linkage between crustacean zooplankton and aquatic bacteria
TRANSCRIPT

AQUATIC MICROBIAL ECOLOGYAquat Microb Ecol
Vol. 61: 261–277, 2010doi: 10.3354/ame01424
Published online August 18
INTRODUCTION
Bacteria and metazoan zooplankton are importantcomponents of the pelagic food web and major contrib-utors to pelagic biodiversity and biogeochemical pro-cesses. Although both inhabit the same environment,they are often treated as separate functional units onlyindirectly connected via nutrient cycling and trophiccascades (Azam & Malfatti 2007). The seeming lack ofphysical structures in the water column gives theimpression that pelagic bacteria are living in a ratherhomogeneous environment, and respond only to pre-
dation, viral attack, and physical–chemical changes inthe surrounding water. This ‘free-living bacteria’ pointof view still dominates the primary literature and textbooks, and microbial ecologists often ignore zooplank-ton and other higher organisms as potential habitatsfor aquatic bacteria except for a few pathogenic spe-cies. Likewise, zooplankton ecologists tend to focusonly on interactions between zooplankton and theirprey or predators, and because bacteria do not usuallyfall within either category, they are often overlookedby zooplankton ecologists. Consequently, microbialecology and zooplankton ecology are taught and prac-
© Inter-Research 2010 · www.int-res.com*Corresponding author. Email: [email protected]
REVIEW
Linkage between crustacean zooplankton andaquatic bacteria
Kam W. Tang1, Valentina Turk2, Hans-Peter Grossart3,*
1Virginia Institute of Marine Science, College of William & Mary, Gloucester Point, Virginia 23062, USA2National Institute of Biology, Marine Biology Station, 6330 Piran, Slovenia
3Leibniz-Institute of Freshwater Ecology and Inland Fisheries, Dept. Limnology of Stratified Lakes, Alte Fischerhuette 2,16775 Stechlin, Germany
ABSTRACT: Bacteria and metazoan zooplankton (mainly crustaceans) are often viewed as 2 separatefunctional groups in the pelagic food webs indirectly linked via nutrient cycling and trophic cascades.Yet a zooplankter’s body carries a high abundance of diverse bacteria, often at an equivalent concen-tration orders of magnitude higher than the ambient bacterial concentration. Zooplankton bodies areorganic-rich micro-environments that support fast bacterial growth. Their physical-chemical con-ditions differ from those in the surrounding water and therefore select for different bacterial commu-nities, including anaerobic bacteria that otherwise may not thrive in a well-oxygenated water col-umn. The zooplankton body provides protection to the associated bacteria from environmentalstresses similar to biofilms. Furthermore, migration by zooplankton enables rapid dispersal of bacte-ria over vast distances and across boundaries such as the pycnocline. In addition to live zooplankton,molts, fecal pellets, and carcasses of zooplankton all influence water column and benthic microbialcommunities in various ways. We review the recent advances in the study of (crustacean) zooplank-ton–bacteria interactions and discuss future research opportunities and challenges. Traditionalaquatic microbial ecology emphasizes free-living bacteria, which represent only a fraction of themicrobial world. By transcending disciplinary boundaries, microbial ecologists and zooplanktonecologists can work together to integrate the two disciplines and advance our understanding inaquatic microbial ecology.
KEY WORDS: Zooplankton . Bacteria . Zooplankton–bacteria interactions . Food web . Bacterialdispersal . Hotspots
Resale or republication not permitted without written consent of the publisher
Contribution to AME Special 4 ‘Progress and perspectives in aquatic microbial ecology’ OPENPEN ACCESSCCESS
Aquat Microb Ecol 61: 261–277, 2010
ticed as separate disciplines, and collaboration be-tween the two is very limited.
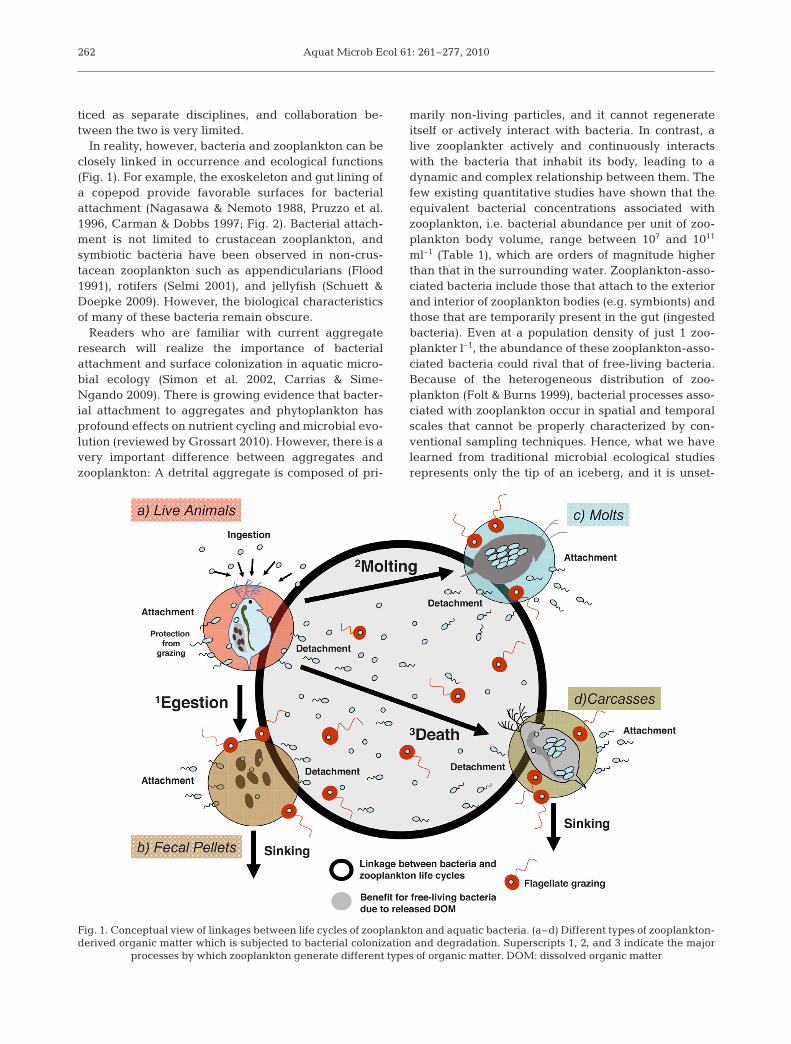
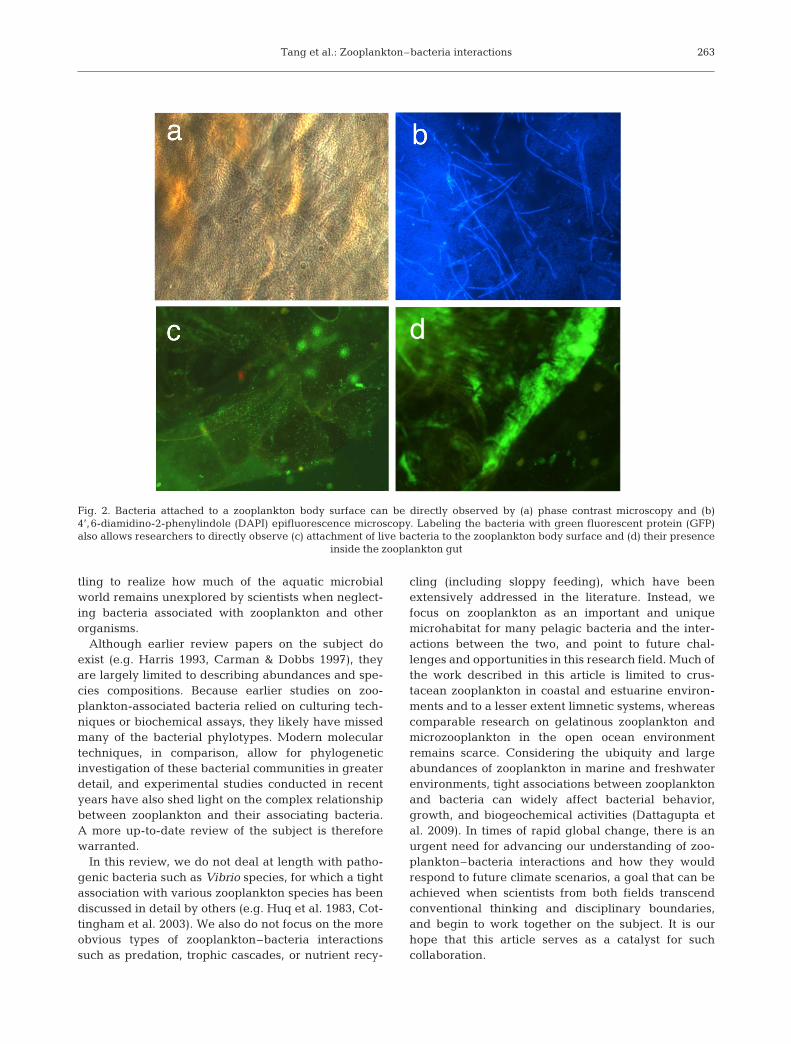
In reality, however, bacteria and zooplankton can beclosely linked in occurrence and ecological functions(Fig. 1). For example, the exoskeleton and gut lining ofa copepod provide favorable surfaces for bacterialattachment (Nagasawa & Nemoto 1988, Pruzzo et al.1996, Carman & Dobbs 1997; Fig. 2). Bacterial attach-ment is not limited to crustacean zooplankton, andsymbiotic bacteria have been observed in non-crus-tacean zooplankton such as appendicularians (Flood1991), rotifers (Selmi 2001), and jellyfish (Schuett &Doepke 2009). However, the biological characteristicsof many of these bacteria remain obscure.
