membrane association of nitric oxide-sensitive guanylyl cyclase in cardiomyocytes
TRANSCRIPT

www.elsevier.com/locate/cardiores
Cardiovascular Research
Dow
nloa
Membrane association of nitric oxide-sensitive guanylyl
cyclase in cardiomyocytesB
Luis Agullo, David Garcia-Dorado*, Noelia Escalona, Marisol Ruiz-Meana,
Maribel Mirabet, Javier Inserte, Jordi Soler-Soler
Servicio de Cardiologıa, Hospital Universitari Vall d’Hebron, Pg Vall d’Hebron 129, 08035 Barcelona, Spain
Received 7 September 2004; received in revised form 11 April 2005; accepted 18 May 2005
Available online 13 June 2005
Time for primary review 19 days
by guest on June 1, 2013http://cardiovascres.oxfordjournals.org/
ded from
AbstractObjective: Although the importance of the cyclic GMP (cGMP) signaling pathway in cardiac myocytes is well established, little is known
about its regulation. Ca2+-dependent translocation of nitric oxide (NO) sensitive guanylyl cyclase (GCNO) to the cell membrane has been
recently proposed to play a role. The aim of this study was to determine the possible functional relevance of GCNO bound to the cardiomyocyte
membrane.
Methods: Cytosolic and particulate fractions of adult rat cardiomyocytes were isolated and blotted, and their GCNO activity was assayed in
parallel experiments.
Results: In untreated cardiomyocytes, approximately 30% of h1-and a1-subunits of GCNO and a similar proportion of GCNO activity were
found in the particulate fraction. The dependence of GCNO activity on pH, Ca2+, GTP and NO donor concentrations was similar in
particulate and cytosolic fractions. Treatment of cardiomyocytes with the ionophore A23187 caused GCNO to translocate to the
sarcolemma, increased GCNO activity in this fraction, and potentiated NO-mediated cGMP synthesis. These effects appeared to be
mediated by Ca2+-dependent changes on the phosphorylation status of GCNO, since they were enhanced by the non-selective inhibitor
staurosporine and by the selective inhibitor of Ca2+/calmodulin-dependent protein kinase KN-93. The effect of drugs increasing
intracellular Ca2+ on cGMP synthesis was clearly correlated with their effects on membrane-associated GCNO activity but not with their
effects on cytosol-associated GCNO.
Conclusion: These results are the first evidence that 1) GCNO is associated with the cell membrane in cardiomyocytes, 2) the regulation of
membrane-associated GCNO differs from that of cytosolic GCNO, and 3) membrane association may have a crucial role in determining the
response of cells to NO.
D 2005 European Society of Cardiology. Published by Elsevier B.V. All rights reserved.
Keywords: Nitric oxide; Calcium (cellular); Signal transduction; Myocytes; Protein phosphorylation
1. Introduction
Cyclic GMP (cGMP) modulates important physiological
functions in the cardiovascular system as vasodilation, Ca2+
cycling, endothelium permeability, or myocardial contrac-
0008-6363/$ - see front matter D 2005 European Society of Cardiology. Publishe
doi:10.1016/j.cardiores.2005.05.021
i Ajay Shah (King’s College, London) served as Guest Editor for this
manuscript
* Corresponding author. Tel.: +34 93 489 40 38; fax: +34 93 489 40 32.
E-mail address: [email protected] (D. Garcia-Dorado).
tility [1]. Abundant evidence indicates that cGMP can
modulate cell death during ischemia-reperfusion [2–7] and
cGMP has been described to mediate late preconditioning in
conscious rabbits [8].
cGMP can be synthesized by two different types of
guanylyl cyclases: a nitric oxide (NO)-sensitive guanylyl
cyclase (GCNO), generally known as cytosolic or soluble
guanylyl cyclase, and guanylyl cyclases that are integral
proteins in the plasmatic membrane of the cell and can be
stimulated by natriuretic peptides. GCNO is constituted of
68 (2005) 65 – 74
d by Elsevier B.V. All rights reserved.

L. Agullo et al. / Cardiovascular Research 68 (2005) 65–7466
by guest ohttp://cardiovascres.oxfordjournals.org/
Dow
nloaded from
two subunits, a and h, and two different isoforms of the a
subunit (a1 and a2) and of the h subunit (h1 and h2) have
been described. The a1h1 heterodimer is predominantly
found in the cardiovascular system, while a2h1 has been
mainly found in brain.
Little is known about how GCNO is modulated in vivo.
Rapid desensitisation of the signal [9] and phosphorylation
by different protein kinases, as protein kinase C (PKC) [11],
cyclic AMP-dependent protein kinase (PKA) [10,11] and
cGMP-dependent protein kinase (PKG) [12], have been
described.
Recent studies have challenged the classical concept of
an exclusive cytosolic location of GCNO. The isoform a2h1
has been found tightly associated to the neuron membrane
through a PSD-95 mediated interaction [13]. A histochem-
ical study suggested the presence of the more amply
distributed heterodimer, a1h1, in the sarcolemmal region
of skeletal muscle fibres [14]. This association to the
particulate fraction of a1 and h1 subunits has been recently
demonstrated in myocardial tissue, endothelial cells and
platelets [15], and in these later cells activation with ADP or
collagen has been correlated with enzyme translocation to
the membrane [15]. However, the mechanism of the
interaction of a1h1 with the membrane was unresolved. A
recent study has described a protein complex formed by
eNOS, Hsp90 and GCNO in endothelial cells, and that
bradykinin and vascular endothelial growth factor potentiate
the formation of this complex [16]. The contribution of
particulate GCNO to the cell response to NO remained
unknown.
In this study, we analyze how association of GCNO to
membrane affects its biochemical properties in cardiomyo-
cytes, the functional relevance of this association, and its
potential regulation by Ca2+.
n June 1, 2013
2. Materials and methods
The animal protocols conformed to the Guide for the
Care and Use of Laboratory Animals published by the US
National Institutes of Health (NIH publication No. 85-23,
revised 1996) and was approved by the Research Commis-
sion on Ethics of Hospital Vall d’Hebron.
2.1. Cardiomyocyte and platelet isolation
Cardiomyocytes were obtained from adult rat hearts as
previously described [17]. At the end of the procedure,
culture dishes contained >85% of quiescent rod-shaped
cells. Rat washed platelets were isolated from venous blood
collected with sodium citrate [18].
2.2. Intracellular cGMP synthesis
After treatment, cells were stimulated for 1 minute with
SNAP 100 AM (unless otherwise indicated) in the presence
of 1 mM 3-isobutyl-1-methylxanthine (IBMX) as inhibitor
of cGMP degradation. cGMP was quantified by radio-
immunoassay using acetylated [3H]cGMP [17].
2.3. Membrane and cytosolic fractions
Following incubation with the different agents, cardio-
myocytes were homogenized with a straight wall grinder
(Radnoti Glass Technology) in ice-cold buffer A [in mM:
Tris.HCl 50 (pH 7.4), sucrose 250, EDTA 0.1, dithiotreitol
1, plus protease inhibitors [17], the protein kinase inhibitor
staurosporin 1�10�3, and the Ser/Thr protein phosphatase
inhibitors okadaic acid 1�10�3 and cypermethrin
5�10�4]. After clearing the homogenates by centrifugation
at 1000 �g for 15 min, the particulate and cytosolic
fractions were obtained by centrifugation (100,000 �g for 1
h). Membrane fractions were homogenized with buffer A
plus 10% glycerol. Triton X-100 solubility of particulate
GCNO was assessed as described [19]. Treated platelets were
centrifuged, resuspended in buffer A, frozen in liquid
nitrogen, thawed slowly on ice and then homogenized with
a straight wall grinder and ultracentrifuged as mentioned for
cardiomyocytes.
