precise circuitry links bilaterally symmetric olfactory maps
TRANSCRIPT

Neuron
Article
Precise Circuitry Links BilaterallySymmetric Olfactory MapsZhiqiang Yan,1,2 Jie Tan,1,2 Chang Qin,2 Yao Lu,2 Cheng Ding,2 and Minmin Luo2,*1Institute of Biophysics, Chinese Academy of Sciences, Beijing, 100101, China2National Institute of Biological Sciences, Beijing, 102206, China
*Correspondence: [email protected] 10.1016/j.neuron.2008.03.012
SUMMARY
Olfactory sensory neurons expressing a commonreceptor gene converge onto one or a few glomeruliwith stereotyped positions within the mouse mainolfactory bulb (MOB), producing a map of �1800olfactory columns representing �1000 odorantreceptors. Here, we report that this precise olfactorymap is maintained over several synapses thatultimately cross MOB hemispheres to link bilateralisofunctional olfactory columns. Focal injection oftracer into genetically identified glomeruli revealedan exquisite topography that involves a bilateral con-nection via the anterior olfactory nucleus parsexterna (AONpE) that links isofunctional olfactorycolumns in the contralateral MOB. Physiologicaland behavioral assays revealed an important rolefor the AONpE in bilateral exchange of odorant-spe-cific information. These results indicate that the inter-bulbar link through the AONpE integrates bilateralolfactory sensory maps and exchanges olfactoryinformation, positioning it as a unique model systemfor studying interhemispheric connections.
INTRODUCTION
All sensory modalities utilize topographically organized sensory
maps to represent features of sensory stimuli (Luo and Flanagan,
2007; O’Leary et al., 1999). Central topography typically reflects
the initial organization of the sensory epithelium, in which recep-
tor neurons are spatially organized based on their types and tun-
ing (Luo and Flanagan, 2007; O’Leary et al., 1999). The olfactory
system is unusual in this regard because olfactory sensory neu-
rons (OSNs) expressing a given receptor gene out of a repertoire
of �1000 are scattered in the olfactory epithelium (Buck and
Axel, 1991; Mombaerts et al., 1996; Ressler et al., 1994; Vassar
et al., 1994; Zhang and Firestein, 2002). This disarray is dramat-
ically transformed to ordered organization at the level of glomer-
ulus in the MOB. Here, OSNs expressing a common receptor
gene converge precisely into one or two glomeruli with a stereo-
typed location in the ipsilateral MOB (Mombaerts et al., 1996;
Ressler et al., 1994; Vassar et al., 1994). In mice, each MOB
has�1800 glomeruli, and in each glomerulus OSNs form synap-
ses with a specific set of mitral/tufted cells (Shepherd et al.,
2004). Like columns in the cerebral cortex, a glomerulus forms
part of a functional unit, or ‘‘olfactory column,’’ consisting of
mitral/tufted cells, periglomerular cells, and underlying granule
cells (Shepherd et al., 2004; Willhite et al., 2006). Isofunctional
olfactory columns are those that are associated with the same
odorant receptors. Each mouse MOB can thus be viewed as
a spatial map containing �1800 olfactory columns with parallel
inputs and outputs.
The stereotyped position of identified glomeruli is preserved
not only between individuals but also between hemispheres
within an individual, creating bilaterally symmetric olfactory
sensory maps (Mombaerts et al., 1996; Ressler et al., 1994;
Rubin and Katz, 1999; Uchida et al., 2000; Vassar et al.,
1994; Wachowiak and Cohen, 2001). Although bilateral olfac-
tory bulbs lack direct connections between them, they can in-
teract indirectly via feedback pathways. A cortical area called
the anterior olfactory nucleus (AON) is known to contain neu-
rons that project to the contralateral MOB (Davis and Macrides,
1981; Haberly and Price, 1978; Schoenfeld and Macrides,
1984; Scott et al., 1985). Within the AON, the anterior olfactory
nucleus pars externa (AONpE) receives input from the ipsilat-
eral MOB and projects exclusively to the granule cell layer of
the contralateral MOB, forming the interbulbar association sys-
tem (Davis and Macrides, 1981; Schoenfeld and Macrides,
1984; Scott et al., 1985). Many important questions remain un-
addressed about the organization of this system. In particular,
do bilateral olfactory maps interact with any specificity at the
level of odorant receptors, and is there a functional role if
such interactions exist?
We studied these questions by utilizing genetically engineered
mice that express green fluorescence protein (GFP) in a specific
set of OSNs that also express the endogenous M71 receptor.
This receptor has a characterized ligand; thus, we were able to
probe both anatomical and functional organization. By targeting
tracers to genetically identified glomeruli, we uncovered an as yet
unknown level of organization in the olfactory system, where
bilateral isofunctional olfactory columns are precisely linked by
neurons in the AONpE. We found that odorant-specific activation
of the contralateral MOB required the AONpE. In addition, we
developed a behavioral assay to establish the important role of
the AONpE in crosslateral transfer of odorant-specific olfactory
memory. These results thus demonstrate that the precision of
the olfactory sensory map is preserved along interhemispheric
circuitry and that these connections contribute to bilateral
exchange of specific olfactory information, including memorized
olfactory associations.
Neuron 58, 613–624, May 22, 2008 ª2008 Elsevier Inc. 613

Neuron
Precise Linking of Bilateral Olfactory Maps
RESULTS
The Interbulbar Association System Links BilateralIsofunctional Olfactory ColumnsTo target tract tracers into a single identified olfactory glomeru-
lus, we first utilized a genetically engineered mouse line in which
GFP is coexpressed with the odorant receptor M71 (Bozza et al.,
2002). In these mice, the lateral GFP-labeled M71 glomerulus is
typically located in the dorsoposterior surface of the MOB.
Guided by GFP fluorescence, we made focal iontophoretic injec-
tions of fluorescence-conjugated dextran amines into the lateral
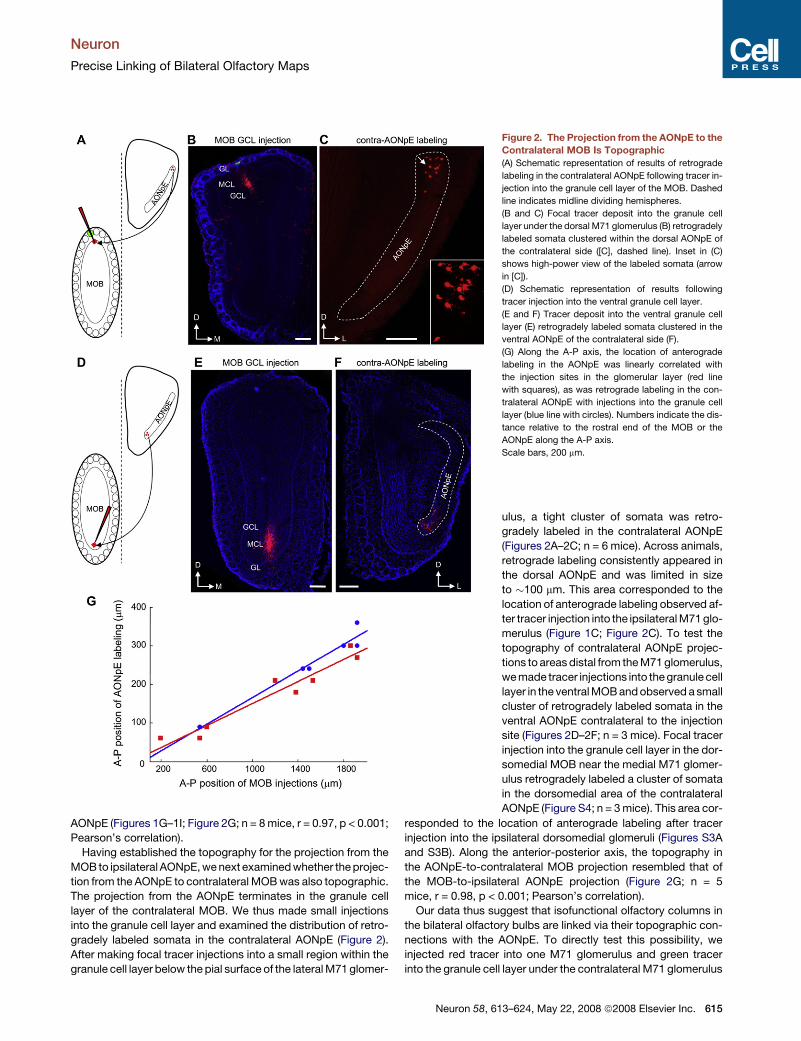
Figure 1. The Projection from the MOB to the Ipsilat-
eral AONpE Is Topographic
(A) Schematic representation of experimental strategy and re-
sulting anterograde labeling. Focal deposit of tract tracer (red
pipette) into a dorsal M71 glomerulus (green) anterogradely
labels a small patch of terminals in the dorsal ipsilateral AONpE.
(B) Focal injection of red tracer into a single dorsal M71 glo-
merulus (green). Inset shows a high-power view of the injec-
tion site (arrow). Glomeruli were delineated by DAPI labeling
(blue).
(C) This injection anterogradely labeled a small patch of axo-
nal terminals (arrow) in layer Ia of the dorsal ipsilateral AONpE
(dashed lines). Inset shows a high-power view of axonal label-
ing (arrow).
(D) Schematic representation of results of injecting dyes into
the mitral/tufted cells in the ventral MOB. Same conventions
as in (A).
(E and F) Injection into the ventral layer of mitral/tufted cells (E)
labeled a patch of axon terminals confined to the ventral
AONpE.
