the innervation of parvalbumin-containing interneurons by vip-immunopositive interneurons in the...
TRANSCRIPT

The innervation of parvalbumin-containing interneuronsby VIP-immunopositive interneurons in the primarysomatosensory cortex of the adult rat
Csaba David,1 Axel Schleicher,2 Werner Zuschratter 3 and Jochen F. Staiger1
1Albert-Ludwigs-University Freiburg, Institute of Anatomy and Cell Biology, Department of Neuroanatomy, D-79001 Freiburg,Germany2Heinrich-Heine-University Dusseldorf, C. & O. Vogt Institute for Brain Research, D-40001 Dusseldorf, Germany3Leibniz-Institute for Neurobiology, D-39008 Magdeburg, Germany
Keywords: barrel cortex, calcium-binding proteins, correlated light and electron microscopy, inhibitory circuitry, neuropeptides
Abstract
c-Aminobutyric acid (GABA)ergic interneurons of neocortex consist of many subgroups with extremely heterogeneous morpholo-gical, physiological and molecular properties. To explore the putative effect of the vasoactive intestinal polypeptide-immunopositive(VIP +) neurons on neocortical circuitry, the number and distribution of VIP + boutons were analysed on somatodendritic domains of272 parvalbumin immunopositive (PV +) 3D-reconstructed neurons. The synaptic nature of 91% of somatic and 76% of dendriticcontacts was verified by electron microscopy. The target PV + neurons were separated in two significantly different groups by meansof cluster analysis. The first group (Cluster 1, 26%) received on average five times more VIP + synapses than those of the secondgroup. The second group (Cluster 2, 74%) contained cells that were poorly innervated by VIP + boutons or did not have eithersomatic or dendritic or any VIP innervation at all. The cells of Cluster 1 had a soma size and total dendritic length significantly smallerthan that of Cluster 2, however, they received three times more dendritic synapses, which resulted in a five times higher VIP +synaptic density on dendrites. Our results showed that although most of the PV + cells are innervated by VIP + boutons at a varyingdegree, some 6% of PV + cells received no input from VIP + interneurons. This suggests a refined morphological basis to influencethe majority of the PV + interneurons, which are very effectively controlling pyramidal cell firing. Together with metabolic andneuromodulatory effects of VIP, this would probably result in an enhanced responsiveness of the latter cell type to tactile stimuli.
Introduction
The heterogeneity of cortical inhibitory interneurons suggests diversefunctional roles for the different neuronal groups. In the neocortex,three main neurochemically identified groups of interneuronshave been proposed, which contain either: (i) parvalbumin (PV);or (ii) calbindin, somatostatin, occasionally neuropeptide-Y; or(iii) calretinin, vasoactive intestinal polypeptide (VIP), and occasion-ally cholecystokinin (Gonchar & Burkhalter, 1997; Kawaguchi &Kubota, 1997).
The PV-immunopositive (PV +) group mainly contains large basketcells and axo-axonic cells. Their inhibitory role is already text bookdata (see, however Szabadics et al., 2006). Both cell types innervatepyramidal cells and the basket cells innervate other interneurons too(Szentagothai, 1975; Somogyi et al., 1983; Staiger et al., 1997).
The VIP-immunopositive (VIP +) group contains neurons thatmostly possess radially orientated dendritic trees, spanning acrossseveral cortical layers (Connor & Peters, 1984; Bayraktar et al., 2000).Both dendrites and axons display very restricted lateral expansion, butare suited as a main source of translaminar inhibition (Kawaguchi &Kubota, 1996; Cauli et al., 1997; Bayraktar et al., 2000). Theirc-aminobutyric acid (GABA)ergic nature was established previously
(Kubota et al., 1994; Bayraktar et al., 1997). A small subpopulation ofVIP + interneurons with basket cell-like morphology, called smallbasket cells, has been described as well (Kawaguchi & Kubota, 1996).In the context of the present study it is important to state the lack ofco-expression of VIP and PV in the same inhibitory interneurons(Kawaguchi & Kubota, 1997; Porter et al., 1999; Cauli et al., 2000;Ekstrand et al., 2001; Toledo-Rodriguez et al., 2005).The VIP + neurons influence the general activity through metabolic
effects of VIP on: (i) blood flow in the nervous system (Cauli et al.,2004); and (ii) the activity of astrocytes (Magistretti et al., 1998).Besides the modulatory effect of VIP (Sessler et al., 1991), their directroles in cortical (Porter et al., 1998, 1999; Staiger et al., 2002, 2004) orthalamocortical (Sun et al., 2003) networks suggest a prominent rolein neuronal communication as well. The VIP + cells innervate otherinterneurons, but relatively few publications appeared describingaccurately the target structures of VIP + interneurons in the neocortex(e.g. Staiger et al., 2004), although it would be important forunderstanding their role in cortical information processing to have thefull connectivity picture available.In the present study, two non-overlapping, neurochemically iden-
tified groups of interneurons were quantitatively investigated: theVIP + and PV + interneurons. The VIP + interneurons are extensivelyinnervated by the PV + interneurons (Staiger et al., 1997), but aputative reciprocal innervation has not been investigated yet. Thespatial distribution of inputs is important in synaptic integration,
Correspondence: C. David, as above.E-mail: [email protected]
Received 10 November 2006, revised 12 February 2007, accepted 21 February 2007
European Journal of Neuroscience, Vol. 25, pp. 2329–2340, 2007 doi:10.1111/j.1460-9568.2007.05496.x
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing Ltd

therefore we analysed the location and density of VIP + terminals on3D-reconstructed somatodendritic domains of PV + cells. Our aimwas to examine the innervation of the PV + subpopulation ofGABAergic interneurons by VIP + boutons on a population level toreveal the overall pattern of connectivity of these cell types.
Materials and methods
Tissue preparation
All procedures were performed according to the German law onanimal welfare. Six male Wistar rats, weighing 250–300 g, were keptin the local animal care facility of the Heinrich-Heine-University understandardized conditions. The animals were deeply anaesthetized withan intraperitoneal pentobarbital injection (250 mg ⁄ kg body weight),then were intracardially perfused with approximately 50 mL 0.9%saline solution followed by 500 mL Zamboni fixative: 4% parafor-maldehyde dissolved in 0.1 m phosphate buffer pH 7.4 (PB) contain-ing 15 Vol-% picric acid. The brains were removed and immersed inthe same fixative for 4 h. After several washes with PB the brains werecut on a Leica VT 1000S vibratome (Leica, Wetzlar, Germany), thenominal section thickness was set to 50 lm. The sectioning began at arandom position rostral to the barrel cortex. Parallel representativeseries through the primary somatosensory cortex were collected inglass vials to be further processed free-floating.The sections were washed again several times with PB and
incubated with cryoprotectant (25% saccharose, 10% glycerol in0.01 m PB) for 40 min. The sections were frozen three times overliquid nitrogen. The cryoprotectant was washed out with PB, and thesections were stored in PB containing 0.05% sodium azide untilfurther processing.
