characterization of stretch-activated ion channels in cultured astrocytes
TRANSCRIPT

GLIA 8:87-96 (1993)
Characterization of Stretch-Activated Ion Channels in Cultured Astrocytes
LEON ISLAS,' HERMINU PASANTES-MORALES,' AND JORGE A. SANCHEZ2 'Institute of Cell Physiology, National A. Uniuersity of Mexico, and "enter of Research and Advanced Studies,
IPN, Mexico City, Mexico
KEY WORDS Volume regulation, Membrane stretch, Potassium channels, Hypos- molarity, Swelling
ABSTRACT The presence of a stretch-activated channel in rat cerebellar astrocytes in culture is described. This stretch-sensitive channel is K+-selective and its open proba- bility increases with suction following a Boltzmann-like distribution with half activation at 45 mm Hg. Kinetic analysis of the single-channel data indicated that there are two open and two closed states and that the shortest time constants of both open and closed states are the most sensitive to suction. A symmetrical two-barrier-one-site permeation model can quantitatively describe the inward rectification of the single-channel current to voltage relations. It is suggested that this stretch-activated channel plays a role in the regulatory volume response of astrocytes to hyposmotic conditions. o 1993 Wiley-Liss, Inc.
INTRODUCTION
A large variety of animal cells are capable of cell volume regulation when challenged by anisosmotic me- dia (Hoffman, 1987; Macknight, 1988; Cala, 1990). This volume regulatory process has also been observed in cultured astrocytes under hyposmotic conditions (Kimel- berg and Frangakis, 1986; Olson et al., 1986; Pasantes- Morales and Schousboe, 1988). When incubated in me- dia of reduced osmolarity, astrocytes respond initially by marked swelling followed by a slower phase of vol- ume recovery that occurs despite the persistence of the anisosmotic conditions. The volume regulatory process is achieved by the activation of transmembrane fluxes of osmotically active intracellular solutes which leave the cell with osmotically obliged water. Potassium, chlo- ride as well as organic molecules such as free amino acids, and polyols have been implicated in cell volume regulation in most cells (Hoffman, 1987; Macknight, 1988; Law and Burg, 1991). The mechanisms involved in the corrective fluxes of osmolytes are apparently dif- ferent in the different cell types. A diffusional process seems responsible for the swelling-activated efflux of amino acids (Sanchez Olea et al., 1991; Schousboe et al., 1992), whereas in the case of inorganic ions, the mechanisms suggested involve either electroneutral cotransporters (Kregenow, 1981; Lauf, 1985; Cala, 1990) 0 1993 Wiley-Liss, Inc
or separate but interdependent conductive pathways for K" and C1- (Grinstein et al., 1984; Hoffman, 1987; Yantorno et al., 1989; Macleod and Hamilton, 1991). The mechanism whereby changes in the cell volume trigger the membrane permeability increase is unknown at present. It may involve effects of mechanical stress in the membrane which result in the opening of stretch- activated channels of the type recently described in a variety of cells (Sachs, 1991). Activated conductances through these channels may serve as signals to elicit the volume corrective pathways.
The term stretch-activated (SA) channels refers to those channels that being silent become active after the direct application of pressure through a patch clamp pipette (Guharay and Sachs, 1984) or by reduction in external osmolarity leading to membrane distension (Sakin, 1989). Cell swelling has been found to induce opening of stretch-activated channels in several types of cells including epithelial cells (Ubl et al., 19881, hepa- tocytes, myocytes, and neuroblastoma cells (Falke and Misler, 1989). A preliminary study has suggested the occurrence of these stretch-activated channels in astro-
Received July 3,1992; accepted October 2,1992. Address reprint requests to Dr. Herminia Pasantes-Morales, Instituto de Fisio-
logia Celular, UNAM, Apartado Postal 70-600,04510, Mexico, D.F. M6xico.

88 ISUS ET AL
cytes (Ding et al., 1988). The aim of the present work was to confirm the occurrence of these channels in cul- tured astrocytes and to characterize some of their prop- erties.
MATERIALS AND METHODS
Primary cultures of astrocytes were obtained from 7-day-old rats according to the procedure of Pate1 and Hunt (1985). Cerebella were mechanically dissociated in Eagle's medium (Gibco, Grand Island, NY) supple- mented with 10% of heat-inactivated fetal calf serum and maintained in an incubator at 37°C in an atmos- phere of 5% C02/95% air. Cells were plated in glass cover-slips coated with poly-L-lysine. Immunohistochem- ical studies in our laboratory show that these cultures are 90-95% pure astrocytes. Cultures of 1-2 weeks old were used in the experiments reported here; in this stage the cells are confluent. The cells were treated with dibutyryl-CAMP (Sigma Chemical Co., St. Louis, MO) 1 day before use; this treatment produces spheri- cal cells, thus enhancing sealing on the astrocyte mem- brane. The treatment produces no changes in the prop- erties of the stretch-activated channels nor in their density, although cells treated expressed a greater num- ber of K' channels.
Single channel recordings were made with fire pol- ished borosilicate glass pipettes as described by Hamill et al. (1981). Recordings were obtained in the inside-out patch configuration using an EPC-7 patch clamp ampli- fier (List Electronics). The composition of pipette and bath solutions are given in each figure. All solutions were buffered with HEPES 10 mM at pH 7.4. The bath temperature was 20-22°C. To enhance single channel activity in stretch-activated channels, pressure was ap- plied through the patch-pipette and was measured si- multaneously with a mercury manometer. In experi- ments involving continuous recording, single channel currents were stored on an analog tape recorder (Hewlett- Packard 3964A) with a bandwidth of 5 kHz and digi- tized off line at 10 kHz for analysis. Currents during voltage step depolarizations, applied from the holding potential (Eh), were also recorded. Pulses were gener- ated by a microcomputer (IBM compatible) through a Tecmar Labmaster board (Scientific Solutions, Inc.) that also digitized the current data with a resolution of 12 bits. Linear membrane currents were subtracted from recordings that contained no obvious single channel ac- tivity.
Data were analyzed by the microcomputer using pCLAMP (Axon Instruments). Openings and closings longer than 500 p s were used for event duration analy- sis. The amplitudes, open durations, and closed dura- tions of single channel events were measured individu- ally and visually checked for accuracy. Scatter plots of event amplitudes versus duration were displayed for each analysis. Open and closed time histograms were constructed from records that contained only one chan- nel and were fitted by exponential functions. Single-
channel activity was scarce in the absence of pressure, making difficult the estimation of the number of chan- nels present at zero pressure. However, the presence of more than one channel in a patch became obvious when pressure was applied. In addition, the statistical test described by Colquhoun and Hawkes (1981) was used to confirm the presence of only one channel in these determinations. In our experiments, about 70% of the patches contained only one channel.
