generation of functional cardiomyocytes from adult mouse spermatogonial stem cells
TRANSCRIPT

Generation of Functional Cardiomyocytes From AdultMouse Spermatogonial Stem Cells
Kaomei Guan,* Stefan Wagner,* Bernhard Unsold,* Lars S. Maier, Diana Kaiser,Bernhard Hemmerlein, Karim Nayernia, Wolfgang Engel, Gerd Hasenfuss
Abstract—Stem cell–based therapy is a promising approach for the treatment of heart failure. Adult stem cells with thepluripotency of embryonic stem cells (ESCs) would be an ideal cell source. Recently, we reported the successfulestablishment of multipotent adult germline stem cells (maGSCs) from mouse testis. These cultured maGSCs showphenotypic characteristics similar to ESCs and can spontaneously differentiate into cells from all 3 germ layers. In thepresent study, we used the hanging drop method to differentiate maGSCs into cardiomyocytes and analyzed theirfunctional properties. Differentiation efficiency of beating cardiomyocytes from maGSCs was similar to that from ESCs.The maGSC-derived cardiomyocytes expressed cardiac-specific L-type Ca2 channels and responded to Ca2
channel–modulating drugs. Cx43 was expressed at cell-to-cell contacts in cardiac clusters, and fluorescence recoveryafter photobleaching assay showed the presence of functional gap junctions among cardiomyocytes. Action potentialanalyses demonstrated the presence of pacemaker-, ventricle-, atrial-, and Purkinje-like cardiomyocytes. Stimulationwith isoproterenol resulted in a significant increase in beating frequency, whereas the addition of cadmium chlorideabolished spontaneous electrical activity. Confocal microscopy analysis of intracellular Ca2 in maGSC-derivedcardiomyocytes showed that calcium increased periodically throughout the cell in a homogenous fashion, pointing to afine regulated Ca2 release from intracellular Ca2 stores. By using line-scan mode, we found rhythmic Ca2 transients.Furthermore, we transplanted maGSCs into normal hearts of mice and found that maGSCs were able to proliferate anddifferentiate. No tumor formation was found up to 1 month after cell transplantation. Taken together, we believe thatmaGSCs provide a new source of distinct types of cardiomyocytes for basic research and potential therapeuticapplication. (Circ Res. 2007;100:1615-1625.)
Key Words: spermatogonial stem cells cardiac differentiation gap junction L-type Ca2 channels cell transplantation
When heart muscle is damaged with myocyte apoptosisor necrosis, functional contracting cardiomyocytes are
replaced by nonfunctional scar tissue. Experimental evidencesuggests that the heart harbors a resident population of stemcells able to differentiate into cardiomyocytes, smooth musclecells, and endothelial cells.1,2 A subset of native cardiacprogenitors has been identified in hearts of newborn mice,rats, and humans.3 Despite the possible existence of thesepopulations and their ability to contribute to cardiac repair,these intrinsic mechanisms are inadequate to restore thecardiac function of a failing heart. Therefore, a potentialtherapeutic approach for treatment of heart failure is toreplace lost cardiomyocytes with new functional ones as analternative to whole-heart transplantation.
Several groups have shown that functional cardiomyocytescan be differentiated from embryonic stem cells (ESCs),derived from the inner cell mass of preimplantation embryos.4
ESC-derived cardiomyocytes have been characterized bytheir developmentally controlled expression of cardiac-specific genes, proteins, and ion channels.5–8 Excitation–contraction coupling and electrophysiologic specializationalso have been observed.9,10 Application of ESCs or ESC-derived cardiomyocytes for treatment of heart failure hasalready been tested in animal models and has demonstratedbeneficial effects.11–14 However, to prevent the rejection ofthe implant, it is necessary to make the cells immunocompat-ible with the recipient. In addition, the use of human ESCshas encountered opposition that has led to considerationsregarding limited availability.
To circumvent these problems, adult stem cells are underinvestigation. Transdifferentiation of adult stem cells in vivointo cardiomyocytes has been shown by some investiga-tors15–17 but not by others.18,19 Moreover, only a few experi-ments have suggested that adult stem cells could generate
Original received July 4, 2006; resubmission received February 21, 2007; revised resubmission received March 30, 2007; accepted April 25, 2007.From the Departments of Cardiology and Pneumology (K.G., S.W., B.U., L.S.M., D.K., G.H.) and Pathology (B.H.) and the Institute of Human
Genetics (K.N., W.E.), Georg-August-University of Gottingen, Germany*These authors contributed equally to this work.Correspondence to Prof Dr Gerd Hasenfuss, Department of Cardiology and Pneumology, Georg-August-University of Gottingen, Robert-Koch-Str. 40,
37075 Gottingen, Germany. E-mail [email protected]© 2007 American Heart Association, Inc.
Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/01.RES.0000269182.22798.d9
1615 by guest on February 23, 2016http://circres.ahajournals.org/Downloaded from by guest on February 23, 2016http://circres.ahajournals.org/Downloaded from by guest on February 23, 2016http://circres.ahajournals.org/Downloaded from by guest on February 23, 2016http://circres.ahajournals.org/Downloaded from by guest on February 23, 2016http://circres.ahajournals.org/Downloaded from by guest on February 23, 2016http://circres.ahajournals.org/Downloaded from by guest on February 23, 2016http://circres.ahajournals.org/Downloaded from by guest on February 23, 2016http://circres.ahajournals.org/Downloaded from by guest on February 23, 2016http://circres.ahajournals.org/Downloaded from by guest on February 23, 2016http://circres.ahajournals.org/Downloaded from by guest on February 23, 2016http://circres.ahajournals.org/Downloaded from by guest on February 23, 2016http://circres.ahajournals.org/Downloaded from by guest on February 23, 2016http://circres.ahajournals.org/Downloaded from

