calcium ion signaling in astrocytes
TRANSCRIPT

Chapter 8 Calcium Ion Signaling in Astrocytes
Joachim W. Deitmer , Karthika Singaravelu , and Christian Lohr
8.1 Introduction
Ca 2+ signaling has been recognized as one of the major second messenger steps in most cell types, including astrocytes, the major macroglial cell type in vertebrate nervous systems. Astrocytes are by no means a homogeneous group of glial cells, but comprise a number of different cell types (see Chap. 1). However, in contrast to a decade ago, when mammalian astrocytes were divided into either protoplasmic type 1 or fibrous type II astrocytes, we assume today that there are many types of astrocytes in different brain regions. Another classification has recognized astro-cytes with a dense distribution of glutamate uptake transporters (EAAT, excitatory amino acid transporter) and poor equipment of ionotropic glutamate receptors, while another type of astrocytes shows a poor expression of EAATs, but prominent distribution of ionotropic glutamate receptors. As with all of these cell type
V. Parpura and P.G. Haydon (eds.), Astrocytes in (Patho)Physiology of the Nervous System, 201DOI: 10.1007/978-0-387-79492-1_8, © Springer Science + Business Media, LLC 2009
J.W. DeitmerAbteilung für Allgemeine Zoologie , FB Biologie, TU Kaiserslautern, Kaiserslautern, Germany. [email protected]
Contents
8.1 Introduction ................................................................................................................... 2018.2 Modes and Mechanisms of Ca2+ Signaling ................................................................... 2028.3 Spontaneous Ca2+ Transients and Oscillations .............................................................. 2068.4 Propagation of Ca2+ Signals .......................................................................................... 2088.5 Ca2+ Responses to Transmitters and Other Signaling Molecules .................................. 2108.6 Ca2+ Responses to Neuronal Activity ............................................................................ 2118.7 Store-Operated Ca2+ Entry and Ca2+ Store Refi lling ..................................................... 2138.8 Ca2+-Induced Release of Gliotransmitters ..................................................................... 2158.9 Functional Signifi cance of Ca2+ Signaling .................................................................... 2168.10 Summary and Conclusion ............................................................................................. 217References ................................................................................................................................ 218Abbreviations ........................................................................................................................... 224

202 J.W. Deitmer et al.
classifications established so far, there are known exemptions, such as the Bergmann glial cells in the cerebellum. Bergmann glia is a radial type of macroglial cell, which is a specialized astrocyte, which has both EAATs and ionotropic alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA)-type glutamate receptors. There are radial-type astrocytes also in the developing cortex, which extend from the ventricular surface to the pial surface. Furthermore, Müller glial cells, the principal glial cell type in the retina, extend across the entire retina from the photoreceptors to the inner retinal surface. Thus, the classification of different types of astrocytes is still in its infancy, and we need to know much more about this major cell type in the brain in order to create a useful taxonomy taking into account the various predominant functions of astrocytes in a given brain region.
Most, if not all, of these different types of astrocytes share the property that many of their cellular functions are related to cytosolic Ca 2+ signaling. The single and repetitive rises of cytosolic Ca 2+ play a complex role for initiating intracellular sign-aling cascades, modulating astrocytic functions and intercellular interaction. Most of these functions and interactions involve other neighboring astrocytes and neurons, but there are also signaling pathways to oligodendrocytes and microglial cells. Astrocytes are endowed with a large number of metabotropic receptors in their cell membrane, most of which are coupled to the release of Ca 2+ from the endoplasmic reticulum (ER) via phospholipase C (PLC)-mediated formation of inositol-trisphosphate (IP
3 ).
Astrocytic Ca 2+ signaling can be a single Ca 2+ transient with or without a shoulder or plateau phase, repetitive Ca 2+ transients, so-called Ca 2+ oscillations, or irregular Ca 2+ rises, depending on the species of primary messenger (neurotransmitter, hormone, growth factor) and its concentration. These Ca 2+ signals may spread along the cell and across cell boundaries to neighboring astrocytes in form of Ca 2+ waves, and can be evoked or modulated by neuronal activity. The spatial and temporal properties of these Ca 2+ signaling modes reflect the versatility of this intracellular messenger system. The signaling pathway leading to a cytosolic Ca 2+ rise, common to many cell types, may be regarded as a type of excitation in electrically nonexcitable cells like astrocytes. Cytosolic Ca 2+ transients may initiate Ca 2+ -dependent release of transmitters (gliotransmitters), affecting neuronal excitability or vasoconstriction/vasodilation of blood vessels in the brain. This chapter reviews types of cytosolic Ca 2+ signaling in astrocytes, their different modes of initiation, and their functional significance for astrocytes and the glia–neuron communication.
8.2 Modes and Mechanisms of Ca 2+ Signaling
Ca 2+ signaling in both excitable and nonexcitable cells is based on the maintenance of a low resting concentration of cytosolic Ca 2+ (<150 nM), as compared with the extracellular compartment (1–2 mM), and intracellular Ca 2+ storage compartments (0.1–1 mM). This differential distribution of Ca 2+ creates a gradient across the membrane of intracellular organelles (ER, mitochondria, calcisomes, lysosomes) and the plasma membrane. Cellular Ca 2+ signaling, manifested as rapid, reversible, and often repeated,

8 Calcium Ion Signaling in Astrocytes 203
intracellular Ca 2+ rises, may therefore result from several Ca 2+ sources. Cytosolic Ca 2+ levels can increase via Ca 2+ influx from the extracellular space across the plasma membrane via Ca 2+ -permeable ion channels, and by Ca 2+ release from intra-cellular stores (Fig. 8.1 ). Calcium ion influx may occur through specific Ca 2+ -permeable, ligand-gated channels, voltage-gated Ca 2+ channels that are activated by membrane depolarization, and store-operated Ca 2+ entry channels that are activated following the depletion of intracellular Ca 2+ stores. At lowered extracellular K + con-centrations, inward rectifier K + channels can become permeable to divalent cations including Ca 2+ , and this novel type of Ca 2+ influx has been observed in astrocytes at K + concentration of 2 mM and below (Dallwig et al., 2000 ; Härtel et al., 2007) .
While electrically excitable cells use the electrochemical gradient across the plasma membrane to effectively increase the intracellular Ca 2+ levels due to the high expression of specific ligand- and voltage-gated ion channels that allow prominent influx of Ca 2+ ions, nonelectrically excitable cells like astrocytes use the Ca 2+ stored in the ER as the main source for cytoplasmic Ca 2+ signaling (reviewed in Deitmer et al., 1998 ; Verkhratsky et al., 1998) . Ionotropic kainate/AMPA receptors, known to be expressed by Bergmann glial cells in the cerebellum, lack the GluR2 subunit, which renders them permeable to Ca 2+ (Müller et al., 1992; Burnashev et al., 1992) . However, Ca 2+ influx through AMPA receptors does not seem to play a major role
Fig. 8.1 Mechanisms of Ca 2+ signaling in astrocytes. Ca 2+ can enter the cell through voltage-operated channels (VOC), ligand-operated channels (LOC), and store-operated cannels (SOC). Activation of G-protein-coupled receptors results in phospholipase C (PLC)-mediated production of IP
3 , which leads to release of Ca 2+ from intracellular stores such as the endoplasmic reticulum
(ER) via IP 3 receptor channels. Ca 2+ is transported out of the cytoplasm into the ER by sarcoplas-
mic-endoplasmic reticulum Ca 2+ -ATPases (SERCA), and into the extracellular space by plasma-lemmal Ca 2+ -ATPases and a sodium-Ca 2+ exchanger (NCX).

204 J.W. Deitmer et al.
in astrocyte Ca 2+ signaling, because in most brain regions, astrocytes express AMPA receptors that contain the GluR2 subunit and hence display a low Ca 2+ permeability (Backus and Berger, 1995 ; Seifert et al., 2003) . Recent studies have also described the presence of ionotropic glutamate receptors of the N-methyl-D-aspartate (NMDA) type in astrocytes in situ, which provide another pathway for Ca 2+ influx (Schipke et al., 2001 ; Lalo et al., 2006 ; Bigini et al., 2006) . Activation of other ligand-gated channels permeable to Ca 2+ such as ionotropic purinergic receptors (P2X receptors; Walz et al., 1994 ; James and Butt, 2001) , or nicotinic acetylcholine receptors con-taining the a7 subunit, has been shown to lead to the gating of cationic conductances and an influx of Ca 2+ in cell culture preparations (Sharma and Vijayaraghavan, 2001) . Ca 2+ influx through ligand-gated channels, however, may not be a major pathway for astrocyte cytosolic Ca 2+ signaling.
Calcium ion influx through voltage-gated Ca 2+ channels in the plasma membrane constitutes another source for intracellular Ca 2+ elevation in different cell types. Astrocytes in culture express a variety of voltage-gated channels, including Ca 2+ channels (Duffy and MacVicar, 1994) ; however, studies performed in situ indicate that functional voltage-gated Ca 2+ channels are mainly expressed by immature astrocytes, if at all (Carmignoto et al., 1998) . In invertebrate glial cells, functional voltage-dependent Ca 2+ channels appear to be the rule, and such Ca 2+ influx may be
Table 8.1 Some receptor ligands capable of evoking Ca 2+ signaling in astrocytes
Ligand Reference
5-Hydroxytryptamine Dave et al., 1991 ; Nilsson et al., 1991 ; Shelton and McCarthy, 2000
Acetylcholine Shao and McCarthy, 1995 ; Sharma and Vijayaraghavan, 2001
Adenosine Peakman and Hill, 1995 ; Porter and McCarthy, 1995a Angiotensin II Wang et al., 1996 ATP P2Y Kastritsis et al., 1992 ; Ho et al., 1995 P2X James and Butt, 2001 Bradykinin Gimpl et al., 1992 Dopamine Parpura and Haydon, 2000 GABA Nilsson et al., 1993 Glutamate AMPA/kainate Enkvist et al., 1989 ; Müller et al., 1992 ; Burnashev et al., 1992 mGluR Jensen and Chiu, 1991 ; Porter and McCarthy, 1995b Histamine Inagaki et al., 1991 ; Shelton and McCarthy, 2000 Met-enkephalin Stiene-Martin et al., 1993 Neuropeptide Y Gimpl et al., 1993 Noradrenaline (norepinephrine) Salm and McCarthy, 1990 ; Duffy and MacVicar, 1995 Tachykinins Delumeau et al., 1991 Thrombin Czubayko and Reiser, 1995 Vasopressin, oytocin Jurzak et al., 1995

