astrocytes produce the antiinflammatory and neuroprotective agent hydrogen sulfide
TRANSCRIPT

A
aicCbcoHcP©
K
1
aubWeHLAos(oc
0d
Neurobiology of Aging 30 (2009) 1523–1534
Rapid communication
Astrocytes produce the antiinflammatory and neuroprotectiveagent hydrogen sulfide
Moonhee Lee, Claudia Schwab, Sheng Yu, Edith McGeer, Patrick L. McGeer ∗Kinsmen Laboratory of Neurological Research, University of British Columbia, 2255 Wesbrook Mall, Vancouver, BC, Canada, V6T 1Z3
Received 14 May 2009; received in revised form 1 June 2009; accepted 5 June 2009Available online 23 July 2009
bstract
Hydrogen sulfide (H2S) is an essential physiological product in brain. We investigated the expression of cystathionine-�-synthase (CBS)nd cystathionine-�-lyase (CGL), the two H2S synthesizing enzymes, in human cell lines and in human brain. Only astrocytes were stronglymmunostained for CBS. Cultured astrocytes synthesized H2S at the rate of 15.06 �mol/g protein/h, which was 7.57 fold higher than microglialells, 10.27 fold higher than SH-SY5Y cells and 11.32 fold higher than NT-2 cells. The H2S synthesis in all these cell types was inhibited by theBS inhibitor hydroxylamine, but not by the CGL inhibitor propargylglycine (PAG). Synthesis of H2S by HUVEC cells was inhibited by PAGut not by hydroxylamine indicating that these vascular cells utilize CGL but not CBS. Inflammatory activation of microglia and astrocytesaused induction of NF�B, release of the inflammatory mediators TNF�, IL-6 and nitrite ions, down-regulation of CBS, and down-regulationf H2S synthesis. There was no effect of such treatment on HUVEC cells. The effects were partially reversed by pretreatment of cells with the
2S releasing agent NaSH. These data indicate that H2S is an endogenous antiinflammatory and neuroprotective agent under the syntheticontrol of CBS. H2S releasing drugs may have therapeutic potential in neurodegenerative disorders of aging such as Alzheimer disease andarkinson disease.2009 Elsevier Inc. All rights reserved.
son dis
e2fl
tsOa4rtmh
eywords: Cystathionine-�-synthase; Microglia; Alzheimer disease; Parkin
. Introduction
Hydrogen sulfide (H2S) is best known as a toxic gas withn extremely unpleasant odor. Yet it is a physiological prod-ct generated by all tissues of the body. Concentrations haveeen estimated to be as high as 50–160 �M (Qu et al., 2008;arenycia et al., 1989). In keeping with such high appar-
nt concentrations, several important biological actions of2S have been identified (for reviews see Kamoun, 2004;owicka and Beltowski, 2007; Qu et al., 2008; Szabo, 2007).t synapses, it is a neuromodulator, facilitating the inductionf hippocampal long-term potentiation (Kimura, 2002). Inmooth muscle, it enhances the relaxing effect of nitric oxide
Hosoki et al., 1997). Pharmacologically, it has a direct effectn K-ATP channel currents (Zhao et al., 2001). In brain cellultures, it stimulates production of cAMP with downstream∗ Corresponding author. Tel.: +1 604 822 7377; fax: +1 604 822 7086.E-mail address: [email protected] (P.L. McGeer).
meiaBy
197-4580/$ – see front matter © 2009 Elsevier Inc. All rights reserved.oi:10.1016/j.neurobiolaging.2009.06.001
ease; Aging; Neuroinflammation; NaSH
ffects on cAMP stimulated intracellular pathways (Kimura,000). So far, a possible role for H2S as an endogenous antiin-ammatory agent in brain has not been extensively evaluated.
Evaluation involves an understanding of how H2S is syn-hesized in brain tissue. There are two differing routes of H2Synthesis, although both involve l-cysteine as the substrate.ne utilizes cystathionine-�-synthase (CBS, EC 4.2.1.22)
nd the other cystathionine-�-lyase (cystathionase, CGL, EC.4.1.1). CBS catalyses conversion of cysteine to serine withelease of H2S, while CGL catalyses conversion of cysteineo pyruvate with release of H2S and NH3. These differing
etabolic routes suggest that the enzymes involved mayave differing biological roles even though they have a com-on end product in H2S. The cells expressing these separate
nzymes may have distinct H2S related functions. This is
ndicated by reports that H2S production in brain is associ-ted with CBS while that in vessels is associated with CGL.rain H2S synthesis is reduced by the CBS inhibitor hydrox-lamine, but not by the CGL inhibitor d,l-propargylglycine
1 y of Ag
(Cikws
ehatofwaeHef
tOc2((gHAtt
2
2
f(aeaa
amscfswatca
dmsbcabO(LTscNts
ibtd(pCit4mtrlaKtP
2
fhRhta
sdwpoD
524 M. Lee et al. / Neurobiolog
PAG) (Abe and Kimura, 1996). In contrast, blockade ofGL by PAG completely abolishes H2S production in arter-
es (Zhao et al., 2001) but not in brain. In mouse models, CBSnockouts cannot synthesize H2S in brain (Eto et al., 2002),hile CGL deletion results in a marked reduction of H2S in
erum, heart and aorta (Yang et al., 2008).To investigate further the role of H2S in brain, we first
xamined the expression of CBS and CGL in post mortemuman brain. We found that CBS is strongly expressed bystrocytes while CGL is expressed only in some vessels. Wehen examined various human cell types for their expressionf CBS and CGL and their ability to synthesize H2S. Weound that astrocytes are the most powerful producers of H2S,ith much smaller quantities being produced by microglia
nd neuroblastoma cells. We further investigated the influ-nce of sodium hydrosulfide (NaSH), a known releaser of2S, on the production of inflammatory mediators, the gen-
ration of neurotoxins, and on the induction of NF�B. Weound that it attenuated all of these effects.
We conclude that H2S is an endogenous antiinflamma-ory and neuroprotective agent. It is a powerful antioxidant.xidation products have been implicated in the aging pro-
ess and longevity (Bonnefoy et al., 2002; Calabrese et al.,008), as well as the pathogenesis of Alzheimer disease (AD)Butterfield et al., 2006; Pratico, 2008), Parkinson diseasePD) (Danielson and Andersen, 2008) and other neurode-enerative diseases (Sayre et al., 2001). A deficiency of2S production could contribute to all of these conditions.ccordingly, we propose that H2S releasing drugs may have
herapeutic potential in CNS disorders such as AD and PDhat are characterized by a neuroinflammatory process.
. Materials and methods
.1. Immunostaining of post mortem human brain tissue
Five cases without neurological disease were selectedrom our brain bank at the University of British Columbia4M, 1F, aged 76–89 years). In all five cases the motor cortexnd hippocampus were examined. In two cases, additionalxamination of the substantia nigra, basal ganglia, thalamus,mygdala, cerebellum, entorhinal cortex, occipital cortex andngular cortex was carried out.
Brain tissues had been fixed in 4% paraformaldehyde,nd, after 3–4 days, transferred to a 15% buffered sucroseaintenance solution. For immunohistochemistry, 30 �m
ections were cut on a freezing microtome (American Opti-al Corporation, Buffalo, NY). The sections were treatedor 30 min with 0.5% H2O2 in 0.01 M phosphate bufferedaline, pH 7.4, containing 0.3% Triton X-100 (PBS-T). Theyere transferred into 5% skim milk in PBS-T for 30 min,
nd incubated either for 72 h at 4 ◦C, or overnight at roomemperature, with anti-cystathionine �-synthase (CBS, M01,lone 3E1, Abnova Corp., Taipei, Taiwan, dilution 1:500) ornti-cystathionase (ab54573, Abcam Inc., Cambridge, MA,
dt1t
ing 30 (2009) 1523–1534
ilution 1:200). Sections were next treated with a rabbit-antiouse biotinylated secondary antibody (DAKO, Missis-
auga, Ontario, 1:2000) for 2 h at room temperature, followedy incubation in avidin-biotinylated horseradish peroxidaseomplex (DAKO, Mississauga, Ontario, 1:10.000) for 1 ht room temperature. Peroxidase labeling was visualizedy incubation in 0.01% 3,3diaminobenzidine (DAB; Sigma,akville, Ontario) containing 1% nickel ammonium sulfate
Fisher Scientific, Ottawa, Ontario), 5 mM imidazole (BDHaboratory Supplies, Poole, UK) and 0.001% H2O2 in 0.05 Mris–HCl buffer, pH 7.6. When a dark purple color developed,ections were washed, mounted on glass slides, air-dried, andoverslipped with Entellan (EMD Biosciences, Gibbstown,J). Controls for immunostaining were performed by omit-
ing the primary antibodies. No staining was observed underuch conditions.
