mtpa: a crustacean metallothionein that affects hepatopancreatic mitochondrial functions
TRANSCRIPT

Available online at www.sciencedirect.com
www.elsevier.com/locate/yabbi
ABBArchives of Biochemistry and Biophysics 467 (2007) 31–40
MTPA: A crustacean metallothionein that affectshepatopancreatic mitochondrial functions
Eduardo Molto 1, Elena Bonzon-Kulichenko 1, Nilda Gallardo, Antonio Andres *
Biochemistry Section, Faculty of Chemistry, Regional Centre for Biomedical Research (CRIB), University of Castilla-La Mancha,
Camilo Jose Cela, 10, 13071 Ciudad Real, Spain
Received 18 April 2007, and in revised form 21 July 2007Available online 29 August 2007
Abstract
Metallothioneins are cysteine-rich proteins, with a high capacity to bind metallic ions, and for which a precise biological role has notbeen established. Here we investigated the effects of MTPA, a metallothionein from the lobster Panulirus argus, on mitochondrial oxygenconsumption and ROS production. An HPLC-RP-ESI-MS analysis of recombinant MTPA showed that despite its extra Cys, MTPAbinds 6 Zn2+ per molecule akin to other crustacean metallothioneins with 18 Cys. The extra Cys is not involved in zinc binding, sinceits side-chain would be oriented to the outside of the molecule according to a preliminary model of the tridimensional structure ofMTPA. MTPA-Zn2+
6 is imported into the hepatopancreatic mitochondria intermembrane space and inhibits mitochondrial oxygen con-sumption, increasing thereby ROS production. Nevertheless, the stimulation of ROS production by MT-bound Zn2+ is weaker com-pared to equivalent amounts of free Zn2+, suggesting that MTPA protects against oxidative stress. This constitutes the first report onmetallothioneins effects on mitochondrial function in invertebrates and agrees with the results described for mammals, suggesting a con-nection between metallothioneins and energy metabolism.� 2007 Elsevier Inc. All rights reserved.
Keywords: Metallothionein; Invertebrate; Crustacea; Mitochondria; ROS; Zinc
Metallothioneins (MTs)2 comprise a superfamily of lowmolecular weight (less than 9 kDa) cysteine-rich proteins,with a high capacity to bind metallic ions. Upon metalbinding, most MTs fold into two domains. In mammals,the N-terminal b-domain has 9 Cys and binds three diva-lent ions, meanwhile the C-terminal a-domain, with 10Cys, some of which in Cys–Cys tandems, binds four diva-lent ions [1,2]. On the other hand, most crustacean MTs
0003-9861/$ - see front matter � 2007 Elsevier Inc. All rights reserved.
doi:10.1016/j.abb.2007.08.016
* Corresponding author. Fax: +34 926 295 318.E-mail address: [email protected] (A. Andres).
1 These authors contributed equally to this work.2 Abbreviations used: MT, metallothionein; MTPA, Panulirus argus
metallothionein; ROS, reactive oxygen species; ETC, electron transportchain; IPTG, isopropyl-b-D-thiogalactopyranoside; GST, glutathione S-transferase; TFA, trifluoracetic acid; MOPS, 3-(N-morpholino) propanesulfonic acid; H2DCFDA, 2 0,7 0-dichlorodihydrofluorescein diacetate;DCF, dichlorofluorescein; SOD, superoxide dismutase.
have been considered as b/b MTs, since each domain bindsthree divalent ions [3,4].
Although the discovery of the first member of this bigfamily took place in 1957, the precise biological role ofthese proteins has not yet been established and only afew works have contributed to clarify metallothioneins’sfunctions in invertebrates. In these animals, as well as invertebrates, the detoxification of non-essential metals isso far the best-established role for these proteins [5,6].Owing to their induction by heavy metals, invertebrateMTs have attracted much attention as pollution markers[7]. However, other MT functions that have been suggestedin mammals, such as the protection against free radicalsand other oxidants, and the regulation of energy metabo-lism [8], have not been analyzed in invertebrates.
It is widely accepted that the physiological role of MTsis closely related to the coordination of metal ions [8].Hence, to shed some light on MTPA function in the lobster

32 E. Molto et al. / Archives of Biochemistry and Biophysics 467 (2007) 31–40
Panulirus argus, we undertook three main approaches.First, in a previous work [9] we analyzed the metal induc-tion of this MT in different tissues from the lobster, demon-strating that the highest MTPA basal levels are present inthe hepatopancreas of the lobster and that Cd or Zn induceits expression. Second, here we obtained pure MTPA byrecombinant synthesis and analyzed its metal bindingcapacity. Finally, taking into account that mammalianMTs could be imported into hepatic mitochondria andaffect respiration [10], together with our previous data [9],here we studied the effects of MTPA on some hepatopan-creatic mitochondrial functions, and whether the proteinwas imported into hepatopancreatic mitochondria andhow far in the mitochondrion it traveled. For the first timein invertebrates we demonstrate that a crustacean MTenters the intermembrane space of hepatopancreatic mito-chondria and affects in a zinc-dependent manner the rate ofelectron transport chain (ETC) and ROS production in thisorganelle. These results could serve as a starting point forfuture studies intending to establish a connection betweenthe metabolic changes underwent by these animals duringreproduction or development and the regulatory elementsof their aerobic metabolism.
Materials and methods
Experimental animals
American lobsters Homarus americanus (0.5 kg each) were purchasedfrom a local commercial dealer the day of the experiment. Before thehepatopancreas extraction, animals were anesthetized on ice and theircarapace was washed with absolute ethanol. The animals were handledaccording to the laws of the European Union and the guidelines of theNational Institutes of Health, and the experimental protocols wereapproved by the institutional committee of bioethics.
Cloning of the open reading frame of MTPA cDNA and
recombinant protein purification
PCR amplification of the MTPA coding sequence was performed usinga clone containing the full MTPA cDNA [9]. The upstream primer:5 0-CCCGGATCCCCTGGTCCCTGCTGCATC-3 0, introduced a BamHIrestriction site immediately upstream of the ATG initiation codon, andthe downstream primer: 5 0-CGGGTCGACCTAAGGGCAGCATTTGCA-3 0, generated a SalI restriction site just after the stop codon. PCRconditions were as described by Molto et al. [9]. The amplified productswere isolated from 1.2% agarose gels and cloned into the pSTBlue-1 vector(Novagen), which was afterwards digested with BamHI and SalI. TheMTPA coding sequence was directionally subcloned into the compatibleBamHI/SalI sites of the pGEX4T-1 expression vector (a kind gift from Dr.S. Atrian). Before the recombinant protein synthesis, the DNA sequencewas confirmed by automated sequencing. JM105 was used as the recipientEscherichia coli strain for cloning and sequencing. To enhance the pro-duction of recombinant protein, the corresponding pGEX recombinantclones were transformed into the protease-deficient BL-21 E. coli strain.Expression and purification of the metal-MTPA complexes were basicallyperformed as described for H. americanus MT [11]. Exponentially growing400 ml LB cultures of the transformed strain were induced with 100 lMIPTG for 3 h in the presence of 300 lM ZnCl2 and 50 lg/ml ampycillin.The GST-MTPA protein was isolated from the cell lysate by affinitychromatography on glutathione-Sepharose 4B (Amersham PharmaciaBiotech) and the not-bound fraction was collected. The fraction that
remained bound to the matrix was digested with thrombin in order toseparate the MTPA portion from the fusion protein (bound fraction). Thisfraction was further purified by Sephadex G-50 (Amersham PharmaciaBiotech) column equilibrated with 10 mM Tris–HCl, pH 8.6, at 0.16 ml/min. About 0.7 ml fractions were collected and analyzed for proteincontent by their absorbance at 226 nm (MTPA fraction). MTPA fractionswere pooled, lyophilized and stored at �20 �C until further use. Theaffinity matrix was regenerated with reduced glutathione 10 mM in 50 mMTris–HCl, pH 8, and the flow through was collected (eluted fraction). Toprevent MTPA oxidation, nitrogen was bubbled in all the steps of thepurification procedure, which were analyzed on 12.5% SDS–PAGECoomassie blue-stained gels.
