference between thegenotypes (see Supplementary material online Figure S3A and B)
34 Cardiomyocyte-selective ATF3deficiency induces cardiac remodelingSince ATF3 is highly induced in cardiomyocytes that are treated withFFA (04 mM palmitate) we studied the expression of ATF3 in heart tis-sue following 15 weeks of a HFD regimen by qRT-PCR ATF3 expres-sion was induced 6-fold in the hearts of HFD-fed ATF3ff mice whencompared to either the NC-fed mice or to HFD-fed ATF3-cKO (Figure2A) We next performed immuno-histochemistry staining of heart slicesderived from NC- and HFD-fed mice with anti-ATF3 antibodies Whileno ATF3 staining was observed in the hearts derived from NC-fed mice
ATF3 expression in cardiomyocytes preserves homeostasis 137D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
or HFD-fed ATF3-cKO mice efficient ATF3 nuclear staining wasobserved in HFD-fed ATF3ff mice (Figure 2B)
In addition western-blot analysis using anti-ATF3 antibodies with car-diomyocyte cell lysate confirmed a high level of ATF3 protein expressionin HFD-fed as compared with NC-fed mice (Figure 2C and D) We nextexamined the cardiac expression levels of various gene specific markersas an indication for cardiac remodeling processes of a stressed heartHFD-fed ATF3-cKO but not ATF3ff mice exhibited significantly higherlevels of the foetal gene programming markers (ANP BNP and bMHC)when compared to the NC-fed wild type counterparts (Figure 2A)Concomitantly a-myosin heavy chain (aMHC) expression was signifi-cantly reduced in HFD-fed mice as compared to NC-fed mice independ-ent of their genotype (Figure 2A) The switch from aMHC to bMHC is anindication of the cardiac remodeling process as well as a cardiac disease
state Heart sections stained with wheat-germ hemagglutinin revealed asignificant increase in the cross sectional area in HFD-fed mice withlarger increased area in ATF3-cKO mice as compared with ATF3ff mice(Figure 2E and F) Taken together cardiac remodeling induced by a HFDis more pronounced in the ATF3-cKO mice than that in the controlATF3ff mice
We next examined fibrosis markers and found elevated expression ofcollagen 1a cTGF and a-SMA in HFD-fed mice as compared to the cor-responding NC-fed mice (Figure 3A) In all cases the ATF3-cKO miceshowed a more pronounced phenotype (Figure 3A) Western-blot analysiswith cell lysates derived from both NC-fed and HFD-fed mice confirmedincreased levels of a-SMA in HFD-fed mice but these high levels weremore noticeable in the lysate derived from HFD-fed ATF3-cKO mice(Figure 3B and C) Another fibrosis marker TGFb3 is significantly elevated
A C
B D
Figure 1 Free fatty acids affect the expression of hypertrophic inflammatory metabolic and fibrosis markers in primary cardiomyocytes Cardiomyocyteswere isolated from adult ATF3ff and ATF3-cKO mice and primary cultures were generated (A) Cardiomyocytes were exposed to 04 mM palmitate orvehicle control for overnight mRNA was extracted and the expression levels of the indicated genes were studied by qRT-PCR The mean ratio of palmitatetreated vs untreated cells for each genotype is shown The results represent the mean ratio plusmn SD (n = 5 independent preparations) (B) ATF3ff cardiomyo-cytes were exposed to 04 mM palmitate for either 1 h or overnight and then fixed and stained with anti-ATF3 (red) and anti-a-actinin (green) antibodiesNuclei were stained with DAPI (blue) (C) Western-blot analysis of cardiomyocytes lysate either palmitate treated overnight or left untreated probed withanti-ATF3 anti-p38 and anti-phospho-p38 antibodies The expression level of a-tubulin was used as loading control Each lane represents a lysate preparedfrom a pool derived from three independent cardiomyocytes culture (D) Isolated cardiomyocytes were treated as described in A Conditioned mediumwas collected and the levels of the indicated cytokines were assessed by ELISA The results represent the mean ratio between palmitate treated tountreatedplusmn SD (n = 5ndash6 independent preparations) P lt_ 005 comparing treated to untreated cells of the same genotype daggerP lt_ 005 comparing palmitate-treated cells between genotypes
138 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
in ATF3-cKO mice but not in the control ATF3ff mice (Figure 3A) againindicating that HFD has a more profound effect on ATF3-cKO mice
We next examined fibrosis histologically by Massonrsquos trichrome stain-ing This analysis confirmed a significant increased fibrosis in the hearts ofHFD-fed mice with ATF3-cKO mice exhibiting the highest fibrosis level(Figure 3D and E) Fibrosis is a result of the deposition of collagen in theECM The net effect of ECM degrading enzymes such as metalloprotei-nases (MMPs) and their inhibitors (TIMPs) determines the level of colla-gen deposition36 The levels of MMP9 were significantly decreased inHFD-fed mice as compared to NC-fed mice independent of the micegenotype (Figure 3A) In contrast TIMP1 levels were significantlyincreased in HFD-fed mice with considerably higher levels in the ATF3-cKO mice consistent with the finding that ATF3-cKO mice displayhigher level of fibrosis (Figure 3A)
Cardiac remodeling processes and diabetes are typically associatedwith increased inflammation We therefore measured several inflamma-tory markers in the hearts of NC- and HFD-fed ATF3ff and ATF3-cKOmice IL-6 expression was highly induced in the hearts of NC- and HFD-fed ATF3-cKO mice as compared with ATF3ff mice Significantly HFD-fed ATF3-cKO mice displayed the highest level (Figure 4A) SimilarlyTNFa was expressed at significantly higher levels in HFD-fed ATF3-cKOmice (Figure 4A) To identify the source of IL-6 and TNFa expression wehave isolated cardiomyocytes from mice fed with HFD for 15 weeks andexamined the cytokinersquos transcription and secretion levels by both qRT-PCR and ELISA respectively Indeed the transcription (Figure 4B) and
secretion (Figure 4C) levels of IL-6 and TNFa were significantly higher incardiomyocytes derived from HFD-fed ATF3-cKO mice as compared totheir ATF3ff cardiomyocytes counterparts To reveal whether theinflammatory response in the heart following a HFD affects circulatingIL-6 and TNFa levels we measured the cytokines in the serum of HFD-fed mice using ELISA Indeed a significant increase in IL-6 and TNFa lev-els were detected in the serum of ATF3-cKO mice (Figure 4D and Erespectively) Collectively we conclude that cardiomyocytes are thecells responsible at least in part for the expression and secretion of IL-6and TNFa in HFD-fed mice with significantly higher circulating levels inATF3 deficient mice
Lastly since inflammation is often associated with a maladaptive meta-bolic response we analysed the expression levels of various metabolicgenes in the heart The level of IRS1 a gene involved in insulin signallingwas significantly reduced in HFD-fed mice The reduction was more pro-nounced in the ATF3-cKO than in control mice (Figure 5A) Similarly thelevel of fatty acid translocase (CD36) was significantly higher in HFD-fedATF3ff mice IRS2 was much higher in the NC-fed ATF3-cKO mice thanin the control counterparts (NC-fed ATF3ff mice) consistent with ourprevious data that its gene expression is suppressed by ATF337
Interestingly IRS2 is induced in HFD-fed ATF3ff mice to comparable levelas HFD-fed ATF3-cKO mice The biological implication of this finding isnot clear One possibility is that induction of IRS2 a pro-survival gene bya HFD is an attempt by the cardiac cells to cope with the HFD stress thisadaptation is not necessary in the ATF3-cKO heart since IRS2 levels are
A
B
C
D
E
F
Figure 2 HFD-fed ATF3-cKO mice display increased expression of cardiac remodeling markers ATF3ff and ATF3-cKO mice were fed with either normalchow (NC) for 25 weeks or switched to a 15-week HFD regimen at 10 weeks of age At endpoint mice were sacrificed and hearts were excised (A) mRNAwas extracted from ventricles and the expression level of ATF3 and the indicated remodeling markers were measured by qRT-PCR Expression levels arepresented as relative values (compared to NC-fed ATF3ff mice determined as 1 for each gene) The results represent the meansplusmn SD (n = 6ndash8group) (B)Immuno-histochemistry analysis of paraformaldehyde fixed heart slices as described in A Representative nuclear staining is indicated by yellow arrowsScale bar = 50lm (C) Western-blot analysis of heart cell lysate probed with anti-ATF3 antibodies The expression level of a-tubulin was used as loadingcontrol (D) Quantification of ATF3 expression from C The results represent the meansplusmn SD (n = 3ndash5group) ratio of ATF3 expression of the indicatedgenotypes relative to NC ATF3ff determined as 1 (E) Ventricles sections were stained with TRITC-labelled wheat germ agglutinin and the cell size was ana-lysed Scale bar = 20lm (F) Quantification of cell size from E The results represent the meansplusmn SD (n = 6ndash8group) P lt_ 005 difference between dietsdaggerP lt_ 005 difference between genotypes
ATF3 expression in cardiomyocytes preserves homeostasis 139D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
already high The expression levels of other lipid and glycolytic regulatorssuch as peroxisome proliferator-activated receptor alpha (PPARa) carni-tine palmitoyltransferase 1 and 2 (CPT12) and pyruvate dehydrogenasekinase 4 (PDK4) were all elevated in HFD-fed mice with no significant dif-ference between the genotypes (Figure 5A) Lastly the level of the glucosetransporter 4 (Glut4) were unaffected by both diet and genotype at themRNA level (Figure 5A) However Glut4 was significantly elevated at theprotein level in HFD-fed mice with no significant difference between thegenotypes (see Supplementary material online Figure S4A and B)Collectively the expression of key metabolic genes was unalteredbetween the various genotypes in response to HFD In view of thechanges observed in metabolic genes in cardiomyocytes treated with pal-mitate (Figure 1A) this result was unexpected Thus we hypothesizedthat ATF3-dependent alterations in metabolic gene expression in cardio-myocytes may be masked by non-cardiomyocytes cells in the heartTherefore we studied the metabolic gene expression changes in isolatedcardiomyocytes derived from HFD- and NCndashfed mice from both ATF3ff
and ATF3-cKO mice (Figure 5B) Indeed the expression of all metabolicgenes tested was augmented in cardiomyocytes derived from HFD-fed ascompared with NC-fed mice Importantly the expression level of CD36CPT1 and CPT2 were significantly upregulated in HFD-fed ATF3-cKOmice as compared with their ATF3ff counterparts (Figure 5B) In contrastthe induction of both IRS1 and IRS2 were significantly lower in cardio-myocytes derived from HFD-fed ATF3-cKO mice (Figure 5B)Collectively the deleterious effects of HFD on cardiac remodeling fibro-sis and inflammation are more pronounced in ATF3-cKO mice suggest-ing that cardiac ATF3 plays a protective role in the HFD paradigm
35 HFD-induced cardiac dysfunctionNext we characterized the effect of HFD and cardiac remodeling oncardiac structure and function in ATF3ff and ATF3-cKO mice by
echocardiography Mice fed with NC had normal heart dimensions andfractional shortening (FS) (Figure 6A and B) HFD significantly reduced FSin both genotypes of HFD-fed mice as compared to corresponding NC-fed mice (Figure 6B) However the reduction was significantly more pro-nounced in the ATF3-cKO mice suggesting deteriorated systolic func-tion in these mice (Figure 6B) The reduced FS in the HFD-fed ATF3-cKO mice seems to be largely driven by an increase in LV internal end-systolic (LVIDs) dimension (Figure 6A) Heart rate was slightly increasedin the HFD-fed mice however this increase cannot explain the cardiacdysfunction observed in ATF3-cKO mice (Figure 6B) In addition weused cardiac MRI the gold standard for LV function and mass asses-sment38 LV mass was significantly higher in the ATF3-cKO mice follow-ing a 15-week HFD regimen (Figure 6C) In addition end-diastolic andend-systolic volumes were significantly larger in ATF3-cKO mice as com-pared to ATF3ff mice Moreover ATF3-cKO mice displayed a signifi-cantly lower ejection fraction as compared to ATF3ff mice (Figure 6D)These results are consistent with the reduced cardiac function found inthe echocardiographic analysis Images of mid-ventricular short-axis(Figure 6E) and long-axis (Figure 6E) slices are demonstrating the cardiaceccentric pattern observed in HFD-fed ATF3-cKO mice Collectivelythe results from the MRI show a greater increase in LV mass and volumein the ATF3-cKO mice as compared to the ATF3ff mice following aHFD suggesting fundamental structural differences in the heart betweenthe ATF3-cKO and ATF3ff mice
36 Impaired glucose tolerance in HFDATF3-cKO miceLastly we measured fasting glucose levels to assess the diabetic state ofthe mice As expected higher glucose levels were observed in HFD-fedmice as compared to NC-fed counterparts Interestingly the
A B
C
D
E
Figure 3 HFD-fed ATF3-cKO mice have higher level of fibrosis in the heart ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) mRNA wasextracted from ventricles and the expression levels of the indicated fibrosis markers were measured by qRT-PCR The expression levels are presented asrelative values (compared to NC-fed ATF3ff mice determined as 1 for each gene n = 6ndash8group) (B) Western-blot analysis of heart cell lysate probed withanti-a-SMA antibody The expression level of a-tubulin was used as loading control (C) Quantification of aSMA expression from B The results representthe meansplusmn SD (n= 3ndash5group) ratio of aSMA expression of the indicated genotypes relative to NC ATF3ff determined as 1 (D) Paraffin-embedded heartsections were stained with Massonrsquos trichrome to visualize fibrosis Scale bar = 100 lm (E) The level of fibrosis () stained by Massonrsquos trichrome was quan-tified and the results represent the meansplusmn SD (n = 6ndash8group) P lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes
140 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
hyperglycemia state of the ATF3-cKO mice was significantly higher thanin the ATF3ff mice following 15 weeks of HFD (Figure 7A) Consistentlythe glucose tolerance test (GTT) revealed that HFD-fed ATF3-cKOmice displayed higher blood glucose levels and longer glucose clearancetime when compared to HFD-fed ATF3ff whereas NC-fed ATF3ff andATF3-cKO mice had normal glucose clearance (Figure 7B)
Interestingly whole body ATF3-KO mice fed with HFD also displayedimpaired glucose clearance (see Supplementary material onlineFigure S5A)39 Previously we reported that decreased glucose tolerancein whole body ATF3-KO mice is at least in part due to reduced insulingene expression in b-cells (see Supplementary material online FigureS5B)39 To determine insulin level in HFD-fed ATF3-cKO mice weanalysed blood insulin levels by ELISA In both HFD-fed ATF3ff andATF3-cKO mice the serum insulin levels were significantly higher ascompared to their NC-fed counterpart mice with the ATF3-cKO micedisplaying the highest insulin levels (Figure 7C) Together these resultssuggest that peripheral tissues of ATF3-cKO mice have reduced ability
to uptake glucose despite their high insulin level in the blood Next wesought to measure the protein levels of components along the insulinpathway Akt and Foxo340 Both Akt and Foxo3 protein levels were sim-ilar in HFD-fed mice independent of the genotype (see Supplementarymaterial online Figure S4A and C) Interestingly phosphorylation of Akt(Ser473) which indicates the activity of the insulin pathway was slightlyelevated in HFD-fed mice however this did not reach statistical signifi-cance and was similar in both of the genotypes (see Supplementary material online Figure S4D) To measure the activity of the insulin signallingpathway following insulin stimulation we used the ratio between phos-phorylated Akt (Ser473) and total Akt expression HFD-fed mice wereinjected with insulin (10 Ukg) and the heart liver and skeletal musclewere harvested 15 min after the injection (Figure 7D and E and seeSupplementary material online Figure S6AndashD) Insulin injection resultedin a significant increase of pAktAkt ratio in the hearts of ATF3ff micewhile no significant change was observed in Akt phosphorylation inATF3-cKO mice (Figure 7D and E) Similarly low Akt phosphorylation
A
D E
B C
Figure 4 HFD-fed ATF3-cKO mice display inflammatory response in the heart (A) ATF3ff and ATF3-cKO mice were fed as described in Figure 2 mRNAwas extracted from ventricles and the expression levels of the indicated inflammatory markers were measured by qRT-PCR Expression levels are presentedas relative values (compared to NC-fed ATF3ff mice determined as 1 for each gene n= 6ndash8group) (B) Mice were fed as described in Figure 2Cardiomyocytes were isolated and primary cultures were generated mRNA was extracted from an overnight culture and the expression levels of IL-6 andTNFa were studied by qRT-PCR The mRNA level obtained in cardiomyocytes derived from NC-fed ATF3ff mice was determined as 1 and the relativeexpression of all other treatment is shown (n = 5ndash6 independent cardiomyocyte preparations) (C) Conditioned media was collected from cardiomyocytesdescribed in B and ELISA assay for the indicated cytokines is shown Cytokines level is presented as relative values to NC-fed ATF3ff mice determined as 1for each cytokine (n = 5ndash6 independent preparations) (D and E) Blood was collected from HFD-fed mice and IL-6 and TNFa serum levels were quantifiedby ELISA (n = 8ndash12group) The results represent the meansplusmn SD P lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes
ATF3 expression in cardiomyocytes preserves homeostasis 141D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
levels were also observed in both the liver and skeletal muscle of theATF3-cKO mice however this did not reach statistically significance (seeSupplementary material online Figure S6AndashD) Consistently palmitate-treated cardiomyocytes derived from ATF3-cKO mice displayedreduced phosphorylation of Akt following 30-min insulin stimulation(see Supplementary material online Figure S6E)
Collectively our findings revealed a surprising phenomenon that car-diac ATF3 deficiency results in increased cardiac remodeling leading topotentiation of local and peripheral insulin resistance
4 Discussion
A HFD is a commonly used regimen to induce T2D The majority of theliterature focuses on diabetic symptoms without addressing the issue ofT2D-induced cardiomyopathy Even when it is addressed there is noclear consensus regarding the extent of cardiac effects imposed byHFD41 Diet-induced obesity in mouse models is associated with globalinsulin resistance and increased myocardial fatty acid utilization how-ever the reported phenotypes on cardiac function vary from extensivehypertrophy and heart failure4 to lack of cardiac dysfunction4243 Thesevariations have been attributed to diet regimens andor sensitivity ofmouse strains to a HFD In this report we present evidence that cardiacATF3 deficiency exacerbates the deleterious effect of a HFD on theheart such as maladaptive cardiac remodeling fibrosis inflammation andcardiac dysfunction Our work may provide a potential molecular insightto variations in the literature regarding the impact of HFD on the heartAs ATF3 is a stress-inducible gene we propose that its proper inductionin the cardiomyocytes is protective and helps the heart to cope withHFD stress It is possible that the induction of ATF3mdashits expressionlevel duration and kineticsmdashvaries in different HFD regimens andormouse strains contributing to the variations observed in the literature
HFD regimen induced a significant higher level of fibrosis in the heartof ATF3-cKO mice We have identified TIMP1 as a possible mechanismto explain the increase in fibrosis in ATF3-cKO cardiomyocytes Severallines of evidence suggest that TIMP1 is possibly an ATF3 target gene
First we observed an increase in TIMP1 transcription in the whole heartand in isolated cardiomyocytes (Figures 1A and 3A) Second TIMP 1 3and 4 were identified as significantly highly expressed genes in the heartof ATF3-KO mice as compared with wild type mice following aorticbanding20 Third a functional AP-1 regulatory element within the TIMP1promoter was described44 Indeed preliminary results in our lab suggestthat ATF3 downregulates the TIMP1 promoter (data not shown) Thusit is possible that TIMP1 is suppressed by ATF3 and therefore results in anet increase in MMPs activity that helps to maintain ECM formationdeg-radation homeostasis
Our findings are consistent with previous studies demonstrating that adeficiency in ATF3 can promote cardiac maladaptive changes which leadto cardiac dilatation and hypertrophy in an aortic banding model1920 Incontrast a different study using a pressure overload model reportedthat ATF3 induces a maladaptive cardiac response17 These seeminglyconflicting data may be explained by the different patterns of ATF3expression induced by the different models In the pressure overloadmodel the acute and transient expression of ATF3 in cardiomyocytesinitiates a cascade of maladaptive changes17 whereas in the aortic band-ing model and the HFD model described here ATF3 expression is longlasting
In this context this is the first report using mice with cardiac-selectivedeficiency of ATF3 (ATF3-cKO) Previous reports on the roles of ATF3in cardiac function were carried out using whole body knockout ofATF325 or transgenic mice ectopically expressing ATF3 in cardiomyo-cytes16 A key advancement of ATF3-cKO mice over the whole bodyknockout mice is that they provide insights into the function of ATF3selectively in cardiomyocytes without the complication of ATF3 defi-ciency in other cells Thus the ATF3-cKO mouse is a useful tool to bet-ter understand the function of cardiac ATF3
Consistent with our previous report25 here we showed that cardiacATF3 deficiency has a more severe HFD-induced diabetic phenotypethan the control wild type mice However while reduced insulin genetranscription in ATF3-deficient b-cells was observed25 our current dataindicates that a lack of insulin is not the explanation for the glucose intol-erance found in the HFD-fed ATF3-cKO mice In contrast these mice
A B
Figure 5 HFD-fed ATF3-cKO mice show metabolic response in the heart ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) mRNA wasextracted from ventricles and the expression levels of the indicated metabolic markers were measured by qRT-PCR Expression levels are presented as rela-tive values (compared to NC-fed ATF3ff mice determined as 1 for each gene n= 6ndash8group) (B) Cardiomyocytes were isolated and plated on laminin for1 h followed by mRNA preparation (n = 4ndash5 independent preparations) The expression levels of the indicated metabolic markers were measured by qRT-PCR Expression levels are presented as relative values (to NC-fed ATF3ff mice determined as 1 for each gene) The results represent the meansplusmn SDP lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes
142 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
had higher blood insulin level (Figure 7C) Therefore upon HFD chal-lenge cardiac-deficiency of ATF3 exacerbates HFD-induced T2D Thislsquoperipheral effectrsquo is surprising since ATF3 is selectively deleted in thecardiomyocytes
The question then is how can we explain the effect of cardiacATF3 deficiency on peripheral glucose intolerance One possibility isthe reduced IRS1 expression in ATF3-deficient cardiomyocytes bothin vivo and in vitro IRS1 suppression may lead to impaired insulin sig-nalling in the heart thereby exacerbating glucose intolerance Indeedwe observed that HFD-fed ATF3-cKO mice displayed insulin resist-ance upon insulin challenge when compared to HFD-fed ATF3ff
mice In addition palmitate-treated cardiomyocytes lacking ATF3 dis-played lower insulin signalling as compared to their wild type coun-terparts Since the heart is only a minor glucose consuming organ incomparison to skeletal muscle cells or the brain the peripheral sys-temic effect cannot be solely based on this observation A case inpoint transgenic mice with insulin receptor (IR) deficiency in theheart do not display a diabetic phenotype4546 Another attractive
possibility for the peripheral effect of cardiac ATF3-deletion is itsimpact on inflammation It is well established that low-grade inflam-mation years in advance of the disease onset is involved in the patho-genic processes causing T2D8 Mediators of inflammation such asTNFa interleukin (IL)-1b the IL-6 family of cytokines IL-18 and cer-tain chemokines have been proposed to be involved in the eventscausing both forms of diabetes (T1D and T2D)8 Interestingly wehave shown that IL-6 a well-known ATF3 target gene47 is known toenhance its transcription and secretion of cytokines such as IL-6 andTNFa in palmitate-treated primary cultures of ATF3-cKO cardio-myocytes as well as in cardiomyocytes derived from HFD-fed ATF3-cKO mice IL-6 has been proposed to play a role in glucose homeo-stasis and metabolism directly and indirectly via its action on skeletalmuscle cells adipocytes hepatocytes pancreatic b-cells and neuro-endocrine cells48
Therefore we hypothesize that elevated levels of inflammatory cyto-kines in ATF3-cKO mice mediate a cross-talk in peripheral tissues leadingto impaired glucose tolerance Although circulating IL-6 and TNFa
A
B
C
D
E
F
Figure 6 HFD-fed ATF3-cKO mice present deteriorated heart function ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) At endpointmice hearts were examined by micro ultrasound The following parameters were measured interventricular septal end diastole (IVSd) left ventricular pos-terior wall end diastole (LVPWd) maximal left ventricular internal end-diastole (LVIDd) end-systole (LVIDs) and heart rate The results represent themeansplusmn SD (n = 6ndash8 in each group as shown in panel B) (B) Heart function was assessed by calculating percentage of fractional shortening (FS) accordingto FS () = [(LVDd-LVDs)LVDd]100 The results represent the meansplusmn SD (n = 6ndash8 in each group) (C and D) Left ventricular cardiac volumes massand function were examined by a cardiac MRI The following parameters were measured left ventricular end-diastolic (LVEDV) and left ventricular end-sys-tolic volume (LVESV) left ventricular (LV) mass and ejection fraction (EF) The results represent the meansplusmn SD (n = 7 in each group) (E and F)Representative images of mid-ventricular short-axis (E) and long-axis (F) slice at peak diastole and systole P lt_ 005 difference between diets daggerP lt_ 005 dif-ference between genotypes
ATF3 expression in cardiomyocytes preserves homeostasis 143D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
cytokines may be derived from different tissues we provide ample evi-dence that ATF3-defficient cardiomyocytes produce and secrete higherlevels of these cytokines as compared to their wild type counterpartsThis indicates a lsquocardiac-to-peripheralrsquo effect in T2D-induced cardiomyop-athy This is a reverse direction from the lsquoperipheral-to-cardiacrsquo effect dis-cussed in the introduction where inflammation in the peripheral tissuesleads to cardiac dysfunction Since inflammatory cytokines are soluble
factors they can exert a systemic effect This is why it makes sense thattheir effect can go both ways Thus diabetes and cardiomyopathy are apart of a vicious cycle Previously it was shown that cardiac specific dele-tion of the thyroid hormone complex mediator MED13 results in aperipheral effect upon HFD Cardiac MED13 deficiency suppresses meta-bolic genes in the heart which leads to a decrease in energy expenditureand an obese phenotype most likely by the secretion of cardiokines that
A
B
C
D
E
Figure 7 HFD-fed ATF3-cKO mice exhibit impaired glucose tolerance ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) Fasting (6 h)blood glucose concentrations (mgdl) The results represent the meansplusmn SD (n = 20ndash30group) P lt_ 005 difference between diets daggerP lt_ 005 differencebetween genotypes (B) Glucose tolerance test (GTT) Values are presented as relative to initial valueplusmn SD (n = 20ndash25group) P lt_ 005 difference betweendiets daggerP lt_ 005 difference between genotypes (C) Serum insulin concentrations were quantified by ELISA Values are presented as relative values (com-pared to NC-fed ATF3ff mice determined as 1 n = 15ndash20group) P lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes (D and E)Effect of HFD on phosphorylated Akt (Ser473 pAkt) to Akt ratio levels in heart lysate following either saline or insulin injection (10 U1 kg) is shownRepresentative blots (D) Densitometric analysis is presented as means ratio (n = 3ndash6group) pAktAkt ratio is presented as relative values (compared toHFD-fed saline injected ATF3ff mice determined as 1) P lt_ 005 insulin vs saline treatment daggerP lt_ 005 difference between genotypes (E)
144 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
mediates the cross talk between the heart and peripheral tissues49ndash51
Here we also observed increased levels of natriuretic peptides (ANP andBNP) which together with IL-6 and TNFa may mediate the cross-talk withthe peripheral tissues to induce glucose intolerance
Does this finding have any relevance to cardiac disease in humansSeveral lines of evidence suggests that congestive heart failure in patientsis associated with an increase in blood glucose levels52 In addition highblood glucose levels in non-diabetic patients with acute myocardialinfarction serves as a prognostic predictor of long-term mortality5354
Thus here we provide evidence that high blood glucose levels are a partof the symptoms of cardiac remodeling process
Collectively our findings suggest that cardiac ATF3 expression isresponsible for an adaptive cardiac response to the stressful environmentunder a HFD (Figure 8) ATF3 dampens the inflammatory response andsuppresses the expression of ECM remodeling genes and thus amelioratescardiac function In HFD-fed mice with ATF3 deficiency or in mice withcardiac remodeling the inflamed heart affects also peripheral tissuesthereby worsening T2D in peripheral tissues resulting in elevated bloodglucose level and turning on a vicious cycle to worsen cardiomyopathy
Supplementary material
Supplementary material is available at Cardiovascular Research online
AcknowledgementsThe authors wish to thank Drs Keren Bitton-Worms and Ran Rostokerand Prof Derek LeRoith from the Rambam Medical Center for their
assistance in GTT and ITT Dr Edith Suss Toby Maya Holdengreber andLior Liba from the BCF Bioimaging Center (Faculty of MedicineTechnion) for their assistance with imaging and image analysis Prof DEMichelle from University of Michigan for cardiomyocytes culture techni-que and Prof Yehiel Zick from the Weizmann institute of Sciences forcritical comments
Conflict of interest none declared
FundingThis work was partially supported by the United States-Israel BinationalGrant 2009179 to AA and TH In addition we wish to thank the MarsPittsburgh Fund for Medical Research to AA the Department of Defensegrant W81XWH-14-1-0179 to TH and to D Dan and Betty Kahn travelgrant support to LK
References
1 Roger VL Go AS Lloyd-Jones DM Benjamin EJ Berry JD Borden WB Bravata DMDai S Ford ES Fox CS Fullerton HJ Gillespie C Hailpern SM Heit JA Howard VJKissela BM Kittner SJ Lackland DT Lichtman JH Lisabeth LD Makuc DM MarcusGM Marelli A Matchar DB Moy CS Mozaffarian D Mussolino ME Nichol GPaynter NP Soliman EZ Sorlie PD Sotoodehnia N Turan TN Virani SS Wong NDWoo D Turner MB American Heart Association Statistics C Stroke Statistics SHeart disease and stroke statisticsndash2012 update a report from the American HeartAssociation Circulation 2012125e2ndashe220
2 Wilson C Diabetes ACCORD 5-year outcomes of intensive glycemic control NatRev Endocrinol 20117314
3 Ernande L Derumeaux G Diabetic cardiomyopathy myth or reality Arch CardiovascDis 2012105218ndash225
4 Battiprolu PK Hojayev B Jiang N Wang ZV Luo X Iglewski M Shelton JM GerardRD Rothermel BA Gillette TG Lavandero S Hill JA Metabolic stress-induced
InsulinGF
Cytokines
FFA
GlucoseATF3
Cardiac
Remodeling
Factors
Paracrine action
Collagens
and ECM
proteins
Fibrosis
Cardiac
Dysfunction
Systemic Effects
Glucose Tolerance
TNFα
IL-6
Cardiomyocyte
ATF3
Homeostasis
Homeostasis
TNFα
IL-6
TIMP1
BNP
ANP
Figure 8 lsquoPeripheral-to-cardiacrsquo and lsquoCardiac-to-peripheralrsquo model scheme Schematic diagram showing the ATF3 adaptive role in maintaining hearthomeostasis following HFD-induced T2D In contrast stressful environment under HFD leads to maladaptive cardiac response in the absence of ATF3Cardiac remodeling caused by ATF3 deficiency induces an increase in cardiac ECM remodeling and inflammatory genes and mediates the communication ofcardiomyocytes with heart resident cells potentially mediated by IL-6 TNFa ANP BNP and TIMP1 In this situation increased cardiac fibrosis and inflamma-tory response worsen cardiac function The inflamed failing heart mediates the communication of cardiomyocytes with distant organs as well thereby affect-ing diabetic outcome and glucose homeostasis
ATF3 expression in cardiomyocytes preserves homeostasis 145D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
activation of FoxO1 triggers diabetic cardiomyopathy in mice J Clin Invest20121221109ndash1118
5 Mandavia CH Aroor AR Demarco VG Sowers JR Molecular and metabolic mecha-nisms of cardiac dysfunction in diabetes Life Sci 201392601ndash608
6 Ventura-Clapier R Garnier A Veksler V Energy metabolism in heart failure J Physiol20045551ndash13
7 Guo S Insulin signaling resistance and the metabolic syndrome insights from mousemodels into disease mechanisms J Endocrinol 2014220T1ndashT23
8 Montane J Cadavez L Novials A Stress and the inflammatory process a major causeof pancreatic cell death in type 2 diabetes Diabetes Metab Syndr Obes 2014725ndash34
9 Ashrafian H Frenneaux MP Opie LH Metabolic mechanisms in heart failureCirculation 2007116434ndash448
10 Tanabe K Liu Y Hasan SD Martinez SC Cras-Meneur C Welling CM Bernal-MizrachiE Tanizawa Y Rhodes CJ Zmuda E Hai T Abumrad NA Permutt MA Glucose andfatty acids synergize to promote B-cell apoptosis through activation of glycogen syn-thase kinase 3beta independent of JNK activation PLoS One 20116e18146
11 Heineke J Molkentin JD Regulation of cardiac hypertrophy by intracellular signallingpathways Nat Rev Mol Cell Biol 20067589ndash600
12 Hai T Wolford CC Chang YS ATF3 a hub of the cellular adaptive-response net-work in the pathogenesis of diseases is modulation of inflammation a unifying com-ponent Gene Expr 2010151ndash11
13 Takii R Inouye S Fujimoto M Nakamura T Shinkawa T Prakasam R Tan K HayashidaN Ichikawa H Hai T Nakai A Heat shock transcription factor 1 inhibits expression ofIL-6 through activating transcription factor 3 J Immunol 20101841041ndash1048
14 Giraldo A Barrett OP Tindall MJ Fuller SJ Amirak E Bhattacharya BS Sugden PHClerk A Feedback regulation by Atf3 in the endothelin-1-responsive transcriptomeof cardiomyocytes Egr1 is a principal Atf3 target Biochem J 2012444343ndash355
15 Hasin T Elhanani O Abassi Z Hai T Aronheim A Angiotensin II signaling up-regulates the immediate early transcription factor ATF3 in the left but not the rightatrium Basic Res Cardiol 2011106175ndash187
16 Koren L Elhanani O Kehat I Hai T Aronheim A Adult cardiac expression of theactivating transcription factor 3 ATF3 promotes ventricular hypertrophy PLoS One20138e68396
17 Koren L Alishekevitz D Elhanani O Nevelsky A Hai T Kehat I Shaked Y AronheimA ATF3-dependent cross-talk between cardiomyocytes and macrophages promotescardiac maladaptive remodeling Int J Cardiol 2015198232ndash240
18 Okamoto Y Chaves A Chen J Kelley R Jones K Weed HG Gardner KL Gangi LYamaguchi M Klomkleaw W Nakayama T Hamlin RL Carnes C Altschuld R BauerJ Hai T Transgenic mice with cardiac-specific expression of activating transcriptionfactor 3 a stress-inducible gene have conduction abnormalities and contractile dys-function Am J Pathol 2001159639ndash650
19 Zhou H Shen DF Bian ZY Zong J Deng W Zhang Y Guo YY Li H Tang QZActivating transcription factor 3 deficiency promotes cardiac hypertrophy dysfunc-tion and fibrosis induced by pressure overload PLoS One 20116e26744
20 Zhou H Guo H Zong J Dai J Yuan Y Bian ZY Tang QZ ATF3 regulates multipletargets and may play a dual role in cardiac hypertrophy and injury Int J Cardiol2014174838ndash839
21 Yang J Yang CJ Yang J Fan ZX ATF3 a promotion effect or a inhibition effect in car-diac maladaptive remodeling Int J Cardiol 2015201245ndash246
22 Koren L Shaked Y Aronheim A Response letter ldquoATF3 a promoter or inhibitor ofcardiac maladaptive remodelingrdquo Int J Cardiol 2015201692
23 Ma ZG Wei WY Xu SC Zhang WB Dai J Tang QZ ATF3 a potential target forcardiac maladaptive remodeling Int J Cardiol 201620250ndash51
24 Zhou H Yuan Y Ni J Guo H Deng W Bian ZY Tang QZ Pleiotropic and puzzlingeffects of ATF3 in maladaptive cardiac remodeling Int J Cardiol 201620687ndash88
25 Zmuda EJ Qi L Zhu MX Mirmira RG Montminy MR Hai T The roles of ATF3 anadaptive-response gene in high-fat-diet-induced diabetes and pancreatic beta-celldysfunction Mol Endocrinol 2010241423ndash1433
26 Lu D Chen J Hai T The regulation of ATF3 gene expression by mitogen-activatedprotein kinases Biochem J 2007401559ndash567
27 Hartman MG Lu D Kim ML Kociba GJ Shukri T Buteau J Wang X Frankel WLGuttridge D Prentki M Grey ST Ron D Hai T Role for activating transcription fac-tor 3 in stress-induced beta-cell apoptosis Mol Cell Biol 2004245721ndash5732
28 Wolford CC McConoughey SJ Jalgaonkar SP Leon M Merchant AS Dominick JLYin X Chang Y Zmuda EJ OrsquoToole SA Millar EK Roller SL Shapiro CL OstrowskiMC Sutherland RL Hai T Transcription factor ATF3 links host adaptive response tobreast cancer metastasis J Clin Iinvest 20131232893ndash2906
29 Agah R Frenkel PA French BA Michael LH Overbeek PA Schneider MD Generecombination in postmitotic cells Targeted expression of Cre recombinase pro-vokes cardiac-restricted site-specific rearrangement in adult ventricular musclein vivo J Clin Invest 1997100169ndash179
30 Livet J Weissman TA Kang H Draft RW Lu J Bennis RA Sanes JR Lichtman JWTransgenic strategies for combinatorial expression of fluorescent proteins in thenervous system Nature 200745056ndash62
31 Davis J Maillet M Miano JM Molkentin JD Lost in transgenesis a userrsquos guidefor genetically manipulating the mouse in cardiac research Circ Res2012111761ndash777
32 Heiberg E Sjogren J Ugander M Carlsson M Engblom H Arheden H Design andvalidation of segment ndash freely available software for cardiovascular image analysisBMC Med Imaging 2010101
33 Kabaeva Z Zhao M Michele DE Blebbistatin extends culture life of adult mouse car-diac myocytes and allows efficient and stable transgene expression Am J Physiol HeartC 2008294H1667ndash1674
34 Subramaniam A Jones WK Gulick J Wert S Neumann J Robbins J Tissue-specificregulation of the alpha-myosin heavy chain gene promoter in transgenic mice J BiolChem 199126624613ndash24620
35 Cunha DA Hekerman P Ladriere L Bazarra-Castro A Ortis F Wakeham MCMoore F Rasschaert J Cardozo AK Bellomo E Overbergh L Mathieu C Lupi R HaiT Herchuelz A Marchetti P Rutter GA Eizirik DL Cnop M Initiation and executionof lipotoxic ER stress in pancreatic beta-cells J Cell Sci 20081212308ndash2318
36 Wynn TA Ramalingam TR Mechanisms of fibrosis therapeutic translation forfibrotic disease Nat Med 2012181028ndash1040
37 Li D Yin X Zmuda EJ Wolford CC Dong X White MF Hai T The repression ofIRS2 gene by ATF3 a stress-inducible gene contributes to pancreatic beta-cell apop-tosis Diabetes 200857635ndash644
38 Myerson SG Bellenger NG Pennell DJ Assessment of left ventricular mass by cardi-ovascular magnetic resonance Hypertension 200239750ndash755
39 Zmuda EJ Viapiano M Grey ST Hadley G Garcia-Ocana A Hai T Deficiency ofAtf3 an adaptive-response gene protects islets and ameliorates inflammation in asyngeneic mouse transplantation model Diabetologia 2010531438ndash1450
40 Riehle C Abel ED Insulin signaling and heart failure Circ Res 20161181151ndash116941 Schreyer SA Wilson DL LeBoeuf RC C57BL6 mice fed high fat diets as models for
diabetes-accelerated atherosclerosis Atherosclerosis 199813617ndash2442 Brainard RE Watson LJ Demartino AM Brittian KR Readnower RD Boakye AA
Zhang D Hoetker JD Bhatnagar A Baba SP Jones SP High fat feeding in mice isinsufficient to induce cardiac dysfunction and does not exacerbate heart failure PLoSOne 20138e83174
43 Gupte AA Minze LJ Reyes M Ren Y Wang X Brunner G Ghosn M Cordero-Reyes AM Ding K Pratico D Morrisett J Shi ZZ Hamilton DJ Lyon CJ Hsueh WAHigh-fat feeding-induced hyperinsulinemia increases cardiac glucose uptake and mito-chondrial function despite peripheral insulin resistance Endocrinology20131542650ndash2662
44 Logan SK Garabedian MJ Campbell CE Werb Z Synergistic transcriptionalactivation of the tissue inhibitor of metalloproteinases-1 promoter via functionalinteraction of AP-1 and Ets-1 transcription factors J Biol Chem1996271774ndash782
45 Belke DD Betuing S Tuttle MJ Graveleau C Young ME Pham M Zhang DCooksey RC McClain DA Litwin SE Taegtmeyer H Severson D Kahn CR AbelED Insulin signaling coordinately regulates cardiac size metabolism and contractileprotein isoform expression J Clin Invest 2002109629ndash639
46 Fernandez AM Kim JK Yakar S Dupont J Hernandez-Sanchez C Castle AL FilmoreJ Shulman GI Le Roith D Functional inactivation of the IGF-I and insulin receptors inskeletal muscle causes type 2 diabetes Genes Dev 2001151926ndash1934
47 Gilchrist M Thorsson V Li B Rust AG Korb M Roach JC Kennedy K Hai TBolouri H Aderem A Systems biology approaches identify ATF3 as a negative regu-lator of Toll-like receptor 4 Nature 2006441173ndash178
48 Kristiansen OP Mandrup-Poulsen T Interleukin-6 and diabetes the good the bad orthe indifferent Diabetes 200554(Suppl 2)S114ndashS124
49 Grueter CE van Rooij E Johnson BA DeLeon SM Sutherland LB Qi X Gautron LElmquist JK Bassel-Duby R Olson EN A cardiac microRNA governs systemic energyhomeostasis by regulation of MED13 Cell 2012149671ndash683
50 Baskin KK Grueter CE Kusminski CM Holland WL Bookout AL Satapati S KongYM Burgess SC Malloy CR Scherer PE Newgard CB Bassel-Duby R Olson ENMED13-dependent signaling from the heart confers leanness by enhancing metabo-lism in adipose tissue and liver EMBO Mol Med 201461610ndash1621
51 Nakamura M Sadoshima J Heart over mind metabolic control of white adipose tis-sue and liver EMBO Mol Med 201461521ndash1524
52 Tenenbaum A Fisman EZ Impaired glucose metabolism in patients with heart failurepathophysiology and possible treatment strategies Am J Cardiovasc Drugs20044269ndash280
53 Suskin N McKelvie RS Burns RJ Latini R Pericak D Probstfield J Rouleau JL SigouinC Solymoss CB Tsuyuki R White M Yusuf S Glucose and insulin abnormalitiesrelate to functional capacity in patients with congestive heart failure Eur Heart J2000211368ndash1375
54 Aronson D Hammerman H Kapeliovich MR Suleiman A Agmon Y Beyar RMarkiewicz W Suleiman M Fasting glucose in acute myocardial infarction incremen-tal value for long-term mortality and relationship with left ventricular systolic func-tion Diabetes Care 200730960ndash966
146 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022

structure and accurate planning of the subsequent scans (ii) Single slicetwo chamber Short Axis view perpendicular to LV walls at the level ofthe papillary muscles and (iii) Contiguous multi slices two chamber ShortAxis view for entire LV coverage
Cardiac MR sequences parameters
i Single slice Long Axis view 1 mm thickness FOV = 3 cm3 cm matrixdimension = 400400 spatial resolution = 7575lm2 repetitionechotime (TRTE) = 6532 ms number of repetitions = 100 10 cardiacmovie frames (cardiac phases)
ii Single slice two chamber Short Axis view 07 mm thicknessFOV=3 cm3 cm matrix dimension=400400 spatial reso-lution=7575lm2 repetitionecho time (TRTE) = 6532 ms numberof repetitions = 150 15 cardiac movie frames (cardiac phases)
iii Multi slices two chamber Short Axis view Contiguous 9ndash12 slicesdepending on heart size 07 mm thickness FOV = 3 cm3 cm matrixdimension = 400400 spatial resolution=7575lm2 repetitionechotime (TRTE) = 81-12032 ms (depending on slices number) number ofrepetitions = 120 15 cardiac movie frames (cardiac phases)
The total acquisition time for each animal was40 min
27 MRI image analysisImage analysis was performed using the freely available Segment soft-ware (Medviso AB Lund Sweden)32 Manual segmentation of the endo-cardium and epicardium of the LV was performed to measure LV massand LV functional parametersmdashend-systolic and end-diastolic (ESV andEDV respectively) volumes Ejection fraction (EF) was calculated as EF() = [(EDV-ESV)EDV]100 The person performing the MRI and thedata analysis was blinded to the mice genotype
28 Heart harvestingAt endpoint mice were anesthetized weighed and sacrificed Heartswere excised and ventricles were then divided into three pieces thatwere used for protein extraction RNA purification and histologicalanalysis
29 mRNA extractionmRNA was purified from ventricles using an Aurum total RNA fatty orfibrous tissue kit (732-6830 Bio-Rad) or from isolated cardiomyocytesusing a High Pure RNA Isolation kit (11828665001 Roche Diagnostics)according to the manufacturerrsquos instructions
210 Quantitative real-time PCR (RT-qPCR)cDNA was synthesized from 800 or 50 ng of purified mRNA derivedfrom the ventricles or isolated from the cardiomyocytes respectivelyPurified mRNA was added to a total reaction mix of high-capacity cDNAreverse transcription kit (4368814 Applied Biosystems) in a final vol-ume of 20 mL Real-time PCR was performed using Rotor-Gene6000TM (Corbett) equipment with absolute blue SYBER green ROXmix (Thermo Scientific AB-4162B) Serial dilutions of a standard samplewere included for each gene to generate a standard curve Values werenormalized to b2 microglobulin expression levels The primer sequencesare shown in Table S1 (see Supplementary material online)
211 Western-blot analysisHarvested tissues were homogenized in RIPA buffer (PBS containing 1NP-40 5 mgmL Na-deoxycholate 01 SDS) and supplemented withprotease inhibitor cocktail (P-8340 SigmandashAldrich) Homogenization
was performed at 4 C using the Bullet Blender homogenizer (BBX24Next advance) according to the manufacturerrsquos instructions as previ-ously described17
212 ImmunohistochemistryTissues were fixed in 4 formaldehyde overnight then embedded inparaffin serially sectioned at 10 lm intervals and then mounted onslides Sections were processed for deparaffinization (xylene 20 min)dehydration (isopropanol) rehydration (H2O) and antigen unmasking(10 mM sodium citrate pH-6 90 C 12 min) Immunostaining was per-formed using Histostain kit (959643 Invitrogen) according to the man-ufacturerrsquos instructions Anti-ATF3 antibodies were diluted 150Following immunostaining nuclei were stained with hematoxylin
213 AntibodiesThe primary antibodies used anti-SMA (Cat A2547) anti-a-tubulin(Cat T-9026) and anti-a-actinin (Cat A7811) which were all pur-chased from SigmandashAldrich Anti-GFP (SC-8334) was purchased fromSanta-Cruz Anti-p38 (Cat 9212) and anti-phospho-p38 (Cat 9211)were purchased from Cell signalling Anti-pAkt (Cat AF887) and anti-Akt (Cat MAB 2055) was purchased from RampD Systems anti-Foxo3(Cat 07-1719) was purchased from Merck Millipore and anti-Glut4(Cat ab654) was purchased from Abcam Lastly anti-ATF3 antibodywas generated in Aronheimrsquos lab15
