crustacean zooplankton communities in lakes recovering from acidification
TRANSCRIPT

Crustacean zooplankton communities in lakes
recovering from acidification
W. Keller, N.D. Yan, K.M. Somers, and J.H. Heneberry
Abstract: Large reductions in sulphur emissions at the Sudbury, Ont., Canada, smelters in recent decades have resultedin decreased lake acidity, and biological improvements have followed. Lakes in the Sudbury area offer a very uniqueopportunity to develop our understanding of the processes regulating biological restructuring in aquatic ecosystems re-covering from acidification. Here, we examine changes in crustacean zooplankton communities that have accompaniedthe chemical recovery of Whitepine and Sans Chambre lakes, near Sudbury, over the last two decades. In both theseformerly acidic lakes, pH has increased to ~6.0, and some zooplankton community recovery has occurred. However,zooplankton communities have not completely recovered based on multivariate comparisons with the community com-position of reference lakes. Although a number of acid-sensitive species have appeared, many did not persist, or didnot achieve abundances typical of the reference lakes. This indicates that zooplankton community recovery will mostlikely depend on biotic and abiotic interactions within these lakes and not on factors affecting species dispersal. Bothchemical and biological factors have large influences on biological recovery processes. Assessing biological recovery isvery important since the restoration of healthy aquatic communities is a major objective of large-scale sulphur emissioncontrol programs.
Résumé: Les réductions importantes des émissions de soufre dans les fonderies de Sudbury, Ont., Canada, au coursdes dernières décennies ont entraîné une diminution de l’acidité des lacs et, par conséquent, une amélioration desconditions biologiques. Les lacs de la région de Sudbury fournissent ainsi une occasion unique de comprendre lesprocessus de restructuration des écosystèmes aquatiques qui se remettent de l’acidification. Nous examinons ici leschangements dans les communautés des crustacés du zooplancton qui ont suivi l’amélioration des conditions chimiquesau cours des 20 dernières années dans les lacs Whitepine et Sans-Chambre, près de Sudbury. Dans ces deux lacsautrefois acidifiés, le pH a augmenté à ~6,0 et une certaine récupération de la communauté du zooplancton s’estproduite. Cependant, des comparaisons multidimensionnelles avec des lacs témoins révèlent que les communautészooplanctoniques ne sont pas totalement rétablies. Bien qu’un nombre d’espèces sensibles à l’acidité soient apparues,plusieurs n’y sont pas demeurées ou n’ont pas réussi à atteindre les densités habituelles des lacs témoins. Cela indiqueque le rétablissement de la communauté zooplanctonique dépendra très probablement d’interactions biotiques et abioti-ques à l’intérieur de ces lacs, plutôt que de facteurs reliés à la dispersion des espèces. Tant les facteurs chimiques quebiologiques agissent fortement sur les processus de récupération biologique. L’évaluation de la récupération biologiqueest très importante, car la restauration de la santé des communautés aquatiques est l’un des objectifs principaux desprogrammes de contrôle à grande échelle des émissions de soufre.
[Traduit par la Rédaction] Keller et al. 735
Introduction
Excessive acid deposition has acidified many thousands oflakes, with resulting damage to their biological communities(Schindler 1988). During the 1980s and 1990s, massive sul-phur emission control programs were implemented in manyareas including Canada, the United States, and Europe tocombat the global problem of acid deposition. With the im-
plementation of emission control programs, it was expectedthat conditions in affected aquatic ecosystems would im-prove, and the focus of acid rain assessment programs beganto change from documenting damage to investigating eco-system recovery.
Other than in the Sudbury, Ont., Canada, area, evidence ofthe chemical recovery of lakes is only beginning to emergefrom regions where sulphur emissions and deposition have
Can. J. Fish. Aquat. Sci.59: 726–735 (2002) DOI: 10.1139/F02-042 © 2002 NRC Canada
726
Received 21 September 2001. Accepted 31 March 2002. Published on the NRC Research Press Web site at http://cjfas.nrc.ca on24 May 2002.J16542
W. Keller 1 and J.H. Heneberry. Ontario Ministry of the Environment, Cooperative Freshwater Ecology Unit,Laurentian University, Ramsey Lake Road, Sudbury, ON P3E 2C6, Canada.N.D. Yan. Biology Department, York University, 4700 Keele Street, Toronto, ON M3J 1P3, Canada.K.M. Somers. Ontario Ministry of the Environment, Dorset Environmental Science Centre, P.O. Box 39, Dorset, ON P0A 1E0,Canada.
1Corresponding author (e-mail: [email protected]).

been reduced (Stoddard et al. 1999). Evidence of biologicalrecovery from lake acidification is even more limited. Lake-scale manipulations (Keller et al. 1990a; Degerman et al.1995; Sampson et al. 1995) indicate that biological improve-ments should follow the water quality improvements ex-pected to result from sulphur emission reductions. However,evidence of the natural recovery of aquatic communities fromacidification is rare.
Some biological recovery, as well as striking chemical re-covery, has been documented in lakes around Sudbury(Keller et al. 1992a, 1999) where emission reductions fromarea metal smelters were particularly large and began in theearly 1970s. However, since recovery is at an early stage inmany Sudbury area lakes, previous studies have not alloweddetailed examination of long-term recovery patterns in lakes,which had reached pH levels that would not be expected tostill severely limit acid-sensitive aquatic biota. A pH ofabout 6.0 appears to be necessary for the protection of manyacid-sensitive taxa (Schindler 1988; Havens et al. 1993).With continuing chemical recovery in the Sudbury area,some lakes have now reached pH ~ 6.0. This chemical re-covery permits evaluation of the natural biological recoverythat has followed water quality improvements in lakes thathave not had direct chemical manipulations. There can, ofcourse, be different definitions of recovery. Here we acceptthat various outcomes may reasonably be expected from thebiological recovery process (Keller and Yan 1998; Keller etal. 1999) and we consider recovery as the return to a typical,not necessarily a pre-stress, condition. Assessments of bio-logical recovery are very important to the evaluation of acidrain controls, since the protection and restoration of aquaticcommunities is a major ultimate objective of emission reduc-tion programs. Empirical documentation of the patterns of re-covery in damaged lakes also offers important opportunitiesto increase our more general understanding of the processesregulating aquatic community structure and biodiversity.
This paper examines long-term patterns in crustaceanplankton communities in two Sudbury area lakes that haveexhibited chemical recovery to pH ~ 6.0, in response tosmelter emission reductions. To assess recovery in the crus-tacean zooplankton communities of these two lakes we havefollowed the approach of Yan et al. (1996), and comparetemporal patterns in univariate and multivariate measures ofcommunity structure over a two-decade period to compara-ble spatial survey data for a set of 47 reference lakes, nearDorset, Ont., that have not shown any general temporaltrends in pH during the study period. The range of variabil-ity in the zooplankton community data for the referencelakes provides our recovery targets.