Analysis of the recombinant zinc-containing MTPA and its
apo-form
Reversed-phase HPLC on-line with Electrospray ionization mass spec-trometry (RP-HPLC-ESI-MS) was used to assess the purity and the metalbinding stoichiometry of the zinc-containing MTPA. ESI-MS spectra wereperformed on a VG AutoSpec API triple-quadrupole instrument (Fison/VGInstruments). ESI-MS conditions for the apo- and holo-forms were thefollowing: capillary counterelectrode voltage, 4 kV; cone potential, 40 V;source temperature, 335 �C; m/z range, 0–2000. Forty microliters of proteinsolution were injected at 0.6 ml/min in a Vydac C4 5 cm · 2 mm HPLCcolumn for the holo-form, and at 0.3 ml/min in a Zorbax C18 300A15 cm · 2.1 mm HPLC for the apo-form. The running buffer were aceto-nitrile (buffer A) and 10 mM NH4Ac (buffer B) for the holo-form analysis,and 0.1% TFA in acetonitrile (buffer A) and 0.1% TFA in water (buffer B) forthe apo-form analysis. Both forms were eluted with a linear gradient from90% B to 5% B within 30 min, and the protein was detected at 220 nm. Massspectra deconvolutions and molecular weight calculation were performedusing the Agilent ChemStation program from the HPLC 100 MSD system(Agilent Technologies). Stoichiometry determination for Zn2+ binding toMTPA was calculated by the molecular weight difference between the apo-and the holo-form and using the formula of Yu et al. [12].
Apo-form purification for mitochondrial assays
Apo MTPA was prepared by passing the holoprotein through aSephadex G-25 (Amersham Pharmacia Biotech) column equilibratedwith deaerated 10 mM HCl. About 0.6 ml fractions were collected andanalyzed for protein content by their absorbance at 220 nm. Protein-containing fractions were pooled, lyophilized and stored at �20 �C untilfurther use.
MTPA quantitation
For MTPA quantitation by amino acid analysis, samples werehydrolyzed in 6 N HCl with 1% phenol for 1 h at 155 �C in vacuum, andanalyzed using a Beckman 6300 system Gold instrument.
Modelling of the three-dimensional structure of MTPA C-terminal
domain
A preliminary homology model of MTPA C-terminal domain three-dimensional structure was generated using the Swiss-Model software(http://swissmodel.expasy.org) on the basis of C-terminal domain PDBstructures of MT-1 from Callinectes sapidus (1dmc) and of MT fromH. americanus (1j5I). For viewing and analysis of the structures, Swiss-PdbViewer (v3.7; http://www.expasy.org/spdbv) and RasMol (v2.6;http://www.umass.edu/microbio/rasmol/) tools were used.
Isolation of hepatopancreatic mitochondria
Mitochondria from H. americanus hepatopancreas were isolatedaccording to the procedure described by Tsokos et al. [13]. All steps were

E. Molto et al. / Archives of Biochemistry and Biophysics 467 (2007) 31–40 33
performed at 4 �C. Since the hemolymph of the lobster is approximatelyisotonic with sea water, a physiological high osmolarity medium was usedfor the isolation of mitochondrial fraction [13]. The mitochondrial pelletwas resuspended to 30–40 mg/ml in a medium containing 83 mM sucrose,600 mM mannitol, 4 mM MOPS, 0.1% BSA, 1 mM PMSF, 10 lM leu-peptin, pH 7.4. The concentration of mitochondrial protein was quantifiedby the Bradford method [14] using BSA as protein standard. Yields wereapproximately 7.5 mg mitochondrial protein per g wet weight of tissue.
Mitochondrial respiration assays
Mitochondrial oxygen consumption was measured at 27 ± 1 �C usinga Clark electrode with stirring (DW1 model, Hansatech Instruments)coupled to a grafic recorder (3066 model, Yokogawa Hokushin Electric),according to the method of Tsokos et al. [13], with some modifications.Briefly, hepatopancreatic mitochondria were added to a 0.5 ml chambercontaining a physiological high osmolarity respiration buffer (RB) thatmet the requirements of marine organisms (65 mM sucrose, 470 mMmannitol, 167 mM KCl, 3.3 mM MgCl2, 4 mM K2HPO4, 4 mM MOPS,0.067% BSA, pH 7.2), to a final protein concentration of 0.40 ± 0.05 mg/ml. MTPA as well as its apo-form (TPA) were dissolved in RB and addedat a final protein concentration of 10 lM in different order respect tosuccinate and ADP. The final concentration for succinate was 5 mM andfor ADP 0.6 mM, and both were added dissolved in water. Additions didnot exceed 10 ll and were made every 3–5 min. The electrode was cali-brated with oxygen-purge RB by addition of Na2S2O4. The rate of oxygenconsumption was calculated from the recorder tracing and expressed asnmol atomic oxygen per min per mg of protein.
Determination of mitochondrial hydrogen peroxide
Mitochondrial hydrogen peroxide was determined spectrofluoromet-rically using H2DCFDA. Hepatopancreatic mitochondria were suspendedin RB without 470 mM mannitol and incubated with succinate (10 mM)and the fluorescent probe (1 lM), with or without 10 lM MTPA, 10 lMTPA or 60 lM ZnCl2. Fluorescence was determined at 528 nm for emis-sion and 485 nm for excitation, with slit widths of 20 nm. Measurementswere performed at 28 �C, every 30 s, during 15 min, on a microplatespectrofluorometer (Synergy HT, Bio-Tek Instruments). The concentra-tion of DCF generated in mitochondria was determined from standardcurves using known concentrations of DCF added to mitochondria(0.5 mg/ml), and are reported as pmol DCF/min/mg mitochondrialprotein.
Import of MTPA into hepatopancreatic mitochondria
Intact coupled hepatopancreatic mitochondria at a protein concen-tration of 40–50 mg/ml in 1 ml RB containing 1 mM PMSF and 10 lMleupeptin, were incubated with 5 mM succinate for 1 min to initiate res-piration, and afterwards with 0.6 mM ADP for 2 min. MTPA was addedto a final concentration of 10 lM and incubated for 5 min. All was doneunder gentle stirring at room temperature.
Mitochondria were collected by centrifugation at 10,000g for 10 min at4 �C, washed twice with 2 ml ice-cold RB lacking BSA (RB-BSA), andresuspended at a concentration of 60–100 mg/ml protein in RB-BSAcontaining 1 mM PMSF and 10 lM leupeptin for submitochondrialfractionation. Additionally, total mitochondrial protein was obtained bysolubilizing overnight with 1% (v/v) Triton X-100 at 4 �C.