214 Cell size analysisSections were stained following deparaffinization with Wheat-germagglutinin TRITC-conjugated (SigmandashAldrich Cat L5266) and diluted toa 1100 phosphate-buffered saline (PBS) Sections were washed 3 timeswith PBS and mounted in Fluoromount-G (Southern BiotechnologyBirmingham AL 0100-01) Sections were then viewed using a ZeissLSM700 confocal microscope (Thornwood NY) equipped with a 40oil objective and solid state 555nm laser
Quantification of the cell size was performed with Image Pro Plus soft-ware Five fields in each slide were photographed Unstained areas werethen identified and segmented using Image Pro Plus software In eachstained area the mean cell perimeter and area was calculated and thenumber of cells was measured
215 Fibrosis stainingHeart tissue was fixed in 4 formaldehyde overnight embedded in par-affin serially sectioned at 10 mm intervals and then mounted on slidesMassonrsquos trichrome staining was performed according to the standardprotocol Images were acquired by using Virtual Microscopy (Olympus)Stained areas were segmented and quantified following calibration usingImage Pro Plus software Five fields were quantified for each slide
216 In vitro cardiomyocyte cultureAdult cardiomyocytes were isolated from 12- to 25-week-old NC- orHFD-fed mice by a modified Langendorff perfusion apparatus asdescribed33 Isolated cells were plated in culture media (MEMmdashEagleHanksrsquo salts) supplemented with penicillin 100 UmL glutamine 2 mMNaHCO3 4 mM HEPES 10 mM bovine serum albumin (BSA) 0225 mM blebbistatin (cat B0560 SigmandashAldrich) and 5 foetal bovineserum Cardiomyocytes were then plated on 35-mm dishes coated withmouse laminin (L2020 SigmandashAldrich) at a density of 105 cellsmL inserum free medium
136 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
217 Palmitate-treated cardiomyocytesForty-five minutes after plating the medium was replaced with a culturemedium containing either 04 mM palmitate or BSA (control) Palmitatestock solution (20 mM) was diluted in 001 M NaOH by heating at 70Cfor 30 min as described10 Following overnight culture conditionedmedium was collected and cardiomyocytes were harvested for RNAextraction In addition overnight palmitate- or BSA-treated cardiomyo-cytes were incubated with 100 nM insulin (I-5500 SigmandashAldrich) for30 min and were then harvested for protein extraction
218 Enzyme-linked immuno-absorbentassayInterleukin 6 (IL-6) and tumour necrosis factor a (TNFa) levels in condi-tioned medium from an overnight culture of primary cardiomyocyteswere measured by ELISA using the DuoSet ELISA kit (RampD Systems)according to the manufacturerrsquos instructions Blood from non-fastingHFD-fed mice was collected from submandibular bleeds and serum IL-6and TNFa levels were measured by ELISA (RampD Systems)
219 StatisticsOur data is expressed as means plusmn SD The statistical test for the GTTwas performed by repeated-measures analysis of variance (ANOVA)The comparison of two means was analysed by the Studentrsquos t-test Thecomparison between several means was analysed by one-way ANOVAfollowed by Tukeyrsquos post hoc analysis All statistical analyses were per-formed using GraphPad Prism 5 software (La Jolla CA) A P value lt_ 005was accepted as statistically significant
3 Results
31 aMHC promoter specificityFirst we verified the tissue specific expression of the Cre recombinaseunder the control of the aMHC promoter by crossing aMHC-Cremouse with the loxP-XFP transgenic (confetti) mouse30 In the confettimouse lox-P specific recombination resulted in the random expressionof one out of four fluorescent proteins (BFP GFP RFP or YFP) Celllysates were prepared from various tissues of either the confettiff mouse(without Cre -) or the double transgenic mouse (containing Cre 1) fol-lowed by western-blot analysis using anti-GFP antibodies As expectedGFP-related proteins were strongly detected in the heart lysate (seeSupplementary material online Figure S1) The lysate derived from lungtissue displayed a lower level of GFP-related proteins as compared tothe heart lysate probably due to low transcription activity of the aMHCpromoter in the lungs34 GFP-related protein expression was undetect-able in all of the other tissues tested (see Supplementary material onlineFigure S1)
32 In vitro analysis of cardiomyocytestreated with free fatty acidsTo reveal the possible role of ATF3 in cardiomyopathy induced by freefatty acids (FFA) (palmitate) we analysed adult cardiomyocyte primarycultures derived from ATF3ff and ATF3-cKO mice (ATF3-cKO geno-type ATF3ffaMHC-Cre) Cardiomyocytes were isolated by theLangendorff perfusion apparatus followed by overnight treatment with04 mM palmitate10 qRT-PCR analysis showed a 4- to 5-fold increase inATF3 induction by palmitate in the ATF3ff cardiomyocytes (Figure 1A)Immuno-fluorescence analysis of cardiomyocytes treated with either 1 h
or overnight palmitate displayed a robust ATF3 nuclear staining (Figure1B) Consistent with a previous study35 western-blot analysis of the car-diomyocytes lysate following overnight palmitate treatment demon-strated a significant induction of the ATF3 protein (Figure 1C) SinceATF3 induction is dependent on the activation of the p38-MAPK signal-ling pathway1026 we measured the p38-MAPK (p38) activation stateusing anti-phospho-p38 (p-p38) While p-p38 levels were increased fol-lowing palmitate treatment p38 remained unchanged independent ofthe treatment (Figure 1C) As expected neither ATF3 transcript (Figure1A) nor ATF3 protein (Figure 1C) was detected in the palmitate-treatedATF3-cKO cardiomyocytes Importantly palmitate treatment inducedthe expression of cardiac hypertrophic markers (ANP and BNP) inflam-mation markers (IL-6 and TNFa) a metabolic marker (PDK4) and anextracellular matrix (ECM) remodeling protein (TIMP1) (Figure 1A)Strikingly significantly higher level of induction of these genes wasobserved in the ATF3-cKO cardiomyocytes as compared with ATF3ff
cardiomyocytes (Figure 1A) In addition while the expression level of theinsulin receptor substrate IRS1 was induced 2-fold by palmitate treat-ment in ATF3ff IRS1 expression was suppressed in ATF3-cKO cardio-myocytes (Figure 1A)
Next we quantified the level of secreted inflammatory cytokines byanalysing the conditioned medium derived from palmitate-treated cardi-omyocytes The levels of IL-6 and TNFa were significantly higher in theconditioned medium derived from palmitate-treated ATF3-cKO cardio-myocytes as compared to their conditioned media levels derived frompalmitate-treated ATF3ff cardiomyocytes (Figure 1D)
Collectively the above-mentioned results support an increase of ATF3expression in cardiomyocytes in response to acute and chronic exposureto FFA The analysis of cardiomyocytes derived from wild type and ATF3-cKO genotypes in vitro is consistent with a possible protective role ofATF3 by dampening the cardiomyocytes remodeling response to FFA
33 The effect of high-fat diet on wild typeand ATF3-cKO miceTo determine whether cardiac ATF3 expression plays a role in responseto a high-fat diet (HFD)-regimen we used mice harbouring acardiomyocyte-selective deletion ATF3-cKO and their wild type (ATF3f
f) counterparts Mice were fed either a normal chow (NC 44 fat) or aHFD (60 fat) The HFD was initiated at 10 weeks of age and continuedfor 15 weeks Both ATF3ff and ATF3-cKO mice fed with a HFD gainedweight at significantly higher rates than NC-fed mice with no significantdifferences between the genotypes (see Supplementary material onlineFigure S2) In addition the levels of circulating total cholesterol and highdensity lipoprotein (HDL) were significantly higher in HFD-fed mice incomparison to NC-fed mice with no significant difference between thegenotypes (see Supplementary material online Figure S3A and B)
34 Cardiomyocyte-selective ATF3deficiency induces cardiac remodelingSince ATF3 is highly induced in cardiomyocytes that are treated withFFA (04 mM palmitate) we studied the expression of ATF3 in heart tis-sue following 15 weeks of a HFD regimen by qRT-PCR ATF3 expres-sion was induced 6-fold in the hearts of HFD-fed ATF3ff mice whencompared to either the NC-fed mice or to HFD-fed ATF3-cKO (Figure2A) We next performed immuno-histochemistry staining of heart slicesderived from NC- and HFD-fed mice with anti-ATF3 antibodies Whileno ATF3 staining was observed in the hearts derived from NC-fed mice
ATF3 expression in cardiomyocytes preserves homeostasis 137D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
or HFD-fed ATF3-cKO mice efficient ATF3 nuclear staining wasobserved in HFD-fed ATF3ff mice (Figure 2B)
In addition western-blot analysis using anti-ATF3 antibodies with car-diomyocyte cell lysate confirmed a high level of ATF3 protein expressionin HFD-fed as compared with NC-fed mice (Figure 2C and D) We nextexamined the cardiac expression levels of various gene specific markersas an indication for cardiac remodeling processes of a stressed heartHFD-fed ATF3-cKO but not ATF3ff mice exhibited significantly higherlevels of the foetal gene programming markers (ANP BNP and bMHC)when compared to the NC-fed wild type counterparts (Figure 2A)Concomitantly a-myosin heavy chain (aMHC) expression was signifi-cantly reduced in HFD-fed mice as compared to NC-fed mice independ-ent of their genotype (Figure 2A) The switch from aMHC to bMHC is anindication of the cardiac remodeling process as well as a cardiac disease
state Heart sections stained with wheat-germ hemagglutinin revealed asignificant increase in the cross sectional area in HFD-fed mice withlarger increased area in ATF3-cKO mice as compared with ATF3ff mice(Figure 2E and F) Taken together cardiac remodeling induced by a HFDis more pronounced in the ATF3-cKO mice than that in the controlATF3ff mice
We next examined fibrosis markers and found elevated expression ofcollagen 1a cTGF and a-SMA in HFD-fed mice as compared to the cor-responding NC-fed mice (Figure 3A) In all cases the ATF3-cKO miceshowed a more pronounced phenotype (Figure 3A) Western-blot analysiswith cell lysates derived from both NC-fed and HFD-fed mice confirmedincreased levels of a-SMA in HFD-fed mice but these high levels weremore noticeable in the lysate derived from HFD-fed ATF3-cKO mice(Figure 3B and C) Another fibrosis marker TGFb3 is significantly elevated
A C
B D
Figure 1 Free fatty acids affect the expression of hypertrophic inflammatory metabolic and fibrosis markers in primary cardiomyocytes Cardiomyocyteswere isolated from adult ATF3ff and ATF3-cKO mice and primary cultures were generated (A) Cardiomyocytes were exposed to 04 mM palmitate orvehicle control for overnight mRNA was extracted and the expression levels of the indicated genes were studied by qRT-PCR The mean ratio of palmitatetreated vs untreated cells for each genotype is shown The results represent the mean ratio plusmn SD (n = 5 independent preparations) (B) ATF3ff cardiomyo-cytes were exposed to 04 mM palmitate for either 1 h or overnight and then fixed and stained with anti-ATF3 (red) and anti-a-actinin (green) antibodiesNuclei were stained with DAPI (blue) (C) Western-blot analysis of cardiomyocytes lysate either palmitate treated overnight or left untreated probed withanti-ATF3 anti-p38 and anti-phospho-p38 antibodies The expression level of a-tubulin was used as loading control Each lane represents a lysate preparedfrom a pool derived from three independent cardiomyocytes culture (D) Isolated cardiomyocytes were treated as described in A Conditioned mediumwas collected and the levels of the indicated cytokines were assessed by ELISA The results represent the mean ratio between palmitate treated tountreatedplusmn SD (n = 5ndash6 independent preparations) P lt_ 005 comparing treated to untreated cells of the same genotype daggerP lt_ 005 comparing palmitate-treated cells between genotypes
138 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
in ATF3-cKO mice but not in the control ATF3ff mice (Figure 3A) againindicating that HFD has a more profound effect on ATF3-cKO mice
We next examined fibrosis histologically by Massonrsquos trichrome stain-ing This analysis confirmed a significant increased fibrosis in the hearts ofHFD-fed mice with ATF3-cKO mice exhibiting the highest fibrosis level(Figure 3D and E) Fibrosis is a result of the deposition of collagen in theECM The net effect of ECM degrading enzymes such as metalloprotei-nases (MMPs) and their inhibitors (TIMPs) determines the level of colla-gen deposition36 The levels of MMP9 were significantly decreased inHFD-fed mice as compared to NC-fed mice independent of the micegenotype (Figure 3A) In contrast TIMP1 levels were significantlyincreased in HFD-fed mice with considerably higher levels in the ATF3-cKO mice consistent with the finding that ATF3-cKO mice displayhigher level of fibrosis (Figure 3A)
Cardiac remodeling processes and diabetes are typically associatedwith increased inflammation We therefore measured several inflamma-tory markers in the hearts of NC- and HFD-fed ATF3ff and ATF3-cKOmice IL-6 expression was highly induced in the hearts of NC- and HFD-fed ATF3-cKO mice as compared with ATF3ff mice Significantly HFD-fed ATF3-cKO mice displayed the highest level (Figure 4A) SimilarlyTNFa was expressed at significantly higher levels in HFD-fed ATF3-cKOmice (Figure 4A) To identify the source of IL-6 and TNFa expression wehave isolated cardiomyocytes from mice fed with HFD for 15 weeks andexamined the cytokinersquos transcription and secretion levels by both qRT-PCR and ELISA respectively Indeed the transcription (Figure 4B) and
secretion (Figure 4C) levels of IL-6 and TNFa were significantly higher incardiomyocytes derived from HFD-fed ATF3-cKO mice as compared totheir ATF3ff cardiomyocytes counterparts To reveal whether theinflammatory response in the heart following a HFD affects circulatingIL-6 and TNFa levels we measured the cytokines in the serum of HFD-fed mice using ELISA Indeed a significant increase in IL-6 and TNFa lev-els were detected in the serum of ATF3-cKO mice (Figure 4D and Erespectively) Collectively we conclude that cardiomyocytes are thecells responsible at least in part for the expression and secretion of IL-6and TNFa in HFD-fed mice with significantly higher circulating levels inATF3 deficient mice
Lastly since inflammation is often associated with a maladaptive meta-bolic response we analysed the expression levels of various metabolicgenes in the heart The level of IRS1 a gene involved in insulin signallingwas significantly reduced in HFD-fed mice The reduction was more pro-nounced in the ATF3-cKO than in control mice (Figure 5A) Similarly thelevel of fatty acid translocase (CD36) was significantly higher in HFD-fedATF3ff mice IRS2 was much higher in the NC-fed ATF3-cKO mice thanin the control counterparts (NC-fed ATF3ff mice) consistent with ourprevious data that its gene expression is suppressed by ATF337
Interestingly IRS2 is induced in HFD-fed ATF3ff mice to comparable levelas HFD-fed ATF3-cKO mice The biological implication of this finding isnot clear One possibility is that induction of IRS2 a pro-survival gene bya HFD is an attempt by the cardiac cells to cope with the HFD stress thisadaptation is not necessary in the ATF3-cKO heart since IRS2 levels are
A
B
C
D
E
F
Figure 2 HFD-fed ATF3-cKO mice display increased expression of cardiac remodeling markers ATF3ff and ATF3-cKO mice were fed with either normalchow (NC) for 25 weeks or switched to a 15-week HFD regimen at 10 weeks of age At endpoint mice were sacrificed and hearts were excised (A) mRNAwas extracted from ventricles and the expression level of ATF3 and the indicated remodeling markers were measured by qRT-PCR Expression levels arepresented as relative values (compared to NC-fed ATF3ff mice determined as 1 for each gene) The results represent the meansplusmn SD (n = 6ndash8group) (B)Immuno-histochemistry analysis of paraformaldehyde fixed heart slices as described in A Representative nuclear staining is indicated by yellow arrowsScale bar = 50lm (C) Western-blot analysis of heart cell lysate probed with anti-ATF3 antibodies The expression level of a-tubulin was used as loadingcontrol (D) Quantification of ATF3 expression from C The results represent the meansplusmn SD (n = 3ndash5group) ratio of ATF3 expression of the indicatedgenotypes relative to NC ATF3ff determined as 1 (E) Ventricles sections were stained with TRITC-labelled wheat germ agglutinin and the cell size was ana-lysed Scale bar = 20lm (F) Quantification of cell size from E The results represent the meansplusmn SD (n = 6ndash8group) P lt_ 005 difference between dietsdaggerP lt_ 005 difference between genotypes
ATF3 expression in cardiomyocytes preserves homeostasis 139D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
already high The expression levels of other lipid and glycolytic regulatorssuch as peroxisome proliferator-activated receptor alpha (PPARa) carni-tine palmitoyltransferase 1 and 2 (CPT12) and pyruvate dehydrogenasekinase 4 (PDK4) were all elevated in HFD-fed mice with no significant dif-ference between the genotypes (Figure 5A) Lastly the level of the glucosetransporter 4 (Glut4) were unaffected by both diet and genotype at themRNA level (Figure 5A) However Glut4 was significantly elevated at theprotein level in HFD-fed mice with no significant difference between thegenotypes (see Supplementary material online Figure S4A and B)Collectively the expression of key metabolic genes was unalteredbetween the various genotypes in response to HFD In view of thechanges observed in metabolic genes in cardiomyocytes treated with pal-mitate (Figure 1A) this result was unexpected Thus we hypothesizedthat ATF3-dependent alterations in metabolic gene expression in cardio-myocytes may be masked by non-cardiomyocytes cells in the heartTherefore we studied the metabolic gene expression changes in isolatedcardiomyocytes derived from HFD- and NCndashfed mice from both ATF3ff
and ATF3-cKO mice (Figure 5B) Indeed the expression of all metabolicgenes tested was augmented in cardiomyocytes derived from HFD-fed ascompared with NC-fed mice Importantly the expression level of CD36CPT1 and CPT2 were significantly upregulated in HFD-fed ATF3-cKOmice as compared with their ATF3ff counterparts (Figure 5B) In contrastthe induction of both IRS1 and IRS2 were significantly lower in cardio-myocytes derived from HFD-fed ATF3-cKO mice (Figure 5B)Collectively the deleterious effects of HFD on cardiac remodeling fibro-sis and inflammation are more pronounced in ATF3-cKO mice suggest-ing that cardiac ATF3 plays a protective role in the HFD paradigm
35 HFD-induced cardiac dysfunctionNext we characterized the effect of HFD and cardiac remodeling oncardiac structure and function in ATF3ff and ATF3-cKO mice by
echocardiography Mice fed with NC had normal heart dimensions andfractional shortening (FS) (Figure 6A and B) HFD significantly reduced FSin both genotypes of HFD-fed mice as compared to corresponding NC-fed mice (Figure 6B) However the reduction was significantly more pro-nounced in the ATF3-cKO mice suggesting deteriorated systolic func-tion in these mice (Figure 6B) The reduced FS in the HFD-fed ATF3-cKO mice seems to be largely driven by an increase in LV internal end-systolic (LVIDs) dimension (Figure 6A) Heart rate was slightly increasedin the HFD-fed mice however this increase cannot explain the cardiacdysfunction observed in ATF3-cKO mice (Figure 6B) In addition weused cardiac MRI the gold standard for LV function and mass asses-sment38 LV mass was significantly higher in the ATF3-cKO mice follow-ing a 15-week HFD regimen (Figure 6C) In addition end-diastolic andend-systolic volumes were significantly larger in ATF3-cKO mice as com-pared to ATF3ff mice Moreover ATF3-cKO mice displayed a signifi-cantly lower ejection fraction as compared to ATF3ff mice (Figure 6D)These results are consistent with the reduced cardiac function found inthe echocardiographic analysis Images of mid-ventricular short-axis(Figure 6E) and long-axis (Figure 6E) slices are demonstrating the cardiaceccentric pattern observed in HFD-fed ATF3-cKO mice Collectivelythe results from the MRI show a greater increase in LV mass and volumein the ATF3-cKO mice as compared to the ATF3ff mice following aHFD suggesting fundamental structural differences in the heart betweenthe ATF3-cKO and ATF3ff mice
36 Impaired glucose tolerance in HFDATF3-cKO miceLastly we measured fasting glucose levels to assess the diabetic state ofthe mice As expected higher glucose levels were observed in HFD-fedmice as compared to NC-fed counterparts Interestingly the
A B
C
D
E
Figure 3 HFD-fed ATF3-cKO mice have higher level of fibrosis in the heart ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) mRNA wasextracted from ventricles and the expression levels of the indicated fibrosis markers were measured by qRT-PCR The expression levels are presented asrelative values (compared to NC-fed ATF3ff mice determined as 1 for each gene n = 6ndash8group) (B) Western-blot analysis of heart cell lysate probed withanti-a-SMA antibody The expression level of a-tubulin was used as loading control (C) Quantification of aSMA expression from B The results representthe meansplusmn SD (n= 3ndash5group) ratio of aSMA expression of the indicated genotypes relative to NC ATF3ff determined as 1 (D) Paraffin-embedded heartsections were stained with Massonrsquos trichrome to visualize fibrosis Scale bar = 100 lm (E) The level of fibrosis () stained by Massonrsquos trichrome was quan-tified and the results represent the meansplusmn SD (n = 6ndash8group) P lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes
140 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
hyperglycemia state of the ATF3-cKO mice was significantly higher thanin the ATF3ff mice following 15 weeks of HFD (Figure 7A) Consistentlythe glucose tolerance test (GTT) revealed that HFD-fed ATF3-cKOmice displayed higher blood glucose levels and longer glucose clearancetime when compared to HFD-fed ATF3ff whereas NC-fed ATF3ff andATF3-cKO mice had normal glucose clearance (Figure 7B)
Interestingly whole body ATF3-KO mice fed with HFD also displayedimpaired glucose clearance (see Supplementary material onlineFigure S5A)39 Previously we reported that decreased glucose tolerancein whole body ATF3-KO mice is at least in part due to reduced insulingene expression in b-cells (see Supplementary material online FigureS5B)39 To determine insulin level in HFD-fed ATF3-cKO mice weanalysed blood insulin levels by ELISA In both HFD-fed ATF3ff andATF3-cKO mice the serum insulin levels were significantly higher ascompared to their NC-fed counterpart mice with the ATF3-cKO micedisplaying the highest insulin levels (Figure 7C) Together these resultssuggest that peripheral tissues of ATF3-cKO mice have reduced ability
to uptake glucose despite their high insulin level in the blood Next wesought to measure the protein levels of components along the insulinpathway Akt and Foxo340 Both Akt and Foxo3 protein levels were sim-ilar in HFD-fed mice independent of the genotype (see Supplementarymaterial online Figure S4A and C) Interestingly phosphorylation of Akt(Ser473) which indicates the activity of the insulin pathway was slightlyelevated in HFD-fed mice however this did not reach statistical signifi-cance and was similar in both of the genotypes (see Supplementary material online Figure S4D) To measure the activity of the insulin signallingpathway following insulin stimulation we used the ratio between phos-phorylated Akt (Ser473) and total Akt expression HFD-fed mice wereinjected with insulin (10 Ukg) and the heart liver and skeletal musclewere harvested 15 min after the injection (Figure 7D and E and seeSupplementary material online Figure S6AndashD) Insulin injection resultedin a significant increase of pAktAkt ratio in the hearts of ATF3ff micewhile no significant change was observed in Akt phosphorylation inATF3-cKO mice (Figure 7D and E) Similarly low Akt phosphorylation
A
D E
B C
Figure 4 HFD-fed ATF3-cKO mice display inflammatory response in the heart (A) ATF3ff and ATF3-cKO mice were fed as described in Figure 2 mRNAwas extracted from ventricles and the expression levels of the indicated inflammatory markers were measured by qRT-PCR Expression levels are presentedas relative values (compared to NC-fed ATF3ff mice determined as 1 for each gene n= 6ndash8group) (B) Mice were fed as described in Figure 2Cardiomyocytes were isolated and primary cultures were generated mRNA was extracted from an overnight culture and the expression levels of IL-6 andTNFa were studied by qRT-PCR The mRNA level obtained in cardiomyocytes derived from NC-fed ATF3ff mice was determined as 1 and the relativeexpression of all other treatment is shown (n = 5ndash6 independent cardiomyocyte preparations) (C) Conditioned media was collected from cardiomyocytesdescribed in B and ELISA assay for the indicated cytokines is shown Cytokines level is presented as relative values to NC-fed ATF3ff mice determined as 1for each cytokine (n = 5ndash6 independent preparations) (D and E) Blood was collected from HFD-fed mice and IL-6 and TNFa serum levels were quantifiedby ELISA (n = 8ndash12group) The results represent the meansplusmn SD P lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes
ATF3 expression in cardiomyocytes preserves homeostasis 141D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
levels were also observed in both the liver and skeletal muscle of theATF3-cKO mice however this did not reach statistically significance (seeSupplementary material online Figure S6AndashD) Consistently palmitate-treated cardiomyocytes derived from ATF3-cKO mice displayedreduced phosphorylation of Akt following 30-min insulin stimulation(see Supplementary material online Figure S6E)
Collectively our findings revealed a surprising phenomenon that car-diac ATF3 deficiency results in increased cardiac remodeling leading topotentiation of local and peripheral insulin resistance
4 Discussion
A HFD is a commonly used regimen to induce T2D The majority of theliterature focuses on diabetic symptoms without addressing the issue ofT2D-induced cardiomyopathy Even when it is addressed there is noclear consensus regarding the extent of cardiac effects imposed byHFD41 Diet-induced obesity in mouse models is associated with globalinsulin resistance and increased myocardial fatty acid utilization how-ever the reported phenotypes on cardiac function vary from extensivehypertrophy and heart failure4 to lack of cardiac dysfunction4243 Thesevariations have been attributed to diet regimens andor sensitivity ofmouse strains to a HFD In this report we present evidence that cardiacATF3 deficiency exacerbates the deleterious effect of a HFD on theheart such as maladaptive cardiac remodeling fibrosis inflammation andcardiac dysfunction Our work may provide a potential molecular insightto variations in the literature regarding the impact of HFD on the heartAs ATF3 is a stress-inducible gene we propose that its proper inductionin the cardiomyocytes is protective and helps the heart to cope withHFD stress It is possible that the induction of ATF3mdashits expressionlevel duration and kineticsmdashvaries in different HFD regimens andormouse strains contributing to the variations observed in the literature
HFD regimen induced a significant higher level of fibrosis in the heartof ATF3-cKO mice We have identified TIMP1 as a possible mechanismto explain the increase in fibrosis in ATF3-cKO cardiomyocytes Severallines of evidence suggest that TIMP1 is possibly an ATF3 target gene
First we observed an increase in TIMP1 transcription in the whole heartand in isolated cardiomyocytes (Figures 1A and 3A) Second TIMP 1 3and 4 were identified as significantly highly expressed genes in the heartof ATF3-KO mice as compared with wild type mice following aorticbanding20 Third a functional AP-1 regulatory element within the TIMP1promoter was described44 Indeed preliminary results in our lab suggestthat ATF3 downregulates the TIMP1 promoter (data not shown) Thusit is possible that TIMP1 is suppressed by ATF3 and therefore results in anet increase in MMPs activity that helps to maintain ECM formationdeg-radation homeostasis
Our findings are consistent with previous studies demonstrating that adeficiency in ATF3 can promote cardiac maladaptive changes which leadto cardiac dilatation and hypertrophy in an aortic banding model1920 Incontrast a different study using a pressure overload model reportedthat ATF3 induces a maladaptive cardiac response17 These seeminglyconflicting data may be explained by the different patterns of ATF3expression induced by the different models In the pressure overloadmodel the acute and transient expression of ATF3 in cardiomyocytesinitiates a cascade of maladaptive changes17 whereas in the aortic band-ing model and the HFD model described here ATF3 expression is longlasting
In this context this is the first report using mice with cardiac-selectivedeficiency of ATF3 (ATF3-cKO) Previous reports on the roles of ATF3in cardiac function were carried out using whole body knockout ofATF325 or transgenic mice ectopically expressing ATF3 in cardiomyo-cytes16 A key advancement of ATF3-cKO mice over the whole bodyknockout mice is that they provide insights into the function of ATF3selectively in cardiomyocytes without the complication of ATF3 defi-ciency in other cells Thus the ATF3-cKO mouse is a useful tool to bet-ter understand the function of cardiac ATF3
Consistent with our previous report25 here we showed that cardiacATF3 deficiency has a more severe HFD-induced diabetic phenotypethan the control wild type mice However while reduced insulin genetranscription in ATF3-deficient b-cells was observed25 our current dataindicates that a lack of insulin is not the explanation for the glucose intol-erance found in the HFD-fed ATF3-cKO mice In contrast these mice
A B
Figure 5 HFD-fed ATF3-cKO mice show metabolic response in the heart ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) mRNA wasextracted from ventricles and the expression levels of the indicated metabolic markers were measured by qRT-PCR Expression levels are presented as rela-tive values (compared to NC-fed ATF3ff mice determined as 1 for each gene n= 6ndash8group) (B) Cardiomyocytes were isolated and plated on laminin for1 h followed by mRNA preparation (n = 4ndash5 independent preparations) The expression levels of the indicated metabolic markers were measured by qRT-PCR Expression levels are presented as relative values (to NC-fed ATF3ff mice determined as 1 for each gene) The results represent the meansplusmn SDP lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes
142 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
had higher blood insulin level (Figure 7C) Therefore upon HFD chal-lenge cardiac-deficiency of ATF3 exacerbates HFD-induced T2D Thislsquoperipheral effectrsquo is surprising since ATF3 is selectively deleted in thecardiomyocytes
The question then is how can we explain the effect of cardiacATF3 deficiency on peripheral glucose intolerance One possibility isthe reduced IRS1 expression in ATF3-deficient cardiomyocytes bothin vivo and in vitro IRS1 suppression may lead to impaired insulin sig-nalling in the heart thereby exacerbating glucose intolerance Indeedwe observed that HFD-fed ATF3-cKO mice displayed insulin resist-ance upon insulin challenge when compared to HFD-fed ATF3ff
mice In addition palmitate-treated cardiomyocytes lacking ATF3 dis-played lower insulin signalling as compared to their wild type coun-terparts Since the heart is only a minor glucose consuming organ incomparison to skeletal muscle cells or the brain the peripheral sys-temic effect cannot be solely based on this observation A case inpoint transgenic mice with insulin receptor (IR) deficiency in theheart do not display a diabetic phenotype4546 Another attractive
possibility for the peripheral effect of cardiac ATF3-deletion is itsimpact on inflammation It is well established that low-grade inflam-mation years in advance of the disease onset is involved in the patho-genic processes causing T2D8 Mediators of inflammation such asTNFa interleukin (IL)-1b the IL-6 family of cytokines IL-18 and cer-tain chemokines have been proposed to be involved in the eventscausing both forms of diabetes (T1D and T2D)8 Interestingly wehave shown that IL-6 a well-known ATF3 target gene47 is known toenhance its transcription and secretion of cytokines such as IL-6 andTNFa in palmitate-treated primary cultures of ATF3-cKO cardio-myocytes as well as in cardiomyocytes derived from HFD-fed ATF3-cKO mice IL-6 has been proposed to play a role in glucose homeo-stasis and metabolism directly and indirectly via its action on skeletalmuscle cells adipocytes hepatocytes pancreatic b-cells and neuro-endocrine cells48
Therefore we hypothesize that elevated levels of inflammatory cyto-kines in ATF3-cKO mice mediate a cross-talk in peripheral tissues leadingto impaired glucose tolerance Although circulating IL-6 and TNFa
A
B
C
D
E
F
Figure 6 HFD-fed ATF3-cKO mice present deteriorated heart function ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) At endpointmice hearts were examined by micro ultrasound The following parameters were measured interventricular septal end diastole (IVSd) left ventricular pos-terior wall end diastole (LVPWd) maximal left ventricular internal end-diastole (LVIDd) end-systole (LVIDs) and heart rate The results represent themeansplusmn SD (n = 6ndash8 in each group as shown in panel B) (B) Heart function was assessed by calculating percentage of fractional shortening (FS) accordingto FS () = [(LVDd-LVDs)LVDd]100 The results represent the meansplusmn SD (n = 6ndash8 in each group) (C and D) Left ventricular cardiac volumes massand function were examined by a cardiac MRI The following parameters were measured left ventricular end-diastolic (LVEDV) and left ventricular end-sys-tolic volume (LVESV) left ventricular (LV) mass and ejection fraction (EF) The results represent the meansplusmn SD (n = 7 in each group) (E and F)Representative images of mid-ventricular short-axis (E) and long-axis (F) slice at peak diastole and systole P lt_ 005 difference between diets daggerP lt_ 005 dif-ference between genotypes
ATF3 expression in cardiomyocytes preserves homeostasis 143D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
cytokines may be derived from different tissues we provide ample evi-dence that ATF3-defficient cardiomyocytes produce and secrete higherlevels of these cytokines as compared to their wild type counterpartsThis indicates a lsquocardiac-to-peripheralrsquo effect in T2D-induced cardiomyop-athy This is a reverse direction from the lsquoperipheral-to-cardiacrsquo effect dis-cussed in the introduction where inflammation in the peripheral tissuesleads to cardiac dysfunction Since inflammatory cytokines are soluble
factors they can exert a systemic effect This is why it makes sense thattheir effect can go both ways Thus diabetes and cardiomyopathy are apart of a vicious cycle Previously it was shown that cardiac specific dele-tion of the thyroid hormone complex mediator MED13 results in aperipheral effect upon HFD Cardiac MED13 deficiency suppresses meta-bolic genes in the heart which leads to a decrease in energy expenditureand an obese phenotype most likely by the secretion of cardiokines that
A
B
C
D
E
Figure 7 HFD-fed ATF3-cKO mice exhibit impaired glucose tolerance ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) Fasting (6 h)blood glucose concentrations (mgdl) The results represent the meansplusmn SD (n = 20ndash30group) P lt_ 005 difference between diets daggerP lt_ 005 differencebetween genotypes (B) Glucose tolerance test (GTT) Values are presented as relative to initial valueplusmn SD (n = 20ndash25group) P lt_ 005 difference betweendiets daggerP lt_ 005 difference between genotypes (C) Serum insulin concentrations were quantified by ELISA Values are presented as relative values (com-pared to NC-fed ATF3ff mice determined as 1 n = 15ndash20group) P lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes (D and E)Effect of HFD on phosphorylated Akt (Ser473 pAkt) to Akt ratio levels in heart lysate following either saline or insulin injection (10 U1 kg) is shownRepresentative blots (D) Densitometric analysis is presented as means ratio (n = 3ndash6group) pAktAkt ratio is presented as relative values (compared toHFD-fed saline injected ATF3ff mice determined as 1) P lt_ 005 insulin vs saline treatment daggerP lt_ 005 difference between genotypes (E)
144 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
mediates the cross talk between the heart and peripheral tissues49ndash51
Here we also observed increased levels of natriuretic peptides (ANP andBNP) which together with IL-6 and TNFa may mediate the cross-talk withthe peripheral tissues to induce glucose intolerance
Does this finding have any relevance to cardiac disease in humansSeveral lines of evidence suggests that congestive heart failure in patientsis associated with an increase in blood glucose levels52 In addition highblood glucose levels in non-diabetic patients with acute myocardialinfarction serves as a prognostic predictor of long-term mortality5354
Thus here we provide evidence that high blood glucose levels are a partof the symptoms of cardiac remodeling process
Collectively our findings suggest that cardiac ATF3 expression isresponsible for an adaptive cardiac response to the stressful environmentunder a HFD (Figure 8) ATF3 dampens the inflammatory response andsuppresses the expression of ECM remodeling genes and thus amelioratescardiac function In HFD-fed mice with ATF3 deficiency or in mice withcardiac remodeling the inflamed heart affects also peripheral tissuesthereby worsening T2D in peripheral tissues resulting in elevated bloodglucose level and turning on a vicious cycle to worsen cardiomyopathy
Supplementary material
Supplementary material is available at Cardiovascular Research online
AcknowledgementsThe authors wish to thank Drs Keren Bitton-Worms and Ran Rostokerand Prof Derek LeRoith from the Rambam Medical Center for their
assistance in GTT and ITT Dr Edith Suss Toby Maya Holdengreber andLior Liba from the BCF Bioimaging Center (Faculty of MedicineTechnion) for their assistance with imaging and image analysis Prof DEMichelle from University of Michigan for cardiomyocytes culture techni-que and Prof Yehiel Zick from the Weizmann institute of Sciences forcritical comments
Conflict of interest none declared
FundingThis work was partially supported by the United States-Israel BinationalGrant 2009179 to AA and TH In addition we wish to thank the MarsPittsburgh Fund for Medical Research to AA the Department of Defensegrant W81XWH-14-1-0179 to TH and to D Dan and Betty Kahn travelgrant support to LK
References
1 Roger VL Go AS Lloyd-Jones DM Benjamin EJ Berry JD Borden WB Bravata DMDai S Ford ES Fox CS Fullerton HJ Gillespie C Hailpern SM Heit JA Howard VJKissela BM Kittner SJ Lackland DT Lichtman JH Lisabeth LD Makuc DM MarcusGM Marelli A Matchar DB Moy CS Mozaffarian D Mussolino ME Nichol GPaynter NP Soliman EZ Sorlie PD Sotoodehnia N Turan TN Virani SS Wong NDWoo D Turner MB American Heart Association Statistics C Stroke Statistics SHeart disease and stroke statisticsndash2012 update a report from the American HeartAssociation Circulation 2012125e2ndashe220
2 Wilson C Diabetes ACCORD 5-year outcomes of intensive glycemic control NatRev Endocrinol 20117314
3 Ernande L Derumeaux G Diabetic cardiomyopathy myth or reality Arch CardiovascDis 2012105218ndash225
4 Battiprolu PK Hojayev B Jiang N Wang ZV Luo X Iglewski M Shelton JM GerardRD Rothermel BA Gillette TG Lavandero S Hill JA Metabolic stress-induced
InsulinGF
Cytokines
FFA
GlucoseATF3
Cardiac
Remodeling
Factors
Paracrine action
Collagens
and ECM
proteins
Fibrosis
Cardiac
Dysfunction
Systemic Effects
Glucose Tolerance
TNFα
IL-6
Cardiomyocyte
ATF3
Homeostasis
Homeostasis
TNFα
IL-6
TIMP1
BNP
ANP
Figure 8 lsquoPeripheral-to-cardiacrsquo and lsquoCardiac-to-peripheralrsquo model scheme Schematic diagram showing the ATF3 adaptive role in maintaining hearthomeostasis following HFD-induced T2D In contrast stressful environment under HFD leads to maladaptive cardiac response in the absence of ATF3Cardiac remodeling caused by ATF3 deficiency induces an increase in cardiac ECM remodeling and inflammatory genes and mediates the communication ofcardiomyocytes with heart resident cells potentially mediated by IL-6 TNFa ANP BNP and TIMP1 In this situation increased cardiac fibrosis and inflamma-tory response worsen cardiac function The inflamed failing heart mediates the communication of cardiomyocytes with distant organs as well thereby affect-ing diabetic outcome and glucose homeostasis
ATF3 expression in cardiomyocytes preserves homeostasis 145D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
activation of FoxO1 triggers diabetic cardiomyopathy in mice J Clin Invest20121221109ndash1118
5 Mandavia CH Aroor AR Demarco VG Sowers JR Molecular and metabolic mecha-nisms of cardiac dysfunction in diabetes Life Sci 201392601ndash608
6 Ventura-Clapier R Garnier A Veksler V Energy metabolism in heart failure J Physiol20045551ndash13
7 Guo S Insulin signaling resistance and the metabolic syndrome insights from mousemodels into disease mechanisms J Endocrinol 2014220T1ndashT23
8 Montane J Cadavez L Novials A Stress and the inflammatory process a major causeof pancreatic cell death in type 2 diabetes Diabetes Metab Syndr Obes 2014725ndash34
9 Ashrafian H Frenneaux MP Opie LH Metabolic mechanisms in heart failureCirculation 2007116434ndash448
10 Tanabe K Liu Y Hasan SD Martinez SC Cras-Meneur C Welling CM Bernal-MizrachiE Tanizawa Y Rhodes CJ Zmuda E Hai T Abumrad NA Permutt MA Glucose andfatty acids synergize to promote B-cell apoptosis through activation of glycogen syn-thase kinase 3beta independent of JNK activation PLoS One 20116e18146
11 Heineke J Molkentin JD Regulation of cardiac hypertrophy by intracellular signallingpathways Nat Rev Mol Cell Biol 20067589ndash600
12 Hai T Wolford CC Chang YS ATF3 a hub of the cellular adaptive-response net-work in the pathogenesis of diseases is modulation of inflammation a unifying com-ponent Gene Expr 2010151ndash11
13 Takii R Inouye S Fujimoto M Nakamura T Shinkawa T Prakasam R Tan K HayashidaN Ichikawa H Hai T Nakai A Heat shock transcription factor 1 inhibits expression ofIL-6 through activating transcription factor 3 J Immunol 20101841041ndash1048
14 Giraldo A Barrett OP Tindall MJ Fuller SJ Amirak E Bhattacharya BS Sugden PHClerk A Feedback regulation by Atf3 in the endothelin-1-responsive transcriptomeof cardiomyocytes Egr1 is a principal Atf3 target Biochem J 2012444343ndash355
15 Hasin T Elhanani O Abassi Z Hai T Aronheim A Angiotensin II signaling up-regulates the immediate early transcription factor ATF3 in the left but not the rightatrium Basic Res Cardiol 2011106175ndash187
16 Koren L Elhanani O Kehat I Hai T Aronheim A Adult cardiac expression of theactivating transcription factor 3 ATF3 promotes ventricular hypertrophy PLoS One20138e68396
17 Koren L Alishekevitz D Elhanani O Nevelsky A Hai T Kehat I Shaked Y AronheimA ATF3-dependent cross-talk between cardiomyocytes and macrophages promotescardiac maladaptive remodeling Int J Cardiol 2015198232ndash240
18 Okamoto Y Chaves A Chen J Kelley R Jones K Weed HG Gardner KL Gangi LYamaguchi M Klomkleaw W Nakayama T Hamlin RL Carnes C Altschuld R BauerJ Hai T Transgenic mice with cardiac-specific expression of activating transcriptionfactor 3 a stress-inducible gene have conduction abnormalities and contractile dys-function Am J Pathol 2001159639ndash650
19 Zhou H Shen DF Bian ZY Zong J Deng W Zhang Y Guo YY Li H Tang QZActivating transcription factor 3 deficiency promotes cardiac hypertrophy dysfunc-tion and fibrosis induced by pressure overload PLoS One 20116e26744
20 Zhou H Guo H Zong J Dai J Yuan Y Bian ZY Tang QZ ATF3 regulates multipletargets and may play a dual role in cardiac hypertrophy and injury Int J Cardiol2014174838ndash839
21 Yang J Yang CJ Yang J Fan ZX ATF3 a promotion effect or a inhibition effect in car-diac maladaptive remodeling Int J Cardiol 2015201245ndash246
22 Koren L Shaked Y Aronheim A Response letter ldquoATF3 a promoter or inhibitor ofcardiac maladaptive remodelingrdquo Int J Cardiol 2015201692
23 Ma ZG Wei WY Xu SC Zhang WB Dai J Tang QZ ATF3 a potential target forcardiac maladaptive remodeling Int J Cardiol 201620250ndash51
24 Zhou H Yuan Y Ni J Guo H Deng W Bian ZY Tang QZ Pleiotropic and puzzlingeffects of ATF3 in maladaptive cardiac remodeling Int J Cardiol 201620687ndash88
25 Zmuda EJ Qi L Zhu MX Mirmira RG Montminy MR Hai T The roles of ATF3 anadaptive-response gene in high-fat-diet-induced diabetes and pancreatic beta-celldysfunction Mol Endocrinol 2010241423ndash1433
26 Lu D Chen J Hai T The regulation of ATF3 gene expression by mitogen-activatedprotein kinases Biochem J 2007401559ndash567
27 Hartman MG Lu D Kim ML Kociba GJ Shukri T Buteau J Wang X Frankel WLGuttridge D Prentki M Grey ST Ron D Hai T Role for activating transcription fac-tor 3 in stress-induced beta-cell apoptosis Mol Cell Biol 2004245721ndash5732
28 Wolford CC McConoughey SJ Jalgaonkar SP Leon M Merchant AS Dominick JLYin X Chang Y Zmuda EJ OrsquoToole SA Millar EK Roller SL Shapiro CL OstrowskiMC Sutherland RL Hai T Transcription factor ATF3 links host adaptive response tobreast cancer metastasis J Clin Iinvest 20131232893ndash2906
29 Agah R Frenkel PA French BA Michael LH Overbeek PA Schneider MD Generecombination in postmitotic cells Targeted expression of Cre recombinase pro-vokes cardiac-restricted site-specific rearrangement in adult ventricular musclein vivo J Clin Invest 1997100169ndash179
30 Livet J Weissman TA Kang H Draft RW Lu J Bennis RA Sanes JR Lichtman JWTransgenic strategies for combinatorial expression of fluorescent proteins in thenervous system Nature 200745056ndash62
31 Davis J Maillet M Miano JM Molkentin JD Lost in transgenesis a userrsquos guidefor genetically manipulating the mouse in cardiac research Circ Res2012111761ndash777
32 Heiberg E Sjogren J Ugander M Carlsson M Engblom H Arheden H Design andvalidation of segment ndash freely available software for cardiovascular image analysisBMC Med Imaging 2010101
33 Kabaeva Z Zhao M Michele DE Blebbistatin extends culture life of adult mouse car-diac myocytes and allows efficient and stable transgene expression Am J Physiol HeartC 2008294H1667ndash1674
34 Subramaniam A Jones WK Gulick J Wert S Neumann J Robbins J Tissue-specificregulation of the alpha-myosin heavy chain gene promoter in transgenic mice J BiolChem 199126624613ndash24620
35 Cunha DA Hekerman P Ladriere L Bazarra-Castro A Ortis F Wakeham MCMoore F Rasschaert J Cardozo AK Bellomo E Overbergh L Mathieu C Lupi R HaiT Herchuelz A Marchetti P Rutter GA Eizirik DL Cnop M Initiation and executionof lipotoxic ER stress in pancreatic beta-cells J Cell Sci 20081212308ndash2318
36 Wynn TA Ramalingam TR Mechanisms of fibrosis therapeutic translation forfibrotic disease Nat Med 2012181028ndash1040
37 Li D Yin X Zmuda EJ Wolford CC Dong X White MF Hai T The repression ofIRS2 gene by ATF3 a stress-inducible gene contributes to pancreatic beta-cell apop-tosis Diabetes 200857635ndash644
38 Myerson SG Bellenger NG Pennell DJ Assessment of left ventricular mass by cardi-ovascular magnetic resonance Hypertension 200239750ndash755