cardiomyocytes under in vitro conditions.20,21 However, thishas also been challenged by other studies.22
We recently showed that stem cells from adult mousetestis, similar to ESCs, are pluripotent.23 Spermatogonialstem cells (SSCs), a unique population of germline stem cellsin adult testis, have the capability to self-renew and toproduce daughter cells destined to differentiate into sperma-tozoa throughout life.24 We have successfully isolated SSCsfrom adult testis and established in vitro culture conditions toconvert SSCs into ESC-like cells, the so called multipotentadult germline stem cells (maGSCs). These maGSCs showphenotypic characteristics similar to mouse ESCs. They canspontaneously differentiate into derivatives of all three germlayers in vitro. These data suggest that SSCs could be a newand promising source of adult stem cells for myocardialregeneration.
The purpose of the present study was to analyze thecomplex functional properties of cardiomyocytes derivedfrom maGSCs in vitro and to analyze the behavior ofundifferentiated maGSCs in normal hearts of mice in vivoafter transplantation. Using molecular, cellular, and physio-logical assays, we found that maGSC-derived cardiomyo-cytes had similar properties to those derived from ESCs. Theyexhibited characteristics typical of heart cells in early stagesof cardiac development. After transplantation of undifferen-tiated maGSCs into normal mouse hearts, they were able toproliferate and differentiate into vascular endothelial andsmooth muscle cells in vivo, and no tumor formation wasfound up to 1 month after cell delivery.
Materials and MethodsIn Vitro Differentiation of maGSCsEstablished maGSC cultures (SSC5, C57BL6 line 9, FVB line 5, and129 line 2) from different mouse strains were cultivated on a feederlayer of mouse embryonic fibroblasts in DMEM (Invitrogen) sup-plemented by 15% FCS (Invitrogen), L-glutamine (Invitrogen,2 mmol/L), -mercaptoethanol (Serva, 50 mol/L), nonessentialamino acids (Invitrogen, stock solution diluted 1:100), and 103
units/mL recombinant human leukemia inhibitory factor (Chemicon)(ie, standard ESC culture conditions) as previously described.23
Standard ESC culture conditions were used for cultivation of mouseMPI-II ESCs.
For differentiation of maGSCs, the hanging drop method de-scribed for mouse ESC differentiation25 was applied for the forma-tion of embryoid bodies (EBs). See the expanded Materials andMethods section in the online data supplement, at http://circres.ahajournals.org, for details on EB differentiation protocols, in vitroresponse of beating EBs to pharmacological agents, isolation ofcardiomyocytes, immunocytochemistry, fluorescence recovery afterphotobleaching assay, action potential (AP) measurements, intracel-lular calcium measurements using confocal laser microscopy, andRT-PCR analysis.
Transplantation of maGSCsThe present study used female C57BL/6 mice obtained from theanimal facility of the University of Gottingen. The investigationconformed to the Guide for the Care and Use of Laboratory Animalspublished by the NIH (NIH Publication No. 85-23, revised 1996) andwas approved by the local government authority. Mouse maGSCs (lineSSC5) labeled with a fluorescent carbocyanine dye CM-DiI(chloromethyl–1,1-dioctadecyl-3,3,3,3-tetramethylindocarbocyanine)(Invitrogen) were injected in the anterolateral wall of the heart or in thejugular vein. Hearts at 2 days and 1 and 4 weeks postinjection wereanalyzed by histochemical and immunohistological staining. Detailed
information is available in the expanded Materials and Methods sectionin the online data supplement.
ResultsInduction of Cardiac DifferentiationOur initial studies show that maGSC lines (SSC5, SSC6, andSSC10) derived from double transgenic mice (Stra8-EGFP/Rosa26) can spontaneously differentiate into contractingcardiac clusters.23 Here we report that maGSCs derived fromwild-type mice of 3 different strains (C57BL6, FVB, and129/Ola) can also spontaneously differentiate into cardio-myocytes by inducing EB formation (Figure 1A through 1C).Spontaneously and rhythmically contracting cells appeared asclusters and were identified in approximately 40% of indi-vidual EBs (n144; 3 independent experiments) derivedfrom C57BL6 line 9 at day 52 (ie, 2 days after plating of5-day-old EBs), increased to as many as 80% of the EBs byday 58, and declined to 50% by day 517 (Figure 1C). Thesize of beating cardiac clusters was 5% to 20% of EBoutgrowths by day 58. The percentages of beating EBsderived from C57BL6 line 9 during differentiation wassimilar to those derived from line SSC5 and ESCs (Figure1C). Similar results were seen in FVB line 5 (Figure 1C).However, lower differentiation efficiency was seen in 129/Ola line 2 in comparison with line SSC5 and ESCs (Figure1C). The remainder of the experiments then focused on EBsor cardiomyocytes derived from C57BL6 line 9 and SSC5.RT-PCR assays showed that cardiomyocytes derived fromC57BL6 line 9, similar to those derived from SSC5,23
expressed cardiac gene products in a developmentally con-trolled manner (data not shown).
Functional L-type Calcium Channels ofmaGSC-Derived CardiomyocytesIn cardiac muscle, where Ca2 influx across the sarcolemmais essential for contraction, the dihydropyridine-sensitiveL-type calcium channel represents the major entry pathway ofextracellular Ca2.26 Using immunofluorescence staining witha specific antibody, we could show that the 1 subunit ofL-type calcium channels was expressed in cardiomyocytes atday 56 in a striated pattern (Figure 1D through 1F). Thefunction of L-type calcium channels was examined in beatingcardiac clusters at day 515 by evaluating chronotropiceffects of cardioactive drugs. The L-type calcium channelactivator (S)-BayK 8644 (1,4 dihydropyridine-type) showeddose-dependent positive chronotropic effects on the beatingfrequency of maGSC-derived cardiomyocytes, whereas theL-type calcium channel blocker diltiazem (1,5-benzothiazepine-type) showed dose-dependent negative chro-notropic effects (Figure 1G). Treatment with 105 mol/L(S)-BayK 8644 doubled the beating frequency. Treatmentwith 105 mol/L diltiazem almost completely blocked thecontractions. Contractions recovered to a normal frequency24 hours after removal of the drug. These results indicate thatfunctional L-type calcium channels exist in the maGSC-derived cardiomyocytes.
Functional Cell-to-Cell CommunicationTo analyze the cell-to-cell coupling in the synchronouslycontracting cardiac clusters, we performed double-staining
1616 Circulation Research June 8, 2007
by guest on February 23, 2016http://circres.ahajournals.org/Downloaded from

using antibodies against either pan-cadherin and -actinin orconnexin 43 (Cx43) and cardiac troponin T. We found thatpan-cadherin localized at cell–cell junctions, indicating thepresence of adhering junctions between cardiomyocytes (Fig-ure 2A through 2C). In addition, Cx43 staining indicated thepresence of gap junctions between cells in cardiac clusters(Figure 2D through 2F). Functional coupling between cellswas confirmed by fluorescence recovery after photobleachinganalysis. Fluorescence recovery of a gap junction–permeabledye (calcein–acetoxy methyl ester [AM]) was observed con-
sistently for cardiomyocytes within 5 minutes after photo-bleaching (Figure 3A). When cells were exposed to carben-oxolone (50 mol/L), a gap junction uncoupler, thefluorescence recovery was disrupted (Figure 3B). Fluores-cence in cardiomyocytes recovered to 25.65.6% (n15)after 30 seconds and carbenoxolone significantly blocked thepercentage of refill to 4.32.4% (n9; P0.05) (Figure 3C).The magnitude of functional gap junctions (or the gapjunction permeability) was assessed by the rate of fluores-cence recovery (k), which reflected the diffusion of
Figure 1. Cardiac differentiation ofmouse maGSCs. A and B, Undifferenti-ated maGSCs (A) differentiated into car-diomyocytes by inducing formation ofEBs (B). C, Percentage of EBs contain-ing beating cardiac clusters during dif-ferentiation. D and E, Double immuno-staining of cardiac cells by antibodiesagainst sarcomeric myosin heavy chain(D) and cardiac-specific L-type calciumchannels (E). F, Overlay of D and E.Nuclear staining with 4,6-diamidino-2-phenylindole (DAPI). G, Effect of diltiazemand (S)-BayK 8644 on the beating fre-quency of cardiomyocytes derived frommaGSCs. *P0.05, **P0.01, ***P0.001.Scale bar: 100 m (A and B); 25 m (Dthrough F).
Guan et al Spermatogonial Stem Cell–Derived Cardiomyocytes 1617
by guest on February 23, 2016http://circres.ahajournals.org/Downloaded from