Submitochondrial fractionation
Submitochondrial fractions were prepared according to the method ofSchnaitman et al. [15] with minor modifications. Digitonin (1 mg/10 mgprotein, 20 min) was used to disrupt the outer mitochondrial membrane(OMM), which was collected by a 144,000g centrifugation for 1 h, obtainingthe intermembrane space (IMS) proteins in the supernatant. The innermitochondrial membrane (IMM) was disrupted by Brij 58 (0.3 mg/mg
protein, 15 min), and after a 144,000g centrifugation for 1 h, IMM wasobtained in the pellet and the mitochondrial matrix (MM) in the superna-tant. The four mitochondrial compartments (OMM, IMS, IMM and MM)were identified by the specific activities of the marker enzymes monoamineoxidase (MAO) for the OMM using the fluorimetric Amplex Red MAOAssay kit (Molecular Probes), succinate dehydrogenase (SDH) for the IMMusing the method of Munujos [16], and malate dehydrogenase (MDH) forthe matrix, according to the method of Parker et al. [17] (data not shown).LDH activity with pyruvate as substrate was measured by the method ofRacher [18] to asses the cytosolic contamination of submitochondrial frac-tions (data not shown). Enzyme activities were expressed as specific activities(nmol of transformed substrate/mg total protein/min).
Western blot analysis
For MTPA assessment, samples were boiled for 3 min in Laemmli’ssample buffer with 19 mM b-mercaptoethanol. Proteins were resolved by12.5% SDS–PAGE and transferred to nitrocellulose membranes. Mem-branes were blocked in PBS–0.1%–Tween 20 containing 5% fat skimmeddry milk and incubated overnight at 4 �C with anti-CgMT antibody (1/500dilution). This antibody was developed against the metallothionein fromthe Pacific oyster Crassostrea gigas (CgMT1) [19] and was a generous giftof Dr. Moraga. The bands were visualized after incubation with goat anti-rabbit-peroxidase conjugated IgG, using the enhanced chemiluminiscencemethod.
Statistical analysis
Statistical significance was calculated using one-way ANOVA followedby Tukey test. Results with p < 0.05 were considered to be statisticallysignificant.
Results
Characterization of the Zn-MTPA complexes obtained by
recombinant technology
According to our previous results [9], MTPA has anextra Cys residue (19 residues) in the portion of thesequence corresponding to the C-terminal domain, com-pared to other crustacean MTs that have 18 Cys (9 in theN-terminal domain and 9 in the C-terminal domain)(Fig. 1). In order to analyze if this extra Cys residueendowed MTPA a higher metal binding capacity comparedto other crustacean MTs, we determined the number ofmetal ions bound per protein molecule in recombinantexpressed MTPA in a zinc-supplemented medium.
The Zn-MTPA complexes obtained by the expressionsystem used, present two extra amino acids at the N-termi-nal (N-Gly-Ser) of the protein, due to the introduction ofthe BamHI restriction site. Nevertheless, it has been previ-ously demonstrated, that this modification does not inter-fere with the metal binding capacity of other MTs [20].Besides, as will be seen further on, in our case this modifi-cation has not affected the protein function in mitochon-dria. The final yield of the purification procedure was0.33 mg/l of bacterial culture, which is within the yieldrange described for other MTs obtained by the sameexpression system [11]. SDS–PAGE (Fig. 2, lane 6) analysisof the Zn-MTPA samples showed a single band of around12 kDa, which should be a dimer of the protein.

Fig. 1. Multiple alignment analysis of MTPA with crustacean MTs. ClustalW-aligned MT sequences of the following organisms: Callinectes sapidus,copper thionein and isoforms 1 and 2, Scylla serrata, isoforms 1 and 2, Astacus astacus, Portunus pelagicus, Potamon potamios, Homarus americanus,Carcinus maenas, Panulirus argus. Fully conserved amino acids are showed in dark gray. In light gray are depicted the conserved positions in all but theCu-MT from C. sapidus. Bold are the MTPA residues that break up with the conservation pattern of the rest of crustacean MTs. Arrows signal theinterdomain region in crustacean MTs and the asterisk the extra non-conserved Cys residue.
Fig. 2. Coomassie blue stained 12.5% SDS–PAGE gels from the different fractions of the purification process. Lanes 1 and 8: pre-stained molecular weightmarkers (BenchMark, Invitrogen). Lane 7: 4 lg insulin (in Laemmli buffer without b-ME). Lanes 2–6: lysate, not-bound, bound, eluted and MTPAfractions, respectively; 25 lg of each fraction were loaded, except for bound and MTPA fractions, where 6 lg were loaded.
34 E. Molto et al. / Archives of Biochemistry and Biophysics 467 (2007) 31–40
Typical RP-HPLC-ESI mass spectra for Zn-MTPA andits apo-form are shown in Fig. 3. The dominant chargestate for Zn-MTPA is +5. However, the apo-form presentsan increased abundance of higher charge states, such as +9,+8, +7 and +6, which agrees with its unfolded conforma-tion, in which the functional groups that can be protonatedare more accessible [12]. The RP-HPLC-ESI mass spectraanalysis for Zn-MTPA and its apo-form were used to cal-culate the stoichiometry for Zn2+ binding to MTPA. Asshown in Table 1, the calculated parent species molecularweight for Zn-MTPA and its apo-form are 6554 ± 3 and6172 ± 4, respectively. Thus, the synthesis of MTPA in azinc-supplemented medium led to the formation of aunique metalated species with 6 Zn2+ ions bound.
Model of the three-dimensional structure of MTPA C-terminal
domain
In order to explain from a structural point of view the stoi-chiometry results obtained by RP-HPLC-ESI-MS, accord-
ing to which the extra Cys in the C-terminal domain didnot increase the metal binding capacity of MPTA, we gener-ated a preliminary model of the tridimensional structure ofthis domain, based on the C-terminal domain of MT-1 fromC. sapidus and MT of H. americanus. As shown in Fig. 4, theextra Cys residue situated in the middle of the triplet of Cys,would be oriented to the outside of the molecule. Its sulphurwould be 7.5 and 7.09 A apart from the sulphurs of the closerCys, respectively, which would be the contiguous cysteines.Since, the mean distance between the sulphurs of the Cys thatcould form a tetrahedral coordination in this model is4.12 ± 0.35 A, it is very improbable that this extra Cys wouldbe involved in Zn coordination.
Effect of Zn-MTPA and its apo-form (TPA) on the oxygen
consumption of hepatopancreatic mitochondria from H.