Fluorescent staining and imaging
For this purpose, the sections were washed in 0.05 m Tris buffer(pH 7.6) containing 0.3% Triton (TBT), then putative unspecificantibody binding was blocked with 10% normal goat serum diluted inTBT for 40 min. The primary antibodies, rabbit anti-VIP (which wasgenerously provided by Dr T. Gorcs and extensively tested previously;Gulyas et al., 1990) and mouse anti-parvalbumin (Sigma, Taufkirchen,Germany), were diluted 1 : 20 000 and 1 : 40 000, respectively, in thesame buffer without normal serum. The sections were incubated withthis cocktail for 2 days at +4 �C, under gentle agitation. After severalwashes with TBT the sections were incubated with a mixture ofsecondary antibodies for 4 h: Alexa 488-conjugated goat anti-rabbitIgG and Alexa 594-conjugated goat anti-mouse IgG (both fromMolecular Probes, Leiden, the Netherlands), diluted at 1 : 200 and1 : 400, respectively, in TBT at room temperature. The unboundantibodies were removed with several washes of TBT, then the buffer
was changed to 0.1 m PB, and the sections were mounted on slidesand coverslipped with Aquapolymount (Polysciences, Warrington, PA,USA). Appropriately diluted normal serum instead of the primaryantibodies served as an immunocytochemical control in our study,which always resulted in the absence of any staining. Because thespecificity of the antibodies was tested and approved by themanufacturers, no further control reactions were carried out.The cortical columns present in the section were identified by the
aggregation of the PV + cells in layer IV, which makes barrels asmorphological correlates of functional columns visible (Fig. 1A). Thewhole volume of the identified cortical columns was automaticallyimaged by the Arivis Browser (Arivis Multiple Image Tools GmbH,Rostock, Germany) on a laser-scanning confocal microscope equippedwith a 63 · multi-immersion objective with a numerical aperture of1.3 (Leica TCS 4D, Bensheim, Germany). Systematic randomsampling was applied to select PV + cells for 3D reconstruction withthe software Neurolucida (Microbrightfield Europe, Magdeburg,Germany) using the confocal module. The reconstruction of samplePV + neurons was performed such that first the soma was recon-structed and then all primary dendrites and their daughter branchesthat could be unambiguously seen originating from the respectivesoma. During reconstruction, the VIP + boutons contacting the PV +cells were marked for further analysis. Only those VIP + boutons weremarked as a putative synaptic contact, which fulfilled two rigorouscriteria: (i) they were in focus in at least two consecutive, 0.6-lm-thickoptical sections; and (ii) there were no black pixels between the VIP +bouton and the PV + soma or dendrite. The raw data were extractedwith the software Neuroexplorer (Microbrightfield Europe) andexported to a spreadsheet.
Correlated light and electron microscopic examination
After the sections were washed in 0.05 m Tris buffer pH 7.6 (TB), theendogenous peroxidase activity was blocked with 0.5% H2O2 (Merck,Darmstadt, Germany) in TB for 30 min. To avoid uneven staining ofthe deep portions of the sections the penetration of the primaryantibodies was enhanced with enzymatic pretreatment optimized fordetection of these antigens during pilot experiments. The sectionswere incubated in 0.5 mg ⁄ mL trypsin solution containing 0.1% CaCl2dissolved in distilled water for 5 min at 37 �C. Before bothconsecutive primary antibody reactions the possible unspecific bindingof antibodies was blocked with 10% normal serum of the host animalof the corresponding secondary antibody diluted in TB for 40 min.The sections were then incubated with rabbit anti-VIP antibody dilutedat 1 : 20 000 in TB for 2 days at 4 �C. In the following step, afterwashing, biotin-conjugated goat anti-rabbit antibody (Vector Labor-atories, Burlingame, CA, USA), diluted at 1 : 200 in TB, was added tothe sections for 2 h followed by washing and incubation withVectastain ABC kit (Vector Laboratories), diluted at 1 : 400 in the
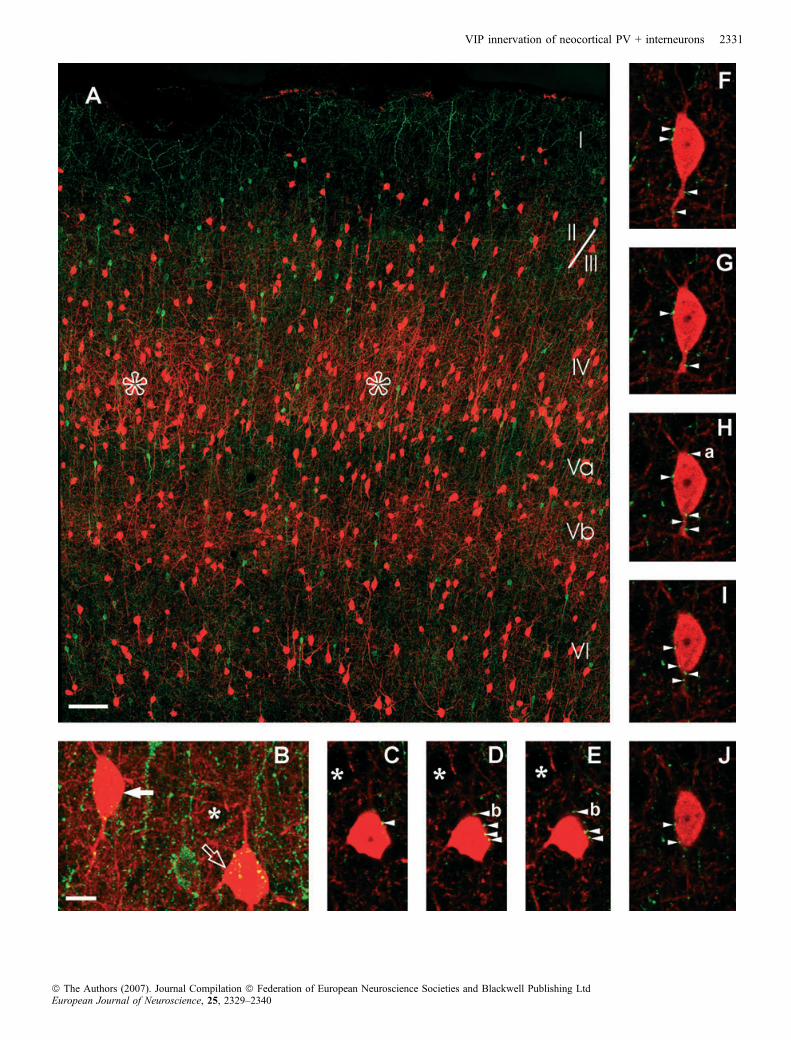
Fig. 1. Laser scanning confocal imaging of somatosensory cortex showing double labelling of VIP+ (green) and PV+ (red) neurons. (A) Typical example ofbarrel-related columns as examined in the present study (maximum intensity projection). The PV immunostaining allows demarcation of the cortical layers (Romannumerals) and two barrels were also visible in layer IV (open asterisks). The exact delineation of the barrel is not easy due to the relatively thin (50 lm) section, butthe higher density of PV+ cell bodies and that of dendrites make it possible to differentiate between the barrels and the interbarrel septa. Scale bar, 100 lm.(B) Maximum intensity projection of two PV+ cells surrounded by numerous VIP+ boutons. The majority of the PV+ cells showed a multipolar morphology(arrow), but some cells had a bipolar (not shown; 14 ⁄ 272) or bitufted (open arrow 74 ⁄ 272) shape. Due to the overlapping structures it is not easy to identify theunlabelled cell surrounded by numerous PV+ boutons (PV+ basket) labelled with an asterisk, but in the single optical sections (C–E) it is clearly visible. Scale bar,10 lm. (also holds for C–J). (C–J) Single optical sections (0.6 lm-thick) as used for the 3D reconstructions. The Arivis automated photographing and image browsersystem made it possible to handle images as large as 1000 · 2000 lm2 containing as many as 60–70 optical sections with a resolution of� 5 pixels ⁄ lm. That meansthat one can see all the stained cells of a section of a cortical column or select only one of them and observe all the boutons attached to it on the same image. (C–E)Single optical sections of the multipolar cell of Fig. 1B. The VIP+ boutons forming close contacts are marked with arrowheads. Bouton b was not counted since thereare black pixels between bouton and PV+ cell body. (F–J) Single optical sections of the bitufted cell of Fig. 1B. The bouton marked with a was not counted because itis in focus only in one optical section.