2.4. Guanylyl cyclase activity
Guanylyl cyclase activity was determined [17] by
measuring cGMP (radioimmunoassay) synthesized after
incubating the particulate and cytosolic extracts with no-
additions (basal) or 100 AM SNAP in assay buffer [final
concentrations (in mM): Tris.HCl 50 (pH 7.4), EGTA 1,
dithiotreitol 1, GTP 2.5, MgCl2 5, phosphocreatine 10,
IBMX 1, plus creatine kinase (50 U/ml)] at 37 -C for 30
min. GCNO activity, calculated by substracting basal
activity, was linear with respect to time in the assay period.
To analyze pH dependence of GCNO activity, Tris.HCl was
substituted by 30 mM PIPES (pH 6.0–6.8) or 30 mM
HEPES (pH 6.8–8.0). Free Ca2+ concentrations were
calculated by a modification of the RECIPC program (S.
Roberston, University of Cincinnati, 1981).
2.5. Western-blotting
Proteins were separated by electrophoresis on a 10% SDS
gel and transferred onto nitrocellulose membrane (Hybond-
ECL, Amersham). Membranes were incubated with rabbit
polyclonal antibodies to h1 (aminoacids 605–619; used at 1 /
2000 dilution; Sigma) and a1 (aminoacids 673–690; 1 /
20,000; Sigma) subunits of soluble guanylyl cyclase. A goat
anti-rabbit IgG horseradish peroxidase conjugated (1 /
50,000; Pierce) was used as secondary antibody. Specificity
of the immunostaining was assessed by displacing the
corresponding bands by incubating in the presence of their
respective immunization peptides (synthesized by Sigma
Genosys). Quantitative chemiluminiscence detection was
performed with SuperSignal West Dura Extended Substrate
L. Agullo et al. / Cardiovascular Research 68 (2005) 65–74 67
Dow
nloaded
(Pierce) and a 16-bit cooled CCD camara system (LAS-
3000, Fujifilm). Equal loading of the different samples was
confirmed by Ponceau Red staining.
2.6. Modulation of the membrane association of GCNO
The effect of increased cytosolic Ca2+ concentration on
the association of GCNO to the membrane fraction was
investigated by incubating cells 1 min with A23187 or 5
min with thapsigargin. Then, cells were: a) stimulated with
SNAP for determining NO-dependent cGMP synthesis, and
b) homogenized for determining content of h1 subunit and
GCNO activity in cytosolic and membrane fractions.
The potential role of changes in GCNO phosphorylation in
this effect was investigated by analyzing its modulation by
the protein kinase inhibitors staurosporin, Go-6976, H-89,
and KN-93. These drugs were added to the incubation media
4 min before stimulation with A23187, and maintained
during the time of incubation with the ionophore.
A
β1
Cyt. Memb. Cyt.
EXP 1 EX
β1
α1
Cytosol Membrane
IB: anti-β1
IB: anti-α1
3 µg
0.6
µg
15 µ
g
100µ
g
3 µg
0.6
µg
15 µ
g
Plq PlqCM C
Plq PlqCM C
B
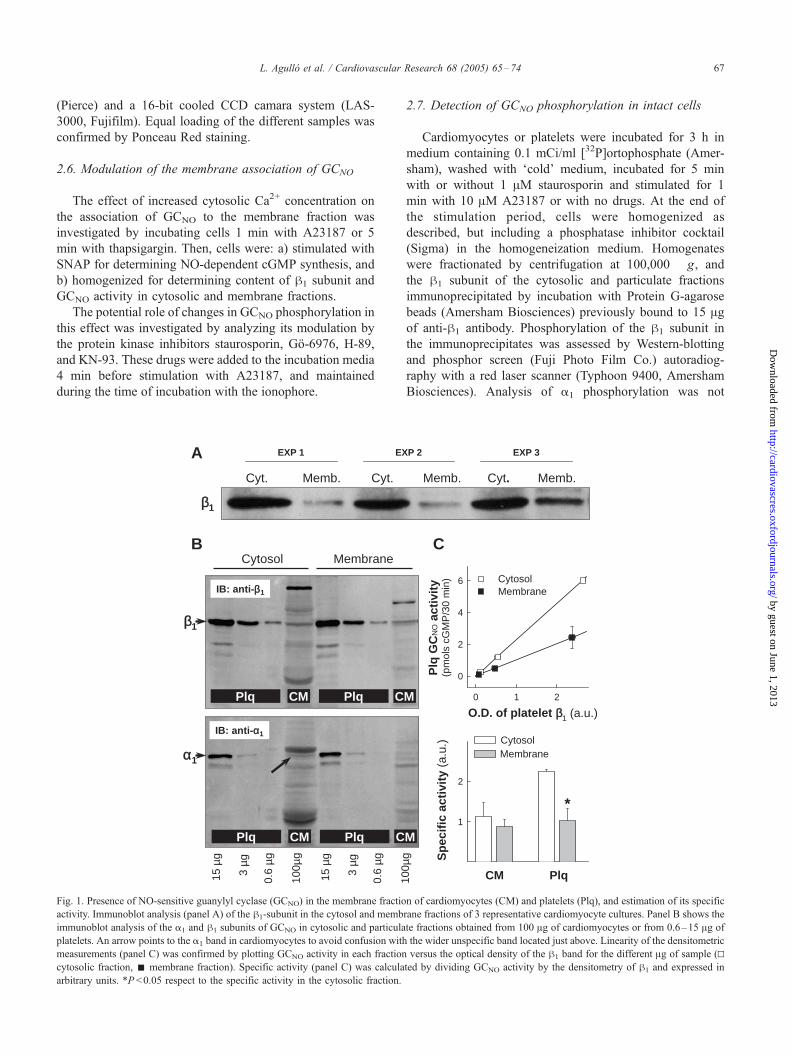
Fig. 1. Presence of NO-sensitive guanylyl cyclase (GCNO) in the membrane fractio
activity. Immunoblot analysis (panel A) of the h1-subunit in the cytosol and memb
immunoblot analysis of the a1 and h1 subunits of GCNO in cytosolic and particula
platelets. An arrow points to the a1 band in cardiomyocytes to avoid confusion with
measurements (panel C) was confirmed by plotting GCNO activity in each fraction
cytosolic fraction, ˝ membrane fraction). Specific activity (panel C) was calcula
arbitrary units. *P <0.05 respect to the specific activity in the cytosolic fraction.
2.7. Detection of GCNO phosphorylation in intact cells
Cardiomyocytes or platelets were incubated for 3 h in
medium containing 0.1 mCi/ml [32P]ortophosphate (Amer-
sham), washed with Fcold_ medium, incubated for 5 min
with or without 1 AM staurosporin and stimulated for 1
min with 10 AM A23187 or with no drugs. At the end of
the stimulation period, cells were homogenized as
described, but including a phosphatase inhibitor cocktail
(Sigma) in the homogeneization medium. Homogenates
were fractionated by centrifugation at 100,000 �g, and
the h1 subunit of the cytosolic and particulate fractions
immunoprecipitated by incubation with Protein G-agarose
beads (Amersham Biosciences) previously bound to 15 Agof anti-h1 antibody. Phosphorylation of the h1 subunit in
the immunoprecipitates was assessed by Western-blotting
and phosphor screen (Fuji Photo Film Co.) autoradiog-
raphy with a red laser scanner (Typhoon 9400, Amersham
Biosciences). Analysis of a1 phosphorylation was not
Memb. Cyt. Memb.
P 2 EXP 3
100µ
g
O.D. of platelet β1 (a.u.)0 1 2
Plq
GC
NO a
ctiv
ity
(pm
ols
cGM
P/3
0 m
in)
0
2
4
6
Sp
ecif
ic a
ctiv
ity
(a.u
.)