Readers who are familiar with current aggregateresearch will realize the importance of bacterialattachment and surface colonization in aquatic micro-bial ecology (Simon et al. 2002, Carrias & Sime-Ngando 2009). There is growing evidence that bacter-ial attachment to aggregates and phytoplankton hasprofound effects on nutrient cycling and microbial evo-lution (reviewed by Grossart 2010). However, there is avery important difference between aggregates andzooplankton: A detrital aggregate is composed of pri-
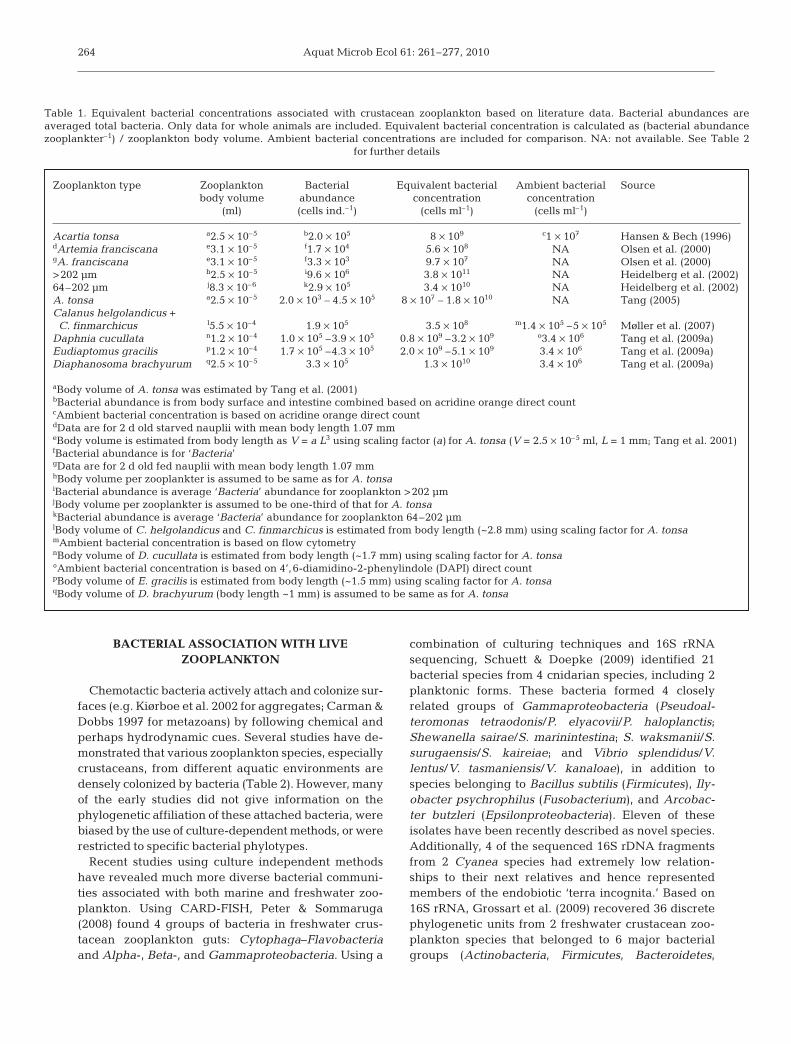
marily non-living particles, and it cannot regenerateitself or actively interact with bacteria. In contrast, alive zooplankter actively and continuously interactswith the bacteria that inhabit its body, leading to adynamic and complex relationship between them. Thefew existing quantitative studies have shown that theequivalent bacterial concentrations associated withzooplankton, i.e. bacterial abundance per unit of zoo-plankton body volume, range between 107 and 1011
ml–1 (Table 1), which are orders of magnitude higherthan that in the surrounding water. Zooplankton-asso-ciated bacteria include those that attach to the exteriorand interior of zooplankton bodies (e.g. symbionts) andthose that are temporarily present in the gut (ingestedbacteria). Even at a population density of just 1 zoo-plankter l–1, the abundance of these zooplankton-asso-ciated bacteria could rival that of free-living bacteria.Because of the heterogeneous distribution of zoo-plankton (Folt & Burns 1999), bacterial processes asso-ciated with zooplankton occur in spatial and temporalscales that cannot be properly characterized by con-ventional sampling techniques. Hence, what we havelearned from traditional microbial ecological studiesrepresents only the tip of an iceberg, and it is unset-
262
Fig. 1. Conceptual view of linkages between life cycles of zooplankton and aquatic bacteria. (a–d) Different types of zooplankton-derived organic matter which is subjected to bacterial colonization and degradation. Superscripts 1, 2, and 3 indicate the major
processes by which zooplankton generate different types of organic matter. DOM: dissolved organic matter
Tang et al.: Zooplankton–bacteria interactions
tling to realize how much of the aquatic microbialworld remains unexplored by scientists when neglect-ing bacteria associated with zooplankton and otherorganisms.
Although earlier review papers on the subject doexist (e.g. Harris 1993, Carman & Dobbs 1997), theyare largely limited to describing abundances and spe-cies compositions. Because earlier studies on zoo-plankton-associated bacteria relied on culturing tech-niques or biochemical assays, they likely have missedmany of the bacterial phylotypes. Modern moleculartechniques, in comparison, allow for phylogeneticinvestigation of these bacterial communities in greaterdetail, and experimental studies conducted in recentyears have also shed light on the complex relationshipbetween zooplankton and their associating bacteria.A more up-to-date review of the subject is thereforewarranted.
In this review, we do not deal at length with patho-genic bacteria such as Vibrio species, for which a tightassociation with various zooplankton species has beendiscussed in detail by others (e.g. Huq et al. 1983, Cot-tingham et al. 2003). We also do not focus on the moreobvious types of zooplankton–bacteria interactionssuch as predation, trophic cascades, or nutrient recy-
cling (including sloppy feeding), which have beenextensively addressed in the literature. Instead, wefocus on zooplankton as an important and uniquemicrohabitat for many pelagic bacteria and the inter-actions between the two, and point to future chal-lenges and opportunities in this research field. Much ofthe work described in this article is limited to crus-tacean zooplankton in coastal and estuarine environ-ments and to a lesser extent limnetic systems, whereascomparable research on gelatinous zooplankton andmicrozooplankton in the open ocean environmentremains scarce. Considering the ubiquity and largeabundances of zooplankton in marine and freshwaterenvironments, tight associations between zooplanktonand bacteria can widely affect bacterial behavior,growth, and biogeochemical activities (Dattagupta etal. 2009). In times of rapid global change, there is anurgent need for advancing our understanding of zoo-plankton–bacteria interactions and how they wouldrespond to future climate scenarios, a goal that can beachieved when scientists from both fields transcendconventional thinking and disciplinary boundaries,and begin to work together on the subject. It is ourhope that this article serves as a catalyst for suchcollaboration.
263
Fig. 2. Bacteria attached to a zooplankton body surface can be directly observed by (a) phase contrast microscopy and (b) 4’,6-diamidino-2-phenylindole (DAPI) epifluorescence microscopy. Labeling the bacteria with green fluorescent protein (GFP)also allows researchers to directly observe (c) attachment of live bacteria to the zooplankton body surface and (d) their presence
inside the zooplankton gut
Aquat Microb Ecol 61: 261–277, 2010
BACTERIAL ASSOCIATION WITH LIVEZOOPLANKTON
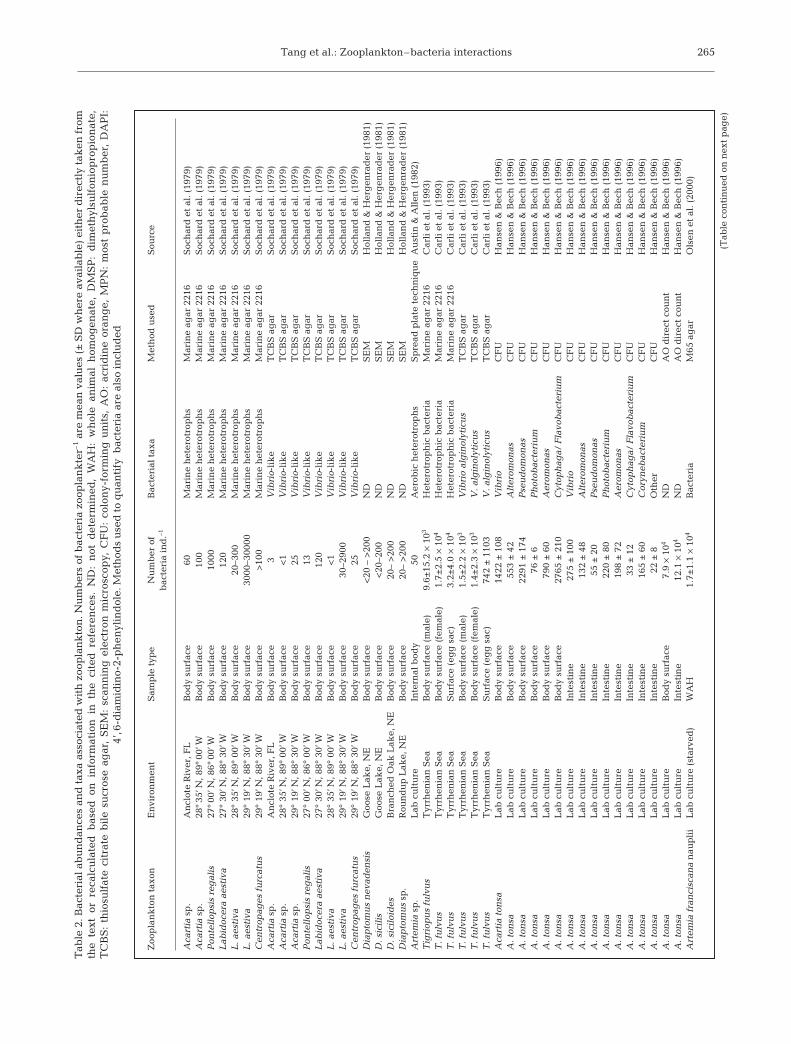
Chemotactic bacteria actively attach and colonize sur-faces (e.g. Kiørboe et al. 2002 for aggregates; Carman &Dobbs 1997 for metazoans) by following chemical andperhaps hydrodynamic cues. Several studies have de-monstrated that various zooplankton species, especiallycrustaceans, from different aquatic environments aredensely colonized by bacteria (Table 2). However, manyof the early studies did not give information on thephylogenetic affiliation of these attached bacteria, werebiased by the use of culture-dependent methods, or wererestricted to specific bacterial phylotypes.