americanus
Taking into account that Zn2+ inhibits mitochondrialrespiration in the lM concentration range [21], we decided
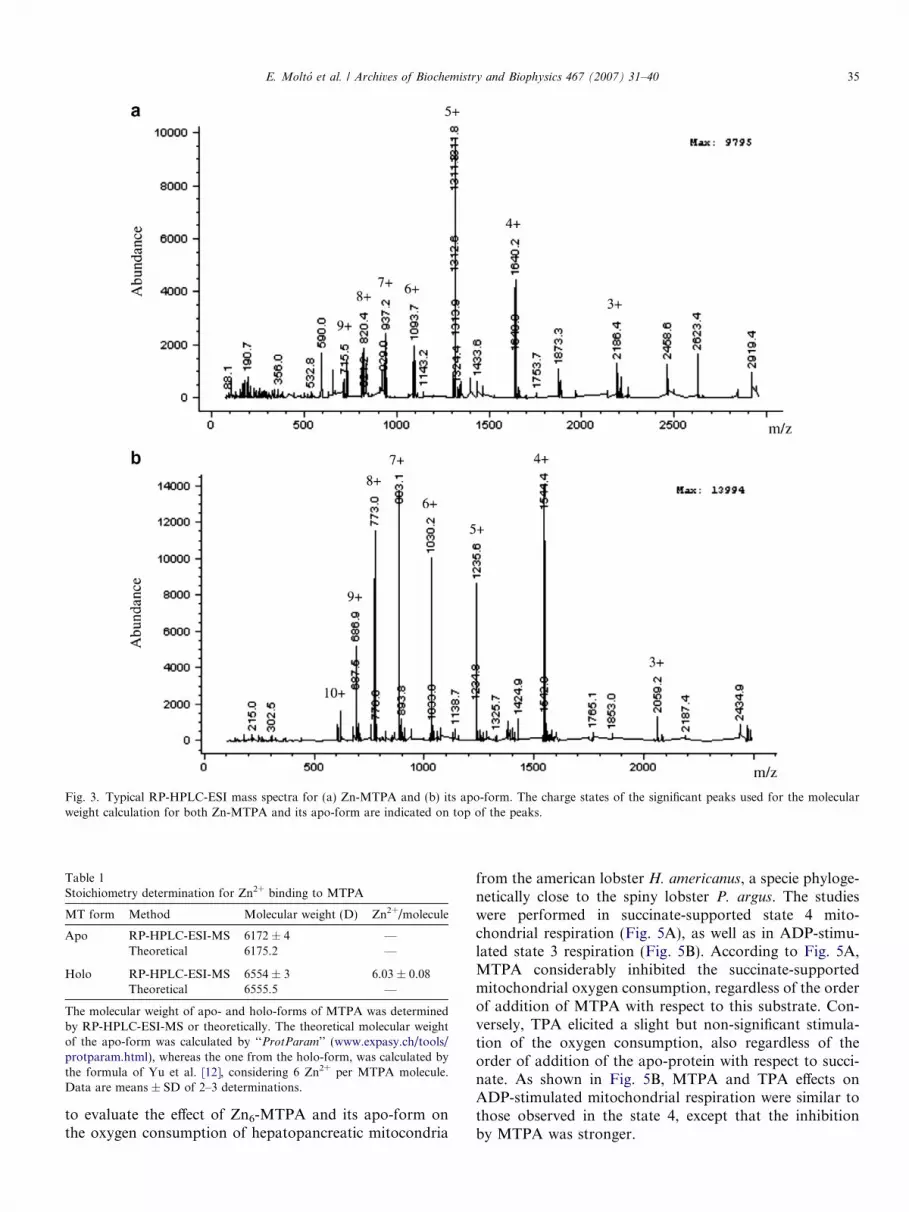
Fig. 3. Typical RP-HPLC-ESI mass spectra for (a) Zn-MTPA and (b) its apo-form. The charge states of the significant peaks used for the molecularweight calculation for both Zn-MTPA and its apo-form are indicated on top of the peaks.
Table 1Stoichiometry determination for Zn2+ binding to MTPA
MT form Method Molecular weight (D) Zn2+/molecule
Apo RP-HPLC-ESI-MS 6172 ± 4 —Theoretical 6175.2 —
Holo RP-HPLC-ESI-MS 6554 ± 3 6.03 ± 0.08Theoretical 6555.5 —
The molecular weight of apo- and holo-forms of MTPA was determinedby RP-HPLC-ESI-MS or theoretically. The theoretical molecular weightof the apo-form was calculated by ‘‘ProtParam’’ (www.expasy.ch/tools/protparam.html), whereas the one from the holo-form, was calculated bythe formula of Yu et al. [12], considering 6 Zn2+ per MTPA molecule.Data are means ± SD of 2–3 determinations.
E. Molto et al. / Archives of Biochemistry and Biophysics 467 (2007) 31–40 35
to evaluate the effect of Zn6-MTPA and its apo-form onthe oxygen consumption of hepatopancreatic mitocondria
from the american lobster H. americanus, a specie phyloge-netically close to the spiny lobster P. argus. The studieswere performed in succinate-supported state 4 mito-chondrial respiration (Fig. 5A), as well as in ADP-stimu-lated state 3 respiration (Fig. 5B). According to Fig. 5A,MTPA considerably inhibited the succinate-supportedmitochondrial oxygen consumption, regardless of the orderof addition of MTPA with respect to this substrate. Con-versely, TPA elicited a slight but non-significant stimula-tion of the oxygen consumption, also regardless of theorder of addition of the apo-protein with respect to succi-nate. As shown in Fig. 5B, MTPA and TPA effects onADP-stimulated mitochondrial respiration were similar tothose observed in the state 4, except that the inhibitionby MTPA was stronger.
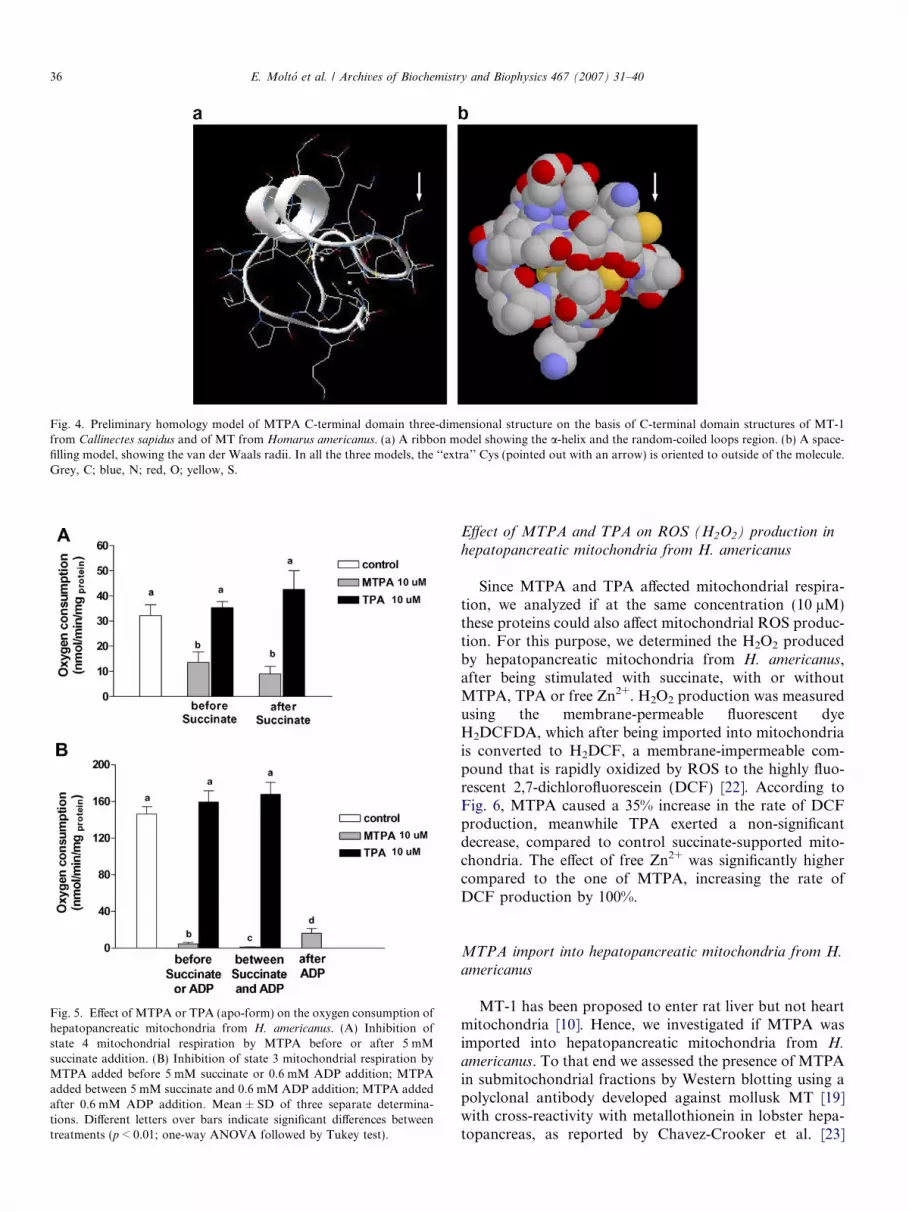
Fig. 4. Preliminary homology model of MTPA C-terminal domain three-dimensional structure on the basis of C-terminal domain structures of MT-1from Callinectes sapidus and of MT from Homarus americanus. (a) A ribbon model showing the a-helix and the random-coiled loops region. (b) A space-filling model, showing the van der Waals radii. In all the three models, the ‘‘extra’’ Cys (pointed out with an arrow) is oriented to outside of the molecule.Grey, C; blue, N; red, O; yellow, S.