Methods
Study lakesThe Sudbury lakes studied (Fig. 1), Whitepine (lat
47°17′N, long 80°50′W) and Sans Chambre (lat 46°43′N,long 81°07′W), are within the large area historically affectedby the Sudbury smelter emissions. A summary of generalphysical and chemical characteristics of the lakes is providedin Table 1. As part of the widespread water quality improve-ments seen in this region (Keller et al. 1992b), these lakes
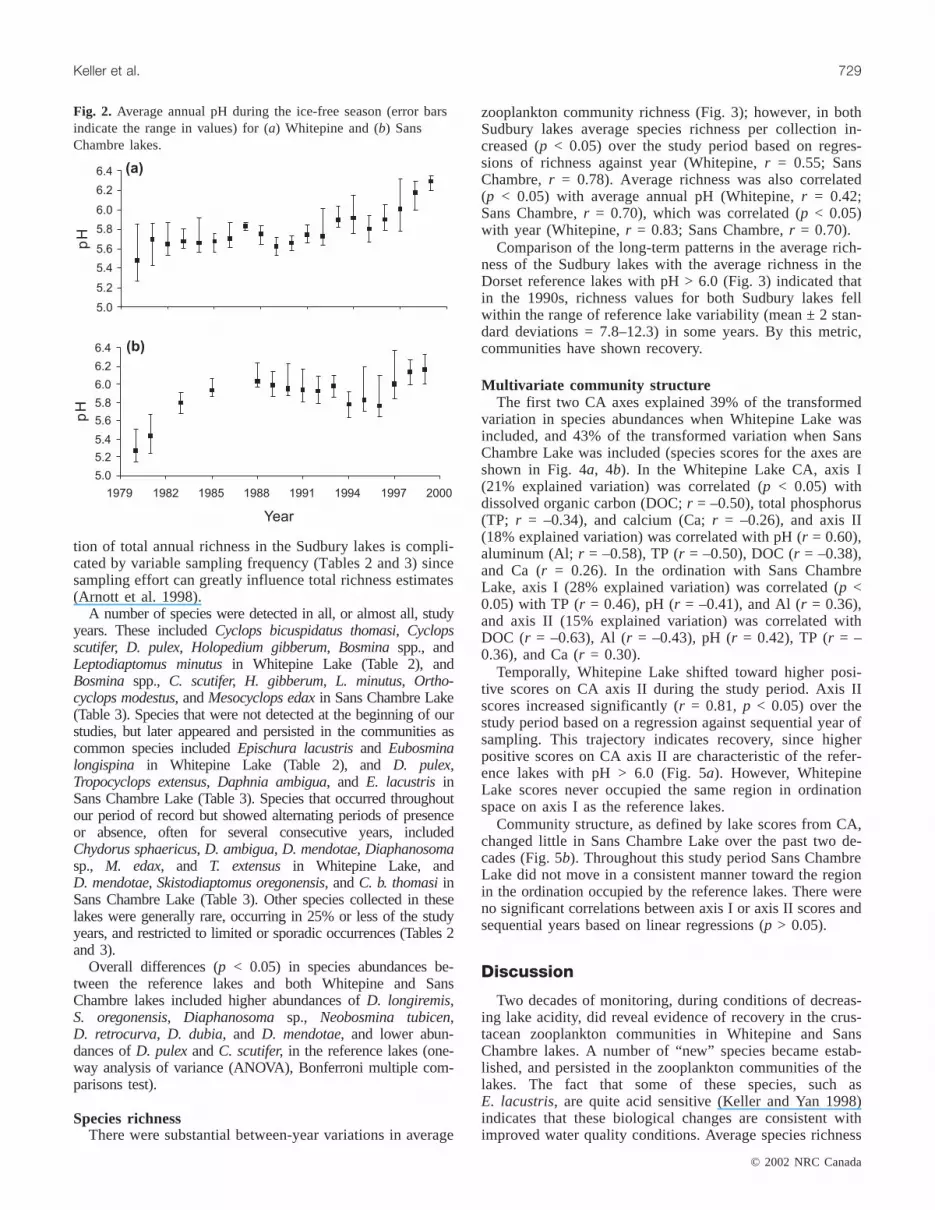
have decreased in acidity following the implementation oflarge reductions (about 90% since the peak in the 1960s) insulphur emissions from the Sudbury area metal smelters dur-ing the 1970s and 1990s. Both these lakes had average pH <5.5 when monitoring commenced in 1980, a pH level atwhich substantial effects on sensitive zooplankton speciesare expected (Keller et al. 1990b; Havens et al. 1993). Bothlakes now have average pH > 6.0 (Fig. 2).
The Dorset reference lakes are located about 200 km south-east of Sudbury (Fig. 1) and are zoogeographically similar tothe Sudbury lakes with respect to zooplankton (Sprules1977). The lakes span a range of chemistry and morpho-metry, which encompasses the conditions in Whitepine andSans Chambre lakes. General physical and chemical charac-teristics of the Dorset lakes are summarized in Table 1. Ad-ditional descriptions of the Dorset lakes are provided in Yanet al. (1996).
Sampling and analysisZooplankton were collected from both of the Sudbury
lakes between 1980 and 1999. Whitepine Lake was sampledin all years, whereas some years (1982, 1984, 1986, 1987)were not included in the monitoring of Sans Chambre Lake.Samples were usually collected monthly during the ice-freeseason; however, sampling frequency was higher in someyears in the early 1980s, and slightly reduced in some yearsin the mid-1990s (Tables 2 and 3). The 47 Dorset referencelakes were sampled monthly during the ice-free season forone year each (1983, 1984, 1987, or 1988).
All zooplankton samples were collected as vertical towsthrough the water column with 12.5-cm-diameter, 80-µm-mesh nets. Metered nets were used for the Dorset lakes andfor the Sudbury lakes after 1981 to allow correction for fil-tration efficiency. Data for Sudbury lakes from 1980 and1981 were adjusted for filtration efficiency by applying themean efficiency for metered net collections in each lake dur-ing the 1980s. Single hauls from 1 m above bottom weretaken in Sans Chambre Lake. In Whitepine Lake and each ofthe Dorset lakes, a series of hauls from three to seven fixeddepths in the water column, at a single site, was taken andcomposited. Depths were chosen so that the composite sam-ple was weighted to account for lake stratum volume.
Samples were preserved with buffered sugar formalin to afinal formalin concentration of 4%. Subsequently, a minimumof 250 crustacean zooplankton were identified and counted.Cladocera and mature Copepoda were identified to speciesand immature Copepoda were identified to suborder. To dealwith changes in nomenclature over the course of the study wehave combined counts of different species ofDiaphanosomato genus, have combinedDaphnia dentifera with Daphniamendotae, and have combinedDaphnia pulicaria and Daph-nia catawba with Daphnia pulex.
