adult generation of glutamatergic olfactory bulb interneurons
TRANSCRIPT

©20
09 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.
1524 VOLUME 12 | NUMBER 12 | dEcEMBER 2009 nature neurOSCIenCe
a r t I C l e S
The presence of neural stem cells (NSCs) in the adult mammalian brain has prompted hope for replacement of degenerating neurons. This potential avenue for repair has been further supported by the observation that neuroblasts originating from these NSCs are recruited to sites of injury1,2. Adult neurogenesis occurs largely in two regions of the forebrain that generate distinct types of neurons: the subgranu-lar zone generating glutamatergic projection neurons of the dentate gyrus and the SEZ generating GABAergic interneurons of the olfac-tory bulb3. The potential of these regions to contribute to endogenous repair may depend on the range of neuronal subtypes that they can produce. Although earlier transplantation studies have proposed a broad potential of NSCs in regard to the generation of different neu-ronal subtypes4, this view has recently been challenged by the demon-stration that there are distinct NSCs with restricted potential regarding subtype specification5. On the other hand, it has been suggested that apoptotic cell death in the cerebral cortex elicits the migration of neuroblasts from the SEZ with the ability to replace glutamatergic projection neurons6,7. Given that SEZ cells have so far been thought to generate only GABAergic neurons, this recruitment either suggests a high degree of plasticity of adult SEZ cells or, alternatively, there might be a yet undefined source of glutamatergic neurons in this region.
NSCs in the adult SEZ generate GABAergic interneurons inte-grating into the granule cell layer (GCL) or glomerular layer of the olfactory bulb8. A large part of these interneurons arise from the lat-eral SEZ, which derives from the ventral telencephalon, producing mostly telencephalic GABAergic neurons during development8,9. Transcription factors that are crucial for the generation of GABAergic interneurons during development, such as Dlx10 and Sp8 (ref. 11), continue to determine the postnatal and adult generation of olfactory bulb interneurons10,11. Transcription factors that are present in the dorsal telencephalon during development, such as Pax6 or Neurog2 (ref. 12), are largely restricted in their expression to the dorsal region of the adult10,13 or postnatal SEZ14. Fate-mapping experiments have shown that the adult dorsal SEZ originates from the Emx1-positive area of the embryonic telencephalon and participates in the genera-tion of specific subtypes of GABAergic olfactory neurons, such as the dopaminergic periglomerular neurons15,16.
However, during embryonic development, dorsal progenitors generate predominantly glutamatergic neurons in a Pax6-dependent manner, including those of the olfactory bulb17,18. Thus, the entire population of olfactory bulb projection neurons is thought to derive from a Pax6-expressing territory19. In the developing cerebral cortex,
1Department of Physiological Genomics, Institute of Physiology, Ludwig-Maximilians University Munich, Munich, Germany. 2Institute for Stem Cell Research, Helmholtz Zentrum München, German Research Center for Environmental Health, Neuherberg, Germany. 3Munich Center for Integrated Protein Science CiPSM, Munich, Germany. 4Cambridge Centre for Brain Repair, Robinson Way, Cambridge, UK. 5Departments of Neurological Surgery and Pathology, University of Washington, Seattle Children’s Hospital Research Institute, Seattle, Washington, USA. 6Laboratory of Molecular Biology and Genetics, Institute of Experimental Medicine, Budapest, Hungary. 7Division of Molecular Neurobiology, Medical Research Council—National Institute for Medical Research, London, UK. 8Insitute of Anatomy and Cell Biology, University of Freiburg, Freiburg, Germany. 9Present address: Brain Research Institute, University of Zürich/ETH, Zürich, Switzerland (O.R.), INSERM U711, Biologie des Interactions Neurones/Glie, Hôpital de la Pitié-Salpêtrière, Bâtiment de la Pharmacie, Paris, France (C.P.). 10These authors contributed equally to this work. Correspondence should be addressed to M.G. ([email protected]), O.R. ([email protected]) or R.F.H. ([email protected]).
Received 15 June; accepted 8 September; published online 1 November 2009; corrected after print 11 December 2009; doi:10.1038/nn.2416
Adult generation of glutamatergic olfactory bulb interneuronsMonika S Brill1–3,10, Jovica Ninkovic2,3,10, Eleanor Winpenny4,10, Rebecca D Hodge5,10, Ilknur Ozen4, Roderick Yang5, Alexandra Lepier1, Sergio Gascón1,2, Ferenc Erdelyi6, Gabor Szabo6, Carlos Parras7,9, Francois Guillemot7, Michael Frotscher8, Benedikt Berninger1,2, Robert F Hevner5, Olivier Raineteau4,9 & Magdalena Götz1–3
The adult mouse subependymal zone (SEZ) harbors neural stem cells that are thought to exclusively generate GABAergic interneurons of the olfactory bulb. We examined the adult generation of glutamatergic juxtaglomerular neurons, which had dendritic arborizations that projected into adjacent glomeruli, identifying them as short-axon cells. Fate mapping revealed that these originate from Neurog2- and Tbr2-expressing progenitors located in the dorsal region of the SEZ. Examination of the progenitors of these glutamatergic interneurons allowed us to determine the sequential expression of transcription factors in these cells that are thought to be hallmarks of glutamatergic neurogenesis in the developing cerebral cortex and adult hippocampus. Indeed, the molecular specification of these SEZ progenitors allowed for their recruitment into the cerebral cortex after a lesion was induced. Taken together, our data indicate that SEZ progenitors not only produce a population of adult-born glutamatergic juxtaglomerular neurons, but may also provide a previously unknown source of progenitors for endogenous repair.

©20
09 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.
nature neurOSCIenCe VOLUME 12 | NUMBER 12 | dEcEMBER 2009 1525
a r t I C l e S
Pax6 regulates Neurog1 and Neurog2 expression and these tran-scription factors are important to specify cortical neurons toward a glutamatergic fate17,18,20. In addition, the transcription factors Tbr1 and Tbr2 (also known as Eomes) are expressed in early postmitotic glutamatergic neurons and their intermediate progenitors21. Given the glutamatergic progeny of the dorsal telencephalon during devel-opment and its contribution to the adult SEZ, we searched for and found progenitors generating glutamatergic neurons in the adult dorsal SEZ.
RESULTSNeurog2 and Tbr2 in a subset of dorsal SEZ progenitorsWe detected Neurog2- and Tbr2-immunoreactive cells in a dorsal sub-region of the SEZ and the rostral migratory stream (RMS) (Fig. 1a–d and Supplementary Fig. 1), whereas Pax6- and Mash1 (also known as Ascl1)-expressing progenitors were distributed in a more wide-spread manner (Supplementary Fig. 2). Consistent with their similar localization, most Neurog2-expressing cells (green fluorescent protein (GFP)-expressing cells in mice in which GFP had been inserted into the Neurog2 locus, referred to Neurog2+/gfp; Supplementary Fig. 1) were either Tbr2 or Tbr1 immunopositive (Tbr2, 96%, 394 Neurog2+/gfp cells; Tbr1, 70%, 151 Neurog2+/gfp cells; Fig. 1e,f). To further examine their identity, we also stained for doublecortin (Dcx), a protein that is exclusively present in neuroblasts and young postmitotic neurons. The majority of Neurog2+/gfp-positive (80 ± 4%, 212 Neurog2+/gfp cells), Tbr2-positive (70 ± 8%, 255 cells) and Tbr1-positive (97 ± 2%, 265 cells) cells were also Dcx positive (Fig. 2a–c). To distinguish postmitotic neuroblasts from proliferating neuronal progenitors, we stained for Ki67, a protein that is present in dividing cells, or provided the DNA-base analog BrdU, which is incorporated in proliferating cells. Tbr1-positive cells lacked Ki67 immunoreactivity and did not incorporate BrdU within a few hours of injection (1 ± 1%, 201 cells; Fig. 2), suggesting that virtually all of the Tbr1-positive cells were post-mitotic neurons. Conversely, 37% of the GFP-positive cells (156 cells) colocalized with Ki67 and 22% were labeled with BrdU (Fig. 2d,g), indicating that one-third of these cells comprise an actively divid-ing progenitor population. Likewise, a small subset of Tbr2-positive cells incorporated BrdU (Fig. 2e,g). During development, Tbr2 is mostly contained in progenitors undergoing a last round of cell divi-sion, whereas Tbr1 is present in postmitotic neurons21,22. To examine whether Tbr1-positive cells arise from proliferating progenitors, we injected BrdU intraperitoneally and waited for 3 d. A quarter of the Tbr1-positive cells labeled with BrdU (Fig. 2g), consistent with Tbr1 being expressed when cells become postmitotic. To ensure that Tbr1-positive neuroblasts originated from Tbr2-positive progenitors, we used Tbr2BAC-gfp mice23 (Supplementary Fig. 3), which allowed us to carry out short-term fate mapping, as GFP persists for longer than
Tbr2 (ref. 24). Indeed, almost all of the Tbr1-immunoreactive cells in the RMS were GFP positive (Supplementary Fig. 3), consistent with the idea that they originate from Tbr2-expressing progenitors.
Notably, none of these transcription factors (Neurog2, Tbr2 or Tbr1) were ever colocalized with GFAP, a protein that is characteristically expressed in astroglia, including NSCs (data not shown). However, we determined that the Tbr-expressing cells were derived from astrocyte-like NSCs by genetic fate mapping with GLAST::creERT2 mediating recombination in GLAST-positive NSCs25 (Supplementary Fig. 4). Consistent with the recombination frequency observed on tamoxifen-induced recombination25,26, more than 60% of the Tbr2-positive cells were GFP positive, that is, derived from astrocyte-like NSCs (64% of the Tbr2-positive cells were Tbr2 and GFP double positive, 104 cells). Thus, an adult, NSC-derived subpopulation of actively dividing pro-genitors and young neuroblasts in the SEZ and RMS express a series of transcription factors that are normally indicative of a glutamatergic neuronal lineage.