For double immunofluorescence staining, sections werencubated in 5% skim milk in PBS-T for 30 min and then incu-ated overnight at room temperature with a combination ofwo primary antibodies. These combinations were: CBS (1:50ilution) plus a rabbit polyclonal antibody to bovine GFAPZ334, Dako, 1:2000); or CGL (1:50 dilution) with a rabbitolyclonal antibody to smooth muscle actin (ab5694, Abcam,ambridge MA, 1:50). After washing, sections were next
ncubated in the dark for 1 h at room temperature with a mix-ure of fluorophore-labeled secondary antibodies Alexa Fluor88 goat anti-mouse to give a green fluorescence for the pri-ary mouse antibodies and AlexaFluor 546 goat anti-rabbit
o give a red fluorescence for the polyclonal antibodies (Invit-ogen, Burlington ON, Canada; both at 1:500). To reduceipofuscin autofluorescence, sections were next treated withsolution of 0.3% Sudan Black B (Gurr Ltd., London, Unitedingdom) in 70% ethanol for 7 min and washed in PBS three
imes. The sections were then air-dried and coverslipped withrolong Gold (Invitrogen).
.2. Cell culture
Human THP-1, NT-2, U373 and U118 cells were obtainedrom the American Type Culture Collection (ATCC). Theuman neuroblastoma SH-SY5Y cell line was a gift from Dr. Ross, Fordham University, NY. Cells were grown underumidified 5% CO2 and 95% air in DMEM/F12 medium con-aining 10% fetal bovine serum (FBS), 100 IU/ml penicillin,nd 100 �g/ml streptomycin (Invitrogen, Carlsbad, CA).
Human astroglial and microglial cells were isolated fromurgically resected temporal lobe tissue and cultured asescribed previously (Klegeris et al., 2006). Briefly, tissuesere rinsed with PBS and chopped into small (<2 mm3)ieces with a sterile scalpel. They were incubated in 10 mlf 0.25% trypsin solution at 37 ◦C for 20 min. SubsequentlyNase I (from bovine pancreas, Pharmacia Biotech, Baie
’Urfé, PQ, Canada) was added to reach a final concentra-ion of 50 �g/ml. Tissues were incubated for an additional0 min at 37 ◦C. After centrifugation at 275 × g for 10 min,he cell pellet was resuspended in the serum-containing
y of Ag
m(siiDacwwohstcupsap
2
dUlc5tteSpUa1pC51b(mba(wpB
mldtT
1ap
2
avscwwMC1Up
Nwc5atdrpnshwta
2
gdsbdio(lPio2m
M. Lee et al. / Neurobiolog
edium and passed through a 100 �m nylon cell strainerBecton Dickinson, Franklin Lakes, NJ). The cell suspen-ion was then centrifuged (275 × g for 10 min), resuspendednto 10 ml of DMEM-F12 with 10% FBS containing gentam-cin (50 �g/ml), pipeted into tissue culture plates (Bectonickinson) and cultured in a humidified 5% CO2, 95% air
tmosphere at 37 ◦C for 2 h to achieve adherence of microglialells. The non-adherent astrocytes along with myelin debrisere transferred into new culture plates. Adherent astrocytesere allowed to grow in these plates by replacing the mediumnce a week. New passages of cells were generated byarvesting confluent astrocyte cultures using trypsin–EDTAolution (0.25% trypsin with EDTA, Invitrogen Canada) andransferring the solution to new culture plates. Human astro-ytes from up to the fifth passage from two surgical cases weresed in the study. Microglia were obtained from the originallating of cells. The two glial species were easily separatedince microglia grow slowly compared with astrocytes. Whenstrocyte confluency has been reached, even after a singleassage, microglial content is less than 0.5%.
.3. Western blotting
Western blotting on cell lysates was performed asescribed by Lee et al. (2008). Briefly, astrocytes, U118,373, microglia, THP-1, NT-2 and SH-SY5Y cells were
ysed with a lysis buffer (150 mM NaCl, 12 mM deoxy-holic acid, 0.1% Nonidet P 40, 0.1% triton X-100 andmM Tris–EDTA, pH 7.4). The protein concentration of
he cell lysates was then determined using a BCA pro-ein assay reagent kit (Pierce, Rockford, IL). Proteins inach sample were loaded onto gels and separated by 10%DS-PAGE (150 V, 1.5 h). The loading quantities of lysateroteins for CBS analysis were: astrocytes, 50 �g; U373 and118 astrocytoma cells 30 �g; microglia, THP-1, SH-SY5Y
nd NT-2 cells 100 �g. For CGL analysis of all cell types,50 �g of protein per lane was loaded. Following SDS-PAGE,roteins were transferred to a PVDF membrane (Bio-Rad,A) at 30 mA for 2 h. The membranes were blocked with% milk in PBS-T (80 mM Na2HPO4, 20 mM NaH2PO4,00 mM NaCl, 0.1% Tween 20, pH 7.5) for 1 h and incu-ated overnight at 4 ◦C with a monoclonal anti-CBS antibodyM01, clone 3E1, Abnova Corp., Taipei, Taiwan, 1:2000) or aonoclonal anti-CGL antibody (ab54573, Abcam Inc., Cam-
ridge, MA, 1:1000). The membranes were then treated withhorseradish peroxidase-conjugated secondary antibody
Sigma, St. Louis, MO, 1:1000) and the bands visualizedith an enhanced chemiluminescence system and exposure tohotographic film (Hyperfilm ECLTM, Amersham Pharmaciaiotech, Little Chalfont, UK).
Tubulin was used as the housekeeping protein for nor-alizing values. The same amounts of lysate protein were
oaded for each cell type as described above. Tubulin wasetected on membranes with a mouse monoclonal anti-�-ubulin antibody (T-6074, Sigma, St. Louis MO, 1:2000).he secondary antibody was anti-mouse IgG (A3682, Sigma,
ooob
ing 30 (2009) 1523–1534 1525
:3000). Primary antibody incubation was overnight at 4 ◦Cnd secondary antibody incubation was for 3 h at room tem-erature.
.4. Inflammatory stimulation of cells
Human microglia and monocytic THP-1 cells, astrocytesnd U118 and U373 astrocytoma cells, and human umbilicalein endothelial cells (HUVECs) (5 × 105 cells each) wereeeded into 24-well plates in 1 ml of DMEM/F12 mediumontaining 5% FBS. Cells were incubated for 2 days at 37 ◦Cith appropriate inflammatory stimulants. These stimulantsere: 1 �g/ml lipopolysaccharide (LPS, Sigma, St. Louis,O Escherichia coli 055:B5) plus 333 U/ml IFN� (Bachemalifornia, Torrance, CA) for microglia and THP-1 cells;50 U/ml IFN� for astrocytes; 500 U/ml IFN� for U118 and373 astrocytoma cells; and 10 �g/ml LPS or 10 �g/ml LPSlus 150 U/ml IFN� for HUVEC cells.
In some experiments cells were first exposed to 10 �MaSH for 8 h. The reason for choosing the 8 h time periodas to provide adequate time for NaSH to accumulate intra-
ellularly. The solutions were centrifuged at 1000 rpm formin, resuspended in PBS for washing through two cyclesnd then resuspended in 800 �l DMEM/F12 medium con-aining 5% FBS with or without the inflammatory stimulantsescribed above. After 48 h incubation, the supernatant wasemoved and the adherent cells lysed with the lysis bufferreviously described for Western blotting. For evaluatingeurotoxicity of stimulated glial cell secretions, the separatedupernatants (400 �l) were transferred to undifferentiateduman neuroblastoma SH-SY5Y cells (2 × 105 cells perell) and the cells incubated for 3 days at 37 ◦C before being
ested for their remaining viability by the LDH and MTTssays.
.5. Cell viability measured by the LDH and MTT assays
Cell viability change was evaluated by the lactic dehydro-enase (LDH) assay, which measures release of LDH fromead cells, and the MTT assay, which measures the conver-ion of the tetrazolium salt to a colored formazan producty living cells. The methods, which have previously beenescribed in detail (Klegeris et al., 1999), provide reciprocalnformation on cell viability and therefore complement eachther. Briefly, for the LDH assay, cell culture supernatants100 �l) were pipetted into the wells of 96-well plates, fol-owed by the addition of 15 �l lactate solution (36 mg/ml inBS) and 15 �l iodonitrotetrazolium (INT) solution (2 mg/ml
n PBS). The enzymatic reaction was started by additionf 15 �l of NAD+/diaphorase solution (3 mg/ml NAD+;.3 mg solid/ml diaphorase). After 1 h, optical densities wereeasured with a Model 450 microplate reader (Bio-Rad Lab-
ratories, Richmond, CA) using a 490 nm filter. The amountf LDH released was expressed as a percentage of the valuebtained in comparative wells where cells were 100% lysedy 1% Triton X-100.
