morphology of the human olfactory epithelium
TRANSCRIPT

THE JOURNAL OF COMPARATIVE NEUROLOGY 297:l-13 (1990)
Morphology of the Human Olfactory Epithelium
EDWARD E. MORRISON AND RICHARD M. COSTANZO Department of Physiology, Medical College of Virginia, Virginia Commonwealth University,
Richmond, Virginia 23298-0551
ABSTRACT The human olfactory epithelium has been previously studied with scanning electron
microscopy; however, most studies have been limited to examining the epithelial surface. In an attempt to examine structures below the surface, we scanned epithelial fractures that occurred during tissue preparation. This made it possible to obtain unique three-dimensional images of cell profiles from the mucosal surface through the full depth of the epithelium. We examined supporting cells, olfactory neurons, basal cells, and a fourth cell type, the microvillar cell. Supporting cells had a microvillar surface and were in close contact with olfactory neurons and their processes. Olfactory neurons were primarily located in the middle and lower epithelial regions. Basal cells occurred alone or in clusters adjacent to the basal lamina. Microvillar cells were always observed in the upper epithelial region. They were flask- or pear-shaped, had a tuft of microvilli that extended into the nasal cavity, and a thin axon-like process that passed basally towards the lamina propria.
This study represents the first comprehensive scanning electron microscopy examination of the human olfactory epithelium. Three-dimensional images obtained for each epithelial cell type allowed us to examine cell processes and their close contacts, especially between supporting cells and olfactory neurons. These results also revealed the irregular and patchy distribution of olfactory receptors within the human nasal cavity. Further studies that examine the detailed morphology of the human olfactory epithelium should provide a better understanding of the physiological mechanism and clinical disorders that affect olfactory function in humans.
Key words: electron microscopy, nervous system, respiratory system, olfactory mucosa, nose
The olfactory epithelium is located in the superior region of the nasal cavity and covers a portion of the septum and superior turbinates. This sensory epithelium plays an impor- tant role in the detection and processing of chemical odor- ants in the inspired air.
The olfactory epithelium of a number of vertebrate species, including man, has been studied by the use of different morphological approaches. Light and transmission electron microscopy studies have shown that the human olfactory epithelium has morphological characteristics simi- lar to those of other vertebrates (Engstrom and Bloom, '53; Naessen, '71; Moran et al., '82a; Pyatkina, '82; Nakashima et al., '84). These studies have shown that the olfactory epithelium of most vertebrates is pseudostratified and consists of supporting cells, olfactory receptors, and basal cells. Recently, a fourth cell type, the microvillar cell, has been shown to be present in the human olfactory epithelium (Moran et al., '82a,b). Although this cell has been observed with both light and transmission electron microscopy in the human olfactory epithelium, its function is unknown.
The scanning electron microscope (SEM) has also been used to study olfactory epithelium of several vertebrates (Graziadei, '75; Graziadei and Monti-Graziadei, '76; Menco, '77; Wang and Halpern, '80; Costanzo and Morrison, '89). Scanning microscopy provides an excellent method for obtaining detailed observations of cell surface structures. In a recent SEM study of the hamster, we used the SEM to obtain detailed observations not only of surface structures, but also the full depth of the epithelium (Costanzo and Morrison, '89; Morrison and Costanzo, '89). With this approach we were able to examine the three-dimensional cell morphology and demonstrate cell contacts that exist between supporting cells and olfactory neurons.
The SEM has also been used to study the human olfactory epithelium (Mygrind, '75; Lenz, '77; Ohno et al., '81; Larsen and Tos, '82; Yamada, '83). Most of these studies, however, concentrated on examining the surface of the epithelium.
Accepted February 12,1990.
o 1990 WILEY-LISS, INC.

2
The purpose of this study was to obtain detailed three- dimensional observations of the human olfactory epithe- lium. Our results represent the first detailed SEM study of the complete human olfactory epithelium. The method of SEM examination used allowed us to view the mucosal surface morphology and the entire depth of the human olfactory epithelium. The results indicate that there is a close structural relationship between supporting cells and olfactory neurons throughout the full extent of the epithe- lium. In addition, the results demonstrate that there is a patchy and irregular distribution of olfactory receptors in the human nasal cavity.
E.E. MORRISON AND R.M. COSTANZO
MATERIALS AND METHODS Olfactory tissue was obtained at autopsy from five sub-
jects (Table I). The skull was opened and the brain re- moved. A block of tissue was obtained by cutting a rectangu- lar piece of bone through the anterior cranial fossa and removing the cribriform plate, nasal septum, and superior and middle turbinates (Fig. 1). The tissue was placed in cold fixative (0.6 %) paraformaldehyde, 2 % glutaraldehyde in sodium cacodylate buffer, pH 7.4) for 1-2 days, then rinsed in buffer and postfixed in 2% OsO, for 2-3 hours. After a buffer rinse, the tissue was dehydrated through graded alcohols then transferred to Freon TF. Samples were critical point dried (Ladd Research Laboratories) with several wash and rinse cycles with carbon dioxide, mounted, and sputter coated for 5-10 minutes at 5 PA. Specimens were examined with a JEOL 810 scanning electron microscope operated a t 5-15 kV.
Small samples of tissue were also processed for light and transmission electron microscopy. Following postfixation osmication (2 hours), olfactory tissue was alcohol dehy- drated and embedded in araldyte epoxy resin. Thick sec- tions for light microscopic examination were stained with toluidine blue. Ultrathin sections were cut on a LKB ultramicrotome, stained with uranyl acetate and lead cit- rate, and photographed with a Zeiss 18-EM operated at 60-80 kV.
RESULTS Our method of tissue preparation produced numerous
epithelial fractures that allowed us to examine the mucosal surface and the full depth of the epithelium (Figs. 2,3). This method offered a three-dimensional view of cell morphology and structural organization of the human olfactory epithe- lium.
Transition region The transition from respiratory to olfactory epithelium
occurred in the superior regions of the nasal cavity. The transition regions were not uniform or well defined. Scan- ning the mucosal surface at low magnification revealed regions of light and dark contrast that formed an irregular boundary between olfactory and non-olfactory tissues (Figs. 4-6). We also observed several regions where patches of respiratory epithelium were found distributed in the olfac- tory area. Distinction between the regions was made by surface morphology. In respiratory regions, cilia were paral- lel and unbranched as they extended into the nasal cavity (Fig. 5). In olfactory regions they were Ionger and formed a dense blanket over the epithelial surface. There were several
types of surface transition regions. Respiratory cells had irregular shapes and short microvilli where nasal gland openings were visible (Fig. 6). Adjacent olfactory areas had long and short sensory cilia. In other regions where the transition zone was irregular, the two epithelial tissues were mixed. There were clusters or individual olfactory receptor neurons among short microvilli non-olfactory cells (Figs. 7 , 8). We observed olfactory dendritic knobs with variable numbers of cilia of different lengths. Cilia arose from a necklace-like swelling on the dendritic knob in these re- gions. Some knobs had many short cilia (Fig. 91, while others had long tapering processes that extended over the epithe- lial surface (Fig. 10).