Fig. 5. Effect of MTPA or TPA (apo-form) on the oxygen consumption ofhepatopancreatic mitochondria from H. americanus. (A) Inhibition ofstate 4 mitochondrial respiration by MTPA before or after 5 mMsuccinate addition. (B) Inhibition of state 3 mitochondrial respiration byMTPA added before 5 mM succinate or 0.6 mM ADP addition; MTPAadded between 5 mM succinate and 0.6 mM ADP addition; MTPA addedafter 0.6 mM ADP addition. Mean ± SD of three separate determina-tions. Different letters over bars indicate significant differences betweentreatments (p < 0.01; one-way ANOVA followed by Tukey test).
36 E. Molto et al. / Archives of Biochemistry and Biophysics 467 (2007) 31–40
Effect of MTPA and TPA on ROS (H2O2) production in
hepatopancreatic mitochondria from H. americanus
Since MTPA and TPA affected mitochondrial respira-tion, we analyzed if at the same concentration (10 lM)these proteins could also affect mitochondrial ROS produc-tion. For this purpose, we determined the H2O2 producedby hepatopancreatic mitochondria from H. americanus,after being stimulated with succinate, with or withoutMTPA, TPA or free Zn2+. H2O2 production was measuredusing the membrane-permeable fluorescent dyeH2DCFDA, which after being imported into mitochondriais converted to H2DCF, a membrane-impermeable com-pound that is rapidly oxidized by ROS to the highly fluo-rescent 2,7-dichlorofluorescein (DCF) [22]. According toFig. 6, MTPA caused a 35% increase in the rate of DCFproduction, meanwhile TPA exerted a non-significantdecrease, compared to control succinate-supported mito-chondria. The effect of free Zn2+ was significantly highercompared to the one of MTPA, increasing the rate ofDCF production by 100%.
MTPA import into hepatopancreatic mitochondria from H.americanus
MT-1 has been proposed to enter rat liver but not heartmitochondria [10]. Hence, we investigated if MTPA wasimported into hepatopancreatic mitochondria from H.
americanus. To that end we assessed the presence of MTPAin submitochondrial fractions by Western blotting using apolyclonal antibody developed against mollusk MT [19]with cross-reactivity with metallothionein in lobster hepa-topancreas, as reported by Chavez-Crooker et al. [23]
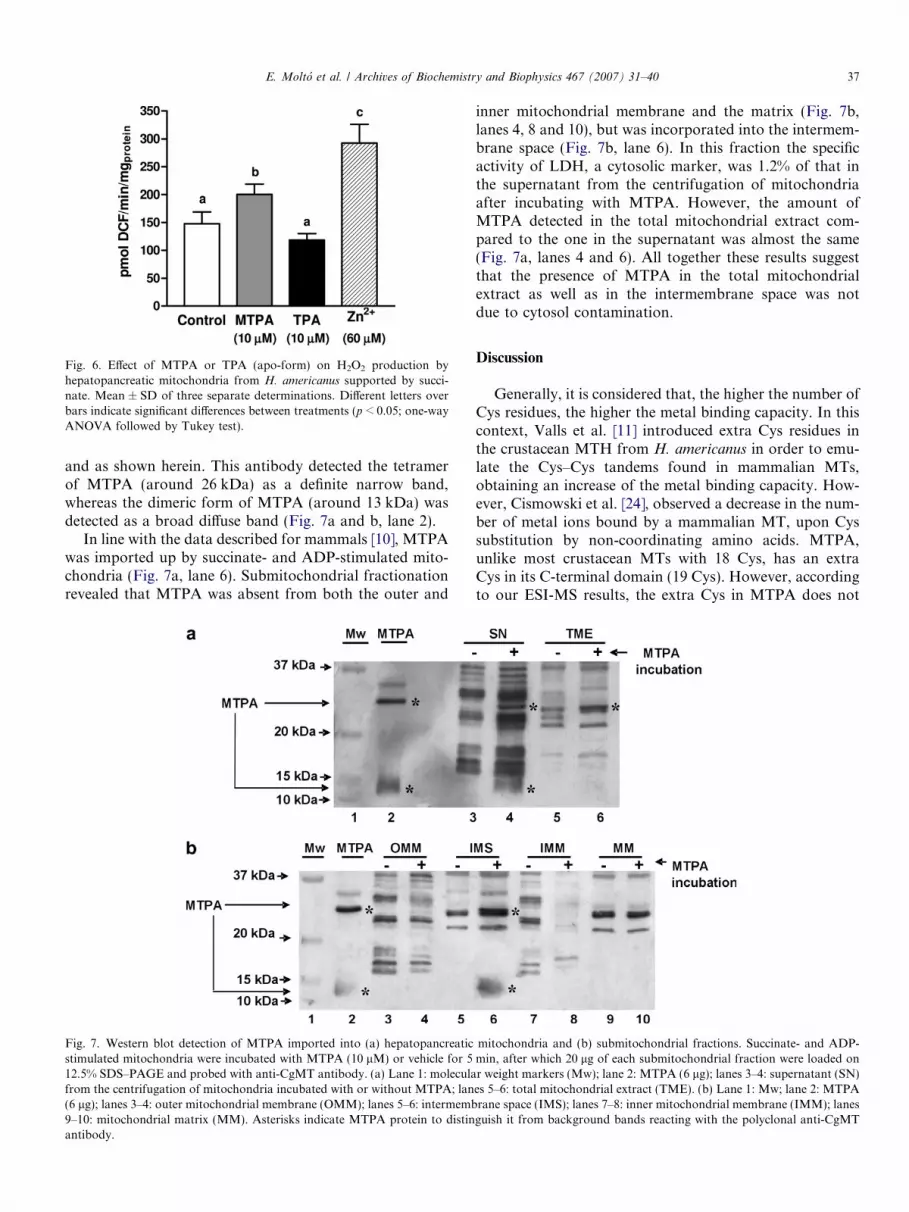
Fig. 6. Effect of MTPA or TPA (apo-form) on H2O2 production byhepatopancreatic mitochondria from H. americanus supported by succi-nate. Mean ± SD of three separate determinations. Different letters overbars indicate significant differences between treatments (p < 0.05; one-wayANOVA followed by Tukey test).
E. Molto et al. / Archives of Biochemistry and Biophysics 467 (2007) 31–40 37
and as shown herein. This antibody detected the tetramerof MTPA (around 26 kDa) as a definite narrow band,whereas the dimeric form of MTPA (around 13 kDa) wasdetected as a broad diffuse band (Fig. 7a and b, lane 2).
In line with the data described for mammals [10], MTPAwas imported up by succinate- and ADP-stimulated mito-chondria (Fig. 7a, lane 6). Submitochondrial fractionationrevealed that MTPA was absent from both the outer and
Fig. 7. Western blot detection of MTPA imported into (a) hepatopancreaticstimulated mitochondria were incubated with MTPA (10 lM) or vehicle for 512.5% SDS–PAGE and probed with anti-CgMT antibody. (a) Lane 1: moleculafrom the centrifugation of mitochondria incubated with or without MTPA; lan(6 lg); lanes 3–4: outer mitochondrial membrane (OMM); lanes 5–6: intermemb9–10: mitochondrial matrix (MM). Asterisks indicate MTPA protein to distinantibody.
inner mitochondrial membrane and the matrix (Fig. 7b,lanes 4, 8 and 10), but was incorporated into the intermem-brane space (Fig. 7b, lane 6). In this fraction the specificactivity of LDH, a cytosolic marker, was 1.2% of that inthe supernatant from the centrifugation of mitochondriaafter incubating with MTPA. However, the amount ofMTPA detected in the total mitochondrial extract com-pared to the one in the supernatant was almost the same(Fig. 7a, lanes 4 and 6). All together these results suggestthat the presence of MTPA in the total mitochondrialextract as well as in the intermembrane space was notdue to cytosol contamination.