Current to voltage relations were fitted to the sim- plest model from Eyring rate theory that involves the presence of one ion-binding site and two energy barri- ers (Hille, 1992). In this model, a saturable binding site within the channel plays a key role in ion permeation. This site must be free before it can be occupied by an- other ion. Binding and unbinding depend on rate con- stants f,, and k,,, respectively. These rate constants are given by Eyring rate theory and depend on a voltage term and a voltage-independent constant as described by Lewis and Stevens (1979):
where v, and v2 are vibration frequencies whose values are 1013/sec and lO''/molar-sec, respectively. Unitary currents carried by the ith ion (Ii) were calculated from the model according to the equations:
hi + k ~ , Po =
k,, t kli + foi + fli
17)
where Po and P, are the probabilities that the site is empty or occupied, respectively. Zi and F have their usual thermodynamic meanings. V is voltage, 6 is the electrical distance, and [XI, and [XIi, are the concentra- tions of ion x in the intracellular and extracellular solu- tions, respectively.
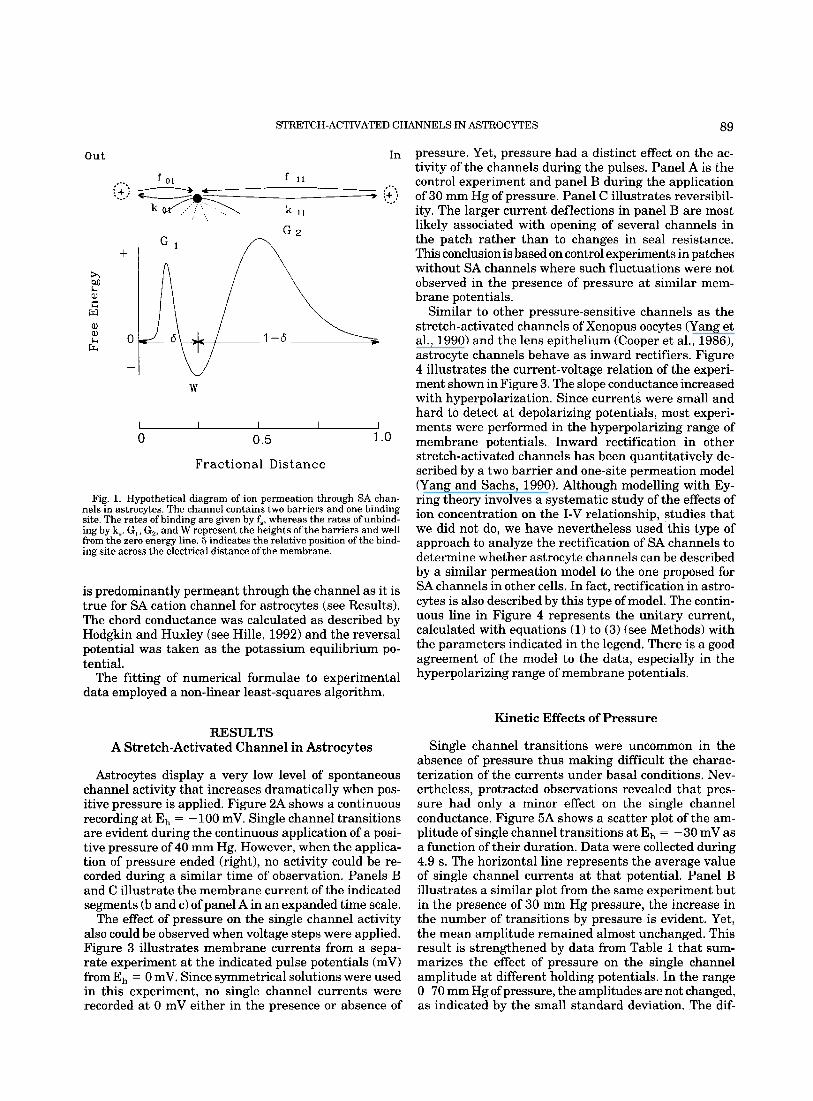
Figure 1 is a representation of the model, in which two energy barriers of heights G, and G2 and an energy well W are shown. The abscissa represents the electri- cal distance across the membrane. 6 represents the rel- ative location of the binding site within the channel.
Modelling of ion permeation can be greatly simplified if it is assumed that only one species of monovalent ions
STRETCH-ACTIVATED CHANNELS IN ASTROCYTES 89
out In
\
W
0 0.5 1 .o
Fractional Distance
Fig. 1. Hypothetical diagram of ion permeation through SA chan- nels in astrocytes. The channel contains two barriers and one binding site. The rates of binding are given by fs, whereas the rates of unbind- ing by k,. G,, G,, and W represent the heights of the barriers and well from the zero energy line. 8 indicates the relative position of the bind- ing site across the electrical distance of the membrane.
is predominantly permeant through the channel as it is true for SA cation channel for astrocytes (see Results). The chord conductance was calculated as described by Hodgkin and Huxley (see Hille, 1992) and the reversal potential was taken as the potassium equilibrium po- tential.
The fitting of numerical formulae to experimental data employed a non-linear least-squares algorithm.
