phenotypic modulation and neuroprotective effects of olfactory
TRANSCRIPT

Phenotypic Modulation and Neuroprotective Effects of OlfactoryEnsheathing Cells: a Promising Tool for Cell Therapy
Rosalia Pellitteri1 & Lidia Cova2 & Damiano Zaccheo3 & Vincenzo Silani2,4 &
Patrizia Bossolasco2
# Springer Science+Business Media New York 2015
Abstract Olfactory Ensheathing Cells (OECs), exhibitingphenotypic characteristics of both astrocytes and SchwannCells, show peculiar plasticity. In vitro, OECs promote axonalgrowth, while in vivo they promote remyelination of damagedaxons. We decided to further investigate OEC potential forregeneration and functional recovery of the damaged CentralNervous System (CNS). To study OEC antigen modulation,OECs prepared from postnatal mouse olfactory bulbs weregrown in different culture conditions: standard or serum-freemedia with/without Growth Factors (GFs) and analyzed fordifferent neural specific markers. OEC functional character-izations were also achieved. Resistance of OECs to the neu-rotoxin 6-hydroxydopamine (6-OHDA) was analyzed byevaluating apoptosis and death. OEC neuroprotective proper-ties were investigated by in vitro co-cultures or by addition ofOEC conditioned medium to the neuroblastoma SH-SY5Ycells exposed to 6-OHDA. We observed: 1) modification ofOEC morphology, reduced cell survival and marker
expression in serum-free medium; 2) GF addition to serum-free medium condition influenced positively survival and re-stored basal marker expression; 3) no OEC apoptosis after aprolonged exposition to 6-OHDA; 4) a clear OEC neuropro-tective tendency, albeit non statistically significant, on 6-OHDA treated SH-SY5Y cells. These peculiar properties ofOECs might render them potential clinical agents able to sup-port injured CNS.
Keywords Olfactory glia . Growth factors .
Immunocytochemistry, flow cytometry . Neuroprotection .
Neurotoxin, cell therapy
Introduction
The olfactory system is a particular area of the Central Ner-vous System (CNS) capable to continuously form new Olfac-tory Receptor Neurons (ORNs) throughout lifetime [1]. TheORNs extend their small unmyelinated axons from the basallamina of the epithelium to the olfactory bulb, crossing theperipheral nervous system-CNS junction [2]. These axonsare protected along their whole length by particular glial cells,so called Olfactory Ensheathing Cells (OECs), which createan adequate environment for their growth. This suggests thatOECs are able to promote the outgrowth of ORN axons driv-ing them towards their correct position in the olfactory bulbs[3].
OECs, first described by Golgi [4] and Blanes [5], sharephenotypic properties with both Schwann Cells (SCs) andastrocytes [6, 7], showing an ability to change in star andfusiform shape [8].
Some peculiar and interesting characteristics of OECsare their plasticity [9] and their ability to secrete severalGrowth Factors (GFs), such as Nerve Growth Factor
Electronic supplementary material The online version of this article(doi:10.1007/s12015-015-9635-3) contains supplementary material,which is available to authorized users.
* Rosalia [email protected]
1 Institute of Neurological Sciences, CNR, Section of Catania,via Paolo Gaifami 18, 95126 Catania, Italy
2 Department of Neurology and Laboratory of Neuroscience, IRCCSIstituto Auxologico Italiano, via Zucchi 18, 20095 CusanoMilanino, Milan, Italy
3 Department of Experimental Medicine, Section of Human Anatomy,University of Genoa, via De Toni 14, 16132 Genoa, Italy
4 Department of Pathophysiology and Transplantation - BDino Ferrari^Center, Università degli Studi di Milano, via Francesco Sforza 35,20122 Milan, Italy
Stem Cell Rev and RepDOI 10.1007/s12015-015-9635-3

(NGF), basic Fibroblast Growth Factor (bFGF), Brain De-rived Neurotrophic Factor (BDNF), Glial Derived Neuro-trophic Factor (GDNF) [10–12], Ciliary NeurotrophicFactor (CNTF) [13], neurotrophins NT4, NT5 andneuregulins [14]. Moreover, OECs express adhesion mol-ecules including laminin, L1, Neuronal Cell AdhesionMolecule (NCAM), fibronectin and collagen IV [15, 16],a n d e x t r a c e l l u l a r ma t r i x mo l e c u l e s , s u ch a smetalloproteinase-2 [17] and amyloid precursor protein[18]. Consequently, OECs have drawn considerable inter-est because they show exceptional ability to promote ax-onal regeneration and functional restoration in the injuredCNS [19–22]. OECs are able to synthesize a myelin con-stituent, the myelin protein zero (P0) [23], that consentthem to promote remyelination of damaged axons [24,25]. In fact, OECs are able to form myelin sheaths arounddemyelinated axons [24], supporting the development ofnodes of Ranvier and the restoration of impulse conduc-tion [26]. The secretion of all these compounds is modu-lated Bon demand^ by the micro-environment, especiallyin the presence of a damage or injured tissue [2, 14, 27].Over the last 15 years, OECs have attracted considerableinterest as a promising approach for repair of spinal cordinjury [28, 29] and axonal growth [30, 31]. Cellular trans-plantations for the treatment of neurodegenerative disor-ders, have been subject of many studies, in fact many celltypes, such as embryonic stem cells, SC [27], mesenchy-mal stem cells (MSCs) [32, 33] and OECs [20] have beenused because able to secrete trophic factors with neuro-protective effects and to promote plasticity in the lesionedarea. In particular, OECs create a favourable regenerativeenvironment secreting many GFs, adhesion molecules in-volved in glia-axon adhesion as well as secreting myelin,thus promoting and guiding axonal growth.
As showed by immunocytochemical studies, OECsalso express several markers, such as Glial FibrillaryAcidic Protein (GFAP), Nerve Growth Factor Receptor(p75), S-100β, oligodendrocyte marker 04, vimentin,nestin [6] and neuropeptide Y [34]. It has been suggestedthat the expression of OEC antigens is variable bothin vivo and in vitro and depends on different factors, suchas the developmental stage, animal species, and cultureconditions [35, 36].
These characteristics have induced to further investigatethe expression of specific OEC markers in different cultureconditions, in particular with and without serum or GF addi-tion. In this study we also evaluated the apoptotic rate of bothOECs and other Neural Stem Cells (NSCs; used as control)exposed to the neurotoxin 6-hydroxydopamine (6-OHDA).Moreover, we investigated the neuroprotective effect of OECson the neuroblastoma cell line SH-SY5Y treated with 6-OHDA in a co-culture system or by addition of OEC condi-tioned medium (OEC-CM).
Materials and Methods
Experiments were performed on P1 mice (FVB strain; Harlan,Italy). Animals were kept in a controlled environment(23 ± 1 °C, 50 ± 5 % humidity) with a 12 h light ⁄dark cyclewith food and water available ad libitum. Experiments werecarried out in compliance with the Italian law on animal careno. 116 ⁄1992 and in accordance with the European Commu-nity Council Directive (86 ⁄ 609 ⁄ EEC). All efforts were madeto minimize animal suffering and to use the fewest number ofanimals possible.