The olfactory epithelium rests upon a thick connective tissue lamina propria containing Bowman’s glands, olfac- tory axon bundles and blood vessels. Blood vessels were observed just below the epithelium and no capillaries were seen within the olfactory epithelium (Fig. 11). Human olfactory epithelium is pseudostratified, consisting of sup- porting cells, olfactory neurons and basal cells. A fourth cell type, the microvillar cell, was occasionally observed (Fig. 12). A description of the epithelial cell types follows.
Supporting cells Supporting cells were tall and columnar, spanning the
depth of the epithelium (Fig. 13). Their upper cellular portion was broad with a microvillar mucosal surface. Their microvilli varied from short and stubby, to long processes (branched and unbranched) extending into the nasal cavity. Fractures often exposed the internal structure of the epithe- lium, revealing that supporting cells have rough surfaces and tapering basal processes. In the lower epithelium sup- porting cells surround and enclose portions of olfactory neuron cell bodies and their processes. Fine processes extend from supporting cells and appear to contact adjacent cells. Some supporting cells taper to a thin basal process that terminates in the basal lamina region.
TABLE 1. Sources of Human Olfactory Epithelium
Hours C a w of death Sex postmortem Age
7 weeks Male a12 Sudden death 32 years Male 12 Heart failure
Liver failure 44 years Male 7 53 years Female
4 Heartfailure Amyotropic lateral sclerosis 67 years Female 3-6
Fig. 1. a: Dorsal view of the anterior cranial fossa outlining the block of tissue removed at autopsy (dotted line). Olfactory nerve fibers enter the cranium through small openings (arrows) in the cribriform plate of the ethmoid bone. b: Ventral view of tissue block removed a t autopsy, looking into the roof of the nasal cavity. Septum (S), superior nasal meatus (*), and lateral wall of nasal cavity (L).
Fig. 2. Low-power scanning micrograph of the surface of the nasal cavity. Note the numerous surface fractures (arrows) that expose the underlying epithelial morphology. x 25.