(G–I) Topography along the anterior-posterior (A-P) axis for
the MOB-to-AONpE projection. (G) Coronal sections showing
injection sites in the glomerular layer of the anterior (red, top
panel) and posterior (green, bottom panel) sections of an
MOB. Distance between the two sections was�720 mm along
the A-P axis. (H and I) Anterograde labeling in the anterior
AONpE was mostly red (H) whereas that in posterior AONpE
was mostly green (I). Distance between the two sections
was �120 mm along the A-P axis.
In this and following figures, all sections are coronal. GL, glo-
merular layer; MCL, mitral cell layer; GCL, granule cell layer.
Scale bars, 200 mm.
M71 glomerulus (Figures 1A and 1B; Figure S1A,
available online). In all cases, we observed a single
patch of intensely labeled axon terminals within
layer Ia of the dorsal AONpE (Figure 1C; n = 7
mice; see Figure S2 for topology of the AONpE).
The location of the axonal patches in the AONpE
was highly stereotyped across animals and was
spatially restricted, spanning only approximately
100 mm at levels approximately 3.2 mm anterior
to Bregma. These results indicate specificity in
the projections from the MOB to the ipsilateral
AONpE. To examine the topographic relationship
between the MOB and AONpE further, we made
tracer injections into different glomeruli in the
MOB (Figure 1; Figure S3). When tracers were in-
jected into the mitral/tufted cells in the ventral
MOB, we observed a distinct cluster of anterogradely labeled
axonal terminals in the ventral AONpE (Figures 1D–1F; n = 5
mice). Tracer injections into the dorsomedial glomeruli near the
medial M71 glomerulus anterogradely labeled a cluster of axonal
terminals within the dorsomedial AONpE (Figures S3A and S3B;
n = 4 mice). Injections into glomeruli located in the middle level of
the medial MOB labeled a small cluster of terminals in ventrome-
dial AONpE (Figures S3C and S3D; n = 3 mice). We next injected
two tracers with different fluorophore into two distinct positions
along the anterior-posterior axis of the dorsal MOB. Anterog-
radely labeled terminal fields were largely separated in the dorsal
614 Neuron 58, 613–624, May 22, 2008 ª2008 Elsevier Inc.
Neuron
Precise Linking of Bilateral Olfactory Maps
AONpE (Figures 1G–1I; Figure 2G; n = 8 mice, r = 0.97, p < 0.001;
Pearson’s correlation).
Having established the topography for the projection from the
MOB to ipsilateral AONpE, we next examined whether the projec-
tion from the AONpE to contralateral MOB was also topographic.
The projection from the AONpE terminates in the granule cell
layer of the contralateral MOB. We thus made small injections
into the granule cell layer and examined the distribution of retro-
gradely labeled somata in the contralateral AONpE (Figure 2).
After making focal tracer injections into a small region within the
granule cell layer below the pial surface of the lateral M71 glomer-
Figure 2. The Projection from the AONpE to the
Contralateral MOB Is Topographic
(A) Schematic representation of results of retrograde
labeling in the contralateral AONpE following tracer in-
jection into the granule cell layer of the MOB. Dashed
line indicates midline dividing hemispheres.
(B and C) Focal tracer deposit into the granule cell
layer under the dorsal M71 glomerulus (B) retrogradely
labeled somata clustered within the dorsal AONpE of
the contralateral side ([C], dashed line). Inset in (C)
shows high-power view of the labeled somata (arrow
in [C]).
(D) Schematic representation of results following
tracer injection into the ventral granule cell layer.
(E and F) Tracer deposit into the ventral granule cell
layer (E) retrogradely labeled somata clustered in the
ventral AONpE of the contralateral side (F).
(G) Along the A-P axis, the location of anterograde
labeling in the AONpE was linearly correlated with
the injection sites in the glomerular layer (red line
with squares), as was retrograde labeling in the con-
tralateral AONpE with injections into the granule cell
layer (blue line with circles). Numbers indicate the dis-
tance relative to the rostral end of the MOB or the
AONpE along the A-P axis.
Scale bars, 200 mm.
ulus, a tight cluster of somata was retro-
gradely labeled in the contralateral AONpE
(Figures 2A–2C; n = 6 mice). Across animals,
retrograde labeling consistently appeared in
the dorsal AONpE and was limited in size
to �100 mm. This area corresponded to the
location of anterograde labeling observed af-
ter tracer injection into the ipsilateral M71 glo-
merulus (Figure 1C; Figure 2C). To test the
topography of contralateral AONpE projec-
tions to areas distal from the M71 glomerulus,
we made tracer injections into the granule cell
layer in the ventral MOB and observed a small
cluster of retrogradely labeled somata in the
ventral AONpE contralateral to the injection
site (Figures 2D–2F; n = 3 mice). Focal tracer
injection into the granule cell layer in the dor-
somedial MOB near the medial M71 glomer-
ulus retrogradely labeled a cluster of somata
in the dorsomedial area of the contralateral
AONpE (Figure S4; n = 3 mice). This area cor-
responded to the location of anterograde labeling after tracer
injection into the ipsilateral dorsomedial glomeruli (Figures S3A
and S3B). Along the anterior-posterior axis, the topography in
the AONpE-to-contralateral MOB projection resembled that of
the MOB-to-ipsilateral AONpE projection (Figure 2G; n = 5
mice, r = 0.98, p < 0.001; Pearson’s correlation).
Our data thus suggest that isofunctional olfactory columns in
the bilateral olfactory bulbs are linked via their topographic con-
nections with the AONpE. To directly test this possibility, we
injected red tracer into one M71 glomerulus and green tracer
into the granule cell layer under the contralateral M71 glomerulus
Neuron 58, 613–624, May 22, 2008 ª2008 Elsevier Inc. 615
Neuron
Precise Linking of Bilateral Olfactory Maps
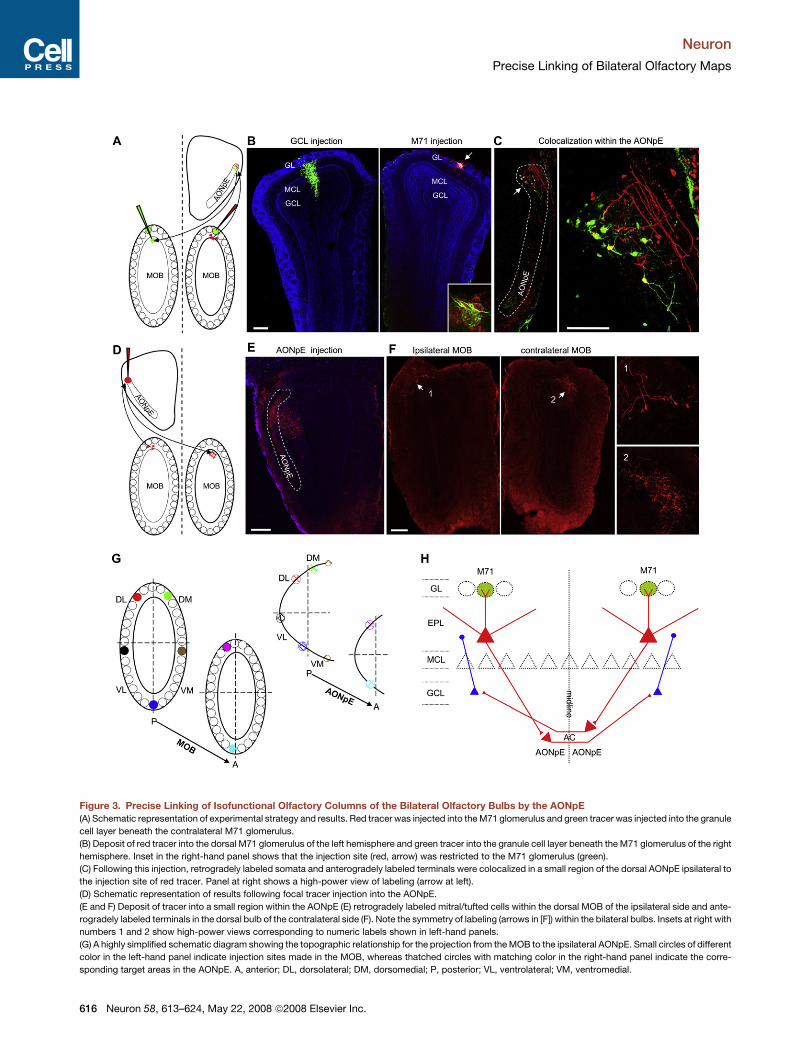
Figure 3. Precise Linking of Isofunctional Olfactory Columns of the Bilateral Olfactory Bulbs by the AONpE
(A) Schematic representation of experimental strategy and results. Red tracer was injected into the M71 glomerulus and green tracer was injected into the granule
cell layer beneath the contralateral M71 glomerulus.
(B) Deposit of red tracer into the dorsal M71 glomerulus of the left hemisphere and green tracer into the granule cell layer beneath the M71 glomerulus of the right
hemisphere. Inset in the right-hand panel shows that the injection site (red, arrow) was restricted to the M71 glomerulus (green).
(C) Following this injection, retrogradely labeled somata and anterogradely labeled terminals were colocalized in a small region of the dorsal AONpE ipsilateral to
the injection site of red tracer. Panel at right shows a high-power view of labeling (arrow at left).
(D) Schematic representation of results following focal tracer injection into the AONpE.
(E and F) Deposit of tracer into a small region within the AONpE (E) retrogradely labeled mitral/tufted cells within the dorsal MOB of the ipsilateral side and ante-
rogradely labeled terminals in the dorsal bulb of the contralateral side (F). Note the symmetry of labeling (arrows in [F]) within the bilateral bulbs. Insets at right with
numbers 1 and 2 show high-power views corresponding to numeric labels shown in left-hand panels.