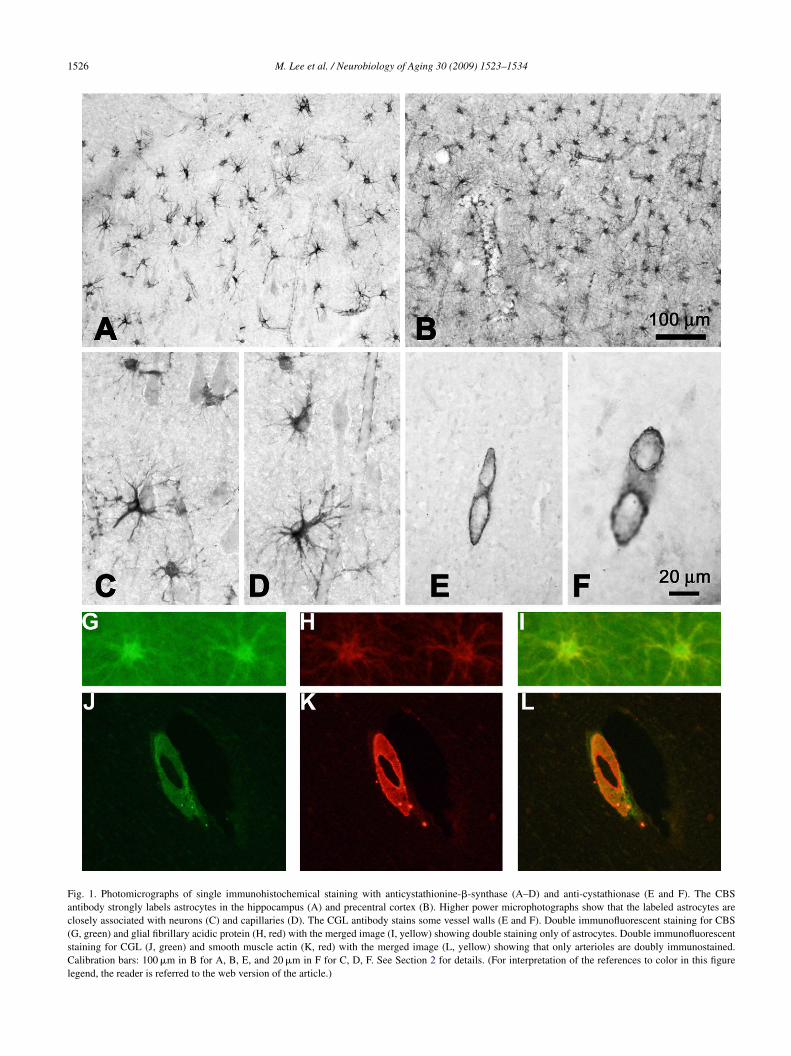
1526 M. Lee et al. / Neurobiology of Aging 30 (2009) 1523–1534
Fig. 1. Photomicrographs of single immunohistochemical staining with anticystathionine-�-synthase (A–D) and anti-cystathionase (E and F). The CBSantibody strongly labels astrocytes in the hippocampus (A) and precentral cortex (B). Higher power microphotographs show that the labeled astrocytes areclosely associated with neurons (C) and capillaries (D). The CGL antibody stains some vessel walls (E and F). Double immunofluorescent staining for CBS(G, green) and glial fibrillary acidic protein (H, red) with the merged image (I, yellow) showing double staining only of astrocytes. Double immunofluorescentstaining for CGL (J, green) and smooth muscle actin (K, red) with the merged image (L, yellow) showing that only arterioles are doubly immunostained.Calibration bars: 100 �m in B for A, B, E, and 20 �m in F for C, D, F. See Section 2 for details. (For interpretation of the references to color in this figurelegend, the reader is referred to the web version of the article.)
y of Ag
drtadsdaav
2
dfip
hhl2la
A2ltm2a
FroETU3ns
M. Lee et al. / Neurobiolog
For the MTT assay, the viability of SH-SY5Y cells wasetermined by adding MTT to the SH-SY5Y cell cultures toeach a final concentration of 1 mg/ml. Following 1 h incuba-ion at 37 ◦C, the dark crystals formed were dissolved bydding SDS/DMF extraction buffer (300 �l, 20% sodiumodecyl sulfate, 50% N,N-dimethylformamide, pH 4.7). Sub-equently plates were incubated overnight at 37 ◦C. Opticalensities at 570 nm were measured by transferring 100 �lliquots to 96-well plates and using a plate reader with anppropriate filter. Data are presented as a percentage of thealue obtained from cells incubated in fresh medium only.
.6. H2S biosynthesis
H2S production levels were measured using a previouslyescribed method (Li et al., 2005). Adherent cells wererst treated with 500 �l ice-cold 100 mM potassium phos-hate buffer (pH 7.4). The mixture was then removed and
apFa
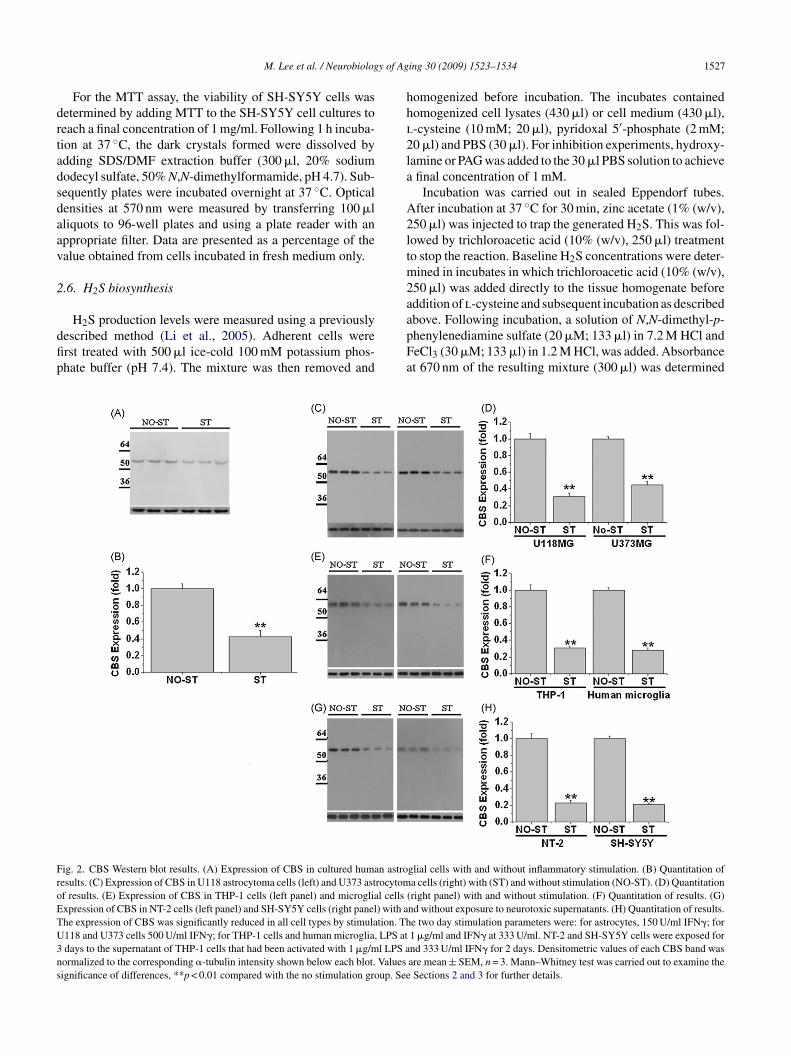
ig. 2. CBS Western blot results. (A) Expression of CBS in cultured human astroesults. (C) Expression of CBS in U118 astrocytoma cells (left) and U373 astrocytomf results. (E) Expression of CBS in THP-1 cells (left panel) and microglial cellsxpression of CBS in NT-2 cells (left panel) and SH-SY5Y cells (right panel) with ahe expression of CBS was significantly reduced in all cell types by stimulation. Th118 and U373 cells 500 U/ml IFN�; for THP-1 cells and human microglia, LPS atdays to the supernatant of THP-1 cells that had been activated with 1 �g/ml LPS aormalized to the corresponding �-tubulin intensity shown below each blot. Valuesignificance of differences, **p < 0.01 compared with the no stimulation group. See
ing 30 (2009) 1523–1534 1527
omogenized before incubation. The incubates containedomogenized cell lysates (430 �l) or cell medium (430 �l),-cysteine (10 mM; 20 �l), pyridoxal 5′-phosphate (2 mM;0 �l) and PBS (30 �l). For inhibition experiments, hydroxy-amine or PAG was added to the 30 �l PBS solution to achievefinal concentration of 1 mM.