RESULTS A Stretch-Activated Channel in Astrocytes
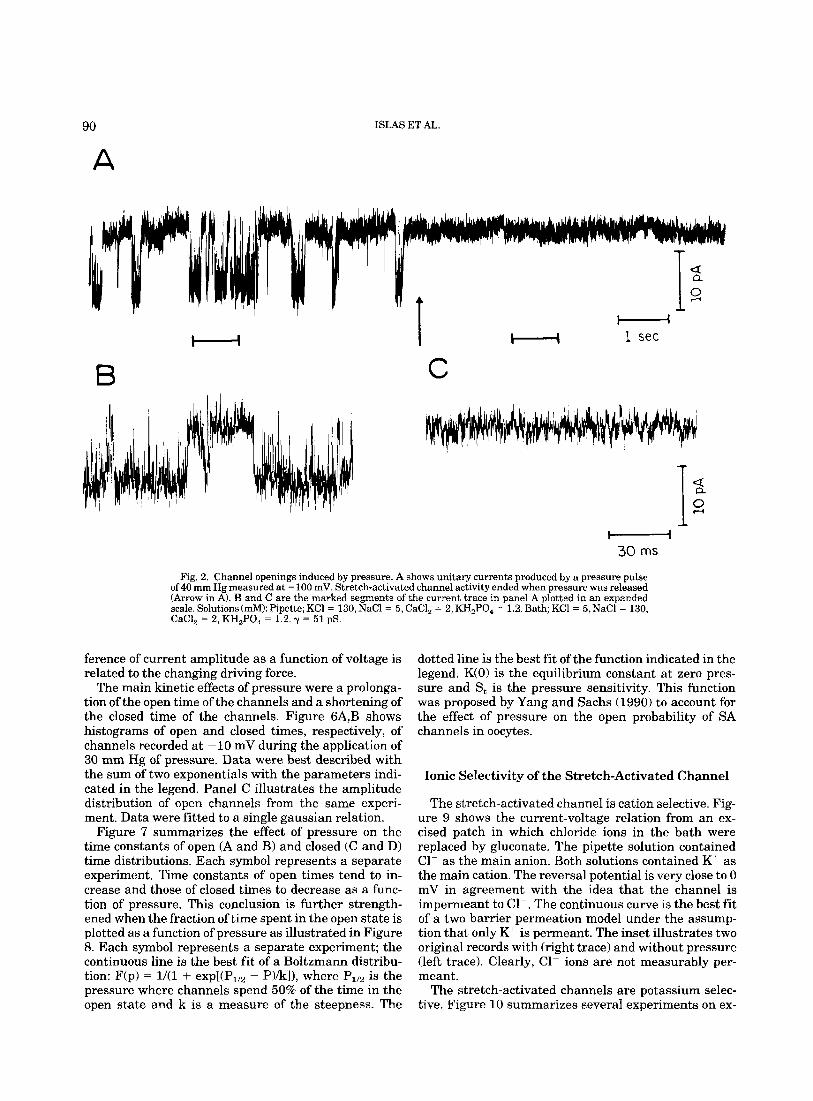
Astrocytes display a very low level of spontaneous channel activity that increases dramatically when pos- itive pressure is applied. Figure 2A shows a continuous recording at Eh = -100 mV. Single channel transitions are evident during the continuous application of a posi- tive pressure of 40 mm Hg. However, when the applica- tion of pressure ended (right), no activity could be re- corded during a similar time of observation. Panels B and C illustrate the membrane current of the indicated segments (b and c) of panel A in an expanded time scale.
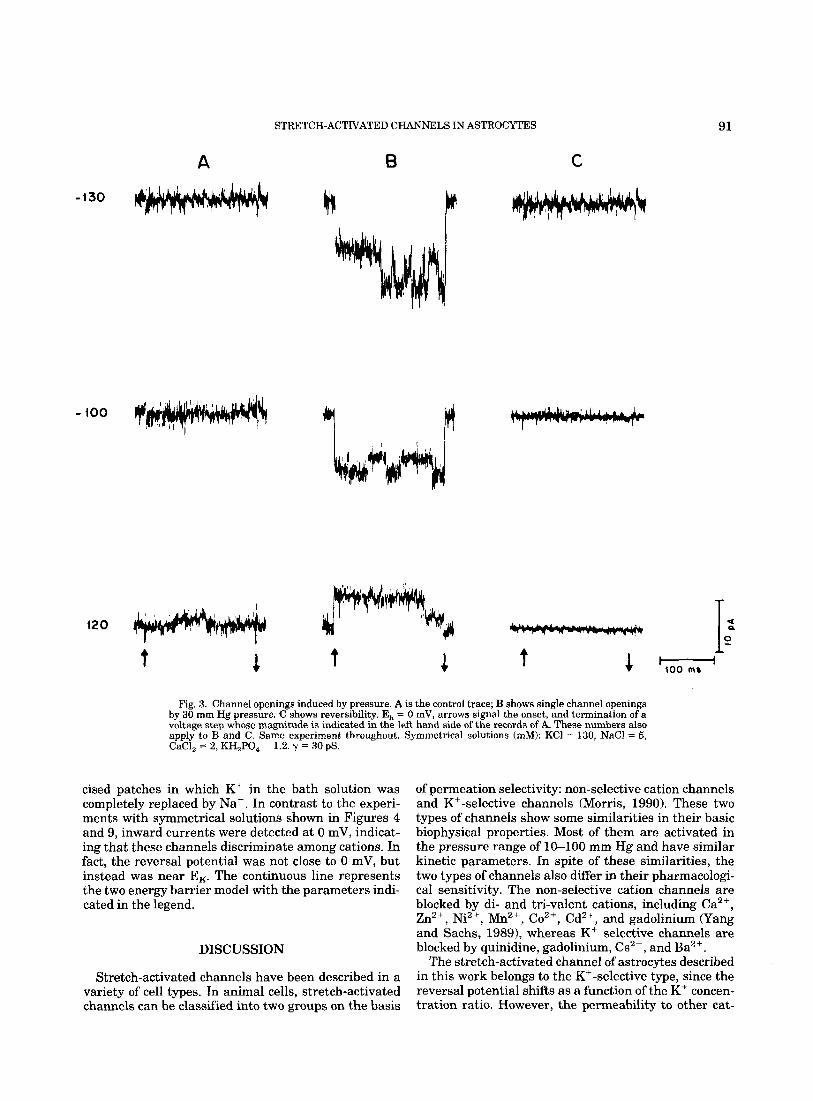
The effect of pressure on the single channel activity also could be observed when voltage steps were applied. Figure 3 illustrates membrane currents from a sepa- rate experiment a t the indicated pulse potentials (mV) from E, = 0 mV. Since symmetrical solutions were used in this experiment, no single channel currents were recorded at 0 mV either in the presence or absence of
pressure. Yet, pressure had a distinct effect on the ac- tivity of the channels during the pulses. Panel A is the control experiment and panel B during the application of 30 mm Hg of pressure. Panel C illustrates reversibil- ity. The larger current deflections in panel B are most likely associated with opening of several channels in the patch rather than to changes in seal resistance. This conclusion is based on control experiments in patches without SA channels where such fluctuations were not observed in the presence of pressure at similar mem- brane potentials.