Recent studies using culture independent methodshave revealed much more diverse bacterial communi-ties associated with both marine and freshwater zoo-plankton. Using CARD-FISH, Peter & Sommaruga(2008) found 4 groups of bacteria in freshwater crus-tacean zooplankton guts: Cytophaga–Flavobacteriaand Alpha-, Beta-, and Gammaproteobacteria. Using a
combination of culturing techniques and 16S rRNAsequencing, Schuett & Doepke (2009) identified 21bacterial species from 4 cnidarian species, including 2planktonic forms. These bacteria formed 4 closelyrelated groups of Gammaproteobacteria (Pseudoal-teromonas tetraodonis/P. elyacovii/P. haloplanctis;Shewanella sairae/S. marinintestina; S. waksmanii/S.surugaensis/S. kaireiae; and Vibrio splendidus/V.lentus/V. tasmaniensis/V. kanaloae), in addition tospecies belonging to Bacillus subtilis (Firmicutes), Ily-obacter psychrophilus (Fusobacterium), and Arcobac-ter butzleri (Epsilonproteobacteria). Eleven of theseisolates have been recently described as novel species.Additionally, 4 of the sequenced 16S rDNA fragmentsfrom 2 Cyanea species had extremely low relation-ships to their next relatives and hence representedmembers of the endobiotic ‘terra incognita.’ Based on16S rRNA, Grossart et al. (2009) recovered 36 discretephylogenetic units from 2 freshwater crustacean zoo-plankton species that belonged to 6 major bacterialgroups (Actinobacteria, Firmicutes, Bacteroidetes,
264
Zooplankton type Zooplankton Bacterial Equivalent bacterial Ambient bacterial Sourcebody volume abundance concentration concentration
(ml) (cells ind.–1) (cells ml–1) (cells ml–1)
Acartia tonsa a2.5 × 10–5 b2.0 × 105 8 × 109 c1 × 107 Hansen & Bech (1996)dArtemia franciscana e3.1 × 10–5 f1.7 × 104 5.6 × 108 NA Olsen et al. (2000)gA. franciscana e3.1 × 10–5 f3.3 × 103 9.7 × 107 NA Olsen et al. (2000)>202 µm h2.5 × 10–5 i9.6 × 106 3.8 × 1011 NA Heidelberg et al. (2002)64–202 µm j8.3 × 10–6 k2.9 × 105 3.4 × 1010 NA Heidelberg et al. (2002)A. tonsa a2.5 × 10–5 2.0 × 103 – 4.5 × 105 8 × 107 – 1.8 × 1010 NA Tang (2005)Calanus helgolandicus +C. finmarchicus l5.5 × 10–4 1.9 × 105 3.5 × 108 m1.4 × 105 –5 × 105 Møller et al. (2007)
Daphnia cucullata n1.2 × 10–4 1.0 × 105 –3.9 × 105 0.8 × 109 –3.2 × 109 o3.4 × 106 Tang et al. (2009a)Eudiaptomus gracilis p1.2 × 10–4 1.7 × 105 –4.3 × 105 2.0 × 109 –5.1 × 109 3.4 × 106 Tang et al. (2009a)Diaphanosoma brachyurum q2.5 × 10–5 3.3 × 105 1.3 × 1010 3.4 × 106 Tang et al. (2009a)
aBody volume of A. tonsa was estimated by Tang et al. (2001)bBacterial abundance is from body surface and intestine combined based on acridine orange direct countcAmbient bacterial concentration is based on acridine orange direct countdData are for 2 d old starved nauplii with mean body length 1.07 mmeBody volume is estimated from body length as V = a L3 using scaling factor (a) for A. tonsa (V = 2.5 × 10–5 ml, L = 1 mm; Tang et al. 2001)fBacterial abundance is for ‘Bacteria’gData are for 2 d old fed nauplii with mean body length 1.07 mmhBody volume per zooplankter is assumed to be same as for A. tonsaiBacterial abundance is average ‘Bacteria’ abundance for zooplankton >202 µmjBody volume per zooplankter is assumed to be one-third of that for A. tonsakBacterial abundance is average ‘Bacteria’ abundance for zooplankton 64–202 µmlBody volume of C. helgolandicus and C. finmarchicus is estimated from body length (~2.8 mm) using scaling factor for A. tonsamAmbient bacterial concentration is based on flow cytometrynBody volume of D. cucullata is estimated from body length (~1.7 mm) using scaling factor for A. tonsa°Ambient bacterial concentration is based on 4’,6-diamidino-2-phenylindole (DAPI) direct countpBody volume of E. gracilis is estimated from body length (~1.5 mm) using scaling factor for A. tonsaqBody volume of D. brachyurum (body length ~1 mm) is assumed to be same as for A. tonsa
Table 1. Equivalent bacterial concentrations associated with crustacean zooplankton based on literature data. Bacterial abundances areaveraged total bacteria. Only data for whole animals are included. Equivalent bacterial concentration is calculated as (bacterial abundancezooplankter–1) / zooplankton body volume. Ambient bacterial concentrations are included for comparison. NA: not available. See Table 2
for further details
Tang et al.: Zooplankton–bacteria interactions 265
Zoo
pla
nk
ton
tax
onE
nvi
ron
men
tS
amp
le t
ype
Nu
mb
er o
f B
acte
rial
tax
aM
eth
od u
sed
Sou
rce
bac
teri
a in
d.–1
Aca
rtia
sp
.A
ncl
ote
Riv
er, F
LB
ody
surf
ace
60M
arin
e h
eter
otro
ph
sM
arin
e ag
ar 2
216
Soc
har
d e
t al
. (19
79)
Aca
rtia
sp
.28
°35’
N, 8
9°00
’WB
ody
surf
ace
100
Mar
ine
het
erot
rop
hs
Mar
ine
agar
221
6S
och
ard
et
al. (
1979
)P
onte
llop
sis
reg
alis
27°0
0’N
, 86°
00’W
Bod
y su
rfac
e10
00M
arin
e h
eter
otro
ph
sM
arin
e ag
ar 2
216
Soc
har
d e
t al
. (19
79)
Lab
idoc
era
aest
iva
27°
30’N
, 88°
30’W
Bod
y su
rfac
e12
0M
arin
e h
eter
otro
ph
sM
arin
e ag
ar 2
216
Soc
har
d e
t al
. (19
79)
L. a
esti
va28
°35
’N, 8
9°00
’WB
ody
surf
ace
20–3
00M
arin
e h
eter
otro
ph
sM
arin
e ag
ar 2
216
Soc
har
d e
t al
. (19
79)
L. a
esti
va29
°19
’N, 8
8°30
’WB
ody
surf
ace
3000
–300
00M
arin
e h
eter
otro
ph
sM
arin
e ag
ar 2
216
Soc
har
d e
t al
. (19
79)
Cen
trop
ages
fu
rcat
us
29°
19’N
, 88°
30’W
Bod
y su
rfac
e>
100
Mar
ine
het
erot
rop
hs
Mar
ine
agar
221
6S
och
ard
et
al. (
1979
)A
cart
ia s
p.