Annual average species richness per collection and abun-dance were determined by averaging monthly data. Wheremore than one sample was available for a month, these val-ues were averaged, to provide a monthly value, before gen-erating annual averages. High numbers of animals (up to~500 and ~700 animals per sample, in Whitepine and SansChambre, respectively) were counted in some samples in afew early years. Because richness is related to the number of
© 2002 NRC Canada
Keller et al. 727

animals counted, richness for these lakes was standardizedto a count of 250 animals, using relationships that had beenestablished previously (Keller and Yan 1991). No correctionwas applied to the Dorset reference lakes because approxi-mately 250 animals were counted in all samples.
Following Yan et al. (1996) we employed correspondenceanalysis (CA) to generate our multivariate metrics of zoo-plankton community structure. Our input to the CA wasaverage annual species abundance data transformed to log(x +1) + 0.2. The factor of 0.2 was applied to further down-weight the influence of uncommon species (K. Somers, un-published data). The rarest species, includingAcantholeberiscurvirostris, Acroperus harpae, Eubosmina coregoni, Eucy-
clops speratus, andAglaodiaptomus leptopus, were excludedfrom the analyses. Analyses were run separately for White-pine Lake with the reference lakes, and for Sans ChambreLake with the reference lakes.
Results
Species occurrences and abundancesDuring the 1980s and 1990s, 31 and 24 species of crusta-
cean plankton were recorded in Whitepine and SansChambre lakes, respectively (Tables 2 and 3). Between 8 and16, and 6 and 16 species were found in any one year inWhitepine and Sans Chambre lakes, respectively, but evalua-
© 2002 NRC Canada
728 Can. J. Fish. Aquat. Sci. Vol. 59, 2002
Fig. 1. Locations of Whitepine and Sans Chambre lakes, near Sudbury, Ont., Canada, and the Dorset reference lakes.
Dorset Sudbury
pH > 6.0 (n = 22) pH < 6.0 (n = 25) Whitepine Sans Chambre
Distance from Sudbury (km) ~200 ~200 89 30Area (ha) 13.1–679.0 7.2–195.0 66.9 14.5Mean depth (m) 5.2–22.0 1.8–12.4 5.9 5.6Max. depth (m) 13.7–61.0 5.8–36.5 22.0 15.0Secchi depth (m) 2.9–7.9 1.3–9.3 6.4 4.5pH 6.0–6.7 5.3–5.9 6.3 6.2Ca (mg·L–1) 2.1–4.0 1.2–2.8 1.8 1.7Total P (µg·L–1) 4–18 4–22 5 8DOC (mg·L–1) 1.5–5.5 1.5–8.7 2.5 2.9
Note: Sudbury chemistry data are ice-free period averages of monthly samples from 1999. Dorset chemistry data are ice-freeperiod averages of monthly samples from one of 1983, 1984, 1987, or 1988. Max, maximum; DOC, dissolved organic carbon.
Table 1. General characteristics of the Sudbury and Dorset study lakes.
© 2002 NRC Canada
Keller et al. 729
tion of total annual richness in the Sudbury lakes is compli-cated by variable sampling frequency (Tables 2 and 3) sincesampling effort can greatly influence total richness estimates(Arnott et al. 1998).
A number of species were detected in all, or almost all, studyyears. These includedCyclops bicuspidatus thomasi, Cyclopsscutifer, D. pulex, Holopedium gibberum, Bosmina spp., andLeptodiaptomus minutus in Whitepine Lake (Table 2), andBosmina spp., C. scutifer, H. gibberum, L. minutus, Ortho-cyclops modestus, andMesocyclops edax in Sans Chambre Lake(Table 3). Species that were not detected at the beginning of ourstudies, but later appeared and persisted in the communities ascommon species includedEpischura lacustris and Eubosminalongispina in Whitepine Lake (Table 2), andD. pulex,Tropocyclops extensus, Daphnia ambigua, and E. lacustris inSans Chambre Lake (Table 3). Species that occurred throughoutour period of record but showed alternating periods of presenceor absence, often for several consecutive years, includedChydorus sphaericus, D. ambigua, D. mendotae, Diaphanosomasp., M. edax, and T. extensus in Whitepine Lake, andD. mendotae, Skistodiaptomus oregonensis, andC. b. thomasi inSans Chambre Lake (Table 3). Other species collected in theselakes were generally rare, occurring in 25% or less of the studyyears, and restricted to limited or sporadic occurrences (Tables 2and 3).
Overall differences (p < 0.05) in species abundances be-tween the reference lakes and both Whitepine and SansChambre lakes included higher abundances ofD. longiremis,S. oregonensis, Diaphanosoma sp., Neobosmina tubicen,D. retrocurva, D. dubia, and D. mendotae, and lower abun-dances ofD. pulex andC. scutifer, in the reference lakes (one-way analysis of variance (ANOVA), Bonferroni multiple com-parisons test).
Species richnessThere were substantial between-year variations in average
zooplankton community richness (Fig. 3); however, in bothSudbury lakes average species richness per collection in-creased (p < 0.05) over the study period based on regres-sions of richness against year (Whitepine,r = 0.55; SansChambre,r = 0.78). Average richness was also correlated(p < 0.05) with average annual pH (Whitepine,r = 0.42;Sans Chambre,r = 0.70), which was correlated (p < 0.05)with year (Whitepine,r = 0.83; Sans Chambre,r = 0.70).
Comparison of the long-term patterns in the average rich-ness of the Sudbury lakes with the average richness in theDorset reference lakes with pH > 6.0 (Fig. 3) indicated thatin the 1990s, richness values for both Sudbury lakes fellwithin the range of reference lake variability (mean ± 2 stan-dard deviations = 7.8–12.3) in some years. By this metric,communities have shown recovery.