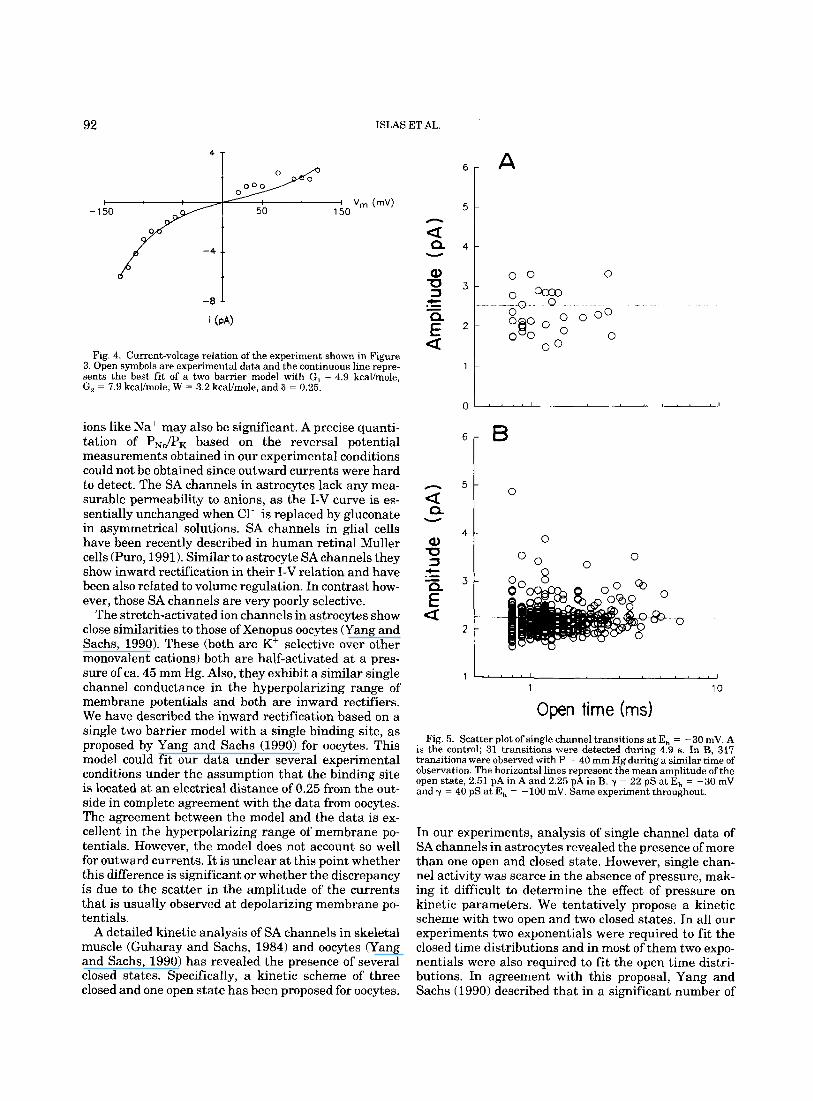
Similar to other pressure-sensitive channels as the stretch-activated channels of Xenopus oocytes (Yang et al., 1990) and the lens epithelium (Cooper et al., 19861, astrocyte channels behave as inward rectifiers. Figure 4 illustrates the current-voltage relation of the experi- ment shown in Figure 3. The slope conductance increased with hyperpolarization. Since currents were small and hard to detect at depolarizing potentials, most experi- ments were performed in the hyperpolarizing range of membrane potentials. Inward rectification in other stretch-activated channels has been quantitatively de- scribed by a two barrier and one-site permeation model (Yang and Sachs, 1990). Although modelling with Ey- ring theory involves a systematic study of the effects of ion concentration on the I-V relationship, studies that we did not do, we have nevertheless used this type of approach to analyze the rectification of SA channels to determine whether astrocyte channels can be described by a similar permeation model to the one proposed for SA channels in other cells. In fact, rectification in astro- cytes is also described by this type of model. The contin- uous line in Figure 4 represents the unitary current, calculated with equations (1) to (3) (see Methods) with the parameters indicated in the legend. There is a good agreement of the model to the data, especially in the hyperpolarizing range of membrane potentials.
Kinetic Effects of Pressure
Single channel transitions were uncommon in the absence of pressure thus making difficult the charac- terization of the currents under basal conditions. Nev- ertheless, protracted observations revealed that pres- sure had only a minor effect on the single channel conductance. Figure 5A shows a scatter plot of the am- plitude of single channel transitions at Eh = - 30 mV as a function of their duration. Data were collected during 4.9 s. The horizontal line represents the average value of single channel currents at that potential. Panel B illustrates a similar plot from the same experiment but in the presence of 30 mm Hg pressure, the increase in the number of transitions by pressure is evident. Yet, the mean amplitude remained almost unchanged. This result is strengthened by data from Table 1 that sum- marizes the effect of pressure on the single channel amplitude at different holding potentials. In the range &70 mm Hg of pressure, the amplitudes are not changed, as indicated by the small standard deviation. The dif-
90
A ISMS ET AL.
H I B
H
C
I
1 sec
Fig. 2. Channel openings induced by pressure. A shows unitary currents produced by a pressure pulse of 40 rnrn Hg measured at - 100 mV. Stretch-activated channel activity ended when pressure was released (Arrow in A). B and C are the marked segments of the current trace in panel A plotted in an expanded scale. Solutions (d): Pipette; KCl = 130, NaCl = 5, CaCl, = 2, KH,PO, = 1.2. Bath; KCl = 5, NaCl = 130, CaCl, = 2, KH,PO, = 1.2. y = 51 pS.
ference of current amplitude as a function of voltage is related to the changing driving force.