calcein-AM from unbleached neighbors into a laser-bleachedcell. The recovery rate in maGSC-derived cardiomyocyteswas 0.470.06 min1 (n15), and the application of carben-oxolone resulted in a significantly lower rate of recovery(k0.040.04 min1; n7; P0.05; Figure 3D). Theseobservations suggest that gap junctions function as a conduitof intercellular communication between cardiomyocytes andplay an important role in cell-to-cell communication, essen-tial for the synchronization of myocardial contractile activityand intact electromechanical coupling.
Action Potential CharacteristicsIn mature cardiac cells, depolarization of the cell membraneduring the AP activates L-type Ca2 channels, leading to Ca2
influx and subsequent release of Ca2 from intracellularcalcium stores. To characterize whether mouse maGSC-derived cardiomyocytes could enter a fully differentiatedcardiac phenotype, the dissociated cardiomyocytes were an-alyzed by patch–clamp. Mouse maGSC-derived cardiomyo-cytes showed spontaneous APs (Figure 4A). We examinedthe shape and properties of APs from 68 single beatingcardiomyocytes. Four major types of APs characteristic for
pacemaker- (n8), ventricle- (n23), atrial- (n9), andPurkinje-like (n9) cells were found (Figure 4A) withdistinct morphologies at day 59. This classification wasbased on the shapes (Figure 4A) and the properties of the APas measured by upstroke velocity (dV/dtmax), AP amplitude(APA), AP duration at 90% and 80% of repolarization, andmaximum diastolic potential as summarized in the supple-mental Table I. Pacemaker-like APs are characterized byprominence of phase 4 depolarization, slow dV/dtmax, and asmaller APA. The ventricle-like APs can be distinguished bythe presence of a significant plateau phase of the AP,resulting in a long duration, and high dV/dtmax and APA. Theatrial-like APs show a triangular shape with a short durationand high dV/dtmax and APA. The Purkinje-like APs arecharacterized by the presence of a notch and plateau-phaseand high dV/dtmax. In addition, a subset of cells (n19)showed an intermediate AP phenotype (Figure 4B), whichexhibited characteristics of both ventricle- and pacemaker-like morphology (slow dV/dtmax, long duration; supplementalTable I).
To determine the functional expression of -adrenergicreceptors in cardiomyocytes, we studied the effects of-adrenergic agonist isoproterenol (1 mol/L) on APs. Theisoproterenol stimulation resulted in the significant increaseof AP frequency from 0.800.15 to 1.620.21 Hz (n5;P0.05; Figure 4C), demonstrating that -adrenergic recep-tors are present in maGSC-derived cardiomyocytes and stim-ulation of these receptors produces a positive chronotropicresponse. Furthermore, cadmium (0.5 mmol/L), a nonspecificblocker of voltage-gated Na and L-type Ca2 channels,completely abolished spontaneous APs (Figure 4D), provingthat Na and Ca2 channels critically contribute to theobserved APs. A further hint for the involvement of fastdepolarizing Na channels are AP upstroke velocities in themagnitude of 30 to 50 V/s1 for ventricle-, Purkinje-, andatrial-like APs (supplemental Table I). In particular,ventricle- and Purkinje-like APs showed plateau phasesobviously caused by Ca2 channel currents.
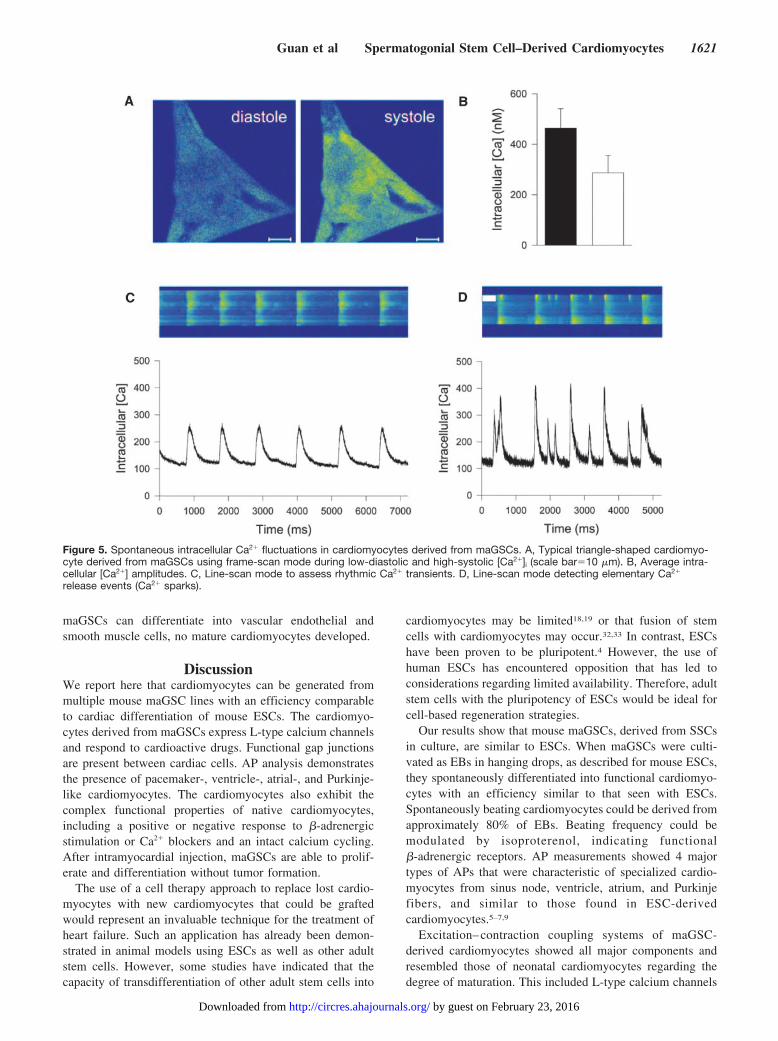
Ca2 Transients and Ca2 SparksWe assessed the spontaneous intracellular Ca2 fluctuationsin maGSC-derived cardiomyocytes using confocal micros-copy. A typical triangle-shaped cardiomyocyte during low-diastolic [Ca2]i and high-systolic [Ca2]i is presented inFigure 5A. Calcium increased homogenously throughout thecell, pointing to a fine-regulated Ca2 release from intracel-lular Ca2 stores, most likely the sarcoplasmic reticulum (SR)(supplemental Figure I; supplemental Video 1). The ampli-tudes of Ca2 transient measured in maGSC-derived cardio-myocytes were 46477 nmol/L (filled bar; n15) versus28769 nmol/L measured in ESC-derived cardiomyocytes(open bar; n5; statistically not significant; Figure 5B). Bothare in the range of amplitudes measured in adult cardiomyo-cytes.27 Using line-scan mode, rhythmic Ca2 transients werefound (Figure 5C) and even small elementary Ca2 releaseevents (Ca2 sparks28; Figure 5D), which are mainly attrib-utable to SR Ca2 release through a cluster of ryanodinereceptors (RyRs), appeared. Ca2 sparks were previously
Figure 2. Double immunostaining of cardiac clusters. A and B,Cardiac cluster (day 511) stained with sarcomeric -actinin (A)and pan-cadherin (B). C, Overlay of A and B. D and E, Cardiaccluster (day-57) stained with cardiac troponin T (D) and Cx43(E). F, Overlay of D and E. Scale bar25 m. Nuclear stainingwith DAPI.
1618 Circulation Research June 8, 2007
by guest on February 23, 2016http://circres.ahajournals.org/Downloaded from

described in increasing numbers at later stages of cardiomyo-cytes derived from ESCs.29
The cardiac RyR2 serves as the major SR calcium-releasechannel to mediate the rapid rise of cytosolic free calcium.We found that the gene encoding RyR2 was expressed duringEB differentiation (Figure 6A). In addition, genes encodingphospholamban, SERCA2a, and Na/Ca2 exchanger(NCX)1, as well as calsequestrin, a high-capacity Ca2-binding protein in the SR, were expressed (Figure 6A). Thisexpression pattern was similar to those described for cardiacdifferentiation of ESCs.5 To study the distribution of RyR2and the Ca2-handling proteins SERCA2a and NCX1throughout cardiomyocytes, we performed double staining inmaGSC-derived cardiomyocytes at day 518. We foundmainly a diffuse distribution of RyR2 in maGSC-derivedcardiomyocytes (Figure 6B). Of note, we observed partialorganization of RyR2 in striated-like structures (Figure 6B,arrow), which are not found in cardiomyocytes until postnatalday 6 when SR begins to organize.30 Immunocytochemicalstaining revealed that SERCA2a and NCX1 were mainlyexpressed in a fine granular, network-like pattern throughoutcardiomyocytes (Figure 6C and 6D) comparable to those ofneonatal cardiomyocytes.30,31 These results suggest that theorganization of the SR is not fully developed in maGSC-derived cardiomyocytes.
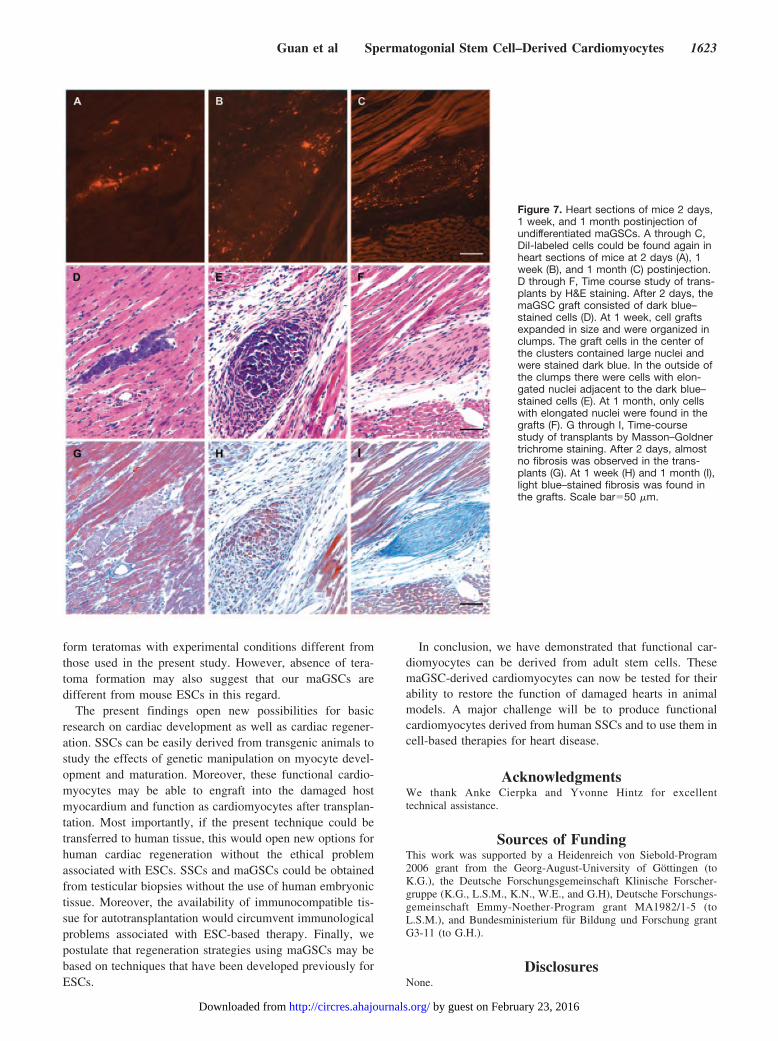
Effect of Cardiac Environment on Development ofmaGSCs In VivoWe further tested the effect of cardiac environment on thedevelopment of maGSCs in vivo. CM-DiI–labeled maGSCs(0.5 to 1106) were injected into the left ventricular free wallof female C57BL/6 mice. Given that the transplanted cellswere DiI-labeled, whereas the myocardium of the recipientmice was not, the fate of donor cells was readily monitored byfluorescence microscopic examination. Two days after cellapplication, DiI-labeled cells could be found again in mice(n5; Figure 7A). Hematoxylin/eosin (H&E) stainingshowed that these cells in small clusters were stained in blue(Figure 7D). One week after cell application, DiI-labeledcells could be found again in the recipients (n4; Figure 7B).This data were confirmed by H&E staining of heart sectionsrevealing that one week post-injection of cells, left ventricularregions of mice frequently contained larger clusters of cellswith large nuclei which were stained dark blue (Figure 7E).One month after cell injection, DiI-labeled cells could still befound in heart sections of mice (n6; Figure 7C). However,they no longer contained large nuclei and were not staineddark blue (Figure 7F). Furthermore, none of the transplantedhearts showed evidence of tumor or teratoma formation onhistological evaluation.
In addition, we tested whether cells could migrate intohearts after intravenous cell delivery. We found that only very
Figure 3. Functional gap junctions in car-diac clusters. A and B, Confocal images ofcardiac clusters loaded with the gap junc-tion permeable dye (calcein-AM) before(Pre), immediately after (arrow, 0 minutes),and 5-minute after photobleaching of a sin-gle cell (scale bar10 m). A, After 5 min-utes of recovery, the decreased fluores-cence intensity of the bleached cell isalmost completely reversed, which is con-sistent with dye refill from adjacent cellsthrough functional gap junctions. B, In thepresence of the gap junction uncoupler car-benoxolone (50 mol/L), the recovery of flu-orescence after photobleaching was almostcompletely abolished. C and D, Mean datafor the magnitude of refill after 30 secondsrecovery (C) and the recovery rate (k) over 5to 10 minutes (D). Compared with vehicle,both the dye refill after 30 seconds recoveryand the recovery rate within 5 to 10 minuteswas significantly reduced in the presence ofcarbenoxolone.
Guan et al Spermatogonial Stem Cell–Derived Cardiomyocytes 1619
by guest on February 23, 2016http://circres.ahajournals.org/Downloaded from