2330 C. David et al.
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2329–2340
VIP innervation of neocortical PV + interneurons 2331
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2329–2340

same buffer for 1 h. For the visualization the sections werepreincubated with 0.5% 3,3¢-diaminobenzidine-hydrochloride (DAB,Sigma) for 10 min, then 0.01% hydrogen peroxide was added to startthe reaction. After reaching optimal colour intensity the reaction wasstopped by washing with TB.The DAB reaction product was intensified by modified silver
impregnation (Gorcs et al., 1986), resulting in a dark brown to blackstaining of the VIP + cells and their processes. Briefly, the sectionswere incubated with 5% thyoglycolic acid in distilled water for 2 h toreduce the intrinsic argyrophily of the nervous tissue. The sectionswere incubated at room temperature in a physical developer containing12.5 mg ⁄ mL ammonium nitrate, 10 mg ⁄ mL silver nitrate, 25 mg ⁄ mLsodium carbonate (Merck, Bruchsal, Germany), 50 mg ⁄ mL silico-tungstic acid (Fluka, Taufkirchen, Germany) and 4 lL ⁄ mL 37%formaldehyde. After the sections reached the optimal intensification,the reaction was blocked by rinsing with 1% acetic acid. After toningin 0.05% gold chloride (Sigma) solution the silver precipitation wasfixed in 3% sodium thiosulphate (Merck) solution.The second immunoreaction was performed similarly as the first
one, but no silver intensification was performed. The primaryantibody, mouse anti-PV (Sigma) and the secondary horse anti-mouseantibody (Vector Laboratories) were diluted at 1 : 40 000 and 1 : 200in TB, respectively. After incubation with the ABC solution, thereaction was visualized either by Vectastain SG-kit (Vector Laborat-ories), which gave blue colour to the target structures, or by DAB(brown), which provided a better contrast in the electron microscope.The sections were treated with 1% OsO4 (Plano, Marburg,
Germany) in distilled water for 1 h, then were dehydrated in anascending series of ethanol (50%, 70%, 90%, 96% each for 10 min,twice for 10 min in 100% and twice for 5 min in propyleneoxide).After the 50% ethanol step the sections were treated with uranylacetate (Merck, Darmstadt, Germany) dissolved in 70% ethanol for45 min. After complete dehydration the sections were incubated inDurcupan ACM (Fluka) for 1 day, mixed according to the manufac-turer’s recommendation. Then the sections were mounted, cover-slipped and placed into a heated chamber. The Durcupan waspolymerized at 56 �C overnight.Six selected PV + neurons were photographed with a digital camera
(CoolSNAP, Visitron, Puchheim, Germany) attached to an Axioplan 2microscope (Zeiss, Gottingen, Germany), and the number of VIP +contacts that were subjected to further electron microscopicalverification as synapses was counted on the soma and on the stainedportion of the dendrites. Image stacks were processed with MetaView4.5 (Universal Imaging, West Chester; PA, USA). The cells were thencut out from the section on the slide with a scalpel blade under a lightmicroscope and placed on a new slide. A Beem capsule was gluedaround each tissue piece, which was then filled with Durcupan andincubated at 56 �C overnight. After polymerization the blocks wereremoved from the slide, the capsules were peeled off and the blockswere trimmed, first with a razor blade and then with a glass knife.Seventy nanometer-thick ultrathin serial sections were made on aLeica Ultracut UCT microtome (Leica, Vienna, Austria). The sectionswere mounted on Formvar-coated single-slot grids, contrasted withlead citrate and analysed in an electron microscope (AB-912, LEO,
Oberkochen, Germany). Photomicrographs were taken with a digitalcamera (Gatan, Munchen, Germany), optimized for contrast, bright-ness and sharpness, and montages were assembled in Photoshop 8.0.1(Adobe Systems, San Jose, CA, USA).
Data analysis
The PV + cells were examined in all cortical layers, except layer Iwhere they are absent. The cells were grouped according to thelocation of the cell body in the following layer compartments: (i)supragranular: layers II ⁄ III; (ii) granular: layer IV; and (iii) infragran-ular: layers Va, Vb and VI (Figs 1A and 2A) to find out if there arelaminar differences in cell connectivity. The following parameters ofeach cell were used for the multiparametric tests: volume of the cellbody; projection area of the cell body parallel to section plane; greatestand smallest diameter of the cell body; number of boutons contactingthe cell body; number of primary dendrites; whole length of dendrites;total number of VIP boutons contacting the dendritic tree; boutondensity on the dendrites; length of the first order dendrite; number ofVIP boutons contacting the first order dendrites; bouton density on thefirst order dendrites.The data were normalized by calculating z-scores, and were
subjected to an exploratory hierarchical cluster analysis (HCA) todetect putative subgroups of neurons in an observer-independentmanner, without any prior assumptions on grouping. HCA is based onthe dissimilarities between cells by calculating the intercellulardistance in a multidimensional space, where each dimension corres-ponds to one of the quantified cellular parameters. The HCA resultedin a cluster tree diagram (dendrogram) revealing a grouping ofthe cells into a number of distinct subgroups or clusters (Fig. 2B). Theshorter the distance between clusters, the more similar are thesubgroups. We used the Euclidean distance as dissimilarity measureand the Ward minimum variance method as linkage procedure (Cauliet al., 2000).The grouping of cells was validated and improved by linear
discriminant analysis (DA, complete; Dombrowski et al., 2001).Classification functions were calculated and the cells were reclassified(grouped) accordingly. This way, the classification of eight out of 372cells was modified. The DA resulted in canonical axes, which are linearcombinations of the variables, weighted by the coefficients of canonicaldiscriminant functions. The cells are depicted in a graph of the first twocanonical axes (Fig. 2C). The most important and characteristicvariables that define the first canonical axis are the following in order ofdecreasing weight: projection area of the cell body parallel to sectionplane; bouton density on dendrites; number of dendrites; number ofboutons on cell body; length of first order dendrites; and number ofboutons on first order dendrites. For the second canonical axis the mostcharacteristic parameters are: greatest diameter of the cell body;projection area of the cell body parallel to section plane; number ofdendrites; bouton density of first order dendrites; smallest diameter ofthe cell body; bouton density on dendrites.The parameters that best separated the cells into their clusters were
identified using a subsequent factor analysis (FA, principal componentmethod) on Clusters 1 and 2 as an independent procedure, which does
Fig. 2. Reconstructions and observer-independent identification of three groups of PV+ cells. (A) The diagram shows all reconstructed cells in the given corticalcolumn. Note the obviously incomplete dendritic trees of the cells of Cluster X (magenta). These cells were excluded from further investigation. Scale bar, 100 lm.All diagrams are color-coded identically: Cluster 1, orange; Custer 2, cyan; Cluster X, magenta. (B) Dendrogram of the HCA indicating the Euclidean distances(horizontal axis) of the three clusters of reconstructed cells (n ¼ 372). (C) Canonical plot of the three initial groups after refinement with DA. Ellipses represent68% confidence intervals of the samples. (D) Scatterplot showing the clear separation of the cells as indicated by FA. The cells were plotted according to thecalculated factors of FA, but individual cells were color labeled identical to the clustering established by the HCA and DA. Ellipses represent 68% confidenceintervals of the samples.
2332 C. David et al.
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2329–2340

VIP innervation of neocortical PV + interneurons 2333
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2329–2340
not use any prior knowledge concerning the number of clusters and thecluster members (for methodological details, see Maddock & Young,1987). Only three factors summarize 75% of the total variability. Themain contributions to factors 1, 2 and 3 were bouton density, cell bodygeometry and dendrite geometry (length and number), respectively.Figure 2D is a scatter plot of factors 1 and 2. To compare the resultswith respect to the previously extracted clusters, both clustering resultswere labelled by identical colours. The graph shows a clear separationof the two clusters, and thus validates the cluster definition from theHCA and its refinement using the DA.Univariate tests were applied to compare the individual parameters
between Clusters 1 and 2. Because most parameters failed thenormality test, the non-parametric Mann–Whitney U-test was applied.The most prominent differences are depicted in Fig. 3. To protect thestatistics against randomly occurring, significant differences, the P-values were multiplied by the number of features tested. Statisticalanalyses were performed in systat V.11 (SPSS, Chicago, IL, USA).Data shown in tables are given as mean ± standard error of the mean.