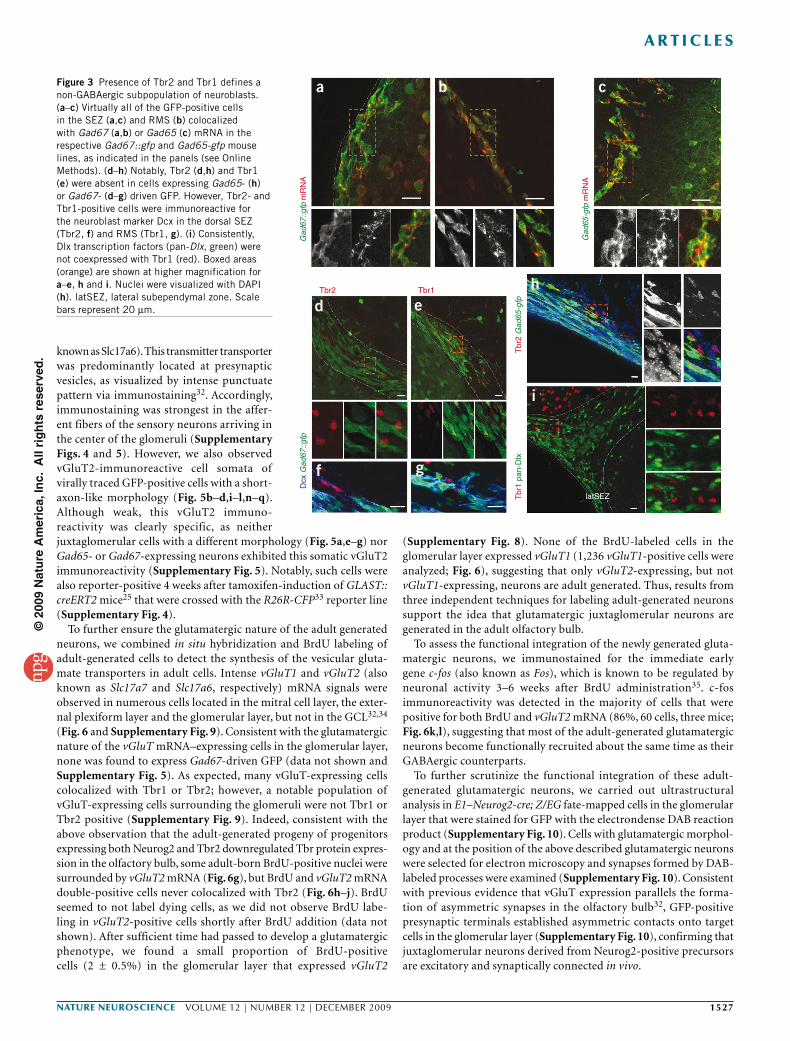
To determine whether Tbr1- or Tbr2-positive neuroblasts may nev-ertheless have GABAergic traits, we analyzed Dcx-positive neuroblasts in the RMS. Although most neuroblasts already express GAD, synthe-size GABA27 and are GFP positive in Gad67 (also known as Gad1)::gfp28 and Gad65 (also known as Gad2)-gfp29 mice in which GFP protein expression is confined to cells expressing Gad67 or Gad65 mRNA (Fig. 3a–c), Tbr1 and Tbr2 double-positive cells define a neuroblast subset that lacks both Gad67- (Fig. 3d–g) and Gad65-driven GFP (Fig. 3h). Consistently, Tbr-positive cells did not contain Dlx (Fig. 3i). Moreover, also in the olfactory bulb, Tbr2-positive cells did not colocalize with any GABAergic interneuron markers, such as tyrosine hydroxylase, calbindin or calretinin (Supplementary Fig. 5).
GFP
LV Third ventricle
300
d e
f
200
100
0GFP positive
Num
ber
of c
ells
per
50-µ
mco
rona
l sec
tion
Tbr2 positive LV
Dorsal
Lateral
Rostral
Ctx Ctx Ctx
LV
LV
LV
Str
RMS
RM
S
RMS
Dcx
GFP Tbr2
GFP Tbr1
Tbr2 Tbr1a b cFigure 1 Neurog2+/gfp, Tbr1 and Tbr2 are expressed in a defined dorsal region of the SEZ. (a–c) Fluorescent micrographs depicting Neurog2+/gfp-, Tbr2- and Tbr1-positive cells as indicated in the dorsal SEZ and RMS. (d) Three-dimensional reconstruction of the localization of Neurog2+/gfp- (green) and Tbr2-positive (red) cells in the adult forebrain indicates that these progenitors are located in the dorsal SEZ, but not in the lateral wall of the lateral ventricle or third ventricle. Coronal views of three different levels are indicated by the black lines. The histogram below depicts the distribution of Neurog2+/gfp- and Tbr2-positive cells along the rostro-caudal axis. Tbr2-positive cells were more widespread than Neurog2+/gfp-positive cells and the expression of both markers dropped rapidly after the cells entered the RMS. (e,f) Neurog2+/gfp cells are immunoreactive for Tbr2 (e) and Tbr1 (f). Nuclei are visualized with DAPI (e,f). Ctx, cortex; LV, lateral ventricle; Str, striatum. Scale bars represent 20 µm.
©20
09 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.
1526 VOLUME 12 | NUMBER 12 | dEcEMBER 2009 nature neurOSCIenCe
a r t I C l e S
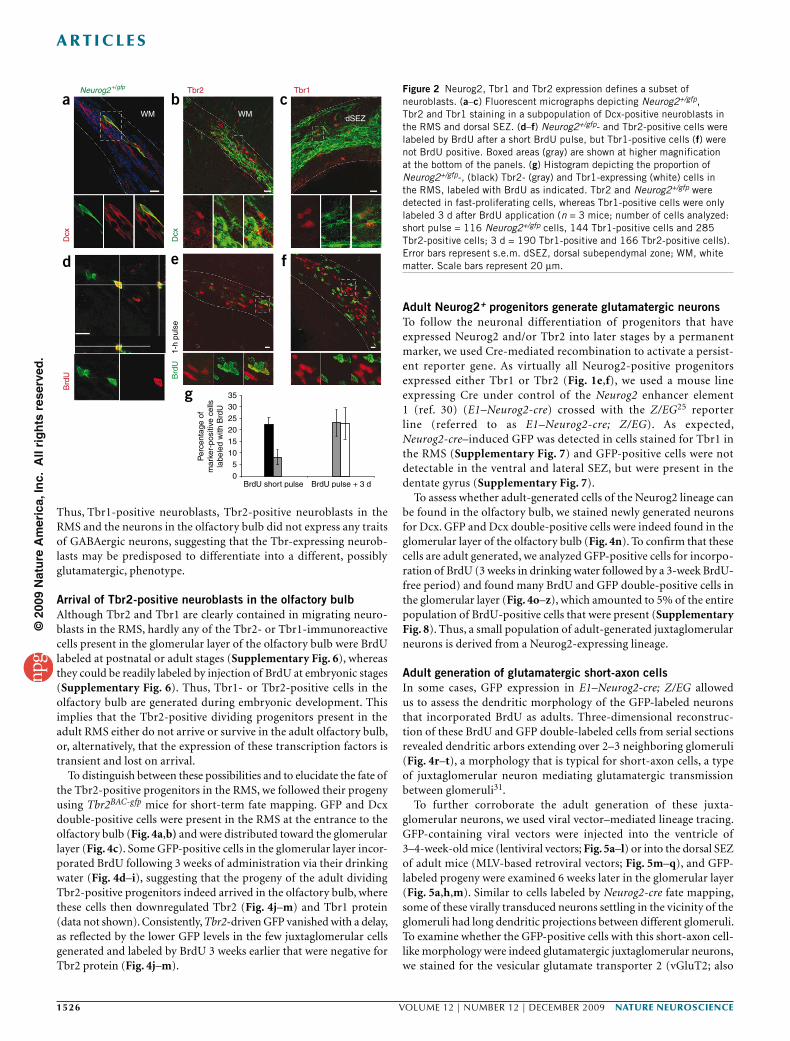
Thus, Tbr1-positive neuroblasts, Tbr2-positive neuroblasts in the RMS and the neurons in the olfactory bulb did not express any traits of GABAergic neurons, suggesting that the Tbr-expressing neurob-lasts may be predisposed to differentiate into a different, possibly glutamatergic, phenotype.
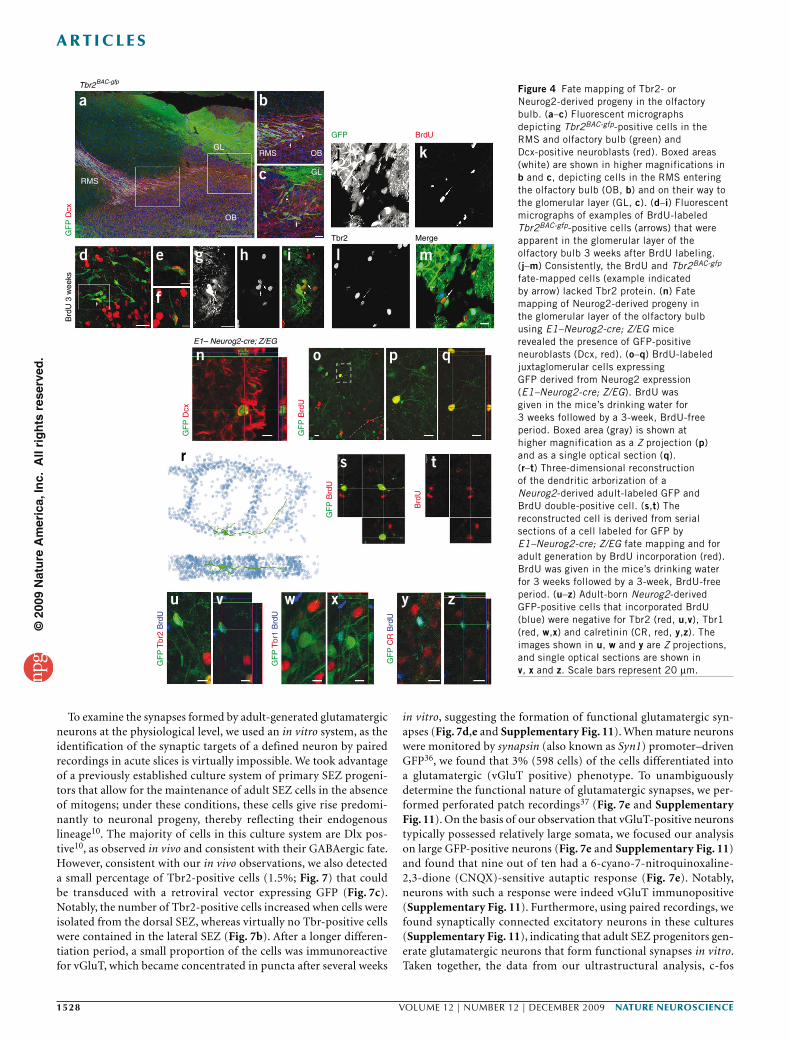
Arrival of Tbr2-positive neuroblasts in the olfactory bulb Although Tbr2 and Tbr1 are clearly contained in migrating neuro-blasts in the RMS, hardly any of the Tbr2- or Tbr1-immunoreactive cells present in the glomerular layer of the olfactory bulb were BrdU labeled at postnatal or adult stages (Supplementary Fig. 6), whereas they could be readily labeled by injection of BrdU at embryonic stages (Supplementary Fig. 6). Thus, Tbr1- or Tbr2-positive cells in the olfactory bulb are generated during embryonic development. This implies that the Tbr2-positive dividing progenitors present in the adult RMS either do not arrive or survive in the adult olfactory bulb, or, alternatively, that the expression of these transcription factors is transient and lost on arrival.
To distinguish between these possibilities and to elucidate the fate of the Tbr2-positive progenitors in the RMS, we followed their progeny using Tbr2BAC-gfp mice for short-term fate mapping. GFP and Dcx double-positive cells were present in the RMS at the entrance to the olfactory bulb (Fig. 4a,b) and were distributed toward the glomerular layer (Fig. 4c). Some GFP-positive cells in the glomerular layer incor-porated BrdU following 3 weeks of administration via their drinking water (Fig. 4d–i), suggesting that the progeny of the adult dividing Tbr2-positive progenitors indeed arrived in the olfactory bulb, where these cells then downregulated Tbr2 (Fig. 4j–m) and Tbr1 protein (data not shown). Consistently, Tbr2-driven GFP vanished with a delay, as reflected by the lower GFP levels in the few juxtaglomerular cells generated and labeled by BrdU 3 weeks earlier that were negative for Tbr2 protein (Fig. 4j–m).