Incubation was carried out in sealed Eppendorf tubes.fter incubation at 37 ◦C for 30 min, zinc acetate (1% (w/v),50 �l) was injected to trap the generated H2S. This was fol-owed by trichloroacetic acid (10% (w/v), 250 �l) treatmento stop the reaction. Baseline H2S concentrations were deter-ined in incubates in which trichloroacetic acid (10% (w/v),
50 �l) was added directly to the tissue homogenate beforeddition of l-cysteine and subsequent incubation as described
bove. Following incubation, a solution of N,N-dimethyl-p-henylenediamine sulfate (20 �M; 133 �l) in 7.2 M HCl andeCl3 (30 �M; 133 �l) in 1.2 M HCl, was added. Absorbancet 670 nm of the resulting mixture (300 �l) was determinedglial cells with and without inflammatory stimulation. (B) Quantitation ofa cells (right) with (ST) and without stimulation (NO-ST). (D) Quantitation
(right panel) with and without stimulation. (F) Quantitation of results. (G)nd without exposure to neurotoxic supernatants. (H) Quantitation of results.e two day stimulation parameters were: for astrocytes, 150 U/ml IFN�; for1 �g/ml and IFN� at 333 U/ml. NT-2 and SH-SY5Y cells were exposed fornd 333 U/ml IFN� for 2 days. Densitometric values of each CBS band wasare mean ± SEM, n = 3. Mann–Whitney test was carried out to examine theSections 2 and 3 for further details.

1 y of Aging 30 (2009) 1523–1534
aCae
2
fTmEd
2
Gfrrp
2
ppkdptr(aBwDtoolas
2
notHceb(
(�m
ol/g
prot
ein/
h)in
hum
anas
troc
ytes
and
mic
rogl
ia,U
373
and
U11
8ce
lls,T
HP-
1ce
lls,S
H-S
Y5Y
and
NT-
2ce
lls,a
ndH
UV
EC
sa .
ition
sA
stro
cyte
sU
373
U11
8M
icro
glia
TH
P-1
SH-S
Y5Y
NT-
2H
UV
EC
nocy
stei
ne)
3.0
±0.
66.
13±
0.64
6.63
±0.
470.
38±
0.04
0.4
±0.
060.
26±
0.04
0.31
±0.
030.
24±
0.04
mM
)15
.06
±1.
0228
.65
±2.
0631
.43
±3.
341.
99±
0.28
2.22
±0.
221.
46±
0.24
1.33
±0.
252.
24±
0.08
mM
)ne
(1m
M)
2.82
±0.
7*3.
84±
0.46
*4.
05±
0.33
*0.
53±
0.05
*0.
7±
0.02
*0.
22±
0.00
4*0.
31±
0.05
*1.
94±
0.08
mM
)PA
G14
.96
±0.
8825
.04
±3.
0530
.63
±2.
272.
05±
0.25
2.42
±0.
241.
38±
0.26
1.32
±0.
040.
64±
0.04
*
mM
)ST
5.74
±0.
42*
11.4
3±
1.84
*10
.63
±1.
05*
0.59
±0.
05*
0.66
±0.
04*
0.44
±0.
04*
0.32
±0.
04*
2.26
±0.
1
mM
)N
aSH
45.7
4±
4.73
*66
.89
±6.
84*
64.8
4±
4.73
*7.
85±
0.63
*8.
52±
0.46
*6.
84±
0.58
*5.
97±
0.63
*–
mM
)N
aSH
mm
ator
yST
11.7
4±
1.05
**19
.95
±2.
05**
20.9
5±
1.95
**2.
84±
0.35
**2.
05±
0.17
**1.
23±
0.25
**1.
42±
0.21
**–
2fo
rtr
eatm
ent
deta
ils.
Val
ues
are
mea
n±
SEM
,n
=4.
The
Man
n–W
hitn
eyte
stw
asin
itial
lyca
rrie
dou
tto
test
the
sign
ifica
nce
ofdi
ffer
ence
sbe
twee
ntr
eatm
ent
grou
ps.
The
sign
ifica
nce
ofth
enad
just
edby
appl
ying
the
Bon
ferr
onit
estf
orm
ultip
leco
mpa
riso
ns.
0.01
for
the
adju
sted
sign
ifica
nce
betw
een
trea
tmen
tgro
up2
and
trea
tmen
tgro
ups
3,4,
5,an
d6.
0.01
for
the
adju
sted
sign
ifica
nce
betw
een
trea
tmen
tgro
ups
6an
d7.
528 M. Lee et al. / Neurobiolog
fter10 min using a 96-well microplate reader (Bio-Rad,A). The H2S concentration of each sample was calculatedgainst a calibration curve of NaSH (1–250 �M) and resultsxpressed as �mol/g protein/h.
.7. Measurement of TNFα and IL-6 release
Cytokine levels were measured in cell-free supernatantsollowing 48 h incubation of microglial cells and astrocytes.he cell stimulation protocols were the same as used foreasuring H2S generation. Quantitation was performed withLISA detection kits (Peprotech, NJ) following protocolsescribed by the manufacturer.
.8. Measurements of nitrite release
Accumulation of NO2− was assayed by the standard
riess reaction. After stimulation of cells for 48 h, the cell-ree supernatants were mixed with an equal volume of Griesseagent (Sigma, St. Louis, MO). Samples were incubated atoom temperature for 15 min. Values were recorded from alate reader (excitation 380 nm, emission 540 nm).
.9. NFκB activation assay
Activation of the NF�B inflammatory transcriptionathway in cell lysates was assessed by induction of phospho-65NF�B (Sakurai and Chiba, 1999) and phospho-p38 MAPinase. Western blot analysis of lysates was carried out asescribed previously. For gel separation 100 �g of lysaterotein was loaded in each lane. Following separation andransfer of the proteins to membranes, detection was car-ied out using as primary antibodies anti-phospho-p65NF�B3031, Cell Signaling, Beverly, MA, 1:1000) and rabbitnti human-phospho-p38 MAP kinase (9211, Cell Signaling,everly, MA, 1:2000). The secondary antibody in each caseas a horseradish peroxidase-conjugated anti-IgG (P0448,AKO, Mississauga, Ontario, CA, 1:2000). Incubation with
he primary antibodies was overnight at 4 ◦C and with the sec-ndary antibodies was 3 h at room temperature. Equalizationf protein loading was assessed independently using tubu-in as the housekeeping protein. The primary antibody wasnti-�-tubulin (T6074, Sigma, St Louis MO, 1:2000) and theecondary antibody anti-mouse IgG (A3682, Sigma, 1:3000).
.10. Data analysis
For two-way comparisons, the data were analyzed for sig-ificance by Mann–Whitney (Figs. 1 and 2 and Table 1) orne-way ANOVA tests (Fig. 3 and Tables 2 and 3). For mul-iple comparisons on the effects of added NaSH or levels of
2S generated by CBS or CGL in stimulated or unstimulatedells, they were analyzed by post-hoc Bonferroni testing. Theffects of hydroxylamine or PAG on levels of H2S generatedy CBS or CGL were similarly analyzed for all cell typesTable 1). Ta
ble
1H
2S
gene
ratio
n
Tre
atm
entc
ond
1.B
asal
leve
ls(
2.C
yste
ine
(10
3.C
yste
ine
(10
hydr
oxyl
ami
4.C
yste
ine
(10
(1m
M)
5.C
yste
ine
(10
infla
mm
ator
y6.