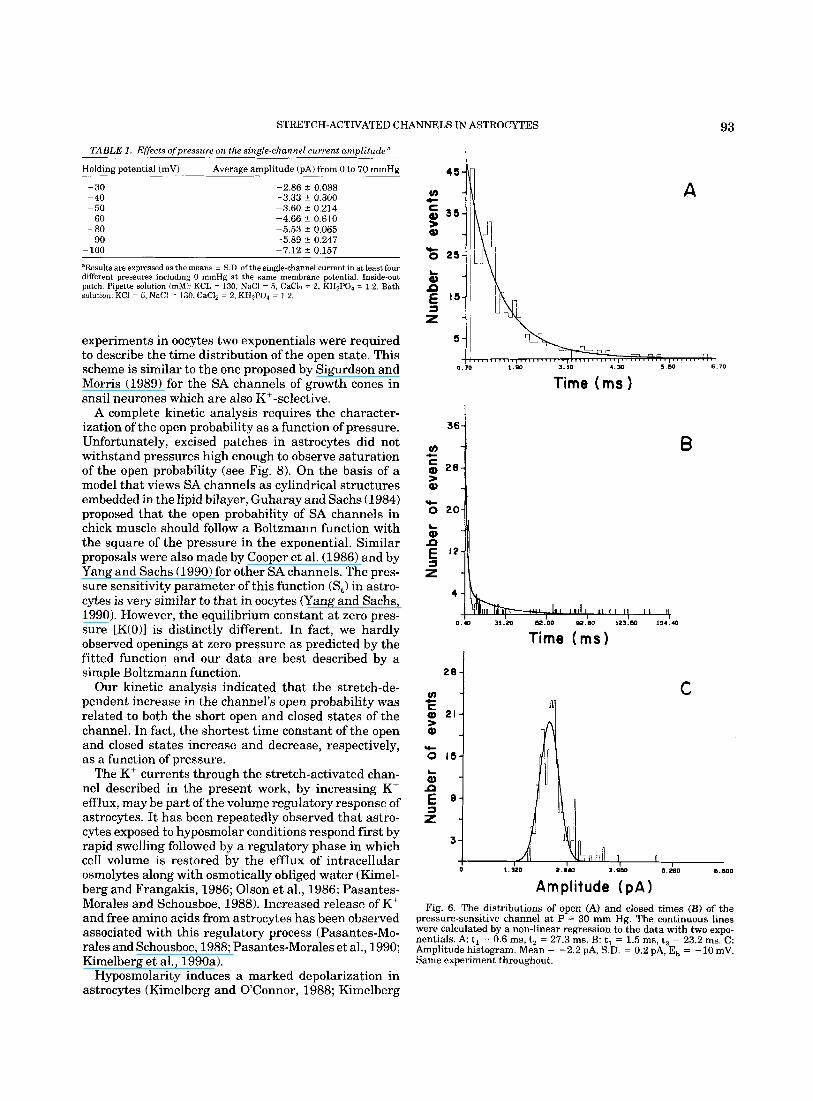
The main kinetic effects of pressure were a prolonga- tion of the open time of the channels and a shortening of the closed time of the channels. Figure 6A,B shows histograms of open and closed times, respectively, of channels recorded at ~ 10 mV during the application of 30 mm Hg of pressure. Data were best described with the sum of two exponentials with the parameters indi- cated in the legend. Panel C illustrates the amplitude distribution of open channels from the same experi- ment. Data were fitted to a single gaussian relation.
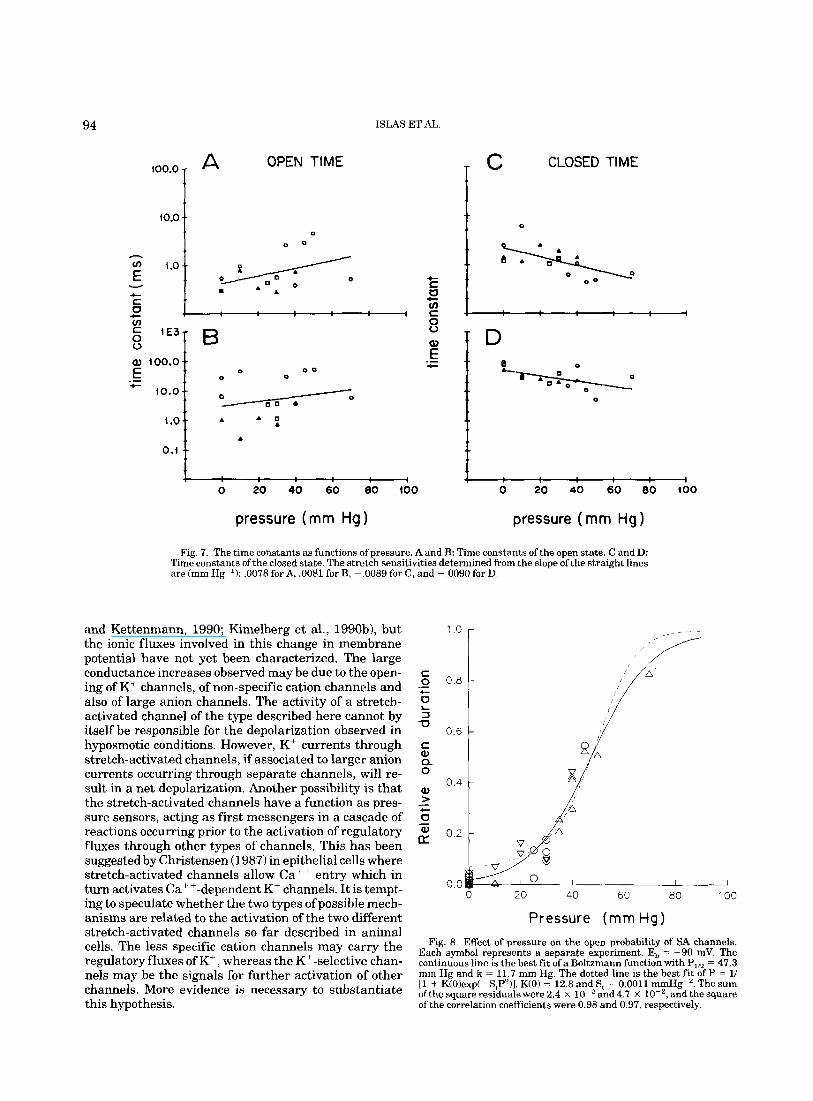
Figure 7 summarizes the effect of pressure on the time constants of open (A and B) and closed (C and D) time distributions. Each symbol represents a separate experiment, Time constants of open times tend to in- crease and those of closed times to decrease as a func- tion of pressure. This conclusion is further strength- ened when the fraction of time spent in the open state is plotted as a function of pressure as illustrated in Figure 8. Each symbol represents a separate experiment; the continuous line is the best fit of a Boltzmann distribu- tion: F(p) = 141 + exp[(P,,2 - P)/kl), where PlIz is the pressure where channels spend 50% of the time in the open state and k is a measure of the steepness. The
dotted line is the best fit of the function indicated in the legend. K(0) is the equilibrium constant at zero pres- sure and s, is the pressure sensitivity. This function was proposed by Yang and Sachs (1990) to account for the effect of pressure on the open probability of SA channels in oocytes.
Ionic Selectivity of the Stretch-Activated Channel
The stretch-activated channel is cation selective. Fig- ure 9 shows the current-voltage relation from an ex- cised patch in which chloride ions in the bath were replaced by gluconate. The pipette solution contained C1- as the main anion. Both solutions contained K+ as the main cation. The reversal potential is very close to 0 mV in agreement with the idea that the channel is impermeant to Cl-. The continuous curve is the best fit of a two barrier permeation model under the assump- tion that only KC is permeant. The inset illustrates two original records with (right trace) and without pressure (left trace). Clearly, C1- ions are not measurably per- meant.