39 Zmuda EJ Viapiano M Grey ST Hadley G Garcia-Ocana A Hai T Deficiency ofAtf3 an adaptive-response gene protects islets and ameliorates inflammation in asyngeneic mouse transplantation model Diabetologia 2010531438ndash1450
40 Riehle C Abel ED Insulin signaling and heart failure Circ Res 20161181151ndash116941 Schreyer SA Wilson DL LeBoeuf RC C57BL6 mice fed high fat diets as models for
diabetes-accelerated atherosclerosis Atherosclerosis 199813617ndash2442 Brainard RE Watson LJ Demartino AM Brittian KR Readnower RD Boakye AA
Zhang D Hoetker JD Bhatnagar A Baba SP Jones SP High fat feeding in mice isinsufficient to induce cardiac dysfunction and does not exacerbate heart failure PLoSOne 20138e83174
43 Gupte AA Minze LJ Reyes M Ren Y Wang X Brunner G Ghosn M Cordero-Reyes AM Ding K Pratico D Morrisett J Shi ZZ Hamilton DJ Lyon CJ Hsueh WAHigh-fat feeding-induced hyperinsulinemia increases cardiac glucose uptake and mito-chondrial function despite peripheral insulin resistance Endocrinology20131542650ndash2662
44 Logan SK Garabedian MJ Campbell CE Werb Z Synergistic transcriptionalactivation of the tissue inhibitor of metalloproteinases-1 promoter via functionalinteraction of AP-1 and Ets-1 transcription factors J Biol Chem1996271774ndash782
45 Belke DD Betuing S Tuttle MJ Graveleau C Young ME Pham M Zhang DCooksey RC McClain DA Litwin SE Taegtmeyer H Severson D Kahn CR AbelED Insulin signaling coordinately regulates cardiac size metabolism and contractileprotein isoform expression J Clin Invest 2002109629ndash639
46 Fernandez AM Kim JK Yakar S Dupont J Hernandez-Sanchez C Castle AL FilmoreJ Shulman GI Le Roith D Functional inactivation of the IGF-I and insulin receptors inskeletal muscle causes type 2 diabetes Genes Dev 2001151926ndash1934
47 Gilchrist M Thorsson V Li B Rust AG Korb M Roach JC Kennedy K Hai TBolouri H Aderem A Systems biology approaches identify ATF3 as a negative regu-lator of Toll-like receptor 4 Nature 2006441173ndash178
48 Kristiansen OP Mandrup-Poulsen T Interleukin-6 and diabetes the good the bad orthe indifferent Diabetes 200554(Suppl 2)S114ndashS124
49 Grueter CE van Rooij E Johnson BA DeLeon SM Sutherland LB Qi X Gautron LElmquist JK Bassel-Duby R Olson EN A cardiac microRNA governs systemic energyhomeostasis by regulation of MED13 Cell 2012149671ndash683
50 Baskin KK Grueter CE Kusminski CM Holland WL Bookout AL Satapati S KongYM Burgess SC Malloy CR Scherer PE Newgard CB Bassel-Duby R Olson ENMED13-dependent signaling from the heart confers leanness by enhancing metabo-lism in adipose tissue and liver EMBO Mol Med 201461610ndash1621
51 Nakamura M Sadoshima J Heart over mind metabolic control of white adipose tis-sue and liver EMBO Mol Med 201461521ndash1524
52 Tenenbaum A Fisman EZ Impaired glucose metabolism in patients with heart failurepathophysiology and possible treatment strategies Am J Cardiovasc Drugs20044269ndash280
53 Suskin N McKelvie RS Burns RJ Latini R Pericak D Probstfield J Rouleau JL SigouinC Solymoss CB Tsuyuki R White M Yusuf S Glucose and insulin abnormalitiesrelate to functional capacity in patients with congestive heart failure Eur Heart J2000211368ndash1375
54 Aronson D Hammerman H Kapeliovich MR Suleiman A Agmon Y Beyar RMarkiewicz W Suleiman M Fasting glucose in acute myocardial infarction incremen-tal value for long-term mortality and relationship with left ventricular systolic func-tion Diabetes Care 200730960ndash966
146 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022

217 Palmitate-treated cardiomyocytesForty-five minutes after plating the medium was replaced with a culturemedium containing either 04 mM palmitate or BSA (control) Palmitatestock solution (20 mM) was diluted in 001 M NaOH by heating at 70Cfor 30 min as described10 Following overnight culture conditionedmedium was collected and cardiomyocytes were harvested for RNAextraction In addition overnight palmitate- or BSA-treated cardiomyo-cytes were incubated with 100 nM insulin (I-5500 SigmandashAldrich) for30 min and were then harvested for protein extraction
218 Enzyme-linked immuno-absorbentassayInterleukin 6 (IL-6) and tumour necrosis factor a (TNFa) levels in condi-tioned medium from an overnight culture of primary cardiomyocyteswere measured by ELISA using the DuoSet ELISA kit (RampD Systems)according to the manufacturerrsquos instructions Blood from non-fastingHFD-fed mice was collected from submandibular bleeds and serum IL-6and TNFa levels were measured by ELISA (RampD Systems)
219 StatisticsOur data is expressed as means plusmn SD The statistical test for the GTTwas performed by repeated-measures analysis of variance (ANOVA)The comparison of two means was analysed by the Studentrsquos t-test Thecomparison between several means was analysed by one-way ANOVAfollowed by Tukeyrsquos post hoc analysis All statistical analyses were per-formed using GraphPad Prism 5 software (La Jolla CA) A P value lt_ 005was accepted as statistically significant
3 Results
31 aMHC promoter specificityFirst we verified the tissue specific expression of the Cre recombinaseunder the control of the aMHC promoter by crossing aMHC-Cremouse with the loxP-XFP transgenic (confetti) mouse30 In the confettimouse lox-P specific recombination resulted in the random expressionof one out of four fluorescent proteins (BFP GFP RFP or YFP) Celllysates were prepared from various tissues of either the confettiff mouse(without Cre -) or the double transgenic mouse (containing Cre 1) fol-lowed by western-blot analysis using anti-GFP antibodies As expectedGFP-related proteins were strongly detected in the heart lysate (seeSupplementary material online Figure S1) The lysate derived from lungtissue displayed a lower level of GFP-related proteins as compared tothe heart lysate probably due to low transcription activity of the aMHCpromoter in the lungs34 GFP-related protein expression was undetect-able in all of the other tissues tested (see Supplementary material onlineFigure S1)
32 In vitro analysis of cardiomyocytestreated with free fatty acidsTo reveal the possible role of ATF3 in cardiomyopathy induced by freefatty acids (FFA) (palmitate) we analysed adult cardiomyocyte primarycultures derived from ATF3ff and ATF3-cKO mice (ATF3-cKO geno-type ATF3ffaMHC-Cre) Cardiomyocytes were isolated by theLangendorff perfusion apparatus followed by overnight treatment with04 mM palmitate10 qRT-PCR analysis showed a 4- to 5-fold increase inATF3 induction by palmitate in the ATF3ff cardiomyocytes (Figure 1A)Immuno-fluorescence analysis of cardiomyocytes treated with either 1 h
or overnight palmitate displayed a robust ATF3 nuclear staining (Figure1B) Consistent with a previous study35 western-blot analysis of the car-diomyocytes lysate following overnight palmitate treatment demon-strated a significant induction of the ATF3 protein (Figure 1C) SinceATF3 induction is dependent on the activation of the p38-MAPK signal-ling pathway1026 we measured the p38-MAPK (p38) activation stateusing anti-phospho-p38 (p-p38) While p-p38 levels were increased fol-lowing palmitate treatment p38 remained unchanged independent ofthe treatment (Figure 1C) As expected neither ATF3 transcript (Figure1A) nor ATF3 protein (Figure 1C) was detected in the palmitate-treatedATF3-cKO cardiomyocytes Importantly palmitate treatment inducedthe expression of cardiac hypertrophic markers (ANP and BNP) inflam-mation markers (IL-6 and TNFa) a metabolic marker (PDK4) and anextracellular matrix (ECM) remodeling protein (TIMP1) (Figure 1A)Strikingly significantly higher level of induction of these genes wasobserved in the ATF3-cKO cardiomyocytes as compared with ATF3ff
cardiomyocytes (Figure 1A) In addition while the expression level of theinsulin receptor substrate IRS1 was induced 2-fold by palmitate treat-ment in ATF3ff IRS1 expression was suppressed in ATF3-cKO cardio-myocytes (Figure 1A)
Next we quantified the level of secreted inflammatory cytokines byanalysing the conditioned medium derived from palmitate-treated cardi-omyocytes The levels of IL-6 and TNFa were significantly higher in theconditioned medium derived from palmitate-treated ATF3-cKO cardio-myocytes as compared to their conditioned media levels derived frompalmitate-treated ATF3ff cardiomyocytes (Figure 1D)
Collectively the above-mentioned results support an increase of ATF3expression in cardiomyocytes in response to acute and chronic exposureto FFA The analysis of cardiomyocytes derived from wild type and ATF3-cKO genotypes in vitro is consistent with a possible protective role ofATF3 by dampening the cardiomyocytes remodeling response to FFA
33 The effect of high-fat diet on wild typeand ATF3-cKO miceTo determine whether cardiac ATF3 expression plays a role in responseto a high-fat diet (HFD)-regimen we used mice harbouring acardiomyocyte-selective deletion ATF3-cKO and their wild type (ATF3f
f) counterparts Mice were fed either a normal chow (NC 44 fat) or aHFD (60 fat) The HFD was initiated at 10 weeks of age and continuedfor 15 weeks Both ATF3ff and ATF3-cKO mice fed with a HFD gainedweight at significantly higher rates than NC-fed mice with no significantdifferences between the genotypes (see Supplementary material onlineFigure S2) In addition the levels of circulating total cholesterol and highdensity lipoprotein (HDL) were significantly higher in HFD-fed mice incomparison to NC-fed mice with no significant difference between thegenotypes (see Supplementary material online Figure S3A and B)
34 Cardiomyocyte-selective ATF3deficiency induces cardiac remodelingSince ATF3 is highly induced in cardiomyocytes that are treated withFFA (04 mM palmitate) we studied the expression of ATF3 in heart tis-sue following 15 weeks of a HFD regimen by qRT-PCR ATF3 expres-sion was induced 6-fold in the hearts of HFD-fed ATF3ff mice whencompared to either the NC-fed mice or to HFD-fed ATF3-cKO (Figure2A) We next performed immuno-histochemistry staining of heart slicesderived from NC- and HFD-fed mice with anti-ATF3 antibodies Whileno ATF3 staining was observed in the hearts derived from NC-fed mice
ATF3 expression in cardiomyocytes preserves homeostasis 137D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
or HFD-fed ATF3-cKO mice efficient ATF3 nuclear staining wasobserved in HFD-fed ATF3ff mice (Figure 2B)
In addition western-blot analysis using anti-ATF3 antibodies with car-diomyocyte cell lysate confirmed a high level of ATF3 protein expressionin HFD-fed as compared with NC-fed mice (Figure 2C and D) We nextexamined the cardiac expression levels of various gene specific markersas an indication for cardiac remodeling processes of a stressed heartHFD-fed ATF3-cKO but not ATF3ff mice exhibited significantly higherlevels of the foetal gene programming markers (ANP BNP and bMHC)when compared to the NC-fed wild type counterparts (Figure 2A)Concomitantly a-myosin heavy chain (aMHC) expression was signifi-cantly reduced in HFD-fed mice as compared to NC-fed mice independ-ent of their genotype (Figure 2A) The switch from aMHC to bMHC is anindication of the cardiac remodeling process as well as a cardiac disease
state Heart sections stained with wheat-germ hemagglutinin revealed asignificant increase in the cross sectional area in HFD-fed mice withlarger increased area in ATF3-cKO mice as compared with ATF3ff mice(Figure 2E and F) Taken together cardiac remodeling induced by a HFDis more pronounced in the ATF3-cKO mice than that in the controlATF3ff mice
We next examined fibrosis markers and found elevated expression ofcollagen 1a cTGF and a-SMA in HFD-fed mice as compared to the cor-responding NC-fed mice (Figure 3A) In all cases the ATF3-cKO miceshowed a more pronounced phenotype (Figure 3A) Western-blot analysiswith cell lysates derived from both NC-fed and HFD-fed mice confirmedincreased levels of a-SMA in HFD-fed mice but these high levels weremore noticeable in the lysate derived from HFD-fed ATF3-cKO mice(Figure 3B and C) Another fibrosis marker TGFb3 is significantly elevated
A C
B D
Figure 1 Free fatty acids affect the expression of hypertrophic inflammatory metabolic and fibrosis markers in primary cardiomyocytes Cardiomyocyteswere isolated from adult ATF3ff and ATF3-cKO mice and primary cultures were generated (A) Cardiomyocytes were exposed to 04 mM palmitate orvehicle control for overnight mRNA was extracted and the expression levels of the indicated genes were studied by qRT-PCR The mean ratio of palmitatetreated vs untreated cells for each genotype is shown The results represent the mean ratio plusmn SD (n = 5 independent preparations) (B) ATF3ff cardiomyo-cytes were exposed to 04 mM palmitate for either 1 h or overnight and then fixed and stained with anti-ATF3 (red) and anti-a-actinin (green) antibodiesNuclei were stained with DAPI (blue) (C) Western-blot analysis of cardiomyocytes lysate either palmitate treated overnight or left untreated probed withanti-ATF3 anti-p38 and anti-phospho-p38 antibodies The expression level of a-tubulin was used as loading control Each lane represents a lysate preparedfrom a pool derived from three independent cardiomyocytes culture (D) Isolated cardiomyocytes were treated as described in A Conditioned mediumwas collected and the levels of the indicated cytokines were assessed by ELISA The results represent the mean ratio between palmitate treated tountreatedplusmn SD (n = 5ndash6 independent preparations) P lt_ 005 comparing treated to untreated cells of the same genotype daggerP lt_ 005 comparing palmitate-treated cells between genotypes
138 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
in ATF3-cKO mice but not in the control ATF3ff mice (Figure 3A) againindicating that HFD has a more profound effect on ATF3-cKO mice
We next examined fibrosis histologically by Massonrsquos trichrome stain-ing This analysis confirmed a significant increased fibrosis in the hearts ofHFD-fed mice with ATF3-cKO mice exhibiting the highest fibrosis level(Figure 3D and E) Fibrosis is a result of the deposition of collagen in theECM The net effect of ECM degrading enzymes such as metalloprotei-nases (MMPs) and their inhibitors (TIMPs) determines the level of colla-gen deposition36 The levels of MMP9 were significantly decreased inHFD-fed mice as compared to NC-fed mice independent of the micegenotype (Figure 3A) In contrast TIMP1 levels were significantlyincreased in HFD-fed mice with considerably higher levels in the ATF3-cKO mice consistent with the finding that ATF3-cKO mice displayhigher level of fibrosis (Figure 3A)
Cardiac remodeling processes and diabetes are typically associatedwith increased inflammation We therefore measured several inflamma-tory markers in the hearts of NC- and HFD-fed ATF3ff and ATF3-cKOmice IL-6 expression was highly induced in the hearts of NC- and HFD-fed ATF3-cKO mice as compared with ATF3ff mice Significantly HFD-fed ATF3-cKO mice displayed the highest level (Figure 4A) SimilarlyTNFa was expressed at significantly higher levels in HFD-fed ATF3-cKOmice (Figure 4A) To identify the source of IL-6 and TNFa expression wehave isolated cardiomyocytes from mice fed with HFD for 15 weeks andexamined the cytokinersquos transcription and secretion levels by both qRT-PCR and ELISA respectively Indeed the transcription (Figure 4B) and
secretion (Figure 4C) levels of IL-6 and TNFa were significantly higher incardiomyocytes derived from HFD-fed ATF3-cKO mice as compared totheir ATF3ff cardiomyocytes counterparts To reveal whether theinflammatory response in the heart following a HFD affects circulatingIL-6 and TNFa levels we measured the cytokines in the serum of HFD-fed mice using ELISA Indeed a significant increase in IL-6 and TNFa lev-els were detected in the serum of ATF3-cKO mice (Figure 4D and Erespectively) Collectively we conclude that cardiomyocytes are thecells responsible at least in part for the expression and secretion of IL-6and TNFa in HFD-fed mice with significantly higher circulating levels inATF3 deficient mice
Lastly since inflammation is often associated with a maladaptive meta-bolic response we analysed the expression levels of various metabolicgenes in the heart The level of IRS1 a gene involved in insulin signallingwas significantly reduced in HFD-fed mice The reduction was more pro-nounced in the ATF3-cKO than in control mice (Figure 5A) Similarly thelevel of fatty acid translocase (CD36) was significantly higher in HFD-fedATF3ff mice IRS2 was much higher in the NC-fed ATF3-cKO mice thanin the control counterparts (NC-fed ATF3ff mice) consistent with ourprevious data that its gene expression is suppressed by ATF337
Interestingly IRS2 is induced in HFD-fed ATF3ff mice to comparable levelas HFD-fed ATF3-cKO mice The biological implication of this finding isnot clear One possibility is that induction of IRS2 a pro-survival gene bya HFD is an attempt by the cardiac cells to cope with the HFD stress thisadaptation is not necessary in the ATF3-cKO heart since IRS2 levels are
A
B
C
D
E
F
Figure 2 HFD-fed ATF3-cKO mice display increased expression of cardiac remodeling markers ATF3ff and ATF3-cKO mice were fed with either normalchow (NC) for 25 weeks or switched to a 15-week HFD regimen at 10 weeks of age At endpoint mice were sacrificed and hearts were excised (A) mRNAwas extracted from ventricles and the expression level of ATF3 and the indicated remodeling markers were measured by qRT-PCR Expression levels arepresented as relative values (compared to NC-fed ATF3ff mice determined as 1 for each gene) The results represent the meansplusmn SD (n = 6ndash8group) (B)Immuno-histochemistry analysis of paraformaldehyde fixed heart slices as described in A Representative nuclear staining is indicated by yellow arrowsScale bar = 50lm (C) Western-blot analysis of heart cell lysate probed with anti-ATF3 antibodies The expression level of a-tubulin was used as loadingcontrol (D) Quantification of ATF3 expression from C The results represent the meansplusmn SD (n = 3ndash5group) ratio of ATF3 expression of the indicatedgenotypes relative to NC ATF3ff determined as 1 (E) Ventricles sections were stained with TRITC-labelled wheat germ agglutinin and the cell size was ana-lysed Scale bar = 20lm (F) Quantification of cell size from E The results represent the meansplusmn SD (n = 6ndash8group) P lt_ 005 difference between dietsdaggerP lt_ 005 difference between genotypes
ATF3 expression in cardiomyocytes preserves homeostasis 139D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
already high The expression levels of other lipid and glycolytic regulatorssuch as peroxisome proliferator-activated receptor alpha (PPARa) carni-tine palmitoyltransferase 1 and 2 (CPT12) and pyruvate dehydrogenasekinase 4 (PDK4) were all elevated in HFD-fed mice with no significant dif-ference between the genotypes (Figure 5A) Lastly the level of the glucosetransporter 4 (Glut4) were unaffected by both diet and genotype at themRNA level (Figure 5A) However Glut4 was significantly elevated at theprotein level in HFD-fed mice with no significant difference between thegenotypes (see Supplementary material online Figure S4A and B)Collectively the expression of key metabolic genes was unalteredbetween the various genotypes in response to HFD In view of thechanges observed in metabolic genes in cardiomyocytes treated with pal-mitate (Figure 1A) this result was unexpected Thus we hypothesizedthat ATF3-dependent alterations in metabolic gene expression in cardio-myocytes may be masked by non-cardiomyocytes cells in the heartTherefore we studied the metabolic gene expression changes in isolatedcardiomyocytes derived from HFD- and NCndashfed mice from both ATF3ff
and ATF3-cKO mice (Figure 5B) Indeed the expression of all metabolicgenes tested was augmented in cardiomyocytes derived from HFD-fed ascompared with NC-fed mice Importantly the expression level of CD36CPT1 and CPT2 were significantly upregulated in HFD-fed ATF3-cKOmice as compared with their ATF3ff counterparts (Figure 5B) In contrastthe induction of both IRS1 and IRS2 were significantly lower in cardio-myocytes derived from HFD-fed ATF3-cKO mice (Figure 5B)Collectively the deleterious effects of HFD on cardiac remodeling fibro-sis and inflammation are more pronounced in ATF3-cKO mice suggest-ing that cardiac ATF3 plays a protective role in the HFD paradigm
35 HFD-induced cardiac dysfunctionNext we characterized the effect of HFD and cardiac remodeling oncardiac structure and function in ATF3ff and ATF3-cKO mice by
echocardiography Mice fed with NC had normal heart dimensions andfractional shortening (FS) (Figure 6A and B) HFD significantly reduced FSin both genotypes of HFD-fed mice as compared to corresponding NC-fed mice (Figure 6B) However the reduction was significantly more pro-nounced in the ATF3-cKO mice suggesting deteriorated systolic func-tion in these mice (Figure 6B) The reduced FS in the HFD-fed ATF3-cKO mice seems to be largely driven by an increase in LV internal end-systolic (LVIDs) dimension (Figure 6A) Heart rate was slightly increasedin the HFD-fed mice however this increase cannot explain the cardiacdysfunction observed in ATF3-cKO mice (Figure 6B) In addition weused cardiac MRI the gold standard for LV function and mass asses-sment38 LV mass was significantly higher in the ATF3-cKO mice follow-ing a 15-week HFD regimen (Figure 6C) In addition end-diastolic andend-systolic volumes were significantly larger in ATF3-cKO mice as com-pared to ATF3ff mice Moreover ATF3-cKO mice displayed a signifi-cantly lower ejection fraction as compared to ATF3ff mice (Figure 6D)These results are consistent with the reduced cardiac function found inthe echocardiographic analysis Images of mid-ventricular short-axis(Figure 6E) and long-axis (Figure 6E) slices are demonstrating the cardiaceccentric pattern observed in HFD-fed ATF3-cKO mice Collectivelythe results from the MRI show a greater increase in LV mass and volumein the ATF3-cKO mice as compared to the ATF3ff mice following aHFD suggesting fundamental structural differences in the heart betweenthe ATF3-cKO and ATF3ff mice
36 Impaired glucose tolerance in HFDATF3-cKO miceLastly we measured fasting glucose levels to assess the diabetic state ofthe mice As expected higher glucose levels were observed in HFD-fedmice as compared to NC-fed counterparts Interestingly the
A B
C
D
E
Figure 3 HFD-fed ATF3-cKO mice have higher level of fibrosis in the heart ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) mRNA wasextracted from ventricles and the expression levels of the indicated fibrosis markers were measured by qRT-PCR The expression levels are presented asrelative values (compared to NC-fed ATF3ff mice determined as 1 for each gene n = 6ndash8group) (B) Western-blot analysis of heart cell lysate probed withanti-a-SMA antibody The expression level of a-tubulin was used as loading control (C) Quantification of aSMA expression from B The results representthe meansplusmn SD (n= 3ndash5group) ratio of aSMA expression of the indicated genotypes relative to NC ATF3ff determined as 1 (D) Paraffin-embedded heartsections were stained with Massonrsquos trichrome to visualize fibrosis Scale bar = 100 lm (E) The level of fibrosis () stained by Massonrsquos trichrome was quan-tified and the results represent the meansplusmn SD (n = 6ndash8group) P lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes
140 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
hyperglycemia state of the ATF3-cKO mice was significantly higher thanin the ATF3ff mice following 15 weeks of HFD (Figure 7A) Consistentlythe glucose tolerance test (GTT) revealed that HFD-fed ATF3-cKOmice displayed higher blood glucose levels and longer glucose clearancetime when compared to HFD-fed ATF3ff whereas NC-fed ATF3ff andATF3-cKO mice had normal glucose clearance (Figure 7B)
Interestingly whole body ATF3-KO mice fed with HFD also displayedimpaired glucose clearance (see Supplementary material onlineFigure S5A)39 Previously we reported that decreased glucose tolerancein whole body ATF3-KO mice is at least in part due to reduced insulingene expression in b-cells (see Supplementary material online FigureS5B)39 To determine insulin level in HFD-fed ATF3-cKO mice weanalysed blood insulin levels by ELISA In both HFD-fed ATF3ff andATF3-cKO mice the serum insulin levels were significantly higher ascompared to their NC-fed counterpart mice with the ATF3-cKO micedisplaying the highest insulin levels (Figure 7C) Together these resultssuggest that peripheral tissues of ATF3-cKO mice have reduced ability
to uptake glucose despite their high insulin level in the blood Next wesought to measure the protein levels of components along the insulinpathway Akt and Foxo340 Both Akt and Foxo3 protein levels were sim-ilar in HFD-fed mice independent of the genotype (see Supplementarymaterial online Figure S4A and C) Interestingly phosphorylation of Akt(Ser473) which indicates the activity of the insulin pathway was slightlyelevated in HFD-fed mice however this did not reach statistical signifi-cance and was similar in both of the genotypes (see Supplementary material online Figure S4D) To measure the activity of the insulin signallingpathway following insulin stimulation we used the ratio between phos-phorylated Akt (Ser473) and total Akt expression HFD-fed mice wereinjected with insulin (10 Ukg) and the heart liver and skeletal musclewere harvested 15 min after the injection (Figure 7D and E and seeSupplementary material online Figure S6AndashD) Insulin injection resultedin a significant increase of pAktAkt ratio in the hearts of ATF3ff micewhile no significant change was observed in Akt phosphorylation inATF3-cKO mice (Figure 7D and E) Similarly low Akt phosphorylation
A
D E
B C
Figure 4 HFD-fed ATF3-cKO mice display inflammatory response in the heart (A) ATF3ff and ATF3-cKO mice were fed as described in Figure 2 mRNAwas extracted from ventricles and the expression levels of the indicated inflammatory markers were measured by qRT-PCR Expression levels are presentedas relative values (compared to NC-fed ATF3ff mice determined as 1 for each gene n= 6ndash8group) (B) Mice were fed as described in Figure 2Cardiomyocytes were isolated and primary cultures were generated mRNA was extracted from an overnight culture and the expression levels of IL-6 andTNFa were studied by qRT-PCR The mRNA level obtained in cardiomyocytes derived from NC-fed ATF3ff mice was determined as 1 and the relativeexpression of all other treatment is shown (n = 5ndash6 independent cardiomyocyte preparations) (C) Conditioned media was collected from cardiomyocytesdescribed in B and ELISA assay for the indicated cytokines is shown Cytokines level is presented as relative values to NC-fed ATF3ff mice determined as 1for each cytokine (n = 5ndash6 independent preparations) (D and E) Blood was collected from HFD-fed mice and IL-6 and TNFa serum levels were quantifiedby ELISA (n = 8ndash12group) The results represent the meansplusmn SD P lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes
ATF3 expression in cardiomyocytes preserves homeostasis 141D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
levels were also observed in both the liver and skeletal muscle of theATF3-cKO mice however this did not reach statistically significance (seeSupplementary material online Figure S6AndashD) Consistently palmitate-treated cardiomyocytes derived from ATF3-cKO mice displayedreduced phosphorylation of Akt following 30-min insulin stimulation(see Supplementary material online Figure S6E)
Collectively our findings revealed a surprising phenomenon that car-diac ATF3 deficiency results in increased cardiac remodeling leading topotentiation of local and peripheral insulin resistance
4 Discussion
A HFD is a commonly used regimen to induce T2D The majority of theliterature focuses on diabetic symptoms without addressing the issue ofT2D-induced cardiomyopathy Even when it is addressed there is noclear consensus regarding the extent of cardiac effects imposed byHFD41 Diet-induced obesity in mouse models is associated with globalinsulin resistance and increased myocardial fatty acid utilization how-ever the reported phenotypes on cardiac function vary from extensivehypertrophy and heart failure4 to lack of cardiac dysfunction4243 Thesevariations have been attributed to diet regimens andor sensitivity ofmouse strains to a HFD In this report we present evidence that cardiacATF3 deficiency exacerbates the deleterious effect of a HFD on theheart such as maladaptive cardiac remodeling fibrosis inflammation andcardiac dysfunction Our work may provide a potential molecular insightto variations in the literature regarding the impact of HFD on the heartAs ATF3 is a stress-inducible gene we propose that its proper inductionin the cardiomyocytes is protective and helps the heart to cope withHFD stress It is possible that the induction of ATF3mdashits expressionlevel duration and kineticsmdashvaries in different HFD regimens andormouse strains contributing to the variations observed in the literature
HFD regimen induced a significant higher level of fibrosis in the heartof ATF3-cKO mice We have identified TIMP1 as a possible mechanismto explain the increase in fibrosis in ATF3-cKO cardiomyocytes Severallines of evidence suggest that TIMP1 is possibly an ATF3 target gene
First we observed an increase in TIMP1 transcription in the whole heartand in isolated cardiomyocytes (Figures 1A and 3A) Second TIMP 1 3and 4 were identified as significantly highly expressed genes in the heartof ATF3-KO mice as compared with wild type mice following aorticbanding20 Third a functional AP-1 regulatory element within the TIMP1promoter was described44 Indeed preliminary results in our lab suggestthat ATF3 downregulates the TIMP1 promoter (data not shown) Thusit is possible that TIMP1 is suppressed by ATF3 and therefore results in anet increase in MMPs activity that helps to maintain ECM formationdeg-radation homeostasis
Our findings are consistent with previous studies demonstrating that adeficiency in ATF3 can promote cardiac maladaptive changes which leadto cardiac dilatation and hypertrophy in an aortic banding model1920 Incontrast a different study using a pressure overload model reportedthat ATF3 induces a maladaptive cardiac response17 These seeminglyconflicting data may be explained by the different patterns of ATF3expression induced by the different models In the pressure overloadmodel the acute and transient expression of ATF3 in cardiomyocytesinitiates a cascade of maladaptive changes17 whereas in the aortic band-ing model and the HFD model described here ATF3 expression is longlasting
In this context this is the first report using mice with cardiac-selectivedeficiency of ATF3 (ATF3-cKO) Previous reports on the roles of ATF3in cardiac function were carried out using whole body knockout ofATF325 or transgenic mice ectopically expressing ATF3 in cardiomyo-cytes16 A key advancement of ATF3-cKO mice over the whole bodyknockout mice is that they provide insights into the function of ATF3selectively in cardiomyocytes without the complication of ATF3 defi-ciency in other cells Thus the ATF3-cKO mouse is a useful tool to bet-ter understand the function of cardiac ATF3
Consistent with our previous report25 here we showed that cardiacATF3 deficiency has a more severe HFD-induced diabetic phenotypethan the control wild type mice However while reduced insulin genetranscription in ATF3-deficient b-cells was observed25 our current dataindicates that a lack of insulin is not the explanation for the glucose intol-erance found in the HFD-fed ATF3-cKO mice In contrast these mice
A B
Figure 5 HFD-fed ATF3-cKO mice show metabolic response in the heart ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) mRNA wasextracted from ventricles and the expression levels of the indicated metabolic markers were measured by qRT-PCR Expression levels are presented as rela-tive values (compared to NC-fed ATF3ff mice determined as 1 for each gene n= 6ndash8group) (B) Cardiomyocytes were isolated and plated on laminin for1 h followed by mRNA preparation (n = 4ndash5 independent preparations) The expression levels of the indicated metabolic markers were measured by qRT-PCR Expression levels are presented as relative values (to NC-fed ATF3ff mice determined as 1 for each gene) The results represent the meansplusmn SDP lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes
142 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
had higher blood insulin level (Figure 7C) Therefore upon HFD chal-lenge cardiac-deficiency of ATF3 exacerbates HFD-induced T2D Thislsquoperipheral effectrsquo is surprising since ATF3 is selectively deleted in thecardiomyocytes
The question then is how can we explain the effect of cardiacATF3 deficiency on peripheral glucose intolerance One possibility isthe reduced IRS1 expression in ATF3-deficient cardiomyocytes bothin vivo and in vitro IRS1 suppression may lead to impaired insulin sig-nalling in the heart thereby exacerbating glucose intolerance Indeedwe observed that HFD-fed ATF3-cKO mice displayed insulin resist-ance upon insulin challenge when compared to HFD-fed ATF3ff
mice In addition palmitate-treated cardiomyocytes lacking ATF3 dis-played lower insulin signalling as compared to their wild type coun-terparts Since the heart is only a minor glucose consuming organ incomparison to skeletal muscle cells or the brain the peripheral sys-temic effect cannot be solely based on this observation A case inpoint transgenic mice with insulin receptor (IR) deficiency in theheart do not display a diabetic phenotype4546 Another attractive
possibility for the peripheral effect of cardiac ATF3-deletion is itsimpact on inflammation It is well established that low-grade inflam-mation years in advance of the disease onset is involved in the patho-genic processes causing T2D8 Mediators of inflammation such asTNFa interleukin (IL)-1b the IL-6 family of cytokines IL-18 and cer-tain chemokines have been proposed to be involved in the eventscausing both forms of diabetes (T1D and T2D)8 Interestingly wehave shown that IL-6 a well-known ATF3 target gene47 is known toenhance its transcription and secretion of cytokines such as IL-6 andTNFa in palmitate-treated primary cultures of ATF3-cKO cardio-myocytes as well as in cardiomyocytes derived from HFD-fed ATF3-cKO mice IL-6 has been proposed to play a role in glucose homeo-stasis and metabolism directly and indirectly via its action on skeletalmuscle cells adipocytes hepatocytes pancreatic b-cells and neuro-endocrine cells48
Therefore we hypothesize that elevated levels of inflammatory cyto-kines in ATF3-cKO mice mediate a cross-talk in peripheral tissues leadingto impaired glucose tolerance Although circulating IL-6 and TNFa
A
B
C
D
E
F
Figure 6 HFD-fed ATF3-cKO mice present deteriorated heart function ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) At endpointmice hearts were examined by micro ultrasound The following parameters were measured interventricular septal end diastole (IVSd) left ventricular pos-terior wall end diastole (LVPWd) maximal left ventricular internal end-diastole (LVIDd) end-systole (LVIDs) and heart rate The results represent themeansplusmn SD (n = 6ndash8 in each group as shown in panel B) (B) Heart function was assessed by calculating percentage of fractional shortening (FS) accordingto FS () = [(LVDd-LVDs)LVDd]100 The results represent the meansplusmn SD (n = 6ndash8 in each group) (C and D) Left ventricular cardiac volumes massand function were examined by a cardiac MRI The following parameters were measured left ventricular end-diastolic (LVEDV) and left ventricular end-sys-tolic volume (LVESV) left ventricular (LV) mass and ejection fraction (EF) The results represent the meansplusmn SD (n = 7 in each group) (E and F)Representative images of mid-ventricular short-axis (E) and long-axis (F) slice at peak diastole and systole P lt_ 005 difference between diets daggerP lt_ 005 dif-ference between genotypes
ATF3 expression in cardiomyocytes preserves homeostasis 143D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
cytokines may be derived from different tissues we provide ample evi-dence that ATF3-defficient cardiomyocytes produce and secrete higherlevels of these cytokines as compared to their wild type counterpartsThis indicates a lsquocardiac-to-peripheralrsquo effect in T2D-induced cardiomyop-athy This is a reverse direction from the lsquoperipheral-to-cardiacrsquo effect dis-cussed in the introduction where inflammation in the peripheral tissuesleads to cardiac dysfunction Since inflammatory cytokines are soluble
factors they can exert a systemic effect This is why it makes sense thattheir effect can go both ways Thus diabetes and cardiomyopathy are apart of a vicious cycle Previously it was shown that cardiac specific dele-tion of the thyroid hormone complex mediator MED13 results in aperipheral effect upon HFD Cardiac MED13 deficiency suppresses meta-bolic genes in the heart which leads to a decrease in energy expenditureand an obese phenotype most likely by the secretion of cardiokines that
A
B
C
D
E
Figure 7 HFD-fed ATF3-cKO mice exhibit impaired glucose tolerance ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) Fasting (6 h)blood glucose concentrations (mgdl) The results represent the meansplusmn SD (n = 20ndash30group) P lt_ 005 difference between diets daggerP lt_ 005 differencebetween genotypes (B) Glucose tolerance test (GTT) Values are presented as relative to initial valueplusmn SD (n = 20ndash25group) P lt_ 005 difference betweendiets daggerP lt_ 005 difference between genotypes (C) Serum insulin concentrations were quantified by ELISA Values are presented as relative values (com-pared to NC-fed ATF3ff mice determined as 1 n = 15ndash20group) P lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes (D and E)Effect of HFD on phosphorylated Akt (Ser473 pAkt) to Akt ratio levels in heart lysate following either saline or insulin injection (10 U1 kg) is shownRepresentative blots (D) Densitometric analysis is presented as means ratio (n = 3ndash6group) pAktAkt ratio is presented as relative values (compared toHFD-fed saline injected ATF3ff mice determined as 1) P lt_ 005 insulin vs saline treatment daggerP lt_ 005 difference between genotypes (E)
144 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
mediates the cross talk between the heart and peripheral tissues49ndash51
Here we also observed increased levels of natriuretic peptides (ANP andBNP) which together with IL-6 and TNFa may mediate the cross-talk withthe peripheral tissues to induce glucose intolerance
Does this finding have any relevance to cardiac disease in humansSeveral lines of evidence suggests that congestive heart failure in patientsis associated with an increase in blood glucose levels52 In addition highblood glucose levels in non-diabetic patients with acute myocardialinfarction serves as a prognostic predictor of long-term mortality5354
Thus here we provide evidence that high blood glucose levels are a partof the symptoms of cardiac remodeling process
Collectively our findings suggest that cardiac ATF3 expression isresponsible for an adaptive cardiac response to the stressful environmentunder a HFD (Figure 8) ATF3 dampens the inflammatory response andsuppresses the expression of ECM remodeling genes and thus amelioratescardiac function In HFD-fed mice with ATF3 deficiency or in mice withcardiac remodeling the inflamed heart affects also peripheral tissuesthereby worsening T2D in peripheral tissues resulting in elevated bloodglucose level and turning on a vicious cycle to worsen cardiomyopathy
Supplementary material
Supplementary material is available at Cardiovascular Research online
AcknowledgementsThe authors wish to thank Drs Keren Bitton-Worms and Ran Rostokerand Prof Derek LeRoith from the Rambam Medical Center for their
assistance in GTT and ITT Dr Edith Suss Toby Maya Holdengreber andLior Liba from the BCF Bioimaging Center (Faculty of MedicineTechnion) for their assistance with imaging and image analysis Prof DEMichelle from University of Michigan for cardiomyocytes culture techni-que and Prof Yehiel Zick from the Weizmann institute of Sciences forcritical comments
Conflict of interest none declared
FundingThis work was partially supported by the United States-Israel BinationalGrant 2009179 to AA and TH In addition we wish to thank the MarsPittsburgh Fund for Medical Research to AA the Department of Defensegrant W81XWH-14-1-0179 to TH and to D Dan and Betty Kahn travelgrant support to LK
References
1 Roger VL Go AS Lloyd-Jones DM Benjamin EJ Berry JD Borden WB Bravata DMDai S Ford ES Fox CS Fullerton HJ Gillespie C Hailpern SM Heit JA Howard VJKissela BM Kittner SJ Lackland DT Lichtman JH Lisabeth LD Makuc DM MarcusGM Marelli A Matchar DB Moy CS Mozaffarian D Mussolino ME Nichol GPaynter NP Soliman EZ Sorlie PD Sotoodehnia N Turan TN Virani SS Wong NDWoo D Turner MB American Heart Association Statistics C Stroke Statistics SHeart disease and stroke statisticsndash2012 update a report from the American HeartAssociation Circulation 2012125e2ndashe220
2 Wilson C Diabetes ACCORD 5-year outcomes of intensive glycemic control NatRev Endocrinol 20117314
3 Ernande L Derumeaux G Diabetic cardiomyopathy myth or reality Arch CardiovascDis 2012105218ndash225
4 Battiprolu PK Hojayev B Jiang N Wang ZV Luo X Iglewski M Shelton JM GerardRD Rothermel BA Gillette TG Lavandero S Hill JA Metabolic stress-induced
InsulinGF
Cytokines
FFA
GlucoseATF3
Cardiac
Remodeling
Factors
Paracrine action
Collagens
and ECM
proteins
Fibrosis
Cardiac
Dysfunction
Systemic Effects
Glucose Tolerance
TNFα
IL-6
Cardiomyocyte
ATF3
Homeostasis
Homeostasis
TNFα
IL-6
TIMP1
BNP
ANP
Figure 8 lsquoPeripheral-to-cardiacrsquo and lsquoCardiac-to-peripheralrsquo model scheme Schematic diagram showing the ATF3 adaptive role in maintaining hearthomeostasis following HFD-induced T2D In contrast stressful environment under HFD leads to maladaptive cardiac response in the absence of ATF3Cardiac remodeling caused by ATF3 deficiency induces an increase in cardiac ECM remodeling and inflammatory genes and mediates the communication ofcardiomyocytes with heart resident cells potentially mediated by IL-6 TNFa ANP BNP and TIMP1 In this situation increased cardiac fibrosis and inflamma-tory response worsen cardiac function The inflamed failing heart mediates the communication of cardiomyocytes with distant organs as well thereby affect-ing diabetic outcome and glucose homeostasis
ATF3 expression in cardiomyocytes preserves homeostasis 145D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
activation of FoxO1 triggers diabetic cardiomyopathy in mice J Clin Invest20121221109ndash1118
5 Mandavia CH Aroor AR Demarco VG Sowers JR Molecular and metabolic mecha-nisms of cardiac dysfunction in diabetes Life Sci 201392601ndash608
6 Ventura-Clapier R Garnier A Veksler V Energy metabolism in heart failure J Physiol20045551ndash13
7 Guo S Insulin signaling resistance and the metabolic syndrome insights from mousemodels into disease mechanisms J Endocrinol 2014220T1ndashT23
8 Montane J Cadavez L Novials A Stress and the inflammatory process a major causeof pancreatic cell death in type 2 diabetes Diabetes Metab Syndr Obes 2014725ndash34
9 Ashrafian H Frenneaux MP Opie LH Metabolic mechanisms in heart failureCirculation 2007116434ndash448
10 Tanabe K Liu Y Hasan SD Martinez SC Cras-Meneur C Welling CM Bernal-MizrachiE Tanizawa Y Rhodes CJ Zmuda E Hai T Abumrad NA Permutt MA Glucose andfatty acids synergize to promote B-cell apoptosis through activation of glycogen syn-thase kinase 3beta independent of JNK activation PLoS One 20116e18146
11 Heineke J Molkentin JD Regulation of cardiac hypertrophy by intracellular signallingpathways Nat Rev Mol Cell Biol 20067589ndash600
12 Hai T Wolford CC Chang YS ATF3 a hub of the cellular adaptive-response net-work in the pathogenesis of diseases is modulation of inflammation a unifying com-ponent Gene Expr 2010151ndash11
13 Takii R Inouye S Fujimoto M Nakamura T Shinkawa T Prakasam R Tan K HayashidaN Ichikawa H Hai T Nakai A Heat shock transcription factor 1 inhibits expression ofIL-6 through activating transcription factor 3 J Immunol 20101841041ndash1048
14 Giraldo A Barrett OP Tindall MJ Fuller SJ Amirak E Bhattacharya BS Sugden PHClerk A Feedback regulation by Atf3 in the endothelin-1-responsive transcriptomeof cardiomyocytes Egr1 is a principal Atf3 target Biochem J 2012444343ndash355
15 Hasin T Elhanani O Abassi Z Hai T Aronheim A Angiotensin II signaling up-regulates the immediate early transcription factor ATF3 in the left but not the rightatrium Basic Res Cardiol 2011106175ndash187
16 Koren L Elhanani O Kehat I Hai T Aronheim A Adult cardiac expression of theactivating transcription factor 3 ATF3 promotes ventricular hypertrophy PLoS One20138e68396
17 Koren L Alishekevitz D Elhanani O Nevelsky A Hai T Kehat I Shaked Y AronheimA ATF3-dependent cross-talk between cardiomyocytes and macrophages promotescardiac maladaptive remodeling Int J Cardiol 2015198232ndash240
18 Okamoto Y Chaves A Chen J Kelley R Jones K Weed HG Gardner KL Gangi LYamaguchi M Klomkleaw W Nakayama T Hamlin RL Carnes C Altschuld R BauerJ Hai T Transgenic mice with cardiac-specific expression of activating transcriptionfactor 3 a stress-inducible gene have conduction abnormalities and contractile dys-function Am J Pathol 2001159639ndash650
19 Zhou H Shen DF Bian ZY Zong J Deng W Zhang Y Guo YY Li H Tang QZActivating transcription factor 3 deficiency promotes cardiac hypertrophy dysfunc-tion and fibrosis induced by pressure overload PLoS One 20116e26744
20 Zhou H Guo H Zong J Dai J Yuan Y Bian ZY Tang QZ ATF3 regulates multipletargets and may play a dual role in cardiac hypertrophy and injury Int J Cardiol2014174838ndash839
21 Yang J Yang CJ Yang J Fan ZX ATF3 a promotion effect or a inhibition effect in car-diac maladaptive remodeling Int J Cardiol 2015201245ndash246
22 Koren L Shaked Y Aronheim A Response letter ldquoATF3 a promoter or inhibitor ofcardiac maladaptive remodelingrdquo Int J Cardiol 2015201692
23 Ma ZG Wei WY Xu SC Zhang WB Dai J Tang QZ ATF3 a potential target forcardiac maladaptive remodeling Int J Cardiol 201620250ndash51
24 Zhou H Yuan Y Ni J Guo H Deng W Bian ZY Tang QZ Pleiotropic and puzzlingeffects of ATF3 in maladaptive cardiac remodeling Int J Cardiol 201620687ndash88
25 Zmuda EJ Qi L Zhu MX Mirmira RG Montminy MR Hai T The roles of ATF3 anadaptive-response gene in high-fat-diet-induced diabetes and pancreatic beta-celldysfunction Mol Endocrinol 2010241423ndash1433
26 Lu D Chen J Hai T The regulation of ATF3 gene expression by mitogen-activatedprotein kinases Biochem J 2007401559ndash567
27 Hartman MG Lu D Kim ML Kociba GJ Shukri T Buteau J Wang X Frankel WLGuttridge D Prentki M Grey ST Ron D Hai T Role for activating transcription fac-tor 3 in stress-induced beta-cell apoptosis Mol Cell Biol 2004245721ndash5732
28 Wolford CC McConoughey SJ Jalgaonkar SP Leon M Merchant AS Dominick JLYin X Chang Y Zmuda EJ OrsquoToole SA Millar EK Roller SL Shapiro CL OstrowskiMC Sutherland RL Hai T Transcription factor ATF3 links host adaptive response tobreast cancer metastasis J Clin Iinvest 20131232893ndash2906
29 Agah R Frenkel PA French BA Michael LH Overbeek PA Schneider MD Generecombination in postmitotic cells Targeted expression of Cre recombinase pro-vokes cardiac-restricted site-specific rearrangement in adult ventricular musclein vivo J Clin Invest 1997100169ndash179