Fig. 3. Higher magnification of fractured region illustrating the three-dimensional images, that are observed following tissue prepara- tion. The mucosal (MI surface and the full depth of the epithelium (E) are exposed. x 280.
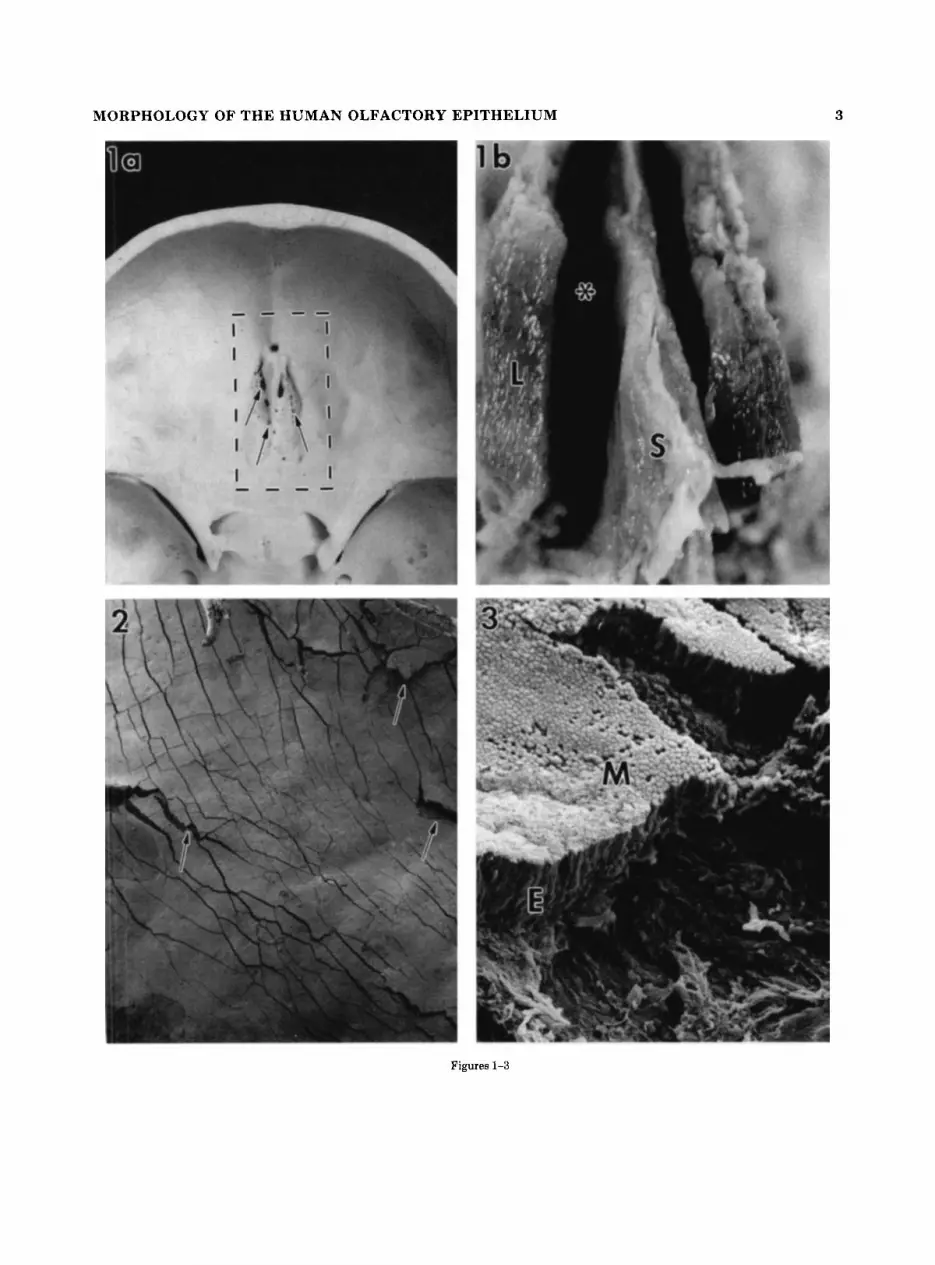
MORPHOLOGY OF THE HUMAN OLFACTORY EPITHELIUM 3
Figures 1-3
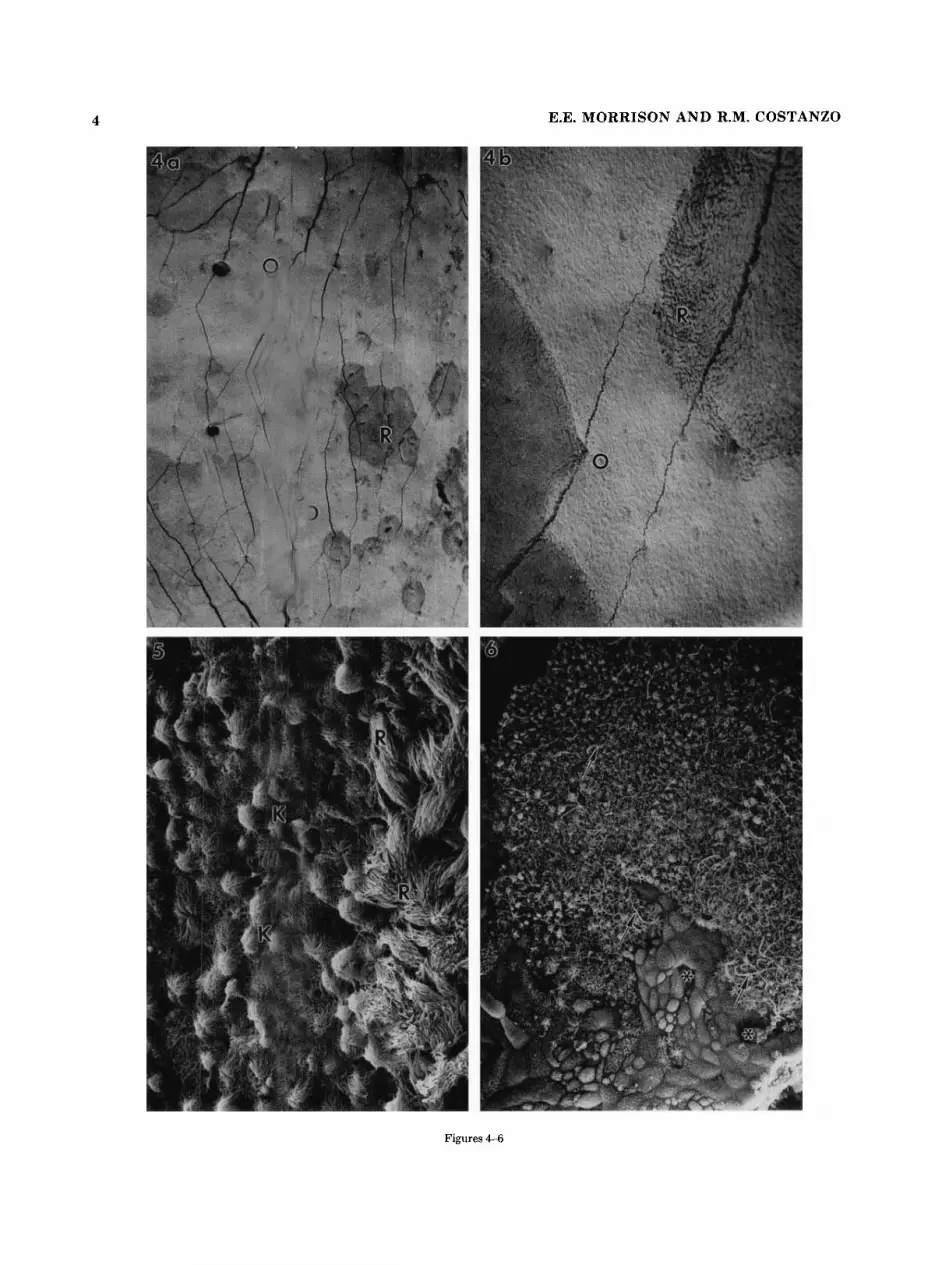
4 E.E. MORRISON AND R.M. COSTANZO
Figures 4-6

MORPHOLOGY OF THE HUMAN OLFACTORY EPITHELIUM 5
Olfactory neurons Human olfactory neurons are bipolar cells with cell bodies
generally located in the middle or lower half of the epithe- lium. Cell bodies were round or oval, 4-6 pm in diameter, with a single unbranched dendrite originating from the apical region. The dendrite was straight or curved, passing along or surrounded by neighboring supporting cells. There were often fine processes observed between dendrites and supporting cells (Fig. 14).
Cell bodies of olfactory neurons were also observed close to the mucosal surface. These neurons were also typically bipolar, but had short, thick dendrites. Olfactory dendrites terminated in ciliated knob-like swellings at the epithelial surface (Fig. 15). Cilia extend from the dendritic knob over the epithelial surface and, in some regions, form a character- istic dense irregular blanket of cilia that obscures the underlying epithelium (Fig. 16).
Olfactory dendritic knobs are spherical or elongated (1-2 pm in diameter), located just above or at the epithelial surface (Fig. 17). They contain typical cytoplasmic or- ganelles, most prominently mitochondria and cilia bearing basal bodies. Dendritic knobs have variable numbers of cilia. These extend from the knob, are thick proximally (0.2-0.3 pm) and taper (0.1 pm) as they extended over the epithelial surface. Olfactory cilia are of variable lengths, from short processes (1-5 Fm) to some that extended for distances greater than 30 pm long.
Dendrites extend to the epithelial surface in a straight or winding manner. Just below the knob, a belt-like junctional complex connects adjacent supporting cells and neurons (Fig. 18). Some dendrites also directly contact or are par- tially enclosed by structures from neighboring supporting cells a t all levels of the epithelium.