The stretch-activated channels are potassium selec- tive. Figure 10 summarizes several experiments on ex-
STRETCH-ACTIVATED CHANNELS IN ASTROCYTES 91
A
-130
-100 ly'
B C
Fig. 3. Channel openings induced by pressure. A is the control trace; B shows single channel openings by 30 rnm Hg pressure. C shows reversibility. E, = 0 mV, arrows signal the onset, and termination of a voltage step whose magnitude is indicated in the left hand side of the records of A. These numbers also apply to B and C. Same experiment throughout. Symmetrical solutions (mM): KCl = 130, NaCl = 5, CaCl, = 2, KH,PO, = 1.2. y = 30 pS.
cised patches in which K+ in the bath solution was completely replaced by Na+. In contrast to the experi- ments with symmetrical solutions shown in Figures 4 and 9, inward currents were detected at 0 mV, indicat- ing that these channels discriminate among cations. In fact, the reversal potential was not close to 0 mV, but instead was near EK. The continuous line represents the two energy barrier model with the parameters indi- cated in the legend.
DISCUSSION
Stretch-activated channels have been described in a variety of cell types. In animal cells, stretch-activated channels can be classified into two groups on the basis
of permeation selectivity: non-selective cation channels and Kt-selective channels (Morris, 1990). These two types of channels show some similarities in their basic biophysical properties. Most of them are activated in the pressure range of 10-100 mm Hg and have similar kinetic parameters. In spite of these similarities, the two types of channels also differ in their pharmacologi- cal sensitivity. The non-selective cation channels are blocked by di- and tri-valent cations, including Ca2+, Zn" , Ni2+, Mn2+, Co2+, Cd", and gadolinium (Yang and Sachs, 19891, whereas K' selective channels are blocked by quinidine, gadolinium, Cs2+, and Ba2+.
The stretch-activated channel of astrocytes described in this work belongs to the K+-selective type, since the reversal potential shifts as a function of the K+ concen- tration ratio. However, the permeability to other cat-
92
4 T
ISMS ET AL.
6 r A
Y T k! -'t
Fig. 4. Current-voltage relation of the experiment shown in Figure 3. Open symbols are experimental data and the continuous line repre- sents the best fit of a two barrier model with G, = 4.9 kcal/mole, G, = 7.9 kcal/mole, W = 3.2 kcal/mole, and 6 L 0.25.
ions like Na+ may also be significant. A precise quanti- tation of P,,/P, based on the reversal potential measurements obtained in our experimental conditions could not be obtained since outward currents were hard to detect. The SA channels in astrocytes lack any mea- surable permeability to anions, as the I-V curve is es- sentially unchanged when C1- is replaced by gluconate in asymmetrical solutions. SA channels in glial cells have been recently described in human retinal Muller cells (Puro, 1991). Similar to astrocyte SA channels they show inward rectification in their I-V relation and have been also related to volume regulation. In contrast how- ever, those SA channels are very poorly selective.
The stretch-activated ion channels in astrocytes show close similarities to those of Xenopus oocytes (Yang and Sachs, 1990). These (both are K+ selective over other monovalent cations) both are half-activated at a pres- sure of ca. 45 mm Hg. Also, they exhibit a similar single channel conductance in the hyperpolarizing range of membrane potentials and both are inward rectifiers. We have described the inward rectification based on a single two barrier model with a single binding site, as proposed by Yang and Sachs (1990) for oocytes. This model could fit our data under several experimental conditions under the assumption that the binding site is located at an electrical distance of 0.25 from the out- side in complete agreement with the data from oocytes. The agreement between the model and the data is ex- cellent in the hyperpolarizing range of membrane po- tentials. However, the model does not account so well for outward currents. It is unclear at this point whether this difference is significant or whether the discrepancy is due to the scatter in the amplitude of the currents that is usually observed at depolarizing membrane po- tentials.
A detailed kinetic analysis of SA channels in skeletal muscle (Guharay and Sachs, 1984) and oocytes (Yang and Sachs, 1990) has revealed the presence of several closed states. Specifically, a kinetic scheme of three closed and one open state has been proposed for oocytes.
5 I c 1 4 U z l
0 0
0 I
1 ' ' " ' 1 I
1 10
Open time (ms) Fig. 5. Scatter plot of single channel transitions at E, = -30 mV. A
is the control; 31 transitions were detected during 4.9 s. In B, 317 transitions were observed with P = 40 mm Hg during a similar time of observation. The horizontal lines represent the mean amplitude of the open state, 2.51 pA in A and 2.25 pA in B. y = 22 pS at E, = -30 mV and y = 40 pS a t E, = -100 mV. Same experiment throughout.
In our experiments, analysis of single channel data of SA channels in astrocytes revealed the presence of more than one open and closed state. However, single chan- nel activity was scarce in the absence of pressure, mak- ing it difficult to determine the effect of pressure on kinetic parameters. We tentatively propose a kinetic scheme with two open and two closed states. In all our experiments two exponentials were required to fit the closed time distributions and in most of them two expo- nentials were also required to fit the open time distri- butions. In agreement with this proposal, Yang and Sachs (1990) described that in a significant number of
STRETCH-ACTIVATED CHANNELS IN ASTROCYTES
a - c w 21- w a -
0 15-
a -
t
rc
L
n E 9-
t - 3-
3
93
nn n fl 1 I
1.320 1.a40 3.am 5.2110 8.600
TABLE 1, Effects of pressure on the sin&-channel current arnulitude
Holding potential (mV) -
-30 - 40 -50
60 ~ 80 - 90 - 100
Average amplitude (PA) from 0 to 70 mmHg
-2 86 2 0 088 -3 33 i 0 300 -3 60 f 0 214 -4 66 t 0 610 -5 53 t 0 065 -5 89 _+ 0 247
- 7 12 t 0 157
'Results are expressed as the means f S.D. ofthe singlc-channel current in at lenst four different pressures including 0 mmHg at the same membrane potential. Inside-out pnlcti. Pipette solution (mM): KCL - 130, NaCl = R , CaCla = 2, KHaPOl = 1.2. Bath solution: KCI ~ 5, NaCl 2 130, CaCl, = 2, KH2PU4 = 1 .?.
experiments in oocytes two exponentials were required to describe the time distribution of the open state. This scheme is similar to the one proposed by Sigurdson and Morris (1989) for the SA channels of growth cones in snail neurones which are also K'-selective.
A complete kinetic analysis requires the character- ization of the open probability as a function of pressure. Unfortunately, excised patches in astrocytes did not withstand pressures high enough to observe saturation of the open probability (see Fig. 8). On the basis of a model that views SA channels as cylindrical structures embedded in the lipid bilayer, Guharay and Sachs (1984) proposed that the open probability of SA channels in chick muscle should follow a Boltzmann function with the square of the pressure in the exponential. Similar proposals were also made by Cooper et al. (1986) and by Yang and Sachs (1990) for other SA channels. The pres- sure sensitivity parameter of this function (St) in astro- cytes is very similar to that in oocytes (Yang and Sachs, 1990). However, the equilibrium constant at zero pres- sure [K(O)l is distinctly different. In fact, we hardly observed openings at zero pressure as predicted by the fitted function and our data are best described by a simple Boltzmann function.