(G) A highly simplified schematic diagram showing the topographic relationship for the projection from the MOB to the ipsilateral AONpE. Small circles of different
color in the left-hand panel indicate injection sites made in the MOB, whereas thatched circles with matching color in the right-hand panel indicate the corre-
sponding target areas in the AONpE. A, anterior; DL, dorsolateral; DM, dorsomedial; P, posterior; VL, ventrolateral; VM, ventromedial.
616 Neuron 58, 613–624, May 22, 2008 ª2008 Elsevier Inc.

Neuron
Precise Linking of Bilateral Olfactory Maps
(Figures 3A and 3B; n = 4 mice). In all four cases, we observed
colocalization of anterogradely labeled terminals and retro-
gradely labeled somata within a highly restricted region of the
AONpE ipsilateral to the injected M71 glomerulus (Figure 3C).
The dendrites of retrogradely labeled somata intermingled with
ramifying axon terminals from the ipsilateral glomerulus injection
(Figure 3C, right-hand panel). To further test whether this homo-
topic linking is specific to the M71 glomerulus alone or applies
more generally to other glomeruli, we carried out tracing exper-
iments targeting lateral mI7/M71 glomeruli in mI7/M71-
IRES-tauGFP mice (Bozza et al., 2002). In these mice, the gene
encoding the M71 receptor is replaced with the gene for the
mouse I7 receptor, and the lateral mI7/M71 glomeruli are
ectopically located in a position anterior to the endogenous lat-
eral M71 glomeruli in the dorsal MOB. Injections into the lateral
mI7/M71 glomerulus anterogradely labeled a cluster of axon
terminals in the dorsal AONpE that was more anterior to the
AONpE area targeted by the lateral M71 glomerulus (Figures
S5A–S5C; n = 4 mice). Small injections into the granule cell layer
below the lateral mI7/M71 glomerulus retrogradely labeled an
area in the contralateral AONpE apparently corresponding to the
area targeted by ipsilateral mI7/M71 glomerulus (Figures S5E
and S5F; n = 4 mice).
Further demonstrating the homotopic linking by the AONpE,
tracer deposit into small regions of the dorsal AONpE labeled
bilaterally symmetric regions in the MOB (Figures 3D–3F; n = 6
mice). Retrogradely labeled somata of mitral/tufted cells within
the ipsilateral bulb and anterogradely labeled axon terminals in
the contralateral bulb occupied highly restricted regions of the
dorsal bulb that were bilaterally symmetric (arrows in Figure 3F).
Confirming previous studies indicating that the AONpE projects
exclusively to the contralateral MOB (Davis and Macrides,
1981; Schoenfeld and Macrides, 1984), we did not observe any
anterograde labeling in cortical regions after tracer injection into
the AONpE.
OSNs expressing common receptors tend to project to just
two glomeruli, one medial and one lateral in the olfactory bulb,
forming two mirror-symmetric isofunctional glomeruli within
each MOB (Mombaerts et al., 1996; Ressler et al., 1994; Vassar
et al., 1994). The two isofunctional glomeruli form precise recip-
rocal connections (Belluscio et al., 2002; Lodovichi et al., 2003),
suggesting tight intrabulbar connections between the medial
and lateral maps. Confirming the precision of the intrabulbar
connections, we observed small axonal patches in the granule
cell layer underlying the medial isofunctional glomeruli in the
ipsilateral MOB when tracers were injected into the lateral M71
or mI7/M71 glomeruli (Figures S1B and S5D). We next tested
whether the medial and lateral maps were represented by
same maps in the AONpE. If the isofunctional glomeruli within
the MOB converge into same areas within the AONpE, then small
tracer injection into the AONpE will retrogradely label two mirror-
symmetric clusters of mitral/tufted cells in the ipsilateral MOB.
Small injections into the AONpE retrogradely labeled only a single
cluster of mitral/tufted cells instead of two mirror-symmetric
clusters in the ipsilateral MOB (Figures 3D–3F; Figure S6; n = 9
mice). Consistently, tracer injections into the medial and lateral
glomeruli labeled distinct clusters of axonal terminals in the
AONpE (Figure 1; Figure S3). These results strongly suggest
that the two mirror-symmetric olfactory maps within the MOB
are represented by spatially distinct areas of the AONpE.
Our data indicate that mitral/tufted cells of a defined glomeru-
lus project topographically to a small region within the ipsilateral
AONpE. In turn, neurons in this AONpE region project to the
contralateral granule cell layer directly beneath the bilaterally
symmetric glomerulus associated with the same odorant recep-
tor. The topographic relationship between the MOB and the
AONpE is illustrated in Figure 3G. Linking of bilateral isofunc-
tional olfactory columns by the AONpE is depicted schematically
in Figure 3H.
Contralateral Relay of Specific Olfactory SensoryInformation Requires the AONpEThe linking of isofunctional olfactory columns in bilateral olfac-
tory bulbs suggested that odorant-specific information might
be relayed across bilateral bulbs by the AONpE. We therefore
tested whether odor-evoked activity within one MOB could be
transferred to the contralateral bulb, and if so, whether such
contralateral activation required the AONpE. Following an estab-
lished stimulation protocol (Schaefer et al., 2001), we used c-Fos
as a marker of odor-evoked neuronal activity within the MOB. We
found that acetophenone (PMK), a known ligand for M71 recep-
tor (Bozza et al., 2002), evoked c-Fos expression in both bulbs
when presented to mice with both nares open. PMK robustly ac-
tivated periglomerular cells and granule cells associated with
each M71 glomerulus and adjacent glomeruli (Figures 4A–4D;
n = 6). When PMK was applied while a single nostril was oc-
cluded, c-Fos expression was absent from the glomerular layer
of the occluded side (Figures 4E–4H). However, c-Fos labeling
was observed in a substantial number of cells within the granule
cell layer of the occluded side, although the overall density of
c-Fos immunopositive cells was significantly reduced (Figures
4E and 4F; 60.5 ± 10.0 cells/0.04 mm2 for both nares open versus
17.8 ± 3.1 cells/0.04 mm2 for ipsilateral naris occluded; mean ±
SD; n = 6 mice, p < 0.001, between-group t test). Activated gran-
ule cells on the occluded side were clustered into areas beneath
the M71 glomerulus and adjacent glomeruli, and formed an acti-
vation map symmetric to that seen in the bulb ipsilateral to the
open naris (Figure 4E). The activation of granule cells within bilat-
erally symmetric regions in contralateral MOB strongly sug-
gested odorant-driven activation of contralateral granule cells
instead of nonselective activation by enhanced mechanosen-
sory input following naris occlusion (Grosmaitre et al., 2007).
We confirmed that granule cells can be activated by contralateral
glomerular activity by electrically stimulating the lateral M71 glo-
merulus. Consistent with odorant-driven c-Fos expression, elec-
trical stimulation activated granule cells below the lateral M71
glomerulus in the contralateral bulb in addition to periglomerular
(H) A schematic diagram summarizing our data on the connectivity patterns linking the MOB to the ipsilateral AONpE and then to the contralateral MOB. AC,
anterior commissure; EPL, external plexiform layer.
Scale bars, 200 mm (B, E, and F); 100 mm (C).
Neuron 58, 613–624, May 22, 2008 ª2008 Elsevier Inc. 617

Neuron
Precise Linking of Bilateral Olfactory Maps
Figure 4. Odorants Passing through One
Naris Activate Isofunctional Olfactory
Columns in the Contralateral MOB
(A) Drawing representing one highly symmetric
activation map as indicated by c-Fos immunore-
activity following application of acetophenone
(PMK). Green spots indicate the M71 glomeruli;
blue dots, c-Fos+ cells. Coronal section contain-
ing the medial M71 glomeruli is shown here to bet-
ter illustrate the symmetry.
(B) Original c-Fos immunoreactivity of region
shown in the dashed box in (A).
(C and D) c-Fos+ periglomerular cells surrounding
the M71 glomeruli (labels and arrows in [B]).
(E–H) A substantial number of granule cells in the
MOB of the occluded side were c-Fos+ following
PMK application to the contraocclusion naris. (E)
c-Fos expression patterns in the bilateral bulbs.
The right-hand naris was occluded in this case.
(F) c-Fos expression pattern in the granule cell
layer beneath the M71 glomerulus of the occluded
side. (G) c-Fos+ periglomerular cells surrounding
the M71 glomerulus of the open side (left arrow
in [E]). (H) Lack of c-Fos immunoreactivity in the
glomerular layer of the occluded side (right arrow
in [E]) indicates effective occlusion.
(I–K) Drawing of c-Fos+ cells following electrical
stimulation of M71 glomerulus on dorsal MOB (I).
This stimulation activated neurons surrounding
the stimulated glomerulus (J) and the granule cells
under the M71 glomerulus in the contralateral
MOB (K).
Scale bars, 200 mm (B and F); 100 mm (J and K).
cells surrounding the M71 glomerulus ipsilateral to stimulation
(Figures 4I–4K; n = 3 mice).