1
2
*
CM Plq
CytosolMembrane
CytosolMembrane
M
M
C
n of cardiomyocytes (CM) and platelets (Plq), and estimation of its specific
rane fractions of 3 representative cardiomyocyte cultures. Panel B shows the
te fractions obtained from 100 Ag of cardiomyocytes or from 0.6–15 Ag of
the wider unspecific band located just above. Linearity of the densitometric
versus the optical density of the h1 band for the different Ag of sample (gted by dividing GCNO activity by the densitometry of h1 and expressed in
by guest on June 1, 2013http://cardiovascres.oxfordjournals.org/
from

L. Agullo et al. / Cardiovascular Research 68 (2005) 65–7468
Dow
nloa
possible since the antibody used against this subunit did
not significantly immunoprecipitate the protein.
2.8. Intracellular Ca2+
Changes induced by the Ca2+ ionophore A23187 and by
thapsigargin were monitored by ratio-fluorescence imaging
in cardiomyocytes loaded with fura 2-acetoxymethyl ester
(Molecular Probes) as previously described [20].
2.9. Data analysis and statistics
Differences between groups were evaluated by means of
paired Student’s t test when appropriated or one-way
analysis of the variance. Individual comparisons between
groups were performed using the Student–Newman Keuls
test. Values are expressed as meanTSEM. Nonlinear fitting
was performed using SigmaPlot (SPSS Inc.).
http://cardiovascres.oxfordjournalsded from
3. Results3.1. Membrane association of GCNO in cardiomyocytes and
platelets
Cytosolic and particulate fractions (100,000 �g) of
adult rat cardiomyocytes were isolated and blotted.
Staining with an antibody against the h1 subunit of the
GCNO, demonstrated that 28T5% of h1 (65.1T0.2 kDa),
n =6, was in the membrane fraction (Fig. 1A). A similar
β1
Cyt. Memb. Cyt. Memb.
+ peptide β1
S
β1
A IB: anti-β1
α1+ peptide α1
B IB: anti-α1
C IB: anti-β1
Plq
Fig. 2. h1 (panel A) and a1 (panel B) immunoreactivity of 65.1 kDa (for the himmunization peptides in cytosolic and particulate fractions of platelets and cardio
kDa in the particulate fraction) were also displaced by peptides. Membrane associa
in a KCl containing buffer, but was completely disrupted by the presence of dete
proportion of the a1 subunit (75.7T2.9 kDa) seemed to be
associated to the particulate fraction, but the limited
sensitivity of the antibody against this subunit in cardio-
myocytes precluded precise analysis (Fig. 1B). Immuniza-
tion peptides displaced h1 and a1 bands both in platelets
and cardiomyocytes (Fig. 2A and B). However, as shown
in Fig. 2, an additional band in the cytosolic fraction (of
110–115 kDa) and in the membrane fraction (of 85–90
kDa) of cardiomyocytes were also displaced. Since the
identity of these other bands is unknown, they were not
used for the quantification of the h1 content. Throughout
the rest of the study only the antibody against the h1-
subunit was used for blot analysis.
Measurement of GCNO activity showed that 33T4% of
the activity was associated to the particulate fraction
(5.1T2.2 and 2.5T1.1 pmol/mg protein x min in the
cytosolic and membrane fractions, respectively, p =6). For
comparison with a cell model in which the association of
GCNO to the membrane had been previously described,
[15,21,22] cytosolic and membrane fractions were
obtained from rat platelets (Fig. 1B). According to
densitometric analysis of blots, platelets contained 100–
150 times more GCNO than cardiomyocytes. Specific
activity of membrane-associated GCNO, calculated by
dividing GCNO activity by the densitometry of h1 in this
fraction (in arbitrary units), was similar in platelets and
cardiomyocytes (Fig. 1C).
To rule-out an unspecific presence of GCNO in the
membrane fraction, cardiomyocyte homogenates were
extensively diluted (15 times) in a homogeneization
Cyt. Memb. Cyt. Memb.
+ peptide β1
Membrane
+KCltandard +TritonX-100
+ peptide α1
CM
1 band) and 75.7 kDa (for the a1 band) were blocked by their respective
myocytes. Other bands (110–115 kDa in the cytosolic fraction and 85–90
tion of GCNO in cardiomyocytes was not altered by diluting the homogenate
rgent (panel C).
by guest on June 1, 2013.org/
Log [GTP], mMC -4 -3
GC
NO a
ctiv
ity
(% G
TP
3 m
M)
0
50
100 A
CytosolMembrane
Log [SNAP], mMC -5 -4 -3
GC
NO a
ctiv
ity
(% S
NA
P 1
mM
)
0
50
100 B
P<0.01
P<0.05
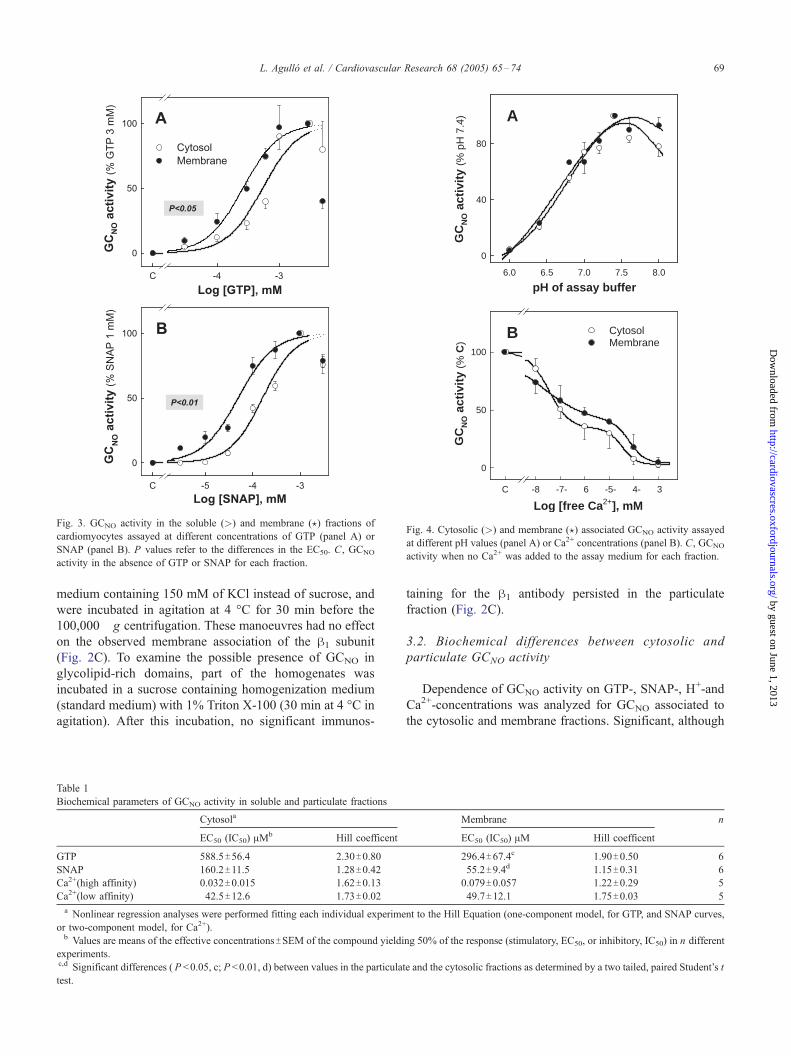
Fig. 3. GCNO activity in the soluble (>) and membrane (?) fractions of
cardiomyocytes assayed at different concentrations of GTP (panel A) or
SNAP (panel B). P values refer to the differences in the EC50. C, GCNO
activity in the absence of GTP or SNAP for each fraction.
Log [free Ca2+], mMC -8 -7- 6 -5- 4- 3
GC
NO a
ctiv
ity
(% C
)0
50
100
B CytosolMembrane
pH of assay buffer6.0 6.5 7.0 7.5 8.0
GC
NO a
ctiv
ity
(% p
H 7
.4)
0
40
80
A
Fig. 4. Cytosolic (>) and membrane (?) associated GCNO activity assayed
at different pH values (panel A) or Ca2+ concentrations (panel B). C, GCNO
activity when no Ca2+ was added to the assay medium for each fraction.