Results
Both the VIP and the PV immunostainings resulted in a labellingpattern similar to previous descriptions (for VIP: Bayraktar et al.,
2000; for PV: Celio, 1986; Staiger et al., 1997). Briefly, most of theVIP + cells were vertically orientated, i.e. their dendritic trees werebipolar or bitufted and the dendrites could be seen in all cortical layers,including layer I (Fig. 1A). There were a few multipolar VIP +neurons in our sample (not shown), which are very likely tocorrespond to small basket cells. The VIP + boutons were quitehomogenously distributed across all the layers, although a higherdensity of VIP + cell bodies was observed in layers II ⁄ III than in otherlayers. (Fig. 1A)In the primary somatosensory cortex, the PV + structures were
numerous in all layers except layer I. There were only a fewdendrites that reached into layer I. The stained neurons were mostlymultipolar (Figs 1B, C–E and 2A), but some bitufted and very rarelybipolar neurons were also detected (Figs 1B, F–J and 2A). The PV +neurons located in layer IV formed cell aggregates, where thedensely positioned stained cells and processes revealed the barrels(Fig. 1A). In the supra- and even more in the infragranular layers, theaverage density of the PV + structures was slightly lower. Anotherdistinguished feature of the PV + staining was the frequentoccurrence of pericellular baskets (Fig. 1C–E), additionally we couldobserve a lower amount of VIP + basket-like structures. Despitemeticulous examination, we did not find any cells that co-localizedVIP and PV.
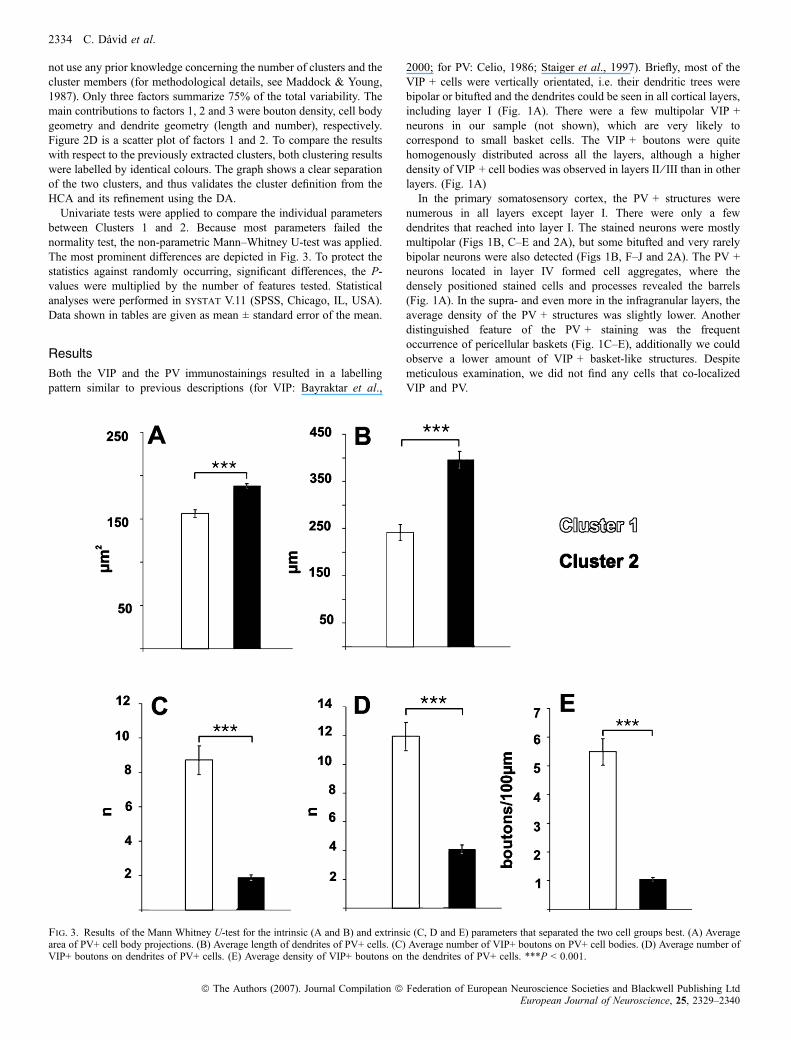
Fig. 3. Results of the Mann Whitney U-test for the intrinsic (A and B) and extrinsic (C, D and E) parameters that separated the two cell groups best. (A) Averagearea of PV+ cell body projections. (B) Average length of dendrites of PV+ cells. (C) Average number of VIP+ boutons on PV+ cell bodies. (D) Average number ofVIP+ boutons on dendrites of PV+ cells. (E) Average density of VIP+ boutons on the dendrites of PV+ cells. ***P < 0.001.
2334 C. David et al.
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2329–2340

In the present work, 1139 PV + cells were observed in the sectionsexamined, from which approximately one-third (372 cells) wereselected systematically, randomly and reconstructed in 3D. Ninety-three supragranular, 111 granular and 186 infragranular PV + cellswere analysed.
Mapping of VIP + boutons onto PV + neurons
HCA indicated three groups of cells (Fig. 2B). After DA wecompared the 3D reconstructions of the cells belonging to the samegroup to find out if they have any common morphological features ordistribution among the cortical layers (for one typical example, seeFig. 2A). Due to the methodological limitations, not all the selectedcells could be reconstructed acceptably. These cells were clearlyseparated from the other cells by the clustering algorithm, thus theywere excluded from the further investigation in an observer-independent manner (Cluster X, Fig. 2A–C). The two remaininggroups labelled as Cluster 1 and Cluster 2 contained 71 and 201cells, respectively, altogether about 23% of the total number of PV +cells (n ¼ 1139) counted in the cortical columns examined in thesample of six animals.
Cluster 1 contained the cells that had more somatic and dendriticcontacts with VIP + boutons despite shorter dendrites in comparisonto Cluster 2. The difference was smallest in the supragranular layercompartment (2.2 times higher) and greater in the granular layer (3.4times higher, Table 1). This difference was most pronounced in thecase of lower order dendrites. The first order dendrites of Cluster 1, inall layer compartments, had five–seven times more VIP + boutonsattached to them than that of Cluster 2. The second and third orderdendrites showed a smaller difference: three–four times more boutonson the cells of Cluster 1 (Table 2). The higher order dendrites showedbarely any difference between the two clusters and the bouton densitywas very low. In the 3D-reconstructed cell sample, 81 cells (22% ofthe reconstructed cells) did not have any bouton on their cell body, 52cells (14%) did not have any boutons on their reconstructed dendritictree. The intersection of these two sets contained 23 cells (6%) thatdisplayed no contacts with VIP + boutons at all. All of these cellswere members of Cluster 2 exclusively. The maximum number ofsomatic contacts is consequently up to 30 in Cluster 1 and up to 10 inCluster 2 in each of the cortical layer compartments (Table 1). Cells inClusters 1 and 2 did not show any significant differences in laminardistribution, though the number of cells of Cluster 2 showed a trend to
Table 1. Statistics on the reconstructed cells grouped by clusters and layers
Supragranular layer Granular layer Infragranular layer
Cluster 1 (n ¼ 71)Soma size (lm2) 154 ± 6.03 (106.9–205.62) 174 ± 9.67 (108–282) 148.89 ± 6.87 (77.3–239.07)Number of primary dendrites 3.74 ± 0.29 (2–7) 4.05 ± 0.28 (2–7) 3.89 ± 0.27 (2–8)Total dendritic length (lm) 262.37 ± 27.6 (32.7–473.4) 246.7 ± 20.99 (105–501) 232.24 ± 26.04 (33–524.4)Number of boutons on cell body 8.48 ± 1.5 (1–30) 11.35 ± 1.77 (1–29) 8.57 ± 1.2 (1–29)Number of boutons on dendrites 12.83 ± 1.35 (1–23) 14.35 ± 2.7 (2–52) 12.71 ± 1.78 (2–48)Total bouton density (boutons ⁄ 100 lm) 5.88 ± 1.19 (1.66–29.99) 5.55 ± 0.87 (1.05–19.05) 6.56 ± 0.74 (0.69–18.83)Length of first order dendrite (lm) 66.71 ± 12.35 (6.2–209.2) 64.43 ± 12.14 (13.3–251.9) 94.54 ± 11.24 (4.9–218.5)Number of boutons on first order dendrite 4.65 ± 0.83 (0–13) 4.25 ± 0.72 (0–13) 6.11 ± 0.67 (0–16)Bouton density on first order dendrite(boutons ⁄ 100 lm)
10.24 ± 2.13 (0–35.71) 7.61 ± 1.1 (0–16) 9.42 ± 2.08 (0–61.22)
Cluster 2 (n ¼ 201)Soma size (lm2) 182.12 ± 5.77 (113.6–297.3) 194.5 ± 4.54 (119–287.5) 185.48 ± 4.89 (85.7–293.1)Number of primary dendrites 5.28 ± 0.18 (3–8) 4.32 ± 0.14 (2–7) 4.67 ± 0.12 (2–7)Total dendritic length (lm) 400.64 ± 32.2 (104.5–937.3) 366.9 ± 25.52 (96.4–1061) 436.98 ± 23.89 (122–1277)Number of boutons on cell body 3.26 ± 0.37 (0–10) 2.65 ± 0.32 (0–11) 1.68 ± 0.24 (0–10)Number of boutons on dendrites 5.77 ± 0.55 (1–18) 4.23 ± 0.62 (0–22) 4.34 ± 0.45 (0–23)Total bouton density (boutons ⁄ 100 lm) 1.67 ± 0.16 (0.35–3.98) 1.13 ± 0.13 (0–5.57) 1.04 ± 0.1 (0–3.77)Length of first order dendrite (lm) 121.9 ± 10.23 (34.4–321.3) 120.8 ± 10.37 (11.1–397.4) 139.67 ± 9.73 (14.9–507.1)Number of boutons on first order dendrite 2.37 ± 0.34 (0–8) 1.88 ± 0.35 (0–18) 2.02 ± 0.33 (0–21)Bouton density on first order dendrite(boutons ⁄ 100 lm)
1.93 ± 0.3 (0–8.77) 1.52 ± 0.24 (0–8.33) 1.33 ± 0.17 (0–8.58)
Data are given as mean ± SEM, with range in parentheses.