Physiological recordings were carried out to further examine
the effect of neuronal activation by olfactory responses of the
contralateral MOB. Bulbar neurons were recorded extracellularly
from urethane-anaesthetized mice. Neuronal responses to odor-
ant pulses were recorded and compared before and after contra-
lateral naris occlusion. Naris occlusion was monitored by local
field potentials recorded from the glomerular layer in the bulb
ipsilateral to the occluded naris (Figure 5A, top traces). Putative
granule cells were selected based on low spontaneous activity,
a bursting firing pattern, and recording sites 100–500 mm ventral
to the dorsal mitral cell layer. In contrast, mitral/tufted cells were
selected based on high spontaneous firing coupled to respiratory
rhythm and recording sites within a thin layer 200–300 mm ventral
to the dorsal surface. Among 29 granule cells that showed strong
olfactory responses to odorants, a substantial number of them
(n = 14/29 or �48%) exhibited significant change in odorant-
evoked responses following contralateral naris occlusion
(p < 0.05, t test). Of the 14 cells with significant changes, a vast
majority showed reversible reduction in odor-evoked responses
following contralateral naris occlusion (Figure 5A; change of nor-
malized response intensity = �54.7% ± 20.8%; n = 12/14 cells),
618 Neuron 58, 613–624, May 22, 2008 ª2008 Elsevier Inc.
although most of these cells remained responsive to ipsilateral
odorant application. A substantial number of putative mitral/
tufted cells (n = 12/25) showed significant changes in their olfac-
tory responses following contralateral naris occlusion as well.
Five of these twelve mitral/tufted cells with significant changes
exhibited a reversible increase in response strength (Figure 5B;
change in response intensity = 70.0% ± 48.9%), whereas the re-
maining seven cells exhibited reversible decreases (Figure 5C;
change in response intensity =�81.3% ± 45.9%; n = 7/12 cells).
Interestingly, we also observed significant changes of spontane-
ous firing rates from a substantial number of mitral/tufted cells
(Figure 5C; n = 8/25 cells). Most of these cells (n = 7/8) showed
a reversible increase of spontaneous firing following contralateral
naris occlusion (8.0 ± 5.3 Hz versus 16.7 ± 4.0 Hz, before and
after naris occlusion, respectively; p < 0.001, within group t
test), suggesting that the contralateral input may regulate both
olfactory responses and spontaneous activity of mitral/tufted
cells.
We then tested if the AONpE played a role in activating granule
cells in the contralateral MOB. Bipolar stimulating electrodes
were targeted to the soma layer of the dorsal AONpE of anaes-
thetized mice (Figure 6A). Following local electrical stimulation
of the dorsal AONpE, significantly more granule cells were

Neuron
Precise Linking of Bilateral Olfactory Maps
Figure 5. Neuronal Olfactory Responses in the Olfactory Bulb Are Modulated by Contralateral Input
(A) A granule cell’s responses to 0.4% valeraldehyde (left) were dramatically reduced following the occlusion of contralateral naris (middle), and the occlusion
effect was reversible (right). Top traces are the local field potentials recorded from the glomerular layer of the contralateral MOB to confirm the occlusion effect.
Bottom histograms are the peristimulus time histogram of firing rate (spikes/bin) averaged over six to ten trials. In this and following panels, bin width was
determined by the mean duration of the animal’s respiratory cycles.
(B) A mitral cell showed a reversible increase in its response to (-)-isopulegol following the occlusion of contralateral naris. Dashed lines and boxes point to zoom-
in views of spiking activity within the second respiratory cycle during odorant delivery.
(C) A mitral cell showed a reversible decrease in its response strength to propyl acetate and a concomitant increase in spontaneous activity following the occlu-
sion of the contralateral naris.
Horizontal bars under the physiological traces indicate 2 s odorant pulses.
c-Fos+ in the dorsal contralateral MOB compared with those of
unstimulated controls (Figures 6B–6D; 12.3 ± 3.0 cells/0.04 mm2
versus 0.6 ± 1.0 cells/0.04 mm2, with versus without stimulation,
respectively; n = 4 mice, p < 0.001, t test, n = 4 mice). To test
whether the AONpE is necessary for the contralateral activation,
we made small unilateral lesions of the AONpE by pressure-
injecting ibotenic acid (Figure 6E), which kills somata in an area
largely restricted to the AONpE but leaves passing fibers intact
(Figure S7). We then occluded the naris contralateral to the
AONpE lesions and examined c-Fos expression patterns in the
bulbs following the presentation of PMK (n = 6 mice). Virtually
no c-Fos expression was observed in the granule cell layer of
the MOB ipsilateral to the occluded naris and contralateral to
the lesioned AONpE (Figures 6F–6I; 1.2 ± 1.8 cells/0.04 mm2
for occluded side following AONpE lesion; n = 6 mice,
p < 0.001, between-group t test for ipsilateral naris occlusion
with and without AONpE lesion), but the pattern of c-Fos expres-
sion on the side of the open naris appeared normal (Figures 6F
and 6I). Thus, activation of the contralateral MOB requires the
AONpE. In accordance with the role of AONpE in contralateral
transfer of olfactory information, application of PMK activated
ipsilateral AONpE regions comparable to the target area of
M71-associated mitral/tufted cells from mice with contralateral
naris occluded (Figures 6J and 6K).
The AONpE Contributes to InterhemisphericTransfer of Olfactory MemoryOur tracing results showed that the AONpE links bilateral iso-
functional olfactory columns. Our results of c-Fos mapping and
physiological recordings indicated that odorant-specific infor-
mation is relayed between the MOB hemispheres by the AONpE.
Since studies have shown that olfactory memory stored in one
hemisphere can be transferred to the contralateral hemisphere
via the anterior commissure in adult rats (Kucharski and Hall,
1987, 1988), we asked whether the intrabulbar association sys-
tem could contribute to transferring olfactory information that
guides behavioral learning. An adult mouse was allowed to
explore freely in two opposing arms of a T-maze and one of
two odorants (PMK and amyl acetate [AA], 0.5% saturated
vapor) was randomly infused into the arm where the mouse
Neuron 58, 613–624, May 22, 2008 ª2008 Elsevier Inc. 619

Neuron
Precise Linking of Bilateral Olfactory Maps
settled (Figure 7A). If the odorant was AA, the animal received
a mild footshock in the same arm after a 10 s delay, whereas
PMK was associated with voltage application to a bottom plate
in the opposite arm. After�100 trials, mice learned to avoid foot-
shock by rapidly moving to the opposite arm following AA appli-
cation but staying in the same arm following PMK, with a perfor-
mance asymptote over 95% correct (Figure 7B). Their responses
to odorant cues were tested without footshock the following day.
After establishing this behavioral paradigm of olfactory learn-
ing, we trained mice with one naris open and the other occluded.
Mice were divided into four groups based on whether their
AONpE was intact or bilaterally lesioned before training and
whether they were tested with the training naris or the previously
occluded naris (Figure 7C; n = 4, 4, 5, and 5 mice for Groups 1–4,
respectively). By mixing the drug ibotenic acid with dextran
tracers, we confirmed that most of the AONpE neurons were
killed, although in some cases the toxin appeared to spread to
small areas adjacent to the AONpE (Figure 7E). Mice with AONpE
lesions could be trained to associate odorants with footshock,
Figure 6. The AONpE Is Sufficient and Nec-
essary for Contralateral Transfer of Activity
Pattern in the MOB
(A–D) Electrical stimulation of dorsal AONpE (A)
activated granule cells within the dorsal area of
the contralateral MOB. (B) A drawing of c-Fos+
cells following electrical stimulation. (C and D)
c-Fos immunoreactivity within the ipsilateral MOB
(C) and the contralateral MOB (D). Arrow in (A)
points to the stimulation site visualized with dex-
tran dye near the tip of the stimulation electrode.
(E–H) Contralateral AONpE lesion abolished c-Fos
immunoreactivity in the granule cell layer following
PMK application to the contralateral naris. (E) Le-
sion site within the AONpE soma layer (dashed
line) as visualized by fluorescent tracers mixed
with ibotenic acid. (F) c-Fos activity was apparently
normal in the left bulb but was eliminated in the
right bulb following lesion of the left AONpE by ibo-
tenic acid and occlusion of the right naris. (G and H)
c-Fos immunoreactivity within the left (G) and right
(H) dashed boxes in (F).
(I) Group data showing the density of the c-Fos+
granule cells beneath M71 glomeruli following
blank control (no odorant), occlusion of ipsilateral
naris and AONpE lesion (ipsi 3 lesion), occlusion
of ipsilateral naris with AONpE intact (ipsi 3), and
PMK application only. Error bars = SD.
(J and K) PMK-evoked c-Fos immunoreactivity
was mostly limited to the dorsal AONpE region.
(K) shows the c-Fos+ neurons within the dashed
box in (J).
Scale bars, 100 mm.
indicating that the AONpE lesion did not
result in general anosmia. When tested
with the training naris, mice performed
equally well with the AONpE either intact
or lesioned (Figure 7D; 97.5% ± 2.5%
and 95.0% ± 3.9% at asymptote for
Groups 1 and 3, respectively; mean ±
SEM for all behavioral assays; p = 0.63, t test), suggesting that
olfactory learning within the ipsilateral hemisphere was indepen-
dent of the AONpE. When tested with the previously occluded
naris, mice performed similarly well with a sham-lesioned
AONpE (Figure 7D; correct ratio = 91.0% ± 5.1%; p = 0.58,
between-group t test for Groups 1 and 2). However, their perfor-
mance dropped to near-chance level when tested with the pre-
viously occluded naris and lesions of the AONpE (Figure 7D; cor-
rect ratio = 60.8% ± 5.0%, p < 0.01, t test for Groups 2 and 4).
Thus, these results are consistent with the concept that the
AONpE contributes to bilateral exchange of information about
odorant identity.