L. Agullo et al. / Cardiovascular Research 68 (2005) 65–74 69
by guest on June 1, 2013http://cardiovascres.oxfordjournals.org/
Dow
nloaded from
medium containing 150 mM of KCl instead of sucrose, and
were incubated in agitation at 4 -C for 30 min before the
100,000 �g centrifugation. These manoeuvres had no effect
on the observed membrane association of the h1 subunit
(Fig. 2C). To examine the possible presence of GCNO in
glycolipid-rich domains, part of the homogenates was
incubated in a sucrose containing homogenization medium
(standard medium) with 1% Triton X-100 (30 min at 4 -C in
agitation). After this incubation, no significant immunos-
Table 1
Biochemical parameters of GCNO activity in soluble and particulate fractions
Cytosola
EC50 (IC50) AMb Hill coefficent
GTP 588.5T56.4 2.30T0.80
SNAP 160.2T11.5 1.28T0.42Ca2+(high affinity) 0.032T0.015 1.62T0.13
Ca2+(low affinity) 42.5T12.6 1.73T0.02a Nonlinear regression analyses were performed fitting each individual experime
or two-component model, for Ca2+).b Values are means of the effective concentrationsTSEM of the compound yieldi
experiments.c,d Significant differences ( P <0.05, c; P <0.01, d) between values in the particulat
test.
taining for the h1 antibody persisted in the particulate
fraction (Fig. 2C).
3.2. Biochemical differences between cytosolic and
particulate GCNO activity
Dependence of GCNO activity on GTP-, SNAP-, H+-and
Ca2+-concentrations was analyzed for GCNO associated to
the cytosolic and membrane fractions. Significant, although
Membrane n
EC50 (IC50) AM Hill coefficent
296.4T67.4c 1.90T0.50 6
55.2T9.4d 1.15T0.31 6
0.079T0.057 1.22T0.29 5
49.7T12.1 1.75T0.03 5
nt to the Hill Equation (one-component model, for GTP, and SNAP curves,
ng 50% of the response (stimulatory, EC50, or inhibitory, IC50) in n different
e and the cytosolic fractions as determined by a two tailed, paired Student’s t
Time (min)0 4
Fu
ra-2
rat
io (
340/
380)
1.0
1.2
1.4
1.6
Time (min)0 5 10 15
Fu
ra-2
rat
io (
340/
380)
1.0
1.2
1.4
1.6
A
B
2
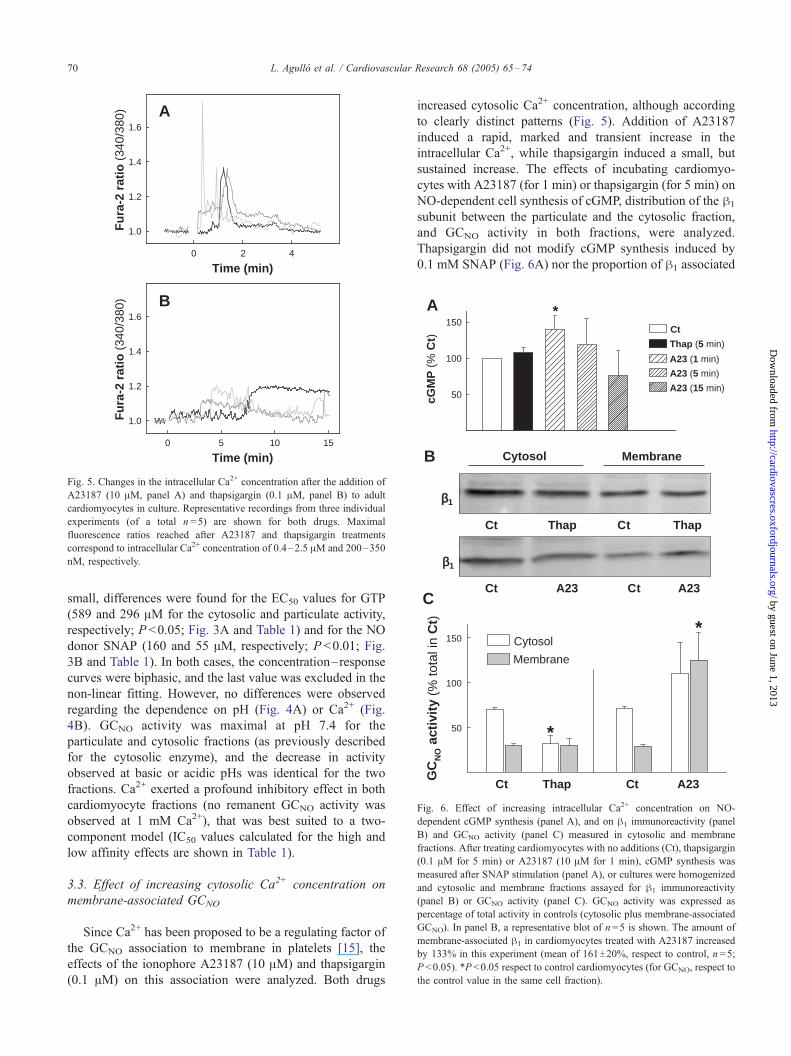
Fig. 5. Changes in the intracellular Ca2+ concentration after the addition of
A23187 (10 AM, panel A) and thapsigargin (0.1 AM, panel B) to adult
cardiomyocytes in culture. Representative recordings from three individual
experiments (of a total n =5) are shown for both drugs. Maximal
fluorescence ratios reached after A23187 and thapsigargin treatments
correspond to intracellular Ca2+ concentration of 0.4–2.5 AM and 200–350
nM, respectively.
B
A
C
cGM
P (
% C
t)
50
100
150*
A23 (15 min)
A23 (5 min)
A23 (1 min)
Thap (5 min)
Ct
Cytosol Membrane
Ct A23 Ct A23
β1
Ct Thap Ct Thap
β1
GC
NO a
ctiv
ity
(% to
tal i
n C
t)
50
100
150
*
*
ThapCt A23Ct
Membrane
Cytosol
Fig. 6. Effect of increasing intracellular Ca2+ concentration on NO-
dependent cGMP synthesis (panel A), and on h1 immunoreactivity (panel
B) and GCNO activity (panel C) measured in cytosolic and membrane
fractions. After treating cardiomyocytes with no additions (Ct), thapsigargin
(0.1 AM for 5 min) or A23187 (10 AM for 1 min), cGMP synthesis was
measured after SNAP stimulation (panel A), or cultures were homogenized
and cytosolic and membrane fractions assayed for h1 immunoreactivity
(panel B) or GCNO activity (panel C). GCNO activity was expressed as
percentage of total activity in controls (cytosolic plus membrane-associated
GCNO). In panel B, a representative blot of n =5 is shown. The amount of
membrane-associated h1 in cardiomyocytes treated with A23187 increased
by 133% in this experiment (mean of 161T20%, respect to control, n =5;
P <0.05). *P <0.05 respect to control cardiomyocytes (for GCNO, respect to
the control value in the same cell fraction).
L. Agullo et al. / Cardiovascular Research 68 (2005) 65–7470
by guest on June 1, 2013http://cardiovascres.oxfordjournals.org/
Dow
nloaded from
small, differences were found for the EC50 values for GTP
(589 and 296 AM for the cytosolic and particulate activity,
respectively; P <0.05; Fig. 3A and Table 1) and for the NO
donor SNAP (160 and 55 AM, respectively; P <0.01; Fig.
3B and Table 1). In both cases, the concentration–response
curves were biphasic, and the last value was excluded in the
non-linear fitting. However, no differences were observed
regarding the dependence on pH (Fig. 4A) or Ca2+ (Fig.