Our kinetic analysis indicated that the stretch-de- pendent increase in the channel's open probability was related to both the short open and closed states of the channel. In fact, the shortest time constant of the open and closed states increase and decrease, respectively, as a function of pressure.
The K+ currents through the stretch-activated chan- nel described in the present work, by increasing Kt efflux, may be part of the volume regulatory response of astrocytes. It has been repeatedly observed that astro- cytes exposed to hyposmolar conditions respond first by rapid swelling followed by a regulatory phase in which cell volume is restored by the efflux of intracellular osmolytes along with osmotically obliged water (Kimel- berg and Frangakis, 1986; Olson et al., 1986; Pasantes- Morales and Schousboe, 1988). Increased release of K+ and free amino acids from astrocytes has been observed associated with this regulatory process (Pasantes-Mo- rales and Schousboe, 1988; Pasantes-Morales et al., 1990; Kimelberg et al., 1990a).
Hyposmolarity induces a marked depolarization in astrocytes (Kimelberg and OConnor, 1988; Kimelberg
Time (ms ) I 1
A
e 28
w aa
B
- 1 0 20
E '2 3 z !I
C
Amplitude ( P A ) Fig. 6. The distributions of open (A) and closed times (B) of the
pressure-sensitive channel at P 30 mm Hg. The continuous lines were calculated by a non-linear regression to the data with two expo- nentials. A t, = 0.6 ms, t, = 27.3 ms. B: t, = 1.5 ms, t, = 23.2 ms. C: Amplitude histogram. Mean = -2.2 PA, S.D. = 0.2 PA, E, = -10 mV. Same experiment throughout.
94 ISMS ET AL.
A OPEN TIME
0 I 0 0
100.0
10.0
4- f
0 '"*Of 0 0 6
O B 1 t t....,.
0 20 40 60 80 100
T C CLOSED TIME
I D 0 f "-.: 0 . 0
i 0 20 40 60 80 100
pressure (mm Hg) pressure (rnm Hg)
Fig. 7. The time constants as functions of pressure. A and B: Time constants of the open state. C and D: Time constants of the closed state. The stretch sensitivities determined from the slope of the straight lines are (mm Hg I): ,0078 for A, .0081 for B, -.0089 for C, and -.0090 for D.
and Kettenmann, 1990; Kimelberg et al., 1990b), but the ionic fluxes involved in this change in membrane potential have not yet been characterized. The large conductance increases observed may be due to the open- ing of K' channels, of non-specific cation channels and also of large anion channels. The activity of a stretch- activated channel of the type described here cannot by itself be responsible for the depolarization observed in hyposmotic conditions. However, K' currents through stretch-activated channels, if associated to larger anion currents occurring through separate channels, will re- sult in a net depolarization. Another possibility i s that the stretch-activated channels have a function as pres- sure sensors, acting as first messengers in a cascade of reactions occurring prior to the activation of regulatory fluxes through other types of channels. This has been suggested by Christensen (1987) in epithelial cells where stretch-activated channels allow Cat+ entry which in turn activates Ca +-dependent K+ channels. It is tempt- ing to speculate whether the two types of possible mech- anisms are related to the activation of the two different stretch-activated channels so far described in animal cells. The less specific cation channels may carry the regulatory fluxes of K', whereas the Kt -selective chan- nels may be the signals for further activation of other channels. More evidence is necessary to substantiate this hypothesis.
c o 0.8 _- c 0 1 I 3 U
c Q, Q 0
0.6
0.4
0.2
0.0
/
0 20 40 60 80 100
Pressure (mm Hg) Fig. 8. Effect of pressure on the open probability of SA channels.
Each symbol represents a separate experiment. E, = -90 mV. The continuous line is the best fit of a Boltzmann function with PI,.. = 47.3 mm Hg and k = 11.7 mm Hg. The dotted line is the best fit of P = 1/ 11 + K(0)exp(-S,P2)J. K(0) = 12.8 and S, = 0.0011 mmHg-'. The sum ofthe square residuals were 2.4 x and4.7 X lo-', and the square of the correlation coefficients were 0.98 and 0.97, respectively.

STRETCH-ACTIVATED CHANNELS IN ASTROCYTES 95
-8 1
Fig. 9. Effect of anion substitution on stretch-activated currents. Pipette solution contained (mM): KCl = 130, NaCl = 5, CaCl, = 2 Bath solution: K-gluconate = 130, NaCl 5, and CaCl, = 2. Open sym- bols are experimental data and the continuous line represents the best fit of a two barrier model with G, = 4.9 kcallmole, G, 6.4 kcallmole, W = 1.6 kcallmole, and S = 0.25. The insert shows membrane currents at -120 mV before (left) and during application of 30 mm Hg pressure (right).
5 1 I I V
8p I -10 1
Fig. 10. Effect of cation substitution on stretch-activated currents. Pipette solution contained (mM): KC1 = 130, NaCl = 5, CaCl, = 2, KH,PO, = 1.2, HEPES = 10. Bath solution: KC1 = 5, NaCl = 130, and CaCI, = 2, KH,PO, = 1.2, HEPES = 10. Open symbols are experi- mental data and the continuous line represents the best fit of a two barrier model with G, = 7.8 kcallmole, G, = 7.7 kcallmole, W = 4.9 kcavmole, and S 0.25. Each symbol represents a separate experi- ment. The arrow indicates the K' equilibrium potential.
ACKNOWLEDGMENTS
The excellent technical assistance of Ms. Claudia Ro- driguez is highly appreciated. The work has been sup- ported by grants from CONACyT (0287N) and DGAPA- UNAM (IN-204589).
REFERENCES
Cala, P.M. (1990) Principles of cell volume regulation ion flux path- ways and the roles of anions. In: Chloride Channels and Carriers in Nerue, Muscle, and Glial Cells. Plenum Press, New York, pp. 67-83.
Christensen, 0. (1987) Mediation of cell volume regulation by Ca'+ infux through stretch-activated channels. Nature, 330:6&68.
Colquhoun, D. and Nawkes, A.G. (1981). The principles of the stochas- tic interpretation of ion-channel mechanisms. In: Single Channel Recording. B. Sakmann and E. Neher, eds. Plenum Press, New York, pp. 135-175.