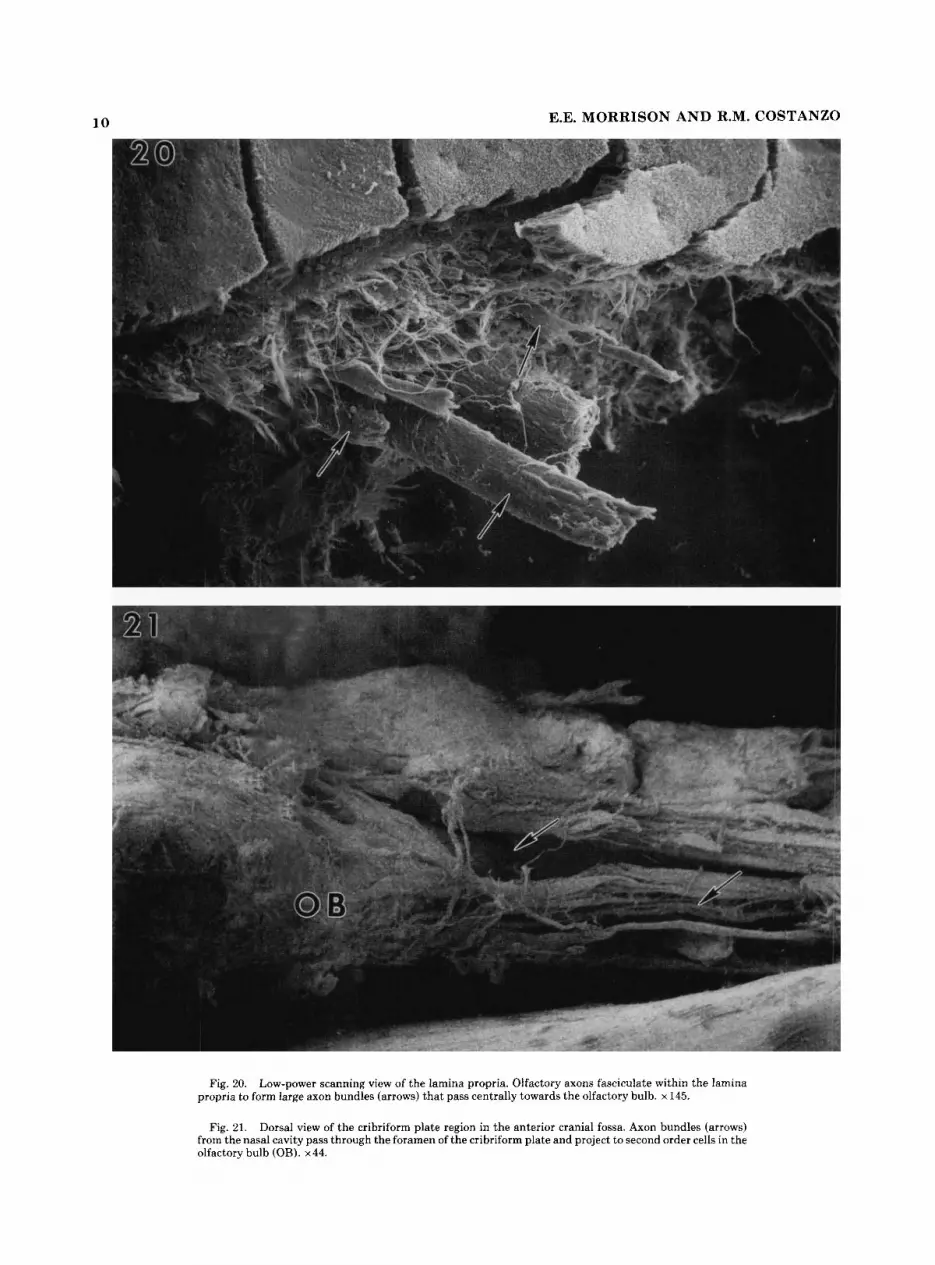
A single axon originates from the basal aspect of olfactory neuron cell bodies. They are 0.1-0.4 pm in diameter, un- branched, and extend to the basal lamina between adjacent supporting cells (Fig. 19). Olfactory axons form small intraep- ithelial fascicles, exit the basal epithelium region, and enter the underlying lamina propria. Within the lamina propria they form larger (20-40 pm) fascicles (fila olfactoria) that pass centrally towards the olfactory bulb (Fig. 20). They then pass through the foramina of the cribriform plate, project to and synapse with second order neurons in the olfactory bulb (Fig. 21).
Fig. 4. a: Low magnification of the surface of the nasal cavity taken from a transition region. Patches of respiratory (R) epithelium (dark areas) can be seen within the olfactory (0) region. x28. b Higher magnification illustrating the contrast in surface texture between respi- ratory (R) and olfactory (0) epithelium. x 188.
Fig. 5. At higher magnification the dendritic knobs (K) and cilia in olfactory regions are visible and appear to form a mat-like surface. In contrast, the respiratory epithelium (R) is characterized by a straight and more parallel arrangement of cilia that extend into the nasal cavity. x781.
Fig. 6. This figure illustrates a different kind of transition region. Cells covered with short stubby microvilli surround nasal duct openings (*). A mixture of long sensory cilia (arrows) and shorter sensory cilia can be seen. x621.
Basal cells Basal cells were generally located in the basal epithelial
region, either alone or in clusters adjacent to the basal lamina. They were rounded or cuboidal (5-7 Wm) and had rough cellular surfaces (Figs. 12,13).
Microvillar cells Some cells resembled microvillar cells. These were typi-
cally close to the mucosal surface (Figs. 22, 23). Microvillar cells are flask- or pear-shaped (5-7 pm), have clear cyto- plasm and a tapered apical surface, from which a tuft or cluster of microvilli extend into the nasal cavity. An axon- like process originates a t the basal region of microvillar cells (Fig. 24). The processes are thin (0.4-0.6 pm), unbranched, and pass basally, often surrounded by the lateral wall of adjacent supporting cells. We were unable to determine if the axon-like processes joined axons (olfactory or microvil- lar) in the epithelium or the lamina propria.
DISCUSSION Previous SEM studies of human olfactory epithelium
were limited to surface morphology of the olfactory mucosa, examining fetal development of the nasal cavity (Yamada, '83) and olfactory knob morphology in adults (Ohno et al., '81). In this study epithelial fractures allowed examination of the full depth of the epithelium as well as the mucosal surface. This also made i t possible to study the three- dimensional organization of the human olfactory epithe- lium.
Transition region The human olfactory epithelial region is often described
as a small area (1-5 cm2) covering part of the superior turbinate, septum, and roof of the nasal cavity (Ganong, '77; Berglund and Lindvall, '82; Moran et al., '82a; Castellucci, '85). The area varies between individuals and the transition from olfactory to respiratory is thought to be well defined. We were able to distinguish respiratory from olfactory epithelium regions of large areas of the nasal cavity, on the basis of surface texture. The transition boundary was not always sharp. Instead, there is an irregular zone where olfactory neurons are in areas of respiratory epithelium. There were also patches of respiratory epithelium in the superior nasal cavity, thought to be a purely olfactory area.
Mixing of respiratory and olfactory epithelial tissues has been previously reported in adult humans (Naessen, '71; Nakashima et al., '84). Naessen ('71) noted that the olfac- tory epithelium in adults was mixed with respiratory epithe- lium and seemed disorganized when compared to fetal tissue. Nakashima et al. ('84) also observed regions of disorganization or degeneration in adult tissue, and found islands of respiratory epithelium in olfactory areas. The regions of disorganization and mixing made it difficult to distinguish respiratory and olfactory epithelium.