An
clot
e R
iver
, FL
Bod
y su
rfac
e3
Vib
rio-
lik
eT
CB
S a
gar
Soc
har
d e
t al
. (19
79)
Aca
rtia
sp
.28
°35
’N, 8
9°00
’WB
ody
surf
ace
<1
Vib
rio-
lik
eT
CB
S a
gar
Soc
har
d e
t al
. (19
79)
Aca
rtia
sp
.29
°19
’N, 8
8°30
’WB
ody
surf
ace
25V
ibri
o-li
ke
TC
BS
ag
arS
och
ard
et
al. (
1979
)P
onte
llop
sis
reg
alis
27°
00’N
, 86°
00’W
Bod
y su
rfac
e13
Vib
rio-
lik
eT
CB
S a
gar
Soc
har
d e
t al
. (19
79)
Lab
idoc
era
aest
iva
27°
30’N
, 88°
30’W
Bod
y su
rfac
e12
0V
ibri
o-li
ke
TC
BS
ag
arS
och
ard
et
al. (
1979
)L
. aes
tiva
28°
35’N
, 89°
00’W
Bod
y su
rfac
e<
1V
ibri
o-li
ke
TC
BS
ag
arS
och
ard
et
al. (
1979
)L
. aes
tiva
29°
19’N
, 88°
30’W
Bod
y su
rfac
e30
–290
0V
ibri
o-li
ke
TC
BS
ag
arS
och
ard
et
al. (
1979
)C
entr
opag
es f
urc
atu
s29
°19
’N, 8
8°30
’WB
ody
surf
ace
25V
ibri
o-li
ke
TC
BS
ag
arS
och
ard
et
al. (
1979
)D
iap
tom
us
nev
aden
sis
Goo
se L
ake,
NE
Bod
y su
rfac
e<
20 –
>20
0N
DS
EM
Hol
lan
d &
Her
gen
rad
er (
1981
)D
. sic
ilis
Goo
se L
ake,
NE
Bod
y su
rfac
e<
20–2
00N
DS
EM
Hol
lan
d &
Her
gen
rad
er (
1981
)D
. sic
iloi
des
Bra
nch
ed O
ak L
ake,
NE
Bod
y su
rfac
e20
– >
200
ND
SE
MH
olla
nd
& H
erg
enra
der
(19
81)
Dia
pto
mu
s sp
.R
oun
du
p L
ake,
NE
Bod
y su
rfac
e20
– >
200
ND
SE
MH
olla
nd
& H
erg
enra
der
(19
81)
Art
emia
sp
.L
ab c
ult
ure
Inte
rnal
bod
y50
Aer
obic
het
erot
rop
hs
Sp
read
pla
te t
ech
niq
ue
Au
stin
& A
llen
(19
82)
Tig
riop
us
fulv
us
Tyr
rhen
ian
Sea
Bod
y su
rfac
e (m
ale)
9.6±
15.2
×10
3H
eter
otro
ph
ic b
acte
ria
Mar
ine
agar
221
6C
arli
et
al. (
1993
)T
. fu
lvu
sT
yrrh
enia
n S
eaB
ody
surf
ace
(fem
ale)
1.7±
2.5
×10
4H
eter
otro
ph
ic b
acte
ria
Mar
ine
agar
221
6C
arli
et
al. (
1993
)T
. fu
lvu
sT
yrrh
enia
n S
eaS
urf
ace
(eg
g s
ac)
3.2±
4.0
×10
4H
eter
otro
ph
ic b
acte
ria
Mar
ine
agar
221
6C
arli
et
al. (
1993
)T
. fu
lvu
sT
yrrh
enia
n S
eaB
ody
surf
ace
(mal
e)1.
5±2.
2 ×
103
Vib
rio
alg
inol
ytic
us
TC
BS
ag
arC
arli
et
al. (
1993
)T
. fu
lvu
sT
yrrh
enia
n S
eaB
ody
surf
ace
(fem
ale)
1.4±
2.3
×10
3V
. alg
inol
ytic
us
TC
BS
ag
arC
arli
et
al. (
1993
)T
. fu
lvu
sT
yrrh
enia
n S
eaS
urf
ace
(eg
g s
ac)
742
±11
03V
. alg
inol
ytic
us
TC
BS
ag
arC
arli
et
al. (
1993
)A
cart
ia t
onsa
Lab
cu
ltu
reB
ody
surf
ace
1422
±10
8V
ibri
oC
FU
Han
sen
& B
ech
(19
96)
A. t
onsa
Lab
cu
ltu
reB
ody
surf
ace
553
±42
Alt
erom
onas
CF
UH
anse
n &
Bec
h (
1996
)A
. ton
saL
ab c
ult
ure
Bod
y su
rfac
e22
91 ±
174
Pse
ud
omon
asC
FU
Han
sen
& B
ech
(19
96)
A. t
onsa
Lab
cu
ltu
reB
ody
surf
ace
76 ±
6P
hot
obac
teri
um
CF
UH
anse
n &
Bec
h (
1996
)A
. ton
saL
ab c
ult
ure
Bod
y su
rfac
e79
0 ±
60A
erom
onas
CF
UH
anse
n &
Bec
h (
1996
)A
. ton
saL
ab c
ult
ure
Bod
y su
rfac
e27
65 ±
210
Cyt
oph
aga/
Fla
vob
acte
riu
mC
FU
Han
sen
& B
ech
(19
96)
A. t
onsa
Lab
cu
ltu
reIn
test
ine
275
±10
0V
ibri
oC
FU
Han
sen
& B
ech
(19
96)
A. t
onsa
Lab
cu
ltu
reIn
test
ine
132
±48
Alt
erom
onas
CF
UH
anse
n &
Bec
h (
1996
)A
. ton
saL
ab c
ult
ure
Inte
stin
e55
±20
Pse
ud
omon
asC
FU
Han
sen
& B
ech
(19
96)
A. t
onsa
Lab
cu
ltu
reIn
test
ine
220
±80
Ph
otob
acte
riu
mC
FU
Han
sen
& B
ech
(19
96)
A. t
onsa
Lab
cu
ltu
reIn
test
ine
198
±72
Aer
omon
asC
FU
Han
sen
& B
ech
(19
96)
A. t
onsa
Lab
cu
ltu
reIn
test
ine
33 ±
12C
ytop
hag
a/ F
lavo
bac
teri
um
CF
UH
anse
n &
Bec
h (
1996
)A
. ton
saL
ab c
ult
ure
Inte
stin
e16
5 ±
60C
oryn
ebac
teri
um
CF
UH
anse
n &
Bec
h (
1996
)A
. ton
saL
ab c
ult
ure
Inte
stin
e22
±8
Oth
erC
FU
Han
sen
& B
ech
(19
96)
A. t
onsa
Lab
cu
ltu
reB
ody
surf
ace
7.9
×10
4N
DA
O d
irec
t co
un
tH
anse
n &
Bec
h (
1996
)A
. ton
saL
ab c
ult
ure
Inte
stin
e12
.1 ×
104
ND
AO
dir
ect
cou
nt
Han
sen
& B
ech
(19
96)
Art
emia
fra
nci
scan
an
aup
lii
Lab
cu
ltu
re (
star
ved
)W
AH
1.7±
1.1
×10
4B
acte
ria
M65
ag
arO
lsen
et
al. (
2000
)
Tab
le 2
. Bac
teri
al a
bu
nd
ance
s an
d t
axa
asso
ciat
ed w
ith
zoo
pla
nk
ton
. Nu
mb
ers
of b
acte
ria
zoop
lan
kte
r–1
are
mea
n v
alu
es (
±S
D w
her
e av
aila
ble
) ei
ther
dir
ectl
y ta
ken
fro
mth
e te
xt o
r re
calc
ula
ted
bas
ed o
n i
nfo
rmat
ion
in
th
e ci
ted
ref
eren
ces.
ND
: n
ot d
eter
min
ed,
WA
H:
wh
ole
anim
al h
omog
enat
e, D
MS
P:
dim
eth
ylsu
lfon
iop
rop
ion
ate,
TC
BS
:th
iosu
lfat
e ci
trat
e b
ile
sucr
ose
agar
, S
EM
: sc
ann
ing
ele
ctro
n m
icro
scop
y, C
FU
: co
lon
y-fo
rmin
g u
nit
s, A
O:
acri
din
e or
ang
e, M
PN
: m
ost
pro
bab
le n
um
ber
, D
AP
I:4’
,6-d
iam
idin
o-2-
ph
enyl
ind
ole.
Met
hod
s u
sed
to
qu
anti
fy b
acte
ria
are
also
incl
ud
ed
(Tab
le c
onti
nu
ed o
n n
ext
pag
e)

Aquat Microb Ecol 61: 261–277, 2010
Alpha-, Beta-, and Gammaproteobacte-ria). Using the same technique, Tang etal. (2009b) identified 37 discrete phylo-genetic units from the marine copepodAcartia tonsa that belonged to 3 majorbacteria groups (Alpha- and Gammapro-teobacteria, Bacteroidetes). Surprisingly,Møller et al. (2007) found only 3 differentRoseobacter species on Calanus spp. inthe North Sea using similar methods.These findings indicate that Alphapro-teobacteria are an important componentof the zooplankton-associated bacterialcommunity, which can vary greatly inrichness and phylotype composition amongzooplankton species, sampling location,and time.
Zooplankton-associated bacteria mustalso exist in a free-living stage if they areto maintain a viable population beyondthe lifetime of the host. For many symbi-otic bacteria such as Vibrio fischeri on thesquid Euprymna scolopes (Koropatnick etal. 2004), it has been shown that the bac-terial symbiont is frequently released intothe surrounding water to allow for colo-nization of other hosts. Earlier studieshave shown that zooplankton bodies andthe surrounding water share similar bac-terial groups but in different proportions(Sochard et al. 1979, Delille & Razouls1994, Hansen & Bech 1996), suggestingan active exchange of bacteria betweenthe 2 compartments, but that their differ-ent physical-chemical conditions tend tofavor different bacterial groups.