8 Calcium Ion Signaling in Astrocytes 205
initiated by depolarizing neurotransmitters (Lohr and Deitmer, 1999, 2006 ; Lohr et al., 2005) . Therefore, unlike neurons and other electrically excitable cells that express a high density of ligand- and voltage-gated channels, which are permeable to Ca 2+ and allow an efficient rise of cytosolic Ca 2+ , astrocytes in the mammalian nervous system usually employ different cellular mechanisms to allow Ca 2+ entry and an increase in their cytosolic Ca 2+ concentration.
Astrocytes express a variety of functional receptors for many neurotransmit-ters, neuromodulators, peptides, etc. This includes glutamate, norepinephrine, gamma-aminobutyric acid (GABA), histamine, adenosine 5¢-triphosphate (ATP), adenosine diphosphate (ADP), adenosine, and acetylcholine (also see later and Table 8.1 ), most of which belong to the metabotropic receptor family. These receptors are associated with G proteins that can stimulate phospholipase C activ-ity, which in turn leads to the formation of the intracellular second messengers IP
3 and diacylglycerol (DAG). Activation of specific, IP
3 -gated receptor channels
in the ER results in release of Ca 2+ from these Ca 2+ stores. This leads to an increase in the cytosolic Ca 2+ concentration, which usually encompasses the entire cell soma and often also cell processes. This mechanism of eliciting an astrocytic Ca 2+ signal represents one important mode of neuron-to-astrocyte com-munication, in which the astrocyte Ca 2+ response is evoked by neurotransmitter(s) released during synaptic activity, activating one or several of the many metabo-tropic receptors in astrocytes.
In addition to IP 3 receptors, the membrane of the endoplasmic reticulum of
most cells also contains ryanodine receptors, a different type of Ca 2+ -permeable receptor that is activated by cytosolic Ca 2+ and/or cyclic ADP-ribose. A rise of the cytosolic Ca 2+ leads to the opening of the ryanodine receptors and hence induces the release of Ca 2+ from the internal stores, a process known as Ca 2+ -induced Ca 2+ release (Sitsapesan et al., 1995 ; Verkhratsky and Shmigol, 1996) . Ryanodine receptors can also be activated by caffeine (Meissner and Henderson, 1987 ; Liu et al., 1989) . The presence of these receptors in astrocytes, however, is controversial; moreover, the lack of caffeine-sensitive Ca 2+ release, as found in astrocytes in culture and in situ, and in contrast to neurons, suggests that the role of these receptors for astrocyte Ca 2+ signaling is either restricted to defined brain areas, or plays a minor role in astrocytes in general (Beck et al., 2004) .
The maintenance of the high intraluminal Ca 2+ concentration in the ER depends on Ca 2+ -ATPases in the ER membrane, which efficiently pump Ca 2+ from the cytosol into the ER, and thereby maintains a high concentration of Ca 2+ by constant refilling of these stores. Since Ca 2+ is also transported out of the cell by Ca 2+ -ATPases and Na + /Ca 2+ exchange carriers in the plasma membrane, Ca 2+ store refilling requires some influx of Ca 2+ from the extracellular compartment, in particular following evoked Ca 2+ release from stores. Ca 2+ entry is using specific Ca 2+ -permeable plasma membrane channels, called store-operated channels, that are regulated by the filling state of the ER and that serve to its replenishment with Ca 2+ . This store-operated Ca 2+ entry (SOCE), also called capacitative Ca 2+ entry, has been observed in many

206 J.W. Deitmer et al.
different cell types and recorded electrophysiologically as a persistent current, called Ca 2+ release-activated current ( I
CRAC ; Hoth and Penner, 1992) . Although the
presence of I CRAC
has not yet been identified in astrocytes, SOCE has been clearly demonstrated in these cells by Ca 2+ imaging studies (Jung et al., 2000 ; Lo et al., 2002 ; Singaravelu et al., 2006) . Because of the importance of this type of Ca 2+ entry in astrocytes, this will be discussed in more detail in Sect. 8.7.
It is the interplay of these different cellular mechanisms, Ca 2+ influx and release, Ca 2+ extrusion and uptake, and Ca 2+ buffering, that controls the cytosolic and organellar Ca 2+ levels, which are crucial for the generation and the shape of astro-cyte Ca 2+ signaling. The spatial organization of Ca 2+ influx may create Ca 2+ micro-domains, i.e., localized regions of Ca 2+ rises, usually near the plasma membrane (Berridge, 2006 ; Oheim et al., 2006) . Mechanisms that are involved in regulating cytosolic Ca 2+ contribute to shaping the kinetics and amplitude of Ca 2+ transients, such as, e.g., Ca 2+ -binding proteins like parvalbumin, calcineurin, and calmodulin that are present in the cytoplasm may serve to buffer cytosolic Ca 2+ concentration. However, this is a low-capacity system that is more effective as mediator of Ca 2+ -dependent cellular processes. Energy-consuming mechanisms are involved in the effective Ca 2+ clearance. Sarco-endoplasmic reticulum Ca 2+ -ATPases (SERCAs) pump Ca 2+ ions into the endoplasmic reticulum using ATP as energy source. Ca 2+ -ATPases and Na + /Ca 2+ exchangers present in the plasma membrane use ATP or the electrochemical gradient of Na + across the plasma membrane, respectively, to extrude Ca 2+ against a steep concentration gradient across the plasma membrane. Finally, mitochondria are dynamic Ca 2+ stores that sequester Ca 2+ through the activity of the Ca 2+ uniporter, with a relatively low affinity, but a large capacity, to store Ca 2+ transiently, and protect the cell from cytosolic Ca 2+ overload, which may lead to cell death. Although other Ca 2+ storing organelles, such as calcisomes and lysosomes, have been reported to contribute to cytosolic Ca 2+ signaling and homeostasis in some cell types, their role in astrocytes has remained largely unknown.
8.3 Spontaneous Ca 2+ Transients and Oscillations
Ca 2+ oscillations are defined as repetitive rises of cytosolic Ca 2+ and may occur intermittently with single Ca 2+ transients and with intervals of up to several minutes (Fig. 8.2 ). Spontaneous Ca 2+ transients and oscillations have been reported in both neurons and glial cells, in culture, in situ, and in vivo (Parri and Crunelli, 2001 ; Nett et al., 2002 ; Morita et al., 2003 ; Zur Nieden and Deitmer, 2006) . Ca 2+ signals can propagate as waves along cell processes and even beyond cell boundaries to neigh-boring glial cells. Ca 2+ transients and Ca 2+ oscillations are usually attributable to release of Ca 2+ from intracellular stores, and can be blocked by inhibiting the Ca 2+ -ATPase of the endoplasmic reticulum (by cyclopiazonic acid or thapsigaragin), and/or by IP
3 receptor antagonists (e.g., 2-APB). These spontaneous Ca 2+ transients
and oscillations occurred in 35% of astrocytes in the hippocampus of juvenile rats, and in 82% of hippocampal astrocytes in juvenile mice, both with a rate of about
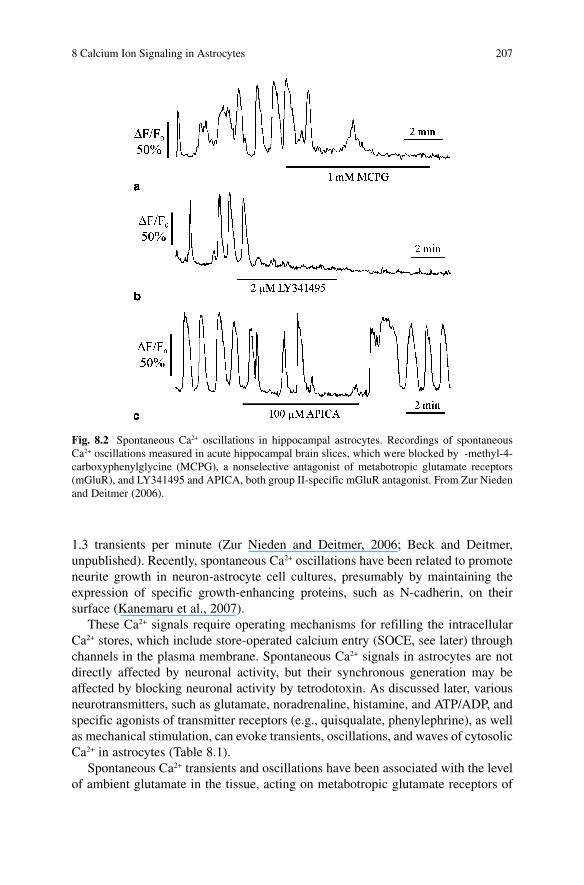
8 Calcium Ion Signaling in Astrocytes 207
1.3 transients per minute (Zur Nieden and Deitmer, 2006 ; Beck and Deitmer, unpublished). Recently, spontaneous Ca 2+ oscillations have been related to promote neurite growth in neuron-astrocyte cell cultures, presumably by maintaining the expression of specific growth-enhancing proteins, such as N-cadherin, on their surface (Kanemaru et al., 2007) .
These Ca 2+ signals require operating mechanisms for refilling the intracellular Ca 2+ stores, which include store-operated calcium entry (SOCE, see later) through channels in the plasma membrane. Spontaneous Ca 2+ signals in astrocytes are not directly affected by neuronal activity, but their synchronous generation may be affected by blocking neuronal activity by tetrodotoxin. As discussed later, various neurotransmitters, such as glutamate, noradrenaline, histamine, and ATP/ADP, and specific agonists of transmitter receptors (e.g., quisqualate, phenylephrine), as well as mechanical stimulation, can evoke transients, oscillations, and waves of cytosolic Ca 2+ in astrocytes (Table 8.1 ).
Spontaneous Ca 2+ transients and oscillations have been associated with the level of ambient glutamate in the tissue, acting on metabotropic glutamate receptors of
Fig. 8.2 Spontaneous Ca 2+ oscillations in hippocampal astrocytes. Recordings of spontaneous Ca 2+ oscillations measured in acute hippocampal brain slices, which were blocked by -methyl-4-carboxyphenylglycine (MCPG), a nonselective antagonist of metabotropic glutamate receptors (mGluR), and LY341495 and APICA, both group II-specific mGluR antagonist. From Zur Nieden and Deitmer (2006).