a few single cells could be detected at 2 days after application(data not shown).
The fate of donor cells following intramyocardial injectionwas determined using immunohistochemical assays. In trans-planted hearts at 1 week after cell injection, immunolabelingwith an antibody specific for Oct4, a germline-specifictranscription factor often used to characterize pluripotentstem cells, revealed large aggregates of labeled cells in themyocardium (Figure 8A). In a parallel section, the Oct4-positive cell aggregates were also stained dark blue by H&Estaining (date not shown), suggesting the presence of undif-ferentiated and nonmature cells in the left ventricles ofrecipients. As shown in Figure 7E, there were cells withelongated nuclei adjacent to the dark blue–stained cells. Theywere Oct4 negative (Figure 8A), suggesting that these cellslost their pluripotency. However, when the sections werestained with an antibody against the cell proliferation markerKi-67, we found that some cells with elongated nuclei werepositive for Ki-67 (Figure 8C), indicating that they still havethe proliferation potential. One month after cell transplanta-tion, no Oct4- or Ki-67–positive cells were found (Figure 8B
and 8D). Immunolabeling for cardiac troponin T showed thatnone of the transplanted cells expressed cardiac troponin T ateither 1 week (supplemental Figure IIA through IIC) or 1month (supplemental Figure IID through IIF). When thesections at 1 month were stained with antibodies against vonWillebrand factor (vWF) and smooth muscle--actin (SM--actin), we found that some of labeled cells were positive forvWF (Figure 8E through 8G) and SM--actin (Figure 8H and8I), indicating that the transplanted maGSCs could differen-tiate into vascular endothelial and smooth muscle cells. Inaddition, Masson–Goldner trichrome staining showed thatfibrosis (light blue staining) developed in the heart at 1 week(Figure 7H) and 1 month (Figure 7I) postinjection of cells,whereas almost no fibrosis was visible in the transplants at 2days (Figure 7G).
Taken together, these findings demonstrate that trans-planted maGSCs are able to proliferate and differentiate inthe normal heart. Some cells lost their pluripotency but stillwere proliferating 1 week after cell delivery. After 1 month,no proliferating cells were observed, and until this time, notumor or teratomas were formed. Although transplanted
Figure 4. AP characteristics ofspontaneously contracting car-diomyocytes at day 59. A,Exemplary original traces of APs.Distinct AP morphologies repre-senting pacemaker-, ventricle-,atrial-, and Purkinje-like cardio-myocytes could be discriminated.B, In a subset of cardiomyocytes,an intermediate phenotypebetween pacemaker- andventricle-like APs was found. C,Application of isoproterenol (Iso)(1 mol/L) resulted in an increasein AP frequency and irregularity.D, Nonspecific blockade ofL-type Ca2 and Na channelswith cadmium (0.5 mmol/L) com-pletely abolished cardiac APs.
1620 Circulation Research June 8, 2007
by guest on February 23, 2016http://circres.ahajournals.org/Downloaded from
maGSCs can differentiate into vascular endothelial andsmooth muscle cells, no mature cardiomyocytes developed.
DiscussionWe report here that cardiomyocytes can be generated frommultiple mouse maGSC lines with an efficiency comparableto cardiac differentiation of mouse ESCs. The cardiomyo-cytes derived from maGSCs express L-type calcium channelsand respond to cardioactive drugs. Functional gap junctionsare present between cardiac cells. AP analysis demonstratesthe presence of pacemaker-, ventricle-, atrial-, and Purkinje-like cardiomyocytes. The cardiomyocytes also exhibit thecomplex functional properties of native cardiomyocytes,including a positive or negative response to -adrenergicstimulation or Ca2 blockers and an intact calcium cycling.After intramyocardial injection, maGSCs are able to prolif-erate and differentiation without tumor formation.
The use of a cell therapy approach to replace lost cardio-myocytes with new cardiomyocytes that could be graftedwould represent an invaluable technique for the treatment ofheart failure. Such an application has already been demon-strated in animal models using ESCs as well as other adultstem cells. However, some studies have indicated that thecapacity of transdifferentiation of other adult stem cells into
cardiomyocytes may be limited18,19 or that fusion of stemcells with cardiomyocytes may occur.32,33 In contrast, ESCshave been proven to be pluripotent.4 However, the use ofhuman ESCs has encountered opposition that has led toconsiderations regarding limited availability. Therefore, adultstem cells with the pluripotency of ESCs would be ideal forcell-based regeneration strategies.
Our results show that mouse maGSCs, derived from SSCsin culture, are similar to ESCs. When maGSCs were culti-vated as EBs in hanging drops, as described for mouse ESCs,they spontaneously differentiated into functional cardiomyo-cytes with an efficiency similar to that seen with ESCs.Spontaneously beating cardiomyocytes could be derived fromapproximately 80% of EBs. Beating frequency could bemodulated by isoproterenol, indicating functional-adrenergic receptors. AP measurements showed 4 majortypes of APs that were characteristic of specialized cardio-myocytes from sinus node, ventricle, atrium, and Purkinjefibers, and similar to those found in ESC-derivedcardiomyocytes.5–7,9
Excitation– contraction coupling systems of maGSC-derived cardiomyocytes showed all major components andresembled those of neonatal cardiomyocytes regarding thedegree of maturation. This included L-type calcium channels
Figure 5. Spontaneous intracellular Ca2 fluctuations in cardiomyocytes derived from maGSCs. A, Typical triangle-shaped cardiomyo-cyte derived from maGSCs using frame-scan mode during low-diastolic and high-systolic [Ca2]i (scale bar10 m). B, Average intra-cellular [Ca2] amplitudes. C, Line-scan mode to assess rhythmic Ca2 transients. D, Line-scan mode detecting elementary Ca2
release events (Ca2 sparks).
Guan et al Spermatogonial Stem Cell–Derived Cardiomyocytes 1621
by guest on February 23, 2016http://circres.ahajournals.org/Downloaded from