Separation of epibionts and gut micro-flora is difficult in practice, but the latterare expected to be more directly affectedby the host’s feeding activity. Harris(1993) proposed a conceptual model thatdifferentiates between transient and res-ident gut microflora associated withinvertebrates: Resident bacteria are per-sistently present in the gut, whereas tran-sient bacteria do not form stable popula-tions within the gut. Empirical evidencein support of this view is provided by 2recent studies. When the copepod Acar-tia tonsa was feeding on different axenicphytoplankton diets, the bacterial diver-sity associated with the copepod de-creased and converged, indicating thepresence of a rather stable resident bac-terial community (Tang et al. 2009b). In
266
Zoo
pla
nk
ton
tax
onE
nvi
ron
men
tS
amp
le t
ype
Nu
mb
er o
f B
acte
rial
tax
aM
eth
od u
sed
Sou
rce
bac
teri
a in
d.–1
A. f
ran
cisc
ana
nau
pli
iL
ab c
ult
ure
(fe
d)
WA
H3.
3±3.
7 ×
103
Bac
teri
aM
65 a
gar
Ols
en e
t al
. (20
00)
A. f
ran
cisc
ana
nau
pli
iL
ab c
ult
ure
(st
arve
d)
WA
H25
50 ±
71H
emol
ytic
bac
teri
aB
lood
ag
arO
lsen
et
al. (
2000
)A
.fra
nci
scan
an
aup
lii
Lab
cu
ltu
re (
fed
)W
AH
960
±11
17H
emol
ytic
bac
teri
aB
lood
ag
arO
lsen
et
al. (
2000
)A
. fra
nci
scan
an
aup
lii
Lab
cu
ltu
re (
star
ved
)W
AH
8.3±
8.1
×10
3V
ibri
oT
CB
S a
gar
Ols
en e
t al
. (20
00)
A. f
ran
cisc
ana
nau
pli
iL
ab c
ult
ure
(fe
d)
WA
H2.
2±2.
7 ×
103
Vib
rio
TC
BS
ag
arO
lsen
et
al. (
2000
)A
cart
ia t
onsa
Lab
cu
ltu
reW
AH
0.9
×10
3–
2.8
×10
4D
MS
P-c
onsu
min
g b
acte
ria
MP
NT
ang
et
al. (
2001
)Z
oop
lan
kto
n >
202
µm
Ch
opta
nk
Riv
er, M
D
WA
H9.
6±23
.3 ×
106
Bac
teri
aF
low
cyt
omet
ryH
eid
elb
erg
et
al. (
2002
)Z
oop
lan
kto
n >
202
µm
Ch
opta
nk
Riv
er, M
DW
AH
1.3±
3.6
×10
6G
amm
apro
teob
acte
ria
Flo
w c
ytom
etry
Hei
del
ber
g e
t al
. (20
02)
Zoo
pla
nk
ton
>20
2 µ
mC
hop
tan
k R
iver
, MD
WA
H6.
2±19
.1 ×
105
Vib
rio-
Ph
otob
acte
riu
mF
low
cyt
omet
ryH
eid
elb
erg
et
al. (
2002
)Z
oop
lan
kto
n >
202
µm
Ch
opta
nk
Riv
er, M
DW
AH
1.3±
2.3
×10
3V
. vu
lnif
icu
sF
low
cyt
omet
ryH
eid
elb
erg
et
al. (
2002
)Z
oop
lan
kto
n >
202
µm
Ch
opta
nk
Riv
er, M
DW
AH
577
±71
4V
. ch
oler
ae –
V. m
imic
us
Flo
w c
ytom
etry
Hei
del
ber
g e
t al
. (20
02)
Zoo
pla
nk
ton
>20
2 µ
mC
hop
tan
k R
iver
, MD
WA
H28
±38
V. c
inci
nn
atie
nsi
sF
low
cyt
omet
ryH
eid
elb
erg
et
al. (
2002
)Z
oop
lan
kto
n 6
4–20
2 µ
mC
hop
tan
k R
iver
, MD
WA
H2.
9±5.
12 ×
105
Bac
teri
aF
low
cyt
omet
ryH
eid
elb
erg
et
al. (
2002
)Z
oop
lan
kto
n 6
4–20
2 µ
mC
hop
tan
k R
iver
, MD
WA
H1.
5±2.
3 ×
104
Gam
map
rote
obac
teri
aF
low
cyt
omet
ryH
eid
elb
erg
et
al. (
2002
)Z
oop
lan
kto
n 6
4–20
2 µ
mC
hop
tan
k R
iver
, MD
WA
H6.
4±10
.5 ×
103
Vib
rio-
Ph
otob
acte
riu
mF
low
cyt
omet
ryH
eid
elb
erg
et
al. (
2002
)Z
oop
lan
kto
n 6
4–20
2 µ
mC
hop
tan
k R
iver
, MD
WA
H22
.6 ±
33.2
V. v
uln
ific
us
Flo
w c
ytom
etry
Hei
del
ber
g e
t al
. (20
02)
Zoo
pla
nk
ton
64–
202
µm
Ch
opta
nk
Riv
er, M
DW
AH
29.7
±38
.6V
. ch
oler
ae –
V. m
imic
us
Flo
w c
ytom
etry
Hei
del
ber
g e
t al
. (20
02)
Zoo
pla
nk
ton
64–
202
µm
Ch
opta
nk
Riv
er, M
DW
AH
1.0
±1.
9V
. cin
cin
nat
ien
sis
Flo
w c
ytom
etry
Hei
del
ber
g e
t al
. (20
02)
Aca
rtia
ton
saL
ab c
ult
ure
WA
H2.
0 ×
103
– 4.
5 ×
105
ND
MP
NT
ang
(20
05)
Cal
anu
s h
elg
olan
dic
us
Nor
th S
eaW
AH
1.9±
0.2
×10
5A
lph
apro
teob
acte
ria
DA
PI
Møl
ler
et a
l. (
2007
)an
d C
. fin
mar
chic
us
Dap
hn
ia c
ucu
llat
aL
ake
Ste
chli
nW
AH
1.0±
0.2
×10
5N
DS
ybrG
old
Tan
g e
t al
. (20
09a)
D.c
ucu
llat
aL
ake
Ste
chli
nW
AH
3.9±
1.5
×10
5N
DS
ybrG
old
Tan
g e
t al
. (20
09a)
Eu
dia
pto
mu
s g
raci
lis
Lak
e S
tech
lin
WA
H1.
7±0.
7 ×
105
ND
Syb
rGol
dT
ang
et
al. (
2009
a)E
. gra
cili
sL
ake
Ste
chli
nW
AH
4.3±
1.4
×10
5N
DS
ybrG
old
Tan
g e
t al
. (20
09a)
Dia
ph
anos
oma
bra
chyu
rum
Lak
e S
tech
lin
WA
H3.
3±0.
2 ×
105
ND
Syb
rGol
dT
ang
et
al. (
2009
a)
Tab
le 2
(co
nti
nu
ed)
Tang et al.: Zooplankton–bacteria interactions
contrast, the bacterial diversity greatly diverged andseveral species such as Pseudoalteromonas, Sulfito-bacter, and Roseobacter only appeared when the cope-pod was feeding on xenic phytoplankton diets, indicat-ing that many of the transient bacteria either activelyattached to the animal’s body surfaces or were pas-sively ingested (Tang et al. 2009b). Similarly, in a fieldstudy, Grossart et al. (2009) observed that the bacterialdiversity associated with freshwater zooplankton(Thermocyclops oithonoides and Bosmina coregoni)decreased after gut evacuation. In addition, theseauthors found that the magnitude of change in bacter-ial diversity induced by gut evacuation was smaller forzooplankton from an oligotrophic lake versus thosefrom a eutrophic lake. This suggests that the residentbacterial communities behave differently even for thesame zooplankton species when living in differentenvironments.
ZOOPLANKTON AS MICROBIAL REACTORS
Both resident and transient bacteria may takeadvantage of the rich organic environments providedby zooplankton and attain higher growth rates thanfree-living bacteria. For example, Carman (1994)demonstrated that nitrogen excretion by copepodsstimulates the growth of epibiotic bacteria. Tang (2005)further developed the concept that live copepods act asa reactor for bacterial colonization and growth. In anutritionally dilute environment, copepods throughfeeding concentrate organic matter into their guts,which their gut microflora can exploit to attain highgrowth rates. For gut bacteria (transient and resident)to exploit the ingested materials, they must first sur-vive digestion by the host. Several studies (e.g. King etal. 1991, Plante & Shriver 1998) have shown that manybacteria, including cyanobacteria (Friedland et al.2005), can survive zooplankton gut passage, and maythen gain access to specific resources that are other-wise limiting in the surrounding water. For example,dimethylsulfoniopropionate (DMSP) is produced bymany marine phytoplankton species (Keller & Korjeff-Bellows 1996) and is a substrate for DMSP-consumingbacteria (DCB; e.g. Diaz et al. 1992, Visscher et al.1992). However, direct release of DMSP from activelygrowing phytoplankton is negligible (Keller & Korjeff-Bellows 1996), and the half-saturation constant forfree-living DCB in coastal to oligotrophic waters (24 to>500 nM) is 5- to 50-fold higher than the dissolvedDMSP concentrations (Ledyard & Dacey 1996), indi-cating substrate limitation in ambient water. Throughfeeding, zooplankton can liberate DMSP from phyto-plankton cells and accumulate it in their guts to µM tomM levels (Tang et al. 1999), which would allow for
maximum turnover by DCB. DCB have indeed beenisolated from copepod bodies, and their abundanceincreased 17 to 30 times when the copepods were fedDMSP-containing food (Tang et al. 2001).