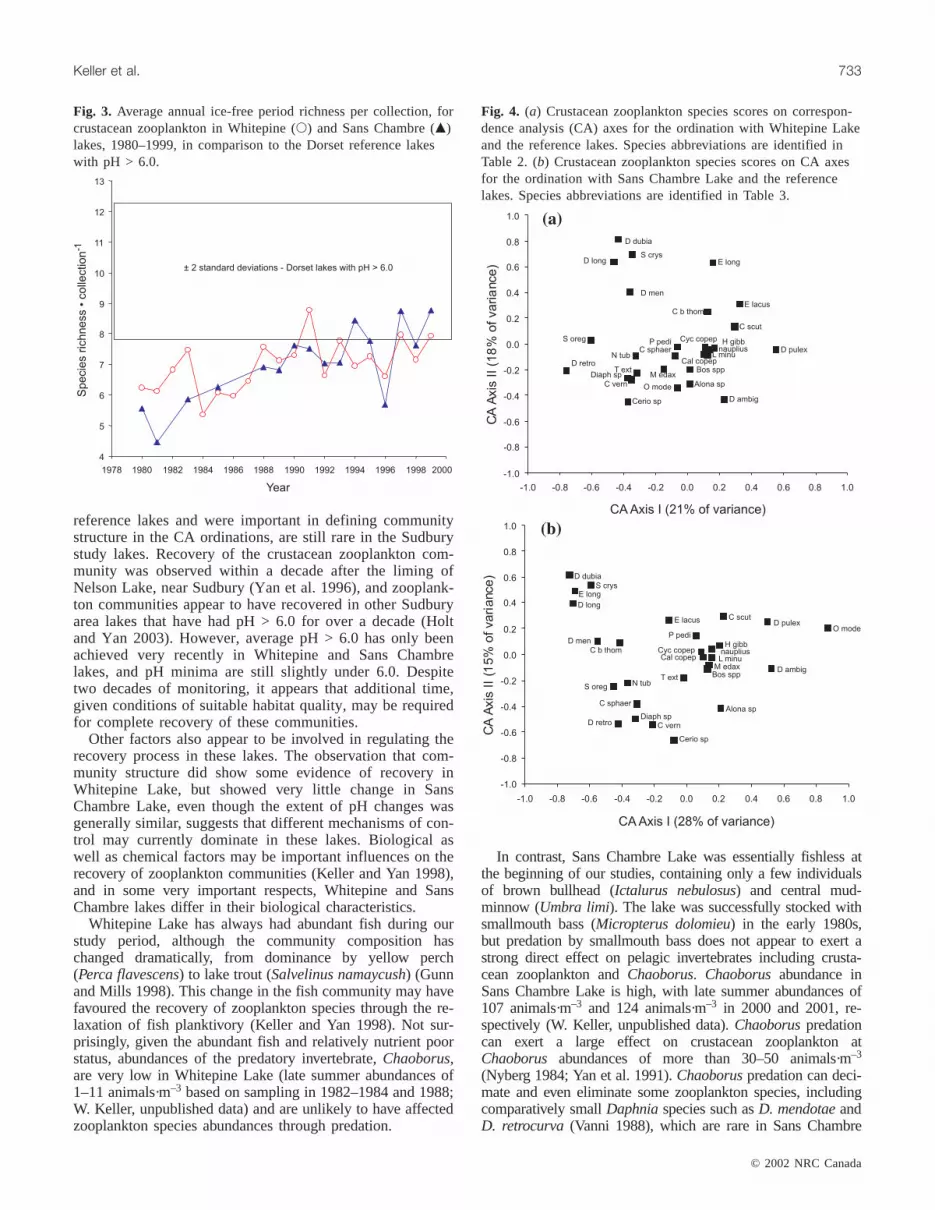
Multivariate community structureThe first two CA axes explained 39% of the transformed
variation in species abundances when Whitepine Lake wasincluded, and 43% of the transformed variation when SansChambre Lake was included (species scores for the axes areshown in Fig. 4a, 4b). In the Whitepine Lake CA, axis I(21% explained variation) was correlated (p < 0.05) withdissolved organic carbon (DOC;r = –0.50), total phosphorus(TP; r = –0.34), and calcium (Ca;r = –0.26), and axis II(18% explained variation) was correlated with pH (r = 0.60),aluminum (Al; r = –0.58), TP (r = –0.50), DOC (r = –0.38),and Ca (r = 0.26). In the ordination with Sans ChambreLake, axis I (28% explained variation) was correlated (p <0.05) with TP (r = 0.46), pH (r = –0.41), and Al (r = 0.36),and axis II (15% explained variation) was correlated withDOC (r = –0.63), Al (r = –0.43), pH (r = 0.42), TP (r = –0.36), and Ca (r = 0.30).
Temporally, Whitepine Lake shifted toward higher posi-tive scores on CA axis II during the study period. Axis IIscores increased significantly (r = 0.81, p < 0.05) over thestudy period based on a regression against sequential year ofsampling. This trajectory indicates recovery, since higherpositive scores on CA axis II are characteristic of the refer-ence lakes with pH > 6.0 (Fig. 5a). However, WhitepineLake scores never occupied the same region in ordinationspace on axis I as the reference lakes.
Community structure, as defined by lake scores from CA,changed little in Sans Chambre Lake over the past two de-cades (Fig. 5b). Throughout this study period Sans ChambreLake did not move in a consistent manner toward the regionin the ordination occupied by the reference lakes. There wereno significant correlations between axis I or axis II scores andsequential years based on linear regressions (p > 0.05).
Discussion
Two decades of monitoring, during conditions of decreas-ing lake acidity, did reveal evidence of recovery in the crus-tacean zooplankton communities in Whitepine and SansChambre lakes. A number of “new” species became estab-lished, and persisted in the zooplankton communities of thelakes. The fact that some of these species, such asE. lacustris, are quite acid sensitive (Keller and Yan 1998)indicates that these biological changes are consistent withimproved water quality conditions. Average species richness
Fig. 2. Average annual pH during the ice-free season (error barsindicate the range in values) for (a) Whitepine and (b) SansChambre lakes.

© 2002 NRC Canada
730 Can. J. Fish. Aquat. Sci. Vol. 59, 2002
per collection in both lakes increased substantially to levelsapproaching, or in some years falling within, the range ofvariability observed in the reference lakes with pH > 6.0.Whether species richness will increase further remains to beseen.
Despite the positive responses to reduced acidity that haveoccurred, zooplankton communities in the two Sudbury lakeshave not truly “recovered” when recovery is defined as thelake positions with respect to our two CA axes. The trajec-tory of community change in Whitepine Lake, determinedby CA, clearly indicates recovery; however, in both White-pine and Sans Chambre lakes, community structure nevercompletely resembled reference lake conditions.
This interpretation of the multivariate patterns in speciescomposition presumes that the range of variability in the ref-erence lake data does provide suitable targets for recovery inour study lakes. The Dorset lakes provide the only compara-bly collected zooplankton reference data set that is availableto help evaluate the responses of our Sudbury lakes. The ref-erence lakes were, however, only sampled during the 1980s,whereas our Sudbury lake data also extend through the
1990s. Therefore, we cannot be completely certain that thesereference data are fully representative of the 1990s, a decadewith, for example, different weather patterns than the 1980s.Such uncertainty underscores the need for additional effortsto establish current biological reference data sets to aid inassessments of the status of aquatic ecosystems affected byacidification and other anthropogenic stressors.