Adult Neurog2+ progenitors generate glutamatergic neuronsTo follow the neuronal differentiation of progenitors that have expressed Neurog2 and/or Tbr2 into later stages by a permanent marker, we used Cre-mediated recombination to activate a persist-ent reporter gene. As virtually all Neurog2-positive progenitors expressed either Tbr1 or Tbr2 (Fig. 1e,f), we used a mouse line expressing Cre under control of the Neurog2 enhancer element 1 (ref. 30) (E1–Neurog2-cre) crossed with the Z/EG25 reporter line (referred to as E1–Neurog2-cre; Z/EG). As expected, Neurog2-cre–induced GFP was detected in cells stained for Tbr1 in the RMS (Supplementary Fig. 7) and GFP-positive cells were not detectable in the ventral and lateral SEZ, but were present in the dentate gyrus (Supplementary Fig. 7).
To assess whether adult-generated cells of the Neurog2 lineage can be found in the olfactory bulb, we stained newly generated neurons for Dcx. GFP and Dcx double-positive cells were indeed found in the glomerular layer of the olfactory bulb (Fig. 4n). To confirm that these cells are adult generated, we analyzed GFP-positive cells for incorpo-ration of BrdU (3 weeks in drinking water followed by a 3-week BrdU-free period) and found many BrdU and GFP double-positive cells in the glomerular layer (Fig. 4o–z), which amounted to 5% of the entire population of BrdU-positive cells that were present (Supplementary Fig. 8). Thus, a small population of adult-generated juxtaglomerular neurons is derived from a Neurog2-expressing lineage.
Adult generation of glutamatergic short-axon cellsIn some cases, GFP expression in E1–Neurog2-cre; Z/EG allowed us to assess the dendritic morphology of the GFP-labeled neurons that incorporated BrdU as adults. Three-dimensional reconstruc-tion of these BrdU and GFP double-labeled cells from serial sections revealed dendritic arbors extending over 2–3 neighboring glomeruli (Fig. 4r–t), a morphology that is typical for short-axon cells, a type of juxtaglomerular neuron mediating glutamatergic transmission between glomeruli31.
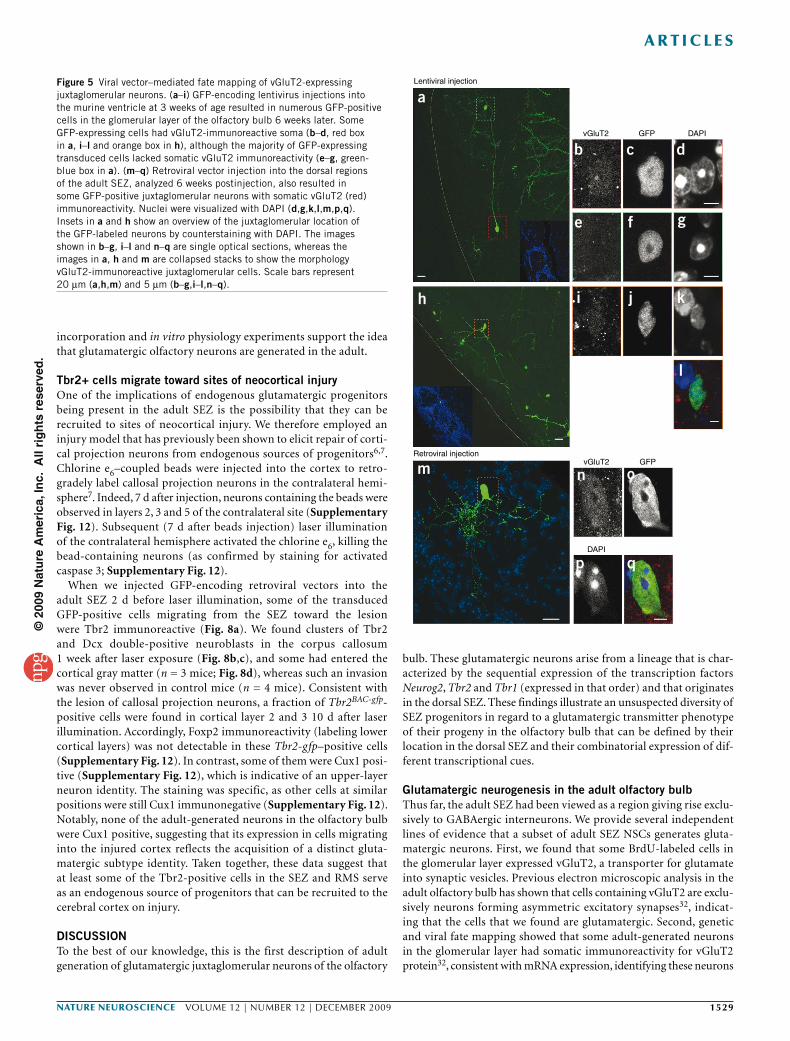
To further corroborate the adult generation of these juxta-glomerular neurons, we used viral vector–mediated lineage tracing. GFP-containing viral vectors were injected into the ventricle of 3–4-week-old mice (lentiviral vectors; Fig. 5a–l) or into the dorsal SEZ of adult mice (MLV-based retroviral vectors; Fig. 5m–q), and GFP-labeled progeny were examined 6 weeks later in the glomerular layer (Fig. 5a,h,m). Similar to cells labeled by Neurog2-cre fate mapping, some of these virally transduced neurons settling in the vicinity of the glomeruli had long dendritic projections between different glomeruli. To examine whether the GFP-positive cells with this short-axon cell-like morphology were indeed glutamatergic juxtaglomerular neurons, we stained for the vesicular glutamate transporter 2 (vGluT2; also
Neurog2+/gfp Tbr2
Dcx
Brd
U Brd
U1-
h pu
lse
Dcx
WM WM dSEZ
Tbr1
35
BrdU short pulse BrdU pulse + 3 d
30
25
20
1510
50
Per
cent
age
ofm
arke
r-po
sitiv
e ce
llsla
bele
d w
ith B
rdU
a
d e
g
f
b cFigure 2 Neurog2, Tbr1 and Tbr2 expression defines a subset of neuroblasts. (a–c) Fluorescent micrographs depicting Neurog2+/gfp, Tbr2 and Tbr1 staining in a subpopulation of Dcx-positive neuroblasts in the RMS and dorsal SEZ. (d–f) Neurog2+/gfp- and Tbr2-positive cells were labeled by BrdU after a short BrdU pulse, but Tbr1-positive cells (f) were not BrdU positive. Boxed areas (gray) are shown at higher magnification at the bottom of the panels. (g) Histogram depicting the proportion of Neurog2+/gfp-, (black) Tbr2- (gray) and Tbr1-expressing (white) cells in the RMS, labeled with BrdU as indicated. Tbr2 and Neurog2+/gfp were detected in fast-proliferating cells, whereas Tbr1-positive cells were only labeled 3 d after BrdU application (n = 3 mice; number of cells analyzed: short pulse = 116 Neurog2+/gfp cells, 144 Tbr1-positive cells and 285 Tbr2-positive cells; 3 d = 190 Tbr1-positive and 166 Tbr2-positive cells). Error bars represent s.e.m. dSEZ, dorsal subependymal zone; WM, white matter. Scale bars represent 20 µm.
©20
09 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.
nature neurOSCIenCe VOLUME 12 | NUMBER 12 | dEcEMBER 2009 1527
a r t I C l e S
known as Slc17a6). This transmitter transporter was predominantly located at presynaptic vesicles, as visualized by intense punctuate pattern via immunostaining32. Accordingly, immunostaining was strongest in the affer-ent fibers of the sensory neurons arriving in the center of the glomeruli (Supplementary Figs. 4 and 5). However, we also observed vGluT2-immunoreactive cell somata of virally traced GFP-positive cells with a short-axon-like morphology (Fig. 5b–d,i–l,n–q). Although weak, this vGluT2 immuno-reactivity was clearly specific, as neither juxtaglomerular cells with a different morphology (Fig. 5a,e–g) nor Gad65- or Gad67-expressing neurons exhibited this somatic vGluT2 immunoreactivity (Supplementary Fig. 5). Notably, such cells were also reporter-positive 4 weeks after tamoxifen-induction of GLAST::creERT2 mice25 that were crossed with the R26R-CFP33 reporter line (Supplementary Fig. 4).
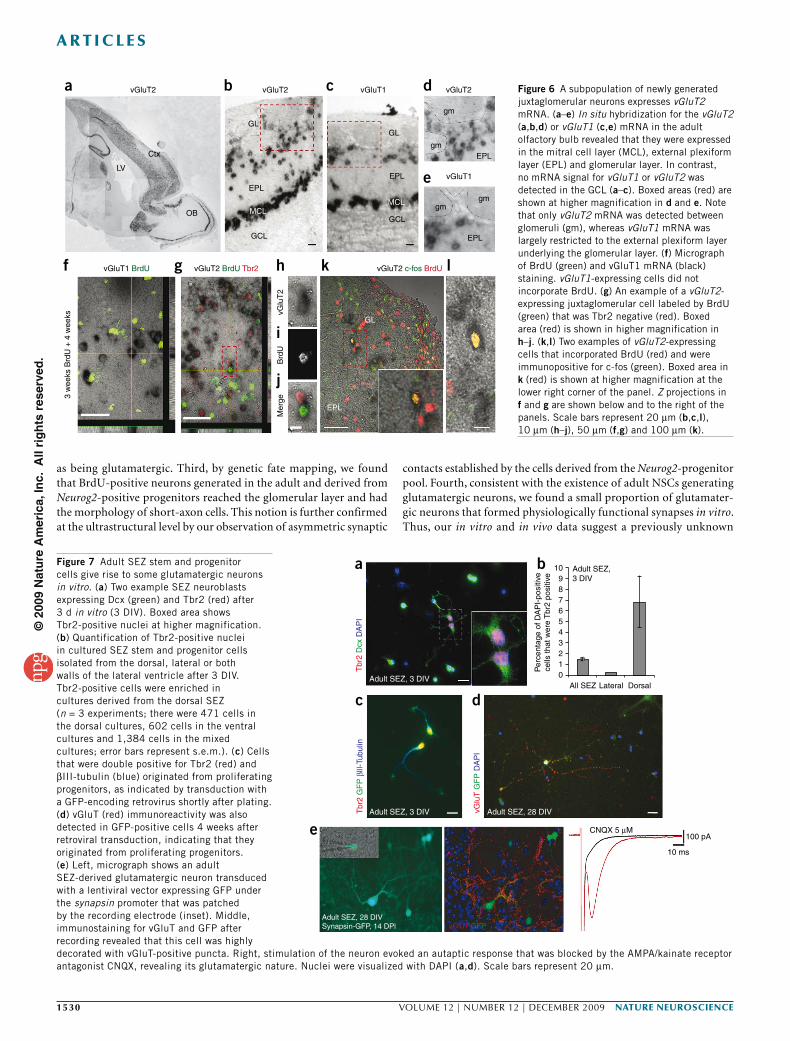
To further ensure the glutamatergic nature of the adult generated neurons, we combined in situ hybridization and BrdU labeling of adult-generated cells to detect the synthesis of the vesicular gluta-mate transporters in adult cells. Intense vGluT1 and vGluT2 (also known as Slc17a7 and Slc17a6, respectively) mRNA signals were observed in numerous cells located in the mitral cell layer, the exter-nal plexiform layer and the glomerular layer, but not in the GCL32,34 (Fig. 6 and Supplementary Fig. 9). Consistent with the glutamatergic nature of the vGluT mRNA–expressing cells in the glomerular layer, none was found to express Gad67-driven GFP (data not shown and Supplementary Fig. 5). As expected, many vGluT-expressing cells colocalized with Tbr1 or Tbr2; however, a notable population of vGluT-expressing cells surrounding the glomeruli were not Tbr1 or Tbr2 positive (Supplementary Fig. 9). Indeed, consistent with the above observation that the adult-generated progeny of progenitors expressing both Neurog2 and Tbr2 downregulated Tbr protein expres-sion in the olfactory bulb, some adult-born BrdU-positive nuclei were surrounded by vGluT2 mRNA (Fig. 6g), but BrdU and vGluT2 mRNA double-positive cells never colocalized with Tbr2 (Fig. 6h–j). BrdU seemed to not label dying cells, as we did not observe BrdU labe-ling in vGluT2-positive cells shortly after BrdU addition (data not shown). After sufficient time had passed to develop a glutamatergic phenotype, we found a small proportion of BrdU-positive cells (2 ± 0.5%) in the glomerular layer that expressed vGluT2
(Supplementary Fig. 8). None of the BrdU-labeled cells in the glomerular layer expressed vGluT1 (1,236 vGluT1-positive cells were analyzed; Fig. 6), suggesting that only vGluT2-expressing, but not vGluT1-expressing, neurons are adult generated. Thus, results from three independent techniques for labeling adult-generated neurons support the idea that glutamatergic juxtaglomerular neurons are generated in the adult olfactory bulb.