Table 2. The densities of VIP+ boutons on PV+ dendrites,
Cluster and layer
Densities of VIP+ boutons on PV+ dendrites (boutons ⁄ 100 lm)
1st order 2nd order 3rd order 4th order 5th order 6th order
Cluster 1Supragranular 10.24 ± 2.13 (86) 5.91 ± 1.58 (104) 5.38 ± 1.16 (54) 2.05 ± 0.62 (22) 2.4 ± 0.58 (12) 0 (2)Granular 7.61 ± 1.10 (81) 5.06 ± 1.07 (102) 4.89 ± 1.12 (48) 2.9 ± 0.87 (17) 0 (6) n.a.Infragranular 9.42 ± 2.08 (109) 4.32 ± 1.21 (96) 6.1 ± 0.91 (42) 0.83 ± 0.35 (12) 2.38 (2) 2.87 (2)
Cluster 2Supragranular 1.93 ± 0.30 (227) 1.75 ± 0.29 (244) 0.78 ± 0.16 (124) 0.08 ± 0.04 (44) 1.03 ± 0.31 (8) n.a.Granular 1.52 ± 0.24 (298) 1.03 ± 0.22 (295) 1.16 ± 0.2 (150) 0.65 ± 0.17 (60) 1.72 ± 0.3 (6) 2.5 (2)Infragranular 1.33 ± 0.17 (411) 1.05 ± 0.2 (370) 0.77 ± 0.1 (162) 1.16 ± 0.27 (28) 0.2 ± 0.04 (8) 0 (2)
Data are given as mean ± SEM, grouped by clusters, layers and dendritic order, and the total number of segments are shown in parentheses. n.a., not applicable.
VIP innervation of neocortical PV + interneurons 2335
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2329–2340

become more abundant in the deeper cortical layers. The relevantmeasured or calculated parameters of the cells in Cluster 1 and Cluster2 are summarized in Tables 1 and 2, and the most significantdifferences were graphed in Fig. 3.
Correlated light and electron microscopic analysis
Six PV + neurons bearing numerous VIP + boutons closely attachedto the cell membrane were selected for correlated light and electronmicroscopic examination (Fig. 4) from all cortical layers except layerI. From the 61 somatic and 29 dendritic light-microscopicallyobserved contacts, 57 somatic and 21 dendritic ones were foundduring the electron microscopic investigation. Most of them [52 ⁄ 57(91%) somatic, 16 ⁄ 21 (76%) dendritic] were verified as symmetricsynapses (Fig. 4E and H). The number of somatic synapses was verylikely not underestimated due to the excellent colour contrast.However, that of the dendritic ones was overestimated by up to25%. Thus, in the present study the light microscopic data areconsidered as a maximum number.Figure 4 shows one of the examined PV + cells located at the border
of layers III and IV. Due to the shape of the cell body and the lightmicroscopically visible landmarks (Fig. 4A and C), the target cell isunambiguously recognizable at the electron microscopic level. Two ofthe 16 stained VIP + boutons, contacting the cell, are shown to formclear synaptic specializations in serial section analysis (Fig. 4D–H),as did the remaining ones not shown here.
Discussion
We have shown a differential innervation pattern of twosignificantly different subgroups of PV + interneurons by VIP +interneurons in the rat primary somatosensory cortex by analysingthe number and distribution of VIP + boutons on 272 PV +3D-reconstructed neurons imaged by high-resolution confocal laser-scanning microscopy. Subsequent correlated light and electronmicroscopy verified the synaptic nature of the light microscopicallycounted contacts. Thus, the following statements can be made.(i) Most but not all of the PV + cells receive a VIP + innervation.(ii) The PV + cell population contains two significantly differentcell clusters. (iii) The cells of both clusters are approximatelyevenly distributed among the cortical layers. (iv) Cluster 1contained more cells that showed in addition to the most frequentmultipolar, also bipolar or bitufted dendritic patterns. The dendritesof the multipolar neurons in Cluster 1 were more numerous butshorter than that of Cluster 2, and the bouton density on thedendrites was on average approximately five times higher. (v) Thecells of Cluster 2 were in general multipolar-shaped cells, possessedfewer but longer dendrites, on which innervation by VIP + boutonswas infrequent or even absent.
Technical considerations
The limitations of the methods and antibodies used for immunostain-ing (e.g. incompleteness of the dendritic tree, uneven antibodypenetration) were described in earlier works (Staiger et al., 1997;Bayraktar et al., 2000). Additionally, due to the penetration problemsof the primary antibodies, we had to apply enzymatic digestion topartially disintegrate the rigidly fixed protein meshwork. Thistreatment and the long incubation times of the whole process resultedin a reduced visibility of the postsynaptic membranes.Due to the high density of the PV + structures it was not always
possible to follow the individual dendrites to the end of the stainedportion. The not acceptably reconstructed cells were selected by theHCA and DA in an observer-independent manner and were excludedfrom the further investigation.