DISCUSSION
In this study, we have integrated tract tracing in transgenic and
wild-type mice, c-Fos labeling, electrophysiology, and behav-
ioral assays to examine the connectivity and functional role of
the interbulbar association system. Our use of transgenic mouse
620 Neuron 58, 613–624, May 22, 2008 ª2008 Elsevier Inc.

Neuron
Precise Linking of Bilateral Olfactory Maps
lines with GFP-labeled glomeruli permitted a level of mapping
that has been impossible in previous studies. By depositing tract
tracers focally into single identified glomeruli, we find that mitral/
tufted cells associated with a specific glomerulus project topo-
graphically to the AONpE, where the AONpE neurons in turn pro-
ject to granule cells underlying the isofunctional glomerulus in the
contralateral MOB. Previous studies observed no topography in
the longitudinal direction and quadrant-to-quadrant topography
in other directions for the interbulbar association system (Davis
and Macrides, 1981; Haberly and Price, 1978; Schoenfeld and
Macrides, 1984; Scott et al., 1985). This conflict is likely a result
of large tracer injection sites in previous studies. Furthermore,
these early studies were carried out prior to the discovery of
odorant receptors and their symmetric representation in the bi-
lateral olfactory bulbs. Because of the lack of receptor identity
for the bilaterally isofunctional glomeruli, it had remained unclear
what the interbulbar association system connected exactly. By
targeted tract tracing, we have uncovered a link between iso-
functional olfactory columns in the bilateral olfactory bulbs by
the interbulbar association system. More importantly, as sug-
Figure 7. Interhemispheric Information Exchange in the
Olfactory System Depends on the AONpE
(A) Method of testing olfactory conditioning in a T-maze. Ten sec-
onds after initiation, amyl acetate (AA) application was paired with
footshock in the same arm and PMK with shock in the opposite
arm. Mice learned to move to the opposite arm only after AA
application.
(B) A raw trace showing a mouse’s movement after training. Note
that the mouse quickly moved to the opposite arm following AA
(curved arrows) whereas it stayed in the same arm following
PMK. Vertical arrows indicate valve opening for odorant applica-
tion.
(C) Paradigms for testing the role of the AONpE in contralateral
memory transfer. Mice were trained with only one naris open
and then tested with either the training naris (Groups 1 and 3) or
the previously occluded naris (Groups 2 and 4) with the AONpE
either intact (Groups 1 and 2) or lesioned (Groups 3 and 4). In
Group2, mice were sham-lesioned by pressure injection of vehicle.
(D) Test scores for Groups 1–4 shown in (C). Error bars = SEM.
(E) A representative lesion site in the AONpE. Dashed line indicates
soma layer of the AONpE as revealed by DAPI labeling. Scale bar,
200 mm.
gested by the exquisite anatomical linkage, we find
that the AONpE mediates bilateral exchange of odor-
ant-specific signals.
Consistent with the precision of integration within
the AONpE, morphological studies have shown that
AONpE pyramidal neurons possess small dendritic
trees (Reyher et al., 1988; Scott et al., 1985), a special-
ization that could permit maintenance of odorant
selectivity originating in the MOB. Future studies on
the synaptic distribution between bulbar neurons
and the dendritic tree of AONpE neurons will allow
precise quantification of the topography and conver-
gence ratio. Nevertheless, the highly topographic
and stereotypical representation of each glomerulus
in the AONpE is reminiscent of the projection neurons
in Drosophila antennal lobes, an area comparable to the mam-
malian olfactory bulb. These projection neurons possess stereo-
typed patterns of axonal distribution in the protocerebrum (Jeffe-
ris et al., 2007; Marin et al., 2002; Wong et al., 2002).
Genetic studies have contributed substantially to our under-
standing of the molecular mechanisms underlying the wiring in
the MOB (Wang et al., 1998; Yoshihara et al., 1997; Yu and
Bargmann, 2001) and the Drosophila antennal lobe (Komiyama
et al., 2007). It will be interesting to examine the developmental
mechanisms underlying the precise linking of�1800 pairs of glo-
meruli by the interbulbar association system. The topographic
precision of the interbulbar association system is comparable
to that of the intrabulbar association system, which precisely
connects the two mirror-symmetric olfactory maps within the
MOB (Belluscio et al., 2002; Liu and Shipley, 1994; Lodovichi
et al., 2003; Schoenfeld et al., 1985). These two intrabulbar mir-
ror-symmetric maps appear to be represented by distinct areas
in the AONpE, suggesting that both receptor identity and glomer-
ular spatial location interact to establish the linking of bilateral
olfactory bulbs. The refinement of the intrabulbar connections
Neuron 58, 613–624, May 22, 2008 ª2008 Elsevier Inc. 621

Neuron
Precise Linking of Bilateral Olfactory Maps
is activity dependent (Marks et al., 2006), a subject that remains
to be explored in the interbulbar association system.
Olfactory memory formed in one hemisphere can be trans-
ferred to the contralateral hemisphere in rats and humans
(Kucharski and Hall, 1987, 1988; Mainland et al., 2002). The
specific neural circuitry underlying this contralateral transfer of
olfactory memory was unknown. Our c-Fos mapping and phys-
iological recordings show that the activity of specific glomeruli
within one hemisphere of the MOB is transferred to the granule
cells underlying the isofunctional glomeruli in the contralateral
hemisphere. In behavioral assays, mice showed clear deficits
in contralateral transfer of olfactory memory when their AONpE
was lesioned with ibotenic acid. Thus, our experiments suggest
that the AONpE is a critical locus for bilateral relay of olfactory
signals, although cortical areas other than the AONpE can also
contribute to this process. We hypothesize the following sce-
nario in olfactory signal propagation between hemispheres: the
activation of a specific glomerulus activates its associated
mitral/tufted cells, which in turn activate their postsynaptic target
neurons in the AONpE, followed in turn by their targets: the gran-
ule cells underlying the isofunctional glomeruli in the contralat-
eral MOB. The activity of these granule cells may in turn enhance
the activity of mitral/tufted cells associated with the isofunctional
glomeruli by mechanisms such as lateral inhibition (Luo and
Katz, 2001; Yokoi et al., 1995). This scenario may be directly
tested by targeted recordings from mitral/tufted cells associated
with a specific glomerulus and optical stimulation of the contra-
lateral isofunctional glomerulus using a recently developed
transgenic mouse line expressing Channelrhodopsin-2 in
mitral/tufted cells (Arenkiel et al., 2007).
The interbulbar association system may be important for
other behaviors as well. Rats can localize olfactory pulses in
space as arriving from either the left or the right side (Rajan
et al., 2006). The linking of isofunctional columns between the
bulbs may contribute to ‘‘stereo’’ olfactory perception by allow-
ing the system to calculate the bilateral differences of odor-
evoked activity patterns between the isofunctional columns. In
addition, human and animal nasal passages exhibit spontane-
ous change in unilateral airflow resistance (nasal-cycle conges-
tion) (Stoksted, 1953). The AONpE may balance the activity of
the bilateral olfactory bulbs when the same odorant generates
sensory signals of distinct intensities in the bilateral olfactory
epithelia during different phases of nasal cycles (Sobel et al.,
1999).
The two symmetric hemispheres of mammalian forebrain are
connected by dense fibers coursing within the corpus callosum
and anterior commissure. The studies on ‘‘split-brains’’ illustrate
the essential role of interhemispheric communication in percep-
tion and sensory learning (Gazzaniga, 1995). For example, ani-
mals with severed corpus callosum have bilaterally separate
‘‘learning centers,’’ with one hemisphere failing to access the
learned information in another hemisphere (Myers and Sperry,
1958). Our use of transgenic mice with a specific glomerulus
expressing GFP permitted unprecedented mapping of bilateral
connections for specific olfactory glomeruli. These data indicate
a highly structured organization of anatomical pathways carrying
specific olfactory information between the bilateral bulbs, sug-
gesting the interbulbar association system as an advantageous
622 Neuron 58, 613–624, May 22, 2008 ª2008 Elsevier Inc.
model for studying the cellular circuits, function, and develop-
ment of interhemispheric connections.
EXPERIMENTAL PROCEDURES
Adult M71-IRES-tauGFP and mI7/M71-IRES-tauGFP mice (7–12 weeks old)
were used in all experiments except for T-maze assays, in which C57BL/6
mice were used.
Tract Tracing and Ibotenic Acid Lesion
Mice were anaesthetized with a mixture of ketamine (200 mg/kg) and xylazine
(25 mg/kg, i.p.). Guided by green fluorescence of the M71 or mI7/M71 glo-
merulus, biotinylated dextran amines (10K MW, BDA, 10%, Invitrogen) mixed
with Texas red dextran amines (10K MW, TRDA, 1%) or fluorescein-
conjugated dextran amines (10K MW, FDA, 10%) were iontophoretically
injected (1 mA, 1–20 s) into target areas through a micropipette. For lesion of
the AONpE, 0.1–0.5 ml of 0.5% ibotenic acid in saline was pressure injected
into the AONpE. Ibotenic acid was mixed with 1% tetramethylrhodamine
dextran amine (TMR-DA) to visualize the lesion sites. Sham lesions were
carried out by pressure injection of vehicle (1% TMR-DA in saline).
Histology
Mice were overdosed with pentobarbital (300 mg/kg, i.p.) and perfused trans-
cardially with 0.1M phosphate buffered saline (PBS) and then 4% paraformal-
dehyde in PBS. Following postfixation and cryoprotection, coronal sections
(60 mm) were prepared on a freezing microtome (Leica CR 1900). BDA tracers
were visualized by fluorescence-conjugated streptavidin. Sections were
mounted in 50% glycerol with DAPI.