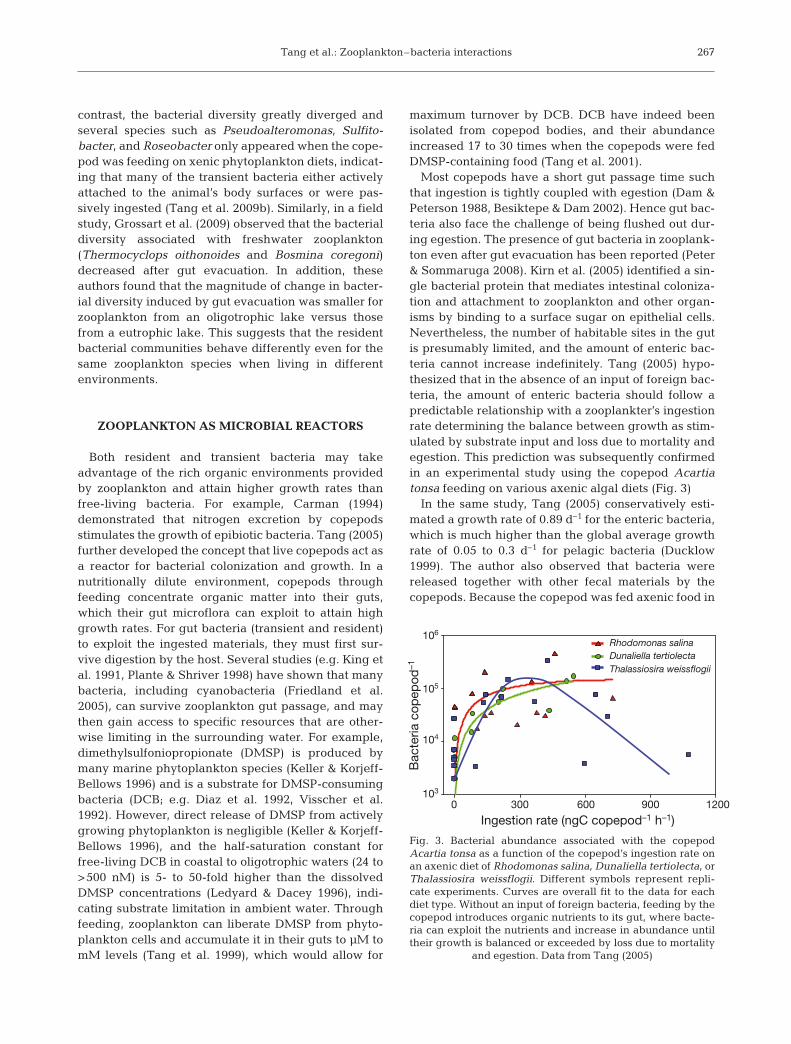
Most copepods have a short gut passage time suchthat ingestion is tightly coupled with egestion (Dam &Peterson 1988, Besiktepe & Dam 2002). Hence gut bac-teria also face the challenge of being flushed out dur-ing egestion. The presence of gut bacteria in zooplank-ton even after gut evacuation has been reported (Peter& Sommaruga 2008). Kirn et al. (2005) identified a sin-gle bacterial protein that mediates intestinal coloniza-tion and attachment to zooplankton and other organ-isms by binding to a surface sugar on epithelial cells.Nevertheless, the number of habitable sites in the gutis presumably limited, and the amount of enteric bac-teria cannot increase indefinitely. Tang (2005) hypo-thesized that in the absence of an input of foreign bac-teria, the amount of enteric bacteria should follow apredictable relationship with a zooplankter’s ingestionrate determining the balance between growth as stim-ulated by substrate input and loss due to mortality andegestion. This prediction was subsequently confirmedin an experimental study using the copepod Acartiatonsa feeding on various axenic algal diets (Fig. 3)
In the same study, Tang (2005) conservatively esti-mated a growth rate of 0.89 d–1 for the enteric bacteria,which is much higher than the global average growthrate of 0.05 to 0.3 d–1 for pelagic bacteria (Ducklow1999). The author also observed that bacteria werereleased together with other fecal materials by thecopepods. Because the copepod was fed axenic food in
267
Ingestion rate (ngC copepod–1 h–1)
Bac
teria
cop
epod
–1
0 300 600 900 1200
106
105
104
103
Rhodomonas salina
Dunaliella tertiolecta
Thalassiosira weissflogii
Fig. 3. Bacterial abundance associated with the copepodAcartia tonsa as a function of the copepod’s ingestion rate onan axenic diet of Rhodomonas salina, Dunaliella tertiolecta, orThalassiosira weissflogii. Different symbols represent repli-cate experiments. Curves are overall fit to the data for eachdiet type. Without an input of foreign bacteria, feeding by thecopepod introduces organic nutrients to its gut, where bacte-ria can exploit the nutrients and increase in abundance untiltheir growth is balanced or exceeded by loss due to mortality
and egestion. Data from Tang (2005)

Aquat Microb Ecol 61: 261–277, 2010
the experiments, the release of bacteria via defecationsuggests that actively feeding zooplankton function asmicroincubators and contribute new bacteria to thewater column. Although this contribution appears tobe small (<0.1%) relative to the ambient bacterialstanding stock, defecation and detachment may serveas important mechanisms for inoculating differentparts of the water column (Grossart et al. 2010).
ZOOPLANKTON AS A SELECTIVE FORCEFOR BACTERIA
The exterior as well as the interior of a zooplankterprovide specific habitats for aquatic bacteria. For ex-ample, bacteria attached to the surface of a zooplank-ter may experience a changing hydrodynamic environ-ment due to the host’s movement (Lawrence &Caldwell 1987) and must also adapt to the specific sur-face property (Dunne 2002). The reactive dissolved or-ganic carbon (DOC) pool in pelagic systems is mainlycomposed of molecules such as carbohydrates, aminoacids, fatty acids, hydrocarbons, and steroids, and theyusually account for a small fraction of total DOC in thewater column (<15%; Benner 2002). In contrast, reac-tive DOC (including amino acids and carbohydrates) inthe vicinity of zooplankton occurs in much higheramounts due to active excretion and sloppy feeding ofthe zooplankton, and this could become available forattached and even free-living bacteria (Peduzzi &Herndl 1992, Hansson & Norrman 1995, Møller et al.2003, Møller 2005). In an experimental mesocosmstudy, however, Kragh et al. (2006) demonstrated thatthe presence of zooplankton leads to an accumulationof less labile aldoses, indicating that zooplankton af-fect the quantity and quality of the DOC pool in thesurrounding water. As a result, release and accumula-tion of specific DOC compounds in the presence ofzooplankton potentially selects for specific bacterialphylotypes, e.g. opportunistic bacteria (Eilers et al.2000, Cottrell & Kirchman 2003).
Enteric bacteria, on the other hand, will experiencehigh organic matter and nutrient concentrations andlarge changes in pH and oxygen availability duringgut passage. The organic-rich, low oxygen environ-ment inside a zooplankter’s gut and fecal pellets mayfavor anaerobic bacterial processes that are otherwisenot feasible in the oxygenated water column. Indeed,an important observation in the past decades is thepresence of strict anaerobes inside the zooplankton gutand fecal pellets (Bianchi et al. 1992, Marty 1993, Proc-tor 1997, Braun et al. 1999). Globally it is estimated that12% of the primary production is consumed, directly orindirectly, by metazoan zooplankton (Calbet 2001).Due to the possible variety of bacterial metabolism
associated with zooplankton, these microenvironmentsrepresent a platform allowing for inter-linkages ofmicrobial processes in processing and degrading alarge fraction of the organic matters in marine andfreshwater environments, and support anaerobic pro-cesses in an otherwise aerobic environments (Stief etal. 2009). As such they have potentially significanteffects on global biogeochemical cycles.
The chemical composition of the zooplankton bodyas a potential source for bacterial substrates substan-tially differs from that of phytoplankton biomass. Dueto the lack of large amounts of carbohydrate storageproducts, carbon content of zooplankton is usuallylower in relation to nitrogen and phosphorus (Beers1966). Hence, the zooplankton body itself is a prefer-ential bacterial substrate (see below under ‘Zooplank-ton carcasses’). The chemical composition and avail-ability of zooplankton-derived particulate and dis-solved organic matters therefore has major implica-tions for bacterial dynamics and phylotype selectionnot only on the zooplankton but also in the surround-ing water.
ZOOPLANKTON AS A REFUGE FOR BACTERIA
Previous studies have shown that potentially patho-genic bacteria, e.g. Vibrio spp., find suitable growthconditions in zooplankton bodies (Colwell 1996). Free-living bacteria are constantly subject to environmentalhazards such as predation, viral lysis, and harmfulradiation and chemicals, and a zooplankton body mayprovide a refuge for bacteria against these externalhazards. Endosymbiotic bacteria that are commonlyfound in protozoa have been shown to survive disinfec-tants that normally kill free-living bacteria, hence pos-ing a great threat to public health (Barker & Brown1994, Bichai et al. 2008).