30 Livet J Weissman TA Kang H Draft RW Lu J Bennis RA Sanes JR Lichtman JWTransgenic strategies for combinatorial expression of fluorescent proteins in thenervous system Nature 200745056ndash62
31 Davis J Maillet M Miano JM Molkentin JD Lost in transgenesis a userrsquos guidefor genetically manipulating the mouse in cardiac research Circ Res2012111761ndash777
32 Heiberg E Sjogren J Ugander M Carlsson M Engblom H Arheden H Design andvalidation of segment ndash freely available software for cardiovascular image analysisBMC Med Imaging 2010101
33 Kabaeva Z Zhao M Michele DE Blebbistatin extends culture life of adult mouse car-diac myocytes and allows efficient and stable transgene expression Am J Physiol HeartC 2008294H1667ndash1674
34 Subramaniam A Jones WK Gulick J Wert S Neumann J Robbins J Tissue-specificregulation of the alpha-myosin heavy chain gene promoter in transgenic mice J BiolChem 199126624613ndash24620
35 Cunha DA Hekerman P Ladriere L Bazarra-Castro A Ortis F Wakeham MCMoore F Rasschaert J Cardozo AK Bellomo E Overbergh L Mathieu C Lupi R HaiT Herchuelz A Marchetti P Rutter GA Eizirik DL Cnop M Initiation and executionof lipotoxic ER stress in pancreatic beta-cells J Cell Sci 20081212308ndash2318
36 Wynn TA Ramalingam TR Mechanisms of fibrosis therapeutic translation forfibrotic disease Nat Med 2012181028ndash1040
37 Li D Yin X Zmuda EJ Wolford CC Dong X White MF Hai T The repression ofIRS2 gene by ATF3 a stress-inducible gene contributes to pancreatic beta-cell apop-tosis Diabetes 200857635ndash644
38 Myerson SG Bellenger NG Pennell DJ Assessment of left ventricular mass by cardi-ovascular magnetic resonance Hypertension 200239750ndash755
39 Zmuda EJ Viapiano M Grey ST Hadley G Garcia-Ocana A Hai T Deficiency ofAtf3 an adaptive-response gene protects islets and ameliorates inflammation in asyngeneic mouse transplantation model Diabetologia 2010531438ndash1450
40 Riehle C Abel ED Insulin signaling and heart failure Circ Res 20161181151ndash116941 Schreyer SA Wilson DL LeBoeuf RC C57BL6 mice fed high fat diets as models for
diabetes-accelerated atherosclerosis Atherosclerosis 199813617ndash2442 Brainard RE Watson LJ Demartino AM Brittian KR Readnower RD Boakye AA
Zhang D Hoetker JD Bhatnagar A Baba SP Jones SP High fat feeding in mice isinsufficient to induce cardiac dysfunction and does not exacerbate heart failure PLoSOne 20138e83174
43 Gupte AA Minze LJ Reyes M Ren Y Wang X Brunner G Ghosn M Cordero-Reyes AM Ding K Pratico D Morrisett J Shi ZZ Hamilton DJ Lyon CJ Hsueh WAHigh-fat feeding-induced hyperinsulinemia increases cardiac glucose uptake and mito-chondrial function despite peripheral insulin resistance Endocrinology20131542650ndash2662
44 Logan SK Garabedian MJ Campbell CE Werb Z Synergistic transcriptionalactivation of the tissue inhibitor of metalloproteinases-1 promoter via functionalinteraction of AP-1 and Ets-1 transcription factors J Biol Chem1996271774ndash782
45 Belke DD Betuing S Tuttle MJ Graveleau C Young ME Pham M Zhang DCooksey RC McClain DA Litwin SE Taegtmeyer H Severson D Kahn CR AbelED Insulin signaling coordinately regulates cardiac size metabolism and contractileprotein isoform expression J Clin Invest 2002109629ndash639
46 Fernandez AM Kim JK Yakar S Dupont J Hernandez-Sanchez C Castle AL FilmoreJ Shulman GI Le Roith D Functional inactivation of the IGF-I and insulin receptors inskeletal muscle causes type 2 diabetes Genes Dev 2001151926ndash1934
47 Gilchrist M Thorsson V Li B Rust AG Korb M Roach JC Kennedy K Hai TBolouri H Aderem A Systems biology approaches identify ATF3 as a negative regu-lator of Toll-like receptor 4 Nature 2006441173ndash178
48 Kristiansen OP Mandrup-Poulsen T Interleukin-6 and diabetes the good the bad orthe indifferent Diabetes 200554(Suppl 2)S114ndashS124
49 Grueter CE van Rooij E Johnson BA DeLeon SM Sutherland LB Qi X Gautron LElmquist JK Bassel-Duby R Olson EN A cardiac microRNA governs systemic energyhomeostasis by regulation of MED13 Cell 2012149671ndash683
50 Baskin KK Grueter CE Kusminski CM Holland WL Bookout AL Satapati S KongYM Burgess SC Malloy CR Scherer PE Newgard CB Bassel-Duby R Olson ENMED13-dependent signaling from the heart confers leanness by enhancing metabo-lism in adipose tissue and liver EMBO Mol Med 201461610ndash1621
51 Nakamura M Sadoshima J Heart over mind metabolic control of white adipose tis-sue and liver EMBO Mol Med 201461521ndash1524
52 Tenenbaum A Fisman EZ Impaired glucose metabolism in patients with heart failurepathophysiology and possible treatment strategies Am J Cardiovasc Drugs20044269ndash280
53 Suskin N McKelvie RS Burns RJ Latini R Pericak D Probstfield J Rouleau JL SigouinC Solymoss CB Tsuyuki R White M Yusuf S Glucose and insulin abnormalitiesrelate to functional capacity in patients with congestive heart failure Eur Heart J2000211368ndash1375
54 Aronson D Hammerman H Kapeliovich MR Suleiman A Agmon Y Beyar RMarkiewicz W Suleiman M Fasting glucose in acute myocardial infarction incremen-tal value for long-term mortality and relationship with left ventricular systolic func-tion Diabetes Care 200730960ndash966
146 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
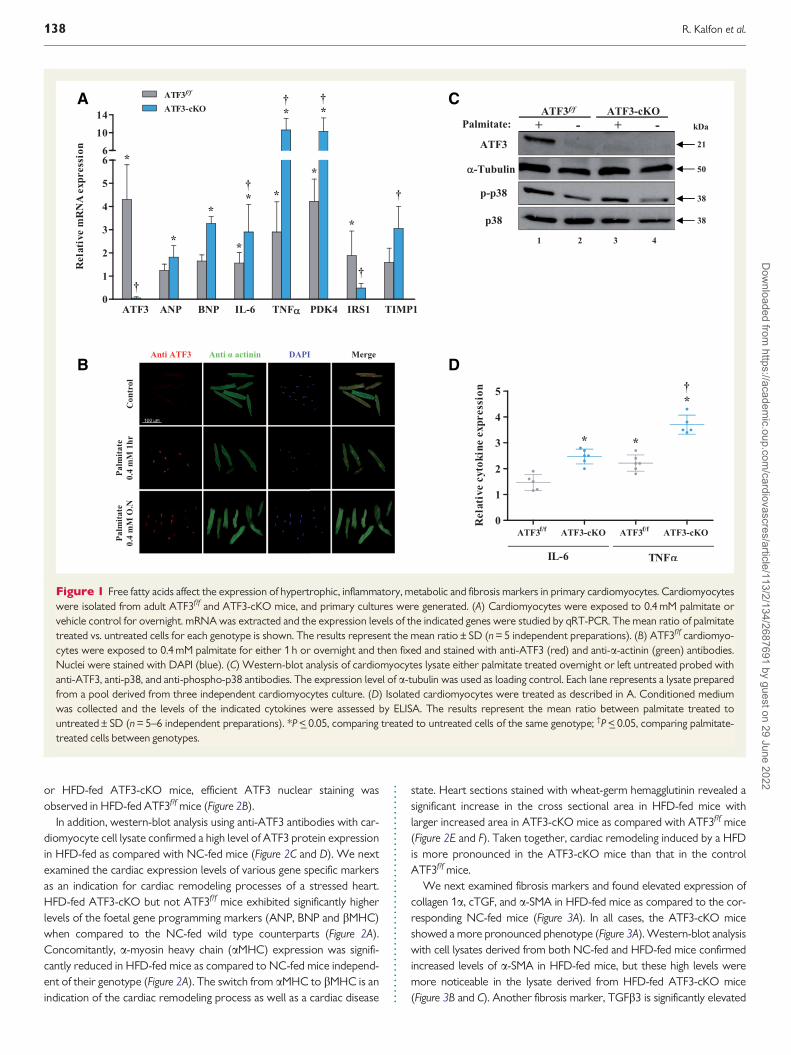
or HFD-fed ATF3-cKO mice efficient ATF3 nuclear staining wasobserved in HFD-fed ATF3ff mice (Figure 2B)
In addition western-blot analysis using anti-ATF3 antibodies with car-diomyocyte cell lysate confirmed a high level of ATF3 protein expressionin HFD-fed as compared with NC-fed mice (Figure 2C and D) We nextexamined the cardiac expression levels of various gene specific markersas an indication for cardiac remodeling processes of a stressed heartHFD-fed ATF3-cKO but not ATF3ff mice exhibited significantly higherlevels of the foetal gene programming markers (ANP BNP and bMHC)when compared to the NC-fed wild type counterparts (Figure 2A)Concomitantly a-myosin heavy chain (aMHC) expression was signifi-cantly reduced in HFD-fed mice as compared to NC-fed mice independ-ent of their genotype (Figure 2A) The switch from aMHC to bMHC is anindication of the cardiac remodeling process as well as a cardiac disease
state Heart sections stained with wheat-germ hemagglutinin revealed asignificant increase in the cross sectional area in HFD-fed mice withlarger increased area in ATF3-cKO mice as compared with ATF3ff mice(Figure 2E and F) Taken together cardiac remodeling induced by a HFDis more pronounced in the ATF3-cKO mice than that in the controlATF3ff mice
We next examined fibrosis markers and found elevated expression ofcollagen 1a cTGF and a-SMA in HFD-fed mice as compared to the cor-responding NC-fed mice (Figure 3A) In all cases the ATF3-cKO miceshowed a more pronounced phenotype (Figure 3A) Western-blot analysiswith cell lysates derived from both NC-fed and HFD-fed mice confirmedincreased levels of a-SMA in HFD-fed mice but these high levels weremore noticeable in the lysate derived from HFD-fed ATF3-cKO mice(Figure 3B and C) Another fibrosis marker TGFb3 is significantly elevated
A C
B D
Figure 1 Free fatty acids affect the expression of hypertrophic inflammatory metabolic and fibrosis markers in primary cardiomyocytes Cardiomyocyteswere isolated from adult ATF3ff and ATF3-cKO mice and primary cultures were generated (A) Cardiomyocytes were exposed to 04 mM palmitate orvehicle control for overnight mRNA was extracted and the expression levels of the indicated genes were studied by qRT-PCR The mean ratio of palmitatetreated vs untreated cells for each genotype is shown The results represent the mean ratio plusmn SD (n = 5 independent preparations) (B) ATF3ff cardiomyo-cytes were exposed to 04 mM palmitate for either 1 h or overnight and then fixed and stained with anti-ATF3 (red) and anti-a-actinin (green) antibodiesNuclei were stained with DAPI (blue) (C) Western-blot analysis of cardiomyocytes lysate either palmitate treated overnight or left untreated probed withanti-ATF3 anti-p38 and anti-phospho-p38 antibodies The expression level of a-tubulin was used as loading control Each lane represents a lysate preparedfrom a pool derived from three independent cardiomyocytes culture (D) Isolated cardiomyocytes were treated as described in A Conditioned mediumwas collected and the levels of the indicated cytokines were assessed by ELISA The results represent the mean ratio between palmitate treated tountreatedplusmn SD (n = 5ndash6 independent preparations) P lt_ 005 comparing treated to untreated cells of the same genotype daggerP lt_ 005 comparing palmitate-treated cells between genotypes
138 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
in ATF3-cKO mice but not in the control ATF3ff mice (Figure 3A) againindicating that HFD has a more profound effect on ATF3-cKO mice
We next examined fibrosis histologically by Massonrsquos trichrome stain-ing This analysis confirmed a significant increased fibrosis in the hearts ofHFD-fed mice with ATF3-cKO mice exhibiting the highest fibrosis level(Figure 3D and E) Fibrosis is a result of the deposition of collagen in theECM The net effect of ECM degrading enzymes such as metalloprotei-nases (MMPs) and their inhibitors (TIMPs) determines the level of colla-gen deposition36 The levels of MMP9 were significantly decreased inHFD-fed mice as compared to NC-fed mice independent of the micegenotype (Figure 3A) In contrast TIMP1 levels were significantlyincreased in HFD-fed mice with considerably higher levels in the ATF3-cKO mice consistent with the finding that ATF3-cKO mice displayhigher level of fibrosis (Figure 3A)
Cardiac remodeling processes and diabetes are typically associatedwith increased inflammation We therefore measured several inflamma-tory markers in the hearts of NC- and HFD-fed ATF3ff and ATF3-cKOmice IL-6 expression was highly induced in the hearts of NC- and HFD-fed ATF3-cKO mice as compared with ATF3ff mice Significantly HFD-fed ATF3-cKO mice displayed the highest level (Figure 4A) SimilarlyTNFa was expressed at significantly higher levels in HFD-fed ATF3-cKOmice (Figure 4A) To identify the source of IL-6 and TNFa expression wehave isolated cardiomyocytes from mice fed with HFD for 15 weeks andexamined the cytokinersquos transcription and secretion levels by both qRT-PCR and ELISA respectively Indeed the transcription (Figure 4B) and
secretion (Figure 4C) levels of IL-6 and TNFa were significantly higher incardiomyocytes derived from HFD-fed ATF3-cKO mice as compared totheir ATF3ff cardiomyocytes counterparts To reveal whether theinflammatory response in the heart following a HFD affects circulatingIL-6 and TNFa levels we measured the cytokines in the serum of HFD-fed mice using ELISA Indeed a significant increase in IL-6 and TNFa lev-els were detected in the serum of ATF3-cKO mice (Figure 4D and Erespectively) Collectively we conclude that cardiomyocytes are thecells responsible at least in part for the expression and secretion of IL-6and TNFa in HFD-fed mice with significantly higher circulating levels inATF3 deficient mice
Lastly since inflammation is often associated with a maladaptive meta-bolic response we analysed the expression levels of various metabolicgenes in the heart The level of IRS1 a gene involved in insulin signallingwas significantly reduced in HFD-fed mice The reduction was more pro-nounced in the ATF3-cKO than in control mice (Figure 5A) Similarly thelevel of fatty acid translocase (CD36) was significantly higher in HFD-fedATF3ff mice IRS2 was much higher in the NC-fed ATF3-cKO mice thanin the control counterparts (NC-fed ATF3ff mice) consistent with ourprevious data that its gene expression is suppressed by ATF337
Interestingly IRS2 is induced in HFD-fed ATF3ff mice to comparable levelas HFD-fed ATF3-cKO mice The biological implication of this finding isnot clear One possibility is that induction of IRS2 a pro-survival gene bya HFD is an attempt by the cardiac cells to cope with the HFD stress thisadaptation is not necessary in the ATF3-cKO heart since IRS2 levels are
A
B
C
D
E
F
Figure 2 HFD-fed ATF3-cKO mice display increased expression of cardiac remodeling markers ATF3ff and ATF3-cKO mice were fed with either normalchow (NC) for 25 weeks or switched to a 15-week HFD regimen at 10 weeks of age At endpoint mice were sacrificed and hearts were excised (A) mRNAwas extracted from ventricles and the expression level of ATF3 and the indicated remodeling markers were measured by qRT-PCR Expression levels arepresented as relative values (compared to NC-fed ATF3ff mice determined as 1 for each gene) The results represent the meansplusmn SD (n = 6ndash8group) (B)Immuno-histochemistry analysis of paraformaldehyde fixed heart slices as described in A Representative nuclear staining is indicated by yellow arrowsScale bar = 50lm (C) Western-blot analysis of heart cell lysate probed with anti-ATF3 antibodies The expression level of a-tubulin was used as loadingcontrol (D) Quantification of ATF3 expression from C The results represent the meansplusmn SD (n = 3ndash5group) ratio of ATF3 expression of the indicatedgenotypes relative to NC ATF3ff determined as 1 (E) Ventricles sections were stained with TRITC-labelled wheat germ agglutinin and the cell size was ana-lysed Scale bar = 20lm (F) Quantification of cell size from E The results represent the meansplusmn SD (n = 6ndash8group) P lt_ 005 difference between dietsdaggerP lt_ 005 difference between genotypes
ATF3 expression in cardiomyocytes preserves homeostasis 139D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
already high The expression levels of other lipid and glycolytic regulatorssuch as peroxisome proliferator-activated receptor alpha (PPARa) carni-tine palmitoyltransferase 1 and 2 (CPT12) and pyruvate dehydrogenasekinase 4 (PDK4) were all elevated in HFD-fed mice with no significant dif-ference between the genotypes (Figure 5A) Lastly the level of the glucosetransporter 4 (Glut4) were unaffected by both diet and genotype at themRNA level (Figure 5A) However Glut4 was significantly elevated at theprotein level in HFD-fed mice with no significant difference between thegenotypes (see Supplementary material online Figure S4A and B)Collectively the expression of key metabolic genes was unalteredbetween the various genotypes in response to HFD In view of thechanges observed in metabolic genes in cardiomyocytes treated with pal-mitate (Figure 1A) this result was unexpected Thus we hypothesizedthat ATF3-dependent alterations in metabolic gene expression in cardio-myocytes may be masked by non-cardiomyocytes cells in the heartTherefore we studied the metabolic gene expression changes in isolatedcardiomyocytes derived from HFD- and NCndashfed mice from both ATF3ff
and ATF3-cKO mice (Figure 5B) Indeed the expression of all metabolicgenes tested was augmented in cardiomyocytes derived from HFD-fed ascompared with NC-fed mice Importantly the expression level of CD36CPT1 and CPT2 were significantly upregulated in HFD-fed ATF3-cKOmice as compared with their ATF3ff counterparts (Figure 5B) In contrastthe induction of both IRS1 and IRS2 were significantly lower in cardio-myocytes derived from HFD-fed ATF3-cKO mice (Figure 5B)Collectively the deleterious effects of HFD on cardiac remodeling fibro-sis and inflammation are more pronounced in ATF3-cKO mice suggest-ing that cardiac ATF3 plays a protective role in the HFD paradigm
35 HFD-induced cardiac dysfunctionNext we characterized the effect of HFD and cardiac remodeling oncardiac structure and function in ATF3ff and ATF3-cKO mice by
echocardiography Mice fed with NC had normal heart dimensions andfractional shortening (FS) (Figure 6A and B) HFD significantly reduced FSin both genotypes of HFD-fed mice as compared to corresponding NC-fed mice (Figure 6B) However the reduction was significantly more pro-nounced in the ATF3-cKO mice suggesting deteriorated systolic func-tion in these mice (Figure 6B) The reduced FS in the HFD-fed ATF3-cKO mice seems to be largely driven by an increase in LV internal end-systolic (LVIDs) dimension (Figure 6A) Heart rate was slightly increasedin the HFD-fed mice however this increase cannot explain the cardiacdysfunction observed in ATF3-cKO mice (Figure 6B) In addition weused cardiac MRI the gold standard for LV function and mass asses-sment38 LV mass was significantly higher in the ATF3-cKO mice follow-ing a 15-week HFD regimen (Figure 6C) In addition end-diastolic andend-systolic volumes were significantly larger in ATF3-cKO mice as com-pared to ATF3ff mice Moreover ATF3-cKO mice displayed a signifi-cantly lower ejection fraction as compared to ATF3ff mice (Figure 6D)These results are consistent with the reduced cardiac function found inthe echocardiographic analysis Images of mid-ventricular short-axis(Figure 6E) and long-axis (Figure 6E) slices are demonstrating the cardiaceccentric pattern observed in HFD-fed ATF3-cKO mice Collectivelythe results from the MRI show a greater increase in LV mass and volumein the ATF3-cKO mice as compared to the ATF3ff mice following aHFD suggesting fundamental structural differences in the heart betweenthe ATF3-cKO and ATF3ff mice
36 Impaired glucose tolerance in HFDATF3-cKO miceLastly we measured fasting glucose levels to assess the diabetic state ofthe mice As expected higher glucose levels were observed in HFD-fedmice as compared to NC-fed counterparts Interestingly the
A B
C
D
E
Figure 3 HFD-fed ATF3-cKO mice have higher level of fibrosis in the heart ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) mRNA wasextracted from ventricles and the expression levels of the indicated fibrosis markers were measured by qRT-PCR The expression levels are presented asrelative values (compared to NC-fed ATF3ff mice determined as 1 for each gene n = 6ndash8group) (B) Western-blot analysis of heart cell lysate probed withanti-a-SMA antibody The expression level of a-tubulin was used as loading control (C) Quantification of aSMA expression from B The results representthe meansplusmn SD (n= 3ndash5group) ratio of aSMA expression of the indicated genotypes relative to NC ATF3ff determined as 1 (D) Paraffin-embedded heartsections were stained with Massonrsquos trichrome to visualize fibrosis Scale bar = 100 lm (E) The level of fibrosis () stained by Massonrsquos trichrome was quan-tified and the results represent the meansplusmn SD (n = 6ndash8group) P lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes
140 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
hyperglycemia state of the ATF3-cKO mice was significantly higher thanin the ATF3ff mice following 15 weeks of HFD (Figure 7A) Consistentlythe glucose tolerance test (GTT) revealed that HFD-fed ATF3-cKOmice displayed higher blood glucose levels and longer glucose clearancetime when compared to HFD-fed ATF3ff whereas NC-fed ATF3ff andATF3-cKO mice had normal glucose clearance (Figure 7B)
Interestingly whole body ATF3-KO mice fed with HFD also displayedimpaired glucose clearance (see Supplementary material onlineFigure S5A)39 Previously we reported that decreased glucose tolerancein whole body ATF3-KO mice is at least in part due to reduced insulingene expression in b-cells (see Supplementary material online FigureS5B)39 To determine insulin level in HFD-fed ATF3-cKO mice weanalysed blood insulin levels by ELISA In both HFD-fed ATF3ff andATF3-cKO mice the serum insulin levels were significantly higher ascompared to their NC-fed counterpart mice with the ATF3-cKO micedisplaying the highest insulin levels (Figure 7C) Together these resultssuggest that peripheral tissues of ATF3-cKO mice have reduced ability
to uptake glucose despite their high insulin level in the blood Next wesought to measure the protein levels of components along the insulinpathway Akt and Foxo340 Both Akt and Foxo3 protein levels were sim-ilar in HFD-fed mice independent of the genotype (see Supplementarymaterial online Figure S4A and C) Interestingly phosphorylation of Akt(Ser473) which indicates the activity of the insulin pathway was slightlyelevated in HFD-fed mice however this did not reach statistical signifi-cance and was similar in both of the genotypes (see Supplementary material online Figure S4D) To measure the activity of the insulin signallingpathway following insulin stimulation we used the ratio between phos-phorylated Akt (Ser473) and total Akt expression HFD-fed mice wereinjected with insulin (10 Ukg) and the heart liver and skeletal musclewere harvested 15 min after the injection (Figure 7D and E and seeSupplementary material online Figure S6AndashD) Insulin injection resultedin a significant increase of pAktAkt ratio in the hearts of ATF3ff micewhile no significant change was observed in Akt phosphorylation inATF3-cKO mice (Figure 7D and E) Similarly low Akt phosphorylation
A
D E
B C
Figure 4 HFD-fed ATF3-cKO mice display inflammatory response in the heart (A) ATF3ff and ATF3-cKO mice were fed as described in Figure 2 mRNAwas extracted from ventricles and the expression levels of the indicated inflammatory markers were measured by qRT-PCR Expression levels are presentedas relative values (compared to NC-fed ATF3ff mice determined as 1 for each gene n= 6ndash8group) (B) Mice were fed as described in Figure 2Cardiomyocytes were isolated and primary cultures were generated mRNA was extracted from an overnight culture and the expression levels of IL-6 andTNFa were studied by qRT-PCR The mRNA level obtained in cardiomyocytes derived from NC-fed ATF3ff mice was determined as 1 and the relativeexpression of all other treatment is shown (n = 5ndash6 independent cardiomyocyte preparations) (C) Conditioned media was collected from cardiomyocytesdescribed in B and ELISA assay for the indicated cytokines is shown Cytokines level is presented as relative values to NC-fed ATF3ff mice determined as 1for each cytokine (n = 5ndash6 independent preparations) (D and E) Blood was collected from HFD-fed mice and IL-6 and TNFa serum levels were quantifiedby ELISA (n = 8ndash12group) The results represent the meansplusmn SD P lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes
ATF3 expression in cardiomyocytes preserves homeostasis 141D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
levels were also observed in both the liver and skeletal muscle of theATF3-cKO mice however this did not reach statistically significance (seeSupplementary material online Figure S6AndashD) Consistently palmitate-treated cardiomyocytes derived from ATF3-cKO mice displayedreduced phosphorylation of Akt following 30-min insulin stimulation(see Supplementary material online Figure S6E)
Collectively our findings revealed a surprising phenomenon that car-diac ATF3 deficiency results in increased cardiac remodeling leading topotentiation of local and peripheral insulin resistance
4 Discussion
A HFD is a commonly used regimen to induce T2D The majority of theliterature focuses on diabetic symptoms without addressing the issue ofT2D-induced cardiomyopathy Even when it is addressed there is noclear consensus regarding the extent of cardiac effects imposed byHFD41 Diet-induced obesity in mouse models is associated with globalinsulin resistance and increased myocardial fatty acid utilization how-ever the reported phenotypes on cardiac function vary from extensivehypertrophy and heart failure4 to lack of cardiac dysfunction4243 Thesevariations have been attributed to diet regimens andor sensitivity ofmouse strains to a HFD In this report we present evidence that cardiacATF3 deficiency exacerbates the deleterious effect of a HFD on theheart such as maladaptive cardiac remodeling fibrosis inflammation andcardiac dysfunction Our work may provide a potential molecular insightto variations in the literature regarding the impact of HFD on the heartAs ATF3 is a stress-inducible gene we propose that its proper inductionin the cardiomyocytes is protective and helps the heart to cope withHFD stress It is possible that the induction of ATF3mdashits expressionlevel duration and kineticsmdashvaries in different HFD regimens andormouse strains contributing to the variations observed in the literature
HFD regimen induced a significant higher level of fibrosis in the heartof ATF3-cKO mice We have identified TIMP1 as a possible mechanismto explain the increase in fibrosis in ATF3-cKO cardiomyocytes Severallines of evidence suggest that TIMP1 is possibly an ATF3 target gene
First we observed an increase in TIMP1 transcription in the whole heartand in isolated cardiomyocytes (Figures 1A and 3A) Second TIMP 1 3and 4 were identified as significantly highly expressed genes in the heartof ATF3-KO mice as compared with wild type mice following aorticbanding20 Third a functional AP-1 regulatory element within the TIMP1promoter was described44 Indeed preliminary results in our lab suggestthat ATF3 downregulates the TIMP1 promoter (data not shown) Thusit is possible that TIMP1 is suppressed by ATF3 and therefore results in anet increase in MMPs activity that helps to maintain ECM formationdeg-radation homeostasis
Our findings are consistent with previous studies demonstrating that adeficiency in ATF3 can promote cardiac maladaptive changes which leadto cardiac dilatation and hypertrophy in an aortic banding model1920 Incontrast a different study using a pressure overload model reportedthat ATF3 induces a maladaptive cardiac response17 These seeminglyconflicting data may be explained by the different patterns of ATF3expression induced by the different models In the pressure overloadmodel the acute and transient expression of ATF3 in cardiomyocytesinitiates a cascade of maladaptive changes17 whereas in the aortic band-ing model and the HFD model described here ATF3 expression is longlasting
In this context this is the first report using mice with cardiac-selectivedeficiency of ATF3 (ATF3-cKO) Previous reports on the roles of ATF3in cardiac function were carried out using whole body knockout ofATF325 or transgenic mice ectopically expressing ATF3 in cardiomyo-cytes16 A key advancement of ATF3-cKO mice over the whole bodyknockout mice is that they provide insights into the function of ATF3selectively in cardiomyocytes without the complication of ATF3 defi-ciency in other cells Thus the ATF3-cKO mouse is a useful tool to bet-ter understand the function of cardiac ATF3
Consistent with our previous report25 here we showed that cardiacATF3 deficiency has a more severe HFD-induced diabetic phenotypethan the control wild type mice However while reduced insulin genetranscription in ATF3-deficient b-cells was observed25 our current dataindicates that a lack of insulin is not the explanation for the glucose intol-erance found in the HFD-fed ATF3-cKO mice In contrast these mice
A B
Figure 5 HFD-fed ATF3-cKO mice show metabolic response in the heart ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) mRNA wasextracted from ventricles and the expression levels of the indicated metabolic markers were measured by qRT-PCR Expression levels are presented as rela-tive values (compared to NC-fed ATF3ff mice determined as 1 for each gene n= 6ndash8group) (B) Cardiomyocytes were isolated and plated on laminin for1 h followed by mRNA preparation (n = 4ndash5 independent preparations) The expression levels of the indicated metabolic markers were measured by qRT-PCR Expression levels are presented as relative values (to NC-fed ATF3ff mice determined as 1 for each gene) The results represent the meansplusmn SDP lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes
142 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
had higher blood insulin level (Figure 7C) Therefore upon HFD chal-lenge cardiac-deficiency of ATF3 exacerbates HFD-induced T2D Thislsquoperipheral effectrsquo is surprising since ATF3 is selectively deleted in thecardiomyocytes
The question then is how can we explain the effect of cardiacATF3 deficiency on peripheral glucose intolerance One possibility isthe reduced IRS1 expression in ATF3-deficient cardiomyocytes bothin vivo and in vitro IRS1 suppression may lead to impaired insulin sig-nalling in the heart thereby exacerbating glucose intolerance Indeedwe observed that HFD-fed ATF3-cKO mice displayed insulin resist-ance upon insulin challenge when compared to HFD-fed ATF3ff
mice In addition palmitate-treated cardiomyocytes lacking ATF3 dis-played lower insulin signalling as compared to their wild type coun-terparts Since the heart is only a minor glucose consuming organ incomparison to skeletal muscle cells or the brain the peripheral sys-temic effect cannot be solely based on this observation A case inpoint transgenic mice with insulin receptor (IR) deficiency in theheart do not display a diabetic phenotype4546 Another attractive
possibility for the peripheral effect of cardiac ATF3-deletion is itsimpact on inflammation It is well established that low-grade inflam-mation years in advance of the disease onset is involved in the patho-genic processes causing T2D8 Mediators of inflammation such asTNFa interleukin (IL)-1b the IL-6 family of cytokines IL-18 and cer-tain chemokines have been proposed to be involved in the eventscausing both forms of diabetes (T1D and T2D)8 Interestingly wehave shown that IL-6 a well-known ATF3 target gene47 is known toenhance its transcription and secretion of cytokines such as IL-6 andTNFa in palmitate-treated primary cultures of ATF3-cKO cardio-myocytes as well as in cardiomyocytes derived from HFD-fed ATF3-cKO mice IL-6 has been proposed to play a role in glucose homeo-stasis and metabolism directly and indirectly via its action on skeletalmuscle cells adipocytes hepatocytes pancreatic b-cells and neuro-endocrine cells48
Therefore we hypothesize that elevated levels of inflammatory cyto-kines in ATF3-cKO mice mediate a cross-talk in peripheral tissues leadingto impaired glucose tolerance Although circulating IL-6 and TNFa
A
B
C
D
E
F
Figure 6 HFD-fed ATF3-cKO mice present deteriorated heart function ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) At endpointmice hearts were examined by micro ultrasound The following parameters were measured interventricular septal end diastole (IVSd) left ventricular pos-terior wall end diastole (LVPWd) maximal left ventricular internal end-diastole (LVIDd) end-systole (LVIDs) and heart rate The results represent themeansplusmn SD (n = 6ndash8 in each group as shown in panel B) (B) Heart function was assessed by calculating percentage of fractional shortening (FS) accordingto FS () = [(LVDd-LVDs)LVDd]100 The results represent the meansplusmn SD (n = 6ndash8 in each group) (C and D) Left ventricular cardiac volumes massand function were examined by a cardiac MRI The following parameters were measured left ventricular end-diastolic (LVEDV) and left ventricular end-sys-tolic volume (LVESV) left ventricular (LV) mass and ejection fraction (EF) The results represent the meansplusmn SD (n = 7 in each group) (E and F)Representative images of mid-ventricular short-axis (E) and long-axis (F) slice at peak diastole and systole P lt_ 005 difference between diets daggerP lt_ 005 dif-ference between genotypes
ATF3 expression in cardiomyocytes preserves homeostasis 143D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
cytokines may be derived from different tissues we provide ample evi-dence that ATF3-defficient cardiomyocytes produce and secrete higherlevels of these cytokines as compared to their wild type counterpartsThis indicates a lsquocardiac-to-peripheralrsquo effect in T2D-induced cardiomyop-athy This is a reverse direction from the lsquoperipheral-to-cardiacrsquo effect dis-cussed in the introduction where inflammation in the peripheral tissuesleads to cardiac dysfunction Since inflammatory cytokines are soluble
factors they can exert a systemic effect This is why it makes sense thattheir effect can go both ways Thus diabetes and cardiomyopathy are apart of a vicious cycle Previously it was shown that cardiac specific dele-tion of the thyroid hormone complex mediator MED13 results in aperipheral effect upon HFD Cardiac MED13 deficiency suppresses meta-bolic genes in the heart which leads to a decrease in energy expenditureand an obese phenotype most likely by the secretion of cardiokines that
A
B
C
D
E
Figure 7 HFD-fed ATF3-cKO mice exhibit impaired glucose tolerance ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) Fasting (6 h)blood glucose concentrations (mgdl) The results represent the meansplusmn SD (n = 20ndash30group) P lt_ 005 difference between diets daggerP lt_ 005 differencebetween genotypes (B) Glucose tolerance test (GTT) Values are presented as relative to initial valueplusmn SD (n = 20ndash25group) P lt_ 005 difference betweendiets daggerP lt_ 005 difference between genotypes (C) Serum insulin concentrations were quantified by ELISA Values are presented as relative values (com-pared to NC-fed ATF3ff mice determined as 1 n = 15ndash20group) P lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes (D and E)Effect of HFD on phosphorylated Akt (Ser473 pAkt) to Akt ratio levels in heart lysate following either saline or insulin injection (10 U1 kg) is shownRepresentative blots (D) Densitometric analysis is presented as means ratio (n = 3ndash6group) pAktAkt ratio is presented as relative values (compared toHFD-fed saline injected ATF3ff mice determined as 1) P lt_ 005 insulin vs saline treatment daggerP lt_ 005 difference between genotypes (E)
144 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
mediates the cross talk between the heart and peripheral tissues49ndash51
Here we also observed increased levels of natriuretic peptides (ANP andBNP) which together with IL-6 and TNFa may mediate the cross-talk withthe peripheral tissues to induce glucose intolerance
Does this finding have any relevance to cardiac disease in humansSeveral lines of evidence suggests that congestive heart failure in patientsis associated with an increase in blood glucose levels52 In addition highblood glucose levels in non-diabetic patients with acute myocardialinfarction serves as a prognostic predictor of long-term mortality5354
Thus here we provide evidence that high blood glucose levels are a partof the symptoms of cardiac remodeling process
Collectively our findings suggest that cardiac ATF3 expression isresponsible for an adaptive cardiac response to the stressful environmentunder a HFD (Figure 8) ATF3 dampens the inflammatory response andsuppresses the expression of ECM remodeling genes and thus amelioratescardiac function In HFD-fed mice with ATF3 deficiency or in mice withcardiac remodeling the inflamed heart affects also peripheral tissuesthereby worsening T2D in peripheral tissues resulting in elevated bloodglucose level and turning on a vicious cycle to worsen cardiomyopathy
Supplementary material
Supplementary material is available at Cardiovascular Research online
AcknowledgementsThe authors wish to thank Drs Keren Bitton-Worms and Ran Rostokerand Prof Derek LeRoith from the Rambam Medical Center for their
assistance in GTT and ITT Dr Edith Suss Toby Maya Holdengreber andLior Liba from the BCF Bioimaging Center (Faculty of MedicineTechnion) for their assistance with imaging and image analysis Prof DEMichelle from University of Michigan for cardiomyocytes culture techni-que and Prof Yehiel Zick from the Weizmann institute of Sciences forcritical comments
Conflict of interest none declared
FundingThis work was partially supported by the United States-Israel BinationalGrant 2009179 to AA and TH In addition we wish to thank the MarsPittsburgh Fund for Medical Research to AA the Department of Defensegrant W81XWH-14-1-0179 to TH and to D Dan and Betty Kahn travelgrant support to LK
References
1 Roger VL Go AS Lloyd-Jones DM Benjamin EJ Berry JD Borden WB Bravata DMDai S Ford ES Fox CS Fullerton HJ Gillespie C Hailpern SM Heit JA Howard VJKissela BM Kittner SJ Lackland DT Lichtman JH Lisabeth LD Makuc DM MarcusGM Marelli A Matchar DB Moy CS Mozaffarian D Mussolino ME Nichol GPaynter NP Soliman EZ Sorlie PD Sotoodehnia N Turan TN Virani SS Wong NDWoo D Turner MB American Heart Association Statistics C Stroke Statistics SHeart disease and stroke statisticsndash2012 update a report from the American HeartAssociation Circulation 2012125e2ndashe220
2 Wilson C Diabetes ACCORD 5-year outcomes of intensive glycemic control NatRev Endocrinol 20117314
3 Ernande L Derumeaux G Diabetic cardiomyopathy myth or reality Arch CardiovascDis 2012105218ndash225
4 Battiprolu PK Hojayev B Jiang N Wang ZV Luo X Iglewski M Shelton JM GerardRD Rothermel BA Gillette TG Lavandero S Hill JA Metabolic stress-induced
InsulinGF
Cytokines
FFA
GlucoseATF3
Cardiac
Remodeling
Factors
Paracrine action
Collagens
and ECM
proteins
Fibrosis
Cardiac
Dysfunction
Systemic Effects
Glucose Tolerance
TNFα
IL-6
Cardiomyocyte
ATF3
Homeostasis
Homeostasis
TNFα
IL-6
TIMP1
BNP
ANP
Figure 8 lsquoPeripheral-to-cardiacrsquo and lsquoCardiac-to-peripheralrsquo model scheme Schematic diagram showing the ATF3 adaptive role in maintaining hearthomeostasis following HFD-induced T2D In contrast stressful environment under HFD leads to maladaptive cardiac response in the absence of ATF3Cardiac remodeling caused by ATF3 deficiency induces an increase in cardiac ECM remodeling and inflammatory genes and mediates the communication ofcardiomyocytes with heart resident cells potentially mediated by IL-6 TNFa ANP BNP and TIMP1 In this situation increased cardiac fibrosis and inflamma-tory response worsen cardiac function The inflamed failing heart mediates the communication of cardiomyocytes with distant organs as well thereby affect-ing diabetic outcome and glucose homeostasis
ATF3 expression in cardiomyocytes preserves homeostasis 145D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
activation of FoxO1 triggers diabetic cardiomyopathy in mice J Clin Invest20121221109ndash1118
5 Mandavia CH Aroor AR Demarco VG Sowers JR Molecular and metabolic mecha-nisms of cardiac dysfunction in diabetes Life Sci 201392601ndash608
6 Ventura-Clapier R Garnier A Veksler V Energy metabolism in heart failure J Physiol20045551ndash13
7 Guo S Insulin signaling resistance and the metabolic syndrome insights from mousemodels into disease mechanisms J Endocrinol 2014220T1ndashT23
8 Montane J Cadavez L Novials A Stress and the inflammatory process a major causeof pancreatic cell death in type 2 diabetes Diabetes Metab Syndr Obes 2014725ndash34
9 Ashrafian H Frenneaux MP Opie LH Metabolic mechanisms in heart failureCirculation 2007116434ndash448
10 Tanabe K Liu Y Hasan SD Martinez SC Cras-Meneur C Welling CM Bernal-MizrachiE Tanizawa Y Rhodes CJ Zmuda E Hai T Abumrad NA Permutt MA Glucose andfatty acids synergize to promote B-cell apoptosis through activation of glycogen syn-thase kinase 3beta independent of JNK activation PLoS One 20116e18146
11 Heineke J Molkentin JD Regulation of cardiac hypertrophy by intracellular signallingpathways Nat Rev Mol Cell Biol 20067589ndash600
12 Hai T Wolford CC Chang YS ATF3 a hub of the cellular adaptive-response net-work in the pathogenesis of diseases is modulation of inflammation a unifying com-ponent Gene Expr 2010151ndash11
13 Takii R Inouye S Fujimoto M Nakamura T Shinkawa T Prakasam R Tan K HayashidaN Ichikawa H Hai T Nakai A Heat shock transcription factor 1 inhibits expression ofIL-6 through activating transcription factor 3 J Immunol 20101841041ndash1048
14 Giraldo A Barrett OP Tindall MJ Fuller SJ Amirak E Bhattacharya BS Sugden PHClerk A Feedback regulation by Atf3 in the endothelin-1-responsive transcriptomeof cardiomyocytes Egr1 is a principal Atf3 target Biochem J 2012444343ndash355
15 Hasin T Elhanani O Abassi Z Hai T Aronheim A Angiotensin II signaling up-regulates the immediate early transcription factor ATF3 in the left but not the rightatrium Basic Res Cardiol 2011106175ndash187
16 Koren L Elhanani O Kehat I Hai T Aronheim A Adult cardiac expression of theactivating transcription factor 3 ATF3 promotes ventricular hypertrophy PLoS One20138e68396
17 Koren L Alishekevitz D Elhanani O Nevelsky A Hai T Kehat I Shaked Y AronheimA ATF3-dependent cross-talk between cardiomyocytes and macrophages promotescardiac maladaptive remodeling Int J Cardiol 2015198232ndash240
18 Okamoto Y Chaves A Chen J Kelley R Jones K Weed HG Gardner KL Gangi LYamaguchi M Klomkleaw W Nakayama T Hamlin RL Carnes C Altschuld R BauerJ Hai T Transgenic mice with cardiac-specific expression of activating transcriptionfactor 3 a stress-inducible gene have conduction abnormalities and contractile dys-function Am J Pathol 2001159639ndash650
19 Zhou H Shen DF Bian ZY Zong J Deng W Zhang Y Guo YY Li H Tang QZActivating transcription factor 3 deficiency promotes cardiac hypertrophy dysfunc-tion and fibrosis induced by pressure overload PLoS One 20116e26744
20 Zhou H Guo H Zong J Dai J Yuan Y Bian ZY Tang QZ ATF3 regulates multipletargets and may play a dual role in cardiac hypertrophy and injury Int J Cardiol2014174838ndash839
21 Yang J Yang CJ Yang J Fan ZX ATF3 a promotion effect or a inhibition effect in car-diac maladaptive remodeling Int J Cardiol 2015201245ndash246
22 Koren L Shaked Y Aronheim A Response letter ldquoATF3 a promoter or inhibitor ofcardiac maladaptive remodelingrdquo Int J Cardiol 2015201692
23 Ma ZG Wei WY Xu SC Zhang WB Dai J Tang QZ ATF3 a potential target forcardiac maladaptive remodeling Int J Cardiol 201620250ndash51
24 Zhou H Yuan Y Ni J Guo H Deng W Bian ZY Tang QZ Pleiotropic and puzzlingeffects of ATF3 in maladaptive cardiac remodeling Int J Cardiol 201620687ndash88
25 Zmuda EJ Qi L Zhu MX Mirmira RG Montminy MR Hai T The roles of ATF3 anadaptive-response gene in high-fat-diet-induced diabetes and pancreatic beta-celldysfunction Mol Endocrinol 2010241423ndash1433
26 Lu D Chen J Hai T The regulation of ATF3 gene expression by mitogen-activatedprotein kinases Biochem J 2007401559ndash567
27 Hartman MG Lu D Kim ML Kociba GJ Shukri T Buteau J Wang X Frankel WLGuttridge D Prentki M Grey ST Ron D Hai T Role for activating transcription fac-tor 3 in stress-induced beta-cell apoptosis Mol Cell Biol 2004245721ndash5732
28 Wolford CC McConoughey SJ Jalgaonkar SP Leon M Merchant AS Dominick JLYin X Chang Y Zmuda EJ OrsquoToole SA Millar EK Roller SL Shapiro CL OstrowskiMC Sutherland RL Hai T Transcription factor ATF3 links host adaptive response tobreast cancer metastasis J Clin Iinvest 20131232893ndash2906
29 Agah R Frenkel PA French BA Michael LH Overbeek PA Schneider MD Generecombination in postmitotic cells Targeted expression of Cre recombinase pro-vokes cardiac-restricted site-specific rearrangement in adult ventricular musclein vivo J Clin Invest 1997100169ndash179
30 Livet J Weissman TA Kang H Draft RW Lu J Bennis RA Sanes JR Lichtman JWTransgenic strategies for combinatorial expression of fluorescent proteins in thenervous system Nature 200745056ndash62
31 Davis J Maillet M Miano JM Molkentin JD Lost in transgenesis a userrsquos guidefor genetically manipulating the mouse in cardiac research Circ Res2012111761ndash777
32 Heiberg E Sjogren J Ugander M Carlsson M Engblom H Arheden H Design andvalidation of segment ndash freely available software for cardiovascular image analysisBMC Med Imaging 2010101
33 Kabaeva Z Zhao M Michele DE Blebbistatin extends culture life of adult mouse car-diac myocytes and allows efficient and stable transgene expression Am J Physiol HeartC 2008294H1667ndash1674
34 Subramaniam A Jones WK Gulick J Wert S Neumann J Robbins J Tissue-specificregulation of the alpha-myosin heavy chain gene promoter in transgenic mice J BiolChem 199126624613ndash24620
35 Cunha DA Hekerman P Ladriere L Bazarra-Castro A Ortis F Wakeham MCMoore F Rasschaert J Cardozo AK Bellomo E Overbergh L Mathieu C Lupi R HaiT Herchuelz A Marchetti P Rutter GA Eizirik DL Cnop M Initiation and executionof lipotoxic ER stress in pancreatic beta-cells J Cell Sci 20081212308ndash2318
36 Wynn TA Ramalingam TR Mechanisms of fibrosis therapeutic translation forfibrotic disease Nat Med 2012181028ndash1040
37 Li D Yin X Zmuda EJ Wolford CC Dong X White MF Hai T The repression ofIRS2 gene by ATF3 a stress-inducible gene contributes to pancreatic beta-cell apop-tosis Diabetes 200857635ndash644
38 Myerson SG Bellenger NG Pennell DJ Assessment of left ventricular mass by cardi-ovascular magnetic resonance Hypertension 200239750ndash755
39 Zmuda EJ Viapiano M Grey ST Hadley G Garcia-Ocana A Hai T Deficiency ofAtf3 an adaptive-response gene protects islets and ameliorates inflammation in asyngeneic mouse transplantation model Diabetologia 2010531438ndash1450
40 Riehle C Abel ED Insulin signaling and heart failure Circ Res 20161181151ndash116941 Schreyer SA Wilson DL LeBoeuf RC C57BL6 mice fed high fat diets as models for
diabetes-accelerated atherosclerosis Atherosclerosis 199813617ndash2442 Brainard RE Watson LJ Demartino AM Brittian KR Readnower RD Boakye AA
Zhang D Hoetker JD Bhatnagar A Baba SP Jones SP High fat feeding in mice isinsufficient to induce cardiac dysfunction and does not exacerbate heart failure PLoSOne 20138e83174
43 Gupte AA Minze LJ Reyes M Ren Y Wang X Brunner G Ghosn M Cordero-Reyes AM Ding K Pratico D Morrisett J Shi ZZ Hamilton DJ Lyon CJ Hsueh WAHigh-fat feeding-induced hyperinsulinemia increases cardiac glucose uptake and mito-chondrial function despite peripheral insulin resistance Endocrinology20131542650ndash2662
44 Logan SK Garabedian MJ Campbell CE Werb Z Synergistic transcriptionalactivation of the tissue inhibitor of metalloproteinases-1 promoter via functionalinteraction of AP-1 and Ets-1 transcription factors J Biol Chem1996271774ndash782
45 Belke DD Betuing S Tuttle MJ Graveleau C Young ME Pham M Zhang DCooksey RC McClain DA Litwin SE Taegtmeyer H Severson D Kahn CR AbelED Insulin signaling coordinately regulates cardiac size metabolism and contractileprotein isoform expression J Clin Invest 2002109629ndash639
46 Fernandez AM Kim JK Yakar S Dupont J Hernandez-Sanchez C Castle AL FilmoreJ Shulman GI Le Roith D Functional inactivation of the IGF-I and insulin receptors inskeletal muscle causes type 2 diabetes Genes Dev 2001151926ndash1934
47 Gilchrist M Thorsson V Li B Rust AG Korb M Roach JC Kennedy K Hai TBolouri H Aderem A Systems biology approaches identify ATF3 as a negative regu-lator of Toll-like receptor 4 Nature 2006441173ndash178
48 Kristiansen OP Mandrup-Poulsen T Interleukin-6 and diabetes the good the bad orthe indifferent Diabetes 200554(Suppl 2)S114ndashS124
49 Grueter CE van Rooij E Johnson BA DeLeon SM Sutherland LB Qi X Gautron LElmquist JK Bassel-Duby R Olson EN A cardiac microRNA governs systemic energyhomeostasis by regulation of MED13 Cell 2012149671ndash683
50 Baskin KK Grueter CE Kusminski CM Holland WL Bookout AL Satapati S KongYM Burgess SC Malloy CR Scherer PE Newgard CB Bassel-Duby R Olson ENMED13-dependent signaling from the heart confers leanness by enhancing metabo-lism in adipose tissue and liver EMBO Mol Med 201461610ndash1621
51 Nakamura M Sadoshima J Heart over mind metabolic control of white adipose tis-sue and liver EMBO Mol Med 201461521ndash1524
52 Tenenbaum A Fisman EZ Impaired glucose metabolism in patients with heart failurepathophysiology and possible treatment strategies Am J Cardiovasc Drugs20044269ndash280
53 Suskin N McKelvie RS Burns RJ Latini R Pericak D Probstfield J Rouleau JL SigouinC Solymoss CB Tsuyuki R White M Yusuf S Glucose and insulin abnormalitiesrelate to functional capacity in patients with congestive heart failure Eur Heart J2000211368ndash1375