sensitive to the calcium channel activator (S)-BayK8644 andthe calcium channel blocker diltiazem, as well as calciumbinding and cycling proteins, such as RyR, calsequestrin,sarcoplasmic reticulum calcium pump, phospholamban, andsarcolemmal sodium calcium exchanger.
We also observed organization of RyRs in a striated-likestructure in maGSC-derived cardiomyocytes, similar to thosein postnatal cardiomyocytes.30 Development of SR and func-tional excitation–contraction coupling is obvious from thepresence of Ca2 sparks and Ca2 transients in maGSC-derived cardiomyocytes. Critically important for cardiacfunction, we show that maGSCs derived cardiomyocytesexpress the gap junction protein Cx43 and develop cell-to-cell coupling. Again, cardiomyocytes derived from maGSCswere structurally and functionally comparable to those ob-tained from ESCs.
The finding that transplanted maGSCs are able to prolif-erate in the heart is consistent with the known self-renewalproperty of the cell source. We observed that maGSCstransplanted into normal hearts lost their proliferating poten-tial at 1 month and differentiated into vascular endothelialand smooth muscle cells but did not differentiate into cardio-myocytes. When maGSCs were cultured as EBs, whichresemble early postimplantation embryos, they were able tospontaneously differentiate into functional cardiomyocytes invitro at day 58.23 However, they could not spontaneously
differentiate into cardiomyocytes in the normal heart at 1month postinjection of cells. It is possible that cardiacdifferentiation may occur at later time points that have notbeen investigated yet. Alternatively, local microenvironmentmight be a critical determinate of the fate of the transplantedmaGSCs. This is also in line with previous studies showingthat in the absence of myocardial infarction, only raretransplanted ESCs remained in the heart at 2 weeks.34
However, undifferentiated mouse ESCs transplanted directlyinto the infarcted heart could differentiate into cardiomyo-cytes and vascular smooth muscle and endothelialcells.11,12,34,35 It will be interesting to investigate whethermaGSCs or predifferentiated maGSCs can contribute to thecardiac and vascular lineages in the absence of fusion aftercellular transplantation into the infarcted heart.
Although maGSCs proliferated after transplantation, underthe present conditions, no teratoma/tumor formation wasobserved, despite the known ability of these cells to formteratomas under certain conditions. This seems consistentwith earlier studies transplanting mouse ESCs into rat ormouse hearts.11,34,35 However, this is in contrast to a recentstudy showing that undifferentiated ESCs formed teratomasin both normal and infarcted hearts of nude or immunocom-petent syngeneic mice. Even allogenic ESCs caused terato-mas, but these were immunologically rejected after severalweeks.36 Of course, we cannot exclude that maGSCs may
Figure 6. Expression of calcium handling proteins.A, RT-PCR analysis of genes encoding RyR2,phospholamban (PLB), SERCA2a, NCX1, andcalsequestrin (CASQ) during differentiation of EBsafter plating at day 5 (d5). M indicates DNAmarker; d0, maGSCs before EB formation; FL,feeder layer. B through D, RyR2 (B), NCX1 (C), andSERCA2a (D) staining in cardiac cells (day 57)stained with sarcomeric -actinin (B through D)(inset, green). Scale bar25 m. Nuclear stainingwith DAPI.
1622 Circulation Research June 8, 2007
by guest on February 23, 2016http://circres.ahajournals.org/Downloaded from
form teratomas with experimental conditions different fromthose used in the present study. However, absence of tera-toma formation may also suggest that our maGSCs aredifferent from mouse ESCs in this regard.
The present findings open new possibilities for basicresearch on cardiac development as well as cardiac regener-ation. SSCs can be easily derived from transgenic animals tostudy the effects of genetic manipulation on myocyte devel-opment and maturation. Moreover, these functional cardio-myocytes may be able to engraft into the damaged hostmyocardium and function as cardiomyocytes after transplan-tation. Most importantly, if the present technique could betransferred to human tissue, this would open new options forhuman cardiac regeneration without the ethical problemassociated with ESCs. SSCs and maGSCs could be obtainedfrom testicular biopsies without the use of human embryonictissue. Moreover, the availability of immunocompatible tis-sue for autotransplantation would circumvent immunologicalproblems associated with ESC-based therapy. Finally, wepostulate that regeneration strategies using maGSCs may bebased on techniques that have been developed previously forESCs.
In conclusion, we have demonstrated that functional car-diomyocytes can be derived from adult stem cells. ThesemaGSC-derived cardiomyocytes can now be tested for theirability to restore the function of damaged hearts in animalmodels. A major challenge will be to produce functionalcardiomyocytes derived from human SSCs and to use them incell-based therapies for heart disease.
AcknowledgmentsWe thank Anke Cierpka and Yvonne Hintz for excellenttechnical assistance.
Sources of FundingThis work was supported by a Heidenreich von Siebold-Program2006 grant from the Georg-August-University of Gottingen (toK.G.), the Deutsche Forschungsgemeinschaft Klinische Forscher-gruppe (K.G., L.S.M., K.N., W.E., and G.H), Deutsche Forschungs-gemeinschaft Emmy-Noether-Program grant MA1982/1-5 (toL.S.M.), and Bundesministerium fur Bildung und Forschung grantG3-11 (to G.H.).
DisclosuresNone.
Figure 7. Heart sections of mice 2 days,1 week, and 1 month postinjection ofundifferentiated maGSCs. A through C,DiI-labeled cells could be found again inheart sections of mice at 2 days (A), 1week (B), and 1 month (C) postinjection.D through F, Time course study of trans-plants by H&E staining. After 2 days, themaGSC graft consisted of dark blue–stained cells (D). At 1 week, cell graftsexpanded in size and were organized inclumps. The graft cells in the center ofthe clusters contained large nuclei andwere stained dark blue. In the outside ofthe clumps there were cells with elon-gated nuclei adjacent to the dark blue–stained cells (E). At 1 month, only cellswith elongated nuclei were found in thegrafts (F). G through I, Time-coursestudy of transplants by Masson–Goldnertrichrome staining. After 2 days, almostno fibrosis was observed in the trans-plants (G). At 1 week (H) and 1 month (I),light blue–stained fibrosis was found inthe grafts. Scale bar50 m.
Guan et al Spermatogonial Stem Cell–Derived Cardiomyocytes 1623
by guest on February 23, 2016http://circres.ahajournals.org/Downloaded from