Changes in the olfactory epithelium have been reported following head trauma, viral infections, and other clinical disorders (Douek et al., '75; Jafek et al., '89). Degenerative changes in the olfactory epithelium have also been observed in young adults (Nakashima et al., '84), suggesting that some changes may be associated with the natural aging process. Our material was obtained postmortem, and the history of olfactory function in our subjects was unknown. I t is possi- ble that our subjects may have been exposed to environmen-
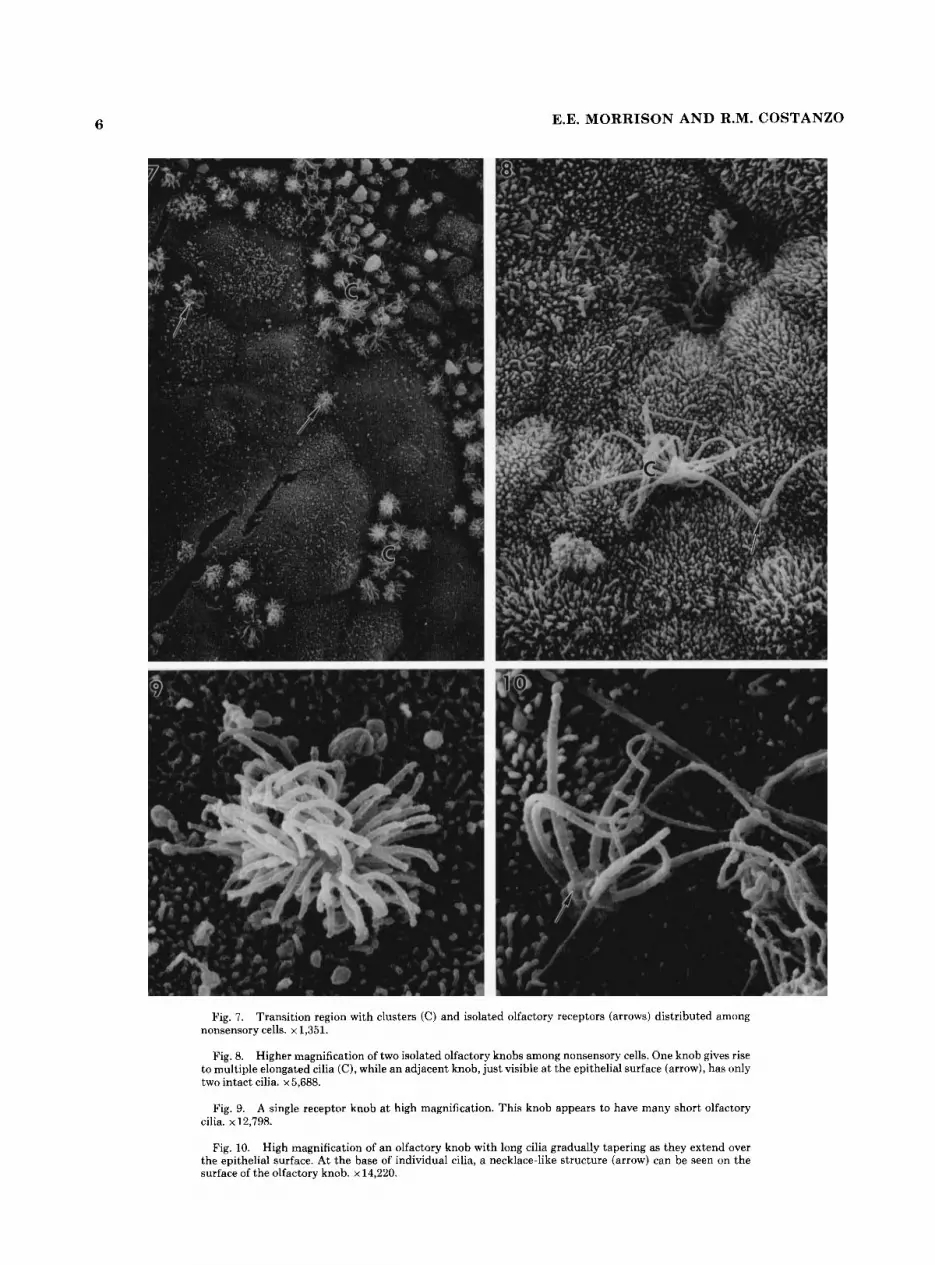
6 E.E. MORRISON A N D R.M. COSTANZO
Fig. 7. Transition region with clusters (C) and isolated olfactory receptors (arrows) distributed among nonsensory cells. x 1,351.
Fig. 8. Higher magnification of two isolated olfactory knobs among nonsensory cells. One knob gives rise to multiple elongated cilia (C), while an adjacent knob, just visible a t the epithelial surface (arrow), has only two intact cilia. x 5,688.
Fig. 9. A single receptor knob a t high magnification. This knob appears to have many short olfactory cilia. x 12,798.
Fig. 10. High magnification of an olfactory knob with long cilia gradually tapering as they extend over the epithelial surface. At the base of individual cilia, a necklace-like structure (arrow) can be seen on the surface of the olfactory knob. x 14,220.

Fig. 11. Low-power three-dimensional scanning view of the olfactory epithelium and lamina propria. The olfactory epithelium (E) overlies a thick connective tissue lamina propria that contains olfactory axon faxicles (Ax) and blood vessels (V). x 248.
Fig. 13. Supporting cells (S) are columnar cells and extend the full depth of the epithelium. An olfactory neuron (0) with its dendrite and a basal cell (B) can be seen among supporting cells. x 1,241.
Fig. 12. Light micrograph of a transverse section through the olfactory epithelium, with supporting cells (S ) , olfactory neurons (O), basal cells (B), and microvillar cells (M). V, blood vessels. x806.

8 E.E. MORRISON AND R.M. COSTANZO
Fig. 14. Cell bodies of olfactory neurons (0) observed in the lower epithelial region. Dendritic processes (D) extend upwards to the epithe- lial surface along the length of adjacent supporting cells. Fine cellular processes (arrows) can be seen extending from supporting cells and appear to contact adjacent dendrites. x 2,964.
Fig. 15. Cell bodies of olfactory neurons (0) observed in the upper region of the epithelium. These neurons have short thick dendrites (arrows) that terminate a t the epithelial surface and give rise to sensory cilia. ~4,000.