To assess the functional integration of the newly generated gluta-matergic neurons, we immunostained for the immediate early gene c-fos (also known as Fos), which is known to be regulated by neuronal activity 3–6 weeks after BrdU administration35. c-fos immunoreactivity was detected in the majority of cells that were positive for both BrdU and vGluT2 mRNA (86%, 60 cells, three mice; Fig. 6k,l), suggesting that most of the adult-generated glutamatergic neurons become functionally recruited about the same time as their GABAergic counterparts.
To further scrutinize the functional integration of these adult- generated glutamatergic neurons, we carried out ultrastructural analysis in E1–Neurog2-cre; Z/EG fate-mapped cells in the glomerular layer that were stained for GFP with the electrondense DAB reaction product (Supplementary Fig. 10). Cells with glutamatergic morphol-ogy and at the position of the above described glutamatergic neurons were selected for electron microscopy and synapses formed by DAB-labeled processes were examined (Supplementary Fig. 10). Consistent with previous evidence that vGluT expression parallels the forma-tion of asymmetric synapses in the olfactory bulb32, GFP-positive presynaptic terminals established asymmetric contacts onto target cells in the glomerular layer (Supplementary Fig. 10), confirming that juxtaglomerular neurons derived from Neurog2-positive precursors are excitatory and synaptically connected in vivo.
Gad
67::g
fp m
RN
A
Gad
65-g
fp m
RN
A
Tbr1Tbr2
Tbr
1 pa
n-D
lx
latSEZ
Tbr
2 G
ad65
-gfp
Dcx
Gad
67::g
fp
a
d eh
i
f g
b cFigure 3 Presence of Tbr2 and Tbr1 defines a non-GABAergic subpopulation of neuroblasts. (a–c) Virtually all of the GFP-positive cells in the SEZ (a,c) and RMS (b) colocalized with Gad67 (a,b) or Gad65 (c) mRNA in the respective Gad67::gfp and Gad65-gfp mouse lines, as indicated in the panels (see Online Methods). (d–h) Notably, Tbr2 (d,h) and Tbr1 (e) were absent in cells expressing Gad65- (h) or Gad67- (d–g) driven GFP. However, Tbr2- and Tbr1-positive cells were immunoreactive for the neuroblast marker Dcx in the dorsal SEZ (Tbr2, f) and RMS (Tbr1, g). (i) Consistently, Dlx transcription factors (pan-Dlx, green) were not coexpressed with Tbr1 (red). Boxed areas (orange) are shown at higher magnification for a–e, h and i. Nuclei were visualized with DAPI (h). latSEZ, lateral subependymal zone. Scale bars represent 20 µm.
©20
09 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.
1528 VOLUME 12 | NUMBER 12 | dEcEMBER 2009 nature neurOSCIenCe
a r t I C l e S
To examine the synapses formed by adult-generated glutamatergic neurons at the physiological level, we used an in vitro system, as the identification of the synaptic targets of a defined neuron by paired recordings in acute slices is virtually impossible. We took advantage of a previously established culture system of primary SEZ progeni-tors that allow for the maintenance of adult SEZ cells in the absence of mitogens; under these conditions, these cells give rise predomi-nantly to neuronal progeny, thereby reflecting their endogenous lineage10. The majority of cells in this culture system are Dlx pos-tive10, as observed in vivo and consistent with their GABAergic fate. However, consistent with our in vivo observations, we also detected a small percentage of Tbr2-positive cells (1.5%; Fig. 7) that could be transduced with a retroviral vector expressing GFP (Fig. 7c). Notably, the number of Tbr2-positive cells increased when cells were isolated from the dorsal SEZ, whereas virtually no Tbr-positive cells were contained in the lateral SEZ (Fig. 7b). After a longer differen-tiation period, a small proportion of the cells was immunoreactive for vGluT, which became concentrated in puncta after several weeks
in vitro, suggesting the formation of functional glutamatergic syn-apses (Fig. 7d,e and Supplementary Fig. 11). When mature neurons were monitored by synapsin (also known as Syn1) promoter–driven GFP36, we found that 3% (598 cells) of the cells differentiated into a glutamatergic (vGluT positive) phenotype. To unambiguously determine the functional nature of glutamatergic synapses, we per-formed perforated patch recordings37 (Fig. 7e and Supplementary Fig. 11). On the basis of our observation that vGluT-positive neurons typically possessed relatively large somata, we focused our analysis on large GFP-positive neurons (Fig. 7e and Supplementary Fig. 11) and found that nine out of ten had a 6-cyano-7-nitroquinoxaline-2,3-dione (CNQX)-sensitive autaptic response (Fig. 7e). Notably, neurons with such a response were indeed vGluT immunopositive (Supplementary Fig. 11). Furthermore, using paired recordings, we found synaptically connected excitatory neurons in these cultures (Supplementary Fig. 11), indicating that adult SEZ progenitors gen-erate glutamatergic neurons that form functional synapses in vitro. Taken together, the data from our ultrastructural analysis, c-fos
Tbr2BAC-gfp
a
d e g h i l m
j k
f
b
c
Brd
U 3
wee
ksG
FP
Dcx
n
r
o
s t
p qE1– Neurog2-cre; Z/EG
GF
P D
cx
u v w x y z
GF
P T
br2
Brd
U
GF
P T
br1
Brd
U
GF
P C
R B
rdU
GF
P B
rdU
GF
P B
rdU
Brd
U
RMS
RMSGL
GL
OB
OB
GFP
Tbr2
BrdU
Merge
Figure 4 Fate mapping of Tbr2- or Neurog2-derived progeny in the olfactory bulb. (a–c) Fluorescent micrographs depicting Tbr2BAC-gfp-positive cells in the RMS and olfactory bulb (green) and Dcx-positive neuroblasts (red). Boxed areas (white) are shown in higher magnifications in b and c, depicting cells in the RMS entering the olfactory bulb (OB, b) and on their way to the glomerular layer (GL, c). (d–i) Fluorescent micrographs of examples of BrdU-labeled Tbr2BAC-gfp-positive cells (arrows) that were apparent in the glomerular layer of the olfactory bulb 3 weeks after BrdU labeling. (j–m) Consistently, the BrdU and Tbr2BAC-gfp fate-mapped cells (example indicated by arrow) lacked Tbr2 protein. (n) Fate mapping of Neurog2-derived progeny in the glomerular layer of the olfactory bulb using E1–Neurog2-cre; Z/EG mice revealed the presence of GFP-positive neuroblasts (Dcx, red). (o–q) BrdU-labeled juxtaglomerular cells expressing GFP derived from Neurog2 expression (E1–Neurog2-cre; Z/EG). BrdU was given in the mice’s drinking water for 3 weeks followed by a 3-week, BrdU-free period. Boxed area (gray) is shown at higher magnification as a Z projection (p) and as a single optical section (q). (r–t) Three-dimensional reconstruction of the dendritic arborization of a Neurog2-derived adult-labeled GFP and BrdU double-positive cell. (s,t) The reconstructed cell is derived from serial sections of a cell labeled for GFP by E1–Neurog2-cre; Z/EG fate mapping and for adult generation by BrdU incorporation (red). BrdU was given in the mice’s drinking water for 3 weeks followed by a 3-week, BrdU-free period. (u–z) Adult-born Neurog2-derived GFP-positive cells that incorporated BrdU (blue) were negative for Tbr2 (red, u,v), Tbr1 (red, w,x) and calretinin (CR, red, y,z). The images shown in u, w and y are Z projections, and single optical sections are shown in v, x and z. Scale bars represent 20 µm.
©20
09 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.
nature neurOSCIenCe VOLUME 12 | NUMBER 12 | dEcEMBER 2009 1529
a r t I C l e S
incorporation and in vitro physiology experiments support the idea that glutamatergic olfactory neurons are generated in the adult.
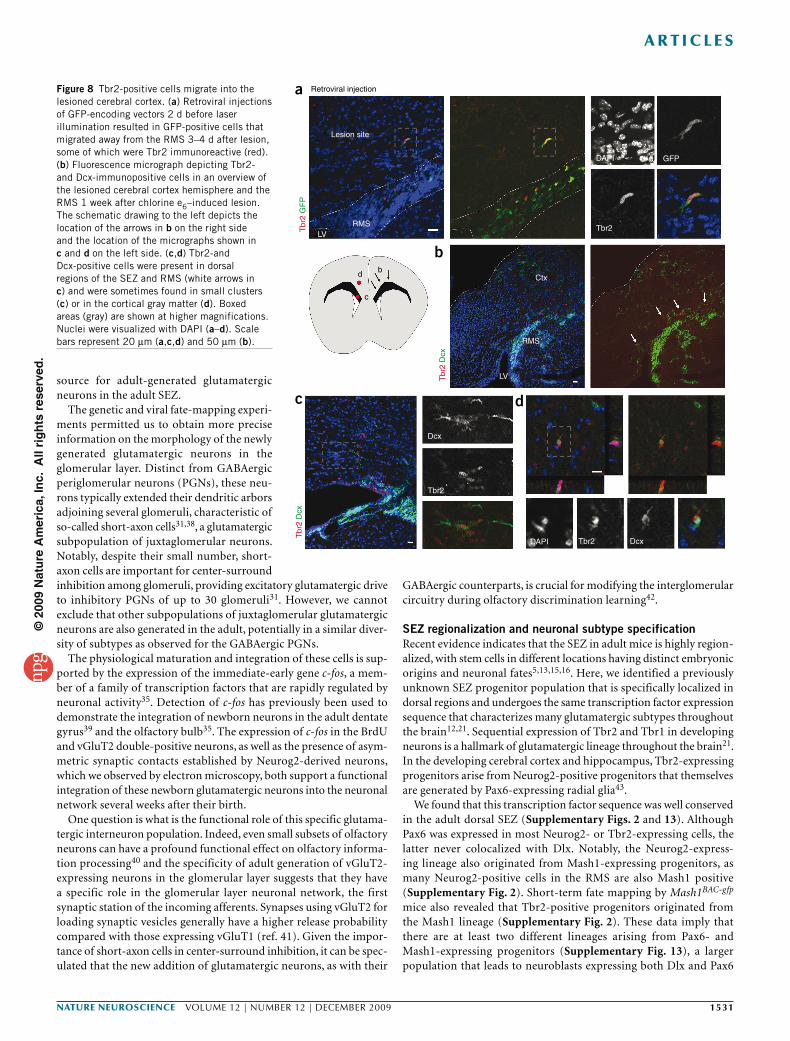
Tbr2+ cells migrate toward sites of neocortical injuryOne of the implications of endogenous glutamatergic progenitors being present in the adult SEZ is the possibility that they can be recruited to sites of neocortical injury. We therefore employed an injury model that has previously been shown to elicit repair of corti-cal projection neurons from endogenous sources of progenitors6,7. Chlorine e6–coupled beads were injected into the cortex to retro-gradely label callosal projection neurons in the contralateral hemi-sphere7. Indeed, 7 d after injection, neurons containing the beads were observed in layers 2, 3 and 5 of the contralateral site (Supplementary Fig. 12). Subsequent (7 d after beads injection) laser illumination of the contralateral hemisphere activated the chlorine e6, killing the bead-containing neurons (as confirmed by staining for activated caspase 3; Supplementary Fig. 12).