OEC Cultures
As previously described [37], OECs were isolated from P1mouse pup olfactory bulbs (an average of ten pups for eachexperiment). Briefly, pups were decapitated and the bulbs re-moved and dissected out in cold (+4 °C) Leibowitz L-15 me-dium (Sigma). Subsequently, they were digested in MediumEssential Medium-H (MEM-H, Sigma) containing 0.03 %collagenase and 0.25 % trypsin (Sigma). Trypsinization wasstopped by adding Dulbecco’s Modified Eagle’s medium(DMEM, Sigma) supplemented with 10 % Foetal Bovine Se-rum (FBS; Sigma). Cells were resuspended and plated inflasks fed with fresh complete medium DMEM/FBS,2 mM L-glutamine, penicillin (50 U/ml) and streptomycin(50 μg/ml). The antimitotic agent, cytosine arabinoside(10−5M) was added 24 h after initial plating to reduce thenumber of dividing fibroblasts. A further procedure of purifi-cation was adopted following the method by Chuah and Au[38]: OEC cultures were processed to an additional step trans-ferring cells from one flask to a new one. This step reducescontaminating cells, hence, during trypsinization, the OECsdetached before fibroblasts that remain on the plastic. In thelast passage, OECs were plated on 25 cm2 flasks and culturedin DMEM/FBS supplemented with bovine pituitary extract(20 μg/ml). Cells were incubated at 37 °C in DMEM/FBSand fed twice a week. Purity of cell cultures was verified usingimmunocytochemistry: the percentage of S-100/p75 positivecells in our cultures was about 85–90 % (data not shown).
Treatment of Cells
Purified OECs were replated in replicate on 16-mm diameterpoly-L-lysine (10 μg/ml, Sigma) coated glass coverslips at afinal density of 0,3x104 cells/coverslip and grown for 12 h inDMEM/FBS. Thereafter, in half of the replicate the mediumwas changed with FBS-free DMEM. Cultures, grown both inFBS and in FBS-free medium, were cultured with the additionof bFGF (10 ng/ml; Sigma) or GDNF (1 ng/ml; Chemicon) fora week with a change of medium. The optimal concentrationand the time of exposure of the cultures to the GFs wereassessed in our previous studies [37]. Control cultures
Stem Cell Rev and Rep

received medium with no added GFs (CTR). After a week,cells were processed for immunocytochemical procedures.
Immunocytochemistry
Cells were fixed by exposure to 4 % paraformaldehyde in0.1 M Phosphate Borate Solution (PBS) for 30 min. After awash in PBS, the cell membranes were permeabilized with5 % Normal Goat Serum in PBS containing 0.1 % Triton X-100 at room temperature for 30 min. OECs were incubatedovernight with the following antibodies: anti-S-100β (mouse,1:100, Sigma), anti-vimentin (mouse, 1:50, Dako), anti-nestin(rabbit, 1:200, AbCam), anti-GFAP (rabbit, 1:1000, Dako),anti-Myelin (mouse, 1:1000, Chemicon), anti-low-affinitynerve growth factor receptor p75 (rabbit, 1:500, Chemicon),anti-Microtubule Associated Protein-2 (MAP2; mouse,1:1000, Covance, Inc) and anti-Protein Gene Product (PGP9.5; rabbit, 1:600, Serotec). Successively, cells were incubatedin secondary antibodies, Cy3 anti-mouse antibody (1:200;Jackson ImmunoResearch, Laboratories, Inc) and goat anti-rabbit antibody IgG Fluorescein Isothiocyanate (FITC, 1:100;Chemicon) to visualize primary antibodies. The immuno-stained coverslips were analyzed on a Zeiss fluorescence mi-croscope and images were captured with an Axiovision Imag-ing System. A direct cell count allowed the evaluation of OECnumber for each specific marker in different grown condition.No non-specific staining of cells was observed in control in-cubations in which the primary antibodies were omitted.
Cell Growth and Fold Increase
In order to quantify the effect of both FBS and GF addition oncell growth, OECs were plated in the same conditions de-scribed above for immunocytochemical analysis, in 24 wellplates. After 7 days of culture, cells were processed bytrypsinization (0.25 % trypsin for 5 min at 37 °C) and countedby means of Trypan blue dye exclusion. Fold increase wasthen calculated (final absolute number of cells after 7 daysof culture, divided by the starting number of cells). Five dif-ferent experiments in quadruplicate were performed.
MTS Assay
Survival of OECs with/without FBS and GF addition wasevaluated by a 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2 H-tetrazolium(MTS) assay (CellTilter 96® Aqueous Assay, Promega, Mad-ison, WI, USA). OECs were seeded at a concentration of 1,5x104 cells/well in a 48 well plate. The seventh day of culture,freshmedium containingMTS solution was added to the wellsand incubated for 2 h at 37 °C. The supernatants were thencollected, centrifuged at 2000 g for 5 min and read at 490 nmwith a microplate reader (Elx800, Bio-Tek Instruments Inc.,
Winooski, VT, USA). Two different experiments in six repli-cated assays were performed.
Cytometric Analysis
A phenotypic characterization by flow cytometric analysiswas performed on OECs harvested with trypsin treatment.Different cytoplasmic or cell surface neural markers were in-vestigated (Suppl. Table 1). For permeabilization, a commer-cial kit was used (BD Cytofix/Cytoperm™ Fixation/Permeabilization Kit) following manufacturer’s instructions.Briefly, cells were first re-suspended and incubated for 20 minat 4 °C in 250 μl of the Fixation/Permeabilization solution.After two washes in 1 ml of 1× BD Perm/Wash™ buffer,staining was performed using specific primary antibody (2,5 μg/ml) for 30 min at 4 °C in the same buffer, in order tokeep cells permeabilized. This procedure was repeated for thesecondary antibody (Alexa fluor 488; 1:400; JacksonImmunoResearch). The cells were acquired by a flowcytometer (FACSCanto II Becton Dickinson) and analyzedusing Diva software. For each marker, 100,000 events wereacquired. Negative control samples consisted of unlabelledcells (for both primary and secondary antibodies) to evaluateauto-fluorescence.
Apoptosis
To evaluate their susceptibility to 6-OHDA, OECs were incu-bated for different times (6 and 12 h) with the neurotoxin (finalconcentration: 100 μM). Both treated and untreated cells wereharvested with trypsin and apoptosis/death were evaluated byFITC Annexin V combined with propidium iodide (PI; Sig-ma) and analyzed by flow cytometry. NSCs, known to beaffected by 6-OHDA, were used as positive controls [39].
CM and CMstim Preparation
OECs (about 100,000) were seeded and cultured in T25 flasksin DMEM with 10%FBS for 7 days. CM was obtained bycentrifugation of the supernatant in order to remove eventualfloating cells and debris. For CMstim, the fifth day, 6-OHDA(100 μM) was added overnight to the culture and the CMstimharvested as previously described for CM. We previously ob-served in an in vitro study that addition of 6-OHDA to MSCsis able to stimulate the production of GFs and increase theirneuroprotective effect [39].