References1. Beltrami AP, Barlucchi L, Torella D, Baker M, Limana F, Chimenti S,
Kasahara H, Rota M, Musso E, Urbanek K, Leri A, Kajstura J, Nadal-Ginard B, Anversa P. Adult cardiac stem cells are multipotent and supportmyocardial regeneration. Cell. 2003;114:763–776.
2. Oh H, Bradfute SB, Gallardo TD, Nakamura T, Gaussin V, Mishina Y,Pocius J, Michael LH, Behringer RR, Garry DJ, Entman ML, SchneiderMD. Cardiac progenitor cells from adult myocardium: homing, differen-tiation, and fusion after infarction. Proc Natl Acad Sci U S A. 2003;100:12313–12318.
3. Laugwitz KL, Moretti A, Lam J, Gruber P, Chen Y, Woodard S, Lin LZ,Cai CL, Lu MM, Reth M, Platoshyn O, Yuan JX, Evans S, Chien KR.Postnatal isl1 cardioblasts enter fully differentiated cardiomyocytelineages. Nature. 2005;433:647–653.
4. Wobus AM, Boheler KR. Embryonic stem cells: prospects for develop-mental biology and cell therapy. Physiol Rev. 2005;85:635–678.
5. Boheler KR, Czyz J, Tweedie D, Yang HT, Anisimov SV, Wobus AM.Differentiation of pluripotent embryonic stem cells into cardiomyocytes.Circ Res. 2002;91:189–201.
6. Wobus AM, Rohwedel J, Maltsev V, Hescheler J. Development of car-diomyocytes expressing cardiac-specific genes, action potentials, andionic channels during embryonic stem cell-derived cardiogenesis. Ann NY Acad Sci. 1995;752:460–469.
7. Hescheler J, Fleischmann BK, Lentini S, Maltsev VA, Rohwedel J,Wobus AM, Addicks K. Embryonic stem cells: a model to study
structural and functional properties in cardiomyogenesis. Cardiovasc Res.1997;36:149–162.
8. Guan K, Furst DO, Wobus AM. Modulation of sarcomere organizationduring embryonic stem cell-derived cardiomyocyte differentiation. EurJ Cell Biol. 1999;78:813–823.
9. Maltsev VA, Rohwedel J, Hescheler J, Wobus AM. Embryonic stem cellsdifferentiate in vitro into cardiomyocytes representing sinusnodal, atrialand ventricular cell types. Mech Dev. 1993;44:41–50.
10. He JQ, Ma Y, Lee Y, Thomson JA, Kamp TJ. Human embryonic stemcells develop into multiple types of cardiac myocytes: action potentialcharacterization. Circ Res. 2003;93:32–39.
11. Hodgson DM, Behfar A, Zingman LV, Kane GC, Perez-Terzic C, Alekseev AE,Puceat M, Terzic A. Stable benefit of embryonic stem cell therapy in myocardialinfarction. Am J Physiol Heart Circ Physiol. 2004;287:H471–H479.
12. Behfar A, Zingman LV, Hodgson DM, Rauzier JM, Kane GC, Terzic A,Puceat M. Stem cell differentiation requires a paracrine pathway in theheart. FASEB J. 2002;16:1558–1566.
13. Klug MG, Soonpaa MH, Koh GY, Field LJ. Genetically selected cardio-myocytes from differentiating embryonic stem cells form stable intra-cardiac grafts. J Clin Invest. 1996;98:216–224.
14. Menard C, Hagege AA, Agbulut O, Barro M, Morichetti MC, BrasseletC, Bel A, Messas E, Bissery A, Bruneval P, Desnos M, Puceat M,Menasche P. Transplantation of cardiac-committed mouse embryonicstem cells to infarcted sheep myocardium: a preclinical study. Lancet.2005;366:1005–1012.
Figure 8. The fate of graft cells following intramyo-cardial injection. A through D, Histological sectionsof engrafted hearts showed Oct4- (A) and Ki-67-positive (C) cells in the clumps at 1 week. At 1month, the cells in the clumps were no longer pos-itive for Oct4 (B) and Ki-67 (D). E through J, Immu-nostaining of the transplants using antibodiesagainst vWF (E through G) and SM--actin (Hthrough J). DiI-labeled transplants (E and H) werepositive for vWF (F) and SM--actin (I) at 1 month.G, Overlay of DiI labeling (E) and vWF staining (F).J, Overlay of DiI labeling (H) and SM--actin stain-ing (I). Scale bar50 m. Nuclear staining withDAPI.
1624 Circulation Research June 8, 2007
by guest on February 23, 2016http://circres.ahajournals.org/Downloaded from

15. Orlic D, Kajstura J, Chimenti S, Limana F, Jakoniuk I, Quaini F, Nadal-Ginard B, Bodine DM, Leri A, Anversa P. Mobilized bone marrow cellsrepair the infarcted heart, improving function and survival. Proc NatlAcad Sci U S A. 2001;98:10344–10349.
16. Orlic D, Kajstura J, Chimenti S, Jakoniuk I, Anderson SM, Li B, PickelJ, McKay R, Nadal-Ginard B, Bodine DM, Leri A, Anversa P. Bonemarrow cells regenerate infarcted myocardium. Nature. 2001;410:701–705.
17. Jackson KA, Majka SM, Wang H, Pocius J, Hartley CJ, Majesky MW,Entman ML, Michael LH, Hirschi KK, Goodell MA. Regeneration ofischemic cardiac muscle and vascular endothelium by adult stem cells.J Clin Invest. 2001;107:1395–1402.
18. Balsam LB, Wagers AJ, Christensen JL, Kofidis T, Weissman IL,Robbins RC. Haematopoietic stem cells adopt mature haematopoieticfates in ischaemic myocardium. Nature. 2004;428:668–673.
19. Murry CE, Soonpaa MH, Reinecke H, Nakajima H, Nakajima HO, RubartM, Pasumarthi KB, Virag JI, Bartelmez SH, Poppa V, Bradford G,Dowell JD, Williams DA, Field LJ. Haematopoietic stem cells do nottransdifferentiate into cardiac myocytes in myocardial infarcts. Nature.2004;428:664–668.
20. Makino S, Fukuda K, Miyoshi S, Konishi F, Kodama H, Pan J, Sano M,Takahashi T, Hori S, Abe H, Hata J, Umezawa A, Ogawa S. Cardiomy-ocytes can be generated from marrow stromal cells in vitro. J Clin Invest.1999;103:697–705.
21. Badorff C, Brandes RP, Popp R, Rupp S, Urbich C, Aicher A, Fleming I,Busse R, Zeiher AM, Dimmeler S. Transdifferentiation of blood-derivedhuman adult endothelial progenitor cells into functionally active cardio-myocytes. Circulation. 2003;107:1024–1032.
22. Gruh I, Beilner J, Blomer U, Schmiedl A, Schmidt-Richter I, Kruse ML,Haverich A, Martin U. No evidence of transdifferentiation of humanendothelial progenitor cells into cardiomyocytes after coculture withneonatal rat cardiomyocytes. Circulation. 2006;113:1326–1334.
23. Guan K, Nayernia K, Maier LS, Wagner S, Dressel R, Lee JH, Nolte J,Wolf F, Li M, Engel W, Hasenfuss G. Pluripotency of spermatogonialstem cells from adult mouse testis. Nature. 2006;440:1199–1203.
24. Spradling A, Drummond-Barbosa D, Kai T. Stem cells find their niche.Nature. 2001;414:98–104.
25. Wobus AM, Guan K, Yang HT, Boheler KR. Embryonic stem cells as amodel to study cardiac, skeletal muscle, and vascular smooth muscle celldifferentiation. Methods Mol Biol. 2002;185:127–156.
26. Bers DM. Cardiac excitation-contraction coupling. Nature. 2002;415:198–205.
27. Maier LS, Zhang T, Chen L, DeSantiago J, Brown JH, Bers DM.Transgenic CaMKIIdeltaC overexpression uniquely alters cardiacmyocyte Ca2 handling: reduced SR Ca2 load and activated SR Ca2release. Circ Res. 2003;92:904–911.
28. Cheng H, Lederer WJ, Cannell MB. Calcium sparks: elementary eventsunderlying excitation-contraction coupling in heart muscle. Science.1993;262:740–744.
29. Sauer H, Theben T, Hescheler J, Lindner M, Brandt MC, Wartenberg M.Characteristics of calcium sparks in cardiomyocytes derived fromembryonic stem cells. Am J Physiol Heart Circ Physiol. 2001;281:H411–H421.
30. Mohler PJ, Gramolini AO, Bennett V. The ankyrin-B C-terminal domaindetermines activity of ankyrin-B/G chimeras in rescue of abnormalinositol 1,4,5-trisphosphate and ryanodine receptor distribution inankyrin-B (-/-) neonatal cardiomyocytes. J Biol Chem. 2002;277:10599–10607.
31. Most P, Boerries M, Eicher C, Schweda C, Volkers M, Wedel T, SollnerS, Katus HA, Remppis A, Aebi U, Koch WJ, Schoenenberger CA.Distinct subcellular location of the Ca2-binding protein S100A1 dif-ferentially modulates Ca2-cycling in ventricular rat cardiomyocytes.J Cell Sci. 2005;118:421–431.
32. Terada N, Hamazaki T, Oka M, Hoki M, Mastalerz DM, Nakano Y,Meyer EM, Morel L, Petersen BE, Scott EW. Bone marrow cells adoptthe phenotype of other cells by spontaneous cell fusion. Nature. 2002;416:542–545.
33. Ying QL, Nichols J, Evans EP, Smith AG. Changing potency by spon-taneous fusion. Nature. 2002;416:545–548.
34. Singla DK, Hacker TA, Ma L, Douglas PS, Sullivan R, Lyons GE, KampTJ. Transplantation of embryonic stem cells into the infarcted mouseheart: formation of multiple cell types. J Mol Cell Cardiol. 2006;40:195–200.
35. Min JY, Yang Y, Converso KL, Liu L, Huang Q, Morgan JP, Xiao YF.Transplantation of embryonic stem cells improves cardiac function inpostinfarcted rats. J Appl Physiol. 2002;92:288–296.
36. Nussbaum J, Minami E, Laflamme MA, Virag JA, Ware CB, Masino A,Muskheli V, Pabon L, Reinecke H, Murry CE. Transplantation of undif-ferentiated murine embryonic stem cells in the heart: teratoma formationand immune response. FASEB J. 2007.
Guan et al Spermatogonial Stem Cell–Derived Cardiomyocytes 1625
by guest on February 23, 2016http://circres.ahajournals.org/Downloaded from