When we injected GFP-encoding retroviral vectors into the adult SEZ 2 d before laser illumination, some of the transduced GFP-positive cells migrating from the SEZ toward the lesion were Tbr2 immunoreactive (Fig. 8a). We found clusters of Tbr2 and Dcx double-positive neuroblasts in the corpus callosum 1 week after laser exposure (Fig. 8b,c), and some had entered the cortical gray matter (n = 3 mice; Fig. 8d), whereas such an invasion was never observed in control mice (n = 4 mice). Consistent with the lesion of callosal projection neurons, a fraction of Tbr2BAC-gfp-positive cells were found in cortical layer 2 and 3 10 d after laser illumination. Accordingly, Foxp2 immunoreactivity (labeling lower cortical layers) was not detectable in these Tbr2-gfp–positive cells (Supplementary Fig. 12). In contrast, some of them were Cux1 posi-tive (Supplementary Fig. 12), which is indicative of an upper-layer neuron identity. The staining was specific, as other cells at similar positions were still Cux1 immunonegative (Supplementary Fig. 12). Notably, none of the adult-generated neurons in the olfactory bulb were Cux1 positive, suggesting that its expression in cells migrating into the injured cortex reflects the acquisition of a distinct gluta-matergic subtype identity. Taken together, these data suggest that at least some of the Tbr2-positive cells in the SEZ and RMS serve as an endogenous source of progenitors that can be recruited to the cerebral cortex on injury.
DISCUSSIONTo the best of our knowledge, this is the first description of adult generation of glutamatergic juxtaglomerular neurons of the olfactory
bulb. These glutamatergic neurons arise from a lineage that is char-acterized by the sequential expression of the transcription factors Neurog2, Tbr2 and Tbr1 (expressed in that order) and that originates in the dorsal SEZ. These findings illustrate an unsuspected diversity of SEZ progenitors in regard to a glutamatergic transmitter phenotype of their progeny in the olfactory bulb that can be defined by their location in the dorsal SEZ and their combinatorial expression of dif-ferent transcriptional cues.
Glutamatergic neurogenesis in the adult olfactory bulbThus far, the adult SEZ had been viewed as a region giving rise exclu-sively to GABAergic interneurons. We provide several independent lines of evidence that a subset of adult SEZ NSCs generates gluta-matergic neurons. First, we found that some BrdU-labeled cells in the glomerular layer expressed vGluT2, a transporter for glutamate into synaptic vesicles. Previous electron microscopic analysis in the adult olfactory bulb has shown that cells containing vGluT2 are exclu-sively neurons forming asymmetric excitatory synapses32, indicat-ing that the cells that we found are glutamatergic. Second, genetic and viral fate mapping showed that some adult-generated neurons in the glomerular layer had somatic immunoreactivity for vGluT2 protein32, consistent with mRNA expression, identifying these neurons
Lentiviral injection
a
b c d
e f g
i
n o
p q
h
m
j k
l
Retroviral injection
vGluT2 GFP DAPI
vGluT2 GFP
DAPI
Figure 5 Viral vector–mediated fate mapping of vGluT2-expressing juxtaglomerular neurons. (a–i) GFP-encoding lentivirus injections into the murine ventricle at 3 weeks of age resulted in numerous GFP-positive cells in the glomerular layer of the olfactory bulb 6 weeks later. Some GFP-expressing cells had vGluT2-immunoreactive soma (b–d, red box in a, i–l and orange box in h), although the majority of GFP-expressing transduced cells lacked somatic vGluT2 immunoreactivity (e–g, green-blue box in a). (m–q) Retroviral vector injection into the dorsal regions of the adult SEZ, analyzed 6 weeks postinjection, also resulted in some GFP-positive juxtaglomerular neurons with somatic vGluT2 (red) immunoreactivity. Nuclei were visualized with DAPI (d,g,k,l,m,p,q). Insets in a and h show an overview of the juxtaglomerular location of the GFP-labeled neurons by counterstaining with DAPI. The images shown in b–g, i–l and n–q are single optical sections, whereas the images in a, h and m are collapsed stacks to show the morphology vGluT2-immunoreactive juxtaglomerular cells. Scale bars represent 20 µm (a,h,m) and 5 µm (b–g,i–l,n–q).
©20
09 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.
1530 VOLUME 12 | NUMBER 12 | dEcEMBER 2009 nature neurOSCIenCe
a r t I C l e S
as being glutamatergic. Third, by genetic fate mapping, we found that BrdU-positive neurons generated in the adult and derived from Neurog2-positive progenitors reached the glomerular layer and had the morphology of short-axon cells. This notion is further confirmed at the ultrastructural level by our observation of asymmetric synaptic
contacts established by the cells derived from the Neurog2-progenitor pool. Fourth, consistent with the existence of adult NSCs generating glutamatergic neurons, we found a small proportion of glutamater-gic neurons that formed physiologically functional synapses in vitro. Thus, our in vitro and in vivo data suggest a previously unknown
vGluT2
Ctx
GL
EPL
GL
EPL
gmgm
gm
gm
EPL
EPL
MCL
GL
EPL
MCL
GCL
GCLOB
LV
vGluT1 BrdU
3 w
eeks
Brd
U +
4 w
eeks
Brd
UvG
luT
2M
erge
vGluT2 BrdU Tbr2 vGluT2 c-fos BrdU
vGluT2 vGluT1 vGluT2
vGluT1
a
f g h k l
i
j
b c d
e
Figure 6 A subpopulation of newly generated juxtaglomerular neurons expresses vGluT2 mRNA. (a–e) In situ hybridization for the vGluT2 (a,b,d) or vGluT1 (c,e) mRNA in the adult olfactory bulb revealed that they were expressed in the mitral cell layer (MCL), external plexiform layer (EPL) and glomerular layer. In contrast, no mRNA signal for vGluT1 or vGluT2 was detected in the GCL (a–c). Boxed areas (red) are shown at higher magnification in d and e. Note that only vGluT2 mRNA was detected between glomeruli (gm), whereas vGluT1 mRNA was largely restricted to the external plexiform layer underlying the glomerular layer. (f) Micrograph of BrdU (green) and vGluT1 mRNA (black) staining. vGluT1-expressing cells did not incorporate BrdU. (g) An example of a vGluT2-expressing juxtaglomerular cell labeled by BrdU (green) that was Tbr2 negative (red). Boxed area (red) is shown in higher magnification in h–j. (k,l) Two examples of vGluT2-expressing cells that incorporated BrdU (red) and were immunopositive for c-fos (green). Boxed area in k (red) is shown at higher magnification at the lower right corner of the panel. Z projections in f and g are shown below and to the right of the panels. Scale bars represent 20 µm (b,c,l), 10 µm (h–j), 50 µm (f,g) and 100 µm (k).
vGluT GFPAdult SEZ, 28 DIVSynapsin-GFP, 14 DPl
CNQX 5 µM100 pA
10 ms
e
Tbr
2 D
cx D
AP
IT
br2
GF
P β
lll-T
ubul
in
vGlu
T G
FP
DA
PI
Adult SEZ, 3 DIV
Adult SEZ, 3 DIV Adult SEZ, 28 DIV
Adult SEZ,3 DIV
All SEZ Lateral Dorsal
Per
cent
age
of D
AP
I-po
sitiv
ece
lls th
at w
ere
Tbr
2 po
sitiv
e
109876543210
a
c d
bFigure 7 Adult SEZ stem and progenitor cells give rise to some glutamatergic neurons in vitro. (a) Two example SEZ neuroblasts expressing Dcx (green) and Tbr2 (red) after 3 d in vitro (3 DIV). Boxed area shows Tbr2-positive nuclei at higher magnification. (b) Quantification of Tbr2-positive nuclei in cultured SEZ stem and progenitor cells isolated from the dorsal, lateral or both walls of the lateral ventricle after 3 DIV. Tbr2-positive cells were enriched in cultures derived from the dorsal SEZ (n = 3 experiments; there were 471 cells in the dorsal cultures, 602 cells in the ventral cultures and 1,384 cells in the mixed cultures; error bars represent s.e.m.). (c) Cells that were double positive for Tbr2 (red) and βIII-tubulin (blue) originated from proliferating progenitors, as indicated by transduction with a GFP-encoding retrovirus shortly after plating. (d) vGluT (red) immunoreactivity was also detected in GFP-positive cells 4 weeks after retroviral transduction, indicating that they originated from proliferating progenitors. (e) Left, micrograph shows an adult SEZ-derived glutamatergic neuron transduced with a lentiviral vector expressing GFP under the synapsin promoter that was patched by the recording electrode (inset). Middle, immunostaining for vGluT and GFP after recording revealed that this cell was highly decorated with vGluT-positive puncta. Right, stimulation of the neuron evoked an autaptic response that was blocked by the AMPA/kainate receptor antagonist CNQX, revealing its glutamatergic nature. Nuclei were visualized with DAPI (a,d). Scale bars represent 20 µm.
©20
09 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.
nature neurOSCIenCe VOLUME 12 | NUMBER 12 | dEcEMBER 2009 1531
a r t I C l e S
source for adult-generated glutamatergic neurons in the adult SEZ.
The genetic and viral fate-mapping experi-ments permitted us to obtain more precise information on the morphology of the newly generated glutamatergic neurons in the glomerular layer. Distinct from GABAergic periglomerular neurons (PGNs), these neu-rons typically extended their dendritic arbors adjoining several glomeruli, characteristic of so-called short-axon cells31,38, a glutamatergic subpopulation of juxtaglomerular neurons. Notably, despite their small number, short-axon cells are important for center-surround inhibition among glomeruli, providing excitatory glutamatergic drive to inhibitory PGNs of up to 30 glomeruli31. However, we cannot exclude that other subpopulations of juxtaglomerular glutamatergic neurons are also generated in the adult, potentially in a similar diver-sity of subtypes as observed for the GABAergic PGNs.