Fig. 16. Low-power scanning view of a region of the upper nasal cavity covered with a characteristic dense mat of olfactory cilia. x 1,716.
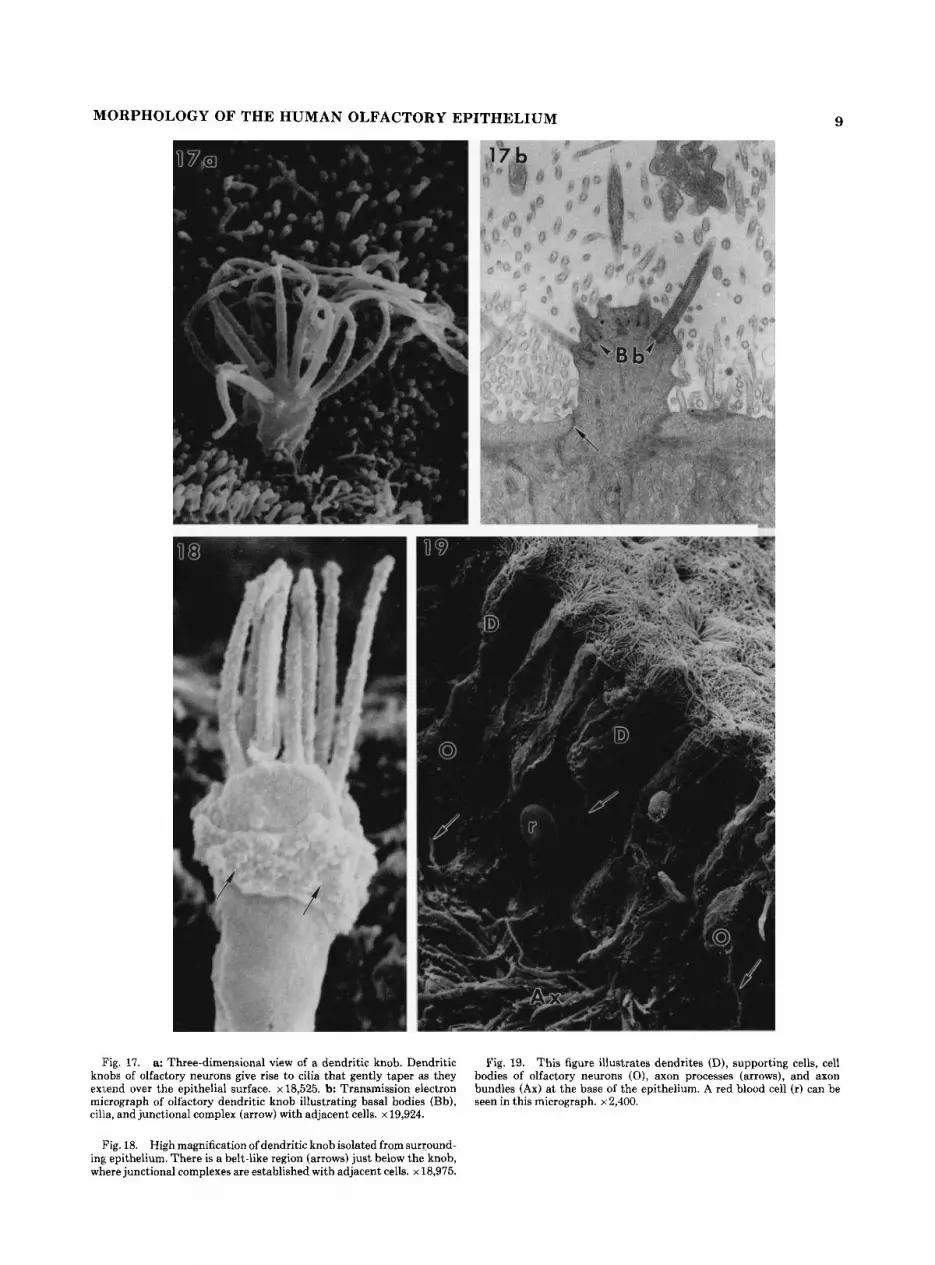
MORPHOLOGY OF THE HUMAN OLFACTORY EPITHELIUM 9
Fig. 17. a: Three-dimensional view of a dendritic knob. Dendritic knobs of olfactory neurons give rise to cilia that gently taper as they exl,end over the epithelial surface. ~18,525. b: Transmission electron micrograph of olfactory dendritic knob illustrating basal bodies (Bh), cilia, and junctional complex (arrow) with adjacent cells. x 19,924.
Fig. 19. This figure illustrates dendrites (D), supporting cells, cell bodies of olfactory neurons ( O ) , axon processes (arrows), and axon bundles (Ax) at the base of the epithelium. A red blood cell (r) can be seen in this micrograph. x 2,400.
Fig. 18. High magnification of dendritic knob isolated from surround- ing epithelium. There is a belt-like region (arrows) just below the knob, where junctional complexes are established with adjacent cells. x 18,975.
10 E.E. MORRISON AND R.M. COSTANZO
Fig. 20. Low-power scanning view of the lamina propria. Olfactory axons fasciculate within the lamina propria to form large axon bundles (arrows) that pass centrally towards the olfactory bulb. x145.
Fig. 21. Dorsal view of the cribriform plate region in the anterior cranial fossa. Axon bundles (arrows) from the nasal cavity pass through the foramen of the cribriform plate and project to second order cells in the olfactory bulb (OB). x44.

MORPHOLOGY OF THE HUMAN OLFACTORY EPITHELIUM 11
Fig. 22. Pear-shaped microvillar cell near the epithelial surface. The apical cell surface appears to have a tuft of short microvilli (M), whereas the adjacent olfactory knob (K) gives rise to long tapering cilia. ~4,590.
Fig. 23. Transmission electron micrograph of a microvillar cell illustrating its characteristic cytoplasm and junctional complex with adjacent cells (arrows). x 10,582.
Fig. 24. a: Low-power magnification of fractured olfactory epithe- lium illustrating the typical location of microvillar cells (M) near the epithelial surface. ~4,131. b: Axon-like processes (arrows) from microvil- lar cells (M) extend basally between supporting cells. ~3,060.

12 E.E. MORRISON AND R.M. COSTANZO
tal agents (antigens, chemical vapors, virus, etc.) resulting in patches of respiratory-like cells. I t is also possible that the epithelial variations we observed, (even at 7 weeks of age) are normal for the human olfactory epithelium.
Human olfactory tissue may not be as clearly defined or as uniform a neuroepithelium as previously believed. Our results illustrate the variability in the distribution of the olfactory region in humans and the potential difficulty in obtaining and interpreting reliable biopsy samples of olfac- tory tissue. We are attempting to map and further define the normal distribution of olfactory areas in the human nasal cavity. Such maps would be valuable to the clinician in the diagnosis and treatment of olfactory disorders.