Neuroprotection
To evaluate neuroprotective effect of OECs, a SH-SY5Y cellline, isolated from the bone marrow of a patient with neuro-blastoma and known to be sensitive to the neurotoxin 6-OHDA [39] was used. The following different conditions
Stem Cell Rev and Rep

were tested: in a first set of experiments, 50,000 cells ofboth OEC and SH-SY5Y were co-cultured in a transwellsystem for 7 days. The sixth day, 6-OHDA was addedovernight to the cultures, and the day after apoptosiswas evaluated by flow cytometry as describe above. In asecond set of experiments, SH-SY5Y were cultured inDMEM with 10%FBS for 7 days as for co-cultures. Thesixth day, contemporaneously to the addition of theneurotoxin, half of the medium was replaced with CMor CMstim. The seventh day, apoptosis was evaluated onSH-SY5Y cells by flow cytometry. The appropriatecontrols (SH-SY5Y cells alone in DMEM with 10%FBSwith and without 6-OHDA addition, OEC cells in allconditions without addition of 6-OHDA) were alsocarried out.
Statistical Analysis
For each culture condition a minimum of 4 experiments inreplicate were carried out, unless otherwise stated. UnpairedStudent t-test was used as statistical analysis for the immuno-characterization by cytofluorimetric analysis.
To evaluate neuroprotective effect of OECs, cell growthand viability after GF addition, statistical analyses were per-formed with GraphPad Instat (GraphPad Software Inc.) usingone way ANOVA test (followed by Bonferroni’s multiplecomparison post hoc test). Values were expressed as themean ± SD. Differences were considered statistically signifi-cant at P < 0.05.
Results
Phenotypic Modifications of OECs Grown in Serum-FreeMedium
OECs from postnatal 1-day (P1) mouse olfactory bulbs weregrown in different conditions: in DMEM/FBS, in FBS-freeDMEM as well as with and without GFs (GDNF and bFGF)addition. For all the conditions, after one week, a characteri-zation was performed both by immunocytochemistry and flowcytometry for the following markers: vimentin, S-100β,nestin, GFAP, Myelin, NCAM, p75, MAP2 and PGP 9.5.
In order to provide a continuous examination of the cellbehaviour, OECs were monitored for a week using phase con-trast microscopy of live cells (Suppl. Figure 1). This allowedthe control of transitional cellular states, rates of cell division,mechanisms ofmovement andmorphological changes. In pre-vious studies we assessed the optimal concentration and thetime of exposure of the cultures to the GFs [37].
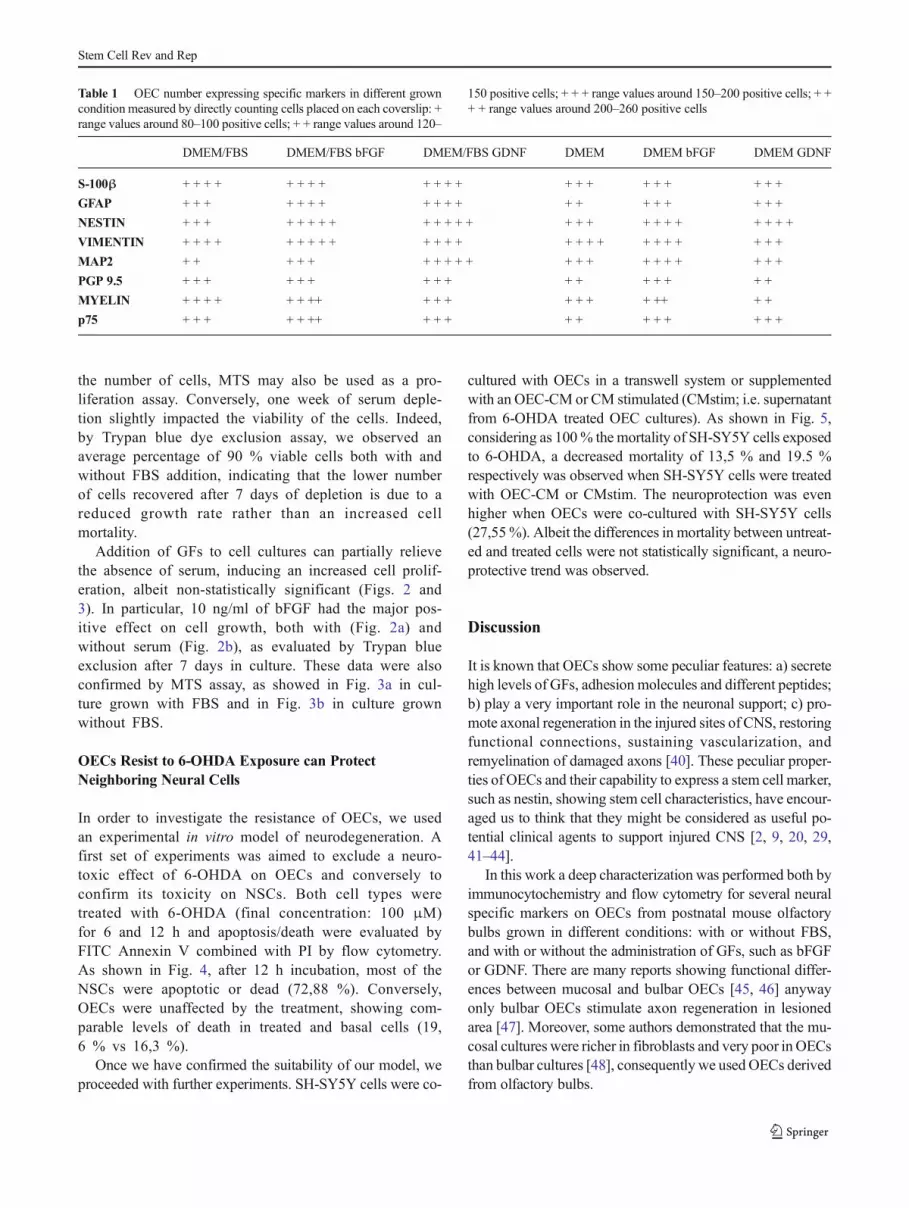
Our results demonstrated that when OECs weregrown in FBS-free medium, a reduced growth rate anda different marker expression were observed, as
summarized in Table 1 and shown by immunocytochem-istry (Fig. 1). The following markers were used: S-100βand p75, both specific markers for OECs, Myelin, PGP9.5 and MAP2 neuronal markers, nestin as specificmarker for stem cells, GFAP and vimentin, markers ofglial differentiation and proliferation respectively. OECnumber expressing specific markers in different growncondition was measured by direct counting, the usedrange was the following: + range values around 80–100, + + range values around 120–150; + + + rangevalues around 150–200; + + + + range values around200–260 (see Table 1). In OECs grown in the absenceof FBS, overall marker expression resulted diminishedby serum withdraw, while the only marker thatremained unchanged was vimentin, that was highlyexpressed in all experimental conditions (Fig. 1).
GFAddition Restores the Standard Biological Propertiesof OECs in Serum-Free Media
When OECs were cultured with the addition of bFGF orGDNF, a positive influence on growth and marker ex-pression was observed (Fig. 1) both in DMEM/FBS andFBS-free DMEM. In fact, in OECs grown in the pres-ence of FBS, both bFGF and GDNF stimulated cellulardivision and marker expression (Table 1), as shown insome representative immunocytochemistry field inFig. 1. In particular, both GFs exerted a positive effecton the GFAP, nestin, Myelin and MAP2 expression,while PGP 9.5 and S-100β were unchanged.
When OECs were grown in FBS-free DMEM, the positiveeffect of GF administration was evident both on cell growthand on OEC antigenic expression, especially for somemarkers, such as GFAP, nestin, PGP 9.5 and MAP2 (Fig. 1and Table 1).