The physiological maturation and integration of these cells is sup-ported by the expression of the immediate-early gene c-fos, a mem-ber of a family of transcription factors that are rapidly regulated by neuronal activity35. Detection of c-fos has previously been used to demonstrate the integration of newborn neurons in the adult dentate gyrus39 and the olfactory bulb35. The expression of c-fos in the BrdU and vGluT2 double-positive neurons, as well as the presence of asym-metric synaptic contacts established by Neurog2-derived neurons, which we observed by electron microscopy, both support a functional integration of these newborn glutamatergic neurons into the neuronal network several weeks after their birth.
One question is what is the functional role of this specific glutama-tergic interneuron population. Indeed, even small subsets of olfactory neurons can have a profound functional effect on olfactory informa-tion processing40 and the specificity of adult generation of vGluT2-expressing neurons in the glomerular layer suggests that they have a specific role in the glomerular layer neuronal network, the first synaptic station of the incoming afferents. Synapses using vGluT2 for loading synaptic vesicles generally have a higher release probability compared with those expressing vGluT1 (ref. 41). Given the impor-tance of short-axon cells in center-surround inhibition, it can be spec-ulated that the new addition of glutamatergic neurons, as with their
GABAergic counterparts, is crucial for modifying the interglomerular circuitry during olfactory discrimination learning42.
SEZ regionalization and neuronal subtype specificationRecent evidence indicates that the SEZ in adult mice is highly region-alized, with stem cells in different locations having distinct embryonic origins and neuronal fates5,13,15,16. Here, we identified a previously unknown SEZ progenitor population that is specifically localized in dorsal regions and undergoes the same transcription factor expression sequence that characterizes many glutamatergic subtypes throughout the brain12,21. Sequential expression of Tbr2 and Tbr1 in developing neurons is a hallmark of glutamatergic lineage throughout the brain21. In the developing cerebral cortex and hippocampus, Tbr2-expressing progenitors arise from Neurog2-positive progenitors that themselves are generated by Pax6-expressing radial glia43.
We found that this transcription factor sequence was well conserved in the adult dorsal SEZ (Supplementary Figs. 2 and 13). Although Pax6 was expressed in most Neurog2- or Tbr2-expressing cells, the latter never colocalized with Dlx. Notably, the Neurog2-express-ing lineage also originated from Mash1-expressing progenitors, as many Neurog2-positive cells in the RMS are also Mash1 positive (Supplementary Fig. 2). Short-term fate mapping by Mash1BAC-gfp mice also revealed that Tbr2-positive progenitors originated from the Mash1 lineage (Supplementary Fig. 2). These data imply that there are at least two different lineages arising from Pax6- and Mash1-expressing progenitors (Supplementary Fig. 13), a larger population that leads to neuroblasts expressing both Dlx and Pax6
Retroviral injection
d b
cT
br2
GF
P
Tbr
2 D
cx
Tbr
2 D
cx
Lesion site
RMS Tbr2
Ctx
RMS
Dcx
Tbr2
DAPI Tbr2 Dcx
LV
DAPI GFP
LV
a
b
c d
Figure 8 Tbr2-positive cells migrate into the lesioned cerebral cortex. (a) Retroviral injections of GFP-encoding vectors 2 d before laser illumination resulted in GFP-positive cells that migrated away from the RMS 3–4 d after lesion, some of which were Tbr2 immunoreactive (red). (b) Fluorescence micrograph depicting Tbr2- and Dcx-immunopositive cells in an overview of the lesioned cerebral cortex hemisphere and the RMS 1 week after chlorine e6–induced lesion. The schematic drawing to the left depicts the location of the arrows in b on the right side and the location of the micrographs shown in c and d on the left side. (c,d) Tbr2-and Dcx-positive cells were present in dorsal regions of the SEZ and RMS (white arrows in c) and were sometimes found in small clusters (c) or in the cortical gray matter (d). Boxed areas (gray) are shown at higher magnifications. Nuclei were visualized with DAPI (a–d). Scale bars represent 20 µm (a,c,d) and 50 µm (b).

©20
09 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.
1532 VOLUME 12 | NUMBER 12 | dEcEMBER 2009 nature neurOSCIenCe
a r t I C l e S
and a smaller population sequentially expressing Pax6, Neurog2 and Tbr2 (Supplementary Fig. 13). Thus, precursors expressing Pax6 and Mash1 can give rise to both GABA and glutamatergic lineages, raising the question of which signals regulate lineage progression. One candidate that might instruct dorsal SEZ progenitors toward a glutamatergic fate is p-catenin–mediated Wnt signaling, which is involved in upregulation of Neurog1 via Lef1 that directly binds to the Neurog1 promoter in vitro in embryonic telencephalic progenitors44. Notably, Neurog1 and Neurog2 are typically expressed together20. Once Neurog2 is upregulated, its functional relevance in the specifi-cation of glutamatergic neurons from adult SEZ cells is evident not only from developmental analysis20,45, but also from the fact that overexpression of Neurog2 in adult-derived neurosphere cells is suf-ficient to upregulate Tbr1 and to direct virtually all neurons toward a functional glutamatergic identity37.
A source for regenerating glutamatergic neuronsAlthough neurogenesis in most regions of the telencephalon, including the striatum and cerebral cortex, ceases at early postnatal stages, the ongoing olfactory neurogenesis serves as a potential source of neuro-blasts after injury. Indeed, many new neuroblasts migrate into the striatum and generate GABAergic neurons after stroke46. Neuroblasts from the SEZ and RMS also migrate into the cerebral cortex, as has been observed after postnatal injury47,48 or adult cerebral cortex lesion with chlorine e6–induced cell death6,7. Using the latter model in conjunction with retroviral lineage tracing, we found that photo-toxic lesion of the cerebral cortex resulted in rerouting of some SEZ progenitors out of the RMS toward the damaged cortex. Some of the recruited SEZ progenitors expressed Tbr2, consistent with their deriva-tion from the glutamatergic lineage. On settling in the damaged cortex, some of these neurons appear to acquire the appropriate layer identity, as suggested by expression of Cux1 in Tbr2-driven GFP-positive cells in upper cortical layers following lesion of callosal projection neurons. In contrast, none of the adult-generated glutamatergic neurons in the glomerular layer were found to express Cux1. These data suggest that progenitors destined to generate glutamatergic juxtaglomerular neurons of the olfactory bulb can be diverted toward a different, in this case a cortical projection neuron, fate6,7. Although the molecular mechanisms underlying such respecification remain unclear, they may involve local cues provided by the injured tissue. Notably, respecifi-cation of SEZ-derived cells toward a different neuronal identity has been described with cells migrating after stroke from the SEZ into the striatum, where they then acquire a Darp32-positive medium spiny neuronal phenotype that is normally not generated in adult olfactory bulb neurogenesis46.
Taken together, our discovery of ongoing generation of gluta-matergic juxtaglomerular neurons highlights not only the diversity of adult-generated olfactory neurons in this region, but also the importance of understanding the molecular code of the distinct subtypes of adult progenitors as a major step toward using these neurons for repair. Indeed, the recent discovery of molecular fate determinants for subsets of cortical projection neurons may allow directing the source of adult progenitors for glutamatergic neu-rons identified here more efficiently toward the repair of cortical projection neurons.
METhODSMethods and any associated references are available in the online version of the paper at http://www.nature.com/natureneuroscience/.
Note: Supplementary information is available on the Nature Neuroscience website.
AcknowledgmentSWe would like to thank F. Bedogni for invaluable advice on methods and we are particularly grateful to Y. Yanagawa for the Gad67::gfp mice. We would also like to thank T. Simon-Ebert, A. Steiner, T. Öztürk and S. Nestel for excellent technical assistance and S. Bauer for retroviral production. We would also like to thank A. Saghatelyan for invaluable discussions and comments on the manuscript. This work was supported by grants from the Deutsche Forschungsgemeinschaft, including the excellence cluster Center for Integrated Protein Science Munich, by European Transcriptome, Regulome and Cellular Commitment Consortium from the European Union and the Bundesministerium für Bildung und Forschung to M.G., and the Bavarian State Ministry of Sciences, Research and the Arts (ForNeuroCell) to B.B. and M.G. E.W. was supported by a fellowship from the Medical Research Council, I.O. by a fellowship from the John & Lucille van Geest Foundation and O.R. by a grant from the Dr. Scholl Foundation. Research in the laboratory of F.G. is supported by funds from the Medical Research Council and grants from the European Commission Research and Technological Development program. C.P. is presently supported by an AVENIR-contract from Institut national de la santé et de la recherché mèdicale and Fondation de France. R.D.H. is supported by a Heart and Stroke Fund of Canada Research Fellowship. Work done in the laboratory of R.F.H. was supported by grants from the US National Institutes of Health (R01 NS050248 and R01 MH080766).
AUtHoR contRIBUtIonSM.S.B., O.R. and R.D.H. made the original observation of glutamatergic progenitors in the SEZ. M.S.B. conducted most of the experiments. J.N., E.W., O.R. and R.D.H. conducted experiments crucial for the identification of adult-generated glutamatergic neurons in the olfactory bulb. I.O. and R.Y. contributed to experiments using Neurog2+/gfp and Tbr2BAC-gfp mice, respectively. A.L. and S.G. designed and produced the viral vectors. F.E. and G.S. provided the Gad65-gfp mice. C.P. contributed with experiments using Ngn2+/gfp and Mash1BAC-gfp mice. F.G. provided the Neurog2gfp/+ and E1–Neurog2-cre mice. M.F. designed the electron-microscopy experiments and analyzed the data. B.B. designed and conducted all of the electrophysiological experiments. R.F.H., O.R. and M.G. supervised the project and designed experiments. M.G. wrote the manuscript. M.S.B., B.B., R.D.H., R.F.H. and O.R. contributed to the writing of the manuscript.
Published online at http://www.nature.com/natureneuroscience/. Reprints and permissions information is available online at http://www.nature.com/reprintsandpermissions/.
1. Kokaia, Z. & Lindvall, O. Neurogenesis after ischaemic brain insults. Curr. Opin. Neurobiol. 13, 127–132 (2003).
2. Sohur, U.S., Emsley, J.G., Mitchell, B.D. & Macklis, J.D. Adult neurogenesis and cellular brain repair with neural progenitors, precursors and stem cells. Phil. Trans. R. Soc. Lond. B 361, 1477–1497 (2006).
3. Ming, G.L. & Song, H. Adult neurogenesis in the mammalian central nervous system. Annu. Rev. Neurosci. 28, 223–250 (2005).
4. Gage, F.H. Mammalian neural stem cells. Science 287, 1433–1438 (2000).5. Merkle, F.T., Mirzadeh, Z. & Alvarez-Buylla, A. Mosaic organization of neural stem
cells in the adult brain. Science 317, 381–384 (2007).6. Chen, J., Magavi, S.S. & Macklis, J.D. Neurogenesis of corticospinal motor neurons
extending spinal projections in adult mice. Proc. Natl. Acad. Sci. USA 101, 16357–16362 (2004).