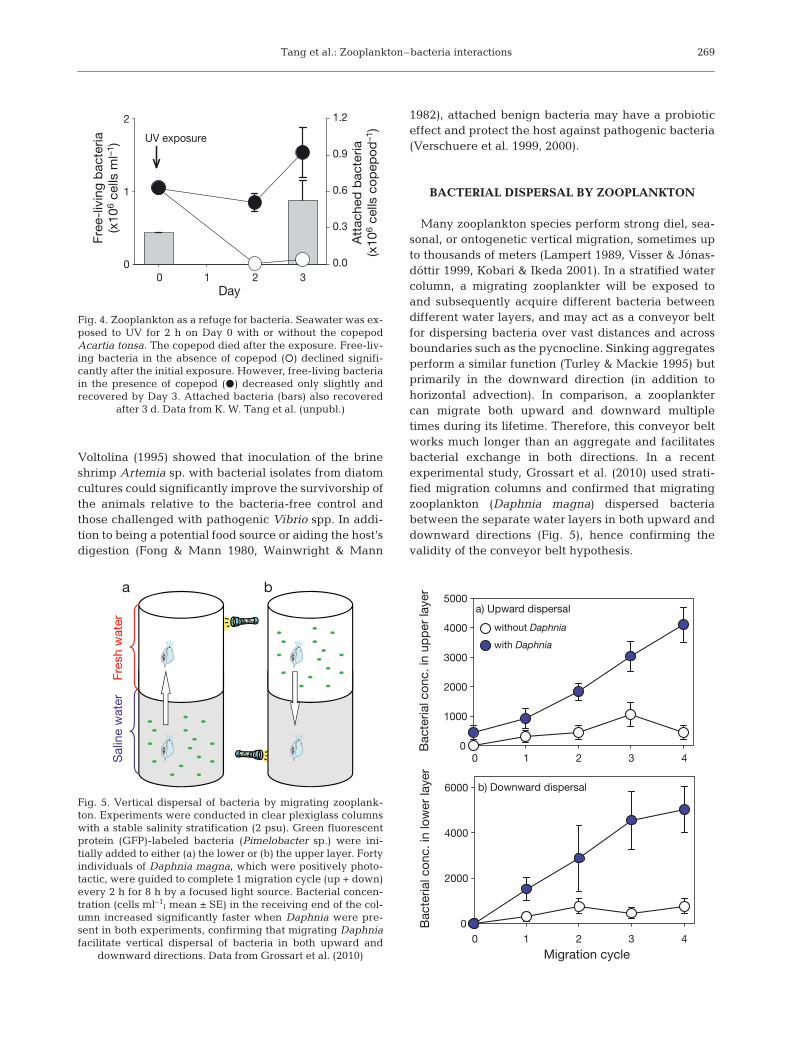
Various disinfection technologies are used commer-cially to treat ballast water in an attempt to curb theglobal spread of harmful organisms. Recent experi-ments by K. W. Tang et al. (unpubl.), however, suggestthat bacteria associated with cladocerans and cope-pods could survive conventional ballast water treat-ments such as heat, UV radiation, and ozonation eventhough free-living bacteria and zooplankton them-selves did not (Fig. 4). Subsequent discharge of treatedballast water into coastal zones may unknowinglyinoculate the local water with potential pathogens car-ried by zooplankton. Protection by zooplankton mayalso affect the evolution of local bacterial communitiesunder the influence of other environmental stresses.
While the zooplankton hosts may provide refuge forthe associated bacteria, the hosts themselves may alsobenefit from these bacteria. For example, Rico-Mora &
268
Tang et al.: Zooplankton–bacteria interactions
Voltolina (1995) showed that inoculation of the brineshrimp Artemia sp. with bacterial isolates from diatomcultures could significantly improve the survivorship ofthe animals relative to the bacteria-free control andthose challenged with pathogenic Vibrio spp. In addi-tion to being a potential food source or aiding the host’sdigestion (Fong & Mann 1980, Wainwright & Mann
1982), attached benign bacteria may have a probioticeffect and protect the host against pathogenic bacteria(Verschuere et al. 1999, 2000).
BACTERIAL DISPERSAL BY ZOOPLANKTON
Many zooplankton species perform strong diel, sea-sonal, or ontogenetic vertical migration, sometimes upto thousands of meters (Lampert 1989, Visser & Jónas-dóttir 1999, Kobari & Ikeda 2001). In a stratified watercolumn, a migrating zooplankter will be exposed toand subsequently acquire different bacteria betweendifferent water layers, and may act as a conveyor beltfor dispersing bacteria over vast distances and acrossboundaries such as the pycnocline. Sinking aggregatesperform a similar function (Turley & Mackie 1995) butprimarily in the downward direction (in addition tohorizontal advection). In comparison, a zooplanktercan migrate both upward and downward multipletimes during its lifetime. Therefore, this conveyor beltworks much longer than an aggregate and facilitatesbacterial exchange in both directions. In a recentexperimental study, Grossart et al. (2010) used strati-fied migration columns and confirmed that migratingzooplankton (Daphnia magna) dispersed bacteriabetween the separate water layers in both upward anddownward directions (Fig. 5), hence confirming thevalidity of the conveyor belt hypothesis.
269
Day
Att
ache
d b
acte
ria(x
106
cells
co
pep
od
–1)
0.0
0.3
0.6
0.9
1.2
Fre
e-liv
ing
bac
teria
(x10
6 ce
lls m
l–1 )
00 1 2 3
1
2
UV exposure
Fig. 4. Zooplankton as a refuge for bacteria. Seawater was ex-posed to UV for 2 h on Day 0 with or without the copepodAcartia tonsa. The copepod died after the exposure. Free-liv-ing bacteria in the absence of copepod (s) declined signifi-cantly after the initial exposure. However, free-living bacteriain the presence of copepod (d) decreased only slightly andrecovered by Day 3. Attached bacteria (bars) also recovered
after 3 d. Data from K. W. Tang et al. (unpubl.)
a b
Bac
teria
l con
c. in
up
per
laye
r
0
1000
2000
3000
4000
5000
without Daphnia
with Daphnia
a) Upward dispersal
b) Downward dispersal
Migration cycle
Bac
teria
l con
c. in
low
er la
yer
Sal
ine
wat
erFr
esh
wat
er
00 1 2 3 4
0 1 2 3 4
2000
4000
6000Fig. 5. Vertical dispersal of bacteria by migrating zooplank-ton. Experiments were conducted in clear plexiglass columnswith a stable salinity stratification (2 psu). Green fluorescentprotein (GFP)-labeled bacteria (Pimelobacter sp.) were ini-tially added to either (a) the lower or (b) the upper layer. Fortyindividuals of Daphnia magna, which were positively photo-tactic, were guided to complete 1 migration cycle (up + down)every 2 h for 8 h by a focused light source. Bacterial concen-tration (cells ml–1; mean ± SE) in the receiving end of the col-umn increased significantly faster when Daphnia were pre-sent in both experiments, confirming that migrating Daphniafacilitate vertical dispersal of bacteria in both upward and
downward directions. Data from Grossart et al. (2010)

Aquat Microb Ecol 61: 261–277, 2010
Research on aggregates has shown that bacteriaattached to surfaces are capable of quorum sensing viahomoserine lactone communication (Gram et al. 2002),which allows them to synchronize specific metabolicprocesses to maximize their metabolic rate and growthefficiency. In the case of probiotic as well as patho-genic bacteria, this will have important consequencesfor the environment. Multidrug resistance can fre-quently be found in bacteria on fish, and it is believedthat the increased tolerance to antibiotics and heavymetals is related to the close proximity of resistant andnon-resistant bacteria, which allows for increased hor-izontal gene exchange (e.g. Pathak & Gopal 2005).Long-range migration by zooplankton and other ani-mal hosts may help the spread of drug-resistant geno-types across a broad geographical range even amongotherwise isolated water masses, posing a hiddenthreat to ecosystem and human health.
OTHER MODES OF ZOOPLANKTON–BACTERIAASSOCIATION
Zooplankton feces
Approximately a third of the food ingested bypelagic organisms is egested as feces (Parsons et al.1977). Zooplankton such as copepods and salps pro-duce fast-sinking fecal pellets, from tens to thousandsm d–1 (Bruland & Silver 1981, Komar et al. 1981, Caronet al. 1989), and thus contribute substantially to the‘biological pump’ (Cherry et al. 1978). On the otherhand, fecal matter may be remineralized by bacteriawithin the water column (Gowing & Silver 1983, Jacob-sen & Azam 1984, Caron et al. 1989), reducing theoverall vertical flux. This process can be furtherenhanced by physical breakage of the pellets by zoo-plankton (Lampitt et al. 1990). Differences in the phys-ical structure of the pellets may also lead to differentremineralization rates. For example, a copepod fecalpellet is bound by a peritrophic membrane and mayremain intact for a longer time, whereas a salp fecalpellet lacks a membrane and tends to be more brittle.