The reference and study areas are nonetheless zoogeo-graphically similar, and lakes in both areas were sampled bycomparable protocols. The reference lakes encompass therange of physico-chemical conditions in the Sudbury lakes,although the Sudbury lakes do tend to lie at the clear, small,low ionic strength end of the distribution of these character-istics in the reference lakes. The major overall differencebetween the Sudbury lakes and the Dorset lakes is in thecomparative scarcity of a number of acid-sensitive zooplank-ton species in the Sudbury lakes, a difference that is attribut-able to the greater degree and duration of acidificationdamage around Sudbury. Collectively, this evidence indi-cates that the reference lakes are suitable for our compari-sons.
Species
Speciesabbreviation
1980
(7)
1981
(13)
1982
(13)
1983
(11)
1984
(5)
1985
(6)
1986
(6)
1987
(6)
1988
(5)Calanoid copepodid Cal copep 100 85 92 91 80 83 100 100 100Cyclopoid copepodid Cyc copep 100 100 100 100 100 100 100 100 100Cyclops bicuspidatus thomasi C b thom 29 31 85 91 60 83 100 83 60Cyclops scutifer C scut 71 77 85 82 60 67 50 67 20Daphnia pulex D pulex 57 85 85 91 60 83 100 67 80Holopedium gibberum H gibb 100 62 69 100 100 100 83 83 60Leptodiaptomus minutus L minu 100 100 100 91 80 100 100 100 100Nauplius nauplius 100 100 100 100 100 100 100 100 100Bosmina spp. Bos spp 86 77 85 100 100 67 33 83 100Epischura lacustris E lacus 0 0 15 27 20 50 83 67 60Bosmina (Eubosmina) longispina E long 0 0 0 0 0 0 0 17 100Mesocyclops edax M edax 57 54 15 64 0 0 0 0 40Daphnia ambigua D ambig 71 62 46 0 40 0 0 17 0Daphnia mendotae D men 14 8 0 9 0 0 0 0 0Tropocyclops extensus T ext 0 8 0 27 0 33 17 17 20Chydorus sphaericus C sphaer 29 8 54 82 0 17 0 0 0Diaphanosoma sp. Diaph sp 0 0 8 0 0 0 0 0 40Alona sp. Alona sp 0 0 0 0 0 0 0 17 20Bosmina (Neobosmina) tubicen N tub 0 0 0 0 0 0 0 17 20Leptodora kindtii L kind 0 23 15 9 0 0 0 17 0Alona affinis A affin 14 15 0 0 0 0 0 0 0Alona guttata A gutt 0 0 8 18 0 0 0 0 0Bosmina (Eubosmina) coregoni E core 0 0 0 0 0 0 0 0 0Daphnia longiremis D long 0 0 0 0 0 0 0 0 0Eucyclops speratus E sper 0 0 8 0 0 0 0 0 0Acantholeberis curvirostris A curv 0 0 0 0 0 0 17 0 0Acroperus harpae A harp 0 0 8 0 0 0 0 0 0Ceriodaphnia sp. Cerio sp 0 0 0 0 0 17 0 0 0Cyclops vernalis C vern 0 0 0 0 0 0 0 0 20Daphnia retrocurva D retro 0 0 8 0 0 0 0 0 0Eucyclops neomacruroides E neom 0 0 0 0 0 0 0 0 0Latona setifera L seti 0 0 0 0 0 0 0 0 0Sida crystallina S crys 0 0 0 9 0 0 0 0 0Skistodiaptomus oregonensis S oreg 0 0 0 0 0 0 17 0 0
Note: Taxa are listed in order of decreasing overall occurrence in years.Daphnia dubia (species abbreviation D dubia) present in Dorset Reference
Table 2. Percent occurrence of crustacean zooplankton in ice-free period samples from Whitepine Lake, 1980–1999.
© 2002 NRC Canada
Keller et al. 731
A number of factors may be involved in regulating biolog-ical recovery in these lakes. Both Whitepine and SansChambre lakes are headwaters, thus it might be expectedthat colonization opportunities from external sources couldbe limited. Invasion rates can be highly variable for differentzooplankton species (Jenkins and Buikema 1998) and vari-ability in factors such as invasion sequence and rate (Patrick1967; Robinson and Edgemon 1988) may account for thevariable temporal trajectories observed in lakes recoveringfrom acidification.
Most zooplankton colonization experiments and studies(e.g., Jenkins and Buikema 1998; Shurin 2000) have beencomparatively short term (one year or less), and small scalewith respect to surface area. They may not completely re-flect the ultimate role of dispersal in determining communitycomposition on the scale of decades in larger natural sys-tems. Extended monitoring studies, as reported here, offer animportant complement to experimental studies examiningfactors regulating aquatic communities, because they reveal theactual long-term, lake-scale patterns of community change.
Long-term monitoring studies also have the advantage that
species have had a long temporal window for colonization,whether the vector of colonization is transport from sourcesexternal to the lake (Maguire 1963; Schlichting and Sides1969) or from the sediment egg bank (De Stasio 1989;Hairston 1996). It is likely that most species in the speciespool will be detected, even if they are rare, when samplingextends through decades. Examination of the two-decade re-cord for Whitepine and Sans Chambre lakes revealed that anumber of acid-sensitive (Keller at al. 1990b; Havens et al.1993) species, includingD. mendotae, D. retrocurva, andS. oregonensis, which were important in defining our CAaxes, were present at times in both lakes, but never becamecommon. This suggests that dispersal was not ultimately a ma-jor limitation to their establishment in the lake communities.
The incomplete recovery, as indicated by multivariate anal-yses of the zooplankton community in Whitepine and SansChambre lakes, may reflect the limitations of time and un-suitable water quality. A pH of at least 6.0 appears to benecessary for the survival of many acid-sensitive zooplank-ton species (Keller at al. 1990b; Havens et al. 1993). A num-ber of these species, which were relatively common in the
1989
(6)
1990
(7)
1991
(6)
1992
(7)
1993
(6)
1994
(6)
1995
(3)
1996
(3)
1997
(5)
1998
(5)
1999
(6)
% of years
(20)100 100 100 100 100 100 100 100 100 100 100 100100 100 100 100 100 100 100 100 100 100 100 10067 71 67 71 67 67 67 67 60 80 67 10083 57 83 57 50 67 67 67 60 40 50 10083 71 100 100 100 100 100 67 100 100 100 10083 100 100 100 100 83 67 100 100 60 100 100
100 100 100 100 100 100 100 100 100 100 100 100100 100 100 100 100 100 100 100 100 100 100 10083 29 33 0 50 33 67 33 60 60 83 9517 43 100 86 100 50 33 67 60 80 50 9083 86 100 100 100 100 100 100 100 100 83 6567 0 33 14 33 17 0 0 60 40 67 650 57 33 0 17 0 0 33 40 0 17 550 14 0 14 17 17 33 33 0 0 0 450 14 0 0 0 0 0 0 40 0 50 450 29 0 0 17 0 33 0 0 0 0 40
17 57 83 14 17 0 0 0 0 0 33 400 0 0 0 0 17 0 0 20 40 0 25
33 14 17 0 0 0 0 0 0 0 0 250 0 33 0 0 0 0 0 0 0 0 250 0 0 0 0 0 0 0 0 0 0 100 0 0 0 0 0 0 0 0 0 0 100 0 0 0 17 17 0 0 0 0 0 100 0 0 0 0 33 33 0 0 0 0 100 0 0 0 0 0 0 0 0 20 0 100 0 0 0 0 0 0 0 0 0 0 50 0 0 0 0 0 0 0 0 0 0 50 0 0 0 0 0 0 0 0 0 0 50 0 0 0 0 0 0 0 0 0 0 50 0 0 0 0 0 0 0 0 0 0 50 0 0 0 0 0 33 0 0 0 0 50 14 0 0 0 0 0 0 0 0 0 50 0 0 0 0 0 0 0 0 0 0 50 0 0 0 0 0 0 0 0 0 0 5
Lakes, but not in either of the Sudbury study lakes. Number in parentheses under year represents the total number of samples (n).