208 J.W. Deitmer et al.
group I and II in the astrocyte membrane (Zur Nieden and Deitmer, 2006) . Changes in ambient glutamate concentration, which are not due to synaptically released glutamate, can, e.g., occur via the cystine-glutamate exchanger (x(c) - or xCT; Ye et al., 1999 ; Shih et al., 2006) or reversed glutamate uptake under pathophysiological conditions (Rossi et al., 2000 ; Allen and Attwell, 2004) . Thus, the frequency of these spontaneous Ca 2+ signals may help to sense the level of extracellular gluta-mate in nervous tissue. Spontaneous Ca 2+ signals itself may induce Ca 2+ -dependent gliotransmitter release (see later), and this might in turn contribute to the ambient glutamate concentration, although this mechanism may be only in action under certain conditions (Pasti et al., 1995, 2001) . It does not seem likely at this point to hypothesize that each astrocytic Ca 2+ signal leads to the release of transmitter. In particular, with the propagation of Ca 2+ transients across the cell processes, the localization of Ca 2+ -dependent gliotransmitter release is still obscure. The elabora-tion of mechanisms and functions of spontaneous Ca 2+ transients and oscillations in astrocytes, and their mode of initiation and maintenance still require more experi-mental analysis.
8.4 Propagation of Ca 2+ Signals
The cytosolic Ca 2+ signals in astrocytes can adopt different modes of temporal and spatial patterns that can be modulated in amplitude, duration, and frequency by different stimuli from other cells. Synaptic activity and the release of neurotrans-mitters and modulators can be functionally important initiators for these glial Ca 2+ signals, which can propagate within and between astrocytes, signaling to different regions of the cell and to different cells. The astrocyte Ca 2+ signal, either spontane-ously generated or evoked by synaptic activity, can be initiated in spatially restricted areas called microdomains (Simpson and Russell, 1997 ; Oheim et al., 2006) . These are often located in the fine, extensive cellular processes, from where the Ca 2+ signals can propagate along the processes to other regions of the astrocyte generating a wave of intracellular Ca 2+ . Elementary events are initiated by a locally restricted group of Ca 2+ channels in the plasma membrane or in the ER (Berridge, 2006) . The spread of these Ca 2+ transients along the cell processes and to neighboring cells can occur with a speed of 5–25 µm s −1 (Peters et al., 2003 ; Fiacco and McCarthy, 2006) . A thorough characterization of the intra- and intercellular Ca 2+ waves, which can be under the control of released modulators and transmitters, still needs to be accomplished.
The intracellular compartmentalization and the control of the propagation of the Ca 2+ signal are highly relevant for cellular functions and the astrocyte-to-neuron communication (Scemes and Giaume, 2006 ; Spät, 2006) . Since gliotransmitters released from astrocytes through Ca 2+ -dependent mechanisms may act as modula-tors of synaptic transmission, the mechanisms controlling the intracellular Ca 2+ wave may not only be of great significance because they will determine the degree of extension of the signal that triggers the neuromodulatory effects of astrocytes,
8 Calcium Ion Signaling in Astrocytes 209
but also because they determine the spatiotemporal response pattern of the cells. Since the Ca 2+ signal evoked by synaptic activity is elicited in restricted regions of the astrocyte, the neuron-to-astrocyte communication is usually a localized event that does not result from uncontrolled spillover of neurotransmitter, but more prob-ably from a spatially highly focussed signaling between synaptic terminals and astrocytic processes (Carmignoto, 2000 ; Fields and Stevens-Graham, 2002) .
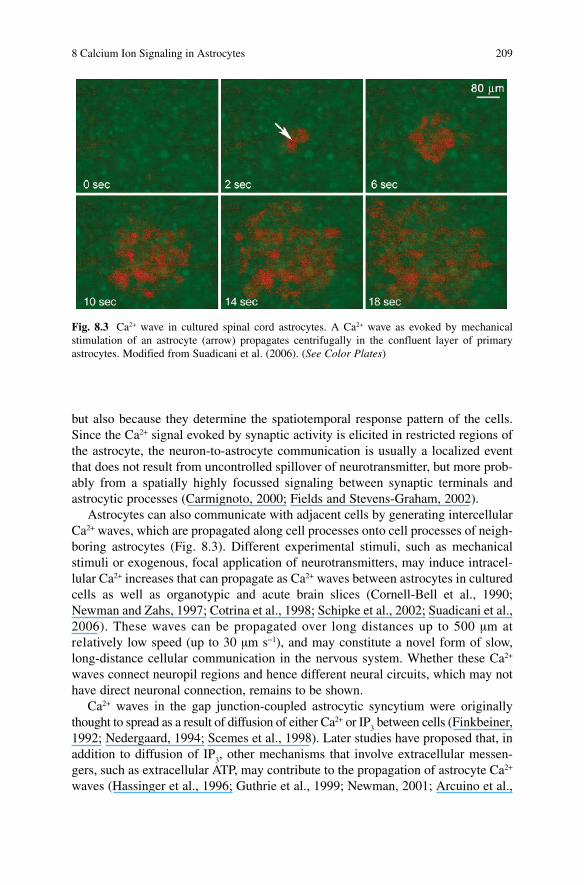
Astrocytes can also communicate with adjacent cells by generating intercellular Ca 2+ waves, which are propagated along cell processes onto cell processes of neigh-boring astrocytes (Fig. 8.3 ). Different experimental stimuli, such as mechanical stimuli or exogenous, focal application of neurotransmitters, may induce intracel-lular Ca 2+ increases that can propagate as Ca 2+ waves between astrocytes in cultured cells as well as organotypic and acute brain slices (Cornell-Bell et al., 1990 ; Newman and Zahs, 1997 ; Cotrina et al., 1998 ; Schipke et al., 2002 ; Suadicani et al., 2006) . These waves can be propagated over long distances up to 500 µm at relatively low speed (up to 30 µm s −1 ), and may constitute a novel form of slow, long-distance cellular communication in the nervous system. Whether these Ca 2+ waves connect neuropil regions and hence different neural circuits, which may not have direct neuronal connection, remains to be shown.
Ca 2+ waves in the gap junction-coupled astrocytic syncytium were originally thought to spread as a result of diffusion of either Ca 2+ or IP
3 between cells (Finkbeiner,
1992 ; Nedergaard, 1994 ; Scemes et al., 1998) . Later studies have proposed that, in addition to diffusion of IP
3 , other mechanisms that involve extracellular messen-
gers, such as extracellular ATP, may contribute to the propagation of astrocyte Ca 2+ waves (Hassinger et al., 1996 ; Guthrie et al., 1999 ; Newman, 2001 ; Arcuino et al.,
Fig. 8.3 Ca 2+ wave in cultured spinal cord astrocytes. A Ca 2+ wave as evoked by mechanical stimulation of an astrocyte (arrow) propagates centrifugally in the confluent layer of primary astrocytes. Modified from Suadicani et al. (2006). (See Color Plates)