4B). GCNO activity was maximal at pH 7.4 for the
particulate and cytosolic fractions (as previously described
for the cytosolic enzyme), and the decrease in activity
observed at basic or acidic pHs was identical for the two
fractions. Ca2+ exerted a profound inhibitory effect in both
cardiomyocyte fractions (no remanent GCNO activity was
observed at 1 mM Ca2+), that was best suited to a two-
component model (IC50 values calculated for the high and
low affinity effects are shown in Table 1).
3.3. Effect of increasing cytosolic Ca2+ concentration on
membrane-associated GCNO
Since Ca2+ has been proposed to be a regulating factor of
the GCNO association to membrane in platelets [15], the
effects of the ionophore A23187 (10 AM) and thapsigargin
(0.1 AM) on this association were analyzed. Both drugs
increased cytosolic Ca2+ concentration, although according
to clearly distinct patterns (Fig. 5). Addition of A23187
induced a rapid, marked and transient increase in the
intracellular Ca2+, while thapsigargin induced a small, but
sustained increase. The effects of incubating cardiomyo-
cytes with A23187 (for 1 min) or thapsigargin (for 5 min) on
NO-dependent cell synthesis of cGMP, distribution of the h1
subunit between the particulate and the cytosolic fraction,
and GCNO activity in both fractions, were analyzed.
Thapsigargin did not modify cGMP synthesis induced by
0.1 mM SNAP (Fig. 6A) nor the proportion of h1 associated
A
C
B
A23Ct A23+Stau
A23Ct A23+Stau
Cytosol Membrane
β1
cGM
P (
% C
t)
50
100
150
200*
*#
Stau + A23
A23 (1 min)
Stau (5 min)
Ct
GC
NO a
ctiv
ity
(% to
tal i
n C
t)
50
100
150
200
#
*
Membrane
Cytosol*
A23Ct A23+StauStau
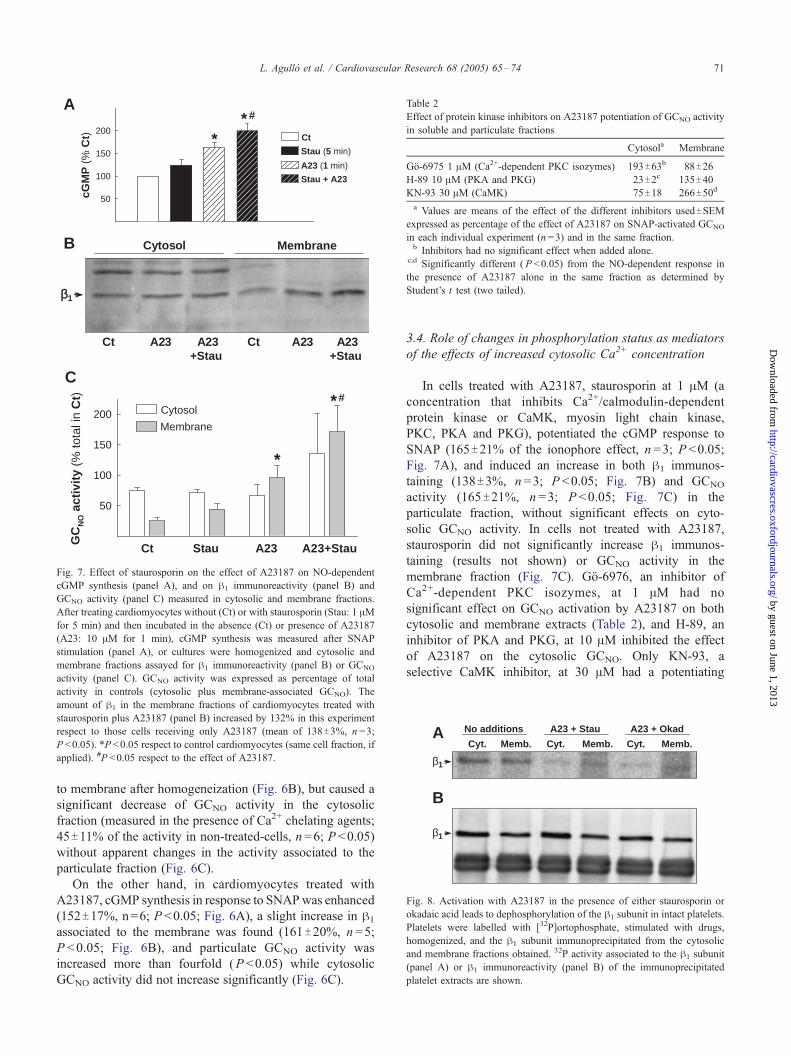
Fig. 7. Effect of staurosporin on the effect of A23187 on NO-dependent
cGMP synthesis (panel A), and on h1 immunoreactivity (panel B) and
GCNO activity (panel C) measured in cytosolic and membrane fractions.
After treating cardiomyocytes without (Ct) or with staurosporin (Stau: 1 AMfor 5 min) and then incubated in the absence (Ct) or presence of A23187
(A23: 10 AM for 1 min), cGMP synthesis was measured after SNAP
stimulation (panel A), or cultures were homogenized and cytosolic and
membrane fractions assayed for h1 immunoreactivity (panel B) or GCNO
activity (panel C). GCNO activity was expressed as percentage of total
activity in controls (cytosolic plus membrane-associated GCNO). The
amount of h1 in the membrane fractions of cardiomyocytes treated with
staurosporin plus A23187 (panel B) increased by 132% in this experiment
respect to those cells receiving only A23187 (mean of 138T3%, n =3;
P <0.05). *P <0.05 respect to control cardiomyocytes (same cell fraction, if
applied). #P <0.05 respect to the effect of A23187.
Table 2
Effect of protein kinase inhibitors on A23187 potentiation of GCNO activity
in soluble and particulate fractions
Cytosola Membrane
Go-6975 1 AM (Ca2+-dependent PKC isozymes) 193T63b 88T26
H-89 10 AM (PKA and PKG) 23T2c 135T40KN-93 30 AM (CaMK) 75T18 266T50d
a Values are means of the effect of the different inhibitors usedTSEMexpressed as percentage of the effect of A23187 on SNAP-activated GCNO
in each individual experiment (n =3) and in the same fraction.b Inhibitors had no significant effect when added alone.
c,d Significantly different ( P <0.05) from the NO-dependent response in
the presence of A23187 alone in the same fraction as determined by
Student’s t test (two tailed).
β1
β1
A
B
Cyt. Memb. Cyt. Memb. Cyt. Memb.
No additions A23 + Stau A23 + Okad
Fig. 8. Activation with A23187 in the presence of either staurosporin or
okadaic acid leads to dephosphorylation of the h1 subunit in intact platelets.
Platelets were labelled with [32P]ortophosphate, stimulated with drugs,
homogenized, and the h1 subunit immunoprecipitated from the cytosolic
and membrane fractions obtained. 32P activity associated to the h1 subunit
(panel A) or h1 immunoreactivity (panel B) of the immunoprecipitated
platelet extracts are shown.
L. Agullo et al. / Cardiovascular Research 68 (2005) 65–74 71
by guest on June 1, 2013http://cardiovascres.oxfordjournals.org/
Dow
nloaded from
to membrane after homogeneization (Fig. 6B), but caused a
significant decrease of GCNO activity in the cytosolic
fraction (measured in the presence of Ca2+ chelating agents;
45T11% of the activity in non-treated-cells, n =6; P <0.05)
without apparent changes in the activity associated to the
particulate fraction (Fig. 6C).
On the other hand, in cardiomyocytes treated with
A23187, cGMP synthesis in response to SNAPwas enhanced
(152T17%, n=6; P <0.05; Fig. 6A), a slight increase in h1
associated to the membrane was found (161T20%, n =5;
P <0.05; Fig. 6B), and particulate GCNO activity was
increased more than fourfold (P <0.05) while cytosolic
GCNO activity did not increase significantly (Fig. 6C).