©2
00
2N
RC
Ca
na
da
73
2C
an.
J.
Fis
h.
Aq
uat.
Sci.
Vo
l.5
9,
20
02
Species
Speciesabbreviation
1980
(4)
1981
(13)
1983
(6)
1985
(6)
1988
(6)
1989
(6)
1990
(6)
1991
(7)
1992
(9)
1993
(9)
1994
(6)
1995
(7)
1996
(4)
1997
(5)
1998
(5)
1999
(6)
% of Years
(16)Bosmina spp. Bos spp 100 85 83 100 50 83 67 71 89 89 100 43 25 100 80 83 100Calanoid copepodid Cal copep 100 100 100 100 100 100 100 100 100 100 100 100 100 100 100 100 100Cyclopoid copepodid Cyc copep 100 100 100 100 100 100 100 100 100 100 100 100 100 100 100 100 100Cyclops scutifer C scut 50 62 100 83 50 83 50 43 44 56 67 86 50 40 40 50 100Holopedium gibberum H gibb 100 100 83 100 83 83 100 100 100 1 00 83 1 00 100 1 00 100 100 100Leptodiaptomus minutus L minu 100 100 100 83 100 100 100 100 100 100 100 100 100 100 100 100 100Mesocyclops edax M edax 100 62 100 67 67 50 33 71 44 89 67 86 50 60 60 67 100Nauplius nauplius 100 100 100 100 100 100 100 100 100 100 100 100 100 100 100 100 100Orthocyclops modestus O mode 100 54 17 0 83 33 83 71 44 56 83 100 25 80 100 83 94Daphnia pulex D pulex 0 0 100 100 100 100 100 100 100 100 83 71 100 80 100 100 88Tropocyclops extensus T ext 0 0 50 33 17 33 50 86 33 22 50 71 50 80 40 83 88Daphnia ambigua D ambig 0 0 0 0 100 67 83 43 11 78 83 43 0 40 40 100 69Epischura lacustris E lacus 0 0 0 0 17 17 17 43 44 11 17 29 25 40 20 0 69Cyclops bicuspidatus
thomasiC b thom 0 8 0 17 17 0 33 29 11 11 17 0 0 0 0 17 56
Daphnia mendotae D men 0 0 0 0 0 0 0 14 11 0 17 14 0 20 0 33 38Skistodiaptomus
oregonensisS oreg 0 0 0 0 0 0 17 14 11 0 17 0 0 20 0 0 31
Aglaodiaptomus leptopus A lept 0 0 0 0 0 0 0 0 0 0 0 0 50 100 60 50 25Daphnia sp. Daph sp 0 0 0 0 17 17 17 0 0 22 0 0 0 0 0 0 25Alona sp. Alona sp 0 0 0 0 0 17 0 0 0 0 0 0 0 0 20 17 19Bosmina (Eubosmina)
longispinaE long 0 0 0 0 0 0 33 0 0 0 17 0 0 0 0 0 13
Chydorus globosus C glob 0 0 0 0 0 0 0 0 0 0 33 29 0 0 0 0 13Chydorus sphaericus C sphaer 0 0 0 0 0 0 0 0 11 0 17 0 0 0 0 0 13Daphnia retrocurva D retro 0 0 0 0 17 0 0 0 11 0 0 0 0 0 0 0 13Polyphemus pediculus P pedi 0 0 17 17 0 0 0 0 0 0 0 0 0 0 0 0 13Bosmina (Neobosmina)
tubicenN tub 0 0 0 0 0 0 0 0 0 0 0 0 0 20 0 0 6
Bosmina (Eubosmina)coregoni
E core 0 0 0 0 0 0 0 0 0 0 0 14 0 0 0 0 6
Sida crystallina S crys 0 8 0 0 0 0 0 0 0 0 0 0 0 0 0 0 6
Note: Taxa are listed in order of decreasing overall occurrence in years.Daphnia dubia (species abbreviation D dubia) present in Dorset Reference Lakes, but not in either of the Sudbury studylakes. Number in parentheses represents the total number of samples (n).
Table 3. Percent occurrence of crustacean zooplankton in ice-free period samples from Sans Chambre Lake, 1980–1999.
© 2002 NRC Canada
Keller et al. 733
reference lakes and were important in defining communitystructure in the CA ordinations, are still rare in the Sudburystudy lakes. Recovery of the crustacean zooplankton com-munity was observed within a decade after the liming ofNelson Lake, near Sudbury (Yan et al. 1996), and zooplank-ton communities appear to have recovered in other Sudburyarea lakes that have had pH > 6.0 for over a decade (Holtand Yan 2003). However, average pH > 6.0 has only beenachieved very recently in Whitepine and Sans Chambrelakes, and pH minima are still slightly under 6.0. Despitetwo decades of monitoring, it appears that additional time,given conditions of suitable habitat quality, may be requiredfor complete recovery of these communities.
Other factors also appear to be involved in regulating therecovery process in these lakes. The observation that com-munity structure did show some evidence of recovery inWhitepine Lake, but showed very little change in SansChambre Lake, even though the extent of pH changes wasgenerally similar, suggests that different mechanisms of con-trol may currently dominate in these lakes. Biological aswell as chemical factors may be important influences on therecovery of zooplankton communities (Keller and Yan 1998),and in some very important respects, Whitepine and SansChambre lakes differ in their biological characteristics.