210 J.W. Deitmer et al.
2002) . ATP released from active astrocytes appears to be an important mediator of long-range Ca 2+ signaling, whereas shorter range signaling may be mediated by gap junctions (Scemes and Giaume, 2006) . In a model of ATP-mediated propagation of Ca 2+ signals from cell to cell, activation of metabotropic P2Y receptors (probably of the subtype 1) leads to the IP
3 -gated Ca 2+ release from intracellular stores, which
in turn results in the release of ATP to the extracellular space, which then excites neighboring cells. However, there is still some controversy about the mechanisms and the location of ATP release from astrocytes.
8.5 Ca 2+ Responses to Transmitters and Other Signaling Molecules
Cytosolic Ca 2+ transients may be triggered by a variety of signaling molecules, including hormones, growth factors, prostaglandins, and many neurotransmitters. As described earlier, astrocytes express a wide variety of functional receptors for most neurotransmitters, and many of them belong to the metabotropic family, i.e., they are coupled to G proteins and second messenger pathways that lead to the IP
3 -mediated Ca 2+ release from intracellular stores. These Ca 2+ signals can be linked
to initiating cellular activity, such as transmitter release, a rise in the K + conductance of the cell membrane, mitochondrial energy production, or activation of enzymes (see later).
Astrocytes may also respond with intracellular Ca 2+ elevations to other signaling molecules such as chemokines, a class of small proteins that bind to G-protein-coupled receptors, which were originally identified as inflammatory mediators of leukocyte chemotaxis, but that have later been shown to be involved also in other functions beyond neuroinflammation. Various types of chemokine receptors are expressed in cells of the nervous system, including astrocytes, where the chemokine stromal cell-derived factor 1 (SDF-1) has been shown to increase astrocytic Ca 2+ through activation of the receptor CXCR4 (Bajetto et al., 1999) , which can lead to glutamate release from astrocytes (Bezzi et al., 2001) .
The list of ligands that activate the release of Ca 2+ from intracellular stores is steadily growing, and some are listed in Table 8.1 . Since some of these ligands, such as glutamate or ATP, may be released by astrocytes themselves in a Ca 2+ -dependent way, autoactivation of astrocytes might be expected. However, if metabotropic receptors linked to the IP
3 second messenger pathway are clustered and/or are
restricted to defined areas, or more or less uniformly distributed on the cell surface, which would render astrocytes as sensors for ambient concentration of ligands, is not known. So far, specialized regions of receptor clustering, as is found at post-synaptic membrane of neurons, have not yet been described for astrocytes, although such clustering may well occur in cell processes facing synaptic and extrasynaptic release sites.
8 Calcium Ion Signaling in Astrocytes 211
8.6 Ca 2+ Responses to Neuronal Activity
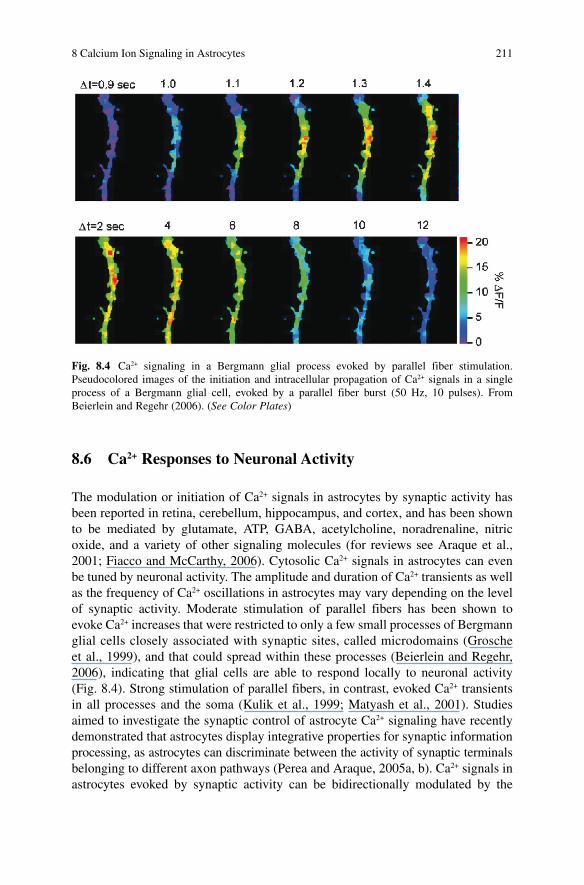
The modulation or initiation of Ca 2+ signals in astrocytes by synaptic activity has been reported in retina, cerebellum, hippocampus, and cortex, and has been shown to be mediated by glutamate, ATP, GABA, acetylcholine, noradrenaline, nitric oxide, and a variety of other signaling molecules (for reviews see Araque et al., 2001 ; Fiacco and McCarthy, 2006) . Cytosolic Ca 2+ signals in astrocytes can even be tuned by neuronal activity. The amplitude and duration of Ca 2+ transients as well as the frequency of Ca 2+ oscillations in astrocytes may vary depending on the level of synaptic activity. Moderate stimulation of parallel fibers has been shown to evoke Ca 2+ increases that were restricted to only a few small processes of Bergmann glial cells closely associated with synaptic sites, called microdomains (Grosche et al., 1999) , and that could spread within these processes (Beierlein and Regehr, 2006) , indicating that glial cells are able to respond locally to neuronal activity (Fig. 8.4 ). Strong stimulation of parallel fibers, in contrast, evoked Ca 2+ transients in all processes and the soma (Kulik et al., 1999 ; Matyash et al., 2001) . Studies aimed to investigate the synaptic control of astrocyte Ca 2+ signaling have recently demonstrated that astrocytes display integrative properties for synaptic information processing, as astrocytes can discriminate between the activity of synaptic terminals belonging to different axon pathways (Perea and Araque, 2005 a, b ). Ca 2+ signals in astrocytes evoked by synaptic activity can be bidirectionally modulated by the
Fig. 8.4 Ca 2+ signaling in a Bergmann glial process evoked by parallel fiber stimulation. Pseudocolored images of the initiation and intracellular propagation of Ca 2+ signals in a single process of a Bergmann glial cell, evoked by a parallel fiber burst (50 Hz, 10 pulses). From Beierlein and Regehr (2006). (See Color Plates)
212 J.W. Deitmer et al.
interaction of different synaptic inputs, being potentiated or depressed depending on the level of synaptic activity. This modulation may also control the intracellular spread of the Ca 2+ signal in form of waves, which may have important consequences on brain function by regulating the spatial extension of a single astrocyte’s impact on different synapses.
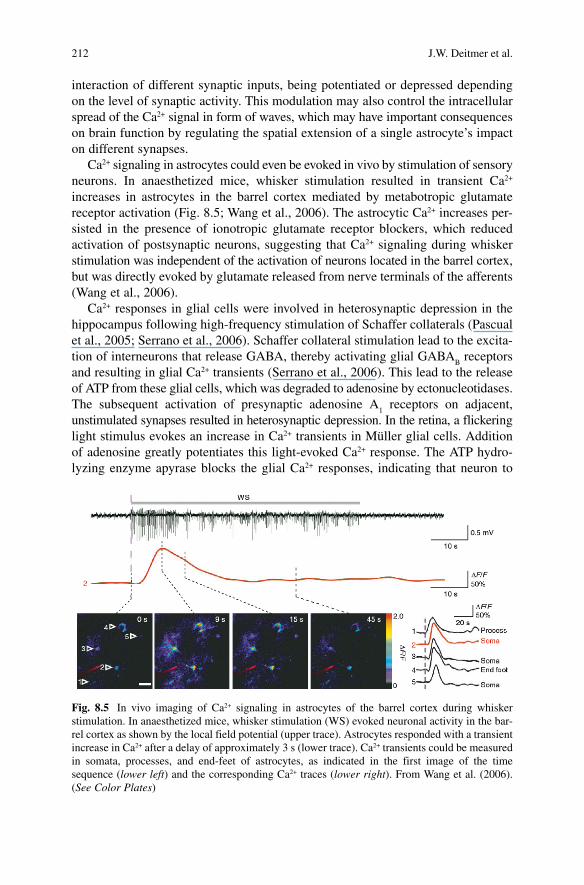
Ca 2+ signaling in astrocytes could even be evoked in vivo by stimulation of sensory neurons. In anaesthetized mice, whisker stimulation resulted in transient Ca 2+ increases in astrocytes in the barrel cortex mediated by metabotropic glutamate receptor activation (Fig. 8.5 ; Wang et al., 2006) . The astrocytic Ca 2+ increases per-sisted in the presence of ionotropic glutamate receptor blockers, which reduced activation of postsynaptic neurons, suggesting that Ca 2+ signaling during whisker stimulation was independent of the activation of neurons located in the barrel cortex, but was directly evoked by glutamate released from nerve terminals of the afferents (Wang et al., 2006) .
Ca 2+ responses in glial cells were involved in heterosynaptic depression in the hippocampus following high-frequency stimulation of Schaffer collaterals (Pascual et al., 2005 ; Serrano et al., 2006) . Schaffer collateral stimulation lead to the excita-tion of interneurons that release GABA, thereby activating glial GABA
B receptors
and resulting in glial Ca 2+ transients (Serrano et al., 2006) . This lead to the release of ATP from these glial cells, which was degraded to adenosine by ectonucleotidases. The subsequent activation of presynaptic adenosine A
1 receptors on adjacent,
unstimulated synapses resulted in heterosynaptic depression. In the retina, a flickering light stimulus evokes an increase in Ca 2+ transients in Müller glial cells. Addition of adenosine greatly potentiates this light-evoked Ca 2+ response. The ATP hydro-lyzing enzyme apyrase blocks the glial Ca 2+ responses, indicating that neuron to
Fig. 8.5 In vivo imaging of Ca 2+ signaling in astrocytes of the barrel cortex during whisker stimulation. In anaesthetized mice, whisker stimulation (WS) evoked neuronal activity in the bar-rel cortex as shown by the local field potential (upper trace). Astrocytes responded with a transient increase in Ca 2+ after a delay of approximately 3 s (lower trace). Ca 2+ transients could be measured in somata, processes, and end-feet of astrocytes, as indicated in the first image of the time sequence ( lower left ) and the corresponding Ca 2+ traces ( lower right ). From Wang et al. (2006). (See Color Plates)

8 Calcium Ion Signaling in Astrocytes 213
glia signaling in the retina is mediated by ATP release from neurons and activation of glial purinergic receptors. On the other hand, Müller cell stimulation can evoke a hyperpolarization in neighboring ganglion cells, which is blocked by the A1 adenosine receptor antagonist DPCPX and is reduced by ecto-ATPase and ectonu-cleotidase inhibitors (Newman, 2005 ; Metea and Newman, 2006) . This suggests that glial cells release ATP, which is converted to adenosine by ectoenzymes, leading to the activation of neuronal adenosine receptors. This demonstrates that there is reciprocal exchange of signals between neurons and glial cells, involving cytosolic Ca 2+ transients and release of transmitters in both types of cells. ATP and its meta-bolites seem to play a prominent and diverse role in mediating different kinds of Ca 2+ signals and their propagation in astrocytes.
8.7 Store-Operated Ca 2+ Entry and Ca 2+ Store Refilling
Store-operated Ca 2+ entry (SOCE) can be triggered by the same mechanisms as those that elicit Ca 2+ release from intracellular stores and thus lead to Ca 2+ store depletion. Because SOCE can be evoked by store depletion, and store refilling is impaired when Ca 2+ entry through SOC channels is prevented, SOCE has been attributed to the refilling of these stores. Although the major function of SOCE is regarded to be the maintenance of Ca 2+ homeostasis of the ER, SOCE-linked cytosolic Ca 2+ transients have been linked to several other Ca 2+ -dependent proc-esses. Among these are vesicular exocytosis, blood vessel contraction and dilation, cell growth and cell death, regulation of enzyme activity, and gene expression (for review see Parekh and Putney, 2005) . There have also been reports linking malfunc-tioning SOCE to Alzheimer’s disease (Putney, 2000 ; Mattson and Chan, 2003) , to severe combined immunodeficiency (Partiseti et al., 1994 ; Feske et al., 2001) , and to acute pancreatitis (Parekh, 2000) .
SOCE can be detected as a sustained, elevated plateau of the intracellular Ca 2+ levels that depends on the previous Ca 2+ depletion of intracellular Ca 2+ stores, and that requires the presence of extracellular Ca 2+ . A putative sensor detects the fall in Ca 2+ content of the stores, which initiates a signaling cascade that relates this infor-mation to specific Ca 2+ -permeable channels in the plasma membrane. The underlying mechanism of this process has remained elusive for over 20 years, but recent studies support a new mechanism for store-operated Ca 2+ signaling (Fig. 8.6 ). In this model, depletion of Ca 2+ stores causes Ca 2+ -independent phospholipase A
2 (iPLA
2 )
to trigger Ca 2+ entry in several cell types including astrocytes (Smani et al., 2004 ; Singaravelu et al., 2006) . A diffusible messenger termed Ca 2+ influx factor (CIF) generated by depleted Ca 2+ stores has been suggested to act through activation of inducible iPLA
2 activity (Smani et al., 2003, 2004) . This enzyme is kept catalytically
inactive by calmodulin, which binds tightly to iPLA 2 under resting conditions. CIF is
proposed to provoke the dissociation of calmodulin and iPLA 2 , which then hydrolyzed
the plasma membrane phospholipids to generate lysophospholipids and arachidonic acid. The specific lysophospholipid products of iPLA
2 , lysophosphotidylcholine

214 J.W. Deitmer et al.
and lysophosphotidylinositol, were shown to trigger SOCE in smooth muscle cells (Smani et al., 2003, 2004) as well as in astrocytes (Singaravelu et al., 2006) , while arachidonic acid, that activates non-SOCE pathways (Shuttleworth and Thompson, 1999) , reduced SOCE in astrocytes (Sergeeva et al., 2003 ; Yang et al., 2005) . The physiological significance of iPLA
2 -mediated Ca 2+ influx for maintaining spontane-
ous Ca 2+ signaling has been demonstrated in cerebellar astrocytes in situ, where the inhibition of iPLA
2 and SOCE channels significantly reduced the occurrence and
frequency of spontaneous Ca 2+ oscillations (Singaravelu et al., 2006) . An exciting advancement in the field of SOCE was provided by the recent iden-
tification of the ER Ca 2+ sensor, STIM1 and the CRAC channel pore subunit, Orai1 (CRACM1). This has enabled significant new insight into the molecular mechanism as to how depletion of Ca 2+ in the ER opens Ca 2+ channels in the plasma membrane. It has been demonstrated that STIM1 was closely associated with the ER at rest, with a N-terminal EF-hand domain facing the lumen of the ER, suggesting the ability to sense the Ca 2+ content of the ER (Roos et al., 2005 ; Liou et al., 2005) . Upon store depletion, STIM1 is rapidly translocated into puncta that accumulated near the plasma membrane and is essential for SOCE (Zhang et al., 2005 ; Liou et al., 2005 ; Roos et al., 2005) . Thus, STIM1 may act as a Ca 2+ influx factor. In addition to being the ER Ca 2+ sensor, STIM1 has also been shown to regulate certain properties of the CRAC channel (Spassova et al., 2005) . Orai1, that codes for a plasma membrane protein with four putative transmembrane-spanning domains, was reported to be an essential pore subunit of the CRAC channel by acting as a Ca 2+ selectivity filter (Vig et al., 2006 ; Prakriya et al., 2006 ; Yeromin et al., 2006) . Combined over-expression of Orai1 and STIM1 in mammalian cells leads to a marked increase in
Fig. 8.6 Proposed mechanism of store-operated Ca 2+ signaling in cerebellar astrocytes. Depletion of intracellular Ca 2+ stores by IP
3 receptor activation leads to the translocation of a Ca 2+ influx
factor (CIF) to membrane-bound Ca 2+ -independent phospholipase A 2 (iPLA
2 ), where CIF releases
the calmodulin (CaM)-dependent block of iPLA 2 . After unblocking iPLA
2 , the enzyme produces
phopholipids such as lysophophoinositol (LPI) and lysophosphocholine (LPC), which would lead to the opening of store-operated Ca 2+ channels (SOC). SERCA sarcoplasmic-endoplasmic reticulum Ca 2+ -ATPase.