Cys
tein
e(1
0(1
0�
M)
7.C
yste
ine
(10
(10
�M
)in
flaa
See
Sect
ion
diff
eren
ces
was
*In
dica
tes
p<
**In
dica
tes
p<
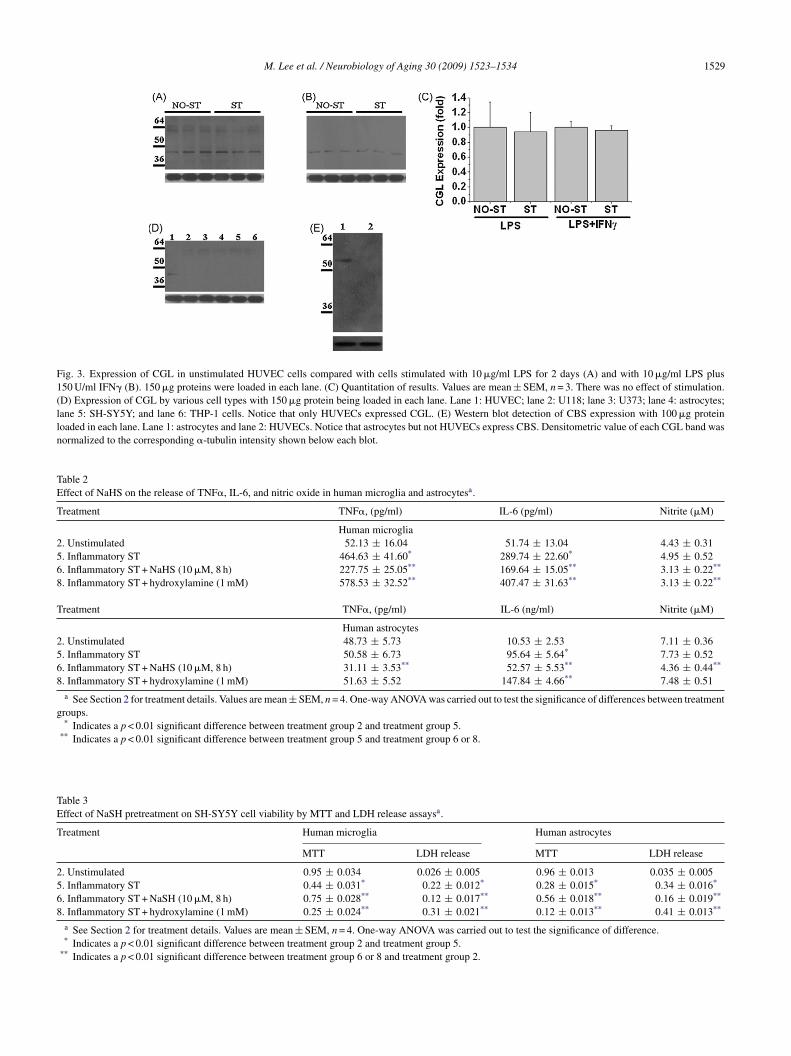
M. Lee et al. / Neurobiology of Aging 30 (2009) 1523–1534 1529
Fig. 3. Expression of CGL in unstimulated HUVEC cells compared with cells stimulated with 10 �g/ml LPS for 2 days (A) and with 10 �g/ml LPS plus150 U/ml IFN� (B). 150 �g proteins were loaded in each lane. (C) Quantitation of results. Values are mean ± SEM, n = 3. There was no effect of stimulation.(D) Expression of CGL by various cell types with 150 �g protein being loaded in each lane. Lane 1: HUVEC; lane 2: U118; lane 3: U373; lane 4: astrocytes;lane 5: SH-SY5Y; and lane 6: THP-1 cells. Notice that only HUVECs expressed CGL. (E) Western blot detection of CBS expression with 100 �g proteinloaded in each lane. Lane 1: astrocytes and lane 2: HUVECs. Notice that astrocytes but not HUVECs express CBS. Densitometric value of each CGL band wasnormalized to the corresponding �-tubulin intensity shown below each blot.
Table 2Effect of NaHS on the release of TNF�, IL-6, and nitric oxide in human microglia and astrocytesa.
Treatment TNF�, (pg/ml) IL-6 (pg/ml) Nitrite (�M)
Human microglia2. Unstimulated 52.13 ± 16.04 51.74 ± 13.04 4.43 ± 0.315. Inflammatory ST 464.63 ± 41.60* 289.74 ± 22.60* 4.95 ± 0.526. Inflammatory ST + NaHS (10 �M, 8 h) 227.75 ± 25.05** 169.64 ± 15.05** 3.13 ± 0.22**
8. Inflammatory ST + hydroxylamine (1 mM) 578.53 ± 32.52** 407.47 ± 31.63** 3.13 ± 0.22**
Treatment TNF�, (pg/ml) IL-6 (ng/ml) Nitrite (�M)
Human astrocytes2. Unstimulated 48.73 ± 5.73 10.53 ± 2.53 7.11 ± 0.365. Inflammatory ST 50.58 ± 6.73 95.64 ± 5.64* 7.73 ± 0.526. Inflammatory ST + NaHS (10 �M, 8 h) 31.11 ± 3.53** 52.57 ± 5.53** 4.36 ± 0.44**
8. Inflammatory ST + hydroxylamine (1 mM) 51.63 ± 5.52 147.84 ± 4.66** 7.48 ± 0.51a See Section 2 for treatment details. Values are mean ± SEM, n = 4. One-way ANOVA was carried out to test the significance of differences between treatment
groups.* Indicates a p < 0.01 significant difference between treatment group 2 and treatment group 5.
** Indicates a p < 0.01 significant difference between treatment group 5 and treatment group 6 or 8.
Table 3Effect of NaSH pretreatment on SH-SY5Y cell viability by MTT and LDH release assaysa.
Treatment Human microglia Human astrocytes
MTT LDH release MTT LDH release
2. Unstimulated 0.95 ± 0.034 0.026 ± 0.005 0.96 ± 0.013 0.035 ± 0.0055. Inflammatory ST 0.44 ± 0.031* 0.22 ± 0.012* 0.28 ± 0.015* 0.34 ± 0.016*
6. Inflammatory ST + NaSH (10 �M, 8 h) 0.75 ± 0.028** 0.12 ± 0.017** 0.56 ± 0.018** 0.16 ± 0.019**
8. Inflammatory ST + hydroxylamine (1 mM) 0.25 ± 0.024** 0.31 ± 0.021** 0.12 ± 0.013** 0.41 ± 0.013**
a See Section 2 for treatment details. Values are mean ± SEM, n = 4. One-way ANOVA was carried out to test the significance of difference.* Indicates a p < 0.01 significant difference between treatment group 2 and treatment group 5.
** Indicates a p < 0.01 significant difference between treatment group 6 or 8 and treatment group 2.

1 y of Ag
3
witia(wsccabwccavaatCiacb
3b
scaSdooufI1iscpsba(tfa(
tsltaannrfssaamC
od4dw(pttoqFetecr
3
tHfsrsccctmfta
530 M. Lee et al. / Neurobiolog
. Results
Fig. 1 shows the result of immunostaining of human brainith antibodies to CBS and CGL. In all brain areas exam-
ned, the antibody against CBS strongly stained astrocyteshroughout white and grey matter (Fig. 1A and B). CBS label-ng was present in the cell bodies and processes, extendingll the way into the astrocytic end feet attached to capillariesFig. 1C). Some astrocytes were found to be in close contactith neurons (Fig. 1D). Astrocytes were stained in the sub-
tantia nigra, hippocampus, amygdala, motor cortex, angularortex, occipital cortex, basal ganglia and thalamus. No brainell type other than astrocytes showed detectable staining inny of these areas. No labeling was found with the CGL anti-ody except for a small subset of medium-caliber vesselsith strongly stained walls (Fig. 1E and F). To verify the
ell types that are immunostained, double immunofluores-ence was carried out with GFAP as the specific marker forstrocytes and smooth muscle actin as the specific marker foressel arterioles. Double immunostaining was found for CBSnd astrocytes (Fig. 1G–I) and for CGL and smooth musclectin (Fig. 1J–L). These data demonstrate that astrocytes arehe dominant cell type in mature human brain expressingBS and that arterioles are the dominant structure express-
ng CGL. CBS and CGL may be expressed by other cell typest levels below those that can be detected by immunohisto-hemistry as indicated by the cell culture results describedelow.
.1. Expression of CBS and CGL proteins by Westernlots
Fig. 2 shows Western blot results of CBS expres-ion in astrocytes (Fig. 2A); U118 and U373 astrocytomaells (Fig. 2C); microglia and THP-1 cells (Fig. 2E);nd NT-2 neuronally committed teratocarcinoma cells andH-SY5Yneuroblastoma cells (Fig. 2G). All cell typesemonstrated a single band consistent with the CBS MWf 63 kDa and all cell types showed a significant diminutionf that band following exposure to an inflammatory stim-lus. For astrocytes the stimulation was 150 U/ml of IFN�or 2 days; for U118 and U373 cells it was 500 U/ml ofFN� for 2 days; for THP-1 and microglial cells it was�g/ml LPS plus 333 U/ml IFN� for 2 days; and for tox-
city to NT-2 and SH-SY5Y cells it was the supernatant fromtimulated THP-1 cells. Below each of these figures, theorresponding tubulin band is shown for normalization ofrotein loading. The quantitative reduction in CBS expres-ion from the non-stimulated level to the stimulated levely each cell type following such stimulation was as follows:strocytes to 0.43 ± 0.07 (Fig. 2B,); U118 cells to 0.31 ± 0.04Fig. 2D); U373 cells to 0.45 ± 0.07 (Fig. 2D); THP-1 cells
o 0.31 ± 0.02 (Fig. 2F); microglia to 0.28 ± 0.03 (Fig. 2F);or supernatant toxicity to NT-2 cells 0.28 ± 0.03 (Fig. 2G);nd for supernatant toxicity to SH-SY5Y cells 0.21 ± 0.03Fig. 2G). In all cases n = 3, p < 0.01. It is noteworthy thatfeat
ing 30 (2009) 1523–1534
he expression of CBS in astrocytoma cells was considerablytronger than in astrocytes despite a lower level of proteinoading (30 versus 50 �g protein in each lane). This indicateshat a stronger inflammatory stimulation is required to bringbout a reduction in CBS. Indeed, when IFN� stimulation ofstrocytoma cells was tested at the same level as astrocytes,o reduction in intensity of the CBS band was observed (dataot shown). NT-2 and SH-SY5Y neuroblastoma cells do notespond to direct inflammatory stimulation, but they are pro-oundly affected by exposure to the toxic supernatants oftimulated THP-1 and microglial cells. In the experimentshown in Fig. 2G and H, the NT-2 and SH-SY5Y cells weressessed after treatment for 3 days with supernatants fromctivated THP-1 cells. Overall, these results show that inflam-atory stimulation typically causes a 2.5–3 fold reduction inBS expression.