3.4. Role of changes in phosphorylation status as mediators
of the effects of increased cytosolic Ca2+ concentration
In cells treated with A23187, staurosporin at 1 AM (a
concentration that inhibits Ca2+/calmodulin-dependent
protein kinase or CaMK, myosin light chain kinase,
PKC, PKA and PKG), potentiated the cGMP response to
SNAP (165T21% of the ionophore effect, n =3; P <0.05;
Fig. 7A), and induced an increase in both h1 immunos-
taining (138T3%, n =3; P <0.05; Fig. 7B) and GCNO
activity (165T21%, n =3; P <0.05; Fig. 7C) in the
particulate fraction, without significant effects on cyto-
solic GCNO activity. In cells not treated with A23187,
staurosporin did not significantly increase h1 immunos-
taining (results not shown) or GCNO activity in the
membrane fraction (Fig. 7C). Go-6976, an inhibitor of
Ca2+-dependent PKC isozymes, at 1 AM had no
significant effect on GCNO activation by A23187 on both
cytosolic and membrane extracts (Table 2), and H-89, an
inhibitor of PKA and PKG, at 10 AM inhibited the effect
of A23187 on the cytosolic GCNO. Only KN-93, a
selective CaMK inhibitor, at 30 AM had a potentiating
L. Agullo et al. / Cardiovascular Research 68 (2005) 65–7472
http://cardiovD
ownloaded from
effect on particulate GCNO activity similar to that of
staurosporin (Table 2). On the other hand, the phospha-
tase inhibitors cypermethrin at 0.05 AM (selective
inhibitor of calcineurin) or okadaic acid at 1 AM(inhibitor of PP1 and PP2A) did not block the potentiat-
ing effect of A23187 on membrane GCNO activity; in
fact, okadaic acid enhanced it (290 T15% of the
ionophore effect, n =3; P <0.05).
We were not able to detect in vivo 32P-labelling of GCNO
in cardiomyocytes neither under control nor after stauro-
sporin plus A23187 treatment. In platelets, incubation with
the ionophore A23187 in the presence of staurosporin
decreased phosphorylation of the h1 subunit compared to
non-treated cells (to about 25% of the initial value; Fig. 8).
Stimulation with A23187 in platelets preincubated with
okadaic acid had a similar effect (Fig. 8).
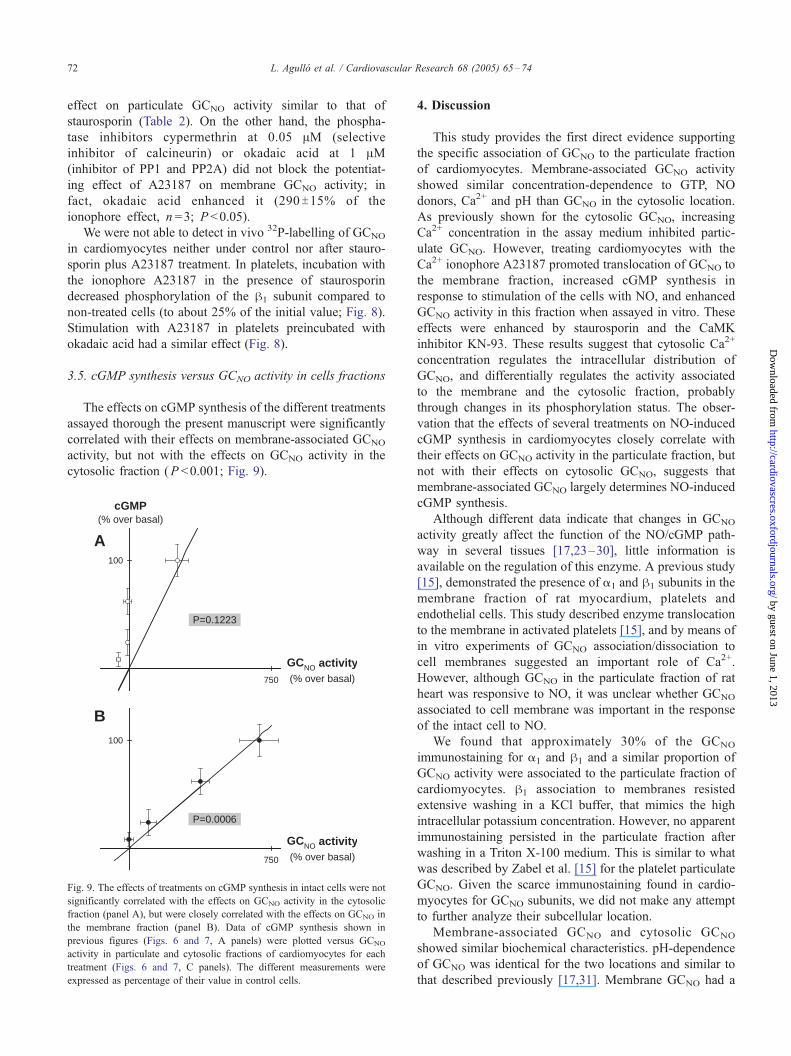
3.5. cGMP synthesis versus GCNO activity in cells fractions
The effects on cGMP synthesis of the different treatments
assayed thorough the present manuscript were significantly
correlated with their effects on membrane-associated GCNO
activity, but not with the effects on GCNO activity in the
cytosolic fraction (P <0.001; Fig. 9).
GCNO activity(% over basal)750
cGMP(% over basal)
100
A
B
750
100
GCNO activity(% over basal)
P=0.1223
P=0.0006
Fig. 9. The effects of treatments on cGMP synthesis in intact cells were not
significantly correlated with the effects on GCNO activity in the cytosolic
fraction (panel A), but were closely correlated with the effects on GCNO in
the membrane fraction (panel B). Data of cGMP synthesis shown in
previous figures (Figs. 6 and 7, A panels) were plotted versus GCNO
activity in particulate and cytosolic fractions of cardiomyocytes for each
treatment (Figs. 6 and 7, C panels). The different measurements were
expressed as percentage of their value in control cells.
by guest on June 1, 2013ascres.oxfordjournals.org/
4. Discussion
This study provides the first direct evidence supporting
the specific association of GCNO to the particulate fraction
of cardiomyocytes. Membrane-associated GCNO activity
showed similar concentration-dependence to GTP, NO
donors, Ca2+ and pH than GCNO in the cytosolic location.
As previously shown for the cytosolic GCNO, increasing
Ca2+ concentration in the assay medium inhibited partic-
ulate GCNO. However, treating cardiomyocytes with the
Ca2+ ionophore A23187 promoted translocation of GCNO to
the membrane fraction, increased cGMP synthesis in
response to stimulation of the cells with NO, and enhanced
GCNO activity in this fraction when assayed in vitro. These
effects were enhanced by staurosporin and the CaMK
inhibitor KN-93. These results suggest that cytosolic Ca2+
concentration regulates the intracellular distribution of
GCNO, and differentially regulates the activity associated
to the membrane and the cytosolic fraction, probably
through changes in its phosphorylation status. The obser-
vation that the effects of several treatments on NO-induced
cGMP synthesis in cardiomyocytes closely correlate with
their effects on GCNO activity in the particulate fraction, but
not with their effects on cytosolic GCNO, suggests that
membrane-associated GCNO largely determines NO-induced
cGMP synthesis.
Although different data indicate that changes in GCNO
activity greatly affect the function of the NO/cGMP path-
way in several tissues [17,23–30], little information is
available on the regulation of this enzyme. A previous study
[15], demonstrated the presence of a1 and h1 subunits in the
membrane fraction of rat myocardium, platelets and
endothelial cells. This study described enzyme translocation
to the membrane in activated platelets [15], and by means of
in vitro experiments of GCNO association/dissociation to
cell membranes suggested an important role of Ca2+.
However, although GCNO in the particulate fraction of rat
heart was responsive to NO, it was unclear whether GCNO
associated to cell membrane was important in the response
of the intact cell to NO.