Moreover, we wanted to confirm the immunocyto-chemical results performing flow cytometric analysison OEC cells cultured in DMEM/FBS or in the samemedium without FBS. A decrease in the expression ofthe different neural markers was observed in OEC cul-tures grown without FBS compared with OECs grownin the presence of FBS. Results are expressed as theaverage percentage of positive cells for each marker(Suppl. Table 2 and representative Suppl. Figure 2).
When OECs were grown in FBS-free medium, a con-siderable cell growth reduction was observed (Suppl.Figure 2). Indeed, the mean number of cells obtainedafter 7 days of culture was 167.600 without serum com-pared to 1.024.400 with serum (a fold increase (FI) ofrespectively 83,8 vs 512,2 (Suppl. Figure 3a). Similarly,very different absorbance values (783,5 vs 2125) wereobserved between the two culture conditions (Suppl.Figure 3b). Since absorbance readings are related to
Stem Cell Rev and Rep
the number of cells, MTS may also be used as a pro-liferation assay. Conversely, one week of serum deple-tion slightly impacted the viability of the cells. Indeed,by Trypan blue dye exclusion assay, we observed anaverage percentage of 90 % viable cells both with andwithout FBS addition, indicating that the lower numberof cells recovered after 7 days of depletion is due to areduced growth rate rather than an increased cellmortality.
Addition of GFs to cell cultures can partially relievethe absence of serum, inducing an increased cell prolif-eration, albeit non-statistically significant (Figs. 2 and3). In particular, 10 ng/ml of bFGF had the major pos-itive effect on cell growth, both with (Fig. 2a) andwithout serum (Fig. 2b), as evaluated by Trypan blueexclusion after 7 days in culture. These data were alsoconfirmed by MTS assay, as showed in Fig. 3a in cul-ture grown with FBS and in Fig. 3b in culture grownwithout FBS.
OECs Resist to 6-OHDA Exposure can ProtectNeighboring Neural Cells
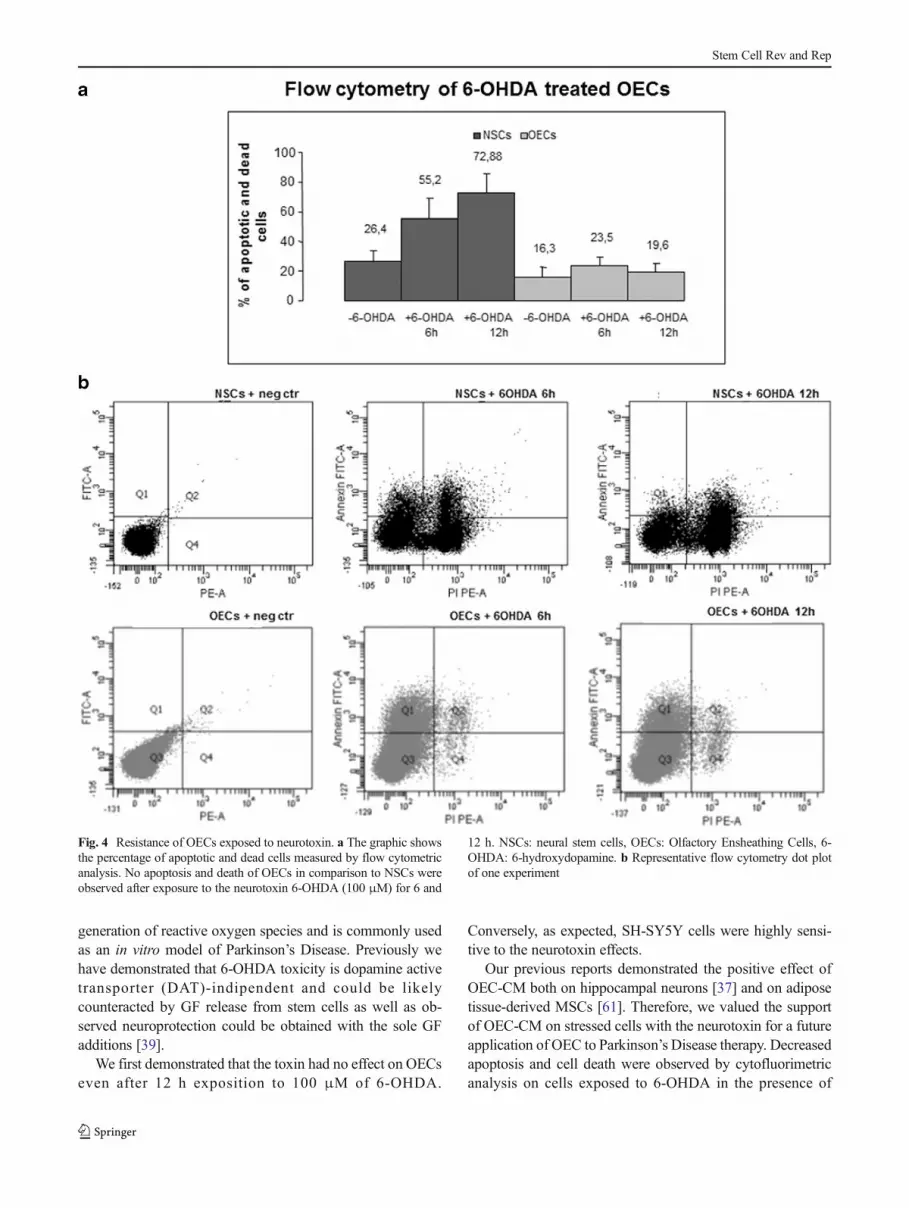
In order to investigate the resistance of OECs, we usedan experimental in vitro model of neurodegeneration. Afirst set of experiments was aimed to exclude a neuro-toxic effect of 6-OHDA on OECs and conversely toconfirm its toxicity on NSCs. Both cell types weretreated with 6-OHDA (final concentration: 100 μM)for 6 and 12 h and apoptosis/death were evaluated byFITC Annexin V combined with PI by flow cytometry.As shown in Fig. 4, after 12 h incubation, most of theNSCs were apoptotic or dead (72,88 %). Conversely,OECs were unaffected by the treatment, showing com-parable levels of death in treated and basal cells (19,6 % vs 16,3 %).
Once we have confirmed the suitability of our model, weproceeded with further experiments. SH-SY5Y cells were co-
cultured with OECs in a transwell system or supplementedwith an OEC-CM or CM stimulated (CMstim; i.e. supernatantfrom 6-OHDA treated OEC cultures). As shown in Fig. 5,considering as 100% the mortality of SH-SY5Y cells exposedto 6-OHDA, a decreased mortality of 13,5 % and 19.5 %respectively was observed when SH-SY5Y cells were treatedwith OEC-CM or CMstim. The neuroprotection was evenhigher when OECs were co-cultured with SH-SY5Y cells(27,55%). Albeit the differences in mortality between untreat-ed and treated cells were not statistically significant, a neuro-protective trend was observed.
Discussion
It is known that OECs show some peculiar features: a) secretehigh levels of GFs, adhesion molecules and different peptides;b) play a very important role in the neuronal support; c) pro-mote axonal regeneration in the injured sites of CNS, restoringfunctional connections, sustaining vascularization, andremyelination of damaged axons [40]. These peculiar proper-ties of OECs and their capability to express a stem cell marker,such as nestin, showing stem cell characteristics, have encour-aged us to think that they might be considered as useful po-tential clinical agents to support injured CNS [2, 9, 20, 29,41–44].