Fig. 3 illustrates very different results from Western blotsf CGL expression. Only HUVEC cells demonstrated aetectable band. It corresponded with the known MW of3.6 kDa (Fig. 3A). This expression was not significantlyown-regulated by pretreatment with 10 �g/ml of LPS orith 10 �g/ml of LPS plus 150 U/ml of IFN� for 2 days
Fig. 3B). The much higher dose of LPS in these cells com-ared with microglia and THP-1 cells was chosen becausehis dose has been shown to be necessary for stimulation ofhese cells (Sun et al., 2008). The results indicate a resistancef this enzyme to inflammatory down-regulation despite ade-uate stimulation of the HUVECs. The cell types shown inig. 2, all of which expressed CBS, were not observed toxpress CGL (Fig. 3C), while HUVECs were not observedo express CBS (Fig. 3D). These data indicate that H2S is gen-rated by a distinctly different mechanism in vascular cellsompared with brain tissue cells, and that they have a differentesponse to inflammatory stimulation.
.2. Generation of H2S
Table 1 records the results of H2S generation in cell cul-ures after various treatments. Treatment 1 shows the basal
2S levels found in unstimulated cells that had been culturedor several days. Presumably these levels represent steadytate concentrations of intracellularly produced H2S. Theseelative concentrations were consistent with the CBS expres-ion levels shown by Western blotting in Fig. 2. The highestoncentrations were found in U118 and U373 astrocytomaells (6.63 and 6.13 �mol/g protein). This was followed byultured astrocytes, which had a base level of 3.0 �mol/g pro-ein. This concentration was 7.9 fold higher than in cultured
icroglial cells, 9.7 fold higher than in NT-2 cells and 11.5old higher than in SH-SY5Y cells. Treatment 2 evaluatedhe rate of H2S synthesis above this background followingddition of 10 mM cysteine substrate and incubation at 37 ◦C
or 30 min. U118 and U373 astrocytoma cells had the high-st synthesis rates. Astrocytes, at 15.06 �mol/g/h, had a ratepproximately one half that of the astrocytoma cell lines. Buthis level was 7.57 fold higher than for microglial cells, 11.3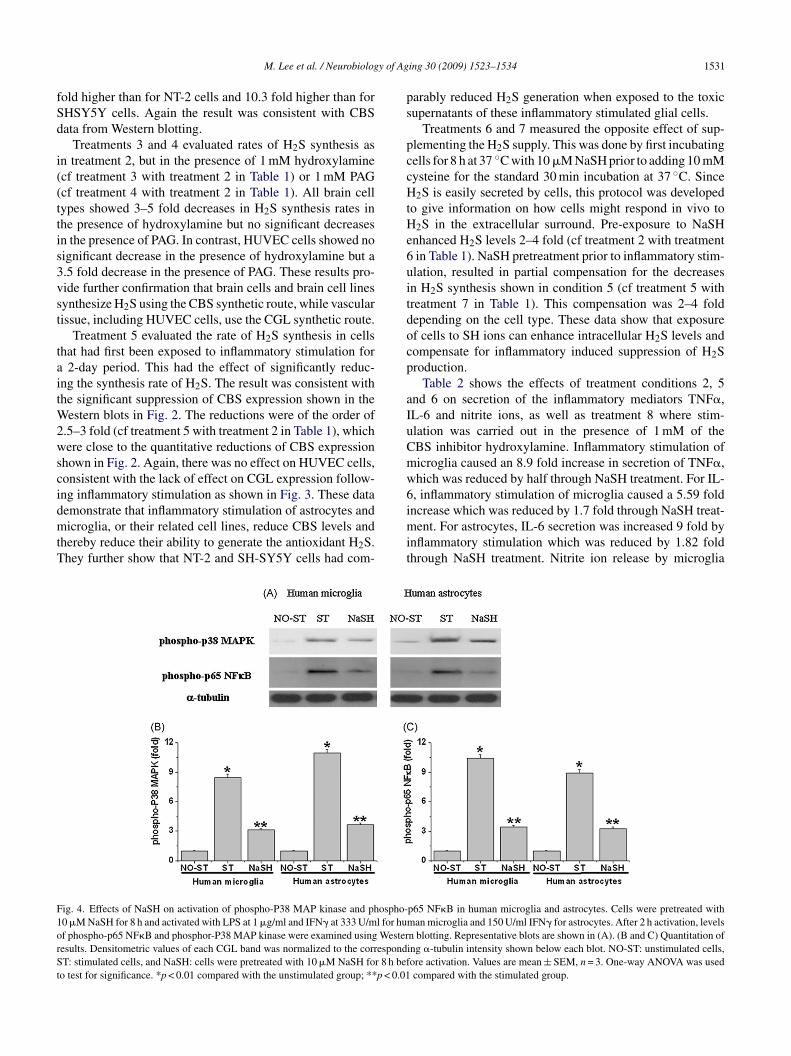
y of Ag
fSd
i((ttis3vst
taitW2wscidmtT
ps
pccHtHe6uitdocp
aIuCmw6
F1orSt
M. Lee et al. / Neurobiolog
old higher than for NT-2 cells and 10.3 fold higher than forHSY5Y cells. Again the result was consistent with CBSata from Western blotting.
Treatments 3 and 4 evaluated rates of H2S synthesis asn treatment 2, but in the presence of 1 mM hydroxylaminecf treatment 3 with treatment 2 in Table 1) or 1 mM PAGcf treatment 4 with treatment 2 in Table 1). All brain cellypes showed 3–5 fold decreases in H2S synthesis rates inhe presence of hydroxylamine but no significant decreasesn the presence of PAG. In contrast, HUVEC cells showed noignificant decrease in the presence of hydroxylamine but a.5 fold decrease in the presence of PAG. These results pro-ide further confirmation that brain cells and brain cell linesynthesize H2S using the CBS synthetic route, while vascularissue, including HUVEC cells, use the CGL synthetic route.
Treatment 5 evaluated the rate of H2S synthesis in cellshat had first been exposed to inflammatory stimulation for
2-day period. This had the effect of significantly reduc-ng the synthesis rate of H2S. The result was consistent withhe significant suppression of CBS expression shown in the
estern blots in Fig. 2. The reductions were of the order of.5–3 fold (cf treatment 5 with treatment 2 in Table 1), whichere close to the quantitative reductions of CBS expression
hown in Fig. 2. Again, there was no effect on HUVEC cells,onsistent with the lack of effect on CGL expression follow-ng inflammatory stimulation as shown in Fig. 3. These data
emonstrate that inflammatory stimulation of astrocytes andicroglia, or their related cell lines, reduce CBS levels andhereby reduce their ability to generate the antioxidant H2S.hey further show that NT-2 and SH-SY5Y cells had com-
imit
ig. 4. Effects of NaSH on activation of phospho-P38 MAP kinase and phospho-0 �M NaSH for 8 h and activated with LPS at 1 �g/ml and IFN� at 333 U/ml for humf phospho-p65 NF�B and phosphor-P38 MAP kinase were examined using Westeresults. Densitometric values of each CGL band was normalized to the correspondT: stimulated cells, and NaSH: cells were pretreated with 10 �M NaSH for 8 h bef
o test for significance. *p < 0.01 compared with the unstimulated group; **p < 0.01
ing 30 (2009) 1523–1534 1531
arably reduced H2S generation when exposed to the toxicupernatants of these inflammatory stimulated glial cells.
Treatments 6 and 7 measured the opposite effect of sup-lementing the H2S supply. This was done by first incubatingells for 8 h at 37 ◦C with 10 �M NaSH prior to adding 10 mMysteine for the standard 30 min incubation at 37 ◦C. Since2S is easily secreted by cells, this protocol was developed
o give information on how cells might respond in vivo to2S in the extracellular surround. Pre-exposure to NaSH
nhanced H2S levels 2–4 fold (cf treatment 2 with treatmentin Table 1). NaSH pretreatment prior to inflammatory stim-lation, resulted in partial compensation for the decreasesn H2S synthesis shown in condition 5 (cf treatment 5 withreatment 7 in Table 1). This compensation was 2–4 foldepending on the cell type. These data show that exposuref cells to SH ions can enhance intracellular H2S levels andompensate for inflammatory induced suppression of H2Sroduction.