7. Magavi, S.S., Leavitt, B.R. & Macklis, J. D. Induction of neurogenesis in the neocortex of adult mice. Nature 405, 951–955 (2000).
8. Lledo, P.M., Alonso, M. & Grubb, M.S. Adult neurogenesis and functional plasticity in neuronal circuits. Nat. Rev. Neurosci. 7, 179–193 (2006).
9. Wonders, C.P. & Anderson, S.A. The origin and specification of cortical interneurons. Nat. Rev. Neurosci. 7, 687–696 (2006).
10. Brill, M.S. et al. A dlx2- and pax6-dependent transcriptional code for periglomerular neuron specification in the adult olfactory bulb. J. Neurosci. 28, 6439–6452 (2008).
11. Waclaw, R.R. et al. The zinc finger transcription factor Sp8 regulates the generation and diversity of olfactory bulb interneurons. Neuron 49, 503–516 (2006).
12. Bertrand, N., Castro, D.S. & Guillemot, F. Proneural genes and the specification of neural cell types. Nat. Rev. Neurosci. 3, 517–530 (2002).
13. Hack, M.A. et al. Neuronal fate determinants of adult olfactory bulb neurogenesis. Nat. Neurosci. 8, 865–872 (2005).
14. Roybon, L., Deierborg, T., Brundin, P. & Li, J.Y. Involvement of Ngn2, Tbr and NeuroD proteins during postnatal olfactory bulb neurogenesis. Eur. J. Neurosci. 29, 232–243 (2009).
15. Young, K.M., Fogarty, M., Kessaris, N. & Richardson, W.D. Subventricular zone stem cells are heterogeneous with respect to their embryonic origins and neurogenic fates in the adult olfactory bulb. J. Neurosci. 27, 8286–8296 (2007).
16. Kohwi, M. et al. A subpopulation of olfactory bulb GABAergic interneurons is derived from Emx1- and Dlx5/6-expressing progenitors. J. Neurosci. 27, 6878–6891 (2007).

©20
09 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.
nature neurOSCIenCe VOLUME 12 | NUMBER 12 | dEcEMBER 2009 1533
a r t I C l e S
17. Kroll, T.T. & O’Leary, D.D. Ventralized dorsal telencephalic progenitors in Pax6 mutant mice generate GABA interneurons of a lateral ganglionic eminence fate. Proc. Natl. Acad. Sci. USA 102, 7374–7379 (2005).
18. Nikoletopoulou, V. et al. Neurotrophin receptor–mediated death of misspecified neurons generated from embryonic stem cells lacking Pax6. Cell Stem Cell 1, 529–540 (2007).
19. Dellovade, T.L., Pfaff, D.W. & Schwanzel-Fukuda, M. Olfactory bulb development is altered in small-eye (Sey) mice. J. Comp. Neurol. 402, 402–418 (1998).
20. Schuurmans, C. et al. Sequential phases of cortical specification involve Neurogenin-dependent and -independent pathways. EMBO J. 23, 2892–2902 (2004).
21. Hevner, R.F., Hodge, R.D., Daza, R.A. & Englund, C. Transcription factors in glutamatergic neurogenesis: conserved programs in neocortex, cerebellum, and adult hippocampus. Neurosci. Res. 55, 223–233 (2006).
22. Hevner, R.F. et al. Tbr1 regulates differentiation of the preplate and layer 6. Neuron 29, 353–366 (2001).
23. Geschwind, D. GENSAT: a genomic resource for neuroscience research. Lancet Neurol. 3, 82 (2004).
24. Kowalczyk, T. et al. Intermediate neuronal progenitors (basal progenitors) produce pyramidal-projection meurons for all layers of cerebral cortex. Cereb. Cortex 19, 2439–2450 (2009).
25. Ninkovic, J., Mori, T. & Gotz, M. Distinct modes of neuron addition in adult mouse neurogenesis. J. Neurosci. 27, 10906–10911 (2007).
26. Mori, T. et al. Inducible gene deletion in astroglia and radial glia—a valuable tool for functional and lineage analysis. Glia 54, 21–34 (2006).
27. Liu, X., Wang, Q., Haydar, T.F. & Bordey, A. Nonsynaptic GABA signaling in postnatal subventricular zone controls proliferation of GFAP-expressing progenitors. Nat. Neurosci. 8, 1179–1187 (2005).
28. Tamamaki, N. et al. Green fluorescent protein expression and colocalization with calretinin, parvalbumin and somatostatin in the GAD67-GFP knock-in mouse. J. Comp. Neurol. 467, 60–79 (2003).
29. López-Bendito, G. et al. Preferential origin and layer destination of GAD65-GFP cortical interneurons. Cereb. Cortex 14, 1122–1133 (2004).
30. Berger, J. et al. E1-Ngn2/Cre is a new line for regional activation of Cre recombinase in the developing CNS. Genesis 40, 195–199 (2004).
31. Aungst, J.L. et al. Centre-surround inhibition among olfactory bulb glomeruli. Nature 426, 623–629 (2003).
32. Gabellec, M.M., Panzanelli, P., Sassoe-Pognetto, M. & Lledo, P.M. Synapse-specific localization of vesicular glutamate transporters in the rat olfactory bulb. Eur. J. Neurosci. 25, 1373–1383 (2007).
33. Srinivas, S. et al. Cre reporter strains produced by targeted insertion of EYFP and ECFP into the ROSA26 locus. BMC Dev. Biol. 1, 4 (2001).
34. Ohmomo, H. et al. Postnatal changes in expression of vesicular glutamate transporters in the main olfactory bulb of the rat. Neuroscience 160, 419–426 (2009).
35. Magavi, S.S., Mitchell, B.D., Szentirmai, O., Carter, B.S. & Macklis, J.D. Adult-born and preexisting olfactory granule neurons undergo distinct experience-dependent modifications of their olfactory responses in vivo. J. Neurosci. 25, 10729–10739 (2005).
36. Gascón, S., Paez-Gomez, J.A., Diaz-Guerra, M., Scheiffele, P. & Scholl, F.G. Dual-promoter lentiviral vectors for constitutive and regulated gene expression in neurons. J. Neurosci. Methods 168, 104–112 (2008).
37. Berninger, B., Guillemot, F. & Gotz, M. Directing neurotransmitter identity of neurones derived from expanded adult neural stem cells. Eur. J. Neurosci. 25, 2581–2590 (2007).
38. Pinching, A.J. & Powell, T.P. The neuron types of the glomerular layer of the olfactory bulb. J. Cell Sci. 9, 305–345 (1971).
39. Jessberger, S. et al. Seizure-associated, aberrant neurogenesis in adult rats characterized with retrovirus-mediated cell labeling. J. Neurosci. 27, 9400–9407 (2007).
40. Eyre, M.D., Kerti, K. & Nusser, Z. Molecular diversity of deep short-axon cells of the rat main olfactory bulb. Eur. J. Neurosci. 29, 1397–1407 (2009).
41. Murphy, G.J. & Isaacson, J.S. Presynaptic cyclic nucleotide–gated ion channels modulate neurotransmission in the mammalian olfactory bulb. Neuron 37, 639–647 (2003).
42. Mouret, A. et al. Learning and survival of newly generated neurons: when time matters. J. Neurosci. 28, 11511–11516 (2008).
43. Scardigli, R., Baumer, N., Gruss, P., Guillemot, F. & Le Roux, I. Direct and concentration-dependent regulation of the proneural gene Neurogenin2 by Pax6. Development 130, 3269–3281 (2003).
44. Israsena, N., Hu, M., Fu, W., Kan, L. & Kessler, J.A. The presence of FGF2 signaling determines whether beta-catenin exerts effects on proliferation or neuronal differentiation of neural stem cells. Dev. Biol. 268, 220–231 (2004).
45. Parras, C.M. et al. Divergent functions of the proneural genes Mash1 and Ngn2 in the specification of neuronal subtype identity. Genes Dev. 16, 324–338 (2002).
46. Arvidsson, A., Collin, T., Kirik, D., Kokaia, Z. & Lindvall, O. Neuronal replacement from endogenous precursors in the adult brain after stroke. Nat. Med. 8, 963–970 (2002).
47. Goings, G.E., Sahni, V. & Szele, F.G. Migration patterns of subventricular zone cells in adult mice change after cerebral cortex injury. Brain Res. 996, 213–226 (2004).
48. Faiz, M. et al. Substantial migration of SVZ cells to the cortex results in the generation of new neurons in the excitotoxically damaged immature rat brain. Mol. Cell. Neurosci. 38, 170–182 (2008).

©20
09 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.
nature neurOSCIenCe doi:10.1038/nn.2416
ONLINE METhODSAnimals. All animal procedures were performed in accordance with protocols approved by the state of Bavaria and in accordance with the UK (Scientific Procedures) Animals Act 1986. For wild-type analysis, we used C57BL/6 mice. In addition, we used mice with an enhanced GFP cassette inserted into either the Neurogenin2 locus (Neurog2+/gfp)49 or the Gad67 locus (Gad67::gfp)28 and mice that expressed the Cre recombinase in the GLAST locus25 crossed with the R26R-CFP reporter33 to track recombined cells (GLAST::creERT2 R26R-CFP). For induction of Cre-mediated recombination, we injected mice injected twice per day with 50 µl tamoxifen (20 mg ml–1, dissolved in corn oil, Sigma) for 5 d consecutively, followed by a 1-week interval and then another 5-d series of injections. We also used BAC transgenic mice expressing GFP under the Mash1 (also known as Ascl1) or Tbr2 genomic regulatory sequences (Mash1BAC-gfp, Tbr2BAC-gfp)23,50, transgenic mice that express GFP under the Gad65 promoter (Gad65-gfp)29 and mice expressing the Cre recombinase under the enhancer ele-ment 1 of Neurogenin2 (E1–Neurog2-cre)30,43 crossed with the Z/EG25 reporter (E1–Neurog2-cre; Z/EG) to follow progeny derived from the Ngn2 derived line-age. All of the mice that we used were adults of 2–3 months of age, with the exception of the mice that we used for lentiviral ventricular injections, which were 3–4 weeks old.
BrdU treatment. For detection of proliferating cells, we injected BrdU (Sigma) intraperitoneally (50–100 mg per kg of body weight) 1–2 h before perfusion to label fast-proliferating cells (short pulse) or placed it in the mice’s drinking water for 3 weeks (1 mg ml–1, complete exchange of water twice per week), which was followed by BrdU-free water for 3–4 weeks to allow full neuronal differentiation of labeled progenitors. For the BrdU chase experiment, a single BrdU pulse was given to the pregnant mother at embryonic day 14 or postnatal day 3 (50 mg per kg) and littermates were killed at postnatal days 16 and 60.
In situ hybridization. The vGluT1 and vGluT2 plasmids that we used for in situ hybridization were the kind gifts of L. Cheng (Chinese Academy of Sciences) and Q. Ma. (Harvard Medical School). The Mash1 and Neurog2 plasmids were provided by F.G. and the Tbr2 and gfp24 were provided by R.F.H.’s laboratory. The Gad65 and Gad67 plasmids were obtained from W. Wurst (Max Plank Institute for Psychiatry).