Mapping Neuronal Activity with c-Fos
For odorant exposure, mice were placed overnight in a chamber (volume = 1.8
liters) with continuous airflow (4 l/min) filtered with active charcoal. PMK pulses
(�1% saturated vapor) were presented for 2 min at 5 min intervals over
a 15 min period. For electrical stimulation, mice were first anaesthetized with
urethane (1.64 g/kg injected as 20% solution i.p.). After 4 hr of exposure to
purified air, bipolar tungsten electrodes (1 MU, Microprobes) were placed
within the M71 glomerulus or the AONpE to generate 240 trains of electric
pulses over 20 min, with each train comprising 100 pulses at 30–50 mA
(100 Hz; pulse duration = 0.1 ms). Mice were transcardially perfused 90 min af-
ter either odorant or electrical stimulation and brains were later processed for
c-Fos immunostaining. The primary antibody was a rabbit polyclonal antibody
against c-Fos (1:500, Santa Cruz) and the secondary antibody was Cy3-
conjugated goat anti-rabbit (1:500, Jackson ImmunoResearch).
Confocal Imaging and Analysis
Sections were imaged with a laser scanning confocal microscope (Zeiss LSM
510). Optical stacks (Z-interval 1.0 mm) were projected into single frames.
Whole-view images were constructed by assembling multiple adjacent frames
of 103 images. Borders and cell layers were identified by DAPI labeling and
autofluorescence. To define c-Fos+ cells, we first measured the mean and
SD of background optical signals. Cells with optical intensities >[mean +
3*SD] were considered c-Fos+. The density of c-Fos+ neurons was calculated
by counting the number of c-Fos+ neurons within a square with side length
equal to 200 mm, which is roughly the size of large glomeruli in the olfactory
bulb.
Electrophysiological Recordings
Animals were anaesthetized with urethane. After the dorsal M71 glomerulus
was identified, a small piece of bone above the glomerulus was removed
and tungsten electrodes were lowered below the M71 glomerulus. Odorants
(1% saturated vapor, 2 s pulse duration) were applied with a custom-made
64-channel robotic olfactometer. Data were band-pass filtered at 0.5–
3.0 KHz, digitized at 6 KHz, and analyzed with a custom-written Matlab pro-
gram. For a neuron to be considered responsive to an odorant, its mean firing
rate during odorant application needed to be significantly different (p < 0.05,
paired t test) from its mean spontaneous rate within 2 s preceding stimulus

Neuron
Precise Linking of Bilateral Olfactory Maps
onset. Its response intensity to an odorant was calculated by subtracting the
spontaneous rate from the firing rate during the odorant application. Change
of normalized response intensity was calculated as ([response intensity after
naris occlusion] / [response intensity before naris occlusion]) � 100%.
Behavioral Assay
A detailed description of this method can be found in Supplementary Online
Material. Briefly, adult C57BL/6 mice were trained in a customized T-maze
to associate AA with footshock in the same arm and PMK with footshock in
the opposite arm. Mice learned to avoid footshock by moving to the opposite
arm following AA or staying within the same arm following PMK. After learning
(100% correct for ten trials), they were tested without actual application of volt-
age. A nostril was occluded by inserting a soft rod (0.8 mm in diameter, 4 mm in
length) and then sealing with a small drop of Vetbond tissue adhesive. Animal
movement was monitored with an infrared CCD camera. Training and testing
were controlled by a custom-written Matlab program.
SUPPLEMENTAL DATA
The Supplemental Data for this article can be found online at http://www.
neuron.org/cgi/content/full/58/4/613/DC1/.
ACKNOWLEDGMENTS
We thank Peter Mombaerts and Stuart Firestein for M71-IRES-tauGFP and
mI7/M71-IRES-tauGFP mice; Liqun Luo, Abigail Person, Kazunari Miyami-
chi, and Charles Zuker for comments on the manuscript; and Ji Hu for assis-
tance with Matlab programming. This work was supported by China Ministry
of Science and Technology 863 Projects, Human Frontier Science Program,
and National Natural Science Foundation of China.
Received: October 5, 2007
Revised: January 24, 2008
Accepted: March 8, 2008
Published: May 21, 2008
REFERENCES
Arenkiel, B.R., Peca, J., Davison, I.G., Feliciano, C., Deisseroth, K., Augustine,
G.J., Ehlers, M.D., and Feng, G. (2007). In vivo light-induced activation of neu-
ral circuitry in transgenic mice expressing channelrhodopsin-2. Neuron 54,
205–218.
Belluscio, L., Lodovichi, C., Feinstein, P., Mombaerts, P., and Katz, L.C.
(2002). Odorant receptors instruct functional circuitry in the mouse olfactory
bulb. Nature 419, 296–300.
Bozza, T., Feinstein, P., Zheng, C., and Mombaerts, P. (2002). Odorant recep-
tor expression defines functional units in the mouse olfactory system.
J. Neurosci. 22, 3033–3043.
Buck, L., and Axel, R. (1991). A novel multigene family may encode odorant
receptors: a molecular basis for odor recognition. Cell 65, 175–187.
Davis, B.J., and Macrides, F. (1981). The organization of centrifugal projections
from the anterior olfactory nucleus, ventral hippocampal rudiment, and piri-
form cortex to the main olfactory bulb in the hamster: an autoradiographic
study. J. Comp. Neurol. 203, 475–493.
Gazzaniga, M.S. (1995). Principles of human brain organization derived from
split-brain studies. Neuron 14, 217–228.
Grosmaitre, X., Santarelli, L.C., Tan, J., Luo, M., and Ma, M. (2007). Dual func-
tions of mammalian olfactory sensory neurons as odor detectors and mechan-
ical sensors. Nat. Neurosci. 10, 348–354.
Haberly, L.B., and Price, J.L. (1978). Association and commissural fiber sys-
tems of the olfactory cortex of the rat. J. Comp. Neurol. 178, 711–740.
Jefferis, G.S., Potter, C.J., Chan, A.M., Marin, E.C., Rohlfing, T., Maurer, C.R.,
Jr., and Luo, L. (2007). Comprehensive maps of Drosophila higher olfactory
centers: spatially segregated fruit and pheromone representation. Cell 128,
1187–1203.
Komiyama, T., Sweeney, L.B., Schuldiner, O., Garcia, K.C., and Luo, L. (2007).
Graded expression of semaphorin-1a cell-autonomously directs dendritic tar-
geting of olfactory projection neurons. Cell 128, 399–410.
Kucharski, D., and Hall, W.G. (1987). New routes to early memories. Science
238, 786–788.
Kucharski, D., and Hall, W.G. (1988). Developmental change in the access to
olfactory memories. Behav. Neurosci. 102, 340–348.
Liu, W.L., and Shipley, M.T. (1994). Intrabulbar associational system in the rat
olfactory bulb comprises cholecystokinin-containing tufted cells that synapse
onto the dendrites of GABAergic granule cells. J. Comp. Neurol. 346, 541–558.
Lodovichi, C., Belluscio, L., and Katz, L.C. (2003). Functional topography of
connections linking mirror-symmetric maps in the mouse olfactory bulb.
Neuron 38, 265–276.
Luo, M., and Katz, L.C. (2001). Response correlation maps of neurons in the
mammalian olfactory bulb. Neuron 32, 1165–1179.
Luo, L., and Flanagan, J.G. (2007). Development of continuous and discrete
neural maps. Neuron 56, 284–300.
Mainland, J.D., Bremner, E.A., Young, N., Johnson, B.N., Khan, R.M., Bensafi,
M., and Sobel, N. (2002). Olfactory plasticity: one nostril knows what the other
learns. Nature 419, 802.
Marin, E.C., Jefferis, G.S., Komiyama, T., Zhu, H., and Luo, L. (2002). Repre-
sentation of the glomerular olfactory map in the Drosophila brain. Cell 109,
243–255.
Marks, C.A., Cheng, K., Cummings, D.M., and Belluscio, L. (2006). Activity-
dependent plasticity in the olfactory intrabulbar map. J. Neurosci. 26,
11257–11266.
Mombaerts, P., Wang, F., Dulac, C., Chao, S.K., Nemes, A., Mendelsohn, M.,
Edmondson, J., and Axel, R. (1996). Visualizing an olfactory sensory map. Cell
87, 675–686.
Myers, R.E., and Sperry, R.W. (1958). Interhemispheric communication
through the corpus callosum: mnemonic carry-over between the hemispheres.
AMA Arch. Neurol. Psychiatry 80, 298–303.
O’Leary, D.D., Yates, P.A., and McLaughlin, T. (1999). Molecular development
of sensory maps: representing sights and smells in the brain. Cell 96, 255–269.
Rajan, R., Clement, J.P., and Bhalla, U.S. (2006). Rats smell in stereo. Science
311, 666–670.
Ressler, K.J., Sullivan, S.L., and Buck, L.B. (1994). Information coding in the
olfactory system: evidence for a stereotyped and highly organized epitope
map in the olfactory bulb. Cell 79, 1245–1255.
Reyher, C.K., Schwerdtfeger, W.K., and Baumgarten, H.G. (1988). Interbulbar
axonal collateralization and morphology of anterior olfactory nucleus neurons
in the rat. Brain Res. Bull. 20, 549–566.
Rubin, B.D., and Katz, L.C. (1999). Optical imaging of odorant representations
in the mammalian olfactory bulb. Neuron 23, 499–511.
Schaefer, M.L., Young, D.A., and Restrepo, D. (2001). Olfactory fingerprints for
major histocompatibility complex-determined body odors. J. Neurosci. 21,
2481–2487.
Schoenfeld, T.A., and Macrides, F. (1984). Topographic organization of con-
nections between the main olfactory bulb and pars externa of the anterior
olfactory nucleus in the hamster. J. Comp. Neurol. 227, 121–135.