In this work a deep characterization was performed both byimmunocytochemistry and flow cytometry for several neuralspecific markers on OECs from postnatal mouse olfactorybulbs grown in different conditions: with or without FBS,and with or without the administration of GFs, such as bFGFor GDNF. There are many reports showing functional differ-ences between mucosal and bulbar OECs [45, 46] anywayonly bulbar OECs stimulate axon regeneration in lesionedarea [47]. Moreover, some authors demonstrated that the mu-cosal cultures were richer in fibroblasts and very poor in OECsthan bulbar cultures [48], consequently we usedOECs derivedfrom olfactory bulbs.
Table 1 OEC number expressing specific markers in different growncondition measured by directly counting cells placed on each coverslip: +range values around 80–100 positive cells; + + range values around 120–
150 positive cells; + + + range values around 150–200 positive cells; + ++ + range values around 200–260 positive cells
DMEM/FBS DMEM/FBS bFGF DMEM/FBS GDNF DMEM DMEM bFGF DMEM GDNF
S-100β + + + + + + + + + + + + + + + + + + + + +
GFAP + + + + + + + + + + + + + + + + + + +
NESTIN + + + + + + + + + + + + + + + + + + + + + + + +
VIMENTIN + + + + + + + + + + + + + + + + + + + + + + + +
MAP2 + + + + + + + + + + + + + + + + + + + +
PGP 9.5 + + + + + + + + + + + + + + + +
MYELIN + + + + + + ++ + + + + + + + ++ + +
p75 + + + + + ++ + + + + + + + + + + +
Stem Cell Rev and Rep
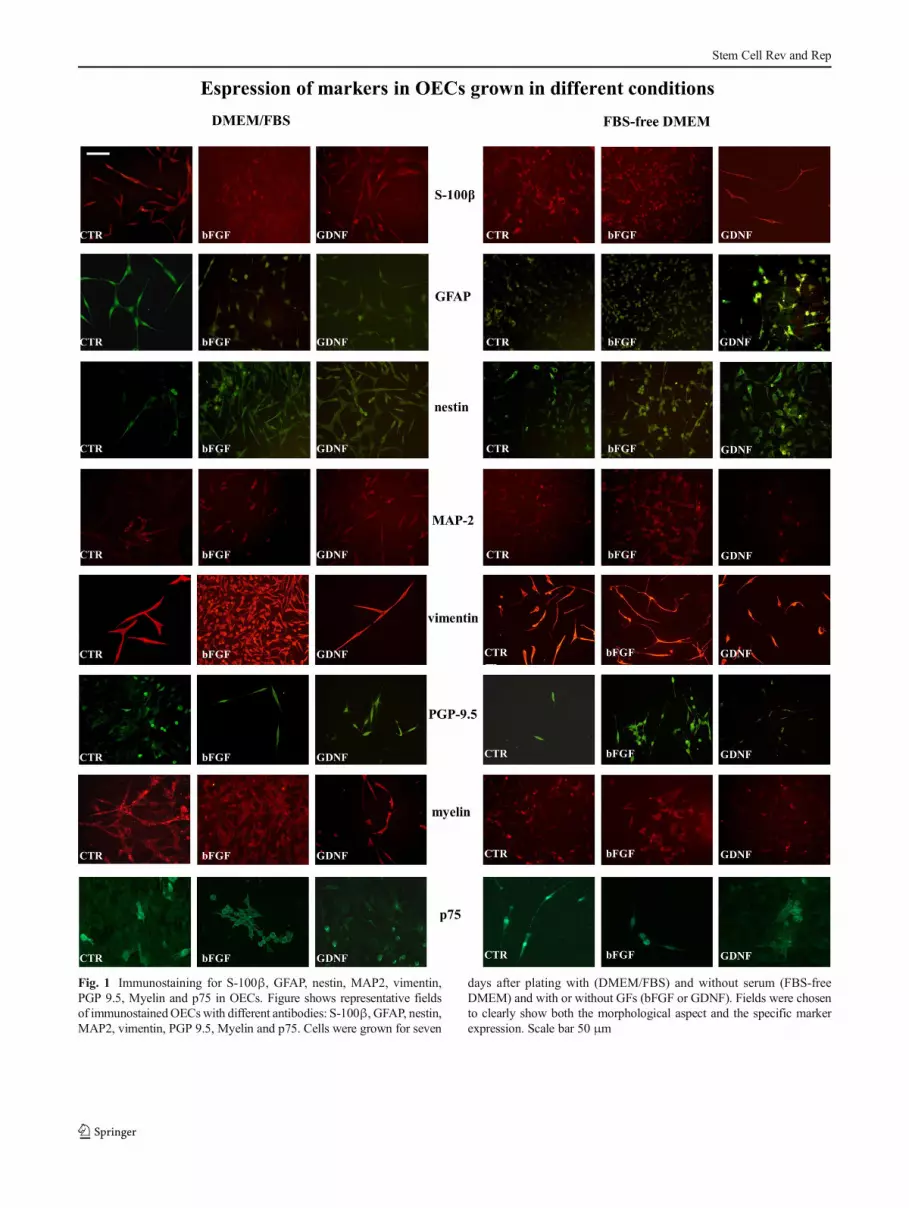
Fig. 1 Immunostaining for S-100β, GFAP, nestin, MAP2, vimentin,PGP 9.5, Myelin and p75 in OECs. Figure shows representative fieldsof immunostainedOECswith different antibodies: S-100β, GFAP, nestin,MAP2, vimentin, PGP 9.5, Myelin and p75. Cells were grown for seven
days after plating with (DMEM/FBS) and without serum (FBS-freeDMEM) and with or without GFs (bFGF or GDNF). Fields were chosento clearly show both the morphological aspect and the specific markerexpression. Scale bar 50 μm
Stem Cell Rev and Rep

Our aim was to analyse the influence of serum-free cultureson OEC phenotype and survival compared to conventionalmedium. This approach was chosen because serum-free culturecondition is mandatory for future therapeutic application. Al-beit the optimal dosage for clinical applications is dependentupon the particular type of human cell and therapy chosen, ahigh number of cells is generally required and therefore in vitrocell expansion step is needed. This may be achieved by addi-tion of bovine or calf sera, which are rich in all the components
required for cell proliferation. However, animal-derived serumstill represents a dangerous health risk. Indeed, it may generateimmune reactions by transplant recipients since cells can retainserum derived components [49]. Although now standardisedGMP bovine serum-containing media for human cell produc-tion could be used, only few suppliers can produce themenough for one blockbuster cell therapy in suitable Internation-al Standards Organization-grade environments [50]. The use ofhuman derived serum is not an alternative strategy as it hasbeen demonstrated that, at least for some cell types, it is lesseffective than its animal counterpart [51], therefore serum-freemedia have to be taken in account.
In this regard, our results highlighted that GFs can partiallysupply the absence of serum stimulating cells growth butcompletely restoring phenotypic features.