8 Calcium Ion Signaling in Astrocytes 215
the CRAC channel activity (Peinelt et al., 2006 ; Soboloff et al., 2006) . Although the involvement of STIM1 and Orai1 may well be of general relevance in cells, the role of either of these proteins for store-operated Ca 2+ signaling in astrocytes or other glial cells still has to be confirmed.
8.8 Ca 2+ -Induced Release of Gliotransmitters
There is growing evidence that elevations of cytosolic Ca 2+ can initiate regulated release of a variety of molecules, including classical transmitters such as glutamate, ATP, and d -serine (Parpura et al, 1994 ; Coco et al., 2003 ; Newman, 2003 ; Mothet et al., 2005 ; Xu et al., 2007) (also see Chaps. 12 and 16). Large rises of cytosolic Ca 2+ as induced by flash photolysis of caged Ca 2+ can trigger the release of these gliotransmitters (Oheim et al., 2006) . Different mechanisms of transmitter release from astrocytes have been postulated: vesicle exocytosis (Bezzi et al., 2004 ; Montana et al., 2006 ; Oheim et al., 2006) , reversed transport (Szatkowski et al., 1990 ; Rossi et al., 2000) , and transmitter efflux through pores such as connexin hemichannels (Ye et al., 2003) , purinergic P2X
7 receptors (Duan and Neary, 2006) ,
and anion channels (Kimelberg et al., 2006) . Many proteins belonging to the exocytotic machinery, including the glutamate
accumulating transporters VGLUT-1 and VGLUT-2, and the soluble N -ethyl maleimide-sensitive fusion protein attachment protein receptor (SNARE) proteins cellubrevin, synaptobrevin II, and SNAP-23 have been found in astrocytes (Zhang et al., 2004 ; Bezzi et al., 2004 ; Montana et al., 2004) . Release of glutamate from cultured astrocytes was reported to be triggered by cytosolic Ca 2+ elevations (Parpura et al., 1994 ; Araque et al., 1998 ; Bezzi et al., 2004) and was reduced when astrocytic Ca 2+ rises were suppressed by the Ca 2+ chelator BAPTA (Araque et al., 1998 ; Zhang et al., 2004) . In addition, inhibition of the endogenous SNARE complex by genetic introduction of an exogenous SNARE motif into astrocytes prevented the release of glutamate and ATP (Zhang et al., 2004 ; Pascual et al., 2005) , emphasizing the involvement of the SNARE complex for the release of gliotransmitters. Recently it was reported that glutamate release from hippocampal astrocytes, initiated by P2Y1 receptor-mediated Ca 2+ signals, strengthens synaptic transmission and hence modu-lates synaptic tuning in neuronal circuits (Jourdain et al., 2007) . Taken together, these findings suggest that transmitters such as glutamate and ATP can be released from astrocytes in a Ca 2+ -dependent manner by vesicular exocytosis, and possibly also by Ca 2+ -independent mechanisms. While vesicular release of gliotransmitters may be the predominant mechanism employed by astrocytes as a response to cytosolic Ca 2+ increases, other mechanisms may mediate release during certain pathophysiological processes, such as cell swelling, which activates anion channels and therefore allows efflux of glutamate or ATP (Kimelberg et al., 2006) , and P2X7 receptor activation by high extracellular ATP levels found during CNS injury (Duan and Neary, 2006) . A more detailed review about vesicular transmitter release from astrocytes is provided by Montana et al. (2006) .

216 J.W. Deitmer et al.
8.9 Functional Significance of Ca 2+ Signaling
There are numerous processes in the nervous system that may be initiated and/or modulated by cytosolic Ca 2+ rises in astrocytes, as has been reported for many other cell types. On the other hand, astrocytic Ca 2+ signaling itself is subject to modula-tion by neuronal activity, in particular at synapses, and therefore is one of the prime mechanisms by which reciprocal neuron–glia signaling is established (Hirase et al., 2004 ; Perea and Araque, 2005a, b ). One of the most consequential processes initiated by Ca 2+ rises in astrocytes is the Ca 2+ -dependent release of transmitters. Some of these gliotransmitters and glia-derived messengers, such as glutamate, ATP, tumor necrosis factor-a, or d -serine, have been shown to modulate neuronal excitability, synaptic transmission, and cerebrovascular microcirculation. Therefore, these glio-transmitters may serve as feedback signals that modulate neuronal activity (see Chap. 15). Astrocyte-induced modulation of synaptic transmission has been observed in cultured cells as well as in tissue slices of several brain areas. Since astrocytes respond to neurotransmitters with Ca 2+ elevations that can extend to relatively large distances and that induce release of gliotransmitters that modulate neurotransmis-sion, astrocyte Ca 2+ signaling may represent a form of brain information pathway that establishes a functional link between distant synaptic areas. Some of these Ca 2+ -dependent processes in astrocytes could also be elicited by photolysis of caged Ca 2+ compounds by UV illumination (Parpura and Haydon, 2000 ; Fellin et al., 2004 ; Kreft et al., 2004 ; Fiacco and McCarthy, 2004) .
Neuronal excitability has also been shown to be modulated by glutamate released from astrocytes both in vitro and in situ. Ca 2+ -dependent glutamate release from astrocytes may have strong impact in brain pathophysiology, because it can lead to the synchronized activity of clusters of neurons, and may be responsible for the generation of epileptiform activity in neurons. However, the role of astrocytic Ca 2+ signaling for the development, maintenance, or recovery from pathophysiolog-ical conditions is still largely unknown.
Other messenger molecules, which have been reported to be released from astrocytes in a Ca 2+ -dependent fashion, are nitric oxide (NO), e.g., mediated by the activation of metabotropic purinergic P2Y receptors. This short-lived volatile mes-senger, which acts both in an ortho- and retrograde way, may be a link to other second messenger cascades in neighbouring neurons and glial cells, such as the cGMP-mediated pathway. Moreover, NO may enhance store-operated Ca 2+ entry (SOCE) and thereby feeds back on the cellular competence to generate Ca 2+ signals due to repeated Ca 2+ release from intracellular stores (Li et al., 2003) . In cultured astrocytes NO caused Ca 2+ -dependent glutamate release from inflammatory acti-vated cells (Bal-Price et al., 2002) . Thus, Ca 2+ signals in astrocytes may be involved both in physiological and pathological processes in the brain.
Ca 2+ signaling in astrocytes has also been associated with the control of cerebral blood flow. Ca 2+ transients and Ca 2+ oscillations are propagated to the astrocytic end-feet, which ensheath part of the endothelial layer of blood capillaries, and can elicit vasodilation and vasoconstriction by the release of lipid metabolites (Zonta et al., 2003 ; Parri and Crunelli, 2003 ; Mulligan and MacVicar, 2004 ; Takano et al.,

8 Calcium Ion Signaling in Astrocytes 217
2006 ; Metea and Newman, 2006) . Inhibition of cyclooxygenase-1 activity blocked astrocyte-triggered vasodilation, supporting the notion that Ca 2+ -dependent release of prostaglandins from astrocytes mediates the control local blood flow. This modu-lation may be initiated by neuronal activity in restricted brain areas, by evoking astrocyte Ca 2+ signals that are propagated toward blood vessels and in turn mediate the control of microcirculation. Astrocytic Ca 2+ signals have been reported to cause either vasoconstriction and vasodilation, and this may be decided by the lipid products involved. As the supply of oxygen and glucose in the brain is affected, these mechanisms may well be an important glial link to the maintenance of brain energy metabolism.
Furthermore, cytosolic Ca 2+ rises in astrocytes may gate or modulate ion channels in the cell membrane, in particular K + channels, which may be involved in K + clearance from the extracellular space following neuronal activity (see Chap. 6). Whether the Ca 2+ signals recorded at low extracellular K + concentration in astro-cytes (Dallwig et al., 2000 ; Härtel et al., 2007) are of physiological and pathological relevance requires further analysis.
The list of cellular functions mediated by Ca 2+ signals in astrocytes will certainly be extended in the years to come. It is the regional control of cellular functions, which may be regulated or modulated by Ca 2+ signals in a unique way. By invading different cellular processes of astrocytes, by being regenerative, and by being a versatile mode of signaling, this form of glial excitation has many assets for allowing a complex dialogue between glial cells and neurons, and may contribute critically to information processing and metabolism in the brain.
8.10 Summary and Conclusion
Cytosolic Ca 2+ signals in astrocytes can be spontaneous or evoked, and are often initiated by activation of metabotropic receptors, resulting in Ca 2+ release from intracellular Ca 2+ stores. While voltage-dependent Ca 2+ influx is rare in astrocytes, the refilling of the intracellular Ca 2+ stores requires store-operated Ca 2+ entry, a major influx pathway of Ca 2+ into astrocytes. Astrocyte Ca 2+ signals are generated in form of transients or oscillations, which can be evoked by neurotransmitters, hormones, chemokines, lipids, and growth factors, often in response to neuronal activity. These Ca 2+ signals are propagated along cellular processes and can travel from the point of origin across the tissue as Ca 2+ waves. When propagated into astrocytic end-feet, they may release cyclooxygenase products to control cerebral blood flow, or modulate neuronal excitability and glia–neuron interactions. The release of transmitters and other molecules elicited by Ca 2+ signaling in astrocytes may be key mechanisms to allow a complex dialogue between neurons and glial cells. Although we have learned a lot about the phenomenology of Ca 2+ signals in astrocytes, and about the potential function it may have, we still lack knowledge about the specificity of astrocytic Ca 2+ signals with respect to the pathophysiological conditions under which they are generated, and the cell function they may elicit in