We found that approximately 30% of the GCNO
immunostaining for a1 and h1 and a similar proportion of
GCNO activity were associated to the particulate fraction of
cardiomyocytes. h1 association to membranes resisted
extensive washing in a KCl buffer, that mimics the high
intracellular potassium concentration. However, no apparent
immunostaining persisted in the particulate fraction after
washing in a Triton X-100 medium. This is similar to what
was described by Zabel et al. [15] for the platelet particulate
GCNO. Given the scarce immunostaining found in cardio-
myocytes for GCNO subunits, we did not make any attempt
to further analyze their subcellular location.
Membrane-associated GCNO and cytosolic GCNO
showed similar biochemical characteristics. pH-dependence
of GCNO was identical for the two locations and similar to
that described previously [17,31]. Membrane GCNO had a

L. Agullo et al. / Cardiovascular Research 68 (2005) 65–74 73
by guest on June 1, 2013http://cardiovascres.oxfordjournals.org/
Dow
nloaded from
significantly lower EC50 value for GTP than the cytosolic,
but the difference was small and its physiological relevance
is doubtful. The difference in sensitivity to the NO donor
SNAP was more clear and similar to that described
previously in heart extracts [15]. However, a very recent
study [21] has suggested that contaminating myoglobin in
cytosolic extracts from heart tissue neutralizes a significant
part of the NO released by NO donors. According to this,
our results may underestimate GCNO activity in the cytosol.
Specific activity in cytosol could be thus higher than in the
particulate fraction, as observed in platelets.
As previously described [32,33], in the present study
Ca2+ inhibited cytosolic GCNO. We found that membrane-
bound GCNO was also inhibited by Ca2+ and that in the two
cell fractions GCNO showed a biphasic pattern very similar
to that recently described in GCNO purified from bovine
lung [34]. IC50 values calculated for the low affinity sites for
Ca2+ were the same for cytosolic and particulate GCNO and
comparable with those previously reported [34], and IC50
values for the high affinity sites of GCNO were also similar
in both cell fractions. But, besides this inhibitory effect of
Ca2+ when added to the assay medium (mediated by a direct
binding of Ca2+ to GCNO), we observed in this study effects
of increasing cytosolic Ca2+ in intact cells that had not been
previously described. These effects are persistent, and can
be detected after cell homogeneization in the presence of
Ca2+ quelating agents. Importantly, the different agents used
to increased cytosolic Ca2+ concentration have different
effects, suggesting distinct roles for different levels of
physiological concentrations of intracellular Ca2+ or for
different subcellular location of these increases. The
moderate and slow Ca2+ increase evoked by thapsigargin
did not alter GCNO activity associated to the particulate
fraction, and inhibited GCNO in the cytosolic fraction, while
the more marked Ca2+ increase elicited by the Ca2+
ionophore A23187 increased several times GCNO activity
in the particulate fraction without significant effects on
cytosolic GCNO. In parallel with the increase in activity in
the particulate fraction, A231287 increased the amount of
h1 subunit associated to this fraction. This is similar to the
translocation previously observed in activated platelets [15].
However, the change in quantity of h1 associated to the
particulate fraction was much smaller than the change
observed in GCNO activity. A critical point is that for the
different conditions assayed, cytosolic and membrane-
associated GCNO were found to respond differentially to
increased cytosolic Ca2+.
A potential explanation for the increase in specific activity
of membrane-associated GCNO induced byA23187 is that the
increase in cytosolic Ca2+ concentration induced by the drug
causes a modification in the phosphorylation status of the
enzyme. Few studies have analyzed GCNO regulation by
phosphorylation with conflicting results. Some studies
suggested that GCNO phosphorylation increases its activity.
Both in vitro phosphorylation by PKC and PKA [11] and in
vivo phosphorylation by PKA [35] have been described to
increase GCNO activity, while dephosphorylation of the h1
subunit has been associated to a decrease in GCNO activity
[23]. Avery recent study has described, in contrast, a decrease
in GCNO activity associated to an increase in GCNO
phosphorylation in response to PKG activation [12]. In the
present study, the protein kinase inhibitors staurosporin and
KN-93 activated membrane-associated GCNO sinergically
with A23187 suggesting that in cardiomyocytes intracellular
Ca2+ potentiates GCNO activity probably by promoting GCNO
dephosphorylation. Although, direct evidence of GCNO
dephosphorylation in response to A23187 could not be
obtained, the evidence obtained in platelets supports this
hypothesis. However, the fact that the protein phosphatase
inhibitor okadaic acid also enhanced the response to A23187
suggest that regulation of GCNO activity by phosphorylation
may be complex. A protein phosphatase activated by
phosphorylation, as found in chromaffin cells [23], or an
additional regulatory site (in the a1 subunit or in some of
regulatory proteins recently described: as Hsp90 [16], Hsp70
[36], or CCTD [37]) that would increase GCNO activity after
phosphorylation could explain the results. To sum up, our
observations suggest that the increase in cytosolic Ca2+
induced by A23187 has two opposite effects on NO-mediated
cGMP synthesis: a direct inhibitory effect and an indirect
stimulatory effect mediated by h1 dephosphorylation result-
ing in membrane-associated GCNO activation.
The present study provides information that strongly
suggests an important functional role of GCNO localized in
the membrane fraction of cardiomyocytes. Our results show
that a profound inhibition of cytosolic GCNO do not
significantly affect the cell response to SNAP, while
activation of the particulate fraction markedly increases it.
As shown in Fig. 9, cGMP synthesis in the entire cell
correlates well with changes in membrane GCNO activity,
but not with changes in cytosolic GCNO activity. This is in
agreement with recent results in bovine aortic endothelial
cells, indicating a decrease cell-response to NO stimulation
when the formation of a membrane-associated protein
complex between eNOS, HSP90 and GCNO is inhibited
[16].
The association of a fraction of GCNO to cell membrane
in cardiomyocytes, the important role of membrane-asso-
ciated GCNO on the cell response to NO, and the fact that
the regulation of its activity differs from that of the cytosolic
enzyme, may be of great relevance for the better under-
standing of pathophysiological conditions in which the NO/
cGMP-pathway is compromised, and in the design of new
therapies for these conditions.
Acknowledgments
Supported by Fondo Investigaciones Sanitarias (Grant
01/3008 and RECAVA), and Comision Interministerial de
Ciencia y Tecnologıa (Grant SAF2002/0759). We thank
Angeles Rojas for her excellent technical work.

L. Agullo et al. / Cardiovascular Research 68 (2005) 65–7474
by guest on June 1, 2013http://cardiovascres.oxfordjournals.org/
Dow
nloaded from
References
[1] Casadei B, Sears CE. Nitric-oxide-mediated regulation of cardiac
contractility and stretch responses. Prog Biophys Mol Biol
2003;82:67–80.
[2] Padilla F, Garcia-Dorado D, Agullo L, Barrabes JA, Inserte J,
Escalona N, et al. Intravenous administration of the natriuretic peptide
urodilatin at low doses during coronary reperfusion limits infarct size
in anesthetized pigs. Cardiovasc Res 2001;51:592–600.
[3] D’Souza SP, Yellon DM, Martin C, Schulz R, Heusch G, Onody A,
et al. B-type natriuretic peptide limits infarct size in rat isolated hearts
via KATP channel opening. Am J Physiol Heart Circ Physiol
2003;284:1592–600.
[4] Ockaili R, Salloum F, Hawkins J, Kukreja RC. Sildenafil (Viagra)
induces powerful cardioprotective effect via opening of mitochondrial
KATP channels in rabbits. Am J Physiol Heart Circ Physiol
2002;283:1263–9.
[5] Ahluwalia A, Foster P, Scotland RS, McLean PG, Mathur A, Perretti
M, et al. Antiinflammatory activity of soluble guanylate cyclase:
cGMP-dependent down-regulation of P-selectin expression and
leukocyte recruitment. Proc Natl Acad Sci U S A 2004;101:1386–91.