Cooper, K.E., Tang, J.M., Rae, J.L., and Eisenberg, R.S. (1986) A cation channel in frog lens epithelium responsive to pressure and calcium. J. Membrane Biol., 93:259-269.
Ding, J.P., Bowman, C.L., Sokabe, M., and Sachs, F. (1989) Mechani- cal transduction in glial cells: SACS and SICS. Biophys. J., 55:244a.
Falke, L.C. and Misler, S. (1989) Activity of ion channels during vol- ume regulation by clonal NlE115 neuroblastoma cells. Proc. Nutl. Acad. Sei. U.S.A., 86:3919-3923.
Grinstein, S., Rothstein, A,, Sarkadi, B., and Gelfand, E.W. (1984) Responses of lymphocytes to anisotonic media: Volume-regulating behavior. Am. J . Physiol., 246, Cell Physiol., 15:C204-C215.
Guharay, F. and Sachs, F. (1984) Stretch-activated single ion currents in tissue cultured embryonic chick skeletal muscle. J. Physiol. (LondJ,
Hamill, O.P., Marty, A,, Neher, E., Sakmann, B., and Sigworth, F.J. (1981) Improved patch-clamp techniques for high-resolution record- ing from cells and cell-free membrane patches. ptlugers Arch.., 391: 8S100.
Hille, B. (1992) Ionic Channels ofExcitabZe Membranes. Sinauer Asso- ciates, Sunderland, MA, 607 pp.
Hoffman, E.K. (1987) Volume regulation in cultured cells. Curr. Topics Membr. Trunsp., 30:125-180.
Kmelberg, H.K., Anderson, E., and Kettenmann, H. (1990b) Swelling- induced changes in electrophysiological properties of cultured astro- cytes and oligodendrocytes. 11. Whole-cell currents. Brain Res., 529: 26Z268.
Kimelberg, H.K. and Frangakis, M.V. (1986) Volume regulation in primary astrocyte cultures. Adu. Biosci., 61:177-186.
Kimelberg, H.K., Goderie, S.K., Higman, S., Pang, S., and Waniewski, R.A. (1990a) Swelling-induced release of glutamate, aspartate and taurine from astrocyte cultures. J . Neurosci., 10:1583-1521.
Kimelberg, H.K. and Kettenmann, H. (1990) Swelling-induced changes in electrophysiological properties of cultured astrocytes and oligo- dendrocytes. I. Effects on membrane potentials, input impedance and cell-cell coupling. Brain Res., 529:255-261.
fimelberg, H.K. and O'Connor, E. (1988) Swelling of astrocytes causes membrane potential depolarization. Gliu, 1:219-224.
Kregenow, F.M. (1981) Osmoregulatory salt transporting mechanisms: control of cell volume in anisotonic media. Annu. Reu. Physiol., 43: 493-505.
Lauf, P.K. (1985) K':Cl- Cotransport: Sulfhydryls, divalent cations,
352:685-701.

ISMS ET AL. 96
and the mechanism of volume activation in a red cell. J. Membrane
Law, R.O. and Burg, M.B. (1991) The role of organic osmolytes in the regulation of mammalian cell volume. Adv. Comp. Environ. Phys-
Lewis, C.A. and Stevens, C.F. (1979) Mechanism of ion permeation through channels in a postsynaptic membrane. In: Membrane Trans- port Processes, Vol. 3. C.F. Stevens and R.W. Tsien, eds. Raven Press, New York, pp. 133-151.
Macknight A.D.C. (1988) Principles of cell volume regulation. Renal Physiol. Biochem., 3 4 1 14-141.
Macleod, R.J. and Hamilton, J.R. (1991) Separate K' and C1- trans- port pathways are activated for regulatory volume decrease in jeju- nal villus cells. Am. J. Physiol., 260:G405-G415.
Morris, C.E. (1990)Mechanosensitive ion channels. J . MembraneBioZ.,
Olson, J.E., Sankar, R., Holzman, D.J., and Fleischacker, D. (1986) Energy-dependent volume regulation in primary culture cerebral astrocytes. J. Cell. Physiol., 128:209-215.
Pasantes-Morales, H., Moran, J . , and Schousboe, A. 11990) Volume- sensitive release of taurine from cultured astrocytes: Properties and mechanism. Glia, 3:427-432.
Pasantes-Morales, H. and Schousboe, A. (1988) Volume regulation in astrocytes: A role for taurine as an osmoeffector. J . Neurosci. Res.,
Bid., 88:l-13.
dol., 9:189-225.
113:93-107.
20:505-509. Patel. A.J. and Hunt, A. (1985) Observations on cell growth and regu-
lation of glutamine synthetase by dexamethasone in primary Lul- tures of forebrain and cerebellar astrocytes. Deu. Brain Res., 18: 175-184.
Puro, D.G. (1891) Stretch-activated channels in human retinal Muller cells. Glia, 4:45&460.
Sachs, F. (1991) Mechanical transduction by membrane ion channels: A mini review. Molec. Cell. Biochem., 104:57-60.
Sakin, H. (1989) A stretch-activated K ' -channel sensitive to cell vol- ume. Proc. Natl. Acad. Sci. U.S.A., 86:1731-1735.
Sanchez-Olea, R., Moran, J . , Schousboe, A., and Pasantes-Morales, H. (1991) Hyposmolarity-activated fluxes of taurine in astrocytes are mediated by diffusion. Neurosci. Lett., 130:23S-236.
Schousboe, A,, Sanchez-Olea, R., Moran, J., and Pasantes-Morales, H. (1991) Hyposmolarity-induced taurine release in cerebellar granule cells is associated with diffusion and not with high-affinity trans- port. J. Neurosci. Re.?., 30:661465.
Sigurdson, W.J. and Morris, C.E. (1989) Stretch-activated ion chan- nels in growth cones of snail neurons. J. Neurosci., 9:2801-2808.
Ubl, J., Murer, H., and Kolb, H.-A. (1988) Ion channels activated by osmotic and mechanical stress in membranes of Opossum kidney cells. J . Membrane B i d , 104:223-232.
Yang, X.-Ch. and Sachs, F. (1990) Characterization of stretch-acti- vated ion channels in Xenopus oocytes. J. Physiol. ~Lond3,431:103- 122.
Yang, X.-Ch. and Sachs, F. (1989) Block of stretch-activated ion chan- nels in Xenopus oocytes by gadolinium and calcium ions. Science, 243:106&1071.
Yantorno, R.E., Coca-Prados, T., Krupin, T., and Civan, M.M. (1989) Volume regulation of cultured, transformed, non-pigmented epithe- lial cells from human ciliary body. Erp. Eye Res., 49:423-437.