The role of VIP + and PV + interneurons in the inhibitory corticalnetwork
Previous investigations revealed that all VIP + interneurons in thebarrel cortex are targets of PV + interneurons (Staiger et al., 1997).This suggests that each member of the VIP + cell population isunder the control of at least one PV + cell. Our findings for the firsttime point out the asymmetric nature of this connection, as theinnervation was not complete in the reciprocal direction. Most of thePV + cells (94%) received inhibitory input from the VIP +population. The cells in Cluster 1 received on average five timesmore synapses from VIP + cells than that of Cluster 2 (Table 1), andthe arrangement of the VIP + boutons around the cell bodiesresembled baskets in many cases. Thus, the cells surrounded bymany VIP + boutons might correspond to one of the putative targetsof VIP-expressing small basket cells (Kawaguchi & Kubota, 1996).The influence of VIP + cells on the sparsely innervated PV + cellsof Cluster 2 that have only a few VIP + contacts may be probablyvery low, although in case of excitation a single synapse can beenough to fire the postsynaptic cell (Gulyas et al., 1993). However, asimilar effectiveness of single inhibitory synapses to veto actionpotential discharge still has to be demonstrated. The inhibitorysynapses most probably have a spatially more restricted effect(Williams & Stuart, 2003; Andrasfalvy & Mody, 2006), thus thesynapses that control action potential generation should be locatedperisomatically, whereas the more distally located dendritic contactsinfluence synaptic plasticity (Froemke et al., 2005).As initially described by Somogyi (1977), the vertically biased or
bitufted form of chandelier cells could be frequently observed infollow-up studies (Kawaguchi, 1995; Kawaguchi & Kubota, 1998;DeFelipe, 1999; Krimer & Goldman-Rakic, 2001; Kawaguchi &Kondo, 2002; Patz et al., 2004; Keros & Hablitz, 2005), thus it can beinferred that the Cluster 1 may contain at least some proportion of thechandelier cells of the PV + population.
Fig. 4. Light and electron microscopic correlation of VIP+ boutons attached to one of the selected PV+ cells in the supragranular layers. (A) The PV+ cellvisualized by DAB has two VIP+ contacts on its soma, which are visualized by silver intensified DAB. (B) Minimum intensity projection of the same cell createdfrom 20 thin optical sections (each 0.6 lm). The boutons b1 and b2 labelled individually, the other light microscopically identified contacts are labelled with openarrows, from which the boutons that could be identified as a synapse in the electron microscope marked with black open arrows. In the case of the boutons labelledwith white open arrows the synaptic specialization has not been found, since the probable synaptic membranes were parallel with the plane of the sections. Noteabsence of pialward dendrites, which were eliminated by the projection algorithm. (C) Electron micrograph of the same cell in the same plane as in Fig. 4A. Besidesthe position and the form of two pialward-directed dendrites (two arrowheads for each), the capillary (Cp) and an unstained cell (asterisk) as landmarks provide asecure identification of the cell. Two VIP+ boutons are visible in close apposition to the soma (b1 and b2). Scale bar, 5 lm (A–C). D–E and F–H show the synapticspecializations of VIP+ boutons b1 and b2, respectively. Rarely a single ultrathin section of the target cells was found, where more than one bouton was visible and inmost cases the synaptic specialization (open arrowheads) was not seen on the same section. Thus it was necessary to follow the boutons in serial sections to the plane,where the synaptic specialization was visible (e.g. Fig. 4E and H). Note the unmistakable black silver granules indicating VIP immunoreaction in the boutons. Scalebar, 0.5 lm ( D–H).
2336 C. David et al.
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2329–2340

VIP innervation of neocortical PV + interneurons 2337
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2329–2340

The heterogeneous innervation pattern reported here suggestsdifferent roles in the cortical circuitry for the members of these twoclusters. The PV + cells of Cluster 1 receive strong innervation andtherefore are very likely to be inhibited by the VIP + cells underappropriate network activity conditions, together with other types ofinterneurons (Staiger et al., 2004). The cells of Cluster 2 receive weakinnervation from the VIP + cell population, thus their inhibitorycontrol could be mediated by other GABAergic cell types as well. TheVIP + interneurons facilitate the excitatory synaptic transmission onpyramidal cells through VIP effects (Haas & Gahwiler, 1992; Cunha-Reis et al., 2004), and connect synaptically to other interneurons in thehippocampus (Acsady et al., 1996), which can be inhibited by theirGABAergic effects (Cunha-Reis et al., 2004). The third group ofinterneurons mentioned in the Introduction containing calbindinreceives a massive innervation from VIP + interneurons (Staigeret al., 2004), thus except for the PV + cells of Cluster 2 most of theinterneurons examined earlier are under the influence of the VIP +system. Even if most interneurons are inhibited due to the VIP + cellactivity, the cells of Cluster 2 could attenuate the pyramidal-activatingand interneuron-inhibiting effect of VIP + cells, which would promotethe basic inhibited state of the cortical column (Welker et al., 1993;Miller et al., 2001)In the primary somatosensory cortex (in rodents also called the
barrel cortex; Woolsey & van der Loos, 1970), the main source ofspecific sensory input is the ventroposteromedial thalamic nucleus(White, 1986). The thalamic fibres in layer IV innervate not only thelocal excitatory neurons (White & Hersch, 1982), but also each of thePV + interneurons (Staiger et al., 1996b). In addition, a substantialportion of the VIP + interneurons in layers II–IV also receiveextensive input from the ventral posteromedial thalamic nucleus(Staiger et al., 1996a). The activation of a barrel-related corticalcolumn occurs only if the thalamic input reaches the excitatory cells inits target layer IV in a synchronized manner (Bruno & Sakmann,2006). In the barrels, a much higher proportion of the inhibitoryinterneurons (� 60%) generate action potentials upon thalamicactivation than do the excitatory ones (< 5%; Porter et al., 2001),which results in a strong and quick damping of the incoming thalamicsignal (Pinto et al., 2003). The PV + neurons can inhibit a largenumber of pyramidal cells in a synchronized manner through theirmultiple perisomatic synapses (Freund, 2003). Although inhibitoryneurotransmission is necessary for the synchronization of corticalnetworks (Cobb et al., 1995; Traub et al., 1996), the inhibitorysynapses between cells belonging to the same type rather desynchro-nize neuronal activity (Gibson et al., 2005), thus the synchronizinginhibitory input should arrive on the PV + cells from inhibitoryneurons of a different type, e.g. VIP + neurons.The VIP + interneurons express postsynaptic nicotinergic acetyl-
choline receptors, and are excited by muscarinic and nicotinergicreceptor agonists, while PV + interneurons are not (Kawaguchi, 1997;Porter et al., 1999). Therefore, they are most probably targets ofnucleus basalis projections, the main source of cholinergic innervationof the neocortex (Struble et al., 1986) In contrast, the PV + cells do notpossess any acetylcholine receptors (Porter et al., 1999), thus they arenot under the direct influence of the cholinergic system (Kawaguchi,1997). The basal nucleus plays a critical role in experience-dependentplasticity (Kilgard & Merzenich, 1998) in the somatosensory cortex(Ego-Stengel et al., 2001) and in the maintenance of attention (Sarteret al., 2005), i.e. higher cortical activity during the examination of newobjects. The size of a whisker representation is also increased afterapplying cholinergic agonists (Penschuck et al., 2002). Nicotinergicagonists, furthermore, enhance thalamocortical synaptic transmission(Gil et al., 1997; Gioanni et al., 1999).
Based on these data we propose the following scheme. The thalamicfibres reaching the barrel cortex innervate both the pyramidal andthe non-pyramidal cells. In case of a synchronized incoming signal thepyramidal cells can be activated before the inhibitory action ofthe PV + cells takes place. Novel environmental information activatesthe basal nucleus, which activates the VIP + interneurons throughcholinergic innervation (see above). Thalamic as well as cholinergicfibres and probably the pyramidal cells as well (Staiger et al., 2002)innervate the VIP + cell population in the barrels, which mayinfluence the cortical activity in at least two ways. (i) The robustVIP + inhibitory innervation of the PV + cells of Cluster 1 maycomplement the activating effect of the thalamic input throughinhibiting the main inhibitors of the excitatory spiny neurons likespiny stellate or pyramidal cells. (ii) It may enhance excitatorysynaptic transmission on pyramidal cells through VIP effects.Therefore the synergistic effect of the sensory input from the thalamusand of the novelty information from the basal nucleus can cause anenhanced activity of the VIP + cells, which results in a temporaryinhibition of the otherwise dominating PV + cell inhibitory activity.The role of the PV + cells of Cluster 2, which have weaker VIP +input, may be to break down the activation chain and bring the corticalcolumn back to a more inhibited state. Our results showed ananatomical basis of this possible mechanism, which would probablyresult in an enhanced activation of the barrels to tactile stimuli.