Table 2 shows the effects of treatment conditions 2, 5nd 6 on secretion of the inflammatory mediators TNF�,L-6 and nitrite ions, as well as treatment 8 where stim-lation was carried out in the presence of 1 mM of theBS inhibitor hydroxylamine. Inflammatory stimulation oficroglia caused an 8.9 fold increase in secretion of TNF�,hich was reduced by half through NaSH treatment. For IL-, inflammatory stimulation of microglia caused a 5.59 fold
ncrease which was reduced by 1.7 fold through NaSH treat-ent. For astrocytes, IL-6 secretion was increased 9 fold bynflammatory stimulation which was reduced by 1.82 foldhrough NaSH treatment. Nitrite ion release by microglia
p65 NF�B in human microglia and astrocytes. Cells were pretreated withan microglia and 150 U/ml IFN� for astrocytes. After 2 h activation, levels
n blotting. Representative blots are shown in (A). (B and C) Quantitation ofing �-tubulin intensity shown below each blot. NO-ST: unstimulated cells,ore activation. Values are mean ± SEM, n = 3. One-way ANOVA was usedcompared with the stimulated group.

1 y of Ag
wbWiibr1TtThadp
6nuemoTrtwcmset5Ho
ttuptltfIIbMkoe
4
H
(cqaeaeCbhpfprZ
o1o1rtihca
isHpnma
tmNotroTp
aworis
532 M. Lee et al. / Neurobiolog
as not affected by stimulation but it was reduced 1.6 foldy NaSH treatment (cf treatments 2, 5 and 6 in Table 2).hen stimulated microglial cells were treated with the CBS
nhibitor hydroxylamine, there was a significant 1.25 foldncrease in TNF�, and a significant 1.37 fold increase in IL-6,ut there was no effect on nitrite ions. There was no effect onelease of TNF� or nitrite ions by astrocytes but a significant.54 fold increase for IL-6 (cf treatments 5 and 8 in Table 2).hese data demonstrate that inhibition of CBS can enhance
he effect of inflammatory stimulation on the production ofNF� and IL-6, indicating that endogenous H2S productionas a protective effect against release of inflammatory medi-tors by stimulated microglia and astrocytes. They furtheremonstrate that externally supplied SH ions can also reduceroduction of inflammatory mediators, including nitrite ions.
Table 3 compares the effects of treatment conditions 2, 5,and 8 on the toxicity of microglial and astrocytic super-
atants towards SH-SY5Y cells. Exposure to supernatants ofnstimulated cells had no effect on SH-SY5Y cells. How-ver exposure to supernatants of inflammatory stimulatedicroglia or astrocytes caused death of approximately 2/3
f the SH-SY5Y cells (cf treatment 5 with treatment 6 inable 3). These toxicity results are consistent with previouseports (Klegeris et al., 2005; Hashioka et al., 2008). Theoxicity was cut by 1.7–2 fold by pretreatment of the cellsith NaSH. These data show that externally applied NaSH
an have a neuroprotective effect through reduction of toxicaterials secreted by glial cells subjected to inflammatory
timuli. The toxicity of the supernatants was significantlynhanced when the stimulated microglia and astrocytes werereated with the CBS inhibitor hydroxylamine (cf treatment
with 8 in Table 3). These data indicate that endogenous2S production has a protective effect against the secretionf toxic materials by stimulated microglia and astrocytes.
Fig. 4 explores by Western blotting the effect of inflamma-ory stimulation of microglia and astrocytes on NF�B induc-ion. This was measured by increased expression upon stim-lation of the specific markers of NF�B induction, phospho-38 MAPK and phospho-p65NF�B. Their ameliorationhrough treatment with NaSH was also measured. Unstimu-ated microglia and astrocytes had barely detectable levels ofhese proteins. But there was a strong induction in microgliaollowing 2 h of exposure to 1 �g/ml LPS plus 333 U/mlFN�, and in astrocytes that had been stimulated by 150 U/mlFN� (Fig. 4A). This induction was significantly reducedy pretreatment with 10 �M NaSH for 8 h for phospho-p38APK (Fig. 4B) and phospho-p65NF�B (Fig. 4C). It is
nown that NF�B is induced by inflammatory stimulationf microglia and astrocytes, and these data demonstrate thatxternally applied NaSH can reduce such activation.
. Discussion
The results described here confirm previous reports that2S is synthesized in brain primarily by the enzyme CBS
asim
ing 30 (2009) 1523–1534
Abe and Kimura, 1996). They further demonstrate that astro-ytes are the most active producers of H2S, with much smalleruantities being generated by microglia and by the NT-2nd SH-SY5Y neuronal cell lines. The relatively low lev-ls of CBS expression by these other cell types probablyccounts for the fact that only astrocytes can be detectedxpressing CBS by immunohistochemistry in human brain.BS immunostaining of radial glia in developing mouserain (Enokido et al., 2005) and in astrocytes in normalumans and Down syndrome cases (Ichinohe et al., 2005) hasreviously been reported. The immunohistochemical resultsurther show that human vessels use a different system forroducing H2S, namely by CGL. This confirms previouseports of CGL expression by arteries (Hosoki et al., 1997;hao et al., 2001; Webb et al., 2008).
Our steady state levels of H2S in cultured cellsf 0.13–1.5 �mol/g protein, and production levels of.46–15 �mol/g protein/h, are consistent with a previ-us report of H2S synthesis in rat brain homogenates of.38 �mol/g/h (Abe and Kimura, 1996). Taken together, theseesults indicate high production of H2S in brain, with a rela-ively rapid turnover rate. H2S, and the SH ions it producesn solution, are readily diffusible with no specific receptorsaving yet been identified. It can therefore act not only intra-ellularly in those cells where it is produced, but it can alsoct on nearby cells by diffusing into the surround.
We have shown here that H2S production is suppressed bynflammatory stimulation of microglia and astrocytes. Thisuppression reduces the natural antiinflammatory effect of2S. We have also shown that exogenously added NaSH canartially counteract this suppression and can ameliorate theeurotoxicity of supernatants of glial cells exposed to inflam-atory stimulation. This shows that H2S has neuroprotective
s well as antiinflammatory properties (Tables 2 and 3).NaSH reduced by almost half the increase in inflamma-
ory cytokine release caused by inflammatory stimulation oficroglia and astrocytes and it ameliorated the induction ofF�B. This further revealed the antiinflammatory capacityf H2S. Moreover, suppression of endogenous H2S produc-ion through administration of hydroxylamine enhanced theelease of inflammatory mediators and enhanced the releasef neurotoxic substances by both microglia and astrocytes.his is further evidence of the antiinflammatory and neuro-rotective effects of endogenously produced H2S.
These actions might result from both direct and indirectctivity of H2S. H2S is a very powerful reducing agent. Itill therefore act as a direct antioxidant as well as an indirectne by keeping other antioxidants such as glutathione in aeduced state. It will also react with nitrite ions, neutraliz-ng the peroxynitrite produced by inflammatory nitric oxideynthase (iNOS) activity.
The mechanism by which H2S acts to inhibit NF�B
ctivation is unknown but it might relate to its ability totimulate cAMP (Kimura, 2000). Such stimulation has annhibitory effect on transcription of inflammatory cytokineRNAs.

y of Ag
ioedrorenm
Hftctrvrnabiiadtiot
A
AoM
i
R
A
B
B
C
D
E
E
H
H
I
K
K
K
K
K
K
L
L
L
L
L
M
P
Q
M. Lee et al. / Neurobiolog
It is of interest that homozygous inheritance of mutationsn CBS can result in homocystinuria, a rare condition in whichne of the end results is profound mental retardation (Muddt al., 1985). CBS condenses homocystine and serine to pro-uce cystathionine and then cysteine, so that homocystinuriaesults from blockade of the first step. In contrast, CBS isverexpressed in Down syndrome due to replication of theegion on chromosome 21 which encodes the gene (Ichinohet al., 2005). Since many other genes are involved, it can-ot be determined whether this overexpression influences theental retardation.The antiinflammatory and neuroprotective properties of
2S suggest there may be considerable therapeutic potentialor H2S releasing drugs in the treatment of neurodegenera-ive diseases of aging such as AD and PD. These diseases areharacterized by the up-regulation of a spectrum of inflamma-ory mediators. Their damaging effects on neurons might beeduced by supplementary H2S that such drugs might pro-ide. For example, we have recently shown that the H2Seleasing NSAIDS S-aspirin and S-diclofenac attenuate theeuroinflammation induced by activation of microglia andstrocytes (Lee et al., 2009). S-diclofenac had previouslyeen found to protect against ischemia-reperfusion injury insolated rabbit heart (Rossoni et al., 2008) and to reduce LPSnflammatory effects (Li et al., 2007). H2S is a vasorelax-nt related to its production by CGL, so such H2S releasingrugs must take into account its consequent actions on bothhe vascular bed and organ tissue such as brain. Where theres synergism, as might be the case for Alzheimer disease andther neurodegenerative disorders, such agents might proveo be particularly valuable.