218 J.W. Deitmer et al.
a given situation. It seems clear that these Ca 2+ signals are instrumental for many processes in astrocytes and for their interaction with other glial cells and neurons.
Acknowledgments Our studies on this topic have been supported by grants from the Deutsche Forschungsgemeinschaft (SFB 530, GRK 845), which we gratefully acknowledge.
References
Allen NJ , Attwell D (2004) The effect of simulated ischaemia on spontaneous GABA release in area CA1 of the juvenile rat hippocampus . J Physiol 561 : 485 – 498 .
Araque A , Sanzgiri RP , Parpura V , Haydon PG (1998) Calcium elevation in astrocytes causes an NMDA receptor-dependent increase in the frequency of miniature synaptic currents in cultured hippocampal neurons . J Neurosci 18 : 6822 – 6829 .
Araque A , Carmignoto G , Haydon PG (2001) Dynamic signaling between astrocytes and neurons . Annu Rev Physiol 63 : 795 – 813 .
Arcuino G , Lin JH , Takano T , Liu C , Jiang L , Gao Q , Kang J , Nedergaard M (2002) Intercellular calcium signaling mediated by point-source burst release of ATP . Proc Natl Acad Sci USA 99 : 9840 – 9845 .
Backus KH , Berger T (1995) Developmental variation of the permeability to Ca 2+ of AMPA recep-tors in presumed hilar glial precursor cells . Pflügers Arch 431 : 244 – 252 .
Bajetto A , Bonavia R , Barbero S , Piccioli P , Costa A , Florio T , Schettini G (1999) Glial and neu-ronal cells express functional chemokine receptor CXCR4 and its natural ligand stromal cell-derived factor 1 . J Neurochem 73 : 2348 – 2357 .
Bal-Price A , Moneer Z , Brown GC (2002) Nitric oxide induces rapid, calcium-dependent release of vesicular glutamate and ATP from cultured rat astrocytes . Glia 40 : 312 – 323 .
Beck A , Zur Nieden R , Schneider HP , Deitmer JW (2004) Calcium release from intracellular stores in rodent astrocytes and neurons in situ . Cell Calcium 35 : 47 – 58 .
Beierlein M , Regehr WG (2006) Brief bursts of parallel fiber activity trigger calcium signals in Bergmann glia . J Neurosci 26 : 6958 – 6967 .
Berridge MJ (2006) Calcium microdomains: organization and function . Cell Calcium 40 : 405 – 412 . Bezzi P , Domercq M , Brambilla L , Galli R , Schols D , De Clercq E , Vescovi A , Bagetta G , Kollias
G , Meldolesi J , Volterra A (2001) CXCR4-activated astrocyte glutamate release via TNFalpha: amplification by microglia triggers neurotoxicity . Nat Neurosci 4 : 702 – 710 .
Bezzi P , Gundersen V , Galbete JL , Seifert G , Steinhauser C , Pilati E , Volterra A (2004) Astrocytes contain a vesicular compartment that is competent for regulated exocytosis of glutamate . Nat Neurosci 7 : 613 – 620 .
Bigini P , Gardoni F , Barbera S , Cagnotto A , Fumagalli E , Longhi A , Corsi MM , Di Luca M , Mennini T (2006) Expression of AMPA and NMDA receptor subunits in the cervical spinal cord of wobbler mice . BMC Neurosci 7 : 71 .
Burnashev N , Khodorova A , Jonas P , Helm PJ , Wisden W , Monyer H , Seeburg PH , Sakmann B (1992) Calcium-permeable AMPA-kainate receptors in fusiform cerebellar glial cells . Science 256 : 1566 – 1570 .
Carmignoto G , Pasti L , Pozzan T (1998) On the role of voltage-dependent calcium channels in calcium signaling of astrocytes in situ . J Neurosci 18 : 4637 – 4645 .
Carmignoto G (2000) Reciprocal communication systems between astrocytes and neurones . Prog Neurobiol 62 : 561 – 581 .
Coco S , Calegari F , Pravettoni E , Pozzi D , Taverna E , Rosa P , Matteoli M , Verderio C (2003) Storage and release of ATP from astrocytes in culture . J Biol Chem 278 : 1354 – 1362 .
Cornell-Bell AH , Finkbeiner SM , Cooper MS , Smith SJ (1990) Glutamate induces calcium waves in cultured astrocytes: long-range glial signaling . Science 247 : 470 – 473 .
Cotrina ML , Kang J , Lin JH , Bueno E , Hansen TW , He L , Liu Y , Nedergaard M (1998) Astrocytic gap junctions remain open during ischemic conditions . J Neurosi 18 : 2520 – 2537 .

8 Calcium Ion Signaling in Astrocytes 219
Czubayko U , Reiser G (1995) [Ca 2+ ] i oscillations in single rat glioma cells induced by thrombin through activation of cell surface receptors . Neuroreport 6 : 1249 – 1252 .
Dallwig R , Vitten H , Deitmer JW (2000) A novel barium-sensitive calcium influx into rat astro-cytes at low external potassium . Cell Calcium 28 : 247 – 259 .
Dave V , Gordon GW , McCarthy KD (1991) Cerebral type 2 astroglia are heterogeneous with respect to their ability to respond to neuroligands linked to calcium mobilization . Glia 4 : 440 – 447 .
Deitmer JW , Verkhratsky A , Lohr C (1998) Calcium signalling in glial cells . Cell Calcium 24 : 405 – 416 .
Delumeau JC , Petitet F , Cordier J , Glowinski J , Premont J (1991) Synergistic regulation of cytosolic Ca 2+ concentration in mouse astrocytes by NK1 tachykinin and adenosine agonists . J Neurochem 57 : 2026 – 2035 .
Duan S , Neary JT (2006) P2X(7) receptors: properties and relevance to CNS function . Glia 54 : 738 – 746 .
Duffy S , MacVicar BA (1994) Potassium-dependent calcium influx in acutely isolated hippocam-pal astrocytes . Neuroscience 61 : 51 – 61 .
Duffy S , MacVicar BA (1995) Adrenergic calcium signaling in astrocyte networks within the hip-pocampal slice . J Neurosci 15 : 5535 – 5550 .
Enkvist MO , Holopainen I , Akerman KE (1989) Glutamate receptor-linked changes in membrane potential and intracellular Ca 2+ in primary rat astrocytes . Glia 2 : 397 – 402 .
Fellin T , Pascual O , Gobbo S , Pozzan T , Haydon PG , Carmignoto G (2004) Neuronal synchrony mediated by astrocytic glutamate through activation of extrasynaptic NMDA receptors . Neuron 43 : 729 – 743 .
Feske S , Giltnane J , Dolmetsch R , Staudt LM , Rao A (2001) Gene regulation mediated by calcium signals in T lymphocytes . Nat Immunol 2 : 316 – 324 .
Fiacco TA , McCarthy KD (2004) Intracellular astrocyte calcium waves in situ increase the frequency of spontaneous AMPA receptor currents in CA1 pyramidal neurons . J Neurosci 24 : 722 – 732 .
Fiacco TA , McCarthy KD (2006) Astrocyte calcium elevations: properties, propagation, and effects on brain signaling . Glia 54 : 676 – 690 .
Fields RD , Stevens-Graham B (2002) New insights into neuron-glia communication . Science 298 : 556 – 562 .
Finkbeiner S (1992) Calcium waves in astrocytes-filling in the gaps . Neuron 8 : 1101 – 1108 . Gimpl G , Walz W , Ohlemeyer C , Kettenmann H (1992) Bradykinin receptors in cultured
astrocytes from neonatal rat brain are linked to physiological responses . Neurosci Lett 144 : 139 – 142 .
Gimpl G , Kirchhoff F , Lang RE , Kettenmann H (1993) dentification of neuropeptide Y receptors in cultured astrocytes from neonatal rat brain . J Neurosci Res 34 : 198 – 205 .
Grosche J , Matyash V , Moller T , Verkhratsky A , Reichenbach A , Kettenmann H (1999) Microdomains for neuron–glia interaction: parallel fiber signaling to Bergmann glial cells . Nat Neurosci 2 : 139 – 143 .
Guthrie PB , Knappenberger J , Segal M , Bennett MV , Charles AC , Kater SB (1999) ATP released from astrocytes mediates glial calcium waves . J Neurosci 19 : 520 – 528 .
Härtel K , Singaravelu K , Kaiser M , Neusch C , Hülsmann S , Deitmer JW (2007) Calcium influx mediated by the inwardly rectifying K + channel Kir4.1 (KCNJ10) at low external K + concen-tration . Cell Calcium 42 : 271 – 280 .
Hassinger TD , Guthrie PB , Atkinson PB , Bennett MV , Kater SB (1996) An extracellular signaling component in propagation of astrocytic calcium waves . Proc Natl Acad Sci USA 93 : 13268 – 13273 .
Hirase H , Qian L , Bartho P , Buzsaki G (2004) Calcium dynamics of cortical astrocytic networks in vivo . PLoS Biol 2 : E96 .
Hirase H , Creso J , Singleton M , Bartho P , Buzsaki G (2004) Two-photon imaging of brain peri-cytes in vivo using dextran-conjugated dyes . Glia 46 : 95 – 100 .
Ho C , Hicks J , Salter MW (1995) A novel P 2 -purinoceptor expressed by a subpopulation of astro-cytes from the dorsal spinal cord of the rat . Br J Pharmacol 116 : 2909 – 2918 .
Hoth M , Penner R (1992) Depletion of intracellular calcium stores activates a calcium current in mast cells . Nature 355 : 353 – 356 .