Discussion
Generally, it is considered that, the higher the number ofCys residues, the higher the metal binding capacity. In thiscontext, Valls et al. [11] introduced extra Cys residues inthe crustacean MTH from H. americanus in order to emu-late the Cys–Cys tandems found in mammalian MTs,obtaining an increase of the metal binding capacity. How-ever, Cismowski et al. [24], observed a decrease in the num-ber of metal ions bound by a mammalian MT, upon Cyssubstitution by non-coordinating amino acids. MTPA,unlike most crustacean MTs with 18 Cys, has an extraCys in its C-terminal domain (19 Cys). However, accordingto our ESI-MS results, the extra Cys in MTPA does not
mitochondria and (b) submitochondrial fractions. Succinate- and ADP-min, after which 20 lg of each submitochondrial fraction were loaded onr weight markers (Mw); lane 2: MTPA (6 lg); lanes 3–4: supernatant (SN)es 5–6: total mitochondrial extract (TME). (b) Lane 1: Mw; lane 2: MTPArane space (IMS); lanes 7–8: inner mitochondrial membrane (IMM); lanesguish it from background bands reacting with the polyclonal anti-CgMT

38 E. Molto et al. / Archives of Biochemistry and Biophysics 467 (2007) 31–40
increase the metal binding capacity, suggesting that notonly the number of Cys residues matters, but also theirposition in the protein sequence. In fact, the preliminarymodel of the tridimensional structure of the C-terminaldomain of MTPA, showed that the extra Cys residue situ-ated in the middle of the triplet of Cys would be oriented tothe outside of the molecule, with its sulphur too far awayfrom the sulphurs of the closer Cys, to form a tetrahedralcoordination. Although the extra Cys is not located inthe a-helix, but in a more flexible region of loops, it is veryimprobable from an energetic point of view, that the side-chains from three contiguous residues could be oriented inthe same direction, that is, in a cis configuration, since thetrans configuration of the peptide bond is more stable,except when a Pro is involved. In fact, in the structuresof C. sapidus and H. americanus this position is occupiedby a Lys, which is also in trans configuration. Hence,assuming the structure showed in Fig. 4, it is highly prob-able that this extra Cys is not involved in Zn coordination,which is consistent with the ESI-MS results.
Cys triplets in MTs have also been found in the Cu-MTfrom C. sapidus [25], in protists [26], in the C-terminaldomain of the human MT-1b, which has 12 Cys insteadof 11 like the rest of mammal MTs [27]; and in the MTfrom the earth worm Eisenia foetida, composed of onlyone domain with 12 Cys, like the C-terminal domain fromthe human MT-1b [28]. To our knowledge, none of theabove mentioned MTs with Cys triplets have been assayedfor their metal binding capacity, except for the MT from E.
foetida. By MS it has been determined that the MT fromthis worm binds 4 Cd2+ ions, akin to the C-terminaldomain of mammal MTs with 11 Cys, suggesting that theCys not involved in Cd binding could be the extra Cys fromthe triplet, since in MTs from other earth worms this posi-tion is occupied by an Ala instead of a Cys [28]. The extraCys residue in the C-terminal domain of MTPA, whichdoes not increase the Zn binding capacity, resembles theresults obtained for the MT from E. foetida. Whether thisextra Cys in MTPA is a result of a late meaningless muta-tion event, as it was suggested for the earth worm or, on thecontrary, could play an important role for example in theinteraction of the MT with other proteins, remains an openquestion.
The role of MTs in non-essential metal detoxification isso far the best-established role for these proteins in verte-brates, as well as in invertebrates [29,6]. Also, other func-tions have been ascribed to MTs such as the regulationof the energetic metabolism and the protection against oxi-dative stress [30–33]. However, in invertebrates these MTfunctions are still poorly explored and only indirect evi-dences exist connecting MTs and protection against oxida-tive [34] and cold stress [35]. Moreover, relatively littleinformation is available on crustacean mitochondrialmetabolism, specially the one from hepatopancreas, anorgan playing a major role in digestion and storage of inor-ganic as well as organic reserves during the molting cycle[36]. Here we have evaluated the effects of recombinant
MTPA on the oxygen consumption and ROS productionof hepatopancreatic mitochondria from the american lob-ster H. americanus.
MTPA-Zn6 added to ADP-stimulated hepatopancreaticmitochondria caused an 89% inhibition of the oxygen con-sumption. Ye et al. [10] obtained an 80% inhibition adding10 lM MT-2-Zn7 from rabbit liver to ADP-stimulatedhepatic mitochondria. In both cases the effects exceededthose caused by MT-1-Zn7 from rabbit liver, which inhib-ited the mitochondrial respiration by only a 30% [10]. Onthe other hand, Simpkins et al. [37] observed a muchweaker inhibition in ADP-stimulated hepatic mitochondriausing the same MTs employed by Ye et al. [10]. The differ-ence between both assays most likely resided in the use bythe group of Simpkins et al. [37] of a commercial MT fromSigma, which is mostly MT-Cd7 [38], and as observedin vitro, Cd2+ inhibits cytochrome bc1 100 times less thanZn2+ [39].
Since 1967 Zn2+ has been known for inhibiting mitochon-drial respiration at micromolar levels, affecting mainly Com-plex III [21,39], where two zinc-binding sites have beenidentified [40]. However, since under physiological condi-tions free zinc is available only at nano–picomolar levels[41], little attention has been paid to the physiological mean-ing of the zinc ability to inhibit mitochondrial respiration. Inthis context, our results showing that MTPA-Zn6 inhibitsrespiration of crustacean hepatopancreatic mitochondriaat physiological micromolar MT concentrations [42],together with the results obtained in mammals [10,37,43],suggest a possible mechanism by which zinc could exert itsinhibitory effects in mitochondria under physiological condi-tions. Moreover, the effects of the apo-form are opposite tothose of the holo-form, suggesting that the MTPA/TPA sys-tem modulates hepatopancreatic mitochondrial respirationin a zinc-dependent manner, once again agreeing with thedata described for mammals [10].
However, an open question remains concerning themechanism by which a MT could exert the describedeffects. According to the zinc Kd of MTs, the holo-form-mediated inhibition of mitochondrial respiration couldnot be due to the free zinc concentration at equilibriumwith MT, but to thermodynamic/kinetic forces, which areknown to condition the direct zinc transfer from MT tozinc acceptor sites [44]. With this regard, Ye et al. [10] dem-onstrated by Western blot that MT localizes in the inter-membrane space (IMS) from liver mitochondria, and weconfirm these results in hepatopancreatic mitochondriafrom the lobster H. americanus. Nevertheless, it is stillintriguing, how a MT, a protein lacking a mitochondrialtargeting sequences neither at its N-terminal, apparentlynor at an internal sequence, could pass through the outermembrane of hepatic or hepatopancreatic mitochondria.This is also the case of other proteins from the mitochon-drial IMS, such as apocytochrome c [45], SOD1 and itscopper chaperone (CCS) [46], which are synthesized inthe cytosol and are imported into the mitochondria by astill unknown mechanism.