Digoxigenin-labeled RNA probes were generated by in vitro transcription (NTP labeling mix, Roche; T3, T7 or SP6 polymerase, Stratagene) and in situ hybridization was performed on 30-µm-thick cryostate sections or 70-µm-thick vibratome sections of perfused brains following standard protocols using α-digoxigenin antibodies (Roche, 1:2,000). For fluorescent in situ hybridization, we used a tyramide signal amplification kit (Perkin Elmer).
Immunohistochemistry. For immunohistochemistry, mice were deeply anaes-thetized using 5% chloralhydrate (wt/vol) in phosphate-buffered saline (PBS, 0.1 ml per 10 g of body weight), and killed by transcardial perfusion with PBS and then 4% paraformaldehyde (wt/vol, PFA) in PBS. Brains were dissected and postfixed overnight in PFA at 4 °C. For cryostate sections, brains were cryoprotected and cut, and immunostainings were carried out at a thickness of 20 µm. Alternatively, free-floating vibratome sections were cut at a thickness of 60 µm after postfixation.
Primary antibodies were diluted in 0.1 M PBS containing 0.5% Triton X-100 (wt/vol) and 10% normal goat serum (vol/vol) or 2% bovine serum albumin (wt/vol). We used primary antibodies to BrdU (rat, 1:400, Abcam or Accurate; sheep, 1:1,000, Fitzgerald Industries), activated caspase-3 (Promega, rabbit, 1:200), c-fos (mouse, 1:200, Chemicon), pan-Dlx (1:500, rabbit, kindly pro-vided by J. Kohtz, Northwestern University), Dcx (1:2,000, rabbit, Abcam; 1:500, goat, Santa Cruz), GFAP (1:250, guinea pig, Advanced Immunochemical; 1:500, mouse, Chemicon), GFP (1:500, rabbit, Invitrogen; 1:1,000 chicken, Abcam), Ki67 (goat, 1:500, Santa Cruz), Mash1 (mouse, 1:200, kindly provided by D. Anderson, California Institute of Technology; mouse, 1:100, BD Biosciences), Neurog2 (1:100, goat, Santa Cruz), Nestin (1:50, rabbit, Chemicon), Pax6 (mouse, 1:1,000, kindly provided by D. Schulte, Developmental Studies Hybridoma Bank; rabbit, 1:300, Covance), PSA-NCAM (1:1,000, mouse, Chemicon), Tbr1 (1:2,000, rabbit, Chemicon or Abcam), Tbr2 (1:2,000, rabbit, Chemicon or Abcam), vGluT1 (1:1,000, rabbit, Synaptic Systems) and vGluT2 (1:1,000, rabbit, Synaptic Systems). Specimens were incubated overnight at 4 °C. After thorough washing,
antibody staining was revealed by appropriate species- or subclass-specific sec-ondary antibodies conjugated to Alexa-488 (1:1,000, Invitrogen), Alexa-568 (1:1,000, Invitrogen), Cy2 or Cy3 (1:1,000, Dianova). Biotinylated secondary antibodies (1:200, Vector Laboratories) were used in combination with the TSA fluorescent amplification kit (Perkin Elmer). Specific labeling was checked by omitting the primary antibody.
BrdU immunohistochemistry was performed as described above after 2 M HCl pretreatment for 45–60 min, followed by incubation in borate buffer (0.1 M, 10 min, pH 8.5). Antigen retrieval for detection of Neurog2 protein was carried out using citrate buffer (0.1 M, pH 6, 20 min, 80 °C).
Viral vector injections. The retro- or lentiviral vectors that we used encoded only GFP, which was driven by a CAG promoter or ubiquitin promoter, respectively. Viral preparations and injections were performed as described previously10,13. Briefly, mice were deeply anesthetized (ketamine (100 mg per kg, CP Pharma) and xylazine (5 mg per kg, Rompun, Bayer)) and injected with 0.5 µl of viral suspen-sion at 0.7 (anteroposterior), 1.2 (mediolateral) and 1.7–1.4 (dorsoventral) for adult SEZ, and on the level of bregma, 0.8 (mediolateral) and –2.0 (dorsoventral) (relative to bregma) for ventricular injections at the age of 3 weeks.
Immunoelectron microscopy. For electron-microscopic analysis, mice were killed by perfusion with 4% PFA containing 0.1% glutaraldehyde (vol/vol) and postfixed overnight at 4 °C. Free-floating, 50-µm-thick vibratome sections were incubated in primary antibody overnight at 4 °C. After thorough wash-ing, primary antibody staining was detected with a biotin-conjugated secondary antibody (1:200, Vector labs), followed by the ABC kit (Vector labs) and DAB (Polysciences) labeling. The intensity of the staining reaction was monitored in the microscope. Control sections were stained with the secondary antibody alone and no staining could be observed. Sections containing DAB-stained cells were dehydrated in ascending series of ethanol and finally embedded in Durcupan. Ultrathin sections were cut on a Reichert Ultratome and viewed in a Philips 100 electron microscope.
Statistical analysis. Stainings were analyzed using an Olympus FV1000, a Leica SPE or Zeiss LSM5 Pascal laser-scanning confocal microscope with optical sections of maximum 1–1.5-µm intervals or with Zeiss AxioImager.Z1 with apotome component. Colocalization with cell type–specific antigens was quanti-fied in single optical sections of the laser-scanning microscopes. Between five and ten sections per mouse were counted per experiment until comparable numbers of stained or GFP-positive cells per mouse or experiment were reached. The total number of cells counted in all of the mice is indicated in the text. One mouse or experiment represents one mean value and standard deviations were calculated between mice or cultures. For the three-dimensional reconstruction of the loca-tion of Neurog2+/gfp-positive and Tbr2+ cells and of the dendritic arborization of E1–Neurog2-cre; Z/EG fate-mapped cells, we used the Neurolucida software (MBF Bioscience).
cultures and electrophysiology. Primary cultures of adult SEZ stem and pro-genitor cells were prepared as described previously10. Briefly, following dissection of the SEZ of adult mice (>8 weeks), cells were directly plated onto poly- d-lysine–coated coverslips in defined medium containing DMEM/F12 (Gibco) and 10 mM HEPES (Gibco), supplemented with B27 (Gibco) in the absence of epidermal growth factor and fibroblast growth factor 2. Dorsal SEZ cultures were obtained by dissecting the upper third of the whole ventricular wall, including the white matter, whereas the lower two-thirds of the ventricular wall were referred to as lateral SEZ cultures. For retroviral labeling, cultures were transduced 2 h after plating. We replaced 50% of the medium with fresh medium 2 d after plat-ing and subsequently maintained the cells for the indicated time periods until electrophysiological analysis or fixation for immunocytochemistry.
For electrophysiology, perforated patch-clamp recordings were performed at 20–25 °C with amphotericin-B (Calbiochem) for perforation37. Micropipettes were made from borosilicate glass capillaries. Pipettes were tip-filled with internal solution and back-filled with internal solution containing 200 µg ml–1 amphotericin-B. The electrodes had resistances of 2–2.5 MΩ. The internal solu-tion contained 136.5 mM potassium gluconate, 17.5 mM KCl, 9 mM NaCl, 1 mM MgCl2, 10 mM HEPES and 0.2 mM EGTA (pH 7.4) at an osmolarity of 300 mOsm. The external solution contained 150 mM NaCl, 3 mM KCl, 3 mM

©20
09 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.
nature neurOSCIenCedoi:10.1038/nn.2416
CaCl2, 2 mM MgCl2, 10 mM HEPES and 5 mM glucose (pH 7.4) at an osmolarity of 310 mOsm. The recording chamber was continuously perfused with external solution at a rate of 0.5 ml min–1. Cells were visualized with a Zeiss Axioskop2. Signals were sampled at 10 kHz with Axopatch 200B patch-clamp amplifiers (Axon Instruments), filtered at 5 kHz and analyzed with Clampfit 9.2 software (Axon Instruments). To assess autaptic connections, we step-polarized single cells in voltage clamp for 1 ms from −70 to +30 mV and recorded responses in the same cell. Responses were considered to be autaptic when they occurred within 3 ms of the step-depolarization. To visualize neurons for electrophysiology, we transduced adult SEZ cultures 14 d after plating with a lentiviral SYN-SYN-GFP vector36 that encodes GFP under the control of the human Synapsin promoter, which allows for specific labeling of neurons.
chlorine e6–induced cortical lesion. Chlorine e6 (Frontier Scientific) was cou-pled to latex beads (Lumaflour) as reported previously6,7. Briefly, 3 ml of 0.01 M phosphate buffer (pH 7.4) were used to dissolve 1.79 mg of chlorine e6 and 5 mg of 1-ethyl-3-(3-dimethylaminopropyl)-carbodiimide (MP Biomedicals) was added. The solution was kept for 30 min at 4 °C. We incubated 25 µl of rhodamine latex beads (Lumafluor) on a shaker for 1 h at 20–25 °C with 1.5 ml of activated chlorine e6 solution. The reaction was stopped by adding 200 µl of 0.1 M glycine buffer (pH 8.0). The coupled latex beads were centrifuged to remove
the supernatant (30 min, 50,000g), washed at least three times with 10 ml of 0.01 M phosphate buffer, resuspended in about 50 µl of 0.01 M phosphate buffer and stored at 4 °C. Injections were performed 5–7 times into the cerebral cortex (maximum depth 0.8 mm, around 100 nl each injection) using a glass capil-lary connected with an air pressure system (WPI). Retrograde transport of latex beads was checked in control mice that did not undergo laser illumination. Laser illumination (633 nm, Schäfter and Kirchhoff) of the contralateral hemisphere was performed directly on the dura after skull removal 2, 3 or 7 d later (5 min) followed by transcardial perfusion 7 or 10 d after that. Different illumination times (20, 10, 5 and 2 min) and different laser intensities (100, 50, 20, 10 and 5 mW) were tested and best results (number and specificity of dying cells) were obtained with 20–30 mW for 4–5 min. For the tracking experiments, retroviral injections were performed in one hemisphere, parallel with chlorine e6–coupled latex beads on the contralateral side, followed by laser illumination 2 d later and perfusion 3–4 d later. Cell death was confirmed by staining for active caspase 3 and NeuN 3 d after laser illumination on the contralateral hemisphere.
49. Seibt, J. et al. Neurogenin2 specifies the connectivity of thalamic neurons by controlling axon responsiveness to intermediate target cues. Neuron 39, 439–452 (2003).
50. Parras, C.M. et al. Mash1 specifies neurons and oligodendrocytes in the postnatal brain. EMBO J. 23, 4495–4505 (2004).

nature neuroscience
e r r ata
Erratum: Adult generation of glutamatergic olfactory bulb interneuronsMonika S Brill, Jovica Ninkovic, Eleanor Winpenny, Rebecca D Hodge, Ilknur Ozen, Roderick Yang, Alexandra Lepier, Sergio Gascón, Ferenc Erdelyi, Gabor Szabo, Carlos Parras, Francois Guillemot, Michael Frotscher, Benedikt Berninger, Robert F Hevner, Olivier Raineteau & Magdalena GötzNat. Neurosci. 12, 1524–1533 (2009); published online 1 November 2009; corrected after print 11 December 2009
In the version of this article initially published, the email address of one of the corresponding authors was misspelled. It should be [email protected]. The error has been corrected in the HTML and PDF versions of the article.
©20
09 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.