220 J.W. Deitmer et al.
Inagaki N , Fukui H , Ito S , Yamatodani A , Wada H (1991) Single type-2 astrocytes show multiple independent sites of Ca 2+ signaling in response to histamine . Proc Natl Acad Sci USA 88 : 4215 – 4219 .
James G , Butt AM (2001) P 2 X and P 2 Y purinoreceptors mediate ATP-evoked calcium signalling in optic nerve glia in situ . Cell Calcium 30 : 251 – 259 .
Jensen AM , Chiu SY (1991) Differential intracellular calcium responses to glutamate in type 1 and type 2 cultured brain astrocytes . J Neurosci 11 : 1674 – 1684 .
Jourdain P , Bergersen LH , Bhaukaurally K , Bezzi P , Santello M , Domercq M , Matute C , Tonello F , Gundersen V , Volterra A (2007) Glutamate exocytosis from astrocytes controls synaptic strength . Nat Neurosci 10 : 331 – 339 .
Jung S , Pfeiffer F , Deitmer JW (2000) Histamine-induced calcium entry in rat cerebellar astro-cytes: evidence for capacitative and non-capacitative mechanisms . J Physiol 61 : 549 – 561 .
Jurzak M , Muller AR , Gerstberger R (1995) Characterization of vasopressin receptors in cultured cells derived from the region of rat brain circumventricular organs . Neuroscience 65 : 1145 – 1159 .
Kanemaru K , Okubo Y , Hirose K , Iino M (2007) Regulation of neurite growth by spontaneous Ca 2+ oscillations in astrocytes . J Neurosci 27 : 8957 – 8966 .
Kastritsis CH , Salm AK , McCarthy K (1992) Stimulation of the P 2 Y purinergic receptor on type 1 astroglia results in inositol phosphate formation and calcium mobilization . J Neurochem 58 : 1277 – 1284 .
Kimelberg HK , Macvicar BA , Sontheimer H (2006) Anion channels in astrocytes: biophysics, pharmacology, and function . Glia 54 : 747 – 757 .
Kreft M , Stenovec M , Rupnik M , Grilc S , Krzan M , Potokar M , Pangrsic T , Haydon PG , Zorec R (2004) Properties of Ca 2+ -dependent exocytosis in cultured astrocytes . Glia 46 : 437 – 445 .
Kulik A , Haentzsch A , Luckermann M , Reichelt W , Ballanyi K (1999) Neuron–glia signaling via 1 adrenoceptor-mediated Ca 2+ release in Bergmann glial cells in situ . J Neurosci 19 : 8401 – 8408 .
Lalo U , Pankratov Y , Kirchhoff F , North RA , Verkhratsky A (2006) NMDA receptors mediate neuron-to-glia signaling in mouse cortical astrocytes . J Neurosci 26 : 2673 – 2683 .
Li N , Sul JY , Haydon PG (2003) A calcium-induced calcium influx factor, nitric oxide, modulates the refilling of calcium stores in astrocytes . J Neurosci 23 : 10302 – 10310 .
Liou J , Kim ML , Heo WD , Jones JT , Myers JW , Ferrell JE , Meyer T (2005) STIM is a Ca 2+ sensor essential for Ca 2+ store depletion triggered Ca 2+ influx . Curr Biol 15 : 1235 – 1241 .
Liu QY , Lai FA , Rousseau E , Jones RV , Meissner G (1989) Multiple conductance states of the purified calcium release channel complex from skeletal sarcoplasmic reticulum . Biophys J 55 : 415 – 424 .
Lo KJ , Luk HN , Chin TY , Chueh SH (2002) Store depletion-induced calcium influx in rat cerebel-lar astrocytes . Br J Pharmacol 135 : 1383 – 1392 .
Lohr C , Deitmer JW (1999) Dendritic calcium transients in the leech giant glial cell . Glia 26 : 109 – 118 .
Lohr C , Heil JE , Deitmer JW (2005) Blockage of voltage-gated calcium signalling impairs migra-tion of glial cells in vivo . Glia 50 : 198 – 211 .
Lohr C , Deitmer JW (2006) Calcium signaling in invertebrate glial cells . Glia 54 : 642 – 649 . Mattson MP , Chan SL (2003) Neuronal and glial calcium signaling in Alzheimer’s disease . Cell
Calcium 34 : 385 – 397 . Matyash V , Filippov V , Mohrhagen K , Kettenmann H (2001) Nitric oxide signals parallel fiber
activity to Bergmann glial cells in the mouse cerebellar slice . Mol Cell Neurosci 18 : 664 – 670 . Meissner G , Henderson JS (1987) Rapid calcium release from cardiac sarcoplasmic reticulum
vesicles is dependent on Ca 2+ and is modulated by Mg 2+ , adenine nucleotide, and calmodulin . J Biol Chem 262 : 3065 – 3073 .
Metea MR , Newman EA (2006) Glial cells dilate and constrict blood vessels: a mechanism of neurovascular coupling . J Neurosci 26 : 2862 – 2870 .
Metea MR , Newman EA (2006) Calcium signaling in specialized glial cells . Glia 54 : 650 – 655 . Montana V , Malarkey EB , Verderio C , Matteoli M , Parpura V (2006) Vesicular transmitter release
from astrocytes . Glia 54 : 700 – 715 .

8 Calcium Ion Signaling in Astrocytes 221
Montana V , Ni Y , Sunjara V , Hua X , Parpura V (2004) Vesicular glutamate transporter dependent glutamate release from astrocytes . J Neurosci 24 : 2633 – 2642 .
Morita M , Higuchi C , Moto T , Kozuka N , Susuki J , Itofusa R , Yamashita J , Kudo Y (2003) Dual regulation of calcium oscillation in astrocytes by growth factors and pro-inflammatory cytokines via the mitogen-activated protein kinase cascade . J Neurosci 23 : 10944 – 10952 .
Mothet JP , Pollegioni L , Ouanounou G , Martineau M , Fossier P , Baux G (2005) Glutamate receptor activation triggers a calcium-dependent and SNARE protein-dependent release of the glio-transmitter d -serine . Proc Natl Acad Sci USA 102 : 5606 – 5611 .
Müller T , Moller T , Berger T , Schnitzer J , Kettenmann H (1992) Calcium entry through kainate recep-tors and resulting potassium-channel blockade in Bergmann glial cells . Science 256 : 1563 – 1566 .
Mulligan SJ , MacVicar BA (2004) Calcium transients in astrocyte endfeet cause cerebrovascular constrictions . Nature 431 : 195 – 199 .
Nedergaard M (1994) Direct signaling from astrocytes to neurons in cultures of mammalian brain cells . Science 263 : 1768 – 1771 .
Nett WJ , Oloff SH , McCarthy KD (2002) Hippocampal astrocytes in situ exhibit calcium oscilla-tions that occur independent of neuronal activity . J Neurophysiol 87 : 528 – 537 .
Newman EA , Zahs KR (1997) Calcium waves in retinal glial cells . Science 275 : 844 – 847 . Newman EA (2001) Propagation of intercellular calcium waves in retinal astrocytes and Muller
cells . J Neurosci 21 : 2215 – 2223 . Newman EA (2003) Glial cell inhibition of neurons by release of ATP . J Neurosci 23 : 1659 – 1666 . Newman EA (2005) Calcium increases in retinal glial cells evoked by light-induced neuronal
activity . J Neurosci 25 : 5502 – 5510 . Nilsson M , Hansson E , Ronnback L (1991) Adrenergic and 5-HT2 receptors on the same astro-
glial cell. A microspectrofluorimetric study on cytosolic Ca 2+ responses in single cells in pri-mary culture . Brain Res Dev Brain Res 63 : 33 – 41 .
Nilsson M , Eriksson PS , Ronnback L , Hansson E (1993) GABA induces Ca 2+ transients in astro-cytes . Neuroscience 54 : 605 – 614 .
Oheim M , Kirchhoff F , Stühmer W (2006) Calcium microdomains in regulated exocytosis . Cell Calcium 40 : 423 – 439 .
Parpura V , Haydon PG (2000) Physiological astrocytic calcium levels stimulate glutamate release to modulate adjacent neurons . Proc Natl Acad Sci USA 97 : 8629 – 8634 .
Parekh AB , Putney JW Jr (2005) Store-operated calcium channels . Physiol Rev 85 : 757 – 810 . Parekh AB (2000) Calcium signaling and acute pancreatitis: specific response to a promiscuous
messenger . Proc Natl Acad Sci USA 97 : 12933 – 12934 . Parpura V , Basarsky TA , Liu F , Jeftinija K , Jeftinija S , Haydon PG (1994) Glutamate-mediated
astrocyte-neuron signalling . Nature 369 : 744 – 747 . Parri HR , Crunelli V (2001) Pacemaker calcium oscillations in thalamic astrocytes in situ .
Neuroreport 12 : 3897 – 3900 . Parri HR , Crunelli V (2003) The role of Ca 2+ in the generation of spontaneous astrocytic Ca 2+
oscillations . Neuroscience 120 : 979 – 992 . Partiseti M , Le Deist F , Hivroz C , Fischer A , Korn H , Choquet D (1994) The calcium current
activated by T cell receptor and store depletion in human lymphocytes is absent in a primary immunodeficiency . J Biol Chem 269 : 32327 – 32335 .
Pascual O , Casper KB , Kubera C , Zhang J , Revilla-Sanchez R , Sul JY , Takano H , Moss SJ , McCarthy K , Haydon PG (2005) Astrocytic purinergic signaling coordinates synaptic net-works . Science 310 : 113 – 116 .
Pasti L , Pozzan T , Carmignoto G (1995) Long-lasting changes of calcium oscillations in astro-cytes. A new form of glutamate-mediated plasticity . J Biol Chem 270 : 15203 – 15210 .
Pasti L , Zonta M , Pozzan T , Vicini S , Carmignoto G (2001) Cytosolic calcium oscillations in astrocytes may regulate exocytotic release of glutamate . J Neurosci 21 : 477 – 484 .
Peakman MC , Hill SJ (1995) Adenosine A1 receptor-mediated changes in basal and histamine-stimulated levels of intracellular calcium in primary rat astrocytes . Br J Pharmacol 115 : 801 – 810 .