Schoenfeld, T.A., Marchand, J.E., and Macrides, F. (1985). Topographic orga-
nization of tufted cell axonal projections in the hamster main olfactory bulb: an
intrabulbar associational system. J. Comp. Neurol. 235, 503–518.
Scott, J.W., Ranier, E.C., Pemberton, J.L., Orona, E., and Mouradian, L.E.
(1985). Pattern of rat olfactory bulb mitral and tufted cell connections to the an-
terior olfactory nucleus pars externa. J. Comp. Neurol. 242, 415–424.
Shepherd, G.M., Chen, W.R., and Greer, C.A. (2004). Olfactory bulb. In The
Synaptic Organization of the Brain, G.M. Shepherd, ed. (New York: Oxford
University Press), pp. 165–216.
Sobel, N., Khan, R.M., Saltman, A., Sullivan, E.V., and Gabrieli, J.D. (1999). The
world smells different to each nostril. Nature 402, 35.
Neuron 58, 613–624, May 22, 2008 ª2008 Elsevier Inc. 623

Neuron
Precise Linking of Bilateral Olfactory Maps
Stoksted, P. (1953). Rhinometric measurements for determination of the nasal
cycle. Acta Otolaryngol. Suppl. 109, 159–175.
Uchida, N., Takahashi, Y.K., Tanifuji, M., and Mori, K. (2000). Odor maps in the
mammalian olfactory bulb: domain organization and odorant structural fea-
tures. Nat. Neurosci. 3, 1035–1043.
Vassar, R., Chao, S.K., Sitcheran, R., Nunez, J.M., Vosshall, L.B., and Axel, R.
(1994). Topographic organization of sensory projections to the olfactory bulb.
Cell 79, 981–991.
Wachowiak, M., and Cohen, L.B. (2001). Representation of odorants by recep-
tor neuron input to the mouse olfactory bulb. Neuron 32, 723–735.
Wang, F., Nemes, A., Mendelsohn, M., and Axel, R. (1998). Odorant receptors
govern the formation of a precise topographic map. Cell 93, 47–60.
Willhite, D.C., Nguyen, K.T., Masurkar, A.V., Greer, C.A., Shepherd, G.M., and
Chen, W.R. (2006). Viral tracing identifies distributed columnar organization in
the olfactory bulb. Proc. Natl. Acad. Sci. USA 103, 12592–12597.
624 Neuron 58, 613–624, May 22, 2008 ª2008 Elsevier Inc.
Wong, A.M., Wang, J.W., and Axel, R. (2002). Spatial representation of the glo-
merular map in the Drosophila protocerebrum. Cell 109, 229–241.
Yokoi, M., Mori, K., and Nakanishi, S. (1995). Refinement of odor molecule tun-
ing by dendrodendritic synaptic inhibition in the olfactory bulb. Proc. Natl.
Acad. Sci. USA 92, 3371–3375.
Yoshihara, Y., Kawasaki, M., Tamada, A., Fujita, H., Hayashi, H., Kagamiyama,
H., and Mori, K. (1997). OCAM: A new member of the neural cell adhesion mol-
ecule family related to zone-to-zone projection of olfactory and vomeronasal
axons. J. Neurosci. 17, 5830–5842.
Yu, T.W., and Bargmann, C.I. (2001). Dynamic regulation of axon guidance.
Nat. Neurosci. Suppl. 4, 1169–1176.
Zhang, X., and Firestein, S. (2002). The olfactory receptor gene superfamily of
the mouse. Nat. Neurosci. 5, 124–133.

Neuron
Previews
Interhemispheric Olfactory Circuitand the Memory Beyond
Takeshi Imai1 and Hitoshi Sakano1,*1Department of Biophysics and Biochemistry, Graduate School of Science, The University of Tokyo, Tokyo 113-0032, Japan*Correspondence: [email protected] 10.1016/j.neuron.2008.05.004
In mammals, olfactory sensory neurons project their axons exclusively to the ipsilateral olfactory bulb. Itremains unclear how odor information interacts between the two hemispheres of the brain. In this issue ofNeuron, Yan et al. describe the precise interbulbar connection through the anterior olfactory nucleus parsexterna (AONpE), which links contralateral isotypic olfactory columns.
Since the discovery of odorant receptors
(ORs) (Buck and Axel, 1991), there has
been tremendous progress in under-
standing how odor information is repre-
sented in the olfactory bulb (OB), the first
center for processing odor information.
Airborne odorants are detected with
�1000 types of ORs expressed by olfac-
tory sensory neurons (OSNs) in the olfac-
tory epithelium (OE). Each
OSN expresses only one
functional OR gene in a mono-
allelic manner, following the
one neuron-one receptor rule
(reviewed by Serizawa et al.,
2004). Furthermore, OSNs
expressing a given type of
OR converge their axons to
a specific pair of glomeruli in
each OB, referred to as the
one glomerulus-one receptor
rule (reviewed by Imai and
Sakano, 2007). The result is a
stereotyped, mirror-symmet-
ric glomerular map (Figure 1).
Thus, the odor information
detected in the OE is con-
verted to a topographic map
of activated glomeruli (re-
viewed by Mori et al., 2006).
In the OB, each glomerulus
with its underlying neurons
forms a functional unit called
the ‘‘olfactory column’’ for
odor processing (reviewed
by Wilson and Mainen,
2006). Local interneurons,
such as periglomerular cells
and granule cells, tune the
odor-derived signals that are
conveyed to second-order
neurons, the mitral/tufted
(M/T) cells. Each M/T cell receives direct
inputs from a single glomerulus and sends
branched axons to several distinct re-
gions in the olfactory cortex (OC), as well
as to other brain regions involved in multi-
modal sensory integration. A recent study
demonstrated that activation of specific
sets of glomeruli in the dorsal domain of
the OB elicits stereotyped innate behav-
iors, suggesting the presence of a hard-
wired neural circuit in the mammalian
main olfactory system (Kobayakawa
et al., 2007). In contrast, discrimination
and associative learning of odor informa-
tion appear to be more complex and
plastic. Fundamental questions regarding
these processes remain mostly unan-
swered. It is yet to be elucidated how
odors are recognized and dis-
criminated and how olfactory
memories are stored and re-
trieved. The first step toward
answering these questions
is to describe the neural cir-
cuitry for respective olfactory
responses from both anatom-
ical and functional aspects.
In this issue of Neuron,
Yan et al. (2008) address an
important question for odor
perception: how are the olfac-
tory inputs from two nostrils
coordinated in the brain? In
contrast to retinal projection
in binocular animals, mam-
malian OSNs project their
axons solely to the ipsilateral
OB: right nostril to the right
OB and left nostril to the
left OB. The same is true for
second-order neurons: the
M/T cells project their axons
to the ipsilateral OC. None-
theless, behavioral studies
demonstrate that the olfac-
tory memory learned by one
nostril can be recalled by
stimuli to either nostril (Ku-
charski and Hall, 1987), sug-
gesting the existence of an
interhemispheric connection
Figure 1. Intrabulbar (Lodovichi et al., 2003) and Interbulbar(Yan et al., 2008) Circuitries in the Mouse Olfactory SystemA horizontal section of the mouse olfactory system is schematically shown.Information flow from one particular lateral glomerulus in the left OB is shown.Isofunctional glomeruli are shown in green. Excitatory and inhibitory neuronsare shown in red and blue, respectively. External tufted cells project axonsto the granule cells in the isofunctional olfactory columns, to form intrabulbarconnections (Lodovichi et al., 2003). M/T cells project axons to the OC, includ-ing the AON. In the AONpE, M/T cells synapse to excitatory neurons, which inturn project axons to the granule cells in the isofunctional olfactory column inthe contralateral OB. This disynaptic circuitry forms the interbulbar connec-tion (Yan et al., 2008). AC, anterior commissure; ETC, external tufted cell; GC,granule cell.
Neuron 58, May 22, 2008 ª2008 Elsevier Inc. 465

Neuron
Previews
for olfactory learning and/or recall. Which
neural circuit mediates the interhemi-
spheric communication? And how?
The anterior olfactory nucleus (AON)
is a region of the OC known to contain
neurons that project to the contralateral
OB. This contralateral projection passes
through the anterior commissure, one of
the fiber bundles connecting the two
hemispheres of the brain. Notably, the
anterior olfactory nucleus pars externa
(AONpE) receives inputs from M/T cells
in the ipsilateral OB and sends fibers
exclusively to the granule cell layer in the
contralateral OB. Among the AON subre-
gions, only the AONpE exhibits topo-
graphic projection to the OB (Davis and
Macrides, 1981). In the present study,
Yan et al. (2008) analyzed whether or not
connections through the AONpE link the
OR-specific olfactory columns of oppo-
site OBs. When fluorescent tracers were
injected into a glomerulus for OR-M71,
anterograde labeling was detected in a
specific spot within the ipsilateral AONpE.
Anterograde labeling from various glo-
merular locations revealed a conservation
of topography between the OB and
AONpE: relative locations of glomeruli in
the OB along the anterior-posterior, dor-
sal-medial, and medial-lateral axes are
all preserved in the AONpE. The projec-
tion of AONpE neurons to the contralat-
eral OB also occurs in a topographically
conserved manner: tracer injection to
the granule cell layer resulted in retro-
grade labeling of a specific spot in the
contralateral AONpE region. To examine
the OR-specific neuronal connection
through the AONpE, Yan et al. (2008) in-
jected red tracers into the M71 glomeru-
lus and green tracers into the granule
cell layer beneath the contralateral M71
glomerulus. They found colocalization of
red and green dyes in a restricted area in
the AONpE. Thus, the two projections,
one from OB to ipsilateral AONpE and
the other from AONpE to contralateral
OB, link the two corresponding OR-spe-
cific ‘‘isofunctional’’ olfactory columns
with high precision (Figure 1).