cknowledgments
This research was supported by a grant from the Pacificlzheimer Research Foundation. We thank Dr. John Maguiref the UBC Department of Pathology and Laboratoryedicine for assistance in obtaining brain tissue specimens.Disclosure statement: The authors report no conflicts of
nterest.
eferences
be, K., Kimura, H., 1996. The possible role of hydrogen sulfide as anendogenous neuromodulator. J. Neurosci. 16, 1066–1071.
onnefoy, M., Drai, J., Kosika, T., 2002. Antioxidants to delay the processof aging: facts and figures. Presse Medicale 31, 1174–1184.
utterfield, D.A., Perluigi, M., Sultana, R., 2006. Oxidative stress inAlzheimer’s disease brain: new insights from redox proteomics. Eur.J. Pharmacol. 545, 39–50.
alabrese, V., Cornelius, C., Mancuso, C., Pennisi, G., Calafato, S., Bellia,F., Bates, T.E., Stella, A.M.G., Schapira, T., Kostova, A.T.D., Albena, T.,Rizarrelli, E., 2008. Cellular stress response: a novel target for chemo-prevention and nutritional neuroprotection in aging, neurodegenerativedisorders and longevity. Neurochem. Res. 33, 2444–2471.
R
ing 30 (2009) 1523–1534 1533
anielson, K., Andersen, J.K., 2008. Oxidative and nitrative proteinmodifications in Parkinson’s disease. Free Radic. Biol. Med. 44,1787–1794.
nokido, Y., Suzuki, E., Iwasawa, K., Kazuhiko, N., Okazawa, H., Kimura,H., 2005. Cystathionine B-synthase, a key enzyme for homocysteinemetabolism, is preferentially expressed in the radial glia/astrocyte lin-eage of developing mouse CNS. FASEB J. 19, 1854–1856.
to, K., Ogasawara, K., Umemura, Y., Nagai, Y., Kimura, H., 2002. Hydro-gen sulfide is produced in response to neuronal stimulation. J. Neurosci.22, 3386–3391.
ashioka, S., Klegeris, A., Schwab, C., McGeer, P.L., 2008. Interferon—dependent cytotoxic activation of human astrocytes and astrocytomacells. Neurobiol. Aging, doi:10.1016/jneurobiolaging.2008.02.019.
osoki, R., Matsuki, N., Kimura, H., 1997. The possible role of hydrogensulfide as an endogenous smooth muscle relaxant in synergy with nitricoxide. Biochem. Biophys. Res. Commun. 237, 527–531.
chinohe, A., Kanaumi, T., Takashima, S., Enokido, Y., Nagai, Y., Kimura,H., 2005. Cystathionine B-synthase is enriched in the brains of Down’spatients. Biochem. Biophys. Res. Commun. 338, 1547–1550.
amoun, P., 2004. Endogenous production of hydrogen sulfide in mammals.Amino Acids 26, 243–254.
imura, H., 2000. Hydrogen sulfide induces cyclic AMP and modu-lates the NMDA receptor. Biochem. Biophys. Res. Commun. 267,129–133.
imura, H., 2002. Hydrogen sulfide as a neuromodulator. Mol. Neurobiol.26, 13–19.
legeris, A., Giasson, B.I., Zhang, H., Maguire, J., Pelech, S., McGeer, P.L.,2006. Alpha-synuclein and its disease-causing mutants induce ICAM-1 and IL-6 in human astrocytes and astrocytoma cells. FASEB J. 20,2000–2008.
legeris, A., Walker, D.G., McGeer, P.L., 1999. Toxicity of humanTHP-1 monocytic cells towards neuron-like cells is reduced by non-steroidal anti-inflammatory drugs (NSAIDs). Neuropharmacology 38,1017–1025.
legeris, A., Bissonnette, C.J., McGeer, P.L., 2005. Modulation of humanmicroglia and THP-1 cell toxicity by cytokines endogenous to the ner-vous system. Neurobiol. Aging 26, 673–682.
ee, M., Lee, S.J., Choi, H.J., Jung, Y.W., Frokiaer, J., Nielsen, S., Kwon,T.H., 2008. Regulation of AQP4 protein expression in rat brain astro-cytes: role of P2X7 receptor activation. Brain Res. 1195, 1–11.
ee, M., Sparatore, A., Soldato, P., McGeer, E.G., McGeer, P.L.,2009. Hydrogen sulfide-releasing NSAIDs attenuate neuroinflamma-tion induced by microglial and astrocytic activation. Glia, in press,doi:10.1002/glia.20905.
i, L., Bhatia, M., Zhu, Y., Zhu, Y.C., Ramath, R.D., Wang, Z.J.,Anaur, F., Whiteman, M., Salto-Tellez, M., Moore, P.K., 2005. Hydro-gen sulfide is a novel mediator of endotoxic shock. FASEB J. 19,1196–1198.
i, L., Rossoni, G., Sparatore, A., Lee, L.C., Del Soldato, P., Moore,P.K., 2007. Anti-inflammatory and gastrointestinal effects of a noveldiclofenac derivative. Free Radic. Biol. Med. 42, 706–719.
owicka, E., Beltowski, J., 2007. Hydrogen sulfide (H2S)—the third gas ofinterest for pharmacologists. Pharmacol. Rep. 59, 4–24.
udd, S.H., Skovby, F., Levy, H.L., Pettigrew, K.D., Wilcken, B., Pyeritz,R.E., Andria, G., Boers, G.H.J., Bromberg, I.L., Cerone, R., Fowler,B., Grobe, H., Schmidt, H., Schweitzer, L., 1985. The natural historyof homocystinuria due to cystathionine beta-synthase deficiency. Am. J.Hum. Genet. 37, 1–31.
ratico, D., 2008. Evidence of oxidative stress in Alzheimer’s disease brainand antioxidant therapy. Ann. N. Y. Acad. Sci. 1147, 70–78.
u, K., Lee, S.W., Bian, J.S., Low, C.M., Wong, P.T., 2008. Hydro-gen sulfide: neurochemistry and neurobiology. Neurochem. Int. 52,
155–165.ossoni, G., Sparatore, A., Tazzari, V., Manfredi, B., Del Soldato, P., Berti, F.,2008. The hydrogen sulphide-releasing derivative of diclofenac protectsagainst ischaemia reperfusion injury in the isolated rabbit heart. Br. J.Pharmacol. 153, 100–109.

1 y of Ag
S
S
S
S
W
W
Y
534 M. Lee et al. / Neurobiolog
akurai, H., Chiba, H., 1999. IkB kinases phosphorylate NF-kB p65 sub-unit on serine 536 in the transactivation domain. J. Biol. Chem. 274,30353–30356.
ayre, M., Smith, M.A., Perry, G., 2001. Chemistry and biochemistry ofoxidative stress in neurodegenerative disease. Curr. Med. Chem. 8,721–738.
un, B., Zou, X., Yueling, C., Zhang, P., Shi, G., 2008. Precondtioning of car-bon monoxide releasing molecule-derived CO attenuates LPS-inducedactivation of HUVEC. Int. J. Biol. Sci. 4, 270–278.
zabo, C., 2007. Hydrogen sulphide and its therapeutic potential. Nat. Rev.Drug Discov. 6, 917–935.
arenycia, M.W., Goodwin, L.R., Benishin, C.G., Reiffenstein, R.J., Fran-com, D.M., Taylor, J.D., Dieken, F.P., 1989. Acute hydrogen sulfidepoisoning. Demonstration of selective uptake of sulfide by the brain-
Z
ing 30 (2009) 1523–1534
stem by measurement of brain sulfide levels. Biochem. Pharmacol. 38,973–981.
ebb, G.D., Lim, L.H., Oh, V.M.S., Yeo, S.B., Cheong, Y.P., Ali, M.Y.,Oakley, R.E., Lee, C.N., Wong, P.S., Caleb, M.G., Salto-Tellez, M., Bha-tia, M., Chan, E.S.Y., Taylor, E.A., Moore, P.K., 2008. Contractile andvasorelaxant effects of hydrogen sulfide and its biosynthesis in the humanmammary artery. J. Pharmacol. Exp. Ther. 324, 876–882.
ang, G., Wu, L., Jiang, B., Yang, W., Qi, J., Cao, K., Meng, Q., Mustala,A.K., Mu, W., Zhang, S., Snyder, S.H., Wang, R., 2008. H2S as a physi-
ologic vasorelaxant: hypertension in mice with deletion of cystathioninegamma-lyase. Science 322, 587–590.hao, W., Zhang, J., Lu, Y., Wang, R., 2001. The vasorelaxant effect of H(2)Sas a novel endogenous gaseous K(ATP) channel opener. EMBO J. 20,6008–6016.