222 J.W. Deitmer et al.
Peinelt C , Vig M , Koomoa DL , Beck A , Nadler MJ , Koblan-Huberson M , Lis A , Fleig A , Penner R , Kinet JP (2006) Amplification of CRAC current by STIM1 and CRACM1 (Orai1) . Nat Cell Biol 8 : 771 – 773 .
Perea G , Araque A (2005a) Properties of synaptically evoked astrocyte calcium signal reveal synaptic information processing by astrocytes . J Neurosci 25:2192–2203. Erratum in J Neurosci 25 : 3022 .
Perea G , Araque A (2005b) Glial calcium signaling and neuron–glia communication . Cell Calcium 38 : 375 – 382 .
Peters O , Schipke CG , Hashimoto Y , Kettenmann H (2003) Different mechanisms promote astrocyte Ca 2+ waves and spreading depression in the mouse neocortex . J Neurosci 23 : 9888 – 9896 .
Porter JT , McCarthy KD (1995a) Adenosine receptors modulate [Ca 2+ ] i in hippocampal astrocytes in situ . J Neurochem 65 : 1515 – 1523 .
Porter JT , McCarthy KD (1995b) GFAP-positive hippocampal astrocytes in situ respond to gluta-matergic neuroligands with increases in [Ca 2+ ] i . Glia 13 : 101 – 112 .
Prakriya M , Feske S , Gwack Y , Srikanth S , Rao A , Hogan PG (2006) Orai1 is an essential pore subunit of the CRAC channel . Nature 443 : 230 – 233 .
Putney JW Jr (2000) Presenilins, Alzheimer’s disease, and capacitative calcium entry . Neuron 27 : 411 – 412 .
Roos J , DiGregorio PJ , Yeromin AV , Ohlsen K , Lioudyno M , Zhang S , Safrina O , Kozak JA , Wagner SL , Cahalan MD , Velicelebi G , Stauderman KA (2005) STIM1, an essential and con-served component of store-operated Ca 2+ channel function . J Cell Biol 169 : 435 – 445 .
Rossi DJ , Oshima T , Attwell D (2000) Glutamate release in severe brain ischaemia is mainly by reversed uptake . Nature 403 : 316 – 321 .
Salm AK , McCarthy KD (1990) Norepinephrine-evoked calcium transients in cultured cerebral type 1 astroglia . Glia 3 : 529 – 538 .
Scemes E , Giaume C (2006) Astrocyte calcium waves: what they are and what they do . Glia 54 : 716 – 725 .
Scemes E , Dermietzel R , Spray DC (1998) Calcium waves between astrocytes from Cx43 knock-out mice . Glia 24 : 65 – 73 .
Schipke CG , Ohlemeyer C , Matyash M , Nolte C , Kettenmann H , Kirchhoff F (2001) Astrocytes of the mouse neocortex express functional N -methyl- d -aspartate receptors . FASEB J 15 : 1270 – 1272 .
Schipke CG , Boucsein C , Ohlemeyer C , Kirchhoff F , Kettenmann H (2002) Astrocyte Ca 2+ waves trigger responses in microglial cells in brain slices . FASEB J 16 : 255 – 257 .
Seifert G , Weber M , Schramm J , Steinhauser C (2003) Changes in splice variant expression and subunit assembly of AMPA receptors during maturation of hippocampal astrocytes . Mol Cell Neurosci 22 : 248 – 258 .
Sergeeva M , Strokin M , Wang H , Ubl JJ , Reiser G (2003) Arachidonic acid in astrocytes blocks Ca 2+ oscillations by inhibiting store-operated Ca 2+ entry, and causes delayed Ca 2+ influx . Cell Calcium 33 : 283 – 292
Serrano A , Haddjeri N , Lacaille JC , Robitaille R (2006) GABAergic network activation of glial cells underlies hippocampal heterosynaptic depression . J Neurosci 26 : 5370 – 5382 .
Shao Y , McCarthy KD (1995) Receptor-mediated calcium signals in astroglia: multiple receptors, common stores and all-or-nothing responses . Cell Calcium 17 : 187 – 196 .
Sharma G , Vijayaraghavan S (2001) Nicotinic cholinergic signaling in hippocampal astrocytes involves calcium-induced calcium release from intracellular stores . Proc Natl Acad Sci USA 98 : 4148 – 4153 .
Shelton MK , McCarthy KD (2000) Hippocampal astrocytes exhibit Ca 2+ -elevating muscarinic cholinergic and histaminergic receptors in situ . J Neurochem 74 : 555 – 563 .
Shih AY , Erb H , Sun X , Toda S , Kalivas PW , Murphy TH (2006) Cystine/glutamate exchange modulates glutathione supply for neuroprotection from oxidative stress and cell proliferation . J Neurosci 26 (41) : 10514 – 10523 .
Shuttleworth TJ , Thompson JL (1999) Discriminating between capacitative and arachidonate-activated Ca 2+ entry pathways in HEK293 cells . J Biol Chem 274 : 31174 – 31178 .

8 Calcium Ion Signaling in Astrocytes 223
Simpson PB , Russell JT (1997) Role of sarcoplasmic/endoplasmic-reticulum Ca 2+ -ATPases in mediating Ca 2+ waves and local Ca 2+ -release microdomains in cultured glia . Biochem J 325 : 239 – 247 .
Singaravelu K , Lohr C , Deitmer JW (2006) Regulation of store-operated calcium entry by calci-um-independent phospholipase A 2 in rat cerebellar astrocytes . J Neurosci 26 : 9579 – 9592 .
Sitsapesan R , McGarry SJ , Williams AJ (1995) Cyclic ADP-ribose, the ryanodine receptor and Ca 2+ release . Trends Pharmacol Sci 16 : 386 – 391 .
Smani T , Zakharov SI , Leno E , Csutora P , Trepakova ES , Bolotina VM (2003) Ca 2+ -independent phospholipase A 2 is a novel determinant of store-operated Ca 2+ entry . J Biol Chem 278 : 11909 – 11915 .
Smani T , Zakharov SI , Leno E , Csutora P , Trepakova ES , Bolotina VM (2004) A novel mechanism for the store-operated calcium influx pathway . Nat Cell Biol 6 : 113 – 120 .
Soboloff J , Spassova MA , Tang XD , Hewavitharana T , Xu W , Gill DL (2006) Orail and STIM reconstitute store-operated calcium channel function . J Biol Chem 281 : 20661 – 20665 .
Spassova MA , Soboloff J , He LP , Xu W , Dziadek MA , Gill DL (2005) STIM1 has a plasma mem-brane role in the activation of store-operated Ca 2+ channels . Proc Natl Acad Sci USA 103 : 4040 – 4045 .
Spät A (2006) Calcium microdomains and the fine control of cell function: an introduction . Cell Calcium 40 : 403 – 404 .
Stiene-Martin A , Mattson MP , Hauser KF (1993) Opiates selectively increase intracellular cal-cium in developing type-1 astrocytes: role of calcium in morphine-induced morphologic dif-ferentiation . Brain Res Dev Brain Res 76 : 189 – 196 .
Suadicani SO , Brosnan CF , Scemes E (2006) P2X7 receptors mediate ATP release and amplifica-tion of astrocytic intercellular Ca 2+ signaling . J Neurosci 26 : 1378 – 1385 .
Szatkowski M , Barbour B , Attwell D (1990) Non-vesicular release of glutamate from glial cells by reversed electrogenic glutamate uptake . Nature 348 : 443 – 446 .
Takano T , Tian GF , Peng W , Lou N , Libionka W , Han X , Nedergaard M (2006) Astrocyte-mediated control of cerebral blood flow . Nat Neurosci 9 : 260 – 267 .
Verkhratsky A , Shmigol A (1996) Calcium-induced calcium release in neurones . Cell Calcium 19 : 1 – 14 . Verkhratsky A , Orkland RK , Kettenmann H (1998) Glial calcium: homeostasis and signaling
function . Physiol Rev 78 : 99 – 141 . Vig M , Peinelt C , Beck A , Koomoa DL , Rabah D , Koblan-Huberson M , Kraft S , Turner H , Fleig
A , Penner R , Kinet JP (2006) CRACM1 is a plasma membrane protein essential for store-operated Ca 2+ entry . Science 312 : 1220 – 1223 .
Walz W , Gimpl G , Ohlemeyer C , Kettenmann H (1994) Extracellular ATP-induced currents in astrocytes: involvement of a cation channel . J Neurosci Res 38 : 12 – 18 .
Wang D , Martens JR , Posner P , Sumners C , Gelband CH (1996) Angiotensin II regulation of intracellular calcium in astroglia cultured from rat hypothalamus and brainstem . J Neurochem 67 : 996 – 1004 .
Wang X , Lou N , Xu Q , Tian GF , Peng WG , Han X , Kang J , Takano T , Nedergaard M (2006) A strocytic Ca 2+ signaling evoked by sensory stimulation in vivo . Nat Neurosci 9 : 816 – 823 .
Xu J , Peng H , Kang N , Zhao Z , Lin JHC , Stanton PK , Kang J (2007) Glutamate-induced exocy-tosis of glutamate from astrocytes . J Biol Chem 282 : 24185 – 24197 .
Yang KT , Chen WP , Chang WL , Su MJ , Tsai KL (2005) Arachidonic acid inhibits capacitative Ca 2+ entry and activates non-capacitative Ca 2+ entry in cultured astrocytes . Biochem Biophys Res Commun 331 : 603 – 613 .
Ye ZC , Rothstein JD , Sontheimer H (1999) Compromised glutamate transport in human glioma cells: reduction-mislocalization of sodium-dependent glutamate transporters and enhanced activity of cystine-glutamate exchange . J Neurosci 19 : 10767 – 10777 .
Ye ZC , Wyeth MS , Baltan-Tekkok S , Ransom BR (2003) Functional hemichannels in astrocytes: a novel mechanism of glutamate release . J Neurosci 23 : 3588 – 3596 .
Yeromin AV , Zhang SL , Jiang W , Yu Y , Safrina O , Cahalan MD (2006) Molecular identification of the CRAC channel by altered ion selectivity in a mutant of Orai . Nature 443 : 226 – 229 .

224 J.W. Deitmer et al.
Zhang Q , Pangrsic T , Kreft M , Krzan M , Li N , Sul JY , Halassa M , Van Bockstaele E , Zorec R , Haydon PG (2004) Fusion-related release of glutamate from astrocytes . J Biol Chem 279 : 12724 – 12733 .
Zhang SL , Yu Y , Roos J , Kozak JA , Deerinck TJ , Ellisman MH , Stauderman KA , Cahalan MD (2005) STIM1 is a Ca 2+ sensor that activates CRAC channels and migrates from the Ca 2+ store to the plasma membrane . Nature 437 : 902 – 905 .
Zonta M , Angulo MC , Gobbo S , Rosengarten B , Hossmann KA , Pozzan T , Carmignoto G (2003) Neuron-to-astrocyte signaling is central to the dynamic control of brain microcirculation . Nat Neurosci 6 : 43 – 50 .
Zur Nieden R , Deitmer JW (2006) The role of metabotropic glutamate receptors for the generation of calcium oscillations in rat hippocampal astrocytes in situ . Cereb Cortex 16 : 676 – 687 .
Abbreviations
2-APB 2-Aminoethoxydiphenyl borate ADP Adenosine diphosphate AMPA Alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid ATP Adenosine 5′-triphosphate BAPTA 1,2-bis(o-aminophenoxy)ethane- N , N , N ′, N ′-tetraacetic acid CIF Ca 2+ influx factor CRAC Ca 2+ release-activated channel DAG Diacylglycerol DPCPX 8-Cyclopentyl-1,3-dipropylxanthine EAAT Excitatory amino acid transporter ER Endoplasmic reticulum GABA Gamma-aminobutyric acid IP
3 Inositol-1,4,5-trisphosphate
iPLA 2 Ca 2+ -independent phopholipase A
2
NMDA N -methyl- D -aspartate P2X Ionotropic purinoceptor P2Y Metabotropic purinoceptor PLC Phospholipase C SERCA Sarcoplasmic-endoplasmic reticulum Ca 2+ -ATPase SNARE Soluble N -ethyl maleimide-sensitive fusion protein attachment protein
receptor SOCE Store-operated Ca 2+ entry STIM1 Stromal interaction molecule 1 VGLUT Vesicular glutamate transporter