[6] Izumi T, Saito Y, Kishimoto I, Harada M, Kuwahara K, Hamanaka I,
et al. Blockade of the natriuretic peptide receptor guanylyl cyclase-A
inhibits NF-nB activation and alleviates myocardial ischemia/reperfu-
sion injury. J Clin Invest 2001;108:203–13.
[7] Hempel AM, Friedrich M, Schluter KD, Forssmann WG, Kuhn M,
Piper HM. ANP protects against reoxygenation-induced hypercon-
tracture in adult cardiomyocytes. Am J Physiol 1997;273:244–9.
[8] Kodani E, Xuan YT, Takano H, Shinmura K, Tang XL, Bolli R. Role
of cyclic guanosine monophosphate in late preconditioning in
conscious rabbits. Circulation 2002;105:3046–52.
[9] Bellamy TC, Garthwaite J. Sub-second kinetics of the nitric oxide
receptor, soluble guanylyl cyclase, in intact cerebellar cells. J Biol
Chem 2001;276:4287–92.
[10] Kostic TS, Tomic M, Andric SA, Stojilkovic SS. Calcium-independent
and cAMP-dependent modulation of soluble guanylyl cyclase activity
by G protein-coupled receptors in pituitary cells. J Biol Chem
2002;277:16412–8.
[11] Zwiller J, Revel MO, Malviya AN. Protein kinase C catalyzes
phosphorylation of guanylate cyclase in vitro. J Biol Chem
1985;260:1350–3.
[12] Murthy KS. Modulation of soluble guanylate cyclase activity by
phosphorylation. Neurochem Int 2004;45:845–51.
[13] Russwurm M, Wittau N, Koesling D. Guanylyl cyclase/PSD-95
interaction. Targeting of the nitric oxide-sensitive a2h1 guanylyl
cyclase to synaptic membranes. J Biol Chem 2001;276:44647–52.
[14] Feussner M, Richter H, Baum O, Gossrau R. Association of soluble
guanylate cyclase with the sarcolemma of mammalian skeletal muscle
fibers. Acta Histochem 2001;103:265–77.
[15] Zabel U, Kleinschnitz C, Oh P, Nedvetsky P, Smolenski A, Muller
H, et al. Calcium-dependent membrane association sensitizes
soluble guanylyl cyclase to nitric oxide. Nat Cell Biol 2002;4:
307–11.
[16] Venema RC, Venema VJ, Ju H, Harris MB, Snead C, Jilling T, et al.
Novel complexes of guanylate cyclase with heat shock protein 90 and
nitric oxide synthase. Am J Physiol Heart Circ Physiol
2003;285:669–78.
[17] Agullo L, Garcia-Dorado D, Escalona N, Ruiz-Meana M, Inserte J,
Soler-Soler J. Effect of ischemia on soluble and particulate guanylyl
cyclase-mediated cGMP synthesis in cardiomyocytes. Am J Physiol
Heart Circ Physiol 2003;284:2170–6.
[18] Massaguer A, Engel P, Perez-del-Pulgar S, Bosch J, Pizcueta P.
Production and characterization of monoclonal antibodies against
conserved epitopes of P-selectin. Tissue Antigens 2000;56:117–28.
[19] Garcıa-Cardena G, Oh P, Liu J, Schnitzer JE, Sessa WC. Targeting of
nitric oxide synthase to endothelial cell caveolae via palmitoylation:
implications for nitric oxide signaling. Proc Natl Acad Sci U S A
1996;93:6448–53.
[20] Ruiz-Meana M, Garcia-Dorado D, Lane S, Pina P, Inserte J, Mirabet
M, et al. Persistence of gap junction communication during myocar-
dial ischemia. Am J Physiol Heart Circ Physiol 2001;280:2563–71.
[21] Wykes V, Garthwaite J. Membrane-association and the sensitivity of
guanylyl cyclase-coupled receptors to nitric oxide. Br J Pharmacol
2004;141:1087–90.
[22] Kempfert J, Behrends. Analysis of nitric oxide-sensitive guanylyl
cyclase in human platelets before and after aggregation. Platelets
2003;14:429–35.
[23] Ferrero R, Rodrıguez-Pascual F, Miras-Portugal MT, Torres M. Nitric
oxide-sensitive guanylyl cyclase activity inhibition through cyclic
GMP-dependent dephosphorylation. J Neurochem 2000;75:2029–39.
[24] Baltrons MA, Saadoun S, Agullo L, Garcıa A. Regulation by calcium
of the nitric oxide/cyclic GMP system in cerebellar granule cells and
astroglia in culture. J Neurosci Res 1997;49:333–41.
[25] Agullo L, Garcia-Dorado D, Escalona N, Inserte J, Ruiz-Meana M,
Barrabes JA, et al. Hypoxia and acidosis impair cGMP synthesis in
microvascular coronary endothelial cells. Am J Physiol Heart Circ
Physiol 2002;283:917–25.
[26] Papapetropoulos A, Abou-Mohamed G, Marczin N, Murad F,
Caldwell RW, Catravas JD. Downregulation of nitrovasodilator-
induced cyclic GMP accumulation in cells exposed to endotoxin or
interleukin-1 beta. Br J Pharmacol 1996;118:1359–66.
[27] Kloss S, Bouloumie A, Mulsch A. Aging and chronic hypertension
decrease expression of rat aortic soluble guanylyl cyclase. Hyper-
tension 2000;35:43–7.
[28] Laber U, Kober T, Schmitz V, Schrammel A, Meyer W, Mayer B, et al.
Effect of hypercholesterolemia on expression and function of vascular
soluble guanylyl cyclase. Circulation 2002;105:855–60.
[29] Mulsch A, Oelze M, Kloss S, Mollnau H, Topfer A, Smolenski A,
et al. Effects of in vivo nitroglycerin treatment on activity and
expression of the guanylyl cyclase and cGMP-dependent protein
kinase and their downstream target vasodilator-stimulated phospho-
protein in aorta. Circulation 2001;103:2188–94.
[30] Sinnaeve P, Chiche JD, Nong Z, Varenne O, Van Pelt N, Gillijns H,
et al. Soluble guanylate cyclase a1 and h1 gene transfer increases NO
responsiveness and reduces neointima formation after balloon injury
in rats via antiproliferative and antimigratory effects. Circ Res
2001;88:103–9.
[31] Grabow M, Chakraborty G, Ledeen RW. Characterization of guanylyl
cyclase in purified myelin. Neurochem Res 1996;21:457–62.
[32] Parkinson SJ, Jovanovic A, Jovanovic S, Wagner F, Terzic A,
Waldman SA. Regulation of nitric oxide-responsive recombinant
soluble guanylyl cyclase by calcium. Biochemistry 1999;38:6441–8.
[33] Serfass L, Carr HS, Aschenbrenner LM, Burstyn JN. Calcium ion
downregulates soluble guanylyl cyclase activity: evidence for a two-
metal ion catalytic mechanism. Arch Biochem Biophys 2001;387:
47–56.
[34] Kazerounian S, Pitari GM, Ruiz-Stewart I, Schulz S, Waldman SA.
Nitric oxide activation of soluble guanylyl cyclase reveals high and
low affinity sites that mediate allosteric inhibition by calcium.
Biochemistry 2002;41:3396–404.
[35] Kostic TS, Andric SA, Stojilkovic SS. Receptor-controlled phosphor-
ylation of a1 soluble guanylyl cyclase enhances nitric oxide-dependent
cGMP production in pituitary cells. Mol Endocrinol 2004;18:458–70.
[36] Balashova N, Chang FJ, Lamothe M, Sun Q, Beuve A. Character-
ization of a novel type of endogenous activator of soluble guanylyl
cyclase. J Biol Chem 2005;280:2186–96.
[37] Hanafy KA, Martin E, Murad F. CCTeta, a novel soluble guanylyl
cyclase-interacting protein. J Biol Chem 2004;279:46946–53.