54 Aronson D Hammerman H Kapeliovich MR Suleiman A Agmon Y Beyar RMarkiewicz W Suleiman M Fasting glucose in acute myocardial infarction incremen-tal value for long-term mortality and relationship with left ventricular systolic func-tion Diabetes Care 200730960ndash966
146 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
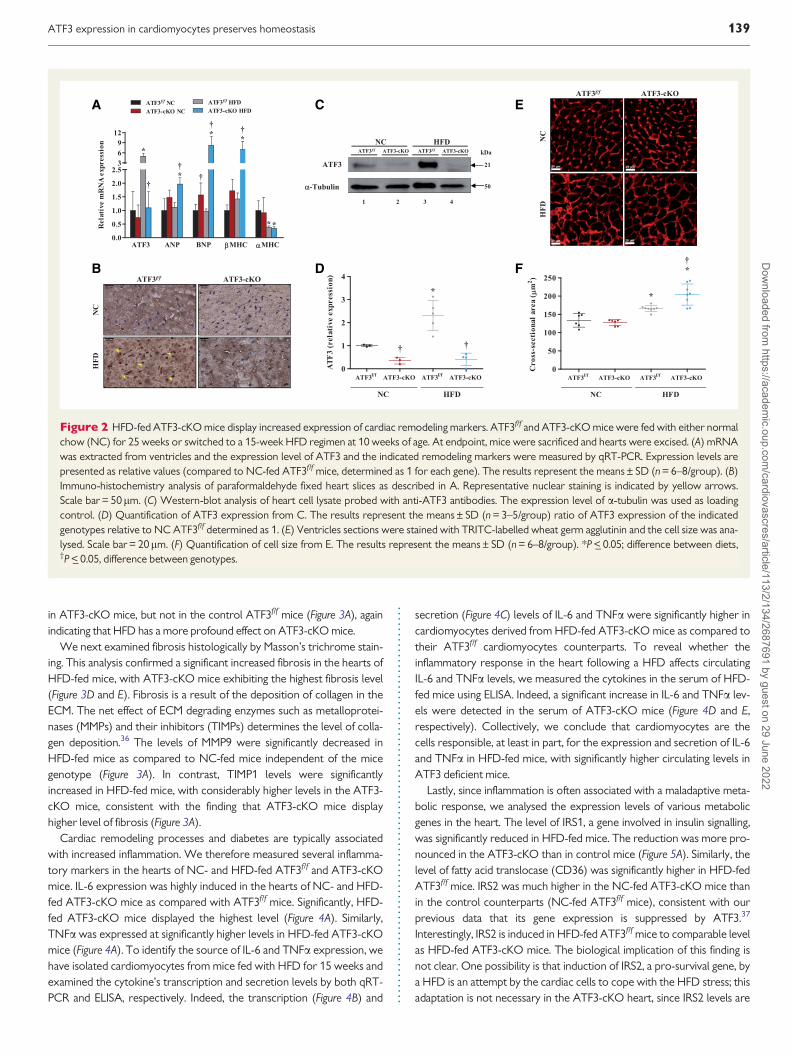
in ATF3-cKO mice but not in the control ATF3ff mice (Figure 3A) againindicating that HFD has a more profound effect on ATF3-cKO mice
We next examined fibrosis histologically by Massonrsquos trichrome stain-ing This analysis confirmed a significant increased fibrosis in the hearts ofHFD-fed mice with ATF3-cKO mice exhibiting the highest fibrosis level(Figure 3D and E) Fibrosis is a result of the deposition of collagen in theECM The net effect of ECM degrading enzymes such as metalloprotei-nases (MMPs) and their inhibitors (TIMPs) determines the level of colla-gen deposition36 The levels of MMP9 were significantly decreased inHFD-fed mice as compared to NC-fed mice independent of the micegenotype (Figure 3A) In contrast TIMP1 levels were significantlyincreased in HFD-fed mice with considerably higher levels in the ATF3-cKO mice consistent with the finding that ATF3-cKO mice displayhigher level of fibrosis (Figure 3A)
Cardiac remodeling processes and diabetes are typically associatedwith increased inflammation We therefore measured several inflamma-tory markers in the hearts of NC- and HFD-fed ATF3ff and ATF3-cKOmice IL-6 expression was highly induced in the hearts of NC- and HFD-fed ATF3-cKO mice as compared with ATF3ff mice Significantly HFD-fed ATF3-cKO mice displayed the highest level (Figure 4A) SimilarlyTNFa was expressed at significantly higher levels in HFD-fed ATF3-cKOmice (Figure 4A) To identify the source of IL-6 and TNFa expression wehave isolated cardiomyocytes from mice fed with HFD for 15 weeks andexamined the cytokinersquos transcription and secretion levels by both qRT-PCR and ELISA respectively Indeed the transcription (Figure 4B) and
secretion (Figure 4C) levels of IL-6 and TNFa were significantly higher incardiomyocytes derived from HFD-fed ATF3-cKO mice as compared totheir ATF3ff cardiomyocytes counterparts To reveal whether theinflammatory response in the heart following a HFD affects circulatingIL-6 and TNFa levels we measured the cytokines in the serum of HFD-fed mice using ELISA Indeed a significant increase in IL-6 and TNFa lev-els were detected in the serum of ATF3-cKO mice (Figure 4D and Erespectively) Collectively we conclude that cardiomyocytes are thecells responsible at least in part for the expression and secretion of IL-6and TNFa in HFD-fed mice with significantly higher circulating levels inATF3 deficient mice
Lastly since inflammation is often associated with a maladaptive meta-bolic response we analysed the expression levels of various metabolicgenes in the heart The level of IRS1 a gene involved in insulin signallingwas significantly reduced in HFD-fed mice The reduction was more pro-nounced in the ATF3-cKO than in control mice (Figure 5A) Similarly thelevel of fatty acid translocase (CD36) was significantly higher in HFD-fedATF3ff mice IRS2 was much higher in the NC-fed ATF3-cKO mice thanin the control counterparts (NC-fed ATF3ff mice) consistent with ourprevious data that its gene expression is suppressed by ATF337
Interestingly IRS2 is induced in HFD-fed ATF3ff mice to comparable levelas HFD-fed ATF3-cKO mice The biological implication of this finding isnot clear One possibility is that induction of IRS2 a pro-survival gene bya HFD is an attempt by the cardiac cells to cope with the HFD stress thisadaptation is not necessary in the ATF3-cKO heart since IRS2 levels are
A
B
C
D
E
F
Figure 2 HFD-fed ATF3-cKO mice display increased expression of cardiac remodeling markers ATF3ff and ATF3-cKO mice were fed with either normalchow (NC) for 25 weeks or switched to a 15-week HFD regimen at 10 weeks of age At endpoint mice were sacrificed and hearts were excised (A) mRNAwas extracted from ventricles and the expression level of ATF3 and the indicated remodeling markers were measured by qRT-PCR Expression levels arepresented as relative values (compared to NC-fed ATF3ff mice determined as 1 for each gene) The results represent the meansplusmn SD (n = 6ndash8group) (B)Immuno-histochemistry analysis of paraformaldehyde fixed heart slices as described in A Representative nuclear staining is indicated by yellow arrowsScale bar = 50lm (C) Western-blot analysis of heart cell lysate probed with anti-ATF3 antibodies The expression level of a-tubulin was used as loadingcontrol (D) Quantification of ATF3 expression from C The results represent the meansplusmn SD (n = 3ndash5group) ratio of ATF3 expression of the indicatedgenotypes relative to NC ATF3ff determined as 1 (E) Ventricles sections were stained with TRITC-labelled wheat germ agglutinin and the cell size was ana-lysed Scale bar = 20lm (F) Quantification of cell size from E The results represent the meansplusmn SD (n = 6ndash8group) P lt_ 005 difference between dietsdaggerP lt_ 005 difference between genotypes
ATF3 expression in cardiomyocytes preserves homeostasis 139D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
already high The expression levels of other lipid and glycolytic regulatorssuch as peroxisome proliferator-activated receptor alpha (PPARa) carni-tine palmitoyltransferase 1 and 2 (CPT12) and pyruvate dehydrogenasekinase 4 (PDK4) were all elevated in HFD-fed mice with no significant dif-ference between the genotypes (Figure 5A) Lastly the level of the glucosetransporter 4 (Glut4) were unaffected by both diet and genotype at themRNA level (Figure 5A) However Glut4 was significantly elevated at theprotein level in HFD-fed mice with no significant difference between thegenotypes (see Supplementary material online Figure S4A and B)Collectively the expression of key metabolic genes was unalteredbetween the various genotypes in response to HFD In view of thechanges observed in metabolic genes in cardiomyocytes treated with pal-mitate (Figure 1A) this result was unexpected Thus we hypothesizedthat ATF3-dependent alterations in metabolic gene expression in cardio-myocytes may be masked by non-cardiomyocytes cells in the heartTherefore we studied the metabolic gene expression changes in isolatedcardiomyocytes derived from HFD- and NCndashfed mice from both ATF3ff
and ATF3-cKO mice (Figure 5B) Indeed the expression of all metabolicgenes tested was augmented in cardiomyocytes derived from HFD-fed ascompared with NC-fed mice Importantly the expression level of CD36CPT1 and CPT2 were significantly upregulated in HFD-fed ATF3-cKOmice as compared with their ATF3ff counterparts (Figure 5B) In contrastthe induction of both IRS1 and IRS2 were significantly lower in cardio-myocytes derived from HFD-fed ATF3-cKO mice (Figure 5B)Collectively the deleterious effects of HFD on cardiac remodeling fibro-sis and inflammation are more pronounced in ATF3-cKO mice suggest-ing that cardiac ATF3 plays a protective role in the HFD paradigm
35 HFD-induced cardiac dysfunctionNext we characterized the effect of HFD and cardiac remodeling oncardiac structure and function in ATF3ff and ATF3-cKO mice by
echocardiography Mice fed with NC had normal heart dimensions andfractional shortening (FS) (Figure 6A and B) HFD significantly reduced FSin both genotypes of HFD-fed mice as compared to corresponding NC-fed mice (Figure 6B) However the reduction was significantly more pro-nounced in the ATF3-cKO mice suggesting deteriorated systolic func-tion in these mice (Figure 6B) The reduced FS in the HFD-fed ATF3-cKO mice seems to be largely driven by an increase in LV internal end-systolic (LVIDs) dimension (Figure 6A) Heart rate was slightly increasedin the HFD-fed mice however this increase cannot explain the cardiacdysfunction observed in ATF3-cKO mice (Figure 6B) In addition weused cardiac MRI the gold standard for LV function and mass asses-sment38 LV mass was significantly higher in the ATF3-cKO mice follow-ing a 15-week HFD regimen (Figure 6C) In addition end-diastolic andend-systolic volumes were significantly larger in ATF3-cKO mice as com-pared to ATF3ff mice Moreover ATF3-cKO mice displayed a signifi-cantly lower ejection fraction as compared to ATF3ff mice (Figure 6D)These results are consistent with the reduced cardiac function found inthe echocardiographic analysis Images of mid-ventricular short-axis(Figure 6E) and long-axis (Figure 6E) slices are demonstrating the cardiaceccentric pattern observed in HFD-fed ATF3-cKO mice Collectivelythe results from the MRI show a greater increase in LV mass and volumein the ATF3-cKO mice as compared to the ATF3ff mice following aHFD suggesting fundamental structural differences in the heart betweenthe ATF3-cKO and ATF3ff mice
36 Impaired glucose tolerance in HFDATF3-cKO miceLastly we measured fasting glucose levels to assess the diabetic state ofthe mice As expected higher glucose levels were observed in HFD-fedmice as compared to NC-fed counterparts Interestingly the
A B
C
D
E
Figure 3 HFD-fed ATF3-cKO mice have higher level of fibrosis in the heart ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) mRNA wasextracted from ventricles and the expression levels of the indicated fibrosis markers were measured by qRT-PCR The expression levels are presented asrelative values (compared to NC-fed ATF3ff mice determined as 1 for each gene n = 6ndash8group) (B) Western-blot analysis of heart cell lysate probed withanti-a-SMA antibody The expression level of a-tubulin was used as loading control (C) Quantification of aSMA expression from B The results representthe meansplusmn SD (n= 3ndash5group) ratio of aSMA expression of the indicated genotypes relative to NC ATF3ff determined as 1 (D) Paraffin-embedded heartsections were stained with Massonrsquos trichrome to visualize fibrosis Scale bar = 100 lm (E) The level of fibrosis () stained by Massonrsquos trichrome was quan-tified and the results represent the meansplusmn SD (n = 6ndash8group) P lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes
140 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
hyperglycemia state of the ATF3-cKO mice was significantly higher thanin the ATF3ff mice following 15 weeks of HFD (Figure 7A) Consistentlythe glucose tolerance test (GTT) revealed that HFD-fed ATF3-cKOmice displayed higher blood glucose levels and longer glucose clearancetime when compared to HFD-fed ATF3ff whereas NC-fed ATF3ff andATF3-cKO mice had normal glucose clearance (Figure 7B)
Interestingly whole body ATF3-KO mice fed with HFD also displayedimpaired glucose clearance (see Supplementary material onlineFigure S5A)39 Previously we reported that decreased glucose tolerancein whole body ATF3-KO mice is at least in part due to reduced insulingene expression in b-cells (see Supplementary material online FigureS5B)39 To determine insulin level in HFD-fed ATF3-cKO mice weanalysed blood insulin levels by ELISA In both HFD-fed ATF3ff andATF3-cKO mice the serum insulin levels were significantly higher ascompared to their NC-fed counterpart mice with the ATF3-cKO micedisplaying the highest insulin levels (Figure 7C) Together these resultssuggest that peripheral tissues of ATF3-cKO mice have reduced ability
to uptake glucose despite their high insulin level in the blood Next wesought to measure the protein levels of components along the insulinpathway Akt and Foxo340 Both Akt and Foxo3 protein levels were sim-ilar in HFD-fed mice independent of the genotype (see Supplementarymaterial online Figure S4A and C) Interestingly phosphorylation of Akt(Ser473) which indicates the activity of the insulin pathway was slightlyelevated in HFD-fed mice however this did not reach statistical signifi-cance and was similar in both of the genotypes (see Supplementary material online Figure S4D) To measure the activity of the insulin signallingpathway following insulin stimulation we used the ratio between phos-phorylated Akt (Ser473) and total Akt expression HFD-fed mice wereinjected with insulin (10 Ukg) and the heart liver and skeletal musclewere harvested 15 min after the injection (Figure 7D and E and seeSupplementary material online Figure S6AndashD) Insulin injection resultedin a significant increase of pAktAkt ratio in the hearts of ATF3ff micewhile no significant change was observed in Akt phosphorylation inATF3-cKO mice (Figure 7D and E) Similarly low Akt phosphorylation
A
D E
B C
Figure 4 HFD-fed ATF3-cKO mice display inflammatory response in the heart (A) ATF3ff and ATF3-cKO mice were fed as described in Figure 2 mRNAwas extracted from ventricles and the expression levels of the indicated inflammatory markers were measured by qRT-PCR Expression levels are presentedas relative values (compared to NC-fed ATF3ff mice determined as 1 for each gene n= 6ndash8group) (B) Mice were fed as described in Figure 2Cardiomyocytes were isolated and primary cultures were generated mRNA was extracted from an overnight culture and the expression levels of IL-6 andTNFa were studied by qRT-PCR The mRNA level obtained in cardiomyocytes derived from NC-fed ATF3ff mice was determined as 1 and the relativeexpression of all other treatment is shown (n = 5ndash6 independent cardiomyocyte preparations) (C) Conditioned media was collected from cardiomyocytesdescribed in B and ELISA assay for the indicated cytokines is shown Cytokines level is presented as relative values to NC-fed ATF3ff mice determined as 1for each cytokine (n = 5ndash6 independent preparations) (D and E) Blood was collected from HFD-fed mice and IL-6 and TNFa serum levels were quantifiedby ELISA (n = 8ndash12group) The results represent the meansplusmn SD P lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes
ATF3 expression in cardiomyocytes preserves homeostasis 141D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
levels were also observed in both the liver and skeletal muscle of theATF3-cKO mice however this did not reach statistically significance (seeSupplementary material online Figure S6AndashD) Consistently palmitate-treated cardiomyocytes derived from ATF3-cKO mice displayedreduced phosphorylation of Akt following 30-min insulin stimulation(see Supplementary material online Figure S6E)
Collectively our findings revealed a surprising phenomenon that car-diac ATF3 deficiency results in increased cardiac remodeling leading topotentiation of local and peripheral insulin resistance
4 Discussion
A HFD is a commonly used regimen to induce T2D The majority of theliterature focuses on diabetic symptoms without addressing the issue ofT2D-induced cardiomyopathy Even when it is addressed there is noclear consensus regarding the extent of cardiac effects imposed byHFD41 Diet-induced obesity in mouse models is associated with globalinsulin resistance and increased myocardial fatty acid utilization how-ever the reported phenotypes on cardiac function vary from extensivehypertrophy and heart failure4 to lack of cardiac dysfunction4243 Thesevariations have been attributed to diet regimens andor sensitivity ofmouse strains to a HFD In this report we present evidence that cardiacATF3 deficiency exacerbates the deleterious effect of a HFD on theheart such as maladaptive cardiac remodeling fibrosis inflammation andcardiac dysfunction Our work may provide a potential molecular insightto variations in the literature regarding the impact of HFD on the heartAs ATF3 is a stress-inducible gene we propose that its proper inductionin the cardiomyocytes is protective and helps the heart to cope withHFD stress It is possible that the induction of ATF3mdashits expressionlevel duration and kineticsmdashvaries in different HFD regimens andormouse strains contributing to the variations observed in the literature
HFD regimen induced a significant higher level of fibrosis in the heartof ATF3-cKO mice We have identified TIMP1 as a possible mechanismto explain the increase in fibrosis in ATF3-cKO cardiomyocytes Severallines of evidence suggest that TIMP1 is possibly an ATF3 target gene
First we observed an increase in TIMP1 transcription in the whole heartand in isolated cardiomyocytes (Figures 1A and 3A) Second TIMP 1 3and 4 were identified as significantly highly expressed genes in the heartof ATF3-KO mice as compared with wild type mice following aorticbanding20 Third a functional AP-1 regulatory element within the TIMP1promoter was described44 Indeed preliminary results in our lab suggestthat ATF3 downregulates the TIMP1 promoter (data not shown) Thusit is possible that TIMP1 is suppressed by ATF3 and therefore results in anet increase in MMPs activity that helps to maintain ECM formationdeg-radation homeostasis
Our findings are consistent with previous studies demonstrating that adeficiency in ATF3 can promote cardiac maladaptive changes which leadto cardiac dilatation and hypertrophy in an aortic banding model1920 Incontrast a different study using a pressure overload model reportedthat ATF3 induces a maladaptive cardiac response17 These seeminglyconflicting data may be explained by the different patterns of ATF3expression induced by the different models In the pressure overloadmodel the acute and transient expression of ATF3 in cardiomyocytesinitiates a cascade of maladaptive changes17 whereas in the aortic band-ing model and the HFD model described here ATF3 expression is longlasting
In this context this is the first report using mice with cardiac-selectivedeficiency of ATF3 (ATF3-cKO) Previous reports on the roles of ATF3in cardiac function were carried out using whole body knockout ofATF325 or transgenic mice ectopically expressing ATF3 in cardiomyo-cytes16 A key advancement of ATF3-cKO mice over the whole bodyknockout mice is that they provide insights into the function of ATF3selectively in cardiomyocytes without the complication of ATF3 defi-ciency in other cells Thus the ATF3-cKO mouse is a useful tool to bet-ter understand the function of cardiac ATF3
Consistent with our previous report25 here we showed that cardiacATF3 deficiency has a more severe HFD-induced diabetic phenotypethan the control wild type mice However while reduced insulin genetranscription in ATF3-deficient b-cells was observed25 our current dataindicates that a lack of insulin is not the explanation for the glucose intol-erance found in the HFD-fed ATF3-cKO mice In contrast these mice
A B
Figure 5 HFD-fed ATF3-cKO mice show metabolic response in the heart ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) mRNA wasextracted from ventricles and the expression levels of the indicated metabolic markers were measured by qRT-PCR Expression levels are presented as rela-tive values (compared to NC-fed ATF3ff mice determined as 1 for each gene n= 6ndash8group) (B) Cardiomyocytes were isolated and plated on laminin for1 h followed by mRNA preparation (n = 4ndash5 independent preparations) The expression levels of the indicated metabolic markers were measured by qRT-PCR Expression levels are presented as relative values (to NC-fed ATF3ff mice determined as 1 for each gene) The results represent the meansplusmn SDP lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes
142 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
had higher blood insulin level (Figure 7C) Therefore upon HFD chal-lenge cardiac-deficiency of ATF3 exacerbates HFD-induced T2D Thislsquoperipheral effectrsquo is surprising since ATF3 is selectively deleted in thecardiomyocytes
The question then is how can we explain the effect of cardiacATF3 deficiency on peripheral glucose intolerance One possibility isthe reduced IRS1 expression in ATF3-deficient cardiomyocytes bothin vivo and in vitro IRS1 suppression may lead to impaired insulin sig-nalling in the heart thereby exacerbating glucose intolerance Indeedwe observed that HFD-fed ATF3-cKO mice displayed insulin resist-ance upon insulin challenge when compared to HFD-fed ATF3ff
mice In addition palmitate-treated cardiomyocytes lacking ATF3 dis-played lower insulin signalling as compared to their wild type coun-terparts Since the heart is only a minor glucose consuming organ incomparison to skeletal muscle cells or the brain the peripheral sys-temic effect cannot be solely based on this observation A case inpoint transgenic mice with insulin receptor (IR) deficiency in theheart do not display a diabetic phenotype4546 Another attractive
possibility for the peripheral effect of cardiac ATF3-deletion is itsimpact on inflammation It is well established that low-grade inflam-mation years in advance of the disease onset is involved in the patho-genic processes causing T2D8 Mediators of inflammation such asTNFa interleukin (IL)-1b the IL-6 family of cytokines IL-18 and cer-tain chemokines have been proposed to be involved in the eventscausing both forms of diabetes (T1D and T2D)8 Interestingly wehave shown that IL-6 a well-known ATF3 target gene47 is known toenhance its transcription and secretion of cytokines such as IL-6 andTNFa in palmitate-treated primary cultures of ATF3-cKO cardio-myocytes as well as in cardiomyocytes derived from HFD-fed ATF3-cKO mice IL-6 has been proposed to play a role in glucose homeo-stasis and metabolism directly and indirectly via its action on skeletalmuscle cells adipocytes hepatocytes pancreatic b-cells and neuro-endocrine cells48
Therefore we hypothesize that elevated levels of inflammatory cyto-kines in ATF3-cKO mice mediate a cross-talk in peripheral tissues leadingto impaired glucose tolerance Although circulating IL-6 and TNFa
A
B
C
D
E
F
Figure 6 HFD-fed ATF3-cKO mice present deteriorated heart function ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) At endpointmice hearts were examined by micro ultrasound The following parameters were measured interventricular septal end diastole (IVSd) left ventricular pos-terior wall end diastole (LVPWd) maximal left ventricular internal end-diastole (LVIDd) end-systole (LVIDs) and heart rate The results represent themeansplusmn SD (n = 6ndash8 in each group as shown in panel B) (B) Heart function was assessed by calculating percentage of fractional shortening (FS) accordingto FS () = [(LVDd-LVDs)LVDd]100 The results represent the meansplusmn SD (n = 6ndash8 in each group) (C and D) Left ventricular cardiac volumes massand function were examined by a cardiac MRI The following parameters were measured left ventricular end-diastolic (LVEDV) and left ventricular end-sys-tolic volume (LVESV) left ventricular (LV) mass and ejection fraction (EF) The results represent the meansplusmn SD (n = 7 in each group) (E and F)Representative images of mid-ventricular short-axis (E) and long-axis (F) slice at peak diastole and systole P lt_ 005 difference between diets daggerP lt_ 005 dif-ference between genotypes
ATF3 expression in cardiomyocytes preserves homeostasis 143D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
cytokines may be derived from different tissues we provide ample evi-dence that ATF3-defficient cardiomyocytes produce and secrete higherlevels of these cytokines as compared to their wild type counterpartsThis indicates a lsquocardiac-to-peripheralrsquo effect in T2D-induced cardiomyop-athy This is a reverse direction from the lsquoperipheral-to-cardiacrsquo effect dis-cussed in the introduction where inflammation in the peripheral tissuesleads to cardiac dysfunction Since inflammatory cytokines are soluble
factors they can exert a systemic effect This is why it makes sense thattheir effect can go both ways Thus diabetes and cardiomyopathy are apart of a vicious cycle Previously it was shown that cardiac specific dele-tion of the thyroid hormone complex mediator MED13 results in aperipheral effect upon HFD Cardiac MED13 deficiency suppresses meta-bolic genes in the heart which leads to a decrease in energy expenditureand an obese phenotype most likely by the secretion of cardiokines that
A
B
C
D
E
Figure 7 HFD-fed ATF3-cKO mice exhibit impaired glucose tolerance ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) Fasting (6 h)blood glucose concentrations (mgdl) The results represent the meansplusmn SD (n = 20ndash30group) P lt_ 005 difference between diets daggerP lt_ 005 differencebetween genotypes (B) Glucose tolerance test (GTT) Values are presented as relative to initial valueplusmn SD (n = 20ndash25group) P lt_ 005 difference betweendiets daggerP lt_ 005 difference between genotypes (C) Serum insulin concentrations were quantified by ELISA Values are presented as relative values (com-pared to NC-fed ATF3ff mice determined as 1 n = 15ndash20group) P lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes (D and E)Effect of HFD on phosphorylated Akt (Ser473 pAkt) to Akt ratio levels in heart lysate following either saline or insulin injection (10 U1 kg) is shownRepresentative blots (D) Densitometric analysis is presented as means ratio (n = 3ndash6group) pAktAkt ratio is presented as relative values (compared toHFD-fed saline injected ATF3ff mice determined as 1) P lt_ 005 insulin vs saline treatment daggerP lt_ 005 difference between genotypes (E)
144 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
mediates the cross talk between the heart and peripheral tissues49ndash51
Here we also observed increased levels of natriuretic peptides (ANP andBNP) which together with IL-6 and TNFa may mediate the cross-talk withthe peripheral tissues to induce glucose intolerance
Does this finding have any relevance to cardiac disease in humansSeveral lines of evidence suggests that congestive heart failure in patientsis associated with an increase in blood glucose levels52 In addition highblood glucose levels in non-diabetic patients with acute myocardialinfarction serves as a prognostic predictor of long-term mortality5354
Thus here we provide evidence that high blood glucose levels are a partof the symptoms of cardiac remodeling process
Collectively our findings suggest that cardiac ATF3 expression isresponsible for an adaptive cardiac response to the stressful environmentunder a HFD (Figure 8) ATF3 dampens the inflammatory response andsuppresses the expression of ECM remodeling genes and thus amelioratescardiac function In HFD-fed mice with ATF3 deficiency or in mice withcardiac remodeling the inflamed heart affects also peripheral tissuesthereby worsening T2D in peripheral tissues resulting in elevated bloodglucose level and turning on a vicious cycle to worsen cardiomyopathy
Supplementary material
Supplementary material is available at Cardiovascular Research online
AcknowledgementsThe authors wish to thank Drs Keren Bitton-Worms and Ran Rostokerand Prof Derek LeRoith from the Rambam Medical Center for their
assistance in GTT and ITT Dr Edith Suss Toby Maya Holdengreber andLior Liba from the BCF Bioimaging Center (Faculty of MedicineTechnion) for their assistance with imaging and image analysis Prof DEMichelle from University of Michigan for cardiomyocytes culture techni-que and Prof Yehiel Zick from the Weizmann institute of Sciences forcritical comments
Conflict of interest none declared
FundingThis work was partially supported by the United States-Israel BinationalGrant 2009179 to AA and TH In addition we wish to thank the MarsPittsburgh Fund for Medical Research to AA the Department of Defensegrant W81XWH-14-1-0179 to TH and to D Dan and Betty Kahn travelgrant support to LK
References
1 Roger VL Go AS Lloyd-Jones DM Benjamin EJ Berry JD Borden WB Bravata DMDai S Ford ES Fox CS Fullerton HJ Gillespie C Hailpern SM Heit JA Howard VJKissela BM Kittner SJ Lackland DT Lichtman JH Lisabeth LD Makuc DM MarcusGM Marelli A Matchar DB Moy CS Mozaffarian D Mussolino ME Nichol GPaynter NP Soliman EZ Sorlie PD Sotoodehnia N Turan TN Virani SS Wong NDWoo D Turner MB American Heart Association Statistics C Stroke Statistics SHeart disease and stroke statisticsndash2012 update a report from the American HeartAssociation Circulation 2012125e2ndashe220
2 Wilson C Diabetes ACCORD 5-year outcomes of intensive glycemic control NatRev Endocrinol 20117314
3 Ernande L Derumeaux G Diabetic cardiomyopathy myth or reality Arch CardiovascDis 2012105218ndash225
4 Battiprolu PK Hojayev B Jiang N Wang ZV Luo X Iglewski M Shelton JM GerardRD Rothermel BA Gillette TG Lavandero S Hill JA Metabolic stress-induced
InsulinGF
Cytokines
FFA
GlucoseATF3
Cardiac
Remodeling
Factors
Paracrine action
Collagens
and ECM
proteins
Fibrosis
Cardiac
Dysfunction
Systemic Effects
Glucose Tolerance
TNFα
IL-6
Cardiomyocyte
ATF3
Homeostasis
Homeostasis
TNFα
IL-6
TIMP1
BNP
ANP
Figure 8 lsquoPeripheral-to-cardiacrsquo and lsquoCardiac-to-peripheralrsquo model scheme Schematic diagram showing the ATF3 adaptive role in maintaining hearthomeostasis following HFD-induced T2D In contrast stressful environment under HFD leads to maladaptive cardiac response in the absence of ATF3Cardiac remodeling caused by ATF3 deficiency induces an increase in cardiac ECM remodeling and inflammatory genes and mediates the communication ofcardiomyocytes with heart resident cells potentially mediated by IL-6 TNFa ANP BNP and TIMP1 In this situation increased cardiac fibrosis and inflamma-tory response worsen cardiac function The inflamed failing heart mediates the communication of cardiomyocytes with distant organs as well thereby affect-ing diabetic outcome and glucose homeostasis
ATF3 expression in cardiomyocytes preserves homeostasis 145D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
activation of FoxO1 triggers diabetic cardiomyopathy in mice J Clin Invest20121221109ndash1118
5 Mandavia CH Aroor AR Demarco VG Sowers JR Molecular and metabolic mecha-nisms of cardiac dysfunction in diabetes Life Sci 201392601ndash608
6 Ventura-Clapier R Garnier A Veksler V Energy metabolism in heart failure J Physiol20045551ndash13
7 Guo S Insulin signaling resistance and the metabolic syndrome insights from mousemodels into disease mechanisms J Endocrinol 2014220T1ndashT23
8 Montane J Cadavez L Novials A Stress and the inflammatory process a major causeof pancreatic cell death in type 2 diabetes Diabetes Metab Syndr Obes 2014725ndash34
9 Ashrafian H Frenneaux MP Opie LH Metabolic mechanisms in heart failureCirculation 2007116434ndash448
10 Tanabe K Liu Y Hasan SD Martinez SC Cras-Meneur C Welling CM Bernal-MizrachiE Tanizawa Y Rhodes CJ Zmuda E Hai T Abumrad NA Permutt MA Glucose andfatty acids synergize to promote B-cell apoptosis through activation of glycogen syn-thase kinase 3beta independent of JNK activation PLoS One 20116e18146
11 Heineke J Molkentin JD Regulation of cardiac hypertrophy by intracellular signallingpathways Nat Rev Mol Cell Biol 20067589ndash600
12 Hai T Wolford CC Chang YS ATF3 a hub of the cellular adaptive-response net-work in the pathogenesis of diseases is modulation of inflammation a unifying com-ponent Gene Expr 2010151ndash11
13 Takii R Inouye S Fujimoto M Nakamura T Shinkawa T Prakasam R Tan K HayashidaN Ichikawa H Hai T Nakai A Heat shock transcription factor 1 inhibits expression ofIL-6 through activating transcription factor 3 J Immunol 20101841041ndash1048
14 Giraldo A Barrett OP Tindall MJ Fuller SJ Amirak E Bhattacharya BS Sugden PHClerk A Feedback regulation by Atf3 in the endothelin-1-responsive transcriptomeof cardiomyocytes Egr1 is a principal Atf3 target Biochem J 2012444343ndash355
15 Hasin T Elhanani O Abassi Z Hai T Aronheim A Angiotensin II signaling up-regulates the immediate early transcription factor ATF3 in the left but not the rightatrium Basic Res Cardiol 2011106175ndash187
16 Koren L Elhanani O Kehat I Hai T Aronheim A Adult cardiac expression of theactivating transcription factor 3 ATF3 promotes ventricular hypertrophy PLoS One20138e68396
17 Koren L Alishekevitz D Elhanani O Nevelsky A Hai T Kehat I Shaked Y AronheimA ATF3-dependent cross-talk between cardiomyocytes and macrophages promotescardiac maladaptive remodeling Int J Cardiol 2015198232ndash240
18 Okamoto Y Chaves A Chen J Kelley R Jones K Weed HG Gardner KL Gangi LYamaguchi M Klomkleaw W Nakayama T Hamlin RL Carnes C Altschuld R BauerJ Hai T Transgenic mice with cardiac-specific expression of activating transcriptionfactor 3 a stress-inducible gene have conduction abnormalities and contractile dys-function Am J Pathol 2001159639ndash650
19 Zhou H Shen DF Bian ZY Zong J Deng W Zhang Y Guo YY Li H Tang QZActivating transcription factor 3 deficiency promotes cardiac hypertrophy dysfunc-tion and fibrosis induced by pressure overload PLoS One 20116e26744
20 Zhou H Guo H Zong J Dai J Yuan Y Bian ZY Tang QZ ATF3 regulates multipletargets and may play a dual role in cardiac hypertrophy and injury Int J Cardiol2014174838ndash839
21 Yang J Yang CJ Yang J Fan ZX ATF3 a promotion effect or a inhibition effect in car-diac maladaptive remodeling Int J Cardiol 2015201245ndash246
22 Koren L Shaked Y Aronheim A Response letter ldquoATF3 a promoter or inhibitor ofcardiac maladaptive remodelingrdquo Int J Cardiol 2015201692
23 Ma ZG Wei WY Xu SC Zhang WB Dai J Tang QZ ATF3 a potential target forcardiac maladaptive remodeling Int J Cardiol 201620250ndash51
24 Zhou H Yuan Y Ni J Guo H Deng W Bian ZY Tang QZ Pleiotropic and puzzlingeffects of ATF3 in maladaptive cardiac remodeling Int J Cardiol 201620687ndash88
25 Zmuda EJ Qi L Zhu MX Mirmira RG Montminy MR Hai T The roles of ATF3 anadaptive-response gene in high-fat-diet-induced diabetes and pancreatic beta-celldysfunction Mol Endocrinol 2010241423ndash1433
26 Lu D Chen J Hai T The regulation of ATF3 gene expression by mitogen-activatedprotein kinases Biochem J 2007401559ndash567
27 Hartman MG Lu D Kim ML Kociba GJ Shukri T Buteau J Wang X Frankel WLGuttridge D Prentki M Grey ST Ron D Hai T Role for activating transcription fac-tor 3 in stress-induced beta-cell apoptosis Mol Cell Biol 2004245721ndash5732
28 Wolford CC McConoughey SJ Jalgaonkar SP Leon M Merchant AS Dominick JLYin X Chang Y Zmuda EJ OrsquoToole SA Millar EK Roller SL Shapiro CL OstrowskiMC Sutherland RL Hai T Transcription factor ATF3 links host adaptive response tobreast cancer metastasis J Clin Iinvest 20131232893ndash2906
29 Agah R Frenkel PA French BA Michael LH Overbeek PA Schneider MD Generecombination in postmitotic cells Targeted expression of Cre recombinase pro-vokes cardiac-restricted site-specific rearrangement in adult ventricular musclein vivo J Clin Invest 1997100169ndash179
30 Livet J Weissman TA Kang H Draft RW Lu J Bennis RA Sanes JR Lichtman JWTransgenic strategies for combinatorial expression of fluorescent proteins in thenervous system Nature 200745056ndash62
31 Davis J Maillet M Miano JM Molkentin JD Lost in transgenesis a userrsquos guidefor genetically manipulating the mouse in cardiac research Circ Res2012111761ndash777
32 Heiberg E Sjogren J Ugander M Carlsson M Engblom H Arheden H Design andvalidation of segment ndash freely available software for cardiovascular image analysisBMC Med Imaging 2010101
33 Kabaeva Z Zhao M Michele DE Blebbistatin extends culture life of adult mouse car-diac myocytes and allows efficient and stable transgene expression Am J Physiol HeartC 2008294H1667ndash1674
34 Subramaniam A Jones WK Gulick J Wert S Neumann J Robbins J Tissue-specificregulation of the alpha-myosin heavy chain gene promoter in transgenic mice J BiolChem 199126624613ndash24620
35 Cunha DA Hekerman P Ladriere L Bazarra-Castro A Ortis F Wakeham MCMoore F Rasschaert J Cardozo AK Bellomo E Overbergh L Mathieu C Lupi R HaiT Herchuelz A Marchetti P Rutter GA Eizirik DL Cnop M Initiation and executionof lipotoxic ER stress in pancreatic beta-cells J Cell Sci 20081212308ndash2318
36 Wynn TA Ramalingam TR Mechanisms of fibrosis therapeutic translation forfibrotic disease Nat Med 2012181028ndash1040
37 Li D Yin X Zmuda EJ Wolford CC Dong X White MF Hai T The repression ofIRS2 gene by ATF3 a stress-inducible gene contributes to pancreatic beta-cell apop-tosis Diabetes 200857635ndash644
38 Myerson SG Bellenger NG Pennell DJ Assessment of left ventricular mass by cardi-ovascular magnetic resonance Hypertension 200239750ndash755
39 Zmuda EJ Viapiano M Grey ST Hadley G Garcia-Ocana A Hai T Deficiency ofAtf3 an adaptive-response gene protects islets and ameliorates inflammation in asyngeneic mouse transplantation model Diabetologia 2010531438ndash1450
40 Riehle C Abel ED Insulin signaling and heart failure Circ Res 20161181151ndash116941 Schreyer SA Wilson DL LeBoeuf RC C57BL6 mice fed high fat diets as models for
diabetes-accelerated atherosclerosis Atherosclerosis 199813617ndash2442 Brainard RE Watson LJ Demartino AM Brittian KR Readnower RD Boakye AA
Zhang D Hoetker JD Bhatnagar A Baba SP Jones SP High fat feeding in mice isinsufficient to induce cardiac dysfunction and does not exacerbate heart failure PLoSOne 20138e83174
43 Gupte AA Minze LJ Reyes M Ren Y Wang X Brunner G Ghosn M Cordero-Reyes AM Ding K Pratico D Morrisett J Shi ZZ Hamilton DJ Lyon CJ Hsueh WAHigh-fat feeding-induced hyperinsulinemia increases cardiac glucose uptake and mito-chondrial function despite peripheral insulin resistance Endocrinology20131542650ndash2662
44 Logan SK Garabedian MJ Campbell CE Werb Z Synergistic transcriptionalactivation of the tissue inhibitor of metalloproteinases-1 promoter via functionalinteraction of AP-1 and Ets-1 transcription factors J Biol Chem1996271774ndash782
45 Belke DD Betuing S Tuttle MJ Graveleau C Young ME Pham M Zhang DCooksey RC McClain DA Litwin SE Taegtmeyer H Severson D Kahn CR AbelED Insulin signaling coordinately regulates cardiac size metabolism and contractileprotein isoform expression J Clin Invest 2002109629ndash639
46 Fernandez AM Kim JK Yakar S Dupont J Hernandez-Sanchez C Castle AL FilmoreJ Shulman GI Le Roith D Functional inactivation of the IGF-I and insulin receptors inskeletal muscle causes type 2 diabetes Genes Dev 2001151926ndash1934
47 Gilchrist M Thorsson V Li B Rust AG Korb M Roach JC Kennedy K Hai TBolouri H Aderem A Systems biology approaches identify ATF3 as a negative regu-lator of Toll-like receptor 4 Nature 2006441173ndash178
48 Kristiansen OP Mandrup-Poulsen T Interleukin-6 and diabetes the good the bad orthe indifferent Diabetes 200554(Suppl 2)S114ndashS124
49 Grueter CE van Rooij E Johnson BA DeLeon SM Sutherland LB Qi X Gautron LElmquist JK Bassel-Duby R Olson EN A cardiac microRNA governs systemic energyhomeostasis by regulation of MED13 Cell 2012149671ndash683
50 Baskin KK Grueter CE Kusminski CM Holland WL Bookout AL Satapati S KongYM Burgess SC Malloy CR Scherer PE Newgard CB Bassel-Duby R Olson ENMED13-dependent signaling from the heart confers leanness by enhancing metabo-lism in adipose tissue and liver EMBO Mol Med 201461610ndash1621
51 Nakamura M Sadoshima J Heart over mind metabolic control of white adipose tis-sue and liver EMBO Mol Med 201461521ndash1524
52 Tenenbaum A Fisman EZ Impaired glucose metabolism in patients with heart failurepathophysiology and possible treatment strategies Am J Cardiovasc Drugs20044269ndash280
53 Suskin N McKelvie RS Burns RJ Latini R Pericak D Probstfield J Rouleau JL SigouinC Solymoss CB Tsuyuki R White M Yusuf S Glucose and insulin abnormalitiesrelate to functional capacity in patients with congestive heart failure Eur Heart J2000211368ndash1375
54 Aronson D Hammerman H Kapeliovich MR Suleiman A Agmon Y Beyar RMarkiewicz W Suleiman M Fasting glucose in acute myocardial infarction incremen-tal value for long-term mortality and relationship with left ventricular systolic func-tion Diabetes Care 200730960ndash966
146 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
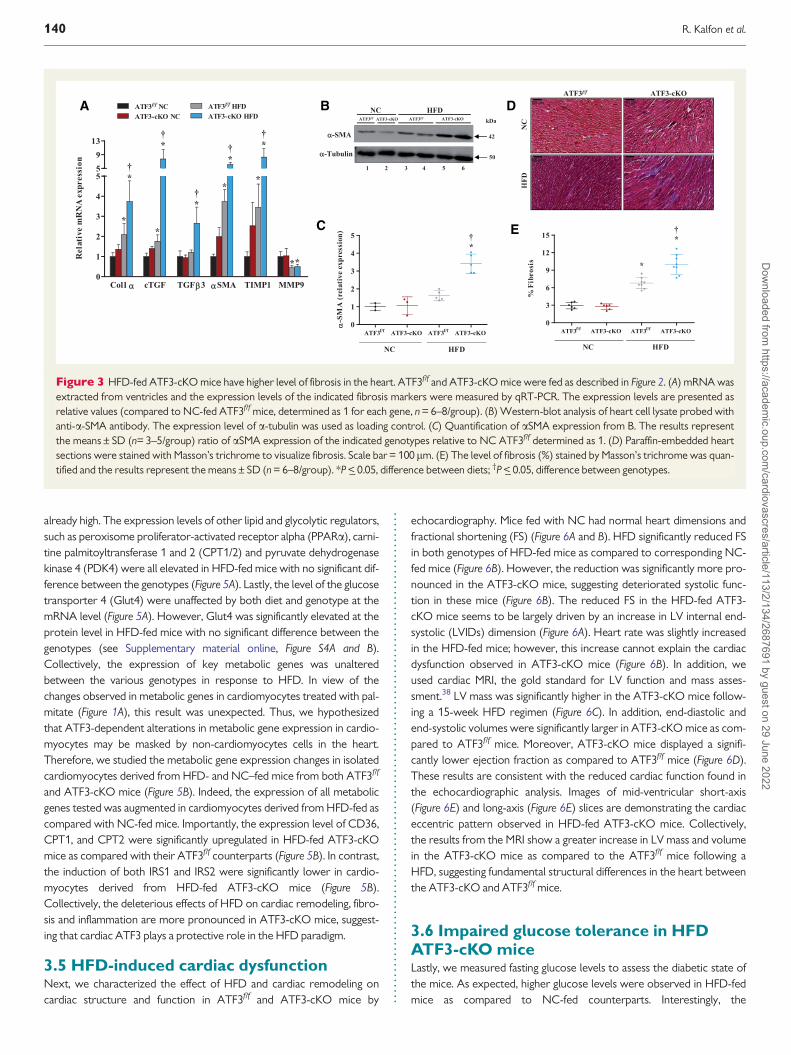
already high The expression levels of other lipid and glycolytic regulatorssuch as peroxisome proliferator-activated receptor alpha (PPARa) carni-tine palmitoyltransferase 1 and 2 (CPT12) and pyruvate dehydrogenasekinase 4 (PDK4) were all elevated in HFD-fed mice with no significant dif-ference between the genotypes (Figure 5A) Lastly the level of the glucosetransporter 4 (Glut4) were unaffected by both diet and genotype at themRNA level (Figure 5A) However Glut4 was significantly elevated at theprotein level in HFD-fed mice with no significant difference between thegenotypes (see Supplementary material online Figure S4A and B)Collectively the expression of key metabolic genes was unalteredbetween the various genotypes in response to HFD In view of thechanges observed in metabolic genes in cardiomyocytes treated with pal-mitate (Figure 1A) this result was unexpected Thus we hypothesizedthat ATF3-dependent alterations in metabolic gene expression in cardio-myocytes may be masked by non-cardiomyocytes cells in the heartTherefore we studied the metabolic gene expression changes in isolatedcardiomyocytes derived from HFD- and NCndashfed mice from both ATF3ff
and ATF3-cKO mice (Figure 5B) Indeed the expression of all metabolicgenes tested was augmented in cardiomyocytes derived from HFD-fed ascompared with NC-fed mice Importantly the expression level of CD36CPT1 and CPT2 were significantly upregulated in HFD-fed ATF3-cKOmice as compared with their ATF3ff counterparts (Figure 5B) In contrastthe induction of both IRS1 and IRS2 were significantly lower in cardio-myocytes derived from HFD-fed ATF3-cKO mice (Figure 5B)Collectively the deleterious effects of HFD on cardiac remodeling fibro-sis and inflammation are more pronounced in ATF3-cKO mice suggest-ing that cardiac ATF3 plays a protective role in the HFD paradigm
35 HFD-induced cardiac dysfunctionNext we characterized the effect of HFD and cardiac remodeling oncardiac structure and function in ATF3ff and ATF3-cKO mice by
echocardiography Mice fed with NC had normal heart dimensions andfractional shortening (FS) (Figure 6A and B) HFD significantly reduced FSin both genotypes of HFD-fed mice as compared to corresponding NC-fed mice (Figure 6B) However the reduction was significantly more pro-nounced in the ATF3-cKO mice suggesting deteriorated systolic func-tion in these mice (Figure 6B) The reduced FS in the HFD-fed ATF3-cKO mice seems to be largely driven by an increase in LV internal end-systolic (LVIDs) dimension (Figure 6A) Heart rate was slightly increasedin the HFD-fed mice however this increase cannot explain the cardiacdysfunction observed in ATF3-cKO mice (Figure 6B) In addition weused cardiac MRI the gold standard for LV function and mass asses-sment38 LV mass was significantly higher in the ATF3-cKO mice follow-ing a 15-week HFD regimen (Figure 6C) In addition end-diastolic andend-systolic volumes were significantly larger in ATF3-cKO mice as com-pared to ATF3ff mice Moreover ATF3-cKO mice displayed a signifi-cantly lower ejection fraction as compared to ATF3ff mice (Figure 6D)These results are consistent with the reduced cardiac function found inthe echocardiographic analysis Images of mid-ventricular short-axis(Figure 6E) and long-axis (Figure 6E) slices are demonstrating the cardiaceccentric pattern observed in HFD-fed ATF3-cKO mice Collectivelythe results from the MRI show a greater increase in LV mass and volumein the ATF3-cKO mice as compared to the ATF3ff mice following aHFD suggesting fundamental structural differences in the heart betweenthe ATF3-cKO and ATF3ff mice
36 Impaired glucose tolerance in HFDATF3-cKO miceLastly we measured fasting glucose levels to assess the diabetic state ofthe mice As expected higher glucose levels were observed in HFD-fedmice as compared to NC-fed counterparts Interestingly the
A B
C
D
E
Figure 3 HFD-fed ATF3-cKO mice have higher level of fibrosis in the heart ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) mRNA wasextracted from ventricles and the expression levels of the indicated fibrosis markers were measured by qRT-PCR The expression levels are presented asrelative values (compared to NC-fed ATF3ff mice determined as 1 for each gene n = 6ndash8group) (B) Western-blot analysis of heart cell lysate probed withanti-a-SMA antibody The expression level of a-tubulin was used as loading control (C) Quantification of aSMA expression from B The results representthe meansplusmn SD (n= 3ndash5group) ratio of aSMA expression of the indicated genotypes relative to NC ATF3ff determined as 1 (D) Paraffin-embedded heartsections were stained with Massonrsquos trichrome to visualize fibrosis Scale bar = 100 lm (E) The level of fibrosis () stained by Massonrsquos trichrome was quan-tified and the results represent the meansplusmn SD (n = 6ndash8group) P lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes
140 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
hyperglycemia state of the ATF3-cKO mice was significantly higher thanin the ATF3ff mice following 15 weeks of HFD (Figure 7A) Consistentlythe glucose tolerance test (GTT) revealed that HFD-fed ATF3-cKOmice displayed higher blood glucose levels and longer glucose clearancetime when compared to HFD-fed ATF3ff whereas NC-fed ATF3ff andATF3-cKO mice had normal glucose clearance (Figure 7B)
Interestingly whole body ATF3-KO mice fed with HFD also displayedimpaired glucose clearance (see Supplementary material onlineFigure S5A)39 Previously we reported that decreased glucose tolerancein whole body ATF3-KO mice is at least in part due to reduced insulingene expression in b-cells (see Supplementary material online FigureS5B)39 To determine insulin level in HFD-fed ATF3-cKO mice weanalysed blood insulin levels by ELISA In both HFD-fed ATF3ff andATF3-cKO mice the serum insulin levels were significantly higher ascompared to their NC-fed counterpart mice with the ATF3-cKO micedisplaying the highest insulin levels (Figure 7C) Together these resultssuggest that peripheral tissues of ATF3-cKO mice have reduced ability
to uptake glucose despite their high insulin level in the blood Next wesought to measure the protein levels of components along the insulinpathway Akt and Foxo340 Both Akt and Foxo3 protein levels were sim-ilar in HFD-fed mice independent of the genotype (see Supplementarymaterial online Figure S4A and C) Interestingly phosphorylation of Akt(Ser473) which indicates the activity of the insulin pathway was slightlyelevated in HFD-fed mice however this did not reach statistical signifi-cance and was similar in both of the genotypes (see Supplementary material online Figure S4D) To measure the activity of the insulin signallingpathway following insulin stimulation we used the ratio between phos-phorylated Akt (Ser473) and total Akt expression HFD-fed mice wereinjected with insulin (10 Ukg) and the heart liver and skeletal musclewere harvested 15 min after the injection (Figure 7D and E and seeSupplementary material online Figure S6AndashD) Insulin injection resultedin a significant increase of pAktAkt ratio in the hearts of ATF3ff micewhile no significant change was observed in Akt phosphorylation inATF3-cKO mice (Figure 7D and E) Similarly low Akt phosphorylation
A
D E
B C
Figure 4 HFD-fed ATF3-cKO mice display inflammatory response in the heart (A) ATF3ff and ATF3-cKO mice were fed as described in Figure 2 mRNAwas extracted from ventricles and the expression levels of the indicated inflammatory markers were measured by qRT-PCR Expression levels are presentedas relative values (compared to NC-fed ATF3ff mice determined as 1 for each gene n= 6ndash8group) (B) Mice were fed as described in Figure 2Cardiomyocytes were isolated and primary cultures were generated mRNA was extracted from an overnight culture and the expression levels of IL-6 andTNFa were studied by qRT-PCR The mRNA level obtained in cardiomyocytes derived from NC-fed ATF3ff mice was determined as 1 and the relativeexpression of all other treatment is shown (n = 5ndash6 independent cardiomyocyte preparations) (C) Conditioned media was collected from cardiomyocytesdescribed in B and ELISA assay for the indicated cytokines is shown Cytokines level is presented as relative values to NC-fed ATF3ff mice determined as 1for each cytokine (n = 5ndash6 independent preparations) (D and E) Blood was collected from HFD-fed mice and IL-6 and TNFa serum levels were quantifiedby ELISA (n = 8ndash12group) The results represent the meansplusmn SD P lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes
ATF3 expression in cardiomyocytes preserves homeostasis 141D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
levels were also observed in both the liver and skeletal muscle of theATF3-cKO mice however this did not reach statistically significance (seeSupplementary material online Figure S6AndashD) Consistently palmitate-treated cardiomyocytes derived from ATF3-cKO mice displayedreduced phosphorylation of Akt following 30-min insulin stimulation(see Supplementary material online Figure S6E)
Collectively our findings revealed a surprising phenomenon that car-diac ATF3 deficiency results in increased cardiac remodeling leading topotentiation of local and peripheral insulin resistance
4 Discussion
A HFD is a commonly used regimen to induce T2D The majority of theliterature focuses on diabetic symptoms without addressing the issue ofT2D-induced cardiomyopathy Even when it is addressed there is noclear consensus regarding the extent of cardiac effects imposed byHFD41 Diet-induced obesity in mouse models is associated with globalinsulin resistance and increased myocardial fatty acid utilization how-ever the reported phenotypes on cardiac function vary from extensivehypertrophy and heart failure4 to lack of cardiac dysfunction4243 Thesevariations have been attributed to diet regimens andor sensitivity ofmouse strains to a HFD In this report we present evidence that cardiacATF3 deficiency exacerbates the deleterious effect of a HFD on theheart such as maladaptive cardiac remodeling fibrosis inflammation andcardiac dysfunction Our work may provide a potential molecular insightto variations in the literature regarding the impact of HFD on the heartAs ATF3 is a stress-inducible gene we propose that its proper inductionin the cardiomyocytes is protective and helps the heart to cope withHFD stress It is possible that the induction of ATF3mdashits expressionlevel duration and kineticsmdashvaries in different HFD regimens andormouse strains contributing to the variations observed in the literature
HFD regimen induced a significant higher level of fibrosis in the heartof ATF3-cKO mice We have identified TIMP1 as a possible mechanismto explain the increase in fibrosis in ATF3-cKO cardiomyocytes Severallines of evidence suggest that TIMP1 is possibly an ATF3 target gene
First we observed an increase in TIMP1 transcription in the whole heartand in isolated cardiomyocytes (Figures 1A and 3A) Second TIMP 1 3and 4 were identified as significantly highly expressed genes in the heartof ATF3-KO mice as compared with wild type mice following aorticbanding20 Third a functional AP-1 regulatory element within the TIMP1promoter was described44 Indeed preliminary results in our lab suggestthat ATF3 downregulates the TIMP1 promoter (data not shown) Thusit is possible that TIMP1 is suppressed by ATF3 and therefore results in anet increase in MMPs activity that helps to maintain ECM formationdeg-radation homeostasis
Our findings are consistent with previous studies demonstrating that adeficiency in ATF3 can promote cardiac maladaptive changes which leadto cardiac dilatation and hypertrophy in an aortic banding model1920 Incontrast a different study using a pressure overload model reportedthat ATF3 induces a maladaptive cardiac response17 These seeminglyconflicting data may be explained by the different patterns of ATF3expression induced by the different models In the pressure overloadmodel the acute and transient expression of ATF3 in cardiomyocytesinitiates a cascade of maladaptive changes17 whereas in the aortic band-ing model and the HFD model described here ATF3 expression is longlasting
In this context this is the first report using mice with cardiac-selectivedeficiency of ATF3 (ATF3-cKO) Previous reports on the roles of ATF3in cardiac function were carried out using whole body knockout ofATF325 or transgenic mice ectopically expressing ATF3 in cardiomyo-cytes16 A key advancement of ATF3-cKO mice over the whole bodyknockout mice is that they provide insights into the function of ATF3selectively in cardiomyocytes without the complication of ATF3 defi-ciency in other cells Thus the ATF3-cKO mouse is a useful tool to bet-ter understand the function of cardiac ATF3
Consistent with our previous report25 here we showed that cardiacATF3 deficiency has a more severe HFD-induced diabetic phenotypethan the control wild type mice However while reduced insulin genetranscription in ATF3-deficient b-cells was observed25 our current dataindicates that a lack of insulin is not the explanation for the glucose intol-erance found in the HFD-fed ATF3-cKO mice In contrast these mice
A B
Figure 5 HFD-fed ATF3-cKO mice show metabolic response in the heart ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) mRNA wasextracted from ventricles and the expression levels of the indicated metabolic markers were measured by qRT-PCR Expression levels are presented as rela-tive values (compared to NC-fed ATF3ff mice determined as 1 for each gene n= 6ndash8group) (B) Cardiomyocytes were isolated and plated on laminin for1 h followed by mRNA preparation (n = 4ndash5 independent preparations) The expression levels of the indicated metabolic markers were measured by qRT-PCR Expression levels are presented as relative values (to NC-fed ATF3ff mice determined as 1 for each gene) The results represent the meansplusmn SDP lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes
142 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
had higher blood insulin level (Figure 7C) Therefore upon HFD chal-lenge cardiac-deficiency of ATF3 exacerbates HFD-induced T2D Thislsquoperipheral effectrsquo is surprising since ATF3 is selectively deleted in thecardiomyocytes
The question then is how can we explain the effect of cardiacATF3 deficiency on peripheral glucose intolerance One possibility isthe reduced IRS1 expression in ATF3-deficient cardiomyocytes bothin vivo and in vitro IRS1 suppression may lead to impaired insulin sig-nalling in the heart thereby exacerbating glucose intolerance Indeedwe observed that HFD-fed ATF3-cKO mice displayed insulin resist-ance upon insulin challenge when compared to HFD-fed ATF3ff
mice In addition palmitate-treated cardiomyocytes lacking ATF3 dis-played lower insulin signalling as compared to their wild type coun-terparts Since the heart is only a minor glucose consuming organ incomparison to skeletal muscle cells or the brain the peripheral sys-temic effect cannot be solely based on this observation A case inpoint transgenic mice with insulin receptor (IR) deficiency in theheart do not display a diabetic phenotype4546 Another attractive
possibility for the peripheral effect of cardiac ATF3-deletion is itsimpact on inflammation It is well established that low-grade inflam-mation years in advance of the disease onset is involved in the patho-genic processes causing T2D8 Mediators of inflammation such asTNFa interleukin (IL)-1b the IL-6 family of cytokines IL-18 and cer-tain chemokines have been proposed to be involved in the eventscausing both forms of diabetes (T1D and T2D)8 Interestingly wehave shown that IL-6 a well-known ATF3 target gene47 is known toenhance its transcription and secretion of cytokines such as IL-6 andTNFa in palmitate-treated primary cultures of ATF3-cKO cardio-myocytes as well as in cardiomyocytes derived from HFD-fed ATF3-cKO mice IL-6 has been proposed to play a role in glucose homeo-stasis and metabolism directly and indirectly via its action on skeletalmuscle cells adipocytes hepatocytes pancreatic b-cells and neuro-endocrine cells48
Therefore we hypothesize that elevated levels of inflammatory cyto-kines in ATF3-cKO mice mediate a cross-talk in peripheral tissues leadingto impaired glucose tolerance Although circulating IL-6 and TNFa
A
B
C
D
E
F
Figure 6 HFD-fed ATF3-cKO mice present deteriorated heart function ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) At endpointmice hearts were examined by micro ultrasound The following parameters were measured interventricular septal end diastole (IVSd) left ventricular pos-terior wall end diastole (LVPWd) maximal left ventricular internal end-diastole (LVIDd) end-systole (LVIDs) and heart rate The results represent themeansplusmn SD (n = 6ndash8 in each group as shown in panel B) (B) Heart function was assessed by calculating percentage of fractional shortening (FS) accordingto FS () = [(LVDd-LVDs)LVDd]100 The results represent the meansplusmn SD (n = 6ndash8 in each group) (C and D) Left ventricular cardiac volumes massand function were examined by a cardiac MRI The following parameters were measured left ventricular end-diastolic (LVEDV) and left ventricular end-sys-tolic volume (LVESV) left ventricular (LV) mass and ejection fraction (EF) The results represent the meansplusmn SD (n = 7 in each group) (E and F)Representative images of mid-ventricular short-axis (E) and long-axis (F) slice at peak diastole and systole P lt_ 005 difference between diets daggerP lt_ 005 dif-ference between genotypes
ATF3 expression in cardiomyocytes preserves homeostasis 143D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
cytokines may be derived from different tissues we provide ample evi-dence that ATF3-defficient cardiomyocytes produce and secrete higherlevels of these cytokines as compared to their wild type counterpartsThis indicates a lsquocardiac-to-peripheralrsquo effect in T2D-induced cardiomyop-athy This is a reverse direction from the lsquoperipheral-to-cardiacrsquo effect dis-cussed in the introduction where inflammation in the peripheral tissuesleads to cardiac dysfunction Since inflammatory cytokines are soluble
factors they can exert a systemic effect This is why it makes sense thattheir effect can go both ways Thus diabetes and cardiomyopathy are apart of a vicious cycle Previously it was shown that cardiac specific dele-tion of the thyroid hormone complex mediator MED13 results in aperipheral effect upon HFD Cardiac MED13 deficiency suppresses meta-bolic genes in the heart which leads to a decrease in energy expenditureand an obese phenotype most likely by the secretion of cardiokines that
A
B
C
D
E
Figure 7 HFD-fed ATF3-cKO mice exhibit impaired glucose tolerance ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) Fasting (6 h)blood glucose concentrations (mgdl) The results represent the meansplusmn SD (n = 20ndash30group) P lt_ 005 difference between diets daggerP lt_ 005 differencebetween genotypes (B) Glucose tolerance test (GTT) Values are presented as relative to initial valueplusmn SD (n = 20ndash25group) P lt_ 005 difference betweendiets daggerP lt_ 005 difference between genotypes (C) Serum insulin concentrations were quantified by ELISA Values are presented as relative values (com-pared to NC-fed ATF3ff mice determined as 1 n = 15ndash20group) P lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes (D and E)Effect of HFD on phosphorylated Akt (Ser473 pAkt) to Akt ratio levels in heart lysate following either saline or insulin injection (10 U1 kg) is shownRepresentative blots (D) Densitometric analysis is presented as means ratio (n = 3ndash6group) pAktAkt ratio is presented as relative values (compared toHFD-fed saline injected ATF3ff mice determined as 1) P lt_ 005 insulin vs saline treatment daggerP lt_ 005 difference between genotypes (E)
144 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
mediates the cross talk between the heart and peripheral tissues49ndash51
Here we also observed increased levels of natriuretic peptides (ANP andBNP) which together with IL-6 and TNFa may mediate the cross-talk withthe peripheral tissues to induce glucose intolerance
Does this finding have any relevance to cardiac disease in humansSeveral lines of evidence suggests that congestive heart failure in patientsis associated with an increase in blood glucose levels52 In addition highblood glucose levels in non-diabetic patients with acute myocardialinfarction serves as a prognostic predictor of long-term mortality5354
Thus here we provide evidence that high blood glucose levels are a partof the symptoms of cardiac remodeling process
Collectively our findings suggest that cardiac ATF3 expression isresponsible for an adaptive cardiac response to the stressful environmentunder a HFD (Figure 8) ATF3 dampens the inflammatory response andsuppresses the expression of ECM remodeling genes and thus amelioratescardiac function In HFD-fed mice with ATF3 deficiency or in mice withcardiac remodeling the inflamed heart affects also peripheral tissuesthereby worsening T2D in peripheral tissues resulting in elevated bloodglucose level and turning on a vicious cycle to worsen cardiomyopathy
Supplementary material
Supplementary material is available at Cardiovascular Research online
AcknowledgementsThe authors wish to thank Drs Keren Bitton-Worms and Ran Rostokerand Prof Derek LeRoith from the Rambam Medical Center for their
assistance in GTT and ITT Dr Edith Suss Toby Maya Holdengreber andLior Liba from the BCF Bioimaging Center (Faculty of MedicineTechnion) for their assistance with imaging and image analysis Prof DEMichelle from University of Michigan for cardiomyocytes culture techni-que and Prof Yehiel Zick from the Weizmann institute of Sciences forcritical comments
Conflict of interest none declared
FundingThis work was partially supported by the United States-Israel BinationalGrant 2009179 to AA and TH In addition we wish to thank the MarsPittsburgh Fund for Medical Research to AA the Department of Defensegrant W81XWH-14-1-0179 to TH and to D Dan and Betty Kahn travelgrant support to LK
References
1 Roger VL Go AS Lloyd-Jones DM Benjamin EJ Berry JD Borden WB Bravata DMDai S Ford ES Fox CS Fullerton HJ Gillespie C Hailpern SM Heit JA Howard VJKissela BM Kittner SJ Lackland DT Lichtman JH Lisabeth LD Makuc DM MarcusGM Marelli A Matchar DB Moy CS Mozaffarian D Mussolino ME Nichol GPaynter NP Soliman EZ Sorlie PD Sotoodehnia N Turan TN Virani SS Wong NDWoo D Turner MB American Heart Association Statistics C Stroke Statistics SHeart disease and stroke statisticsndash2012 update a report from the American HeartAssociation Circulation 2012125e2ndashe220
2 Wilson C Diabetes ACCORD 5-year outcomes of intensive glycemic control NatRev Endocrinol 20117314
3 Ernande L Derumeaux G Diabetic cardiomyopathy myth or reality Arch CardiovascDis 2012105218ndash225
4 Battiprolu PK Hojayev B Jiang N Wang ZV Luo X Iglewski M Shelton JM GerardRD Rothermel BA Gillette TG Lavandero S Hill JA Metabolic stress-induced
InsulinGF
Cytokines
FFA
GlucoseATF3
Cardiac
Remodeling
Factors
Paracrine action
Collagens
and ECM
proteins
Fibrosis
Cardiac
Dysfunction
Systemic Effects
Glucose Tolerance
TNFα
IL-6
Cardiomyocyte
ATF3
Homeostasis
Homeostasis
TNFα
IL-6
TIMP1
BNP
ANP
Figure 8 lsquoPeripheral-to-cardiacrsquo and lsquoCardiac-to-peripheralrsquo model scheme Schematic diagram showing the ATF3 adaptive role in maintaining hearthomeostasis following HFD-induced T2D In contrast stressful environment under HFD leads to maladaptive cardiac response in the absence of ATF3Cardiac remodeling caused by ATF3 deficiency induces an increase in cardiac ECM remodeling and inflammatory genes and mediates the communication ofcardiomyocytes with heart resident cells potentially mediated by IL-6 TNFa ANP BNP and TIMP1 In this situation increased cardiac fibrosis and inflamma-tory response worsen cardiac function The inflamed failing heart mediates the communication of cardiomyocytes with distant organs as well thereby affect-ing diabetic outcome and glucose homeostasis
ATF3 expression in cardiomyocytes preserves homeostasis 145D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
activation of FoxO1 triggers diabetic cardiomyopathy in mice J Clin Invest20121221109ndash1118
5 Mandavia CH Aroor AR Demarco VG Sowers JR Molecular and metabolic mecha-nisms of cardiac dysfunction in diabetes Life Sci 201392601ndash608
6 Ventura-Clapier R Garnier A Veksler V Energy metabolism in heart failure J Physiol20045551ndash13
7 Guo S Insulin signaling resistance and the metabolic syndrome insights from mousemodels into disease mechanisms J Endocrinol 2014220T1ndashT23
8 Montane J Cadavez L Novials A Stress and the inflammatory process a major causeof pancreatic cell death in type 2 diabetes Diabetes Metab Syndr Obes 2014725ndash34
9 Ashrafian H Frenneaux MP Opie LH Metabolic mechanisms in heart failureCirculation 2007116434ndash448
10 Tanabe K Liu Y Hasan SD Martinez SC Cras-Meneur C Welling CM Bernal-MizrachiE Tanizawa Y Rhodes CJ Zmuda E Hai T Abumrad NA Permutt MA Glucose andfatty acids synergize to promote B-cell apoptosis through activation of glycogen syn-thase kinase 3beta independent of JNK activation PLoS One 20116e18146
11 Heineke J Molkentin JD Regulation of cardiac hypertrophy by intracellular signallingpathways Nat Rev Mol Cell Biol 20067589ndash600
12 Hai T Wolford CC Chang YS ATF3 a hub of the cellular adaptive-response net-work in the pathogenesis of diseases is modulation of inflammation a unifying com-ponent Gene Expr 2010151ndash11
13 Takii R Inouye S Fujimoto M Nakamura T Shinkawa T Prakasam R Tan K HayashidaN Ichikawa H Hai T Nakai A Heat shock transcription factor 1 inhibits expression ofIL-6 through activating transcription factor 3 J Immunol 20101841041ndash1048
14 Giraldo A Barrett OP Tindall MJ Fuller SJ Amirak E Bhattacharya BS Sugden PHClerk A Feedback regulation by Atf3 in the endothelin-1-responsive transcriptomeof cardiomyocytes Egr1 is a principal Atf3 target Biochem J 2012444343ndash355
15 Hasin T Elhanani O Abassi Z Hai T Aronheim A Angiotensin II signaling up-regulates the immediate early transcription factor ATF3 in the left but not the rightatrium Basic Res Cardiol 2011106175ndash187
16 Koren L Elhanani O Kehat I Hai T Aronheim A Adult cardiac expression of theactivating transcription factor 3 ATF3 promotes ventricular hypertrophy PLoS One20138e68396
17 Koren L Alishekevitz D Elhanani O Nevelsky A Hai T Kehat I Shaked Y AronheimA ATF3-dependent cross-talk between cardiomyocytes and macrophages promotescardiac maladaptive remodeling Int J Cardiol 2015198232ndash240
18 Okamoto Y Chaves A Chen J Kelley R Jones K Weed HG Gardner KL Gangi LYamaguchi M Klomkleaw W Nakayama T Hamlin RL Carnes C Altschuld R BauerJ Hai T Transgenic mice with cardiac-specific expression of activating transcriptionfactor 3 a stress-inducible gene have conduction abnormalities and contractile dys-function Am J Pathol 2001159639ndash650
19 Zhou H Shen DF Bian ZY Zong J Deng W Zhang Y Guo YY Li H Tang QZActivating transcription factor 3 deficiency promotes cardiac hypertrophy dysfunc-tion and fibrosis induced by pressure overload PLoS One 20116e26744
20 Zhou H Guo H Zong J Dai J Yuan Y Bian ZY Tang QZ ATF3 regulates multipletargets and may play a dual role in cardiac hypertrophy and injury Int J Cardiol2014174838ndash839
21 Yang J Yang CJ Yang J Fan ZX ATF3 a promotion effect or a inhibition effect in car-diac maladaptive remodeling Int J Cardiol 2015201245ndash246
22 Koren L Shaked Y Aronheim A Response letter ldquoATF3 a promoter or inhibitor ofcardiac maladaptive remodelingrdquo Int J Cardiol 2015201692
23 Ma ZG Wei WY Xu SC Zhang WB Dai J Tang QZ ATF3 a potential target forcardiac maladaptive remodeling Int J Cardiol 201620250ndash51
24 Zhou H Yuan Y Ni J Guo H Deng W Bian ZY Tang QZ Pleiotropic and puzzlingeffects of ATF3 in maladaptive cardiac remodeling Int J Cardiol 201620687ndash88
25 Zmuda EJ Qi L Zhu MX Mirmira RG Montminy MR Hai T The roles of ATF3 anadaptive-response gene in high-fat-diet-induced diabetes and pancreatic beta-celldysfunction Mol Endocrinol 2010241423ndash1433
26 Lu D Chen J Hai T The regulation of ATF3 gene expression by mitogen-activatedprotein kinases Biochem J 2007401559ndash567
27 Hartman MG Lu D Kim ML Kociba GJ Shukri T Buteau J Wang X Frankel WLGuttridge D Prentki M Grey ST Ron D Hai T Role for activating transcription fac-tor 3 in stress-induced beta-cell apoptosis Mol Cell Biol 2004245721ndash5732
28 Wolford CC McConoughey SJ Jalgaonkar SP Leon M Merchant AS Dominick JLYin X Chang Y Zmuda EJ OrsquoToole SA Millar EK Roller SL Shapiro CL OstrowskiMC Sutherland RL Hai T Transcription factor ATF3 links host adaptive response tobreast cancer metastasis J Clin Iinvest 20131232893ndash2906
29 Agah R Frenkel PA French BA Michael LH Overbeek PA Schneider MD Generecombination in postmitotic cells Targeted expression of Cre recombinase pro-vokes cardiac-restricted site-specific rearrangement in adult ventricular musclein vivo J Clin Invest 1997100169ndash179
30 Livet J Weissman TA Kang H Draft RW Lu J Bennis RA Sanes JR Lichtman JWTransgenic strategies for combinatorial expression of fluorescent proteins in thenervous system Nature 200745056ndash62
31 Davis J Maillet M Miano JM Molkentin JD Lost in transgenesis a userrsquos guidefor genetically manipulating the mouse in cardiac research Circ Res2012111761ndash777
32 Heiberg E Sjogren J Ugander M Carlsson M Engblom H Arheden H Design andvalidation of segment ndash freely available software for cardiovascular image analysisBMC Med Imaging 2010101
33 Kabaeva Z Zhao M Michele DE Blebbistatin extends culture life of adult mouse car-diac myocytes and allows efficient and stable transgene expression Am J Physiol HeartC 2008294H1667ndash1674
34 Subramaniam A Jones WK Gulick J Wert S Neumann J Robbins J Tissue-specificregulation of the alpha-myosin heavy chain gene promoter in transgenic mice J BiolChem 199126624613ndash24620
35 Cunha DA Hekerman P Ladriere L Bazarra-Castro A Ortis F Wakeham MCMoore F Rasschaert J Cardozo AK Bellomo E Overbergh L Mathieu C Lupi R HaiT Herchuelz A Marchetti P Rutter GA Eizirik DL Cnop M Initiation and executionof lipotoxic ER stress in pancreatic beta-cells J Cell Sci 20081212308ndash2318
36 Wynn TA Ramalingam TR Mechanisms of fibrosis therapeutic translation forfibrotic disease Nat Med 2012181028ndash1040
37 Li D Yin X Zmuda EJ Wolford CC Dong X White MF Hai T The repression ofIRS2 gene by ATF3 a stress-inducible gene contributes to pancreatic beta-cell apop-tosis Diabetes 200857635ndash644
38 Myerson SG Bellenger NG Pennell DJ Assessment of left ventricular mass by cardi-ovascular magnetic resonance Hypertension 200239750ndash755
39 Zmuda EJ Viapiano M Grey ST Hadley G Garcia-Ocana A Hai T Deficiency ofAtf3 an adaptive-response gene protects islets and ameliorates inflammation in asyngeneic mouse transplantation model Diabetologia 2010531438ndash1450
40 Riehle C Abel ED Insulin signaling and heart failure Circ Res 20161181151ndash116941 Schreyer SA Wilson DL LeBoeuf RC C57BL6 mice fed high fat diets as models for
diabetes-accelerated atherosclerosis Atherosclerosis 199813617ndash2442 Brainard RE Watson LJ Demartino AM Brittian KR Readnower RD Boakye AA
Zhang D Hoetker JD Bhatnagar A Baba SP Jones SP High fat feeding in mice isinsufficient to induce cardiac dysfunction and does not exacerbate heart failure PLoSOne 20138e83174
43 Gupte AA Minze LJ Reyes M Ren Y Wang X Brunner G Ghosn M Cordero-Reyes AM Ding K Pratico D Morrisett J Shi ZZ Hamilton DJ Lyon CJ Hsueh WAHigh-fat feeding-induced hyperinsulinemia increases cardiac glucose uptake and mito-chondrial function despite peripheral insulin resistance Endocrinology20131542650ndash2662
44 Logan SK Garabedian MJ Campbell CE Werb Z Synergistic transcriptionalactivation of the tissue inhibitor of metalloproteinases-1 promoter via functionalinteraction of AP-1 and Ets-1 transcription factors J Biol Chem1996271774ndash782
45 Belke DD Betuing S Tuttle MJ Graveleau C Young ME Pham M Zhang DCooksey RC McClain DA Litwin SE Taegtmeyer H Severson D Kahn CR AbelED Insulin signaling coordinately regulates cardiac size metabolism and contractileprotein isoform expression J Clin Invest 2002109629ndash639
46 Fernandez AM Kim JK Yakar S Dupont J Hernandez-Sanchez C Castle AL FilmoreJ Shulman GI Le Roith D Functional inactivation of the IGF-I and insulin receptors inskeletal muscle causes type 2 diabetes Genes Dev 2001151926ndash1934
47 Gilchrist M Thorsson V Li B Rust AG Korb M Roach JC Kennedy K Hai TBolouri H Aderem A Systems biology approaches identify ATF3 as a negative regu-lator of Toll-like receptor 4 Nature 2006441173ndash178
48 Kristiansen OP Mandrup-Poulsen T Interleukin-6 and diabetes the good the bad orthe indifferent Diabetes 200554(Suppl 2)S114ndashS124
49 Grueter CE van Rooij E Johnson BA DeLeon SM Sutherland LB Qi X Gautron LElmquist JK Bassel-Duby R Olson EN A cardiac microRNA governs systemic energyhomeostasis by regulation of MED13 Cell 2012149671ndash683
50 Baskin KK Grueter CE Kusminski CM Holland WL Bookout AL Satapati S KongYM Burgess SC Malloy CR Scherer PE Newgard CB Bassel-Duby R Olson ENMED13-dependent signaling from the heart confers leanness by enhancing metabo-lism in adipose tissue and liver EMBO Mol Med 201461610ndash1621
51 Nakamura M Sadoshima J Heart over mind metabolic control of white adipose tis-sue and liver EMBO Mol Med 201461521ndash1524
52 Tenenbaum A Fisman EZ Impaired glucose metabolism in patients with heart failurepathophysiology and possible treatment strategies Am J Cardiovasc Drugs20044269ndash280
53 Suskin N McKelvie RS Burns RJ Latini R Pericak D Probstfield J Rouleau JL SigouinC Solymoss CB Tsuyuki R White M Yusuf S Glucose and insulin abnormalitiesrelate to functional capacity in patients with congestive heart failure Eur Heart J2000211368ndash1375
54 Aronson D Hammerman H Kapeliovich MR Suleiman A Agmon Y Beyar RMarkiewicz W Suleiman M Fasting glucose in acute myocardial infarction incremen-tal value for long-term mortality and relationship with left ventricular systolic func-tion Diabetes Care 200730960ndash966
146 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
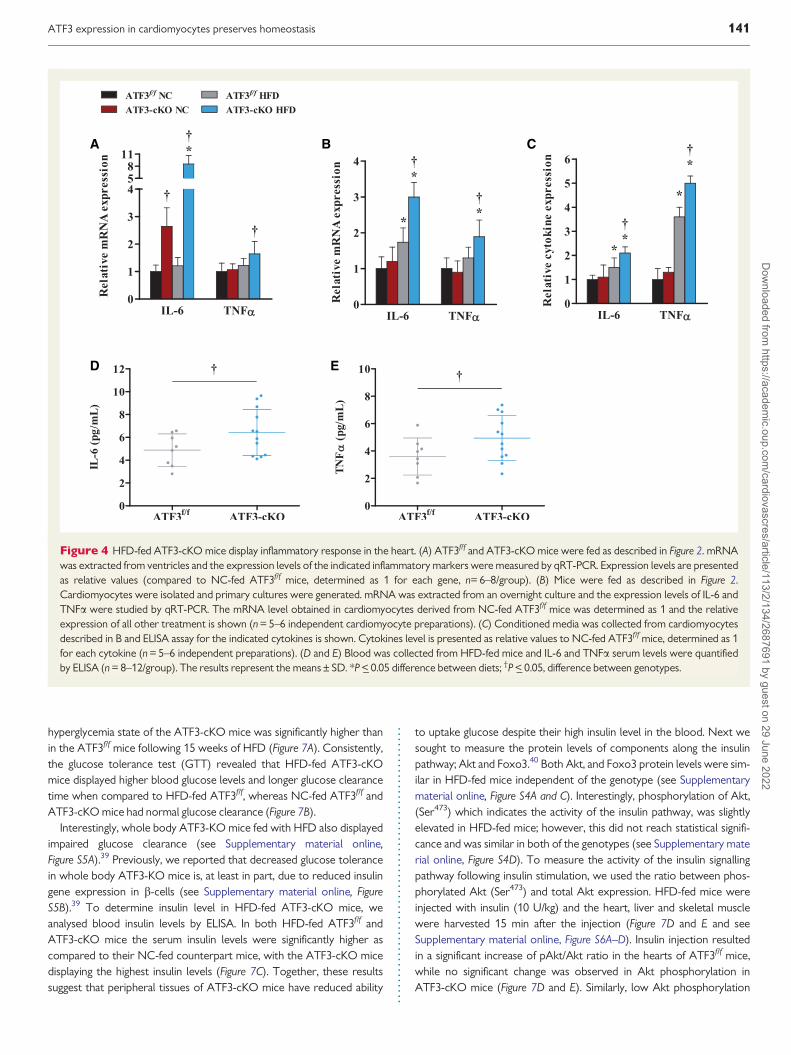
hyperglycemia state of the ATF3-cKO mice was significantly higher thanin the ATF3ff mice following 15 weeks of HFD (Figure 7A) Consistentlythe glucose tolerance test (GTT) revealed that HFD-fed ATF3-cKOmice displayed higher blood glucose levels and longer glucose clearancetime when compared to HFD-fed ATF3ff whereas NC-fed ATF3ff andATF3-cKO mice had normal glucose clearance (Figure 7B)
Interestingly whole body ATF3-KO mice fed with HFD also displayedimpaired glucose clearance (see Supplementary material onlineFigure S5A)39 Previously we reported that decreased glucose tolerancein whole body ATF3-KO mice is at least in part due to reduced insulingene expression in b-cells (see Supplementary material online FigureS5B)39 To determine insulin level in HFD-fed ATF3-cKO mice weanalysed blood insulin levels by ELISA In both HFD-fed ATF3ff andATF3-cKO mice the serum insulin levels were significantly higher ascompared to their NC-fed counterpart mice with the ATF3-cKO micedisplaying the highest insulin levels (Figure 7C) Together these resultssuggest that peripheral tissues of ATF3-cKO mice have reduced ability
to uptake glucose despite their high insulin level in the blood Next wesought to measure the protein levels of components along the insulinpathway Akt and Foxo340 Both Akt and Foxo3 protein levels were sim-ilar in HFD-fed mice independent of the genotype (see Supplementarymaterial online Figure S4A and C) Interestingly phosphorylation of Akt(Ser473) which indicates the activity of the insulin pathway was slightlyelevated in HFD-fed mice however this did not reach statistical signifi-cance and was similar in both of the genotypes (see Supplementary material online Figure S4D) To measure the activity of the insulin signallingpathway following insulin stimulation we used the ratio between phos-phorylated Akt (Ser473) and total Akt expression HFD-fed mice wereinjected with insulin (10 Ukg) and the heart liver and skeletal musclewere harvested 15 min after the injection (Figure 7D and E and seeSupplementary material online Figure S6AndashD) Insulin injection resultedin a significant increase of pAktAkt ratio in the hearts of ATF3ff micewhile no significant change was observed in Akt phosphorylation inATF3-cKO mice (Figure 7D and E) Similarly low Akt phosphorylation
A
D E
B C
Figure 4 HFD-fed ATF3-cKO mice display inflammatory response in the heart (A) ATF3ff and ATF3-cKO mice were fed as described in Figure 2 mRNAwas extracted from ventricles and the expression levels of the indicated inflammatory markers were measured by qRT-PCR Expression levels are presentedas relative values (compared to NC-fed ATF3ff mice determined as 1 for each gene n= 6ndash8group) (B) Mice were fed as described in Figure 2Cardiomyocytes were isolated and primary cultures were generated mRNA was extracted from an overnight culture and the expression levels of IL-6 andTNFa were studied by qRT-PCR The mRNA level obtained in cardiomyocytes derived from NC-fed ATF3ff mice was determined as 1 and the relativeexpression of all other treatment is shown (n = 5ndash6 independent cardiomyocyte preparations) (C) Conditioned media was collected from cardiomyocytesdescribed in B and ELISA assay for the indicated cytokines is shown Cytokines level is presented as relative values to NC-fed ATF3ff mice determined as 1for each cytokine (n = 5ndash6 independent preparations) (D and E) Blood was collected from HFD-fed mice and IL-6 and TNFa serum levels were quantifiedby ELISA (n = 8ndash12group) The results represent the meansplusmn SD P lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes
ATF3 expression in cardiomyocytes preserves homeostasis 141D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
levels were also observed in both the liver and skeletal muscle of theATF3-cKO mice however this did not reach statistically significance (seeSupplementary material online Figure S6AndashD) Consistently palmitate-treated cardiomyocytes derived from ATF3-cKO mice displayedreduced phosphorylation of Akt following 30-min insulin stimulation(see Supplementary material online Figure S6E)
Collectively our findings revealed a surprising phenomenon that car-diac ATF3 deficiency results in increased cardiac remodeling leading topotentiation of local and peripheral insulin resistance
4 Discussion
A HFD is a commonly used regimen to induce T2D The majority of theliterature focuses on diabetic symptoms without addressing the issue ofT2D-induced cardiomyopathy Even when it is addressed there is noclear consensus regarding the extent of cardiac effects imposed byHFD41 Diet-induced obesity in mouse models is associated with globalinsulin resistance and increased myocardial fatty acid utilization how-ever the reported phenotypes on cardiac function vary from extensivehypertrophy and heart failure4 to lack of cardiac dysfunction4243 Thesevariations have been attributed to diet regimens andor sensitivity ofmouse strains to a HFD In this report we present evidence that cardiacATF3 deficiency exacerbates the deleterious effect of a HFD on theheart such as maladaptive cardiac remodeling fibrosis inflammation andcardiac dysfunction Our work may provide a potential molecular insightto variations in the literature regarding the impact of HFD on the heartAs ATF3 is a stress-inducible gene we propose that its proper inductionin the cardiomyocytes is protective and helps the heart to cope withHFD stress It is possible that the induction of ATF3mdashits expressionlevel duration and kineticsmdashvaries in different HFD regimens andormouse strains contributing to the variations observed in the literature
HFD regimen induced a significant higher level of fibrosis in the heartof ATF3-cKO mice We have identified TIMP1 as a possible mechanismto explain the increase in fibrosis in ATF3-cKO cardiomyocytes Severallines of evidence suggest that TIMP1 is possibly an ATF3 target gene
First we observed an increase in TIMP1 transcription in the whole heartand in isolated cardiomyocytes (Figures 1A and 3A) Second TIMP 1 3and 4 were identified as significantly highly expressed genes in the heartof ATF3-KO mice as compared with wild type mice following aorticbanding20 Third a functional AP-1 regulatory element within the TIMP1promoter was described44 Indeed preliminary results in our lab suggestthat ATF3 downregulates the TIMP1 promoter (data not shown) Thusit is possible that TIMP1 is suppressed by ATF3 and therefore results in anet increase in MMPs activity that helps to maintain ECM formationdeg-radation homeostasis
Our findings are consistent with previous studies demonstrating that adeficiency in ATF3 can promote cardiac maladaptive changes which leadto cardiac dilatation and hypertrophy in an aortic banding model1920 Incontrast a different study using a pressure overload model reportedthat ATF3 induces a maladaptive cardiac response17 These seeminglyconflicting data may be explained by the different patterns of ATF3expression induced by the different models In the pressure overloadmodel the acute and transient expression of ATF3 in cardiomyocytesinitiates a cascade of maladaptive changes17 whereas in the aortic band-ing model and the HFD model described here ATF3 expression is longlasting
In this context this is the first report using mice with cardiac-selectivedeficiency of ATF3 (ATF3-cKO) Previous reports on the roles of ATF3in cardiac function were carried out using whole body knockout ofATF325 or transgenic mice ectopically expressing ATF3 in cardiomyo-cytes16 A key advancement of ATF3-cKO mice over the whole bodyknockout mice is that they provide insights into the function of ATF3selectively in cardiomyocytes without the complication of ATF3 defi-ciency in other cells Thus the ATF3-cKO mouse is a useful tool to bet-ter understand the function of cardiac ATF3
Consistent with our previous report25 here we showed that cardiacATF3 deficiency has a more severe HFD-induced diabetic phenotypethan the control wild type mice However while reduced insulin genetranscription in ATF3-deficient b-cells was observed25 our current dataindicates that a lack of insulin is not the explanation for the glucose intol-erance found in the HFD-fed ATF3-cKO mice In contrast these mice
A B
Figure 5 HFD-fed ATF3-cKO mice show metabolic response in the heart ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) mRNA wasextracted from ventricles and the expression levels of the indicated metabolic markers were measured by qRT-PCR Expression levels are presented as rela-tive values (compared to NC-fed ATF3ff mice determined as 1 for each gene n= 6ndash8group) (B) Cardiomyocytes were isolated and plated on laminin for1 h followed by mRNA preparation (n = 4ndash5 independent preparations) The expression levels of the indicated metabolic markers were measured by qRT-PCR Expression levels are presented as relative values (to NC-fed ATF3ff mice determined as 1 for each gene) The results represent the meansplusmn SDP lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes
142 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
had higher blood insulin level (Figure 7C) Therefore upon HFD chal-lenge cardiac-deficiency of ATF3 exacerbates HFD-induced T2D Thislsquoperipheral effectrsquo is surprising since ATF3 is selectively deleted in thecardiomyocytes
The question then is how can we explain the effect of cardiacATF3 deficiency on peripheral glucose intolerance One possibility isthe reduced IRS1 expression in ATF3-deficient cardiomyocytes bothin vivo and in vitro IRS1 suppression may lead to impaired insulin sig-nalling in the heart thereby exacerbating glucose intolerance Indeedwe observed that HFD-fed ATF3-cKO mice displayed insulin resist-ance upon insulin challenge when compared to HFD-fed ATF3ff
mice In addition palmitate-treated cardiomyocytes lacking ATF3 dis-played lower insulin signalling as compared to their wild type coun-terparts Since the heart is only a minor glucose consuming organ incomparison to skeletal muscle cells or the brain the peripheral sys-temic effect cannot be solely based on this observation A case inpoint transgenic mice with insulin receptor (IR) deficiency in theheart do not display a diabetic phenotype4546 Another attractive
possibility for the peripheral effect of cardiac ATF3-deletion is itsimpact on inflammation It is well established that low-grade inflam-mation years in advance of the disease onset is involved in the patho-genic processes causing T2D8 Mediators of inflammation such asTNFa interleukin (IL)-1b the IL-6 family of cytokines IL-18 and cer-tain chemokines have been proposed to be involved in the eventscausing both forms of diabetes (T1D and T2D)8 Interestingly wehave shown that IL-6 a well-known ATF3 target gene47 is known toenhance its transcription and secretion of cytokines such as IL-6 andTNFa in palmitate-treated primary cultures of ATF3-cKO cardio-myocytes as well as in cardiomyocytes derived from HFD-fed ATF3-cKO mice IL-6 has been proposed to play a role in glucose homeo-stasis and metabolism directly and indirectly via its action on skeletalmuscle cells adipocytes hepatocytes pancreatic b-cells and neuro-endocrine cells48
Therefore we hypothesize that elevated levels of inflammatory cyto-kines in ATF3-cKO mice mediate a cross-talk in peripheral tissues leadingto impaired glucose tolerance Although circulating IL-6 and TNFa
A
B
C
D
E
F
Figure 6 HFD-fed ATF3-cKO mice present deteriorated heart function ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) At endpointmice hearts were examined by micro ultrasound The following parameters were measured interventricular septal end diastole (IVSd) left ventricular pos-terior wall end diastole (LVPWd) maximal left ventricular internal end-diastole (LVIDd) end-systole (LVIDs) and heart rate The results represent themeansplusmn SD (n = 6ndash8 in each group as shown in panel B) (B) Heart function was assessed by calculating percentage of fractional shortening (FS) accordingto FS () = [(LVDd-LVDs)LVDd]100 The results represent the meansplusmn SD (n = 6ndash8 in each group) (C and D) Left ventricular cardiac volumes massand function were examined by a cardiac MRI The following parameters were measured left ventricular end-diastolic (LVEDV) and left ventricular end-sys-tolic volume (LVESV) left ventricular (LV) mass and ejection fraction (EF) The results represent the meansplusmn SD (n = 7 in each group) (E and F)Representative images of mid-ventricular short-axis (E) and long-axis (F) slice at peak diastole and systole P lt_ 005 difference between diets daggerP lt_ 005 dif-ference between genotypes
ATF3 expression in cardiomyocytes preserves homeostasis 143D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
cytokines may be derived from different tissues we provide ample evi-dence that ATF3-defficient cardiomyocytes produce and secrete higherlevels of these cytokines as compared to their wild type counterpartsThis indicates a lsquocardiac-to-peripheralrsquo effect in T2D-induced cardiomyop-athy This is a reverse direction from the lsquoperipheral-to-cardiacrsquo effect dis-cussed in the introduction where inflammation in the peripheral tissuesleads to cardiac dysfunction Since inflammatory cytokines are soluble
factors they can exert a systemic effect This is why it makes sense thattheir effect can go both ways Thus diabetes and cardiomyopathy are apart of a vicious cycle Previously it was shown that cardiac specific dele-tion of the thyroid hormone complex mediator MED13 results in aperipheral effect upon HFD Cardiac MED13 deficiency suppresses meta-bolic genes in the heart which leads to a decrease in energy expenditureand an obese phenotype most likely by the secretion of cardiokines that
A
B
C
D
E
Figure 7 HFD-fed ATF3-cKO mice exhibit impaired glucose tolerance ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) Fasting (6 h)blood glucose concentrations (mgdl) The results represent the meansplusmn SD (n = 20ndash30group) P lt_ 005 difference between diets daggerP lt_ 005 differencebetween genotypes (B) Glucose tolerance test (GTT) Values are presented as relative to initial valueplusmn SD (n = 20ndash25group) P lt_ 005 difference betweendiets daggerP lt_ 005 difference between genotypes (C) Serum insulin concentrations were quantified by ELISA Values are presented as relative values (com-pared to NC-fed ATF3ff mice determined as 1 n = 15ndash20group) P lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes (D and E)Effect of HFD on phosphorylated Akt (Ser473 pAkt) to Akt ratio levels in heart lysate following either saline or insulin injection (10 U1 kg) is shownRepresentative blots (D) Densitometric analysis is presented as means ratio (n = 3ndash6group) pAktAkt ratio is presented as relative values (compared toHFD-fed saline injected ATF3ff mice determined as 1) P lt_ 005 insulin vs saline treatment daggerP lt_ 005 difference between genotypes (E)
144 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
mediates the cross talk between the heart and peripheral tissues49ndash51
Here we also observed increased levels of natriuretic peptides (ANP andBNP) which together with IL-6 and TNFa may mediate the cross-talk withthe peripheral tissues to induce glucose intolerance
Does this finding have any relevance to cardiac disease in humansSeveral lines of evidence suggests that congestive heart failure in patientsis associated with an increase in blood glucose levels52 In addition highblood glucose levels in non-diabetic patients with acute myocardialinfarction serves as a prognostic predictor of long-term mortality5354
Thus here we provide evidence that high blood glucose levels are a partof the symptoms of cardiac remodeling process
Collectively our findings suggest that cardiac ATF3 expression isresponsible for an adaptive cardiac response to the stressful environmentunder a HFD (Figure 8) ATF3 dampens the inflammatory response andsuppresses the expression of ECM remodeling genes and thus amelioratescardiac function In HFD-fed mice with ATF3 deficiency or in mice withcardiac remodeling the inflamed heart affects also peripheral tissuesthereby worsening T2D in peripheral tissues resulting in elevated bloodglucose level and turning on a vicious cycle to worsen cardiomyopathy
Supplementary material
Supplementary material is available at Cardiovascular Research online
AcknowledgementsThe authors wish to thank Drs Keren Bitton-Worms and Ran Rostokerand Prof Derek LeRoith from the Rambam Medical Center for their
assistance in GTT and ITT Dr Edith Suss Toby Maya Holdengreber andLior Liba from the BCF Bioimaging Center (Faculty of MedicineTechnion) for their assistance with imaging and image analysis Prof DEMichelle from University of Michigan for cardiomyocytes culture techni-que and Prof Yehiel Zick from the Weizmann institute of Sciences forcritical comments
Conflict of interest none declared
FundingThis work was partially supported by the United States-Israel BinationalGrant 2009179 to AA and TH In addition we wish to thank the MarsPittsburgh Fund for Medical Research to AA the Department of Defensegrant W81XWH-14-1-0179 to TH and to D Dan and Betty Kahn travelgrant support to LK
References
1 Roger VL Go AS Lloyd-Jones DM Benjamin EJ Berry JD Borden WB Bravata DMDai S Ford ES Fox CS Fullerton HJ Gillespie C Hailpern SM Heit JA Howard VJKissela BM Kittner SJ Lackland DT Lichtman JH Lisabeth LD Makuc DM MarcusGM Marelli A Matchar DB Moy CS Mozaffarian D Mussolino ME Nichol GPaynter NP Soliman EZ Sorlie PD Sotoodehnia N Turan TN Virani SS Wong NDWoo D Turner MB American Heart Association Statistics C Stroke Statistics SHeart disease and stroke statisticsndash2012 update a report from the American HeartAssociation Circulation 2012125e2ndashe220
2 Wilson C Diabetes ACCORD 5-year outcomes of intensive glycemic control NatRev Endocrinol 20117314
3 Ernande L Derumeaux G Diabetic cardiomyopathy myth or reality Arch CardiovascDis 2012105218ndash225
4 Battiprolu PK Hojayev B Jiang N Wang ZV Luo X Iglewski M Shelton JM GerardRD Rothermel BA Gillette TG Lavandero S Hill JA Metabolic stress-induced
InsulinGF
Cytokines
FFA
GlucoseATF3
Cardiac
Remodeling
Factors
Paracrine action
Collagens
and ECM
proteins
Fibrosis
Cardiac
Dysfunction
Systemic Effects
Glucose Tolerance
TNFα
IL-6
Cardiomyocyte
ATF3
Homeostasis
Homeostasis
TNFα
IL-6
TIMP1
BNP
ANP
Figure 8 lsquoPeripheral-to-cardiacrsquo and lsquoCardiac-to-peripheralrsquo model scheme Schematic diagram showing the ATF3 adaptive role in maintaining hearthomeostasis following HFD-induced T2D In contrast stressful environment under HFD leads to maladaptive cardiac response in the absence of ATF3Cardiac remodeling caused by ATF3 deficiency induces an increase in cardiac ECM remodeling and inflammatory genes and mediates the communication ofcardiomyocytes with heart resident cells potentially mediated by IL-6 TNFa ANP BNP and TIMP1 In this situation increased cardiac fibrosis and inflamma-tory response worsen cardiac function The inflamed failing heart mediates the communication of cardiomyocytes with distant organs as well thereby affect-ing diabetic outcome and glucose homeostasis
ATF3 expression in cardiomyocytes preserves homeostasis 145D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
activation of FoxO1 triggers diabetic cardiomyopathy in mice J Clin Invest20121221109ndash1118
5 Mandavia CH Aroor AR Demarco VG Sowers JR Molecular and metabolic mecha-nisms of cardiac dysfunction in diabetes Life Sci 201392601ndash608
6 Ventura-Clapier R Garnier A Veksler V Energy metabolism in heart failure J Physiol20045551ndash13
7 Guo S Insulin signaling resistance and the metabolic syndrome insights from mousemodels into disease mechanisms J Endocrinol 2014220T1ndashT23
8 Montane J Cadavez L Novials A Stress and the inflammatory process a major causeof pancreatic cell death in type 2 diabetes Diabetes Metab Syndr Obes 2014725ndash34
9 Ashrafian H Frenneaux MP Opie LH Metabolic mechanisms in heart failureCirculation 2007116434ndash448
10 Tanabe K Liu Y Hasan SD Martinez SC Cras-Meneur C Welling CM Bernal-MizrachiE Tanizawa Y Rhodes CJ Zmuda E Hai T Abumrad NA Permutt MA Glucose andfatty acids synergize to promote B-cell apoptosis through activation of glycogen syn-thase kinase 3beta independent of JNK activation PLoS One 20116e18146
11 Heineke J Molkentin JD Regulation of cardiac hypertrophy by intracellular signallingpathways Nat Rev Mol Cell Biol 20067589ndash600
12 Hai T Wolford CC Chang YS ATF3 a hub of the cellular adaptive-response net-work in the pathogenesis of diseases is modulation of inflammation a unifying com-ponent Gene Expr 2010151ndash11
13 Takii R Inouye S Fujimoto M Nakamura T Shinkawa T Prakasam R Tan K HayashidaN Ichikawa H Hai T Nakai A Heat shock transcription factor 1 inhibits expression ofIL-6 through activating transcription factor 3 J Immunol 20101841041ndash1048
14 Giraldo A Barrett OP Tindall MJ Fuller SJ Amirak E Bhattacharya BS Sugden PHClerk A Feedback regulation by Atf3 in the endothelin-1-responsive transcriptomeof cardiomyocytes Egr1 is a principal Atf3 target Biochem J 2012444343ndash355
15 Hasin T Elhanani O Abassi Z Hai T Aronheim A Angiotensin II signaling up-regulates the immediate early transcription factor ATF3 in the left but not the rightatrium Basic Res Cardiol 2011106175ndash187