Olfactory neurons Olfactory neurons are bipolar cells with apical dendrites
that terminate in ciliated knobs a t or above the epithelial surface. Knobs above the surface had up to 30 cilia, while those among supporting cell microvilli a t the surface gener- ally had less than 10. These observations are in agreement with previous human SEM (Ohno et al., '81; Yamada, '83) and TEM (Moran et al., '82a) studies. Areas with fewer cilia may represent young neurons or cells exposed to environmen- tal agents.
Olfactory axons pass basally, enter the lamina propria forming fila olfactoria, and project centrally through the cribriform plate of the ethmoid bone. They then enter the CNS, synapsing with second-order neurons in the olfactory bulb. The region where the fila olfactoria enter the CNS is susceptible to injury. Intracranial movement of the brain in head trauma can generate significant shearing forces. This can damage or tear the thin olfactory axons, disrupting the nerve pathway and affecting the sense of smell (Zusho, '82; Costanzo and Becker, '86; Jafek et al., '89). Head injury patients are reported to have a reduction (hyposmia) or loss (anosmia) of odor detection and identification ability (Cos- tanzo et al., '86). This may be due to a severing of the olfactory axons (axotomy) a t the cribriform plate, resulting in retrograde degeneration of olfactory neurons. In nonhu- man primates, olfactory axotomy causes retrograde degener- ation of the mature olfactory neurons in the epithelium (Monti-Graziadei et al., '80). Reconnection with the olfac- tory bulb follows recovery (neurogenesis and replacement of olfactory neurons) (Graziadei and Monti-Graziadei, '79; Costanzo, '84). Some head injury patients have been re- ported to recover some olfactory function (Sumner, '67; Costanzo and Becker, '86). It is possible that replacement of olfactory neurons and their functional connections with the olfactory bulb also occurs in humans.
Supporting cells Olfactory supporting cells span the epithelium, taper
basally, and attach near the basal lamina. Yamada ('83) observed different types of supporting cells during human development. We also find variation among supporting cells but were unable to determine if distinct cell types exist, such as those in nonhuman vertebrates (Rafols and Getchell, '83; Costanzo and Morrison, '89). Supporting cells probably provide physical support and insulation for receptor neu- rons. These cells have been observed partially or completely surrounding the neurons and their processes (Graziadei, '71; Graziadei and Monti-Graziadei, '76; Costanzo and Morri- son, '89). Our observations of human supporting cells are similar to those in other vertebrates. Extensive contacts
between olfactory neurons and supporting cells have also been described in other vertebrates (Rafols and Getchell, '83; Morrison and Costanzo, '89). In the human olfactory epithelium, we observed contacts between supporting cells and olfactory neurons resembling those of glial cells with neural elements in the CNS (Connors et al., '84). I t may be possible that supporting cells of the human olfactory epithe- lium are analogous to glia within the CNS.
In the upper epithelial region supporting cells are of broad columnar shape with a microvillar surface. We occasionally observed ballooning from the apical cell surface. Apical swelling has been observed by others in rapidly fixed human olfactory tissue and may reflect supporting cell sensitivity to fixation or postmortem changes (Graziadei, '75; Saini and Breipohl, '76; Moran et al., '82a).
Microvillar cell Moran e t al. ('82a) described the fine structure of human
olfactory epithelium, In addition to the three cell types observed in most vertebrates, they consistently found a fourth type, the microvillar cell (Moran et al., '82b). These cells were located near the epithelial surface, had clear cytoplasm, and displayed fine structure characteristics simi- lar to neurons. Although the function of these cells is unknown, Moran et al. ('82a,b) speculated that these cells may be secondary chemoreceptors.
The chemosensory receptor cells of the vomeronasal organ are also microvillar and play an important role in odor detection and reproductive function (Halpern, '87). They are primary sensory neurons derived from neural ectoderm (Graziadei and Tucker, '70; Cuschieri and Banister, '75). Microvillar cells with morphological characteristics of neu- rons have also been reported in the nasal cavity (Menco, '80; Moran et al., '82b; Menco and Farbman, '85), trachea (Jeffery and Reid, '75), and lung (Meyrick and Reid, '68).
Our SEM observations demonstrate microvillar cells in the human olfactory epithelium, similar to those reported by Moran et al. ('82b). They were typically observed near the epithelial surface. A tuft of microvilli extends into the nasal cavity, while an axon process passes basally between and within nearby supporting cells. I t was unclear whether these axon-like structures fasciculated with olfactory axons within the epithelium or the lamina propria. I t should be pointed out that the number of microvillar cells observed in the human olfactory epithelium was considerably less than the other epithelial cell types. I t also remains unclear what function, if any, these cells perform in the human olfactory epithelium.
In summary, we have described the detailed cellular morphology of the human olfactory epithelium. There has been increased interest in clinical disorders of olfactory function resulting from chemical exposure, airborne viruses and head trauma (Twomey, et al., '79; Scott, '89; Jafek et al., '89). Our findings, describing the normal morphology of the human olfactory epithelium, could be important in under- standing both clinical disorders as well as basic physiologi- cal mechanisms that affect olfactory function in humans.
ACKNOWLEDGMENTS We thank Drs. Steven Price, Pasquale P.C. Graziadei, and
Nitya Ghatak for their comments and suggestions during the preparation of this manuscript. We also thank Judy

MORPHOLOGY OF THE HUMAN OLFACTORY EPITHELIUM 13
Williamson, Peter Heywood, and Nancy Koster for their technical assistance and Jeanette Acree for her invaluable assistance in the preparation of this manuscript. This work was supported by NIH grant DC 00165 from the National Institute on Deafness and Other Communicative Disorders (NIDCD).
LITERATURE CITED I3erglund, B., and B. Lindvall (1982) Olfaction. In D.F. Proctor and I.
Anderson (eds): The Nose Upper Airway Physiology and the Atmo- spheric Environment. New York Elsevier, pp. 279-298.
Castellucci, U.F. (1985) The Chemical senses: Taste and smell. In E.R. Kandel and J.H. Schwartz (eds): Principles of Neural Science. New York Elsevier, pp. 409-425.
Connors, B.W., L.S. Benardo, and D.A. Price (1984) Carbon dioxide sensitiv- ity of dye coupling among glia and neurons of the neocortex. J. Neurosci. 4t1324-1330.
Costanzo, R.M. (1984) Comparison of neurogenesis and cell replacement in the hamster olfactory system with and without a target (olfactory bulb). Brain Res. 307:295-301.