E. Molto et al. / Archives of Biochemistry and Biophysics 467 (2007) 31–40 39
In the case of the cytochrome c hemo lyase from Neu-
rospora crassa, a mitochondrial enzyme that guaranteesthe covalent binding of the hemo group to the cyto-chrome c, the internal mitochondrial targeting sequenceis part of the sequence probably involved in the hemogroup binding [45]. The 30% of this sequence consistsof positively or negatively charged residues. The 25%of MTPA residues are charged, and the number andpositions of the Lys are conserved throughout all MTs,including those from mammals. In mammals it has beendemonstrated that these Lys play a key role in MT inter-nalization into mitochondria [45,47]. Hence, it could bepossible that some of this MTPA Lys could be part ofan internal signal sequence guaranteeing the import ofthis protein into mitochondria. Alternatively, MTPAcould be imported into the IMS through the specificmitochondrial machinery for import and assembly ofIMS, termed MIA, recently identified [48]. The proteinsimported into the IMS contain repetitive Cys motifs,and it has been suggested that a cascade of disulfidebonds is formed during the import of these small pro-teins by the MIA pathway [49]. In this context, sinceMT are small cysteine-rich proteins, it is tempting tospeculate that MTPA could be imported using this spe-cific mechanism. Nevertheless, further work has to bedone before a conclusion may be drawn.
Additionally, since the effects of the apo- and theholo-forms of MTPA on mitochondrial respiration areopposite, we suggest that the holo-form does not looseits Zn2+ ions in order to enter mitochondria, and conse-quently, is translocated through the outer mitochondrialmembrane without loosing its native conformation. Themechanism or transporter involved in MTPA transloca-tion and retention into the mitochondrion remains tobe discovered. Additionally, the ability of MTPA to inhi-bit the respiration of crustacean hepatopancreatic, as wellas mouse hepatic mitochondria (data not shown), indi-cates that the mechanisms of MT import into hepaticmitochondria and ETC inhibition, remained conservedduring evolution.
Zn2+ is able to decrease ADP-stimulated mitochondrialrespiration not only by inhibiting the ETC, but also bycausing mitochondrial swelling [50], which is in turn asso-ciated with the opening of the mitochondrial permeabilitytransition pore, loss of mitochondrial membrane potential,and uncoupling of the oxidative phosphorylation fromETC [51]. However, the addition of at least 1 mM Mg2+
to the mitochondrial respiration buffer, prevents for20 min the zinc-induced mitochondrial swelling [52]. Tak-ing this into account, we included Mg2+ in the respirationbuffer at 3.3 mM in the assay with hepatopancreatic mito-chondria, and at 5 mM, with hepatic mitochondria.Besides, the longest incubation with MTPA (added beforesuccinate) did not exceed 12 min. Hence, MTPA-Zn-induced inhibition of mitochondrial respiration is due toan inhibition of the ETC and not to changes in mitochon-drial integrity, since an uncoupling would have caused an
increase in oxygen consumption of succinate-supportedmitochondria.
Additionally, our results concerning ROS productionsupport the notion that MTPA acts as an inhibitor of theETC and not as an uncoupler, since, as it has been demon-strated in mammal mitochondria, uncouplers suppressROS formation in respiration sate 4 [53], whereas ETCinhibitors increase ROS formation by causing an accumu-lation of the transporters in a reduced state before the siteof inhibition, thereby favoring the production of superoxide radicals [54].
The addition of free Zn2+ in amounts equivalent tothose bound by MTPA, increases 4 fold mitochondrialH2O2 production. Hence, by binding Zn, MTPA preventsthe generation of higher levels of ROS by the free formof the metal, suggesting that MTPA exerts a protectiveeffect against ROS production. These results agree wellwith the reports in mammals [32] and in mollusks [34].
The different effects of free or MTPA-bound Zn2+ couldbe explained by its site of action. After being transferred byMTPA to the mitochondrial IMS, zinc would bind mainlyto Complex III, and would be retained in this mitochon-drial compartment without entering the matrix. However,when added in its free form, zinc not only would bind toComplex III inhibiting ETC, but could also somehowreach the mitochondrial matrix, inhibiting enzymes of theoxidative stress defense [55], thereby causing higher levelsof ROS formation. With this regard, the results of Costelloet al. [56] point at the existence of a putative zinc trans-porter associated with the inner mitochondrial membranethat is important in the zinc trafficking into the mitochon-drial matrix. Finally, the protective role of Zn-MTPA act-ing by some mechanism of radical scavenging cannot beruled out.
To our knowledge, our study constitutes the first reporton a MT affecting crustacean hepatopancreatic mitochon-drial functions, suggesting a connection between MTs andenergetic metabolism in invertebrates. Such a connectionhas already been reported in mammals, since MT-null micebecome mildly obese [57] and endotoxin treated MT �/�mice show lower blood and liver lactate levels comparedto normal MT +/+ mice [33]. On the other hand, in a typ-ical crustacean, considerable amounts of metals (Ca, Zn,Cu) are stored in the hepatopancreas in the premolt stage[36]. The ratios of Cu/Zn bound to the MTs fractionchanges from predominantly Cu during intermolt to Znduring premolt, and then back to Cu after ecdysis, suggest-ing an accumulation of Zn in MTs prior to the molt [58].Additionally, according to our previous results [9], MTPAis induced in the hepatopancreas by high zinc concentra-tions. Taken together, these findings suggest that duringthe premolt stage of P. argus hepatopancreatic MTPA lev-els could increase, which in the light of the data presentedherein, could led to a decrease in the metabolic rate, allow-ing the lobster to endure the soft shell stage, when feeding islimited and metabolic energy should be preserved. Thishypothesis needs confirmation in future studies.

40 E. Molto et al. / Archives of Biochemistry and Biophysics 467 (2007) 31–40
Acknowledgments
This work was supported in part by Research GrantsBFU2005-07647-C03-03 from Ministerio de Ciencia y Tec-nologıa and GC-04-008 from Junta de Comunidades deCastilla-La Mancha (JCCM), Spain. E.M. was supportedby a Research Training Grant PAC-05-003-1 from JCCM,and E.B.-K. by a fellowship from JCCM, Spain. We thankDr. Satrustegui for the advice and discussion on mitochon-drial assays, Dr. Moraga for providing the anti-CgMTantibody, and Dr. Gomez-Puertas for his assistance with3D models.
References
[1] A.H. Robbins, D.E. McRee, M. Williamson, S.A. Collett, N.H.Xuong, W.F. Furey, B.C. Wang, C.D. Stout, J. Mol. Biol. 221 (1991)1269–1293.
[2] K. Zangger, G. Oz, J.D. Otvos, I.M. Armitage, Protein Sci. 8 (1999)2630–2638.
[3] S.S. Narula, M. Brouwer, Y. Hua, I.M. Armitage, Biochemistry 34(1995) 620–631.
[4] A. Munoz, F.H. Forsterling, C.F. Shaw 3rd, D.H. Petering, J. Biol.Inorg. Chem. 7 (2002) 713–724.
[5] R. Dallinger, B. Berger, P. Hunziker, J.H. Kagi, Nature 388 (1997)237–238.
[6] S.C. Swain, K. Keusekotten, R. Baumeister, S.R. Sturzenbaum, J.Mol. Biol. 341 (2004) 951–959.
[7] R. Dallinger, Appl. Biochem. Biotechnol. 48 (1994) 27–31.[8] M. Vasak, D.W. Hasler, Curr. Opin. Chem. Biol. 4 (2000) 177–183.[9] E. Molto, E. Bonzon-Kulichenko, A. del Arco, D.M. Lopez-Alanon,
O. Carrillo, N. Gallardo, A. Andres, Gene 361 (2005) 140–148.[10] B. Ye, W. Maret, B.L. Vallee, Proc. Natl. Acad. Sci. USA 98 (2001)
2317–2322.[11] M. Valls, R. Bofill, R. Gonzalez-Duarte, P. Gonzalez-Duarte, M.