Acknowledgements
We are most grateful to Ulrich Opfermann-Emmerich for excellent technicalassistance and for his expertise in digital image processing, and to Drs OrsolyaKantor and Akos Kulik for critically reading the manuscript. This work wassupported by DFG (Sta 431 ⁄ 2-4).
Abbreviations
DA, discriminant analysis; DAB, 3,3¢-diaminobenzidine-hydrochloride; FA,factor analysis; GABA, c-aminobutyric acid; HCA, hierarchical clusteranalysis; PB, phosphate buffer; PV, parvalbumin; PV +, parvalbumin-immu-nopositive; TBT, Tris buffer containing 0.3% Triton X-100; VIP, vasoactiveintestinal polypeptide; VIP +, vasoactive intestinal polypeptide-immunopo-sitive.
References
Acsady, L., Gorcs, T.J. & Freund, T.F. (1996) Different populations ofvasoactive intestinal polypeptide-immunoreactive interneurons are specia-lized to control pyramidal cells or interneurons in the hippocampus.Neuroscience, 73, 317–334.
Andrasfalvy, B.K. & Mody, I. (2006) Differences between the scaling ofminiature IPSCs and EPSCs recorded in the dendrites of CA1 mousepyramidal neurons. J. Physiol., 576, 191–196.
Bayraktar, T., Staiger, J.F., Acsady, L., Cozzari, C., Freund, T.F. & Zilles, K.(1997) Co-localization of vasoactive intestinal polypeptide, gamma-amino-butyric acid and choline acetyltransferase in neocortical interneurons of theadult rat. Brain Res., 757, 209–217.
Bayraktar, T., Welker, E., Freund, T.F., Zilles, K. & Staiger, J.F. (2000) Neuronsimmunoreactive for vasoactive intestinal polypeptide in the rat primarysomatosensory cortex: morphology and spatial relationship to barrel-relatedcolumns. J. Comp. Neurol., 420, 291–304.
Bruno, R.M. & Sakmann, B. (2006) Cortex is driven by weak butsynchronously active thalamocortical synapses. Science, 312, 1622–1627.
Cauli, B., Audinat, E., Lambolez, B., Angulo, M.C., Ropert, N., Tsuzuki, K.,Hestrin, S. & Rossier, J. (1997) Molecular and physiological diversity ofcortical nonpyramidal cells. J. Neurosci., 17, 3894–3906.
Cauli, B., Porter, J.T., Tsuzuki, K., Lambolez, B., Rossier, J., Quenet, B. &Audinat, E. (2000) Classification of fusiform neocortical interneuronsbased on unsupervised clustering. Proc. Natl Acad. Sci. USA, 97, 6144–6149.
2338 C. David et al.
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2329–2340

Cauli, B., Tong, X.K., Rancillac, A., Serluca, N., Lambolez, B., Rossier, J. &Hamel, E. (2004) Cortical GABA interneurons in neurovascular coupling:relays for subcortical vasoactive pathways. J. Neurosci., 24, 8940–8949.
Celio, M.R. (1986) Parvalbumin in most gamma-aminobutyric acid-containingneurons of the rat cerebral cortex. Science, 231, 995–997.
Cobb, S.R., Buhl, E.H., Halasy, K., Paulsen, O. & Somogyi, P. (1995)Synchronization of neuronal activity in hippocampus by individualGABAergic interneurons. Nature, 378, 75–78.
Connor, J.R. & Peters, A. (1984) Vasoactive intestinal polypeptide-immunor-eactive neurons in rat visual cortex. Neuroscience, 12, 1027–1044.
Cunha-Reis, D., Sebastiao, A.M., Wirkner, K., Illes, P. & Ribeiro, J.A. (2004)VIP enhances both pre- and postsynaptic GABAergic transmission tohippocampal interneurones leading to increased excitatory synaptic transmis-sion to CA1 pyramidal cells. Br. J. Pharmacol., 143, 733–744.
DeFelipe, J. (1999) Chandelier cells and epilepsy. Brain, 122, 1807–1822.Dombrowski, S.M., Hilgetag, C.C. & Barbas, H. (2001) Quantitative
architecture distinguishes prefrontal cortical systems in the rhesus monkey.Cereb. Cortex, 11, 975–988.
Ego-Stengel, V., Shulz, D.E., Haidarliu, S., Sosnik, R. & Ahissar, E. (2001)Acetylcholine-dependent induction and expression of functional plasticity inthe barrel cortex of the adult rat. J. Neurophysiol., 86, 422–437.
Ekstrand, J.J., Domroese, M.E., Feig, S.L., Illig, K.R. & Haberly, L.B. (2001)Immunocytochemical analysis of basket cells in rat piriform cortex. J. Comp.Neurol., 434, 308–328.
Freund, T.F. (2003) Interneuron diversity series: rhythm and mood inperisomatic inhibition. Trends Neurosci., 26, 489–495.
Froemke, R.C., Poo, M.M. & Dan, Y. (2005) Spike-timing-dependent synapticplasticity depends on dendritic location. Nature, 434, 221–225.
Gibson, J.R., Beierlein, M. & Connors, B.W. (2005) Functional properties ofelectrical synapses between inhibitory interneurons of neocortical layer 4.J. Neurophysiol., 93, 467–480.
Gil, Z., Connors, B.W. & Amitai, Y. (1997) Differential regulation ofneocortical synapses by neuromodulators and activity. Neuron, 19, 679–686.
Gioanni, Y., Rougeot, C., Clarke, P.B.S., Lepouse, C., Thierry, A.M. & Vidal,C. (1999) Nicotinic receptors in the rat prefrontal cortex: increase inglutamate release and facilitation of mediodorsal thalamo-cortical transmis-sion. Eur. J. Neurosci., 11, 18–30.
Gonchar, Y. & Burkhalter, A. (1997) Three distinct families of GABAergicneurons in rat visual cortex. Cereb. Cortex, 7, 347–358.
Gorcs, T.J., Leranth, C. & Maclusky, N.J. (1986) The use of gold-substitutedsilver-intensified diaminobenzidine (DAB) and non-intensified DAB forsimultaneous electron microscopic immunoperoxidase labeling of tyrosinehydroxylase and glutamic acid decarboxylase immunoreactivity in the ratmedial preoptic area. J. Histochem. Cytochem., 34, 1439–1447.
Gulyas, A.I., Gorcs, T.J. & Freund, T.F. (1990) Innervation of different peptide-containing neurons in the hippocampus by GABAergic septal afferents.Neuroscience, 37, 31–44.
Gulyas, A.I., Miles, R., Sik, A., Toth, K., Tamamaki, N. & Freund, T.F. (1993)Hippocampal pyramidal cells excite inhibitory neurons through a singlerelease site. Nature, 366, 683–687.
Haas, H.L. & Gahwiler, B.H. (1992) Vasoactive intestinal polypeptidemodulates neuronal excitability in hippocampal slices of the rat.Neuroscience, 47, 273–277.
Kawaguchi, Y. (1995) Physiological subgroups of nonpyramidal cells withspecific morphological characteristics in layer II ⁄ III of rat frontal cortex. J.Neurosci., 15, 2638–2655.
Kawaguchi, Y. (1997) Selective cholinergic modulation of cortical GABAergiccell subtypes. J. Neurophysiol., 78, 1743–1747.
Kawaguchi, Y. & Kondo, S. (2002) Parvalbumin, somatostatin and cholecys-tokinin as chemical markers for specific GABAergic interneuron types in therat frontal cortex. J. Neurocytol., 31, 277–287.
Kawaguchi, Y. & Kubota, Y. (1996) Physiological and morphologicalidentification of somatostatin- or vasoactive intestinal polypeptide-containingcells among GABAergic cell subtypes in rat frontal cortex. J. Neurosci., 16,2701–2715.
Kawaguchi, Y. & Kubota, Y. (1997) GABAergic cell subtypes and theirsynaptic connections in rat frontal cortex. Cereb. Cortex, 7, 476–486.