Costanzo, R.M., and D.P. Becker (1986) Smell and taste disorders in head injury and neurosurgery patients. In H.L. Meiselman and R.S. Rivilin (eds): Clinical Measurement of Taste and Smell. New York: MacMillian Pub. Co., pp. 565-578.
Costanzo, R.M., P.G. Heywood, J.D. Ward, and H.F. Young (1986) Neurolog- ical applications of clinical olfactory assessment. Ann. NY Acad. Sci. 510:242-244.
Costanzo, R.M., and E.E. Morrison (1989) Three-dimensional scanning electron microscopic study of the normal hamster olfactory epithelium. J. Neurocytol. 18:381-391.
Cuschieri, A., and L. Bannister (1975) The development of the olfactory mucosa in the mouse: Light microscopy. J. Anat. 119:277-286.
Douek, E., L.H. Bannister, and H.C. Dodson (1975) Recent advances in the pathology of olfaction. Proc. R. SOC. Med. 68t467-470.
Engstrom, H., and G. Bloom (1953) The structure of the olfactory region in man. Acta Otolaryngol. 43:ll-21.
Ganong, W.F. (1977) The Nervous System. California: Lange Medical Publications, p. 114.
Graziadei, P.P.C., and D. Tucker (1970) Vomeronasal receptors in turtles. Z. Zellforsck. 105:498-514.
Graziadei, P.P.C. (1971) Topographical relations between olfactory neurons. Z. Zellforsck. Mikrosh. Anat. I18t449-466.
Graziadei, P.P.C. (1975) Application of scanning electron microscopy and autoradiography in the study of olfactory mucosa. In D.G. Moulton, A. Turk and J.W. Johnston (eds): Methods in Olfactory Research. New York: Academic Press, pp. 191-240.
Graziadei, P.P.C., and G.A. Monti-Graziadei (1976) Olfactory epithelium of Necturus maculosus and Ambystoma tigrinum. J. Neurocytol. 5~11-32.
Graziadei, P.P.C., and G.A. Monti-Graziadei (1979) Neurogenesis and neuron regeneration in the olfactory system of mammals. I. Morphological aspects of differentiation and structural organization of the olfactory sensory neurons. J. Neurocytol. 8:l-18.
Halpern, M. (1987) The organization and function of the vomeronasal system. Annu. Rev. Neurosci. ZOt325-362.
Jafek, B.W., P.M. Eller, B.A. Esses, and D.T. Moran (1989) Post-traumatic anosmia. Arch. Neurol. 46~300-304.
Jeffery, P.K., and L. Reid (1975) New observations of rat airway epithelium: a quantitative and electron microscopic study. J. Anat. 120:295-320.
Larsen, P., and M. Tos (1982) Development of the ciliary epithelium in the human nose. Rhinology 200:138-143.
Lenz, H. (1977) Surface of the olfactory region in man observed by scanning electron microscope. Acta Otolaryngol. 84:145-154.
Menco, B.Ph.M. (1977) A qualitative and quantitative investigation of olfactory and respiratory nasal surfaces of cow and sheep based on various ultrastructural and biochemical methods. Communs. Agric. Univ. Wa- geningen. 77-13:l-157.
Menco, B.Ph.M. (1980) Qualitative and quantitative freeze fracture studies on olfactory and nasal respiratory structures of frog, ox, rat and dog. Cell Tissue Res. 207t183-209.
Menco, B.P., and A.I. Farbman (1985) Genesis of cilia and microvilli of rat nasal epithelia during pre-natal development. I. Olfactory epithelium, qualitative studies, J. Cell Sci. 78:283-310.
Meyrick, B., and L. Reid (1968) The alveolar brush cell in rat lung-a third pneumocyte. J. Ultrastructure Res. 23:71-80.
Monti-Graziadei, G.A., M.S. Karlan, J.J. Bernstein, and P.P.C. Graziadei (1980) Reinnervation of the olfactory bulb after section of the olfactory nerves in monkey (Sarniri Sciureus). Brain Res. 189t343-354.
Moran, D.T., J.C. Rowley, and B.W. Jafek (1982a) The fine structure of olfactory mucosa in man. J. Neurocytol. 11:721-746.
Moran, D.T., J.C. Rowley, and B.W. Jafek (1982b) Electron microscopy of human olfactory epithelium reveals a new cell type: The microvillar cell. Brain Res. 253:3946.
Morrison, E.E., and R.M. Costanzo (1989) Scanning electron microscopic study of degeneration and regeneration in the olfactory epithelium after axotomy. J. Neurocytol. 18t393-405.
Mygrind, N. (1975) Scanning electron microscopy of the human nasal mucosa. Rhinology 1657-75.
Naessen, R. (1971) An enquiry on the morphological characteristics and possible changes with age in the olfactory region of man. Acta Otolaryn- gol. 71:49-62.
Nakashima, T., C.P. Kimmelman, and J.B. Snow (1984) Structure of human fetal and adult olfactory neuroepithelium. Arch Otolaryngol. 110:641- 646.
Ohno, I., M. Ohyma, Y. Hanamure, and K. Ogawa (1981) Comparative Anatomy of olfactory epithelium. Biomed. Res. 2:455-458.
Pyatkina, G.A. (1982) Development of olfactory epithelium in man. Z. Mikrosk. Anat. Forsch. 96t361-372.
Rafols, J.A., and T.V. Getchell (1983) Morphological relations between the receptor neurons, sustentacular cells and Schwann cells in the olfactory mucosa of the salamander. Anat. Rec. 206:87-101.
Saini, K.D., and W. Breipohl (1976) Surface morphology in the olfactory epithelium of normal male and female rhesus monkeys. Am. J. Anat. 147:433-446.
Scott, A. (1989) Medical management of taste and smell disorders. Ear Nose Throat J. 68t386-392.
Sumner, D. (1967) Post-traumatic ageusia. Brain 90:187-202. Twomey, J.A., C.M. Barker, G. Robinson, and D. Howell (1979) Olfactory
mucosa in herpes simplex encephalitis. J. Neurol. Neurosurg. Psych. 42t983-987.
Wang, R.T., and M. Halpern (1980) Scanning electron microscopic studies of the surface morphology of the vomeronasal epithelium and olfactory epithelium of garter snakes. Am. J. Anat. 157t399-428.
Yamada, S. (1983) Scanning electron microscopic study of olfactory epithelia. J. Clin. Electron Microsc. 16t95-108.
Zusho, H. (1982) Posttraumatic anosmia. Arch. Otolaryngol. 108:90-92.