Capdevila, S. Atrian, J. Biol. Chem. 276 (2001) 32385–32843.[12] X. Yu, M. Wojciechowski, C. Fenselau, Anal. Chem. 65 (1993) 1355–
1359.[13] J. Tsokos, R. Kreisberg, A. Michaels, B. Komm, J. Linton, Arch.
Biochem. Biophys. 224 (1983) 707–717.[14] M.M. Bradford, Anal. Biochem. 72 (1976) 248–254.[15] C. Schnaitman, V.G. Erwin, J.W. Greenawalt, J. Cell Biol. 32 (1967)
719–735.[16] P. Munujos, J. Coll-Canti, F. Gonzalez-Sastre, F.J. Gella, Anal.
Biochem. 212 (1993) 506–509.[17] D.M. Parker, A. Lodola, J.J. Holbrook, Biochem. J. 173 (1978) 959–
967.[18] A. Racher, LDH assay, in: A. Doyle, J.B. Griffiths (Eds.), Cell and
Tissue Culture: Laboratory Procedures in Biotechnology, John Wiley& Sons, Chichester, New York, Weinheim, 1998, pp. 71–75.
[19] I. Boutet, A. Tanguy, M. Auffret, R. Riso, D. Moraga, Environ.Toxicol. Chem. 21 (2002) 1009–1014.
[20] M. Capdevila, N. Cols, N. Romero-Isart, R. Gonzalez-Duarte, S.Atrian, R. Gonzalez-Duarte, Cell Mol. Life Sci. 53 (1997) 681–688.
[21] V.P. Skulachev, V.V. Chistyakov, A.A. Jasaitis, E.G. Smirnova,Biochem. Biophys. Res. Commun. 26 (1967) 1–6.
[22] H. Sauer, B. Klimm, J. Hescheler, FASEB J. 15 (2001) 2539–2541.[23] P. Chavez-Crooker, P. Pozo, H. Castro, M.S. Dice, I. Boutet, A.
Tanguy, D. Moraga, G.A. Ahearn, Comp. Biochem. Physiol. C 136(2003) 213–224.
[24] M.J. Cismowski, S.S. Narula, I.M. Armitage, M.L. Chernaik, P.C.Huang, J. Biol. Chem. 266 (1991) 24390–24397.
[25] R.A. Syring, T. Hoexum-Brouwer, M. Brouwer, Comp. Biochem.Physiol. C. Toxicol. Pharmacol. 125 (2000) 325–332.
[26] E. Piccinni, W. Staudenmann, V. Albergoni, R. De Gabrieli, P.James, Eur. J. Biochem. 226 (1994) 853–859.
[27] A. Heguy, A. West, R.I. Richards, M. Karin, Mol. Cell Biol. 6 (1986)2149–2157.
[28] C. Gruber, S. Sturzenbaum, P. Gehrig, R. Sack, P. Hunziker, B.Berger, R. Dallinger, Eur. J. Biochem. 267 (2000) 573–582.
[29] C.D. Klaassen, J. Liu, S. Choudhuri, Annu. Rev. Pharmacol.Toxicol. 39 (1999) 267–294.
[30] J.H. Beattie, D.J. Black, A.M. Wood, P. Trayhurn, Am. J. Physiol.270 (1996) R971–R977.
[31] J.S. Lazo, Y. Kondo, D. Dellapiazza, A.E. Michalska, K.H. Choo,B.R. Pitt, J. Biol. Chem. 270 (1995) 5506–5510.
[32] J.S. Lazo, S.M. Kuo, E.S. Woo, B.R. Pitt, Chem. Biol. Interact. 111–112 (1998) 255–262.
[33] A.M. Rofe, J.C. Philcox, P. Coyle, Biol. Trace Elem. Res. 75 (2000)87–97.
[34] A. Viarengo, B. Burlando, M. Cavaletto, B. Marchi, E. Ponzano, J.Blasco, Am. J. Physiol. 277 (1999) R1612–R1619.
[35] T.E. English, K.B. Storey, J. Exp. Biol. 206 (2003) 2517–2524.[36] R. Gibson, P.L. Barker, Oceanogr. Mar. Biol. Ann. Rev. 17 (1979)
285–346.[37] C. Simpkins, S. Balderman, E. Mensah, J. Surg. Res. 80 (1998) 16–
21.[38] H. Chassaigne, R. Lobinski, Anal. Chem. 70 (1998) 2536–2543.[39] T.A. Link, G. von Jagow, J. Biol. Chem. 270 (1995) 25001–
25006.[40] E.A. Berry, Z. Zhang, H.D. Bellamy, L. Huang, Biochim. Biophys.
Acta 1459 (2000) 440–448.[41] T.J. Simons, J. Membr. Biol. 123 (1991) 63–71.[42] Z.A. Shaikh, in: J.F. Riordan, B.L. Vallee (Eds.), Methods in
Enzymology, Academic Press, New York, 1991, p. 27.[43] C.O. Simpkins, H.L. Zhao, C.A. Torrence, Life Sci. 55 (1994) 221–
226.[44] W. Maret, B.L. Vallee, Proc. Natl. Acad. Sci. USA 95 (1998) 3478–
3482.[45] K. Diekert, G. Kispal, B. Guiard, R. Lill, Proc. Natl. Acad. Sci. USA
96 (1999) 11752–11757.[46] L.A. Sturtz, K. Diekert, L.T. Jensen, R. Lill, V.C. Culotta, J. Biol.
Chem. 276 (2001) 38084–38089.[47] K. Diekert, A.I. de Kroon, U. Ahting, B. Niggemeyer, W. Neupert,
B. de Kruijff, R. Lill, EMBO J. 20 (2001) 5626–5635.[48] A. Chacinska, S. Pfannschmidt, N. Wiedemann, V. Kozjak, L.K.
Sanjuan-Szklarz, A. Schulze-Specking, K.N. truscott, B. Guiard, C.Meisinger, N. Pfanner, EMBO J. 23 (2004) 3735–3746.
[49] N. Mesecke, N. Terziyska, C. Kozany, F. Baumann, W. Neupert, K.Hell, J.M. Herrmann, Cell 121 (2005) 1059–1069.
[50] W.D. Cash, H.L. Aanning, H.E. Carlson, S.W. Cox, E.A. Ekong,Arch. Biochem. Biophys. 128 (1968) 456–459.
[51] M. Zoratti, I. Szabo, Biochim. Biophys. Acta 1241 (1995) 139–176.
[52] J. Wudarczyk, G. Debska, E. Lenartowicz, Arch. Biochem. Biophys.363 (1999) 1–8.
[53] A. Boveris, B. Chance, Biochem. J. 134 (1973) 707–716.[54] J.F. Turrens, J. Physiol. 552 (2003) 335–344.[55] M.P. Rigobello, M.T. Callegaro, E. Barzon, M. Benetti, A. Bindoli,
Free Radic. Biol. Med. 24 (1998) 370–376.[56] L.C. Costello, Z.H. Guan, R.B. Franklin, P. Feng, J. Inorg. Biochem.
98 (2004) 664–666.[57] J.H. Beattie, A.M. Wood, A.M. Newman, I. Bremner, K.H. Choo,
A.E. Michalska, J.S. Duncan, P. Trayhurn, Proc. Natl. Acad. Sci.USA 95 (1998) 358–363.
[58] D.W. Engel, M. Brouwer, R. Mercado-Allen, Mar. Environ. Res. 52(2001) 257–269.