Whitepine Lake has always had abundant fish during ourstudy period, although the community composition haschanged dramatically, from dominance by yellow perch(Perca flavescens) to lake trout (Salvelinus namaycush) (Gunnand Mills 1998). This change in the fish community may havefavoured the recovery of zooplankton species through the re-laxation of fish planktivory (Keller and Yan 1998). Not sur-prisingly, given the abundant fish and relatively nutrient poorstatus, abundances of the predatory invertebrate,Chaoborus,are very low in Whitepine Lake (late summer abundances of1–11 animals·m–3 based on sampling in 1982–1984 and 1988;W. Keller, unpublished data) and are unlikely to have affectedzooplankton species abundances through predation.
In contrast, Sans Chambre Lake was essentially fishless atthe beginning of our studies, containing only a few individualsof brown bullhead (Ictalurus nebulosus) and central mud-minnow (Umbra limi). The lake was successfully stocked withsmallmouth bass (Micropterus dolomieu) in the early 1980s,but predation by smallmouth bass does not appear to exert astrong direct effect on pelagic invertebrates including crusta-cean zooplankton andChaoborus. Chaoborus abundance inSans Chambre Lake is high, with late summer abundances of107 animals·m–3 and 124 animals·m–3 in 2000 and 2001, re-spectively (W. Keller, unpublished data).Chaoborus predationcan exert a large effect on crustacean zooplankton atChaoborus abundances of more than 30–50 animals·m–3
(Nyberg 1984; Yan et al. 1991).Chaoborus predation can deci-mate and even eliminate some zooplankton species, includingcomparatively smallDaphnia species such asD. mendotae andD. retrocurva (Vanni 1988), which are rare in Sans Chambre
Fig. 3. Average annual ice-free period richness per collection, forcrustacean zooplankton in Whitepine (s) and Sans Chambre (m)lakes, 1980–1999, in comparison to the Dorset reference lakeswith pH > 6.0.
Fig. 4. (a) Crustacean zooplankton species scores on correspon-dence analysis (CA) axes for the ordination with Whitepine Lakeand the reference lakes. Species abbreviations are identified inTable 2. (b) Crustacean zooplankton species scores on CA axesfor the ordination with Sans Chambre Lake and the referencelakes. Species abbreviations are identified in Table 3.

Lake. The current characteristics of the Sans Chambre Lakezooplankton community, including scarcity of smallDaphnia,absence ofDiaphanosoma birgei, dominance byD. pulex, andthe frequent occurrence ofA. leptopus and O. modestus, areconsistent with communities in fishless lakes exhibiting highChaoborus predation (Vanni 1988; Arnott and Vanni 1993;Keller and Conlon 1994).
Thus, biological resistance (Nyberg 1984; Keller and Yan1998) through intensive invertebrate predation is probablylimiting the recovery of zooplankton in Sans Chambre Lake.Chaoborus abundance varies widely in the reference lakes(Persaud and Yan 2001) but none of the near-neutral refer-ence lakes are fishless. Based on its size of 14.5 ha, SansChambre Lake was probably also not fishless prior to acidi-fication (Matuszek et al. 1990), and therefore our recoverytargets based on reference data would be valid for that lake.Substantial recovery of zooplankton communities in thislake may require not just suitable water quality, but biologi-cal intervention through stocking of planktivorous fish. It islikely that a reduction in invertebrate predation pressure will
be needed before the zooplankton community can respond toreduced acidity.
The recovery of damaged aquatic ecosystems after the re-moval of stress is a very complex, poorly understood process(Cairns 1990; Power 1999). In general, the factors affectingbiological recovery fall into two categories: (i) chemical,physical, and biological factors related to habitat quality, and(ii) factors related to the dispersal of organisms. Understand-ing the relative roles of processes operating within lakes ver-sus processes related to dispersal is essential to thedevelopment of our knowledge of aquatic ecosystem recov-ery, and is also fundamental to addressing broader ecologicalquestions about the factors regulating biodiversity in aquaticsystems. Whether these different processes are termed localversus regional (e.g., Lukaszewski et al. 1999; Shurin et al.2000) or internal versus external (e.g., Keller and Yan 1998),a key question is if community structure is primarily regu-lated by factors affecting the dispersal of species, or by fac-tors affecting the survival and persistence of species.
The re-establishment of some zooplankton species includ-ing hypolimnetic forms and the “glacial opportunists” is ex-pected to be limited by dispersal abilities (Schindler 1987;Keller and Yan 1998) and the regional species pool will havean overriding effect on local species assemblages. However,increasing experimental evidence is emerging that demon-strates the importance of “local” processes in determiningzooplankton community structure (Arnott and Vanni 1993;Lukaszewski et al. 1999; Shurin 2000). Different mecha-nisms appear to be the primary current controls on zooplank-ton community recovery in Whitepine (duration of suitablehabitat conditions) and Sans Chambre (biological resistance)lakes. However, both these examples provide empirical evi-dence for the importance of within-lake factors as controlson biological recovery. Our results suggest that the success-ful establishment of most common zooplankton species inthese lakes as they recover from acidification will mainly de-pend on biotic and abiotic interactions, not on factors affect-ing species dispersal.
Acknowledgements
This paper is a contribution from the Aquatic RestorationGroup of the Cooperative Freshwater Ecology Unit, a part-nership between Laurentian University, the Ontario Ministryof the Environment, the Ontario Ministry of Natural Re-sources, Inco Limited, Falconbridge Limited, and Environ-ment Canada. We thank Shelley Arnott and the Journalreferees for helpful comments on the manuscript, and PeggyGale and Jim Carbone for their many years of help collect-ing zooplankton samples.
References
Arnott, S.E., and Vanni, M.J. 1993. Zooplankton assemblages infishless bog lakes: influence of biotic and abiotic factors. Ecol-ogy, 74: 2361–2380.
Arnott, S.E., Magnuson, J.J., and Yan, N.D. 1998. Crustacean zoo-plankton species richness—single and multiple-year estimates.Can. J. Fish. Aquat. Sci.55: 1573–1582.
Cairns, J. 1990. Lack of a theoretical basis for predicting rate andpathways of recovery. Environ. Manag.14: 517–526.
© 2002 NRC Canada
734 Can. J. Fish. Aquat. Sci. Vol. 59, 2002
Fig. 5. (a) Lake scores on correspondence analysis (CA) axes forthe ordination with Whitepine Lake and the reference lakes.(b) Lake scores on CA axes for the ordination with SansChambre Lake and the reference lakes.

© 2002 NRC Canada
Keller et al. 735
Degerman, E., Henrikson, L., Hermann, J., and Nyberg, P. 1995.The effects of liming on aquatic fauna.In Liming of acidified sur-face waters.Edited by L. Henrikson and W.Y. Brodin. Springer-Verlag, Heidelberg. pp. 221–282.
De Stasio, B.T., Jr. 1989. The seed bank of a freshwater crustacean:copepodology for the plant ecologist. Ecology,70: 1377–1389.