Here we reported a reduced expression for most of themarkers analyzed both by immocytochemistry and flow cy-tometry, as well as modifications of the cell morphology inOEC cultures depleted of serum. Different phenotypes, withand without serum, have been reported for several markers inother cell types, probably due to the composition of the serumthat might contain some factors capable of modulating antigenexpression. For example, the expression of CD105 was sig-nificantly lower when serum was omitted from MSCs [52].Similarly, some authors observed a higher expression level ofstem cell markers when different glioblastoma cell line-derived spheres were grown with serum compared to aserum-free medium [53]. We demonstrated that the additionof specific GFs (bFGF or GDNF) was able to increase OECmarker expression, in particular in serum-free condition, re-storing the original phenotype of cells. Modulation of label-ling intensity in serum depleted condition was observed inconjunction with cell growth alterations, which were partiallyovercome by GF addition. Interestingly, our study also showsthat GF addition is able to improve cell growth even in thepresence of serum. This result is in agreement with studieswhich have reported that some GFs are mitogenic for OECsand are in able to exert a positive effect on proliferation anddifferentiation [54]. In addition, we here demonstrate that se-rum deprivation in OECs acts more on cell growth than on cellviability/apoptosis induction.
GFs have also been reported to be potentially major tools intherapeutic treatment of neurological diseases [55, 56]. More-over, some papers already reported clinical trials, in particularfor the repair of neurological damage in human spinal cordinjury, demonstrating the feasibility and safety of transplanta-tion of autologous OECs [42, 57–59]. In agreement with ourresults, a frequent procedure was to remove serum several daysbefore transplantation and to add GFs in the cultures [60].
In this study, we also evaluated the neuroprotective effectof OECs on 6-OHDA treated SH-SY5Y cells in order to ver-ify if these cells may be suitable for cell therapy in neurode-generative diseases. The 6-OHDA neurotoxin induces the
Fig. 2 GF effect on cell growth evaluated by Trypan blue exclusion after7 days in culture with (a) or without (b) FBS and 10 ng/ml bFGF or1 ng/ml GDNF addition in the medium
Fig. 3 GF effect onOECs evaluated byMTS assay after 7 days in culturewith (a) or without (b) FBS and 10 ng/ml bFGF or 1 ng/ml GDNFaddition in the medium
Stem Cell Rev and Rep
generation of reactive oxygen species and is commonly usedas an in vitro model of Parkinson’s Disease. Previously wehave demonstrated that 6-OHDA toxicity is dopamine activetransporter (DAT)-indipendent and could be likelycounteracted by GF release from stem cells as well as ob-served neuroprotection could be obtained with the sole GFadditions [39].
We first demonstrated that the toxin had no effect on OECseven after 12 h exposition to 100 μM of 6-OHDA.
Conversely, as expected, SH-SY5Y cells were highly sensi-tive to the neurotoxin effects.
Our previous reports demonstrated the positive effect ofOEC-CM both on hippocampal neurons [37] and on adiposetissue-derived MSCs [61]. Therefore, we valued the supportof OEC-CM on stressed cells with the neurotoxin for a futureapplication of OEC to Parkinson’s Disease therapy. Decreasedapoptosis and cell death were observed by cytofluorimetricanalysis on cells exposed to 6-OHDA in the presence of
Fig. 4 Resistance of OECs exposed to neurotoxin. a The graphic showsthe percentage of apoptotic and dead cells measured by flow cytometricanalysis. No apoptosis and death of OECs in comparison to NSCs wereobserved after exposure to the neurotoxin 6-OHDA (100 μM) for 6 and
12 h. NSCs: neural stem cells, OECs: Olfactory Ensheathing Cells, 6-OHDA: 6-hydroxydopamine. b Representative flow cytometry dot plotof one experiment
Stem Cell Rev and Rep

OEC-CM, OEC-CMstim or in transwell co-culture withOECs. The neuroprotective effect of CMstim was higher thanunstimulated CM. We previously reported comparable resultsusing MSCs, showing that noxious conditions may influenceGF secretion [39]. The most effective protection was exertedby co-culture of OECs with SH-SY5Y cells. This may be dueto the simultaneous presence of both cell types under effect of6-OHDA and therefore continuous stimulation of the cells bythe toxic environment. OEC neuroprotective effects have alsobeen suggested in vivo by co-transplantation of NSCs andOECs in a 6-OHDA induced rat model, and in vitro but usingrat-derived OEC-CM, albeit CMstim was not investigated[62, 63]. It is known that OECs are a source of severalmolecules, such as GFs, chemokines, matrix metallopro-teinases, complement factors as well as extracellular andmatricellular proteins [64] responsible for their
neuroprotective effects after oxidative damage by en-hanced survival pathways [63].
Finally, we are aware that our findings could not be directlytransferred to human OECs for therapeutic purposes sinceinterspecies differences may play distinct roles in cell behav-ior. Nevertheless, our encouraging results on murine OECssupport further investigation to assess the phenotype andproperties of human OECs in GF added-serum free media.
Conclusions
The present results provide a further supportive evidence thatOECs could represent a suitable tool for cellular therapy indifferent neurological disorders and injured CNS. Furthermore,they could be expanded in vitro without serum for clinical
Fig. 5 Neuroprotective capacity of OECs. a The graphic shows thepercentage of apoptotic and dead SH-SY5Y cells exposed to 6-OHDAand treated with OECs, OEC-CM or CMstim to test OEC neuroprotective
potential. OECs: Olfactory Ensheathing Cells, OEC-CM: OECconditioned medium, CMstim: CM previously stimulated by exposureto 6-OHDA. b Representative flow cytometry dot plot of one experiment
Stem Cell Rev and Rep

purpose by the only addition of GFs. Finally, OECs have thepeculiar and unique characteristic to intermingle with astrocyteexerting a neuroprotective role by reducing astrocytic gliosis[65]. They also show the exceptional ability to promote axonalregeneration and functional restoration in ubiquitous lesionedarea of CNS.
Acknowledgments We would like to thank Dr. Luigi Castoro for hisuseful technical help in the laboratory.
Compliance with Ethical Standards
Conflict of Interest The authors declare that they have no conflict ofinterest.
References
1. Graziadei, P. P. C., &Monti-Graziadei, G. A. (1979). Neurogenesisand neuron regeneration in the olfactory system of mammals. I.Morphological Aspects of Differentiation and StructuralOrganization of the Olfactory Sensory Neurons. Journal ofNeurocytology, 8, 1–18.
2. Fairless, R., & Barnett, S. (2005). Olfactory ensheathing cells: theirrole in central nervous system repair. The International Journal ofBiochemistry and Cell Biology, 37, 693–699.
3. Doucette, R. (1990). Glial influences on axonal growth in the pri-mary olfactory system. Glia, 3, 433–449.
4. Golgi, C. (1875). Sulla fina anatomia dei bulbi olfattorii. RivistaSperimentale di Freniatria, 1, 403–425.
5. Blanes, T. (1898). Sobre algunos puntos dudosos de la estructuradel bulbo olfatorio. Revista Trimestral Micrografica, 3, 99–127.
6. Ramon-Cueto, A., & Avila, J. (1998). Olfactory ensheathing cells:properties and function. Brain Research Bulletin, 46, 175–187.
7. Wewetzer, K., Verdù, E., Angelov, D. N., & Navarro, X. (2002).Olfactory ensheathing glia and Schwann cells: two of a kind?Cellular Tissue Research, 309, 337–345.