16 Koren L Elhanani O Kehat I Hai T Aronheim A Adult cardiac expression of theactivating transcription factor 3 ATF3 promotes ventricular hypertrophy PLoS One20138e68396
17 Koren L Alishekevitz D Elhanani O Nevelsky A Hai T Kehat I Shaked Y AronheimA ATF3-dependent cross-talk between cardiomyocytes and macrophages promotescardiac maladaptive remodeling Int J Cardiol 2015198232ndash240
18 Okamoto Y Chaves A Chen J Kelley R Jones K Weed HG Gardner KL Gangi LYamaguchi M Klomkleaw W Nakayama T Hamlin RL Carnes C Altschuld R BauerJ Hai T Transgenic mice with cardiac-specific expression of activating transcriptionfactor 3 a stress-inducible gene have conduction abnormalities and contractile dys-function Am J Pathol 2001159639ndash650
19 Zhou H Shen DF Bian ZY Zong J Deng W Zhang Y Guo YY Li H Tang QZActivating transcription factor 3 deficiency promotes cardiac hypertrophy dysfunc-tion and fibrosis induced by pressure overload PLoS One 20116e26744
20 Zhou H Guo H Zong J Dai J Yuan Y Bian ZY Tang QZ ATF3 regulates multipletargets and may play a dual role in cardiac hypertrophy and injury Int J Cardiol2014174838ndash839
21 Yang J Yang CJ Yang J Fan ZX ATF3 a promotion effect or a inhibition effect in car-diac maladaptive remodeling Int J Cardiol 2015201245ndash246
22 Koren L Shaked Y Aronheim A Response letter ldquoATF3 a promoter or inhibitor ofcardiac maladaptive remodelingrdquo Int J Cardiol 2015201692
23 Ma ZG Wei WY Xu SC Zhang WB Dai J Tang QZ ATF3 a potential target forcardiac maladaptive remodeling Int J Cardiol 201620250ndash51
24 Zhou H Yuan Y Ni J Guo H Deng W Bian ZY Tang QZ Pleiotropic and puzzlingeffects of ATF3 in maladaptive cardiac remodeling Int J Cardiol 201620687ndash88
25 Zmuda EJ Qi L Zhu MX Mirmira RG Montminy MR Hai T The roles of ATF3 anadaptive-response gene in high-fat-diet-induced diabetes and pancreatic beta-celldysfunction Mol Endocrinol 2010241423ndash1433
26 Lu D Chen J Hai T The regulation of ATF3 gene expression by mitogen-activatedprotein kinases Biochem J 2007401559ndash567
27 Hartman MG Lu D Kim ML Kociba GJ Shukri T Buteau J Wang X Frankel WLGuttridge D Prentki M Grey ST Ron D Hai T Role for activating transcription fac-tor 3 in stress-induced beta-cell apoptosis Mol Cell Biol 2004245721ndash5732
28 Wolford CC McConoughey SJ Jalgaonkar SP Leon M Merchant AS Dominick JLYin X Chang Y Zmuda EJ OrsquoToole SA Millar EK Roller SL Shapiro CL OstrowskiMC Sutherland RL Hai T Transcription factor ATF3 links host adaptive response tobreast cancer metastasis J Clin Iinvest 20131232893ndash2906
29 Agah R Frenkel PA French BA Michael LH Overbeek PA Schneider MD Generecombination in postmitotic cells Targeted expression of Cre recombinase pro-vokes cardiac-restricted site-specific rearrangement in adult ventricular musclein vivo J Clin Invest 1997100169ndash179
30 Livet J Weissman TA Kang H Draft RW Lu J Bennis RA Sanes JR Lichtman JWTransgenic strategies for combinatorial expression of fluorescent proteins in thenervous system Nature 200745056ndash62
31 Davis J Maillet M Miano JM Molkentin JD Lost in transgenesis a userrsquos guidefor genetically manipulating the mouse in cardiac research Circ Res2012111761ndash777
32 Heiberg E Sjogren J Ugander M Carlsson M Engblom H Arheden H Design andvalidation of segment ndash freely available software for cardiovascular image analysisBMC Med Imaging 2010101
33 Kabaeva Z Zhao M Michele DE Blebbistatin extends culture life of adult mouse car-diac myocytes and allows efficient and stable transgene expression Am J Physiol HeartC 2008294H1667ndash1674
34 Subramaniam A Jones WK Gulick J Wert S Neumann J Robbins J Tissue-specificregulation of the alpha-myosin heavy chain gene promoter in transgenic mice J BiolChem 199126624613ndash24620
35 Cunha DA Hekerman P Ladriere L Bazarra-Castro A Ortis F Wakeham MCMoore F Rasschaert J Cardozo AK Bellomo E Overbergh L Mathieu C Lupi R HaiT Herchuelz A Marchetti P Rutter GA Eizirik DL Cnop M Initiation and executionof lipotoxic ER stress in pancreatic beta-cells J Cell Sci 20081212308ndash2318
36 Wynn TA Ramalingam TR Mechanisms of fibrosis therapeutic translation forfibrotic disease Nat Med 2012181028ndash1040
37 Li D Yin X Zmuda EJ Wolford CC Dong X White MF Hai T The repression ofIRS2 gene by ATF3 a stress-inducible gene contributes to pancreatic beta-cell apop-tosis Diabetes 200857635ndash644
38 Myerson SG Bellenger NG Pennell DJ Assessment of left ventricular mass by cardi-ovascular magnetic resonance Hypertension 200239750ndash755
39 Zmuda EJ Viapiano M Grey ST Hadley G Garcia-Ocana A Hai T Deficiency ofAtf3 an adaptive-response gene protects islets and ameliorates inflammation in asyngeneic mouse transplantation model Diabetologia 2010531438ndash1450
40 Riehle C Abel ED Insulin signaling and heart failure Circ Res 20161181151ndash116941 Schreyer SA Wilson DL LeBoeuf RC C57BL6 mice fed high fat diets as models for
diabetes-accelerated atherosclerosis Atherosclerosis 199813617ndash2442 Brainard RE Watson LJ Demartino AM Brittian KR Readnower RD Boakye AA
Zhang D Hoetker JD Bhatnagar A Baba SP Jones SP High fat feeding in mice isinsufficient to induce cardiac dysfunction and does not exacerbate heart failure PLoSOne 20138e83174
43 Gupte AA Minze LJ Reyes M Ren Y Wang X Brunner G Ghosn M Cordero-Reyes AM Ding K Pratico D Morrisett J Shi ZZ Hamilton DJ Lyon CJ Hsueh WAHigh-fat feeding-induced hyperinsulinemia increases cardiac glucose uptake and mito-chondrial function despite peripheral insulin resistance Endocrinology20131542650ndash2662
44 Logan SK Garabedian MJ Campbell CE Werb Z Synergistic transcriptionalactivation of the tissue inhibitor of metalloproteinases-1 promoter via functionalinteraction of AP-1 and Ets-1 transcription factors J Biol Chem1996271774ndash782
45 Belke DD Betuing S Tuttle MJ Graveleau C Young ME Pham M Zhang DCooksey RC McClain DA Litwin SE Taegtmeyer H Severson D Kahn CR AbelED Insulin signaling coordinately regulates cardiac size metabolism and contractileprotein isoform expression J Clin Invest 2002109629ndash639
46 Fernandez AM Kim JK Yakar S Dupont J Hernandez-Sanchez C Castle AL FilmoreJ Shulman GI Le Roith D Functional inactivation of the IGF-I and insulin receptors inskeletal muscle causes type 2 diabetes Genes Dev 2001151926ndash1934
47 Gilchrist M Thorsson V Li B Rust AG Korb M Roach JC Kennedy K Hai TBolouri H Aderem A Systems biology approaches identify ATF3 as a negative regu-lator of Toll-like receptor 4 Nature 2006441173ndash178
48 Kristiansen OP Mandrup-Poulsen T Interleukin-6 and diabetes the good the bad orthe indifferent Diabetes 200554(Suppl 2)S114ndashS124
49 Grueter CE van Rooij E Johnson BA DeLeon SM Sutherland LB Qi X Gautron LElmquist JK Bassel-Duby R Olson EN A cardiac microRNA governs systemic energyhomeostasis by regulation of MED13 Cell 2012149671ndash683
50 Baskin KK Grueter CE Kusminski CM Holland WL Bookout AL Satapati S KongYM Burgess SC Malloy CR Scherer PE Newgard CB Bassel-Duby R Olson ENMED13-dependent signaling from the heart confers leanness by enhancing metabo-lism in adipose tissue and liver EMBO Mol Med 201461610ndash1621
51 Nakamura M Sadoshima J Heart over mind metabolic control of white adipose tis-sue and liver EMBO Mol Med 201461521ndash1524
52 Tenenbaum A Fisman EZ Impaired glucose metabolism in patients with heart failurepathophysiology and possible treatment strategies Am J Cardiovasc Drugs20044269ndash280
53 Suskin N McKelvie RS Burns RJ Latini R Pericak D Probstfield J Rouleau JL SigouinC Solymoss CB Tsuyuki R White M Yusuf S Glucose and insulin abnormalitiesrelate to functional capacity in patients with congestive heart failure Eur Heart J2000211368ndash1375
54 Aronson D Hammerman H Kapeliovich MR Suleiman A Agmon Y Beyar RMarkiewicz W Suleiman M Fasting glucose in acute myocardial infarction incremen-tal value for long-term mortality and relationship with left ventricular systolic func-tion Diabetes Care 200730960ndash966
146 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
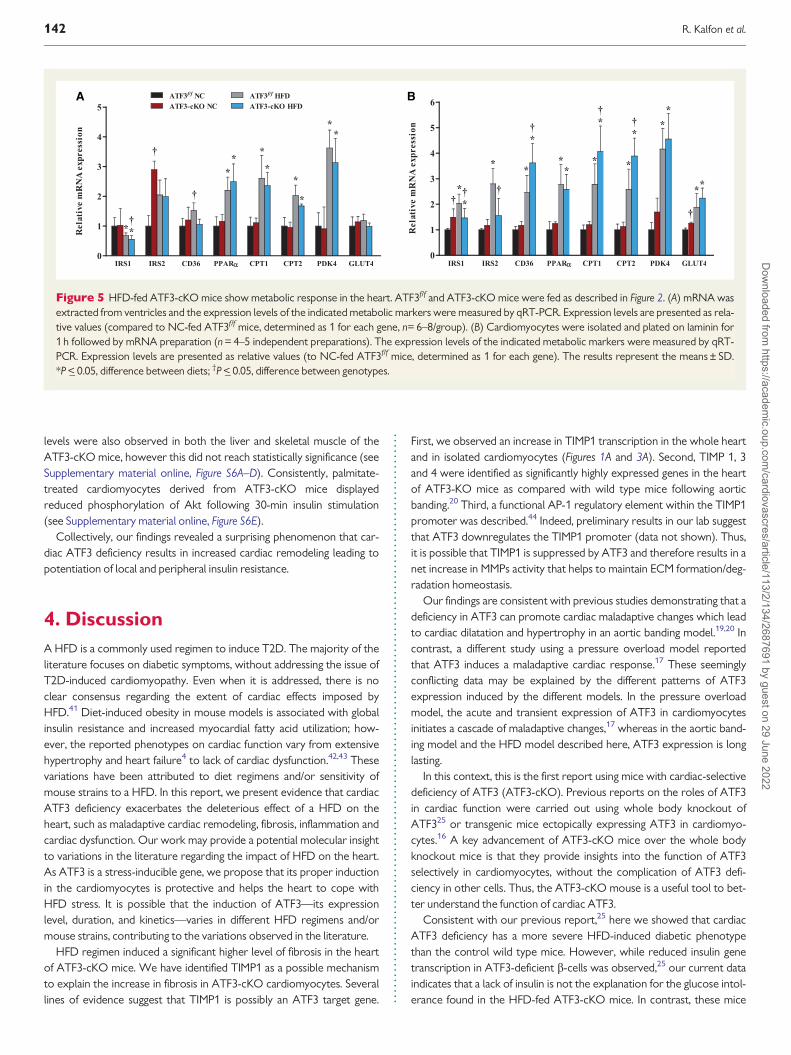
levels were also observed in both the liver and skeletal muscle of theATF3-cKO mice however this did not reach statistically significance (seeSupplementary material online Figure S6AndashD) Consistently palmitate-treated cardiomyocytes derived from ATF3-cKO mice displayedreduced phosphorylation of Akt following 30-min insulin stimulation(see Supplementary material online Figure S6E)
Collectively our findings revealed a surprising phenomenon that car-diac ATF3 deficiency results in increased cardiac remodeling leading topotentiation of local and peripheral insulin resistance
4 Discussion
A HFD is a commonly used regimen to induce T2D The majority of theliterature focuses on diabetic symptoms without addressing the issue ofT2D-induced cardiomyopathy Even when it is addressed there is noclear consensus regarding the extent of cardiac effects imposed byHFD41 Diet-induced obesity in mouse models is associated with globalinsulin resistance and increased myocardial fatty acid utilization how-ever the reported phenotypes on cardiac function vary from extensivehypertrophy and heart failure4 to lack of cardiac dysfunction4243 Thesevariations have been attributed to diet regimens andor sensitivity ofmouse strains to a HFD In this report we present evidence that cardiacATF3 deficiency exacerbates the deleterious effect of a HFD on theheart such as maladaptive cardiac remodeling fibrosis inflammation andcardiac dysfunction Our work may provide a potential molecular insightto variations in the literature regarding the impact of HFD on the heartAs ATF3 is a stress-inducible gene we propose that its proper inductionin the cardiomyocytes is protective and helps the heart to cope withHFD stress It is possible that the induction of ATF3mdashits expressionlevel duration and kineticsmdashvaries in different HFD regimens andormouse strains contributing to the variations observed in the literature
HFD regimen induced a significant higher level of fibrosis in the heartof ATF3-cKO mice We have identified TIMP1 as a possible mechanismto explain the increase in fibrosis in ATF3-cKO cardiomyocytes Severallines of evidence suggest that TIMP1 is possibly an ATF3 target gene
First we observed an increase in TIMP1 transcription in the whole heartand in isolated cardiomyocytes (Figures 1A and 3A) Second TIMP 1 3and 4 were identified as significantly highly expressed genes in the heartof ATF3-KO mice as compared with wild type mice following aorticbanding20 Third a functional AP-1 regulatory element within the TIMP1promoter was described44 Indeed preliminary results in our lab suggestthat ATF3 downregulates the TIMP1 promoter (data not shown) Thusit is possible that TIMP1 is suppressed by ATF3 and therefore results in anet increase in MMPs activity that helps to maintain ECM formationdeg-radation homeostasis
Our findings are consistent with previous studies demonstrating that adeficiency in ATF3 can promote cardiac maladaptive changes which leadto cardiac dilatation and hypertrophy in an aortic banding model1920 Incontrast a different study using a pressure overload model reportedthat ATF3 induces a maladaptive cardiac response17 These seeminglyconflicting data may be explained by the different patterns of ATF3expression induced by the different models In the pressure overloadmodel the acute and transient expression of ATF3 in cardiomyocytesinitiates a cascade of maladaptive changes17 whereas in the aortic band-ing model and the HFD model described here ATF3 expression is longlasting
In this context this is the first report using mice with cardiac-selectivedeficiency of ATF3 (ATF3-cKO) Previous reports on the roles of ATF3in cardiac function were carried out using whole body knockout ofATF325 or transgenic mice ectopically expressing ATF3 in cardiomyo-cytes16 A key advancement of ATF3-cKO mice over the whole bodyknockout mice is that they provide insights into the function of ATF3selectively in cardiomyocytes without the complication of ATF3 defi-ciency in other cells Thus the ATF3-cKO mouse is a useful tool to bet-ter understand the function of cardiac ATF3
Consistent with our previous report25 here we showed that cardiacATF3 deficiency has a more severe HFD-induced diabetic phenotypethan the control wild type mice However while reduced insulin genetranscription in ATF3-deficient b-cells was observed25 our current dataindicates that a lack of insulin is not the explanation for the glucose intol-erance found in the HFD-fed ATF3-cKO mice In contrast these mice
A B
Figure 5 HFD-fed ATF3-cKO mice show metabolic response in the heart ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) mRNA wasextracted from ventricles and the expression levels of the indicated metabolic markers were measured by qRT-PCR Expression levels are presented as rela-tive values (compared to NC-fed ATF3ff mice determined as 1 for each gene n= 6ndash8group) (B) Cardiomyocytes were isolated and plated on laminin for1 h followed by mRNA preparation (n = 4ndash5 independent preparations) The expression levels of the indicated metabolic markers were measured by qRT-PCR Expression levels are presented as relative values (to NC-fed ATF3ff mice determined as 1 for each gene) The results represent the meansplusmn SDP lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes
142 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
had higher blood insulin level (Figure 7C) Therefore upon HFD chal-lenge cardiac-deficiency of ATF3 exacerbates HFD-induced T2D Thislsquoperipheral effectrsquo is surprising since ATF3 is selectively deleted in thecardiomyocytes
The question then is how can we explain the effect of cardiacATF3 deficiency on peripheral glucose intolerance One possibility isthe reduced IRS1 expression in ATF3-deficient cardiomyocytes bothin vivo and in vitro IRS1 suppression may lead to impaired insulin sig-nalling in the heart thereby exacerbating glucose intolerance Indeedwe observed that HFD-fed ATF3-cKO mice displayed insulin resist-ance upon insulin challenge when compared to HFD-fed ATF3ff
mice In addition palmitate-treated cardiomyocytes lacking ATF3 dis-played lower insulin signalling as compared to their wild type coun-terparts Since the heart is only a minor glucose consuming organ incomparison to skeletal muscle cells or the brain the peripheral sys-temic effect cannot be solely based on this observation A case inpoint transgenic mice with insulin receptor (IR) deficiency in theheart do not display a diabetic phenotype4546 Another attractive
possibility for the peripheral effect of cardiac ATF3-deletion is itsimpact on inflammation It is well established that low-grade inflam-mation years in advance of the disease onset is involved in the patho-genic processes causing T2D8 Mediators of inflammation such asTNFa interleukin (IL)-1b the IL-6 family of cytokines IL-18 and cer-tain chemokines have been proposed to be involved in the eventscausing both forms of diabetes (T1D and T2D)8 Interestingly wehave shown that IL-6 a well-known ATF3 target gene47 is known toenhance its transcription and secretion of cytokines such as IL-6 andTNFa in palmitate-treated primary cultures of ATF3-cKO cardio-myocytes as well as in cardiomyocytes derived from HFD-fed ATF3-cKO mice IL-6 has been proposed to play a role in glucose homeo-stasis and metabolism directly and indirectly via its action on skeletalmuscle cells adipocytes hepatocytes pancreatic b-cells and neuro-endocrine cells48
Therefore we hypothesize that elevated levels of inflammatory cyto-kines in ATF3-cKO mice mediate a cross-talk in peripheral tissues leadingto impaired glucose tolerance Although circulating IL-6 and TNFa
A
B
C
D
E
F
Figure 6 HFD-fed ATF3-cKO mice present deteriorated heart function ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) At endpointmice hearts were examined by micro ultrasound The following parameters were measured interventricular septal end diastole (IVSd) left ventricular pos-terior wall end diastole (LVPWd) maximal left ventricular internal end-diastole (LVIDd) end-systole (LVIDs) and heart rate The results represent themeansplusmn SD (n = 6ndash8 in each group as shown in panel B) (B) Heart function was assessed by calculating percentage of fractional shortening (FS) accordingto FS () = [(LVDd-LVDs)LVDd]100 The results represent the meansplusmn SD (n = 6ndash8 in each group) (C and D) Left ventricular cardiac volumes massand function were examined by a cardiac MRI The following parameters were measured left ventricular end-diastolic (LVEDV) and left ventricular end-sys-tolic volume (LVESV) left ventricular (LV) mass and ejection fraction (EF) The results represent the meansplusmn SD (n = 7 in each group) (E and F)Representative images of mid-ventricular short-axis (E) and long-axis (F) slice at peak diastole and systole P lt_ 005 difference between diets daggerP lt_ 005 dif-ference between genotypes
ATF3 expression in cardiomyocytes preserves homeostasis 143D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
cytokines may be derived from different tissues we provide ample evi-dence that ATF3-defficient cardiomyocytes produce and secrete higherlevels of these cytokines as compared to their wild type counterpartsThis indicates a lsquocardiac-to-peripheralrsquo effect in T2D-induced cardiomyop-athy This is a reverse direction from the lsquoperipheral-to-cardiacrsquo effect dis-cussed in the introduction where inflammation in the peripheral tissuesleads to cardiac dysfunction Since inflammatory cytokines are soluble
factors they can exert a systemic effect This is why it makes sense thattheir effect can go both ways Thus diabetes and cardiomyopathy are apart of a vicious cycle Previously it was shown that cardiac specific dele-tion of the thyroid hormone complex mediator MED13 results in aperipheral effect upon HFD Cardiac MED13 deficiency suppresses meta-bolic genes in the heart which leads to a decrease in energy expenditureand an obese phenotype most likely by the secretion of cardiokines that
A
B
C
D
E
Figure 7 HFD-fed ATF3-cKO mice exhibit impaired glucose tolerance ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) Fasting (6 h)blood glucose concentrations (mgdl) The results represent the meansplusmn SD (n = 20ndash30group) P lt_ 005 difference between diets daggerP lt_ 005 differencebetween genotypes (B) Glucose tolerance test (GTT) Values are presented as relative to initial valueplusmn SD (n = 20ndash25group) P lt_ 005 difference betweendiets daggerP lt_ 005 difference between genotypes (C) Serum insulin concentrations were quantified by ELISA Values are presented as relative values (com-pared to NC-fed ATF3ff mice determined as 1 n = 15ndash20group) P lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes (D and E)Effect of HFD on phosphorylated Akt (Ser473 pAkt) to Akt ratio levels in heart lysate following either saline or insulin injection (10 U1 kg) is shownRepresentative blots (D) Densitometric analysis is presented as means ratio (n = 3ndash6group) pAktAkt ratio is presented as relative values (compared toHFD-fed saline injected ATF3ff mice determined as 1) P lt_ 005 insulin vs saline treatment daggerP lt_ 005 difference between genotypes (E)
144 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
mediates the cross talk between the heart and peripheral tissues49ndash51
Here we also observed increased levels of natriuretic peptides (ANP andBNP) which together with IL-6 and TNFa may mediate the cross-talk withthe peripheral tissues to induce glucose intolerance
Does this finding have any relevance to cardiac disease in humansSeveral lines of evidence suggests that congestive heart failure in patientsis associated with an increase in blood glucose levels52 In addition highblood glucose levels in non-diabetic patients with acute myocardialinfarction serves as a prognostic predictor of long-term mortality5354
Thus here we provide evidence that high blood glucose levels are a partof the symptoms of cardiac remodeling process
Collectively our findings suggest that cardiac ATF3 expression isresponsible for an adaptive cardiac response to the stressful environmentunder a HFD (Figure 8) ATF3 dampens the inflammatory response andsuppresses the expression of ECM remodeling genes and thus amelioratescardiac function In HFD-fed mice with ATF3 deficiency or in mice withcardiac remodeling the inflamed heart affects also peripheral tissuesthereby worsening T2D in peripheral tissues resulting in elevated bloodglucose level and turning on a vicious cycle to worsen cardiomyopathy
Supplementary material
Supplementary material is available at Cardiovascular Research online
AcknowledgementsThe authors wish to thank Drs Keren Bitton-Worms and Ran Rostokerand Prof Derek LeRoith from the Rambam Medical Center for their
assistance in GTT and ITT Dr Edith Suss Toby Maya Holdengreber andLior Liba from the BCF Bioimaging Center (Faculty of MedicineTechnion) for their assistance with imaging and image analysis Prof DEMichelle from University of Michigan for cardiomyocytes culture techni-que and Prof Yehiel Zick from the Weizmann institute of Sciences forcritical comments
Conflict of interest none declared
FundingThis work was partially supported by the United States-Israel BinationalGrant 2009179 to AA and TH In addition we wish to thank the MarsPittsburgh Fund for Medical Research to AA the Department of Defensegrant W81XWH-14-1-0179 to TH and to D Dan and Betty Kahn travelgrant support to LK
References
1 Roger VL Go AS Lloyd-Jones DM Benjamin EJ Berry JD Borden WB Bravata DMDai S Ford ES Fox CS Fullerton HJ Gillespie C Hailpern SM Heit JA Howard VJKissela BM Kittner SJ Lackland DT Lichtman JH Lisabeth LD Makuc DM MarcusGM Marelli A Matchar DB Moy CS Mozaffarian D Mussolino ME Nichol GPaynter NP Soliman EZ Sorlie PD Sotoodehnia N Turan TN Virani SS Wong NDWoo D Turner MB American Heart Association Statistics C Stroke Statistics SHeart disease and stroke statisticsndash2012 update a report from the American HeartAssociation Circulation 2012125e2ndashe220
2 Wilson C Diabetes ACCORD 5-year outcomes of intensive glycemic control NatRev Endocrinol 20117314
3 Ernande L Derumeaux G Diabetic cardiomyopathy myth or reality Arch CardiovascDis 2012105218ndash225
4 Battiprolu PK Hojayev B Jiang N Wang ZV Luo X Iglewski M Shelton JM GerardRD Rothermel BA Gillette TG Lavandero S Hill JA Metabolic stress-induced
InsulinGF
Cytokines
FFA
GlucoseATF3
Cardiac
Remodeling
Factors
Paracrine action
Collagens
and ECM
proteins
Fibrosis
Cardiac
Dysfunction
Systemic Effects
Glucose Tolerance
TNFα
IL-6
Cardiomyocyte
ATF3
Homeostasis
Homeostasis
TNFα
IL-6
TIMP1
BNP
ANP
Figure 8 lsquoPeripheral-to-cardiacrsquo and lsquoCardiac-to-peripheralrsquo model scheme Schematic diagram showing the ATF3 adaptive role in maintaining hearthomeostasis following HFD-induced T2D In contrast stressful environment under HFD leads to maladaptive cardiac response in the absence of ATF3Cardiac remodeling caused by ATF3 deficiency induces an increase in cardiac ECM remodeling and inflammatory genes and mediates the communication ofcardiomyocytes with heart resident cells potentially mediated by IL-6 TNFa ANP BNP and TIMP1 In this situation increased cardiac fibrosis and inflamma-tory response worsen cardiac function The inflamed failing heart mediates the communication of cardiomyocytes with distant organs as well thereby affect-ing diabetic outcome and glucose homeostasis
ATF3 expression in cardiomyocytes preserves homeostasis 145D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
activation of FoxO1 triggers diabetic cardiomyopathy in mice J Clin Invest20121221109ndash1118
5 Mandavia CH Aroor AR Demarco VG Sowers JR Molecular and metabolic mecha-nisms of cardiac dysfunction in diabetes Life Sci 201392601ndash608
6 Ventura-Clapier R Garnier A Veksler V Energy metabolism in heart failure J Physiol20045551ndash13
7 Guo S Insulin signaling resistance and the metabolic syndrome insights from mousemodels into disease mechanisms J Endocrinol 2014220T1ndashT23
8 Montane J Cadavez L Novials A Stress and the inflammatory process a major causeof pancreatic cell death in type 2 diabetes Diabetes Metab Syndr Obes 2014725ndash34
9 Ashrafian H Frenneaux MP Opie LH Metabolic mechanisms in heart failureCirculation 2007116434ndash448
10 Tanabe K Liu Y Hasan SD Martinez SC Cras-Meneur C Welling CM Bernal-MizrachiE Tanizawa Y Rhodes CJ Zmuda E Hai T Abumrad NA Permutt MA Glucose andfatty acids synergize to promote B-cell apoptosis through activation of glycogen syn-thase kinase 3beta independent of JNK activation PLoS One 20116e18146
11 Heineke J Molkentin JD Regulation of cardiac hypertrophy by intracellular signallingpathways Nat Rev Mol Cell Biol 20067589ndash600
12 Hai T Wolford CC Chang YS ATF3 a hub of the cellular adaptive-response net-work in the pathogenesis of diseases is modulation of inflammation a unifying com-ponent Gene Expr 2010151ndash11
13 Takii R Inouye S Fujimoto M Nakamura T Shinkawa T Prakasam R Tan K HayashidaN Ichikawa H Hai T Nakai A Heat shock transcription factor 1 inhibits expression ofIL-6 through activating transcription factor 3 J Immunol 20101841041ndash1048
14 Giraldo A Barrett OP Tindall MJ Fuller SJ Amirak E Bhattacharya BS Sugden PHClerk A Feedback regulation by Atf3 in the endothelin-1-responsive transcriptomeof cardiomyocytes Egr1 is a principal Atf3 target Biochem J 2012444343ndash355
15 Hasin T Elhanani O Abassi Z Hai T Aronheim A Angiotensin II signaling up-regulates the immediate early transcription factor ATF3 in the left but not the rightatrium Basic Res Cardiol 2011106175ndash187
16 Koren L Elhanani O Kehat I Hai T Aronheim A Adult cardiac expression of theactivating transcription factor 3 ATF3 promotes ventricular hypertrophy PLoS One20138e68396
17 Koren L Alishekevitz D Elhanani O Nevelsky A Hai T Kehat I Shaked Y AronheimA ATF3-dependent cross-talk between cardiomyocytes and macrophages promotescardiac maladaptive remodeling Int J Cardiol 2015198232ndash240
18 Okamoto Y Chaves A Chen J Kelley R Jones K Weed HG Gardner KL Gangi LYamaguchi M Klomkleaw W Nakayama T Hamlin RL Carnes C Altschuld R BauerJ Hai T Transgenic mice with cardiac-specific expression of activating transcriptionfactor 3 a stress-inducible gene have conduction abnormalities and contractile dys-function Am J Pathol 2001159639ndash650
19 Zhou H Shen DF Bian ZY Zong J Deng W Zhang Y Guo YY Li H Tang QZActivating transcription factor 3 deficiency promotes cardiac hypertrophy dysfunc-tion and fibrosis induced by pressure overload PLoS One 20116e26744
20 Zhou H Guo H Zong J Dai J Yuan Y Bian ZY Tang QZ ATF3 regulates multipletargets and may play a dual role in cardiac hypertrophy and injury Int J Cardiol2014174838ndash839
21 Yang J Yang CJ Yang J Fan ZX ATF3 a promotion effect or a inhibition effect in car-diac maladaptive remodeling Int J Cardiol 2015201245ndash246
22 Koren L Shaked Y Aronheim A Response letter ldquoATF3 a promoter or inhibitor ofcardiac maladaptive remodelingrdquo Int J Cardiol 2015201692
23 Ma ZG Wei WY Xu SC Zhang WB Dai J Tang QZ ATF3 a potential target forcardiac maladaptive remodeling Int J Cardiol 201620250ndash51
24 Zhou H Yuan Y Ni J Guo H Deng W Bian ZY Tang QZ Pleiotropic and puzzlingeffects of ATF3 in maladaptive cardiac remodeling Int J Cardiol 201620687ndash88
25 Zmuda EJ Qi L Zhu MX Mirmira RG Montminy MR Hai T The roles of ATF3 anadaptive-response gene in high-fat-diet-induced diabetes and pancreatic beta-celldysfunction Mol Endocrinol 2010241423ndash1433
26 Lu D Chen J Hai T The regulation of ATF3 gene expression by mitogen-activatedprotein kinases Biochem J 2007401559ndash567
27 Hartman MG Lu D Kim ML Kociba GJ Shukri T Buteau J Wang X Frankel WLGuttridge D Prentki M Grey ST Ron D Hai T Role for activating transcription fac-tor 3 in stress-induced beta-cell apoptosis Mol Cell Biol 2004245721ndash5732
28 Wolford CC McConoughey SJ Jalgaonkar SP Leon M Merchant AS Dominick JLYin X Chang Y Zmuda EJ OrsquoToole SA Millar EK Roller SL Shapiro CL OstrowskiMC Sutherland RL Hai T Transcription factor ATF3 links host adaptive response tobreast cancer metastasis J Clin Iinvest 20131232893ndash2906
29 Agah R Frenkel PA French BA Michael LH Overbeek PA Schneider MD Generecombination in postmitotic cells Targeted expression of Cre recombinase pro-vokes cardiac-restricted site-specific rearrangement in adult ventricular musclein vivo J Clin Invest 1997100169ndash179
30 Livet J Weissman TA Kang H Draft RW Lu J Bennis RA Sanes JR Lichtman JWTransgenic strategies for combinatorial expression of fluorescent proteins in thenervous system Nature 200745056ndash62
31 Davis J Maillet M Miano JM Molkentin JD Lost in transgenesis a userrsquos guidefor genetically manipulating the mouse in cardiac research Circ Res2012111761ndash777
32 Heiberg E Sjogren J Ugander M Carlsson M Engblom H Arheden H Design andvalidation of segment ndash freely available software for cardiovascular image analysisBMC Med Imaging 2010101
33 Kabaeva Z Zhao M Michele DE Blebbistatin extends culture life of adult mouse car-diac myocytes and allows efficient and stable transgene expression Am J Physiol HeartC 2008294H1667ndash1674
34 Subramaniam A Jones WK Gulick J Wert S Neumann J Robbins J Tissue-specificregulation of the alpha-myosin heavy chain gene promoter in transgenic mice J BiolChem 199126624613ndash24620
35 Cunha DA Hekerman P Ladriere L Bazarra-Castro A Ortis F Wakeham MCMoore F Rasschaert J Cardozo AK Bellomo E Overbergh L Mathieu C Lupi R HaiT Herchuelz A Marchetti P Rutter GA Eizirik DL Cnop M Initiation and executionof lipotoxic ER stress in pancreatic beta-cells J Cell Sci 20081212308ndash2318
36 Wynn TA Ramalingam TR Mechanisms of fibrosis therapeutic translation forfibrotic disease Nat Med 2012181028ndash1040
37 Li D Yin X Zmuda EJ Wolford CC Dong X White MF Hai T The repression ofIRS2 gene by ATF3 a stress-inducible gene contributes to pancreatic beta-cell apop-tosis Diabetes 200857635ndash644
38 Myerson SG Bellenger NG Pennell DJ Assessment of left ventricular mass by cardi-ovascular magnetic resonance Hypertension 200239750ndash755
39 Zmuda EJ Viapiano M Grey ST Hadley G Garcia-Ocana A Hai T Deficiency ofAtf3 an adaptive-response gene protects islets and ameliorates inflammation in asyngeneic mouse transplantation model Diabetologia 2010531438ndash1450
40 Riehle C Abel ED Insulin signaling and heart failure Circ Res 20161181151ndash116941 Schreyer SA Wilson DL LeBoeuf RC C57BL6 mice fed high fat diets as models for
diabetes-accelerated atherosclerosis Atherosclerosis 199813617ndash2442 Brainard RE Watson LJ Demartino AM Brittian KR Readnower RD Boakye AA
Zhang D Hoetker JD Bhatnagar A Baba SP Jones SP High fat feeding in mice isinsufficient to induce cardiac dysfunction and does not exacerbate heart failure PLoSOne 20138e83174
43 Gupte AA Minze LJ Reyes M Ren Y Wang X Brunner G Ghosn M Cordero-Reyes AM Ding K Pratico D Morrisett J Shi ZZ Hamilton DJ Lyon CJ Hsueh WAHigh-fat feeding-induced hyperinsulinemia increases cardiac glucose uptake and mito-chondrial function despite peripheral insulin resistance Endocrinology20131542650ndash2662
44 Logan SK Garabedian MJ Campbell CE Werb Z Synergistic transcriptionalactivation of the tissue inhibitor of metalloproteinases-1 promoter via functionalinteraction of AP-1 and Ets-1 transcription factors J Biol Chem1996271774ndash782
45 Belke DD Betuing S Tuttle MJ Graveleau C Young ME Pham M Zhang DCooksey RC McClain DA Litwin SE Taegtmeyer H Severson D Kahn CR AbelED Insulin signaling coordinately regulates cardiac size metabolism and contractileprotein isoform expression J Clin Invest 2002109629ndash639
46 Fernandez AM Kim JK Yakar S Dupont J Hernandez-Sanchez C Castle AL FilmoreJ Shulman GI Le Roith D Functional inactivation of the IGF-I and insulin receptors inskeletal muscle causes type 2 diabetes Genes Dev 2001151926ndash1934
47 Gilchrist M Thorsson V Li B Rust AG Korb M Roach JC Kennedy K Hai TBolouri H Aderem A Systems biology approaches identify ATF3 as a negative regu-lator of Toll-like receptor 4 Nature 2006441173ndash178
48 Kristiansen OP Mandrup-Poulsen T Interleukin-6 and diabetes the good the bad orthe indifferent Diabetes 200554(Suppl 2)S114ndashS124
49 Grueter CE van Rooij E Johnson BA DeLeon SM Sutherland LB Qi X Gautron LElmquist JK Bassel-Duby R Olson EN A cardiac microRNA governs systemic energyhomeostasis by regulation of MED13 Cell 2012149671ndash683
50 Baskin KK Grueter CE Kusminski CM Holland WL Bookout AL Satapati S KongYM Burgess SC Malloy CR Scherer PE Newgard CB Bassel-Duby R Olson ENMED13-dependent signaling from the heart confers leanness by enhancing metabo-lism in adipose tissue and liver EMBO Mol Med 201461610ndash1621
51 Nakamura M Sadoshima J Heart over mind metabolic control of white adipose tis-sue and liver EMBO Mol Med 201461521ndash1524
52 Tenenbaum A Fisman EZ Impaired glucose metabolism in patients with heart failurepathophysiology and possible treatment strategies Am J Cardiovasc Drugs20044269ndash280
53 Suskin N McKelvie RS Burns RJ Latini R Pericak D Probstfield J Rouleau JL SigouinC Solymoss CB Tsuyuki R White M Yusuf S Glucose and insulin abnormalitiesrelate to functional capacity in patients with congestive heart failure Eur Heart J2000211368ndash1375
54 Aronson D Hammerman H Kapeliovich MR Suleiman A Agmon Y Beyar RMarkiewicz W Suleiman M Fasting glucose in acute myocardial infarction incremen-tal value for long-term mortality and relationship with left ventricular systolic func-tion Diabetes Care 200730960ndash966
146 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
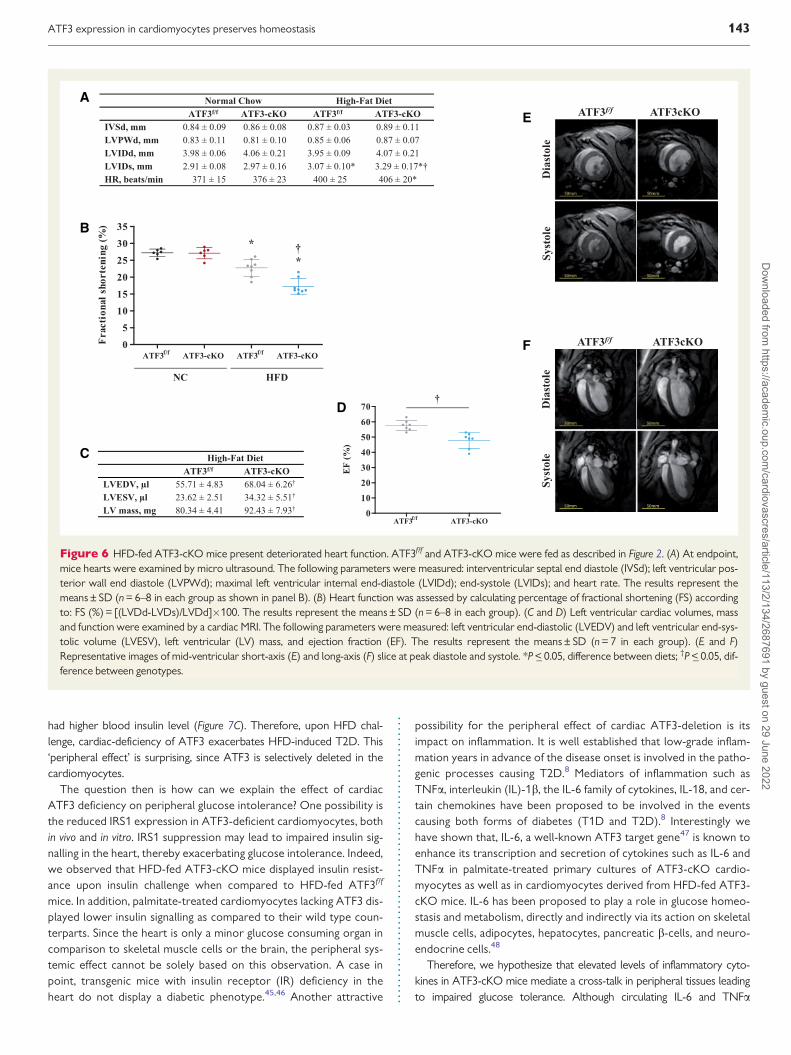
had higher blood insulin level (Figure 7C) Therefore upon HFD chal-lenge cardiac-deficiency of ATF3 exacerbates HFD-induced T2D Thislsquoperipheral effectrsquo is surprising since ATF3 is selectively deleted in thecardiomyocytes
The question then is how can we explain the effect of cardiacATF3 deficiency on peripheral glucose intolerance One possibility isthe reduced IRS1 expression in ATF3-deficient cardiomyocytes bothin vivo and in vitro IRS1 suppression may lead to impaired insulin sig-nalling in the heart thereby exacerbating glucose intolerance Indeedwe observed that HFD-fed ATF3-cKO mice displayed insulin resist-ance upon insulin challenge when compared to HFD-fed ATF3ff
mice In addition palmitate-treated cardiomyocytes lacking ATF3 dis-played lower insulin signalling as compared to their wild type coun-terparts Since the heart is only a minor glucose consuming organ incomparison to skeletal muscle cells or the brain the peripheral sys-temic effect cannot be solely based on this observation A case inpoint transgenic mice with insulin receptor (IR) deficiency in theheart do not display a diabetic phenotype4546 Another attractive
possibility for the peripheral effect of cardiac ATF3-deletion is itsimpact on inflammation It is well established that low-grade inflam-mation years in advance of the disease onset is involved in the patho-genic processes causing T2D8 Mediators of inflammation such asTNFa interleukin (IL)-1b the IL-6 family of cytokines IL-18 and cer-tain chemokines have been proposed to be involved in the eventscausing both forms of diabetes (T1D and T2D)8 Interestingly wehave shown that IL-6 a well-known ATF3 target gene47 is known toenhance its transcription and secretion of cytokines such as IL-6 andTNFa in palmitate-treated primary cultures of ATF3-cKO cardio-myocytes as well as in cardiomyocytes derived from HFD-fed ATF3-cKO mice IL-6 has been proposed to play a role in glucose homeo-stasis and metabolism directly and indirectly via its action on skeletalmuscle cells adipocytes hepatocytes pancreatic b-cells and neuro-endocrine cells48
Therefore we hypothesize that elevated levels of inflammatory cyto-kines in ATF3-cKO mice mediate a cross-talk in peripheral tissues leadingto impaired glucose tolerance Although circulating IL-6 and TNFa
A
B
C
D
E
F
Figure 6 HFD-fed ATF3-cKO mice present deteriorated heart function ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) At endpointmice hearts were examined by micro ultrasound The following parameters were measured interventricular septal end diastole (IVSd) left ventricular pos-terior wall end diastole (LVPWd) maximal left ventricular internal end-diastole (LVIDd) end-systole (LVIDs) and heart rate The results represent themeansplusmn SD (n = 6ndash8 in each group as shown in panel B) (B) Heart function was assessed by calculating percentage of fractional shortening (FS) accordingto FS () = [(LVDd-LVDs)LVDd]100 The results represent the meansplusmn SD (n = 6ndash8 in each group) (C and D) Left ventricular cardiac volumes massand function were examined by a cardiac MRI The following parameters were measured left ventricular end-diastolic (LVEDV) and left ventricular end-sys-tolic volume (LVESV) left ventricular (LV) mass and ejection fraction (EF) The results represent the meansplusmn SD (n = 7 in each group) (E and F)Representative images of mid-ventricular short-axis (E) and long-axis (F) slice at peak diastole and systole P lt_ 005 difference between diets daggerP lt_ 005 dif-ference between genotypes
ATF3 expression in cardiomyocytes preserves homeostasis 143D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
cytokines may be derived from different tissues we provide ample evi-dence that ATF3-defficient cardiomyocytes produce and secrete higherlevels of these cytokines as compared to their wild type counterpartsThis indicates a lsquocardiac-to-peripheralrsquo effect in T2D-induced cardiomyop-athy This is a reverse direction from the lsquoperipheral-to-cardiacrsquo effect dis-cussed in the introduction where inflammation in the peripheral tissuesleads to cardiac dysfunction Since inflammatory cytokines are soluble
factors they can exert a systemic effect This is why it makes sense thattheir effect can go both ways Thus diabetes and cardiomyopathy are apart of a vicious cycle Previously it was shown that cardiac specific dele-tion of the thyroid hormone complex mediator MED13 results in aperipheral effect upon HFD Cardiac MED13 deficiency suppresses meta-bolic genes in the heart which leads to a decrease in energy expenditureand an obese phenotype most likely by the secretion of cardiokines that
A
B
C
D
E
Figure 7 HFD-fed ATF3-cKO mice exhibit impaired glucose tolerance ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) Fasting (6 h)blood glucose concentrations (mgdl) The results represent the meansplusmn SD (n = 20ndash30group) P lt_ 005 difference between diets daggerP lt_ 005 differencebetween genotypes (B) Glucose tolerance test (GTT) Values are presented as relative to initial valueplusmn SD (n = 20ndash25group) P lt_ 005 difference betweendiets daggerP lt_ 005 difference between genotypes (C) Serum insulin concentrations were quantified by ELISA Values are presented as relative values (com-pared to NC-fed ATF3ff mice determined as 1 n = 15ndash20group) P lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes (D and E)Effect of HFD on phosphorylated Akt (Ser473 pAkt) to Akt ratio levels in heart lysate following either saline or insulin injection (10 U1 kg) is shownRepresentative blots (D) Densitometric analysis is presented as means ratio (n = 3ndash6group) pAktAkt ratio is presented as relative values (compared toHFD-fed saline injected ATF3ff mice determined as 1) P lt_ 005 insulin vs saline treatment daggerP lt_ 005 difference between genotypes (E)
144 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
mediates the cross talk between the heart and peripheral tissues49ndash51
Here we also observed increased levels of natriuretic peptides (ANP andBNP) which together with IL-6 and TNFa may mediate the cross-talk withthe peripheral tissues to induce glucose intolerance
Does this finding have any relevance to cardiac disease in humansSeveral lines of evidence suggests that congestive heart failure in patientsis associated with an increase in blood glucose levels52 In addition highblood glucose levels in non-diabetic patients with acute myocardialinfarction serves as a prognostic predictor of long-term mortality5354
Thus here we provide evidence that high blood glucose levels are a partof the symptoms of cardiac remodeling process
Collectively our findings suggest that cardiac ATF3 expression isresponsible for an adaptive cardiac response to the stressful environmentunder a HFD (Figure 8) ATF3 dampens the inflammatory response andsuppresses the expression of ECM remodeling genes and thus amelioratescardiac function In HFD-fed mice with ATF3 deficiency or in mice withcardiac remodeling the inflamed heart affects also peripheral tissuesthereby worsening T2D in peripheral tissues resulting in elevated bloodglucose level and turning on a vicious cycle to worsen cardiomyopathy
Supplementary material
Supplementary material is available at Cardiovascular Research online
AcknowledgementsThe authors wish to thank Drs Keren Bitton-Worms and Ran Rostokerand Prof Derek LeRoith from the Rambam Medical Center for their
assistance in GTT and ITT Dr Edith Suss Toby Maya Holdengreber andLior Liba from the BCF Bioimaging Center (Faculty of MedicineTechnion) for their assistance with imaging and image analysis Prof DEMichelle from University of Michigan for cardiomyocytes culture techni-que and Prof Yehiel Zick from the Weizmann institute of Sciences forcritical comments
Conflict of interest none declared
FundingThis work was partially supported by the United States-Israel BinationalGrant 2009179 to AA and TH In addition we wish to thank the MarsPittsburgh Fund for Medical Research to AA the Department of Defensegrant W81XWH-14-1-0179 to TH and to D Dan and Betty Kahn travelgrant support to LK
References
1 Roger VL Go AS Lloyd-Jones DM Benjamin EJ Berry JD Borden WB Bravata DMDai S Ford ES Fox CS Fullerton HJ Gillespie C Hailpern SM Heit JA Howard VJKissela BM Kittner SJ Lackland DT Lichtman JH Lisabeth LD Makuc DM MarcusGM Marelli A Matchar DB Moy CS Mozaffarian D Mussolino ME Nichol GPaynter NP Soliman EZ Sorlie PD Sotoodehnia N Turan TN Virani SS Wong NDWoo D Turner MB American Heart Association Statistics C Stroke Statistics SHeart disease and stroke statisticsndash2012 update a report from the American HeartAssociation Circulation 2012125e2ndashe220
2 Wilson C Diabetes ACCORD 5-year outcomes of intensive glycemic control NatRev Endocrinol 20117314
3 Ernande L Derumeaux G Diabetic cardiomyopathy myth or reality Arch CardiovascDis 2012105218ndash225
4 Battiprolu PK Hojayev B Jiang N Wang ZV Luo X Iglewski M Shelton JM GerardRD Rothermel BA Gillette TG Lavandero S Hill JA Metabolic stress-induced
InsulinGF
Cytokines
FFA
GlucoseATF3
Cardiac
Remodeling
Factors
Paracrine action
Collagens
and ECM
proteins
Fibrosis
Cardiac
Dysfunction
Systemic Effects
Glucose Tolerance
TNFα
IL-6
Cardiomyocyte
ATF3
Homeostasis
Homeostasis
TNFα
IL-6
TIMP1
BNP
ANP
Figure 8 lsquoPeripheral-to-cardiacrsquo and lsquoCardiac-to-peripheralrsquo model scheme Schematic diagram showing the ATF3 adaptive role in maintaining hearthomeostasis following HFD-induced T2D In contrast stressful environment under HFD leads to maladaptive cardiac response in the absence of ATF3Cardiac remodeling caused by ATF3 deficiency induces an increase in cardiac ECM remodeling and inflammatory genes and mediates the communication ofcardiomyocytes with heart resident cells potentially mediated by IL-6 TNFa ANP BNP and TIMP1 In this situation increased cardiac fibrosis and inflamma-tory response worsen cardiac function The inflamed failing heart mediates the communication of cardiomyocytes with distant organs as well thereby affect-ing diabetic outcome and glucose homeostasis
ATF3 expression in cardiomyocytes preserves homeostasis 145D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
activation of FoxO1 triggers diabetic cardiomyopathy in mice J Clin Invest20121221109ndash1118
5 Mandavia CH Aroor AR Demarco VG Sowers JR Molecular and metabolic mecha-nisms of cardiac dysfunction in diabetes Life Sci 201392601ndash608
6 Ventura-Clapier R Garnier A Veksler V Energy metabolism in heart failure J Physiol20045551ndash13
7 Guo S Insulin signaling resistance and the metabolic syndrome insights from mousemodels into disease mechanisms J Endocrinol 2014220T1ndashT23
8 Montane J Cadavez L Novials A Stress and the inflammatory process a major causeof pancreatic cell death in type 2 diabetes Diabetes Metab Syndr Obes 2014725ndash34
9 Ashrafian H Frenneaux MP Opie LH Metabolic mechanisms in heart failureCirculation 2007116434ndash448
10 Tanabe K Liu Y Hasan SD Martinez SC Cras-Meneur C Welling CM Bernal-MizrachiE Tanizawa Y Rhodes CJ Zmuda E Hai T Abumrad NA Permutt MA Glucose andfatty acids synergize to promote B-cell apoptosis through activation of glycogen syn-thase kinase 3beta independent of JNK activation PLoS One 20116e18146
11 Heineke J Molkentin JD Regulation of cardiac hypertrophy by intracellular signallingpathways Nat Rev Mol Cell Biol 20067589ndash600
12 Hai T Wolford CC Chang YS ATF3 a hub of the cellular adaptive-response net-work in the pathogenesis of diseases is modulation of inflammation a unifying com-ponent Gene Expr 2010151ndash11
13 Takii R Inouye S Fujimoto M Nakamura T Shinkawa T Prakasam R Tan K HayashidaN Ichikawa H Hai T Nakai A Heat shock transcription factor 1 inhibits expression ofIL-6 through activating transcription factor 3 J Immunol 20101841041ndash1048
14 Giraldo A Barrett OP Tindall MJ Fuller SJ Amirak E Bhattacharya BS Sugden PHClerk A Feedback regulation by Atf3 in the endothelin-1-responsive transcriptomeof cardiomyocytes Egr1 is a principal Atf3 target Biochem J 2012444343ndash355
15 Hasin T Elhanani O Abassi Z Hai T Aronheim A Angiotensin II signaling up-regulates the immediate early transcription factor ATF3 in the left but not the rightatrium Basic Res Cardiol 2011106175ndash187
16 Koren L Elhanani O Kehat I Hai T Aronheim A Adult cardiac expression of theactivating transcription factor 3 ATF3 promotes ventricular hypertrophy PLoS One20138e68396
17 Koren L Alishekevitz D Elhanani O Nevelsky A Hai T Kehat I Shaked Y AronheimA ATF3-dependent cross-talk between cardiomyocytes and macrophages promotescardiac maladaptive remodeling Int J Cardiol 2015198232ndash240
18 Okamoto Y Chaves A Chen J Kelley R Jones K Weed HG Gardner KL Gangi LYamaguchi M Klomkleaw W Nakayama T Hamlin RL Carnes C Altschuld R BauerJ Hai T Transgenic mice with cardiac-specific expression of activating transcriptionfactor 3 a stress-inducible gene have conduction abnormalities and contractile dys-function Am J Pathol 2001159639ndash650
19 Zhou H Shen DF Bian ZY Zong J Deng W Zhang Y Guo YY Li H Tang QZActivating transcription factor 3 deficiency promotes cardiac hypertrophy dysfunc-tion and fibrosis induced by pressure overload PLoS One 20116e26744
20 Zhou H Guo H Zong J Dai J Yuan Y Bian ZY Tang QZ ATF3 regulates multipletargets and may play a dual role in cardiac hypertrophy and injury Int J Cardiol2014174838ndash839
21 Yang J Yang CJ Yang J Fan ZX ATF3 a promotion effect or a inhibition effect in car-diac maladaptive remodeling Int J Cardiol 2015201245ndash246
22 Koren L Shaked Y Aronheim A Response letter ldquoATF3 a promoter or inhibitor ofcardiac maladaptive remodelingrdquo Int J Cardiol 2015201692
23 Ma ZG Wei WY Xu SC Zhang WB Dai J Tang QZ ATF3 a potential target forcardiac maladaptive remodeling Int J Cardiol 201620250ndash51