Kawaguchi, Y. & Kubota, Y. (1998) Neurochemical features and synapticconnections of large physiologically-identified GABAergic cells in the ratfrontal cortex. Neuroscience, 85, 677–701.
Keros, S. & Hablitz, J.J. (2005) Ectopic action potential generation in corticalinterneurons during synchronized GABA responses. Neuroscience, 131,833–842.
Kilgard, M.P. & Merzenich, M.M. (1998) Cortical map reorganization enabledby nucleus basalis activity. Science, 279, 1714–1718.
Krimer, L.S. & Goldman-Rakic, P.S. (2001) Prefrontal microcircuits:membrane properties and excitatory input of local, medium, and wide arborinterneurons. J. Neurosci., 21, 3788–3796.
Kubota, Y., Hattori, R. & Yui, Y. (1994) Three distinct subpopulations ofGABAergic neurons in rat frontal agranular cortex. Brain Res., 649, 159–173.
Maddock, L. & Young, J.Z. (1987) Quantitative differences among the brainsof cephalopods. J. Zool. Lond., 212, 739–767.
Magistretti, P.J., Cardinaux, J.R. & Martin, J.L. (1998) VIP and PACAP in theCNS: regulators of glial energy metabolism and modulators of glutamatergicsignaling. Ann. N.Y. Acad. Sci., 865, 213–225.
Miller, K.D., Pinto, D.J. & Simons, D.J. (2001) Processing in layer 4 of theneocortical circuit: new insights from visual and somatosensory cortex. Curr.Opin. Neurobiol., 11, 488–497.
Patz, S., Grabert, J., Gorba, T., Wirth, M.J. & Wahle, P. (2004) Parvalbuminexpression in visual cortical interneurons depends on neuronal activity andTrkB ligands during an early period of postnatal development. Cereb.Cortex, 14, 342–351.
Penschuck, S., Chen-Bee, C.H., Prakash, N. & Frostig, R.D. (2002) In vivomodulation of a cortical functional sensory representation shortly aftertopical cholinergic agent application. J. Comp. Neurol., 452, 38–50.
Pinto, D.J., Hartings, J.A. & Simons, D.J. (2003) Cortical damping: analysis ofthalamocortical response transformations in rodent barrel cortex. Cereb.Cortex, 13, 33–44.
Porter, J.T., Cauli, B., Staiger, J.F., Lambolez, B., Rossier, J. & Audinat, E.(1998) Properties of bipolar VIPergic interneurons and their excitation bypyramidal neurons in the rat neocortex. Eur. J. Neurosci., 10, 3617–3628.
Porter, J.T., Cauli, B., Tsuzuki, K., Lambolez, B., Rossier, J. & Audinat, E.(1999) Selective excitation of subtypes of neocortical interneurons bynicotinic receptors. J. Neurosci., 19, 5228–5235.
Porter, J.T., Johnson, C.K. & Agmon, A. (2001) Diverse types of interneuronsgenerate thalamus-evoked feedforward inhibition in the mouse barrel cortex.J. Neurosci., 21, 2699–2710.
Sarter, M., Hasselmo, M.E., Bruno, J.P. & Givens, B. (2005) Unraveling theattentional functions of cortical cholinergic inputs: interactions betweensignal-driven and cognitive modulation of signal detection. Brain Res. BrainRes. Rev., 48, 98–111.
Sessler, F.M., Grady, S.M., Waterhouse, B.D. & Moises, H.C. (1991)Electrophysiological actions of VIP in rat somatosensory cortex. Peptides,12, 715–721.
Somogyi, P. (1977) A specific ‘axo-axonal’ interneuron in the visual cortex ofthe rat. Brain Res., 136, 345–350.
Somogyi, P., Nunzi, M.G., Gorio, A. & Smith, A.D. (1983) A new type ofspecific interneuron in the monkey hippocampus forming synapsesexclusively with the axon initial segments of pyramidal cells. Brain Res.,259, 137–142.
Staiger, J.F., Freund, T.F. & Zilles, K. (1997) Interneurons immunoreactive forvasoactive intestinal polypeptide (VIP) are extensively innervated byparvalbumin-containing boutons in rat primary somatosensory cortex. Eur.J. Neurosci., 9, 2259–2268.
Staiger, J.F., Masanneck, C., Schleicher, A. & Zuschratter, W. (2004)Calbindin-containing interneurons are a target for VIP-immunoreactivesynapses in rat primary somatosensory cortex. J. Comp. Neurol., 468,179–189.
Staiger, J.F., Schubert, D., Zuschratter, W., Kotter, R., Luhmann, H.J. & Zilles,K. (2002) Innervation of interneurons immunoreactive for VIP byintrinsically bursting pyramidal cells and fast-spiking interneurons ininfragranular layers of juvenile rat neocortex. Eur. J. Neurosci., 16, 11–20.
Staiger, J.F., Zilles, K. & Freund, T.F. (1996a) Innervation of VIP-immunoreactive neurons by the ventroposteromedial thalamic nucleus inthe barrel cortex of the rat. J. Comp. Neurol., 367, 194–204.
Staiger, J.F., Zilles, K. & Freund, T.F. (1996b) Distribution of GABAergicelements postsynaptic to ventroposteromedial thalamic projections in layerIV of rat barrel cortex. Eur. J. Neurosci., 8, 2273–2285.
Struble, R.G., Lehmann, J., Mitchell, S.J., McKinney, M., Price, D.L.,Coyle, J.T. & DeLong, M.R. (1986) Basal forebrain neurons providemajor cholinergic innervation of primate neocortex. Neurosci. Lett., 66,215–220.
Sun, Q.Q., Prince, D.A. & Huguenard, J.R. (2003) Vasoactive intestinalpolypeptide and pituitary adenylate cyclase-activating polypeptide activatehyperpolarization-activated cationic current and depolarize thalamocorticalneurons in vitro. J. Neurosci., 23, 2751–2758.
Szabadics, J., Varga, C., Molnar, G., Olah, S., Barzo, P. & Tamas, G. (2006)Excitatory effect of GABAergic axo-axonic cells in cortical microcircuits.Science, 311, 233–235.
VIP innervation of neocortical PV + interneurons 2339
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2329–2340

Szentagothai, J. (1975) The ‘module-concept’ in cerebral cortex architecture.Brain Res., 95, 475–496.
Toledo-Rodriguez, M., Goodman, P., Illic, M., Wu, C.Z. & Markram, H. (2005)Neuropeptide and calcium-binding protein gene expression profiles predictneuronal anatomical type in the juvenile rat. J. Physiol. (Lond.), 567, 401–413.
Traub, R.D., Whittington, M.A., Colling, S.B., Buzsaki, G. & Jefferys, J.G.R.(1996) Analysis of gamma rhythms in the rat hippocampus in vitro and invivo. J. Physiol. (Lond.), 493, 471–484.
Welker, E., Armstrong-James, M.A., van der Loos, H. & Kraftsik, R. (1993)The mode of activation of a barrel column: response properties of singleunits in the somatosensory cortex of the mouse upon whisker deflection. Eur.J. Neurosci., 5, 691–712.
White, E.L. (1986) Termination of thalamic afferents in the cerebral cortex. InJones, E.G. & Peters, A. (Eds), Sensory-Motor Areas and Aspects of CorticalConnectivity. Plenum Press, New York, pp. 271–289.
White, E.L. & Hersch, S.M. (1982) A quantitative study of thalamocortical andother synapses involving the apical dendrites of corticothalamic projectioncells in mouse SmI cortex. J. Neurocytol., 11, 137–157.
Williams, S.R. & Stuart, G.J. (2003) Voltage- and site-dependent controlof the somatic impact of dendritic IPSPs. J. Neurosci., 23, 7358–7367.
Woolsey, T.A. & van der Loos, H. (1970) The structural organization of layerIV in the somatosensory region (SI) of mouse cerebral cortex. Brain Res., 17,205–242.
2340 C. David et al.
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2329–2340