8. Van den Pol, A. N., & Santarelli, J. G. (2003). Olfactoryensheathing cells: time lapse imaging of cellular interactions, axo-nal support, rapid morphologic shifts, and mitosis. Journal ofComparative Neurology, 458, 175–194.
9. Yang, H., He, B. R., & Hao, D. J. (2014). Biological roles of olfac-tory ensheathing cells in facilitating neural regeneration: a system-atic review. Molecular Neurobiology, 51(1), 168–179.
10. Lipson, A. C.,Widenfalk, J., Lindqvist, E., Ebendal, T., &Olson, L.(2003). Neurotrophic properties of olfactory ensheathing glia.Experimental Neurology, 180, 167–171.
11. Mackay-Sim, A., & Chuah, M. I. (2000). Neurotrophic factors in theprimary olfactory pathway. Progress in Neurobiology, 62, 527–559.
12. Woodhall, E., West, A. K., & Chuah, M. I. (2001). Cultured olfac-tory ensheathing cells express nerve growth factor, brain-derivedneurotrophic factor, glia cell line-derived neurotrophic factor andtheir receptors. Molecular Brain Research, 88, 203–213.
13. Wewetzer, K., Grothe, C., & Claus, P. (2001). In vitro expressionand regulation of ciliary neurotrophic factor and its α receptor sub-unit in neonatal rat olfactory ensheathing cells. NeuroscienceLetters, 306, 165–168.
14. Boruch, A. V., Conners, J. J., Pipitone, M., Deadwyler, G., Storer, P.D., Devries, et al. (2001). Neurotrophic and migratory properties ofan olfactory ensheathing cell line. Glia, 33, 225–229.
15. Doucette, R. (1996). Immunohistochemical localization of laminin,fibronectin and collagen type IV in the nerve fiber layer of the
olfactory bulb. International Journal of DevelopmentalNeuroscience, 14, 945–959.
16. Franceschini, I. A., & Barnett, S. C. (1996). Low-affinity NGF-receptor and E-N-CAM expression define two types of olfactorynerve ensheathing cells that share a common lineage.Developmental Biology, 173, 327–343.
17. Pastrana, E., Moreno-Flores, M. T., Gurzov, E. N., Ávila, J.,Wandosell, F., & Díaz-Nido, J. (2006). Genes associated with adultaxon regeneration promoted by olfactory ensheathing cells: a newrole for matrix metalloproteinase 2. Journal of Neuroscience, 26,5347–5359.
18. Moreno-Flores, M. T., Lim, F., Martín-Bermejo, M. J., Díaz-Nido,J., Ávila, J., & Wandosell, F. (2003). High level of amyloid precur-sor protein expression in neurite-promoting olfactory ensheathingglia (OEG) and OEG-derived cell lines. Journal of NeuroscienceResearch, 71, 871–881.
19. Barnett, S. C., & Riddell, J. S. (2004). Olfactory ensheathing cells(OECs) and the treatment of CNS injury: advantages and possiblecaveats. Journal of Anatomy, 204, 57–67.
20. Franssen, E. H., de Bree, F. M., & Verhaagen, J. (2007). Olfactoryensheathing glia: their contribution to primary olfactory nervoussystem regeneration and their regenerative potential followingtransplantation into the injured spinal cord. Brian Research, 56,236–258.
21. Raisman, G. (2001). Olfactory ensheathing cells – another miracle curefor spinal cord injury? Nature Reviews Neuroscience, 2, 369–375.
22. Ramon-Cueto, A., Corsero, M. I., Santos-Benito, F. F., & Avila, J.(2000). Functional recovery of paraplegic rats and motor axon re-generation in their spinal cords by olfactory ensheathing cells.Neuron, 25, 425–435.
23. Lee, M. J., Calle, E., Brennan, A., Ahmed, S., Sviderskaya, E.,Jessen, K. R., et al. (2001). In early development of the rat mRNAfor themajor myelin protein P(0) is expressed in non-sensory areas ofthe embryonic inner ear, notochord, enteric nervous system, andolfactory ensheathing cells. Developmental Dynamics, 222, 40–51.
24. Franklin, R. J. M. (2003). Remyelination by transplanted olfactoryensheathing cells. The Anatomical Record, 271B, 71–76.
25. Sasaki, M., Lankford, K. L., Radtke, C., Honmou, O., &Kocsis, J. D. (2011). Remyelination after olfactoryensheathing cell transplantation into diverse demyelinatingenvironments. Experimental Neurology, 229(1), 88–98.
26. Sasaki, M., Hains, B., Lankford, K. L.,Waxman, S. G., & Kocsis, J.D. (2006). Protection of corticospinal tract neurons after dorsalspinal cord transaction and engraftment of olfactory ensheathingcells. Glia, 53, 352–359.
27. Oudega, M., & Xu, X. M. (2006). Schwann cell transplantation forrepair of the adult spinal cord. Journal of Neurotrauma, 23, 453–467.
28. Lindsay, S. L., Riddell, J. S., & Barnett, S. C. (2010). Olfactorymucosa for transplant mediated repair: a complex tissue for a com-plex injury? Glia, 58, 125–134.
29. Raisman, G., & Li, Y. (2007). Repair of neural pathways by olfac-tory ensheathing cells. Nature Reviews Neuroscience, 8, 312–319.
30. Chehrehasa, F., Windus, L. C., Ekberg, J. A., Scott, S. E., Amaya,D., Mackay-Sim, A., et al. (2010). Olfactory glia enhance neonatalaxon regeneration. Molecular and Cellular Neuroscience, 45(3),277–288.
31. Su, Z., & He, C. (2010). Olfactory ensheathing cells: biology inneural development and regeneration. Progress in Neurobiology,92(4), 517–532.
32. Cova, L., Armentero, M. T., Zennaro, E., Calzarossa, C.,Bossolasco, P., Busca G., et al. (2010). Multiple neurogenic andneurorescue effects of human mesenchymal stem cell after trans-plantation in an experimental model of Parkinson’s disease. BrainResearch, 1311, 12–27.
Stem Cell Rev and Rep

33. Blandini, F., Cova, L., Armentero,M. T., Zennaro, E., Levandis, G.,Bossolasco, P., et al. (2010). Transplantation of undifferentiatedhuman mesenchymal s tem cel ls protec ts agains t 6-hydroxydopamine neurotoxicity in the rat. Cell Transplantation,19(2), 203–217.
34. Ubink, R., & Hökfelt, T. (2000). Expression of neuropeptide Y inolfactory ensheathing cells during prenatal development. Journal ofComparative Neurology, 423, 13–25.
35. Ebel, C., Brandes, G., Radtke, C., Rohn, K., & Wewetzer,K. (2013). Clonal in vitro analysis of neurotrophin receptorp75-immunofluorescent cells reveals phenotypic plasticity ofprimary rat olfactory ensheathing cells. NeurochemistryResearch, 38(5), 1078–1087.
36. Vincent, A. J., West, A. K., & Chuah, M. I. (2005). Morphologicaland functional plasticity of olfactory ensheathing cells. Journal ofNeurocytology, 34, 65–80.
37. Pellitteri, R., Spatuzza, M., Russo, A., Zaccheo, D., & Stanzani, S.(2009). Olfactory ensheathing cells represent an optimal substratefor hippocampal neurons: an in vitro study. International Journal ofDevelopmental Neuroscience, 27(5), 453–458.