The next question was, does this circuit
regulate the neuronal activity of the con-
tralateral OB? Using c-Fos expression as
a measure of activity, Yan et al. (2008) an-
alyzed the odor-evoked neuronal activity
in the OB. Odorant stimuli induced c-Fos
expression in the periglomerular cells
466 Neuron 58, May 22, 2008 ª2008 Elsevie
and granule cells in the OB. When one
nostril was occluded, c-Fos signals were
dramatically reduced on the occluded
side. Interestingly, however, significant
levels of signal were detected in granule
cells. The activities of the underlying
granule cells were caused by excitatory
inputs from the contralateral AONpE.
When AONpE neurons on the contralat-
eral (open) side were killed by isobutenic
acid, c-Fos signals in granule cells on the
occluded side were not observed. These
results indicate that M/T cells activate
the ipsilateral AONpE neurons, which in
turn lead to the excitation of granule cells
in the contralateral OB. This notion was
also confirmed by electrophysiological
recordings of granule cell activities.
In each OB, mirror-symmetrical maps
are formed in the medial and lateral
halves, and reciprocal projections within
the OB link the isotypic olfactory columns
(Lodovichi et al., 2003). Connectivity of
the interbulbar circuitry appears to be
analogous to the intrabulbar circuitry. It
has been found that external tufted cells
(excitatory neurons) project to granule
cells within the isotypic olfactory column
on the other side of the OB (Figure 1).
However, it is still unknown how the acti-
vated granule cells modulate M/T cells,
and how the intrabulbar association
tunes olfactory signals as a whole. Be-
cause granule cells are GABAergic neu-
rons, both intrabulbar and interbulbar
connections appear to be inhibitory.
These systems might be used to syn-
chronize the odor-evoked neuronal activ-
ity and transmit the robust signals to the
OC. Alternatively, interbulbar reciprocal
connections may enhance the difference
in inputs between the two nostrils, allow-
ing the ‘‘stereo’’ sensation of odors. In-
deed, there is a report demonstrating
that bilateral sniffing is essential to sense
the direction of the odor source in rat
(Rajan et al., 2006).
Yan et al. (2008) assume a different
scenario for the function of interbulbar
connection. They examined the functional
requirement of interbulbar circuitry for
the exchange of olfactory information
between hemispheres. It has been re-
ported that unilateral olfactory learning
can be recalled by stimulating either of
the two nostrils, trained or untrained,
and that transection of the anterior
commissure impairs the memory recall
r Inc.
by the untrained nostril (Kucharski and
Hall, 1987). Yan et al. (2008) specifically
eliminated the AONpE neurons by focal
injection of isobutenic acid and assessed
its impact on bilateral communication.
They used a unilateral olfactory learning/
recall paradigm in which both ANOpE-le-
sioned and sham-lesioned mice were
trained to associate footshock with a par-
ticular odor through a single nostril and
then tested for the olfactory memory
either with the trained or untrained nostril.
Sham-lesioned mice could avoid the odor
with either nostril. Even when the AONpE
was lesioned, the mice still successfully
avoided the associated odor with the
trained nostril, suggesting that olfactory
learning and recall in the ipsilateral brain
was not affected by the surgery. In con-
trast, the AONpE-lesioned mice did not
avoid the odor with the untrained nostril.
Because the AONpE neurons project ex-
clusively to the contralateral OB and not
to other brain regions (Davis and Macr-
ides, 1981), these results indicate that
the projection from AONpE to contralat-
eral OB is essential for interhemispheric
communication in the olfactory learning
and/or recall. This finding is quite amaz-
ing; however, one must be cautious in
interpreting the data of surgical lesions.
In the vicinity of the AONpE, other sub-
regions are closely packed, some of
which also project to the contralateral
hemisphere (Davis and Macrides, 1981).
Therefore, one cannot exclude the possi-
bility that the AONpE lesion may have
also damaged other subregions, which
might instead mediate the learning and/
or memory recall between the two hemi-
spheres. To address these issues, spe-
cific elimination or silencing of AONpE
neurons using genetic methods (e.g., Ko-
bayakawa et al., 2007) will be useful for
future studies.
The present study by Yan et al. (2008)
also raises some important questions. If
the interbulbar circuitry indeed mediates
the learning and/or recall, does the activa-
tion of granule cells in the contralateral OB
elicit the memory? In order to learn or to
recall the olfactory memory in the brain,
regulation of M/T cells appears to be es-
sential, because they are the only projec-
tion neurons in the OB. Yan et al. (2008) in-
deed find that the neuronal activities of
some M/T cells are changed (either ex-
cited or inhibited) by contralateral inputs.

Neuron
Previews
However, it is puzzling that the activation
of the underlying GABAergic granule cells
leads to such changes. Future studies
using genetic methods that label a specific
subset of neurons or stimulate them by
light-gated ion channels will help describe
the local neural circuitry in detail.
Interbulbar circuit formation is an in-
triguing issue in developmental neurobi-
ology. Recent studies have shed light
on the molecular basis of olfactory map
formation in the OB: OR-derived cAMP
signals direct the axonal projection of
OSNs by regulating the gene expression
of axon guidance/sorting molecules (re-
viewed by Imai and Sakano, 2007). Spon-
taneous signaling from ORs, rather than
odor-evoked activity, appears to be im-
portant in mammalian OSN projection,
whereas the intrabulbar projection of
external tufted cells is highly dependent
on neuronal activity (Marks et al., 2006).
Although the intrabulbar connection is
monosynaptic, the interbulbar circuit de-
scribed in the present study is disynaptic,
which may require more complicated
processing during development. The
commissural fibers play an important
Au Naturel
Garrett B. Stanley1,*1Coulter Department of Biomedical Engineerin*Correspondence: [email protected] 10.1016/j.neuron.2008.05.003
Although adaptation is a ubiquitousquences in the natural visual enviroa comprehensive set of in vivo expof adaptation that have been well stenvironment and are predictable froensembles.
Nature does nothing uselessly.
—Aristotle, 384–322 BC
It is this compelling idea that has driven
neuroscientists for decades to ponder the
evolution, development, and function of
the brain in the context of the natural envi-
role in exchanging higher-order informa-
tion between hemispheres, and studies
on the interbulbar circuitry will provide
new insights into the molecular mecha-
nisms of the precise wiring between
hemispheres.
Another issue raised in the present
study is the interhemispheric exchange
of olfactory information. The reported
failure of interhemispheric communica-
tion in the behavioral experiments on the
AONpE-lesioned mice may be due to the
inability to either ‘‘form’’ the memory or
to ‘‘transfer’’ it to the contralateral hemi-
sphere. Alternatively, it may be due to
the deficit of the ‘‘recall’’ of memory
stored in the contralateral hemisphere.
These possibilities are not mutually exclu-
sive and can be dissected and tested with
genetic tools to silence the AON in a re-
versible manner. It is still not well under-
stood where and how olfactory memory
is stored and what kinds of neuronal activ-
ities lead to learning and recall of olfactory
information. The interbulbar circuitry de-
scribed in the present study will continue
to serve as an excellent tool for the study
of olfactory memory.
g, Georgia Institute of Technology, and Emoryh.edu
property of neurons in the early visnment are unknown. In this issue oferiments in the visual thalamus, thaudied with artificial probes capture tm properties of the visual scene that
ronment within which we exist. Simply
put, to understand the brain, we cannot
ignore our surroundings. Although the
key to the mysteries of the endless com-
plexity of the anatomy and function may
indeed lie at the interface between the
individual and the world, scientific explo-
Neuro
REFERENCES
Buck, L., and Axel, R. (1991). Cell 65, 175–187.
Davis, B.J., and Macrides, F. (1981). J. Comp.Neurol. 203, 475–493.
Imai, T., and Sakano, H. (2007). Curr. Opin. Neuro-biol. 17, 507–515.
Kobayakawa, K., Kobayakawa, R., Matsumoto, H.,Oka, Y., Imai, T., Ikawa, M., Okabe, M., Ikeda, T.,Itohara, S., Kikusui, T., et al. (2007). Nature 450,503–508.
Kucharski, D., and Hall, W.G. (1987). Science 238,786–788.
Lodovichi, C., Belluscio, L., and Katz, L.C. (2003).Neuron 38, 265–276.
Marks, C.A., Cheng, K., Cummings, D.M., and Bel-luscio, L. (2006). J. Neurosci. 26, 11257–11266.
Mori, K., Takahashi, Y.K., Igarashi, K.M., andYamaguchi, M. (2006). Physiol. Rev. 86, 409–433.
Rajan, R., Clement, J.P., and Bhalla, U.S. (2006).Science 311, 666–670.
Serizawa, S., Miyamichi, K., and Sakano, H. (2004).Trends Genet. 20, 648–653.
Wilson, R.I., and Mainen, Z.F. (2006). Annu. Rev.Neurosci. 29, 163–201.
Yan, Z., Tan, J., Qin, C., Lu, Y., Ding, C., and Luo, M.(2008). Neuron 58, this issue, 613–624.
University, Atlanta, GA 30332, USA
ual pathway, the functional conse-Neuron, Mante et al. show, throught the basic functional mechanismshe neuronal response in the naturalmay be represented by local neural
ration of the brain from this perspective
is a vexing task. The idea does not lend
itself well to carefully controlled experi-
ments that normally constitute scientific
investigation. Nevertheless, confronting
this issue may help us move from what
can the brain do? to what does the brain
n 58, May 22, 2008 ª2008 Elsevier Inc. 467