Hemmerlein, Karim Nayernia, Wolfgang Engel and Gerd HasenfussKaomei Guan, Stefan Wagner, Bernhard Unsöld, Lars S. Maier, Diana Kaiser, Bernhard
Generation of Functional Cardiomyocytes From Adult Mouse Spermatogonial Stem Cells
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 2007 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/01.RES.0000269182.22798.d92007;100:1615-1625; originally published online May 3, 2007;Circ Res.
http://circres.ahajournals.org/content/100/11/1615World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org/content/suppl/2007/05/06/01.RES.0000269182.22798.d9.DC1.htmlData Supplement (unedited) at:
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Researchin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on February 23, 2016http://circres.ahajournals.org/Downloaded from

1
Materials and Methods
Cell Culture and Differentiation of Embryoid Bodies
Established maGSC cultures (SSC5, C57BL6 line 9, FVB line 5 and 129 line 2) from different
mouse strains were cultivated on a feeder layer of primary mouse embryonic fibroblasts on
gelatin (0.1%)-coated petri dishes (Nunc) in Dulbecco’s modified Eagle’s minimal essential
medium (DMEM, 4.5 g/L glucose; Invitrogen) supplemented by 15% heat-inactivated fetal
calf serum (FCS, selected batches, Invitrogen), L-glutamine (Invitrogen, 2 mmol/L), β-
mercaptoethanol (β-ME, Serva, final concentration 50 µmol/L), non-essential amino acids
(NEAA, Invitrogen, stock solution diluted 1:100) and 103 Units/ml recombinant human
leukemia inhibitory factor (LIF; ESGRO, Chemicon) (= standard ESC culture conditions) as
previously described.1 Standard ESC culture conditions were used for cultivation of mouse
MPI-II ESCs.
For differentiation of maGSCs, mouse maGSCs were cultivated as embryoid bodies (EBs) in
hanging drops in Iscove’s modified Dulbecco’s medium (IMDM, Invitrogen) supplemented
with 20% FCS, L-glutamine, NEAA and α-monothioglycerol 3-mercapto-1,2-propandiol
(MTG, final concentration 450 µmol/L, Sigma) as described for standard mouse ESC
differentiation.2 Briefly, cells (n = 400) in 20 µL differentiation medium were placed on the lids
of petri dishes filled with phosphate buffered saline (PBS) and cultivated in hanging drops for
2 days and in bacteriological petri dishes for 3 days. EBs were plated separately onto gelatin-
coated 24-microwell or 6-cm culture plates at day 5. For the determination of cardiac
differentiation efficiency, spontaneously beating cardiomyocytes were investigated from the
outgrowths of EBs cultured in 24-microwell plates at various stages during differentiation.
The percentage of EBs containing beating cardiomyocytes was used as a measure of the
efficiency of cardiomyocyte differentiation. At least three independent experiments (n = 48
EBs per experiment) were performed.

2
In vitro Response of Beating EBs to Pharmacological Agents
A day before the experiment, cells were fed with 1 mL/well differentiation medium and
beating areas were photographed and marked for later identification. The next day, the
beating frequency for each area was measured by visual inspection before adding the drugs.
To examine pharmacological responses, drugs (all from Sigma) were added to the culture at
the lowest dose. The beating frequency was monitored after cultures were incubated with
drugs at 37°C for 5 min. The procedure was repeated several times by sequentially adding
additional doses of the drug followed by monitoring the beating frequency. The results are
presented as the mean beating frequency ± standard error of the mean measured for at least
15 beating areas.
Isolation of Cardiomyocytes
Cardiomyocytes were isolated as single cells from the beating areas of EBs mechanically
using a micro-scalpel under an inverted microscope. Tissues were collected in Tyrode’s
solution with (in mmol/L) 137 NaCl, 5,4 KCl, 1,2 MgSO4, 1,2 Na2HPO4, 10 glucose, 20
HEPES (pH 7.4 with NaOH). The isolated clusters were digested in collagenase type 2 (310
U/mL, Worthington) supplemented Tyrode’s solution with 30 µmol/L CaCl2 at 37oC for 30
minutes. For the isolation of small cardiac clusters, the incubation time was shortened to 15
minutes. The cells were then resuspended and dissociated in 100 µL of KB medium2 at 37oC
for 30 minutes. The cell suspensions were transferred into tissue culture plates containing
gelatin-coated cover slips and incubated in differentiation medium at 37oC overnight.
Cardiomyocytes began rhythmical contractions after 24 hours and were used for
electrophyosiological assays and immunostaining 2-3 days later.
Immunocytochemistry
For immunostaining, isolated cardiomyocytes were first incubated with the antibodies against
sarcomeric α-actinin (rabbit polyclonal antibody, clone 653, kindly from Prof. D.O. Fürst,
University of Potsdam, Germany), cardiac troponin T (Ab-1, mouse monoclonal antibody,

3
clone 13-11, Lab Vision), L-type Ca2+ channel α (Cav1.2a, rabbit polyclonal antibody,
Alomone labs), connexin 43 (rabbit polyclonal antibody, Lab Vision), pan-cadherin (mouse
monoclonal antibody, clone CH-19, Sigma), sarcoplasmic/endoplasmic reticulum Ca2+-
ATPase (SERCA2a; mouse monoclonal antibody, clone 2A7-A1, Affinity Bioreagents),
sodium/calcium exchanger 1 (NCX1; mouse monoclonal antibody, clone 6H2, Chemicon),
and ryanodine receptor (RyR2; mouse monoclonal antibody, clone 34C, Affinity
Bioreagents). Afterwards, specimens were incubated with the fluorescent antibodies: FITC-
conjugated goat anti-rabbit (Dianova) for α-actinin; Cy3-conjugated goat anti-mouse IgG +
IgM (Dianova) for L-type Ca2+ channel α, SERCA2a, NCX1, and RyR2; FITC-conjugated
goat anti-mouse IgG (Dianova) for troponin T; Cy3-conjugated goat anti-rabbit (Dianova) for
connexin 43. Specimens were count stained with 4,6-diamino-2-phenylindole (DAPI; Sigma)
and analyzed using a fluorescence microscope.
Fluorescence Recovery after Photobleaching (FRAP) Assay
Functional analysis of gap junctions in isolated cardiac clusters was performed by measuring
the cell-to-cell diffusion of a fluorescent dye using a FRAP assay at room temperature. Briefly,
cardiac clusters were loaded with the membrane-permeant fluorescent dye calcein AM (5
µmol/L; Invitrogen) in Ca2+-free Tyrode solution for 20 min at 37oC. After washing away the
excess extracellular fluorescent dye to prevent further loading, the cultures were bathed in
Ca2+-free Tyrode solution and placed on the stage of a Zeiss LSM 5 PASCAL laser-scanning
confocal microscope. Using Zeiss software, a rectangular region encompassing 30-50% of a
single cell within a cell cluster was selected and its fluorescence was bleached by a high-
intensity laser pulse (488 nm, 3-5 s duration). This caused immediate loss of calcein
fluorescence emission recorded through a 505 nm long-pass emission filter. Calcein
redistribution from adjacent unbleached cells through connexin pores into the bleached
region of interest (ROI) was recorded in subsequent confocal images acquired at 30 s
intervals for up to 10 min using a low-intensity laser pulse. Fluorescence recovery within the
ROI was plotted as a function of time and fit to a single exponential function: IROI (t) = A [1 –