38. Chuah, M. I., & Au, C. (1993). Cultures of ensheathing cells fromneonatal rat olfactory bulbs. Brain Research, 601(1–2), 213–220.
39. Cova, L., Bossolasco, P., Armentero, M. T., Diana, V., Zennaro, E.,Mellone, M., et al. (2012). Neuroprotective effects of human mes-enchymal stem cells on neural cultures exposed to 6-hydroxydopamine: implications for reparative therapy inParkinson’s disease. Apoptosis, 17(3), 289–304.
40. Pellitteri, R., Spatuzza, M., Stanzani, S., & Zaccheo, D. (2010).Biomarkers expression in rat olfactory ensheathing cells.Frontiers in Bioscience, 2, 289–298.
41. Mackay-Sim, A., & St. John, J. A. (2010). Olfactory ensheathingcells from the nose: clinical application in human spinal cord inju-ries. Experimental Neurology, 229, 174–180.
42. Rao, Y., Zhu, W., Liu, H., Jia, C., Zhao, Q., & Wang, Y. (2013).Clinical application of olfactory ensheathing cells in the treatmentof spinal cord injury. Journal of International Medical Research,41(2), 473–481.
43. Sethi, R., Sethi, R., Redmond, A., & Lavik, E. (2014). Olfactoryensheathing cells promote differentiation of neural stem cells and ro-bust neurite extension. StemCell Reviews and Report, 10(6), 772–785.
44. Siddiqui, A. M., Khazaei, M., & Fehlings, M. G. (2015).Translating mechanisms of neuroprotection, regeneration, and re-pair to treatment of spinal cord injury. Progress in Brain Research,218, 15–54.
45. Guérout, N., Derambure, C., Drouot, L., Bon-Mardion, N., Duclos,C., Boyer, O., et al. (2010). Comparative gene expression profilingof olfactory ensheathing cells from olfactory bulb and olfactorymucosa. Glia, 58(13), 1570–1580.
46. Kueh, J. L., Raisman, G., Li, Y., Stevens, R., & Li, D.(2011). Comparison of bulbar and mucosal olfactoryensheathing cells using FACS and simultaneous antigenicbivariate cell cycle analysis. Glia, 59(11), 1658–1671.
47. Tabakow, P., Raisman, G., Fortuna, W., Czyz, M., Huberm, J., Li,D., et al. (2014). Functional regeneration of supraspinal connectionsin a patient with transected spinal cord following transplantation ofbulbar olfactory ensheathing cells with peripheral nerve bridging.Cell Transplantation, 23(12), 1631–1655.
48. Yamamoto, M., Raisman, G., Li, D., & Li, Y. (2009).Transplanted olfactory mucosal cells restore paw reachingfunction without regeneration of severed corticospinal tractfibres across the lesion. Brain Research, 1303, 26–31.
49. Spees, J. L., Gregory, C. A., Singh, H., Tucker, H. A., Peister, A.,Lynch, et al. (2004). Internalized antigens must be removed to pre-pare hypoimmunogenic mesenchymal stem cells for cell and genetherapy. Molecular Therapy, 9, 747–756.
50. Brindley, D. A., Davie, N. L., Culme-Seymour, E. J.,Mason, C., Smith, D. W., & Rowley, J. A. (2012). Peakserum: implications of serum supply for cell therapymanufacturing. Regenerative Medicine, 7(1), 7–13.
51. Kuznetsov, S. A., Mankani, M. H., & Robey, P. G. (2000). Effect ofserum on human bone marrow stromal cells: ex vivo expansion andin vivo bone formation. Transplantation, 70, 1780–1787.
52. Mark, P., Kleinsorge, M., Gaebel, R., Lux, C. A., Toelk, A.,Pittermann, E., et al. (2013). Human mesenchymal stem cells dis-play reduced expression of CD105 after culture in serum-free me-dium. Stem Cells International, 2013, 698076.
53. Hong, X., Chedid, K., & Kalkanis, S. N. (2012). Glioblastoma cellline-derived spheres in serum containingmedium versus serum-freemedium: a comparison of cancer stem cell properties. InternationalJournal of Oncology, 41(5), 1693–1700.
54. Alexander, C. L., Fitzgerald, U. F., & Barnett, S. C. (2002).Identification of growth factors that promote long-term prolifera-tion of olfactory ensheathing cells and modulate their antigenicphenotype. Glia, 37, 349–364.
55. Sharma, H. S. (2007). Neurotrophic factors in combination: a pos-sible new therapeutic strategy to influence pathophysiology of spi-nal cord injury and repair mechanisms. Current PharmaceuticalDesign, 13(18), 1841–1874.
56. Duan, D., & Lu, M. (2015). Olfactory mucosa: a rich source of celltherapy for central nervous system repair. Reviews in theNeurosciences, 26(3), 281–293.
57. Féron, F., Perry, C., Cochrane, J., Licina, P., Nowitzke, A.,Urquhart, S., et al. (2005). Autologous olfactory ensheathing celltransplantation in human spinal cord injury. Brain, 128, 2951–2960.
58. Huang, H., Chen, L., Wang, H., Xiu, B., Li, B., Wang, R.,et al. (2003). Influence of patients' age on functional recov-ery after transplantation of olfactory ensheathing cells intoinjured spinal cord injury. Chinese Medical Journal, 116(10),1488–1491.
59. Wu, J., Sun, T., Ye, C., Yao, J., Zhu, B., & He, H. (2012).Clinical observation of fetal olfactory ensheathing glia trans-plantation (OEGT) in patients with complete chronic spinalcord injury. Cell Transplantation, 21(Suppl 1), S33–S37.
60. Rosner, J., Avalos, P., Acosta, F., Liu, J., & Drazin, D. (2012). Thepotential for cellular therapy combined with growth factors in spinalcord injury. Stem Cells International, 2012, 826754.
61. Lo Furno, D., Pellitteri, R., Graziano, A., Giuffrida, R., Vancheri,C., Gili, E., et al. (2013). Differentiation of human adipose stemcells into neural phenotype by neuroblastoma-or olfactoryensheathing cells-conditioned medium. Journal of CellularPhysiology, 228, 2109–2118.
62. Shukla, S., Chaturvedi, R. K., Seth, K., Roy, N. S., & Agrawal, A.K. (2009). Enhanced survival and function of neural stem cells-derived dopaminergic neurons under influence of olfactoryensheathing cells in parkinsonian rats. Journal of Neurochemistry,109, 436–451.
63. Shukla, A., Mohapatra, T. M., Parmar, D., & Seth, K.(2014). Neuroprotective potentials of neurotrophin rich ol-factory ensheathing cells conditioned media against6OHDA-induced oxidative damage. Free Radical Research,48(5), 560–571.
64. Roet, K. C., Bossers, K., Franssen, E. H., Ruitenberg, M. J., &Verhaagen, J. (2011). A meta-analysis of microarray-based geneexpression studies of olfactory bulb-derived olfactory ensheathingcells. Experimental Neurology, 229(1), 10–45.
65. Lakatos, A., Franklin, R. J. M., & Barnett, S. C. (2000).Olfactory ensheating cells and Schwann cells differ in theirin vitro interactions with astrocytes. Glia, 32, 214–225.
Stem Cell Rev and Rep