24 Zhou H Yuan Y Ni J Guo H Deng W Bian ZY Tang QZ Pleiotropic and puzzlingeffects of ATF3 in maladaptive cardiac remodeling Int J Cardiol 201620687ndash88
25 Zmuda EJ Qi L Zhu MX Mirmira RG Montminy MR Hai T The roles of ATF3 anadaptive-response gene in high-fat-diet-induced diabetes and pancreatic beta-celldysfunction Mol Endocrinol 2010241423ndash1433
26 Lu D Chen J Hai T The regulation of ATF3 gene expression by mitogen-activatedprotein kinases Biochem J 2007401559ndash567
27 Hartman MG Lu D Kim ML Kociba GJ Shukri T Buteau J Wang X Frankel WLGuttridge D Prentki M Grey ST Ron D Hai T Role for activating transcription fac-tor 3 in stress-induced beta-cell apoptosis Mol Cell Biol 2004245721ndash5732
28 Wolford CC McConoughey SJ Jalgaonkar SP Leon M Merchant AS Dominick JLYin X Chang Y Zmuda EJ OrsquoToole SA Millar EK Roller SL Shapiro CL OstrowskiMC Sutherland RL Hai T Transcription factor ATF3 links host adaptive response tobreast cancer metastasis J Clin Iinvest 20131232893ndash2906
29 Agah R Frenkel PA French BA Michael LH Overbeek PA Schneider MD Generecombination in postmitotic cells Targeted expression of Cre recombinase pro-vokes cardiac-restricted site-specific rearrangement in adult ventricular musclein vivo J Clin Invest 1997100169ndash179
30 Livet J Weissman TA Kang H Draft RW Lu J Bennis RA Sanes JR Lichtman JWTransgenic strategies for combinatorial expression of fluorescent proteins in thenervous system Nature 200745056ndash62
31 Davis J Maillet M Miano JM Molkentin JD Lost in transgenesis a userrsquos guidefor genetically manipulating the mouse in cardiac research Circ Res2012111761ndash777
32 Heiberg E Sjogren J Ugander M Carlsson M Engblom H Arheden H Design andvalidation of segment ndash freely available software for cardiovascular image analysisBMC Med Imaging 2010101
33 Kabaeva Z Zhao M Michele DE Blebbistatin extends culture life of adult mouse car-diac myocytes and allows efficient and stable transgene expression Am J Physiol HeartC 2008294H1667ndash1674
34 Subramaniam A Jones WK Gulick J Wert S Neumann J Robbins J Tissue-specificregulation of the alpha-myosin heavy chain gene promoter in transgenic mice J BiolChem 199126624613ndash24620
35 Cunha DA Hekerman P Ladriere L Bazarra-Castro A Ortis F Wakeham MCMoore F Rasschaert J Cardozo AK Bellomo E Overbergh L Mathieu C Lupi R HaiT Herchuelz A Marchetti P Rutter GA Eizirik DL Cnop M Initiation and executionof lipotoxic ER stress in pancreatic beta-cells J Cell Sci 20081212308ndash2318
36 Wynn TA Ramalingam TR Mechanisms of fibrosis therapeutic translation forfibrotic disease Nat Med 2012181028ndash1040
37 Li D Yin X Zmuda EJ Wolford CC Dong X White MF Hai T The repression ofIRS2 gene by ATF3 a stress-inducible gene contributes to pancreatic beta-cell apop-tosis Diabetes 200857635ndash644
38 Myerson SG Bellenger NG Pennell DJ Assessment of left ventricular mass by cardi-ovascular magnetic resonance Hypertension 200239750ndash755
39 Zmuda EJ Viapiano M Grey ST Hadley G Garcia-Ocana A Hai T Deficiency ofAtf3 an adaptive-response gene protects islets and ameliorates inflammation in asyngeneic mouse transplantation model Diabetologia 2010531438ndash1450
40 Riehle C Abel ED Insulin signaling and heart failure Circ Res 20161181151ndash116941 Schreyer SA Wilson DL LeBoeuf RC C57BL6 mice fed high fat diets as models for
diabetes-accelerated atherosclerosis Atherosclerosis 199813617ndash2442 Brainard RE Watson LJ Demartino AM Brittian KR Readnower RD Boakye AA
Zhang D Hoetker JD Bhatnagar A Baba SP Jones SP High fat feeding in mice isinsufficient to induce cardiac dysfunction and does not exacerbate heart failure PLoSOne 20138e83174
43 Gupte AA Minze LJ Reyes M Ren Y Wang X Brunner G Ghosn M Cordero-Reyes AM Ding K Pratico D Morrisett J Shi ZZ Hamilton DJ Lyon CJ Hsueh WAHigh-fat feeding-induced hyperinsulinemia increases cardiac glucose uptake and mito-chondrial function despite peripheral insulin resistance Endocrinology20131542650ndash2662
44 Logan SK Garabedian MJ Campbell CE Werb Z Synergistic transcriptionalactivation of the tissue inhibitor of metalloproteinases-1 promoter via functionalinteraction of AP-1 and Ets-1 transcription factors J Biol Chem1996271774ndash782
45 Belke DD Betuing S Tuttle MJ Graveleau C Young ME Pham M Zhang DCooksey RC McClain DA Litwin SE Taegtmeyer H Severson D Kahn CR AbelED Insulin signaling coordinately regulates cardiac size metabolism and contractileprotein isoform expression J Clin Invest 2002109629ndash639
46 Fernandez AM Kim JK Yakar S Dupont J Hernandez-Sanchez C Castle AL FilmoreJ Shulman GI Le Roith D Functional inactivation of the IGF-I and insulin receptors inskeletal muscle causes type 2 diabetes Genes Dev 2001151926ndash1934
47 Gilchrist M Thorsson V Li B Rust AG Korb M Roach JC Kennedy K Hai TBolouri H Aderem A Systems biology approaches identify ATF3 as a negative regu-lator of Toll-like receptor 4 Nature 2006441173ndash178
48 Kristiansen OP Mandrup-Poulsen T Interleukin-6 and diabetes the good the bad orthe indifferent Diabetes 200554(Suppl 2)S114ndashS124
49 Grueter CE van Rooij E Johnson BA DeLeon SM Sutherland LB Qi X Gautron LElmquist JK Bassel-Duby R Olson EN A cardiac microRNA governs systemic energyhomeostasis by regulation of MED13 Cell 2012149671ndash683
50 Baskin KK Grueter CE Kusminski CM Holland WL Bookout AL Satapati S KongYM Burgess SC Malloy CR Scherer PE Newgard CB Bassel-Duby R Olson ENMED13-dependent signaling from the heart confers leanness by enhancing metabo-lism in adipose tissue and liver EMBO Mol Med 201461610ndash1621
51 Nakamura M Sadoshima J Heart over mind metabolic control of white adipose tis-sue and liver EMBO Mol Med 201461521ndash1524
52 Tenenbaum A Fisman EZ Impaired glucose metabolism in patients with heart failurepathophysiology and possible treatment strategies Am J Cardiovasc Drugs20044269ndash280
53 Suskin N McKelvie RS Burns RJ Latini R Pericak D Probstfield J Rouleau JL SigouinC Solymoss CB Tsuyuki R White M Yusuf S Glucose and insulin abnormalitiesrelate to functional capacity in patients with congestive heart failure Eur Heart J2000211368ndash1375
54 Aronson D Hammerman H Kapeliovich MR Suleiman A Agmon Y Beyar RMarkiewicz W Suleiman M Fasting glucose in acute myocardial infarction incremen-tal value for long-term mortality and relationship with left ventricular systolic func-tion Diabetes Care 200730960ndash966
146 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
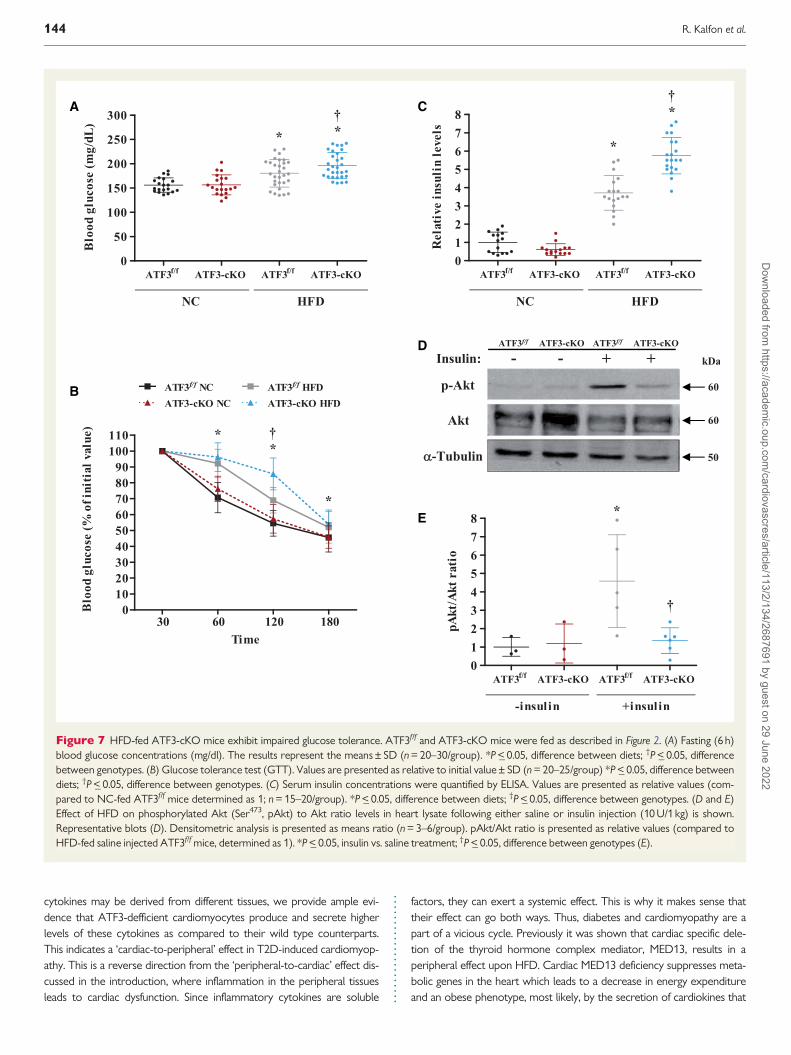
cytokines may be derived from different tissues we provide ample evi-dence that ATF3-defficient cardiomyocytes produce and secrete higherlevels of these cytokines as compared to their wild type counterpartsThis indicates a lsquocardiac-to-peripheralrsquo effect in T2D-induced cardiomyop-athy This is a reverse direction from the lsquoperipheral-to-cardiacrsquo effect dis-cussed in the introduction where inflammation in the peripheral tissuesleads to cardiac dysfunction Since inflammatory cytokines are soluble
factors they can exert a systemic effect This is why it makes sense thattheir effect can go both ways Thus diabetes and cardiomyopathy are apart of a vicious cycle Previously it was shown that cardiac specific dele-tion of the thyroid hormone complex mediator MED13 results in aperipheral effect upon HFD Cardiac MED13 deficiency suppresses meta-bolic genes in the heart which leads to a decrease in energy expenditureand an obese phenotype most likely by the secretion of cardiokines that
A
B
C
D
E
Figure 7 HFD-fed ATF3-cKO mice exhibit impaired glucose tolerance ATF3ff and ATF3-cKO mice were fed as described in Figure 2 (A) Fasting (6 h)blood glucose concentrations (mgdl) The results represent the meansplusmn SD (n = 20ndash30group) P lt_ 005 difference between diets daggerP lt_ 005 differencebetween genotypes (B) Glucose tolerance test (GTT) Values are presented as relative to initial valueplusmn SD (n = 20ndash25group) P lt_ 005 difference betweendiets daggerP lt_ 005 difference between genotypes (C) Serum insulin concentrations were quantified by ELISA Values are presented as relative values (com-pared to NC-fed ATF3ff mice determined as 1 n = 15ndash20group) P lt_ 005 difference between diets daggerP lt_ 005 difference between genotypes (D and E)Effect of HFD on phosphorylated Akt (Ser473 pAkt) to Akt ratio levels in heart lysate following either saline or insulin injection (10 U1 kg) is shownRepresentative blots (D) Densitometric analysis is presented as means ratio (n = 3ndash6group) pAktAkt ratio is presented as relative values (compared toHFD-fed saline injected ATF3ff mice determined as 1) P lt_ 005 insulin vs saline treatment daggerP lt_ 005 difference between genotypes (E)
144 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
mediates the cross talk between the heart and peripheral tissues49ndash51
Here we also observed increased levels of natriuretic peptides (ANP andBNP) which together with IL-6 and TNFa may mediate the cross-talk withthe peripheral tissues to induce glucose intolerance
Does this finding have any relevance to cardiac disease in humansSeveral lines of evidence suggests that congestive heart failure in patientsis associated with an increase in blood glucose levels52 In addition highblood glucose levels in non-diabetic patients with acute myocardialinfarction serves as a prognostic predictor of long-term mortality5354
Thus here we provide evidence that high blood glucose levels are a partof the symptoms of cardiac remodeling process
Collectively our findings suggest that cardiac ATF3 expression isresponsible for an adaptive cardiac response to the stressful environmentunder a HFD (Figure 8) ATF3 dampens the inflammatory response andsuppresses the expression of ECM remodeling genes and thus amelioratescardiac function In HFD-fed mice with ATF3 deficiency or in mice withcardiac remodeling the inflamed heart affects also peripheral tissuesthereby worsening T2D in peripheral tissues resulting in elevated bloodglucose level and turning on a vicious cycle to worsen cardiomyopathy
Supplementary material
Supplementary material is available at Cardiovascular Research online
AcknowledgementsThe authors wish to thank Drs Keren Bitton-Worms and Ran Rostokerand Prof Derek LeRoith from the Rambam Medical Center for their
assistance in GTT and ITT Dr Edith Suss Toby Maya Holdengreber andLior Liba from the BCF Bioimaging Center (Faculty of MedicineTechnion) for their assistance with imaging and image analysis Prof DEMichelle from University of Michigan for cardiomyocytes culture techni-que and Prof Yehiel Zick from the Weizmann institute of Sciences forcritical comments
Conflict of interest none declared
FundingThis work was partially supported by the United States-Israel BinationalGrant 2009179 to AA and TH In addition we wish to thank the MarsPittsburgh Fund for Medical Research to AA the Department of Defensegrant W81XWH-14-1-0179 to TH and to D Dan and Betty Kahn travelgrant support to LK
References
1 Roger VL Go AS Lloyd-Jones DM Benjamin EJ Berry JD Borden WB Bravata DMDai S Ford ES Fox CS Fullerton HJ Gillespie C Hailpern SM Heit JA Howard VJKissela BM Kittner SJ Lackland DT Lichtman JH Lisabeth LD Makuc DM MarcusGM Marelli A Matchar DB Moy CS Mozaffarian D Mussolino ME Nichol GPaynter NP Soliman EZ Sorlie PD Sotoodehnia N Turan TN Virani SS Wong NDWoo D Turner MB American Heart Association Statistics C Stroke Statistics SHeart disease and stroke statisticsndash2012 update a report from the American HeartAssociation Circulation 2012125e2ndashe220
2 Wilson C Diabetes ACCORD 5-year outcomes of intensive glycemic control NatRev Endocrinol 20117314
3 Ernande L Derumeaux G Diabetic cardiomyopathy myth or reality Arch CardiovascDis 2012105218ndash225
4 Battiprolu PK Hojayev B Jiang N Wang ZV Luo X Iglewski M Shelton JM GerardRD Rothermel BA Gillette TG Lavandero S Hill JA Metabolic stress-induced
InsulinGF
Cytokines
FFA
GlucoseATF3
Cardiac
Remodeling
Factors
Paracrine action
Collagens
and ECM
proteins
Fibrosis
Cardiac
Dysfunction
Systemic Effects
Glucose Tolerance
TNFα
IL-6
Cardiomyocyte
ATF3
Homeostasis
Homeostasis
TNFα
IL-6
TIMP1
BNP
ANP
Figure 8 lsquoPeripheral-to-cardiacrsquo and lsquoCardiac-to-peripheralrsquo model scheme Schematic diagram showing the ATF3 adaptive role in maintaining hearthomeostasis following HFD-induced T2D In contrast stressful environment under HFD leads to maladaptive cardiac response in the absence of ATF3Cardiac remodeling caused by ATF3 deficiency induces an increase in cardiac ECM remodeling and inflammatory genes and mediates the communication ofcardiomyocytes with heart resident cells potentially mediated by IL-6 TNFa ANP BNP and TIMP1 In this situation increased cardiac fibrosis and inflamma-tory response worsen cardiac function The inflamed failing heart mediates the communication of cardiomyocytes with distant organs as well thereby affect-ing diabetic outcome and glucose homeostasis
ATF3 expression in cardiomyocytes preserves homeostasis 145D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
activation of FoxO1 triggers diabetic cardiomyopathy in mice J Clin Invest20121221109ndash1118
5 Mandavia CH Aroor AR Demarco VG Sowers JR Molecular and metabolic mecha-nisms of cardiac dysfunction in diabetes Life Sci 201392601ndash608
6 Ventura-Clapier R Garnier A Veksler V Energy metabolism in heart failure J Physiol20045551ndash13
7 Guo S Insulin signaling resistance and the metabolic syndrome insights from mousemodels into disease mechanisms J Endocrinol 2014220T1ndashT23
8 Montane J Cadavez L Novials A Stress and the inflammatory process a major causeof pancreatic cell death in type 2 diabetes Diabetes Metab Syndr Obes 2014725ndash34
9 Ashrafian H Frenneaux MP Opie LH Metabolic mechanisms in heart failureCirculation 2007116434ndash448
10 Tanabe K Liu Y Hasan SD Martinez SC Cras-Meneur C Welling CM Bernal-MizrachiE Tanizawa Y Rhodes CJ Zmuda E Hai T Abumrad NA Permutt MA Glucose andfatty acids synergize to promote B-cell apoptosis through activation of glycogen syn-thase kinase 3beta independent of JNK activation PLoS One 20116e18146
11 Heineke J Molkentin JD Regulation of cardiac hypertrophy by intracellular signallingpathways Nat Rev Mol Cell Biol 20067589ndash600
12 Hai T Wolford CC Chang YS ATF3 a hub of the cellular adaptive-response net-work in the pathogenesis of diseases is modulation of inflammation a unifying com-ponent Gene Expr 2010151ndash11
13 Takii R Inouye S Fujimoto M Nakamura T Shinkawa T Prakasam R Tan K HayashidaN Ichikawa H Hai T Nakai A Heat shock transcription factor 1 inhibits expression ofIL-6 through activating transcription factor 3 J Immunol 20101841041ndash1048
14 Giraldo A Barrett OP Tindall MJ Fuller SJ Amirak E Bhattacharya BS Sugden PHClerk A Feedback regulation by Atf3 in the endothelin-1-responsive transcriptomeof cardiomyocytes Egr1 is a principal Atf3 target Biochem J 2012444343ndash355
15 Hasin T Elhanani O Abassi Z Hai T Aronheim A Angiotensin II signaling up-regulates the immediate early transcription factor ATF3 in the left but not the rightatrium Basic Res Cardiol 2011106175ndash187
16 Koren L Elhanani O Kehat I Hai T Aronheim A Adult cardiac expression of theactivating transcription factor 3 ATF3 promotes ventricular hypertrophy PLoS One20138e68396
17 Koren L Alishekevitz D Elhanani O Nevelsky A Hai T Kehat I Shaked Y AronheimA ATF3-dependent cross-talk between cardiomyocytes and macrophages promotescardiac maladaptive remodeling Int J Cardiol 2015198232ndash240
18 Okamoto Y Chaves A Chen J Kelley R Jones K Weed HG Gardner KL Gangi LYamaguchi M Klomkleaw W Nakayama T Hamlin RL Carnes C Altschuld R BauerJ Hai T Transgenic mice with cardiac-specific expression of activating transcriptionfactor 3 a stress-inducible gene have conduction abnormalities and contractile dys-function Am J Pathol 2001159639ndash650
19 Zhou H Shen DF Bian ZY Zong J Deng W Zhang Y Guo YY Li H Tang QZActivating transcription factor 3 deficiency promotes cardiac hypertrophy dysfunc-tion and fibrosis induced by pressure overload PLoS One 20116e26744
20 Zhou H Guo H Zong J Dai J Yuan Y Bian ZY Tang QZ ATF3 regulates multipletargets and may play a dual role in cardiac hypertrophy and injury Int J Cardiol2014174838ndash839
21 Yang J Yang CJ Yang J Fan ZX ATF3 a promotion effect or a inhibition effect in car-diac maladaptive remodeling Int J Cardiol 2015201245ndash246
22 Koren L Shaked Y Aronheim A Response letter ldquoATF3 a promoter or inhibitor ofcardiac maladaptive remodelingrdquo Int J Cardiol 2015201692
23 Ma ZG Wei WY Xu SC Zhang WB Dai J Tang QZ ATF3 a potential target forcardiac maladaptive remodeling Int J Cardiol 201620250ndash51
24 Zhou H Yuan Y Ni J Guo H Deng W Bian ZY Tang QZ Pleiotropic and puzzlingeffects of ATF3 in maladaptive cardiac remodeling Int J Cardiol 201620687ndash88
25 Zmuda EJ Qi L Zhu MX Mirmira RG Montminy MR Hai T The roles of ATF3 anadaptive-response gene in high-fat-diet-induced diabetes and pancreatic beta-celldysfunction Mol Endocrinol 2010241423ndash1433
26 Lu D Chen J Hai T The regulation of ATF3 gene expression by mitogen-activatedprotein kinases Biochem J 2007401559ndash567
27 Hartman MG Lu D Kim ML Kociba GJ Shukri T Buteau J Wang X Frankel WLGuttridge D Prentki M Grey ST Ron D Hai T Role for activating transcription fac-tor 3 in stress-induced beta-cell apoptosis Mol Cell Biol 2004245721ndash5732
28 Wolford CC McConoughey SJ Jalgaonkar SP Leon M Merchant AS Dominick JLYin X Chang Y Zmuda EJ OrsquoToole SA Millar EK Roller SL Shapiro CL OstrowskiMC Sutherland RL Hai T Transcription factor ATF3 links host adaptive response tobreast cancer metastasis J Clin Iinvest 20131232893ndash2906
29 Agah R Frenkel PA French BA Michael LH Overbeek PA Schneider MD Generecombination in postmitotic cells Targeted expression of Cre recombinase pro-vokes cardiac-restricted site-specific rearrangement in adult ventricular musclein vivo J Clin Invest 1997100169ndash179
30 Livet J Weissman TA Kang H Draft RW Lu J Bennis RA Sanes JR Lichtman JWTransgenic strategies for combinatorial expression of fluorescent proteins in thenervous system Nature 200745056ndash62
31 Davis J Maillet M Miano JM Molkentin JD Lost in transgenesis a userrsquos guidefor genetically manipulating the mouse in cardiac research Circ Res2012111761ndash777
32 Heiberg E Sjogren J Ugander M Carlsson M Engblom H Arheden H Design andvalidation of segment ndash freely available software for cardiovascular image analysisBMC Med Imaging 2010101
33 Kabaeva Z Zhao M Michele DE Blebbistatin extends culture life of adult mouse car-diac myocytes and allows efficient and stable transgene expression Am J Physiol HeartC 2008294H1667ndash1674
34 Subramaniam A Jones WK Gulick J Wert S Neumann J Robbins J Tissue-specificregulation of the alpha-myosin heavy chain gene promoter in transgenic mice J BiolChem 199126624613ndash24620
35 Cunha DA Hekerman P Ladriere L Bazarra-Castro A Ortis F Wakeham MCMoore F Rasschaert J Cardozo AK Bellomo E Overbergh L Mathieu C Lupi R HaiT Herchuelz A Marchetti P Rutter GA Eizirik DL Cnop M Initiation and executionof lipotoxic ER stress in pancreatic beta-cells J Cell Sci 20081212308ndash2318
36 Wynn TA Ramalingam TR Mechanisms of fibrosis therapeutic translation forfibrotic disease Nat Med 2012181028ndash1040
37 Li D Yin X Zmuda EJ Wolford CC Dong X White MF Hai T The repression ofIRS2 gene by ATF3 a stress-inducible gene contributes to pancreatic beta-cell apop-tosis Diabetes 200857635ndash644
38 Myerson SG Bellenger NG Pennell DJ Assessment of left ventricular mass by cardi-ovascular magnetic resonance Hypertension 200239750ndash755
39 Zmuda EJ Viapiano M Grey ST Hadley G Garcia-Ocana A Hai T Deficiency ofAtf3 an adaptive-response gene protects islets and ameliorates inflammation in asyngeneic mouse transplantation model Diabetologia 2010531438ndash1450
40 Riehle C Abel ED Insulin signaling and heart failure Circ Res 20161181151ndash116941 Schreyer SA Wilson DL LeBoeuf RC C57BL6 mice fed high fat diets as models for
diabetes-accelerated atherosclerosis Atherosclerosis 199813617ndash2442 Brainard RE Watson LJ Demartino AM Brittian KR Readnower RD Boakye AA
Zhang D Hoetker JD Bhatnagar A Baba SP Jones SP High fat feeding in mice isinsufficient to induce cardiac dysfunction and does not exacerbate heart failure PLoSOne 20138e83174
43 Gupte AA Minze LJ Reyes M Ren Y Wang X Brunner G Ghosn M Cordero-Reyes AM Ding K Pratico D Morrisett J Shi ZZ Hamilton DJ Lyon CJ Hsueh WAHigh-fat feeding-induced hyperinsulinemia increases cardiac glucose uptake and mito-chondrial function despite peripheral insulin resistance Endocrinology20131542650ndash2662
44 Logan SK Garabedian MJ Campbell CE Werb Z Synergistic transcriptionalactivation of the tissue inhibitor of metalloproteinases-1 promoter via functionalinteraction of AP-1 and Ets-1 transcription factors J Biol Chem1996271774ndash782
45 Belke DD Betuing S Tuttle MJ Graveleau C Young ME Pham M Zhang DCooksey RC McClain DA Litwin SE Taegtmeyer H Severson D Kahn CR AbelED Insulin signaling coordinately regulates cardiac size metabolism and contractileprotein isoform expression J Clin Invest 2002109629ndash639
46 Fernandez AM Kim JK Yakar S Dupont J Hernandez-Sanchez C Castle AL FilmoreJ Shulman GI Le Roith D Functional inactivation of the IGF-I and insulin receptors inskeletal muscle causes type 2 diabetes Genes Dev 2001151926ndash1934
47 Gilchrist M Thorsson V Li B Rust AG Korb M Roach JC Kennedy K Hai TBolouri H Aderem A Systems biology approaches identify ATF3 as a negative regu-lator of Toll-like receptor 4 Nature 2006441173ndash178
48 Kristiansen OP Mandrup-Poulsen T Interleukin-6 and diabetes the good the bad orthe indifferent Diabetes 200554(Suppl 2)S114ndashS124
49 Grueter CE van Rooij E Johnson BA DeLeon SM Sutherland LB Qi X Gautron LElmquist JK Bassel-Duby R Olson EN A cardiac microRNA governs systemic energyhomeostasis by regulation of MED13 Cell 2012149671ndash683
50 Baskin KK Grueter CE Kusminski CM Holland WL Bookout AL Satapati S KongYM Burgess SC Malloy CR Scherer PE Newgard CB Bassel-Duby R Olson ENMED13-dependent signaling from the heart confers leanness by enhancing metabo-lism in adipose tissue and liver EMBO Mol Med 201461610ndash1621
51 Nakamura M Sadoshima J Heart over mind metabolic control of white adipose tis-sue and liver EMBO Mol Med 201461521ndash1524
52 Tenenbaum A Fisman EZ Impaired glucose metabolism in patients with heart failurepathophysiology and possible treatment strategies Am J Cardiovasc Drugs20044269ndash280
53 Suskin N McKelvie RS Burns RJ Latini R Pericak D Probstfield J Rouleau JL SigouinC Solymoss CB Tsuyuki R White M Yusuf S Glucose and insulin abnormalitiesrelate to functional capacity in patients with congestive heart failure Eur Heart J2000211368ndash1375
54 Aronson D Hammerman H Kapeliovich MR Suleiman A Agmon Y Beyar RMarkiewicz W Suleiman M Fasting glucose in acute myocardial infarction incremen-tal value for long-term mortality and relationship with left ventricular systolic func-tion Diabetes Care 200730960ndash966
146 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
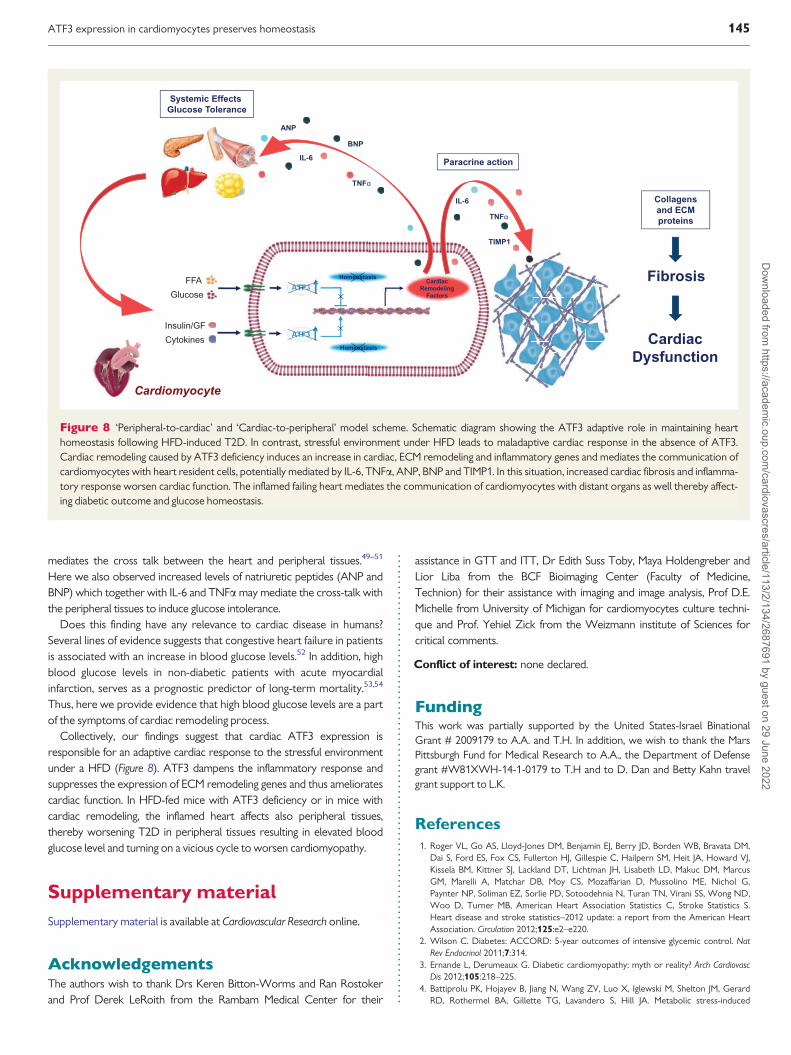
mediates the cross talk between the heart and peripheral tissues49ndash51
Here we also observed increased levels of natriuretic peptides (ANP andBNP) which together with IL-6 and TNFa may mediate the cross-talk withthe peripheral tissues to induce glucose intolerance
Does this finding have any relevance to cardiac disease in humansSeveral lines of evidence suggests that congestive heart failure in patientsis associated with an increase in blood glucose levels52 In addition highblood glucose levels in non-diabetic patients with acute myocardialinfarction serves as a prognostic predictor of long-term mortality5354
Thus here we provide evidence that high blood glucose levels are a partof the symptoms of cardiac remodeling process
Collectively our findings suggest that cardiac ATF3 expression isresponsible for an adaptive cardiac response to the stressful environmentunder a HFD (Figure 8) ATF3 dampens the inflammatory response andsuppresses the expression of ECM remodeling genes and thus amelioratescardiac function In HFD-fed mice with ATF3 deficiency or in mice withcardiac remodeling the inflamed heart affects also peripheral tissuesthereby worsening T2D in peripheral tissues resulting in elevated bloodglucose level and turning on a vicious cycle to worsen cardiomyopathy
Supplementary material
Supplementary material is available at Cardiovascular Research online
AcknowledgementsThe authors wish to thank Drs Keren Bitton-Worms and Ran Rostokerand Prof Derek LeRoith from the Rambam Medical Center for their
assistance in GTT and ITT Dr Edith Suss Toby Maya Holdengreber andLior Liba from the BCF Bioimaging Center (Faculty of MedicineTechnion) for their assistance with imaging and image analysis Prof DEMichelle from University of Michigan for cardiomyocytes culture techni-que and Prof Yehiel Zick from the Weizmann institute of Sciences forcritical comments
Conflict of interest none declared
FundingThis work was partially supported by the United States-Israel BinationalGrant 2009179 to AA and TH In addition we wish to thank the MarsPittsburgh Fund for Medical Research to AA the Department of Defensegrant W81XWH-14-1-0179 to TH and to D Dan and Betty Kahn travelgrant support to LK
References
1 Roger VL Go AS Lloyd-Jones DM Benjamin EJ Berry JD Borden WB Bravata DMDai S Ford ES Fox CS Fullerton HJ Gillespie C Hailpern SM Heit JA Howard VJKissela BM Kittner SJ Lackland DT Lichtman JH Lisabeth LD Makuc DM MarcusGM Marelli A Matchar DB Moy CS Mozaffarian D Mussolino ME Nichol GPaynter NP Soliman EZ Sorlie PD Sotoodehnia N Turan TN Virani SS Wong NDWoo D Turner MB American Heart Association Statistics C Stroke Statistics SHeart disease and stroke statisticsndash2012 update a report from the American HeartAssociation Circulation 2012125e2ndashe220
2 Wilson C Diabetes ACCORD 5-year outcomes of intensive glycemic control NatRev Endocrinol 20117314
3 Ernande L Derumeaux G Diabetic cardiomyopathy myth or reality Arch CardiovascDis 2012105218ndash225
4 Battiprolu PK Hojayev B Jiang N Wang ZV Luo X Iglewski M Shelton JM GerardRD Rothermel BA Gillette TG Lavandero S Hill JA Metabolic stress-induced
InsulinGF
Cytokines
FFA
GlucoseATF3
Cardiac
Remodeling
Factors
Paracrine action
Collagens
and ECM
proteins
Fibrosis
Cardiac
Dysfunction
Systemic Effects
Glucose Tolerance
TNFα
IL-6
Cardiomyocyte
ATF3
Homeostasis
Homeostasis
TNFα
IL-6
TIMP1
BNP
ANP
Figure 8 lsquoPeripheral-to-cardiacrsquo and lsquoCardiac-to-peripheralrsquo model scheme Schematic diagram showing the ATF3 adaptive role in maintaining hearthomeostasis following HFD-induced T2D In contrast stressful environment under HFD leads to maladaptive cardiac response in the absence of ATF3Cardiac remodeling caused by ATF3 deficiency induces an increase in cardiac ECM remodeling and inflammatory genes and mediates the communication ofcardiomyocytes with heart resident cells potentially mediated by IL-6 TNFa ANP BNP and TIMP1 In this situation increased cardiac fibrosis and inflamma-tory response worsen cardiac function The inflamed failing heart mediates the communication of cardiomyocytes with distant organs as well thereby affect-ing diabetic outcome and glucose homeostasis
ATF3 expression in cardiomyocytes preserves homeostasis 145D
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022
activation of FoxO1 triggers diabetic cardiomyopathy in mice J Clin Invest20121221109ndash1118
5 Mandavia CH Aroor AR Demarco VG Sowers JR Molecular and metabolic mecha-nisms of cardiac dysfunction in diabetes Life Sci 201392601ndash608
6 Ventura-Clapier R Garnier A Veksler V Energy metabolism in heart failure J Physiol20045551ndash13
7 Guo S Insulin signaling resistance and the metabolic syndrome insights from mousemodels into disease mechanisms J Endocrinol 2014220T1ndashT23
8 Montane J Cadavez L Novials A Stress and the inflammatory process a major causeof pancreatic cell death in type 2 diabetes Diabetes Metab Syndr Obes 2014725ndash34
9 Ashrafian H Frenneaux MP Opie LH Metabolic mechanisms in heart failureCirculation 2007116434ndash448
10 Tanabe K Liu Y Hasan SD Martinez SC Cras-Meneur C Welling CM Bernal-MizrachiE Tanizawa Y Rhodes CJ Zmuda E Hai T Abumrad NA Permutt MA Glucose andfatty acids synergize to promote B-cell apoptosis through activation of glycogen syn-thase kinase 3beta independent of JNK activation PLoS One 20116e18146
11 Heineke J Molkentin JD Regulation of cardiac hypertrophy by intracellular signallingpathways Nat Rev Mol Cell Biol 20067589ndash600
12 Hai T Wolford CC Chang YS ATF3 a hub of the cellular adaptive-response net-work in the pathogenesis of diseases is modulation of inflammation a unifying com-ponent Gene Expr 2010151ndash11
13 Takii R Inouye S Fujimoto M Nakamura T Shinkawa T Prakasam R Tan K HayashidaN Ichikawa H Hai T Nakai A Heat shock transcription factor 1 inhibits expression ofIL-6 through activating transcription factor 3 J Immunol 20101841041ndash1048
14 Giraldo A Barrett OP Tindall MJ Fuller SJ Amirak E Bhattacharya BS Sugden PHClerk A Feedback regulation by Atf3 in the endothelin-1-responsive transcriptomeof cardiomyocytes Egr1 is a principal Atf3 target Biochem J 2012444343ndash355
15 Hasin T Elhanani O Abassi Z Hai T Aronheim A Angiotensin II signaling up-regulates the immediate early transcription factor ATF3 in the left but not the rightatrium Basic Res Cardiol 2011106175ndash187
16 Koren L Elhanani O Kehat I Hai T Aronheim A Adult cardiac expression of theactivating transcription factor 3 ATF3 promotes ventricular hypertrophy PLoS One20138e68396
17 Koren L Alishekevitz D Elhanani O Nevelsky A Hai T Kehat I Shaked Y AronheimA ATF3-dependent cross-talk between cardiomyocytes and macrophages promotescardiac maladaptive remodeling Int J Cardiol 2015198232ndash240
18 Okamoto Y Chaves A Chen J Kelley R Jones K Weed HG Gardner KL Gangi LYamaguchi M Klomkleaw W Nakayama T Hamlin RL Carnes C Altschuld R BauerJ Hai T Transgenic mice with cardiac-specific expression of activating transcriptionfactor 3 a stress-inducible gene have conduction abnormalities and contractile dys-function Am J Pathol 2001159639ndash650
19 Zhou H Shen DF Bian ZY Zong J Deng W Zhang Y Guo YY Li H Tang QZActivating transcription factor 3 deficiency promotes cardiac hypertrophy dysfunc-tion and fibrosis induced by pressure overload PLoS One 20116e26744
20 Zhou H Guo H Zong J Dai J Yuan Y Bian ZY Tang QZ ATF3 regulates multipletargets and may play a dual role in cardiac hypertrophy and injury Int J Cardiol2014174838ndash839
21 Yang J Yang CJ Yang J Fan ZX ATF3 a promotion effect or a inhibition effect in car-diac maladaptive remodeling Int J Cardiol 2015201245ndash246
22 Koren L Shaked Y Aronheim A Response letter ldquoATF3 a promoter or inhibitor ofcardiac maladaptive remodelingrdquo Int J Cardiol 2015201692
23 Ma ZG Wei WY Xu SC Zhang WB Dai J Tang QZ ATF3 a potential target forcardiac maladaptive remodeling Int J Cardiol 201620250ndash51
24 Zhou H Yuan Y Ni J Guo H Deng W Bian ZY Tang QZ Pleiotropic and puzzlingeffects of ATF3 in maladaptive cardiac remodeling Int J Cardiol 201620687ndash88
25 Zmuda EJ Qi L Zhu MX Mirmira RG Montminy MR Hai T The roles of ATF3 anadaptive-response gene in high-fat-diet-induced diabetes and pancreatic beta-celldysfunction Mol Endocrinol 2010241423ndash1433
26 Lu D Chen J Hai T The regulation of ATF3 gene expression by mitogen-activatedprotein kinases Biochem J 2007401559ndash567
27 Hartman MG Lu D Kim ML Kociba GJ Shukri T Buteau J Wang X Frankel WLGuttridge D Prentki M Grey ST Ron D Hai T Role for activating transcription fac-tor 3 in stress-induced beta-cell apoptosis Mol Cell Biol 2004245721ndash5732
28 Wolford CC McConoughey SJ Jalgaonkar SP Leon M Merchant AS Dominick JLYin X Chang Y Zmuda EJ OrsquoToole SA Millar EK Roller SL Shapiro CL OstrowskiMC Sutherland RL Hai T Transcription factor ATF3 links host adaptive response tobreast cancer metastasis J Clin Iinvest 20131232893ndash2906
29 Agah R Frenkel PA French BA Michael LH Overbeek PA Schneider MD Generecombination in postmitotic cells Targeted expression of Cre recombinase pro-vokes cardiac-restricted site-specific rearrangement in adult ventricular musclein vivo J Clin Invest 1997100169ndash179
30 Livet J Weissman TA Kang H Draft RW Lu J Bennis RA Sanes JR Lichtman JWTransgenic strategies for combinatorial expression of fluorescent proteins in thenervous system Nature 200745056ndash62
31 Davis J Maillet M Miano JM Molkentin JD Lost in transgenesis a userrsquos guidefor genetically manipulating the mouse in cardiac research Circ Res2012111761ndash777
32 Heiberg E Sjogren J Ugander M Carlsson M Engblom H Arheden H Design andvalidation of segment ndash freely available software for cardiovascular image analysisBMC Med Imaging 2010101
33 Kabaeva Z Zhao M Michele DE Blebbistatin extends culture life of adult mouse car-diac myocytes and allows efficient and stable transgene expression Am J Physiol HeartC 2008294H1667ndash1674
34 Subramaniam A Jones WK Gulick J Wert S Neumann J Robbins J Tissue-specificregulation of the alpha-myosin heavy chain gene promoter in transgenic mice J BiolChem 199126624613ndash24620
35 Cunha DA Hekerman P Ladriere L Bazarra-Castro A Ortis F Wakeham MCMoore F Rasschaert J Cardozo AK Bellomo E Overbergh L Mathieu C Lupi R HaiT Herchuelz A Marchetti P Rutter GA Eizirik DL Cnop M Initiation and executionof lipotoxic ER stress in pancreatic beta-cells J Cell Sci 20081212308ndash2318
36 Wynn TA Ramalingam TR Mechanisms of fibrosis therapeutic translation forfibrotic disease Nat Med 2012181028ndash1040
37 Li D Yin X Zmuda EJ Wolford CC Dong X White MF Hai T The repression ofIRS2 gene by ATF3 a stress-inducible gene contributes to pancreatic beta-cell apop-tosis Diabetes 200857635ndash644
38 Myerson SG Bellenger NG Pennell DJ Assessment of left ventricular mass by cardi-ovascular magnetic resonance Hypertension 200239750ndash755
39 Zmuda EJ Viapiano M Grey ST Hadley G Garcia-Ocana A Hai T Deficiency ofAtf3 an adaptive-response gene protects islets and ameliorates inflammation in asyngeneic mouse transplantation model Diabetologia 2010531438ndash1450
40 Riehle C Abel ED Insulin signaling and heart failure Circ Res 20161181151ndash116941 Schreyer SA Wilson DL LeBoeuf RC C57BL6 mice fed high fat diets as models for
diabetes-accelerated atherosclerosis Atherosclerosis 199813617ndash2442 Brainard RE Watson LJ Demartino AM Brittian KR Readnower RD Boakye AA
Zhang D Hoetker JD Bhatnagar A Baba SP Jones SP High fat feeding in mice isinsufficient to induce cardiac dysfunction and does not exacerbate heart failure PLoSOne 20138e83174
43 Gupte AA Minze LJ Reyes M Ren Y Wang X Brunner G Ghosn M Cordero-Reyes AM Ding K Pratico D Morrisett J Shi ZZ Hamilton DJ Lyon CJ Hsueh WAHigh-fat feeding-induced hyperinsulinemia increases cardiac glucose uptake and mito-chondrial function despite peripheral insulin resistance Endocrinology20131542650ndash2662
44 Logan SK Garabedian MJ Campbell CE Werb Z Synergistic transcriptionalactivation of the tissue inhibitor of metalloproteinases-1 promoter via functionalinteraction of AP-1 and Ets-1 transcription factors J Biol Chem1996271774ndash782
45 Belke DD Betuing S Tuttle MJ Graveleau C Young ME Pham M Zhang DCooksey RC McClain DA Litwin SE Taegtmeyer H Severson D Kahn CR AbelED Insulin signaling coordinately regulates cardiac size metabolism and contractileprotein isoform expression J Clin Invest 2002109629ndash639
46 Fernandez AM Kim JK Yakar S Dupont J Hernandez-Sanchez C Castle AL FilmoreJ Shulman GI Le Roith D Functional inactivation of the IGF-I and insulin receptors inskeletal muscle causes type 2 diabetes Genes Dev 2001151926ndash1934
47 Gilchrist M Thorsson V Li B Rust AG Korb M Roach JC Kennedy K Hai TBolouri H Aderem A Systems biology approaches identify ATF3 as a negative regu-lator of Toll-like receptor 4 Nature 2006441173ndash178
48 Kristiansen OP Mandrup-Poulsen T Interleukin-6 and diabetes the good the bad orthe indifferent Diabetes 200554(Suppl 2)S114ndashS124
49 Grueter CE van Rooij E Johnson BA DeLeon SM Sutherland LB Qi X Gautron LElmquist JK Bassel-Duby R Olson EN A cardiac microRNA governs systemic energyhomeostasis by regulation of MED13 Cell 2012149671ndash683
50 Baskin KK Grueter CE Kusminski CM Holland WL Bookout AL Satapati S KongYM Burgess SC Malloy CR Scherer PE Newgard CB Bassel-Duby R Olson ENMED13-dependent signaling from the heart confers leanness by enhancing metabo-lism in adipose tissue and liver EMBO Mol Med 201461610ndash1621
51 Nakamura M Sadoshima J Heart over mind metabolic control of white adipose tis-sue and liver EMBO Mol Med 201461521ndash1524
52 Tenenbaum A Fisman EZ Impaired glucose metabolism in patients with heart failurepathophysiology and possible treatment strategies Am J Cardiovasc Drugs20044269ndash280
53 Suskin N McKelvie RS Burns RJ Latini R Pericak D Probstfield J Rouleau JL SigouinC Solymoss CB Tsuyuki R White M Yusuf S Glucose and insulin abnormalitiesrelate to functional capacity in patients with congestive heart failure Eur Heart J2000211368ndash1375
54 Aronson D Hammerman H Kapeliovich MR Suleiman A Agmon Y Beyar RMarkiewicz W Suleiman M Fasting glucose in acute myocardial infarction incremen-tal value for long-term mortality and relationship with left ventricular systolic func-tion Diabetes Care 200730960ndash966
146 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022

activation of FoxO1 triggers diabetic cardiomyopathy in mice J Clin Invest20121221109ndash1118
5 Mandavia CH Aroor AR Demarco VG Sowers JR Molecular and metabolic mecha-nisms of cardiac dysfunction in diabetes Life Sci 201392601ndash608
6 Ventura-Clapier R Garnier A Veksler V Energy metabolism in heart failure J Physiol20045551ndash13
7 Guo S Insulin signaling resistance and the metabolic syndrome insights from mousemodels into disease mechanisms J Endocrinol 2014220T1ndashT23
8 Montane J Cadavez L Novials A Stress and the inflammatory process a major causeof pancreatic cell death in type 2 diabetes Diabetes Metab Syndr Obes 2014725ndash34
9 Ashrafian H Frenneaux MP Opie LH Metabolic mechanisms in heart failureCirculation 2007116434ndash448
10 Tanabe K Liu Y Hasan SD Martinez SC Cras-Meneur C Welling CM Bernal-MizrachiE Tanizawa Y Rhodes CJ Zmuda E Hai T Abumrad NA Permutt MA Glucose andfatty acids synergize to promote B-cell apoptosis through activation of glycogen syn-thase kinase 3beta independent of JNK activation PLoS One 20116e18146
11 Heineke J Molkentin JD Regulation of cardiac hypertrophy by intracellular signallingpathways Nat Rev Mol Cell Biol 20067589ndash600
12 Hai T Wolford CC Chang YS ATF3 a hub of the cellular adaptive-response net-work in the pathogenesis of diseases is modulation of inflammation a unifying com-ponent Gene Expr 2010151ndash11
13 Takii R Inouye S Fujimoto M Nakamura T Shinkawa T Prakasam R Tan K HayashidaN Ichikawa H Hai T Nakai A Heat shock transcription factor 1 inhibits expression ofIL-6 through activating transcription factor 3 J Immunol 20101841041ndash1048
14 Giraldo A Barrett OP Tindall MJ Fuller SJ Amirak E Bhattacharya BS Sugden PHClerk A Feedback regulation by Atf3 in the endothelin-1-responsive transcriptomeof cardiomyocytes Egr1 is a principal Atf3 target Biochem J 2012444343ndash355
15 Hasin T Elhanani O Abassi Z Hai T Aronheim A Angiotensin II signaling up-regulates the immediate early transcription factor ATF3 in the left but not the rightatrium Basic Res Cardiol 2011106175ndash187
16 Koren L Elhanani O Kehat I Hai T Aronheim A Adult cardiac expression of theactivating transcription factor 3 ATF3 promotes ventricular hypertrophy PLoS One20138e68396
17 Koren L Alishekevitz D Elhanani O Nevelsky A Hai T Kehat I Shaked Y AronheimA ATF3-dependent cross-talk between cardiomyocytes and macrophages promotescardiac maladaptive remodeling Int J Cardiol 2015198232ndash240
18 Okamoto Y Chaves A Chen J Kelley R Jones K Weed HG Gardner KL Gangi LYamaguchi M Klomkleaw W Nakayama T Hamlin RL Carnes C Altschuld R BauerJ Hai T Transgenic mice with cardiac-specific expression of activating transcriptionfactor 3 a stress-inducible gene have conduction abnormalities and contractile dys-function Am J Pathol 2001159639ndash650
19 Zhou H Shen DF Bian ZY Zong J Deng W Zhang Y Guo YY Li H Tang QZActivating transcription factor 3 deficiency promotes cardiac hypertrophy dysfunc-tion and fibrosis induced by pressure overload PLoS One 20116e26744
20 Zhou H Guo H Zong J Dai J Yuan Y Bian ZY Tang QZ ATF3 regulates multipletargets and may play a dual role in cardiac hypertrophy and injury Int J Cardiol2014174838ndash839
21 Yang J Yang CJ Yang J Fan ZX ATF3 a promotion effect or a inhibition effect in car-diac maladaptive remodeling Int J Cardiol 2015201245ndash246
22 Koren L Shaked Y Aronheim A Response letter ldquoATF3 a promoter or inhibitor ofcardiac maladaptive remodelingrdquo Int J Cardiol 2015201692
23 Ma ZG Wei WY Xu SC Zhang WB Dai J Tang QZ ATF3 a potential target forcardiac maladaptive remodeling Int J Cardiol 201620250ndash51
24 Zhou H Yuan Y Ni J Guo H Deng W Bian ZY Tang QZ Pleiotropic and puzzlingeffects of ATF3 in maladaptive cardiac remodeling Int J Cardiol 201620687ndash88
25 Zmuda EJ Qi L Zhu MX Mirmira RG Montminy MR Hai T The roles of ATF3 anadaptive-response gene in high-fat-diet-induced diabetes and pancreatic beta-celldysfunction Mol Endocrinol 2010241423ndash1433
26 Lu D Chen J Hai T The regulation of ATF3 gene expression by mitogen-activatedprotein kinases Biochem J 2007401559ndash567
27 Hartman MG Lu D Kim ML Kociba GJ Shukri T Buteau J Wang X Frankel WLGuttridge D Prentki M Grey ST Ron D Hai T Role for activating transcription fac-tor 3 in stress-induced beta-cell apoptosis Mol Cell Biol 2004245721ndash5732
28 Wolford CC McConoughey SJ Jalgaonkar SP Leon M Merchant AS Dominick JLYin X Chang Y Zmuda EJ OrsquoToole SA Millar EK Roller SL Shapiro CL OstrowskiMC Sutherland RL Hai T Transcription factor ATF3 links host adaptive response tobreast cancer metastasis J Clin Iinvest 20131232893ndash2906
29 Agah R Frenkel PA French BA Michael LH Overbeek PA Schneider MD Generecombination in postmitotic cells Targeted expression of Cre recombinase pro-vokes cardiac-restricted site-specific rearrangement in adult ventricular musclein vivo J Clin Invest 1997100169ndash179
30 Livet J Weissman TA Kang H Draft RW Lu J Bennis RA Sanes JR Lichtman JWTransgenic strategies for combinatorial expression of fluorescent proteins in thenervous system Nature 200745056ndash62
31 Davis J Maillet M Miano JM Molkentin JD Lost in transgenesis a userrsquos guidefor genetically manipulating the mouse in cardiac research Circ Res2012111761ndash777
32 Heiberg E Sjogren J Ugander M Carlsson M Engblom H Arheden H Design andvalidation of segment ndash freely available software for cardiovascular image analysisBMC Med Imaging 2010101
33 Kabaeva Z Zhao M Michele DE Blebbistatin extends culture life of adult mouse car-diac myocytes and allows efficient and stable transgene expression Am J Physiol HeartC 2008294H1667ndash1674
34 Subramaniam A Jones WK Gulick J Wert S Neumann J Robbins J Tissue-specificregulation of the alpha-myosin heavy chain gene promoter in transgenic mice J BiolChem 199126624613ndash24620
35 Cunha DA Hekerman P Ladriere L Bazarra-Castro A Ortis F Wakeham MCMoore F Rasschaert J Cardozo AK Bellomo E Overbergh L Mathieu C Lupi R HaiT Herchuelz A Marchetti P Rutter GA Eizirik DL Cnop M Initiation and executionof lipotoxic ER stress in pancreatic beta-cells J Cell Sci 20081212308ndash2318
36 Wynn TA Ramalingam TR Mechanisms of fibrosis therapeutic translation forfibrotic disease Nat Med 2012181028ndash1040
37 Li D Yin X Zmuda EJ Wolford CC Dong X White MF Hai T The repression ofIRS2 gene by ATF3 a stress-inducible gene contributes to pancreatic beta-cell apop-tosis Diabetes 200857635ndash644
38 Myerson SG Bellenger NG Pennell DJ Assessment of left ventricular mass by cardi-ovascular magnetic resonance Hypertension 200239750ndash755
39 Zmuda EJ Viapiano M Grey ST Hadley G Garcia-Ocana A Hai T Deficiency ofAtf3 an adaptive-response gene protects islets and ameliorates inflammation in asyngeneic mouse transplantation model Diabetologia 2010531438ndash1450
40 Riehle C Abel ED Insulin signaling and heart failure Circ Res 20161181151ndash116941 Schreyer SA Wilson DL LeBoeuf RC C57BL6 mice fed high fat diets as models for
diabetes-accelerated atherosclerosis Atherosclerosis 199813617ndash2442 Brainard RE Watson LJ Demartino AM Brittian KR Readnower RD Boakye AA
Zhang D Hoetker JD Bhatnagar A Baba SP Jones SP High fat feeding in mice isinsufficient to induce cardiac dysfunction and does not exacerbate heart failure PLoSOne 20138e83174
43 Gupte AA Minze LJ Reyes M Ren Y Wang X Brunner G Ghosn M Cordero-Reyes AM Ding K Pratico D Morrisett J Shi ZZ Hamilton DJ Lyon CJ Hsueh WAHigh-fat feeding-induced hyperinsulinemia increases cardiac glucose uptake and mito-chondrial function despite peripheral insulin resistance Endocrinology20131542650ndash2662
44 Logan SK Garabedian MJ Campbell CE Werb Z Synergistic transcriptionalactivation of the tissue inhibitor of metalloproteinases-1 promoter via functionalinteraction of AP-1 and Ets-1 transcription factors J Biol Chem1996271774ndash782
45 Belke DD Betuing S Tuttle MJ Graveleau C Young ME Pham M Zhang DCooksey RC McClain DA Litwin SE Taegtmeyer H Severson D Kahn CR AbelED Insulin signaling coordinately regulates cardiac size metabolism and contractileprotein isoform expression J Clin Invest 2002109629ndash639
46 Fernandez AM Kim JK Yakar S Dupont J Hernandez-Sanchez C Castle AL FilmoreJ Shulman GI Le Roith D Functional inactivation of the IGF-I and insulin receptors inskeletal muscle causes type 2 diabetes Genes Dev 2001151926ndash1934
47 Gilchrist M Thorsson V Li B Rust AG Korb M Roach JC Kennedy K Hai TBolouri H Aderem A Systems biology approaches identify ATF3 as a negative regu-lator of Toll-like receptor 4 Nature 2006441173ndash178
48 Kristiansen OP Mandrup-Poulsen T Interleukin-6 and diabetes the good the bad orthe indifferent Diabetes 200554(Suppl 2)S114ndashS124
49 Grueter CE van Rooij E Johnson BA DeLeon SM Sutherland LB Qi X Gautron LElmquist JK Bassel-Duby R Olson EN A cardiac microRNA governs systemic energyhomeostasis by regulation of MED13 Cell 2012149671ndash683
50 Baskin KK Grueter CE Kusminski CM Holland WL Bookout AL Satapati S KongYM Burgess SC Malloy CR Scherer PE Newgard CB Bassel-Duby R Olson ENMED13-dependent signaling from the heart confers leanness by enhancing metabo-lism in adipose tissue and liver EMBO Mol Med 201461610ndash1621
51 Nakamura M Sadoshima J Heart over mind metabolic control of white adipose tis-sue and liver EMBO Mol Med 201461521ndash1524
52 Tenenbaum A Fisman EZ Impaired glucose metabolism in patients with heart failurepathophysiology and possible treatment strategies Am J Cardiovasc Drugs20044269ndash280
53 Suskin N McKelvie RS Burns RJ Latini R Pericak D Probstfield J Rouleau JL SigouinC Solymoss CB Tsuyuki R White M Yusuf S Glucose and insulin abnormalitiesrelate to functional capacity in patients with congestive heart failure Eur Heart J2000211368ndash1375
54 Aronson D Hammerman H Kapeliovich MR Suleiman A Agmon Y Beyar RMarkiewicz W Suleiman M Fasting glucose in acute myocardial infarction incremen-tal value for long-term mortality and relationship with left ventricular systolic func-tion Diabetes Care 200730960ndash966
146 R Kalfon et alD
ownloaded from
httpsacademicoupcom
cardiovascresarticle11321342687691 by guest on 29 June 2022