4
exp (–kt)], where IROI (t) is the ROI fluorescence intensity at time t, A is the amplitude of
fluorescence recovery, and k is the rate of recovery. The latter is considered a measure of
gap junction permeability.3 For the inhibitor studies, the cells were incubated in Ca2+-free
Tyrode solution containing the gap junction uncoupler 3β-hydroxy-11-oxoolean-12-en-30-oic
acid 3-hemisuccinate (carbenoxolone, 50 µmol/L; Sigma).
Action Potential Measurements
The membrane potential (Em) of single cardiomyocytes was measured at room temperature
with ruptured-patch whole cell current clamp as described previously.1 Fire-polished glass
microelectrodes of >10 MΩ resistance when filled with pipette solution were used. Pipette
solution contained (in mmol/L) 120 potassium aspartate, 8 KCl, 7 NaCl, 1 MgCl2, 10 HEPES,
5 Mg-ATP, 0.3 Li-GTP (pH 7.2 with KOH). A normal Tyrode’s solution with 2 mmol/L CaCl2
served as bath solution. Liquid junction potentials were corrected before GΩ seal
establishment. Access resistance was typically ~20 MΩ after patch rupture. Spontaneous
action potentials (APs) were recorded immediately after patch rupture and followed for about
2 min. Signals were filtered with 2.9 and 10 kHz Bessel filters, and recorded with an EPC10
amplifier (HEKA Elektronik, Lambrecht/Pfalz, Germany) using the Patchmaster software.
Action potentials were averaged (~10 APs per cell) and analysis was done using MS Excel®
to determine the maximum rate of rise of the AP upstroke (dV/dtmax), AP amplitude (APA), AP
duration at 90/80% of repolarisation (APD 90/80) and the maximum diastolic potential (MDP).
In some experiments isoproterenol (1 µmol/L) or cadmium chloride (0.5 mmol/L) were added
to the bath solution during recording. Signals were averaged and results visualized using
Graphpad PrismTM and SPSS SigmaPlot®.
Intracellular Calcium Measurements using Confocal Laser Microscopy
Intracellular calcium ([Ca2+]i) signals were recorded after incubating cells with 10 µmol/L fluo-
4 acetoxymethylester (Mobitec, Göttingen, Germany) for 15 min on a laser scanning confocal
microscope (Zeiss LSM 5 PASCAL, Göttingen, Germany). Cells were washed with Tyrode’s

5
solution. Fluo-4 was excited via an argon laser (488 nm; 30 mW) and emitted fluorescence
(F) was collected through a 505 nm long-pass emission filter. Changes in fluo-4 fluorescence
(indicating fluctuation in cytosolic Ca2+) were recorded in frame and line scan mode while the
cells were beating spontaneously. The images were acquired and analyzed using Zeiss
software and fluorescence signals were normalized to basal cell fluorescence after fluo-4
loading (F0). Intracellular Ca2+ was assessed using line scan modus and calibrated by the
following pseudo-ratio equation2 [Ca]i=Kd(F/F0)/(Kd/[Ca]i-rest+1–F/F0) with Kd=1100 nmol/L and
[Ca]i-rest=100 nmol/L.
RT-PCR Analysis
Differentiating EBs at various stages were collected for total RNA isolation (Promega). 150
ng of DNase-treated RNA was used for first-strand cDNA synthesis. One-tenth of the
resulting cDNA was taken as PCR template and amplified for 25-35 cycles. GAPDH was
used as an internal control. Sequences of oligonucleotide primers used for RT-PCR analyses
are available from the authors on request.
Transplantation of maGSCs
The present study used female C57BL/6 mice obtained from the animal facility of the
University of Goettingen. The investigation conforms to the Guide for the Care and Use of
Laboratory Animals published by the US National Institutes of Health (NIH Publication No.
85-23, revised 1996) and was approved by the local government authority.
Immunosuppression with cyclosporine A (Sandimmun, Novartis) was applied to all animals
used. Cyclosporine A was diluted 1:50 in 5% glucose and stored at -20°C. The dilution was
injected once daily intraperitoneally so that the resulting dose was 10 mg/kg body weight.
Intramyocardial Injection of maGSCs
Mice were anesthetized with isoflurane, intubated with a 22 gauge (G) plastic cannula and
ventilated with a mixture of 2% isoflurane in ambient air (150 breaths/min, tidal volume 150

6
µl) by use of a MiniVent (Type 845, Harvard Apparatus). The heart was exposed by a left
lateral thoracotomy via the 4th intercostal space and the pericardium was opened. Five
hundred thousand to one million cells (SSC5) labelled with a fluorescent carbocyanine dye
CM-DiI (Invitrogen) were injected in a volume of 50 µl by use of a 30G steel cannula
connected to a 50 µl Hamilton syringe via a PE10 tube. The total volume was applied in 5-7
injections of 7-10 µl each covering the area of the anterolateral wall of the heart that was
accessible via the thoracotomy. Afterwards, the thorax was closed with single sutures before
the skin was adapted with polypropylene 6-0 sutures. The mice were placed on a heating
pad to recover until fully awake.
Intravenous Application of maGSCs
Mice were anesthetized and ventilated as described above. The jugular vein was exposed
via a median cervical incision. Cannulation of the vein was performed with a 30G steel
cannula connected to an insulin syringe with a 29G needle via a PE10 tube. One million
maGSCs labelled with CM-DiI were injected in a volume of 100 µl. After the injection the skin
was closed and the mice were allowed to recover as described above.
Histochemical and Immunohistological Analysis of Hearts
Hearts were harvested, fixed in 4% buffered-formalin, embedded in paraffin and sectioned at
6 µm on a microtome. Haematoxylin-Eosin (H&E) staining and Masson-Goldner trichrome
staining of the sections was performed on an automated immunostainer (Ventana Medical
Systems, Tucson, AZ) according to the manufacturer's protocol. For immunohistology, non-
specific immunoreactivity was blocked by incubation of sections in 4% BSA. The antibody
panel used included monoclonal mouse anti-Ki-67 (Dako) and anti-Oct3/4 (Santa Cruz
Biotechnology) at dilutions of 1 : 50; monoclonal mouse anti-cardiac troponin T (cTnT;
LabVision, Fremont, CA) at a dilution of 1 : 1000; monoclonal mouse anti-smooth muscle-α-
actin (SM-α-actin; Sigma) at a dilution of 1:8000; polyclonal rabbit anti-von Willebrand factor
(vWF; Dako) at a dilution of 1:100. The antibodies Ki-67 and Oct3/4 were detected with

7
commercial available SuperPicTure-Plus Kit (Broad Spectrum-DAB; from Invitrogen).
Sections incubated with cTnT, vWF and SM-α-actin antibodies were reacted with a
fluorescein-conjugated secondary antibody.
Data Analysis and Statistics
All data was expressed as mean value ± standard error of the mean (SEM). Student’s
unpaired t-test was used to test for significant difference. Double sided P-values of P<0.05
were considered significant.
1. Guan K, Nayernia K, Maier LS, Wagner S, Dressel R, Lee JH, Nolte J, Wolf F, Li M,
Engel W, Hasenfuss G. Pluripotency of spermatogonial stem cells from adult mouse
testis. Nature. 2006;440:1199-1203.
2. Wobus AM, Guan K, Yang HT, Boheler KR. Embryonic stem cells as a model to study
cardiac, skeletal muscle, and vascular smooth muscle cell differentiation. Methods
Mol Biol. 2002;185:127-156.
3. Wade MH, Trosko JE, Schindler M. A fluorescence photobleaching assay of gap
junction-mediated communication between human cells. Science. 1986;232:525-528.

Supplementary Table 1 Characteristics of APs in maGSC-derived single cardiomyocytes
Pacemaker-like Ventricle-like Atrial-like Purkinje-like Intermediate
CM (pF) [n] 25.1±4.0 [8] 31.0±3.3 [23] 22.5±2.7 [9] 34.4±4.9 [9] 31.2±4.8 [19]
AP Amplitude
(mV) 62.8±6.9 93.1±3.0 88.3±5.6 96.4±5.3 73.6±3.2
APD 90
(ms) 227.7±12.4 314.8±21.3 162.8±19.1 287.6±23.9 338.8±15.8
APD 80
(ms) 203.3±12.3 281.7±20.8 120.2±14.9 185.8±17.3 310.4±15.1
Max. dV/dt (Vs-1) 2.5±0.6 31.6±3.8 43.5±5.4 49.4±6.2 2.9±0.6
MDP (mV) -38.1±3.4 -54.5±2.0 -56.9±2.0 -57.4±2.4 -42.6±2.3
Data are mean±SEM. n indicates the cell number; APA, AP amplitude; APD90/APD80, AP duration measeured at 90% or 80% repolarization; Max.
dV/dt, maximum rate of rise of AP; and MDP, maximum diastolic potential.

1
Supplementary Figure Legends
Supplementary Figure 1
Real time [Ca2+]i transients. Frames (recorded every ~149 ms) of one [Ca2+]i transient cycle
of the cardiomyocyte shown in Figure 5A are presented.
Supplementary Figure 2
Absence of cardiac differentiation of maGSCs after transplantion into the normal heart.
Immunostaining of the transplants using an antibody against cardiac Troponin T (cTnT)
showed that DiI-labeled transplants (A, D) were negative for cTnT at 1 week (B) and 1 month
(E). (C) Overlay of A and B. (F) overlay of D and E. Scale bar, 25 µm. Nuclear staining with
DAPI.
Supplementary Video Legends
Supplementary Video 1
Short video sequence of real time [Ca2+]i transients as detected by fluo-4 fluorescent dye and
recorded every 149 ms.