Gunn, J.M., and Mills, K.H. 1998. The potential for restoration ofacid-damaged lake trout lakes. Restor. Ecol.6: 390–397.
Hairston, N.G., Jr. 1996. Zooplankton egg banks as biotic reservoirsin changing environments. Limnol. Oceanogr.41: 1087–1092.
Havens, K.E., Yan, N.D., and Keller, W. 1993. Lake acidification:effects on crustacean zooplankton populations. Environ. Sci.Technol.27: 1621–1624.
Holt, C.A., and Yan, N.D. 2003. Recovery of zooplankton commu-nities from acidification in Killarney Park, Ontario, 1971–2000:pH 6 as a recovery goal. Ambio. In press.
Jenkins, D.J., and Buikema, A.L., Jr. 1998. Do similar communi-ties develop in similar sites? A test with zooplankton structureand function. Ecol. Monogr.68: 421–443.
Keller, W., and Conlon, M. 1994. Crustacean zooplankton commu-nities and lake morphometry in Precambrian Shield lakes. Can.J. Fish. Aquat. Sci.51: 2424–2434.
Keller, W., and Yan, N.D. 1991. Recovery of crustacean zooplank-ton species richness in Sudbury area lakes following water qual-ity improvements. Can. J. Fish. Aquat. Sci.48: 1635–1644.
Keller, W., and Yan, N.D. 1998. Biological recovery from acidifi-cation: zooplankton communities as a model of patterns andprocesses. Restor. Ecol.6: 364–375.
Keller, W., Dodge, D.P., and Booth, G.M. 1990a. Experimentallake neutralization program: overview of neutralization studiesin Ontario. Can. J. Fish. Aquat. Sci.47: 410–411.
Keller, W., Yan, N.D., Holtze, K.E., and Pitblado, J.R. 1990b. In-ferred effects of lake acidification onDaphnia galeata mendotae.Environ. Sci. Technol.24: 1259–1261.
Keller, W., Gunn, J.M., and Yan, N.D. 1992a. Evidence of biologi-cal recovery in acid stressed lakes near Sudbury, Canada. Envi-ron. Pollut.78: 79–85.
Keller, W., Pitblado, J.R., and Carbone, J. 1992b. Chemical re-sponses of acidic lakes in the Sudbury, Ontario, area to reducedsmelter emissions, 1981–1989. Can. J. Fish. Aquat. Sci.49(Suppl. 1): 25–32.
Keller, W., Gunn, J., and Yan, N. 1999. Acid rain: perspectives onlake recovery. J. Aquat. Ecosyst. Stress Recovery,6: 207–216.
Lukaszewski, Y., Arnott, S.E., and Frost, T.M. 1999. Regional ver-sus local processes in determining zooplankton community com-position of Little Rock Lake, Wisconsin, U.S.A. J. PlanktonRes.21: 991–1003.
Maguire, B., Jr. 1963. The passive dispersal of small aquatic organ-isms and their colonization of isolated bodies of water. Ecol.Monogr. 33: 61–185.
Matuszek, J.E., Goodier, J., and Wales, D.L. 1990. The occurrenceof cyprinidae and other small fish species in relation to pH ofOntario lakes. Trans. Am. Fish. Soc.119: 850–861.
Nyberg, P. 1984. Impact ofChaoborus predation on planktoniccrustacean communities in some acidified and limed forest lakesin Sweden. Rep. Inst. Freshw. Res. Drottningholm,61: 154–166.
Patrick, R. 1967. The effect of invasion rate, species pool, and sizeof area on the structure of the diatom community. Proc. Natl.Acad. Sci. U.S.A.,58: 1335–1342.
Persaud, A.D., and Yan, N.D. 2001. Accounting for spatial vari-ability in the design of sampling programmes forChaoborus lar-vae. J. Plankton Res.23: 279–285.
Power, M. 1999. Recovery in aquatic ecosytems: an overview ofknowledge and needs. J. Aquat. Ecosyst. Stress Recovery,6:253–257.
Robinson, J.V., and Edgemon, M.A. 1988. An experimental evalua-tion of the effect of invasion history on community structure.Ecology,69: 1410–1417.
Sampson, C.J., Brezonik, P.L., Frost, T.M., Webster, K.E., andSimonson, T.D. 1995. Experimental acidification of Little RockLake, Wisconsin: the first four years of chemical and biologicalrecovery. Water Air Soil Pollut.85: 1713–1719.
Schindler, D.W. 1987. Detecting ecosystem responses to anthropo-genic stresses. Can. J. Fish. Aquat. Sci.44(Suppl. 1): 6–25.
Schindler, D.W. 1988. Effects of acid rain on freshwater ecosys-tems. Science (Washington, D.C.),239: 149–157.
Schlichting, H.E., Jr., and Sides, S.L. 1969. The passive transportof aquatic microoorganisms by selected Hemiptera. J. Ecol.57:759–764.
Shurin, J.B. 2000. Dispersal limitation, invasion resistance andthe structure of pond zooplankton communities. Ecology,81:3062–3073.
Shurin, J.B., Liebold, M.A., Havel, J.E., and Pinel-Alloul, B. 2000.Local and regional species richness: a scale-independent test forsaturation. Ecology,81: 3074–3086.
Sprules, W.G. 1977. Crustacean zooplankton communities as indi-cators of limnological conditions: an approach using principalcomponent analysis. J. Fish. Res. Board Can.34: 389–395.
Stoddard, J.L., Jeffries, D.S., Lukewille, A., Clair, T.A., Dillon,P.J., Driscoll, C.T., Forsius, M., Johannessen, M., Kahl, J.S.,Kellogg, J.H., Kemp, A., Mannio, J., Monteith, D.T., Murdoch,P.S., Patrick, S., Rebsdorf, A., Skjelkvale, B.L., Stainton, M.P.,Traen, T., van Dam, H., Webster, K.E., and Wilander, A. 1999.Regional trends in aquatic recovery from acidification in NorthAmerica and Europe. Nature (London),401: 575–578.
Vanni, M.J. 1988. Freshwater zooplankton community structure:introduction of large invertebrate predators and large herbivoresto a small species community. Can. J. Fish. Aquat. Sci.45:1758–1770.
Yan, N.D., Keller, W., MacIsaac, H.J., and McEachern, L.J. 1991.Regulation of zooplankton community structure in an acidiclake by Chaoborus. Ecol. Appl. 1: 52–65.
Yan, N.D., Keller, W., Somers, K.M., Pawson, T.W., and Girard,R.G. 1996. Recovery of crustacean zooplankton communitiesfrom acid and metal contamination: comparing manipulated andreference lakes. Can. J. Fish. Aquat. Sci.53: 1301–1327.