Fecal matter released by zooplankton often alreadycarries large amounts of bacteria (Turner 1979). Forexample, the reported equivalent total bacterial con-centrations in copepod fecal pellets are 1010 to 1011
cells ml–1 for Acartia tonsa (Hansen & Bech 1996, Thoret al. 2003) and 103 to 105 cells ml–1 for Temora stylifera(Delille & Razouls 1994). The latter study also showedthat the frequency of dividing bacterial cells and meanbacterial cell volumes increased, respectively, from3% and 0.12 µm3 in seawater to >6% and 0.24 µm3 infecal pellets. Previously, it had been thought that bac-terial colonization of zooplankton fecal pellets is
mainly initiated from the outside by chemotactic bacte-ria in the surrounding water (Honjo & Roman 1978,Turner 1979, Jacobsen & Azam 1984). However, Gow-ing & Silver (1983) demonstrated that most of the bac-terial colonization is from the inside by enteric anddigestion-resistant bacteria. The presence of bacteriain fecal materials produced by copepods feeding onaxenic food also confirmed that enteric bacteria can bepassed onto fecal pellets (Tang 2005). On the otherhand, a study with deposit-feeders showed that bacte-rial proliferation on egesta was faster than could beexplained by growth alone (Plante & Wilde 2001), indi-cating that colonization by external bacteria must havealso occurred at the same time.
Zooplankton molts
Chitin is the second-most abundant biogenic poly-saccharide on earth after cellulose and hence is amongthe most abundant and important sources of nutrientsand energy in the marine environment (Gooday 1990).Chitin is a major component of the cuticles andexoskeletons of worms, mollusks, and arthropods (Jeu-niaux 1982). The annual biosynthesis of chitin hasbeen estimated at 1.4 billion tons in all aquatic envi-ronments combined (Cauchie 2002). The dry mass ofchitin produced by the planktonic crustacean Euphau-sia pacifica through molting alone is ca. 5 to 12 milliontons annually (Jerde & Lasker 1966, Jeuniaux 1971).Yu et al. (1991) asserted that chitin production by cope-pods and other organisms leads to a continuous ‘rain’of particulate organic matter. Furthermore, molts (exu-viae) of crustacean zooplankton significantly contri-bute to the formation of macroscopic organic aggre-gates in marine (Gardner 1977) and freshwater (Grossartet al. 1997, Grossart & Simon 1998) systems. From anecological point of view, chitin plays a key role in thebiogeochemical cycles of both C and N, and the ratesof chitin production and degradation influence C andN pools and their availability (Poulicek et al. 1998,Pruzzo et al. 2008).
In his pioneering in situ study of chitin degradation(practical grade chitin from crustacean shells) using themesh bag technique, Kirchner (1995) revealed that 90%of the initial substrate was lost after 3 mo of incubationin seawater at 10 to 18°C. Other studies also foundthat several marine bacteria are capable of degradingchitin (Bassler et al. 1991, Keyhani & Roseman 1999,Ramaiah et al. 2000, Tilly et al. 2001, Itoi et al. 2006), butnone of them used natural zooplankton molts as asubstrate. Studying bacterial colonization of lake snow inLake Constance, Weiss et al. (1996) showed thataggregates containing a high fraction of zooplanktonexoskeleton during the clear water phase were mainly
270

Tang et al.: Zooplankton–bacteria interactions
colonized by Proteobacteria of the Alpha (5 to 32%), Beta(20 to 60%), and Gamma subclasses (6 to 42%). Inthe Delaware estuary, Cottrell & Kirchman (2003) foundchitinase genes in a handful of bacterial groups in-cluding Enterobacteriaceae, Alteromondaceae, Vibrio-ceae, and Alphaproteobacteria of the Roseobacter clade.However, except for Vibrio spp., almost nothing isknown about bacterial colonization of zooplanktonmolts. Tamplin et al. (1990) showed that 4 of 5 clinicalV. cholerae O1 strains and endogenous bacteria pre-ferentially attached to zooplankton molts rather thanwhole animals. V. cholerae growing on natural chitinsurfaces show coordinated gene expression involvedwith chitin chemotaxis and adherence as well as thetransport and assimilation of N-acetylglucosamine(GlcNAc; Meibom et al. 2004), suggesting that thisspecies is well adapted to using this highly abundantcarbon and nitrogen source in aquatic environments.
Using methylumbelliferyl-N, N’-diacetyl-chitobio-side, and 14C-chitin, Kirchman & White (1999) showedthat bacterial hydrolysis of chitin was always fasterthan mineralization, resulting in the release of GlcNActo the surrounding water. On average, however, chitinhydrolysis and uptake accounts for only 10% of thetotal bacterial production (Kirchman & White 1999).Interestingly, Riemann & Azam (2002) showed not onlythat roughly a third of all marine bacteria take up Glc-NAc, but also that these bacteria are predominantlyfacultative anaerobes. The capability of fermentativemetabolism could be an important adaptation to lowoxygen microenvironments in the pelagial associatedwith molts, fecal pellets, and zooplankton gut.
Zooplankton carcasses
Traditional zooplankton sampling tends to ignore thepresence of carcasses due to methodological difficultiesin separating live and dead animals. Nevertheless, zoo-plankton carcasses have been observed worldwide(Tang et al. 2009a), and simple staining methods arenow available for distinguishing between live and deadcrustacean zooplankton in freshwater and marine sys-tems (Bickel et al. 2009, Elliott & Tang 2009). Usingthese staining methods, it has been shown that up to40% of the zooplankton (copepods and daphnids) canbe dead in situ (Bickel et al. 2009, Elliott & Tang 2009).This is of particular interest since carcasses of bothcrustacean and non-crustacean zooplankton speciesrepresent concentrated reservoirs of labile organic sub-strates for water column bacteria (Tang et al. 2006a,Titelman et al. 2006, Bickel & Tang in press). Previousexperiments have shown that zooplankton carcassesare rapidly colonized and decomposed by bacteriamainly from the outside (Harding 1973, Bickel & Tang
in press), with the caveat that some of the epibiontscould have been killed by the treatment that producedthe zooplankton carcasses, leading to an underesti-mation of the contribution to the decomposition processby epibionts. The initial increase in bacterial abun-dance during the carcass decomposition process istemperature-dependent but is less dependent on aero-bic versus anaerobic conditions (Tang et al. 2006b).Tang et al. (2009a) observed that cladoceran carcasseswere colonized by bacteria faster than copepod car-casses both in the laboratory and in the field. In addi-tion, carcasses suspended in a eutrophic lake had ahigher average carbon loss rate than those suspendedin an oligotrophic lake. These differences suggest thatexploitation of zooplankton carcasses by bacteria de-pends on carcass type and environmental conditions.
In addition to bacteria, fungi are also able to colonizeand decompose zooplankton carcasses, especiallywhen bacterial activities are suppressed (Tang et al.2006b). The exoskeleton of zooplankton contains alarge fraction of chitin and can be densely colonized byaquatic fungi. Chitin is often used for isolation and cul-tivation of a variety of fungi, and fungal chitinases arewell known to efficiently degrade chitin (reviewed byWurzbacher et al. 2010). Hence zooplankton molts andcarcasses are important organic carbon sources notsolely for bacteria but also for fungi.
Although these carcass-colonizing microbes exhibitboth very high protease and lipase activities, carcassprotein decomposition is faster than lipid decomposi-tion (Bickel & Tang in press), resulting in differentialremineralization and preservation of different bio-chemical components of the carcasses. Analysis of sim-ilarity (ANOSIM) of denaturing gradient gel elec-trophoresis (DGGE) banding patterns revealed thatbacterial communities on decomposing zooplanktoncarcasses rapidly diverged from those in the surround-ing water, but remained similar among different typesof zooplankton carcasses (Tang et al. 2009a). Theseobservations demonstrate that zooplankton carcassesare decomposed by similar bacterial groups and serveas important microbial microenvironments whererapid and efficient local selection takes place.
In many lakes, crustacean zooplankton populationscan suffer high and abrupt mortality due to starvationand diseases resulting in the phenomenon of ‘mid-summer decline’ (Gries & Güde 1999, Hülsmann &Weiler 2000, Hülsmann & Voigt 2002). Massive die-offof zooplankton will produce abundant zooplanktoncarcasses as microbial hotspots to fuel water columnand benthic bacterial production. Unfortunately, inmost traditional microbial studies, these hotspots havebeen excluded by pre-filtration of water samples.
There are very few published studies on the poten-tially important aspect of jellyfish blooms, i.e. the fate
271

Aquat Microb Ecol 61: 261–277, 2010
of jellyfish-derived organic matter. Jellyfish carcassescan impact microbial processes, causing significantchanges to nitrogen and oxygen dynamics in the sur-rounding environment (Pitt et al. 2009, West et al.2009) as well as the composition and activity of theambient bacterial community (Riemann et al. 2006,Titelman et al. 2006, Tinta et al. 2010). Some prelimi-nary results of the bacterial community structure asmeasured by DGGE indicated differences in thebacterial community response between ecosystemswhere medusae occur throughout the year and areaswhere they occur only seasonally (Tinta et al. 2010).Mass deposition of jellyfish and salp carcasses hasalso been hypothesized to fuel the microbial foodweb in the seabed (Billet et al. 2006, Lebrato & Jones2009).
FUTURE CHALLENGES AND OPPORTUNITIES
The available, albeit limited, quantitative data in theliterature (Table 2) indicate large variability in zoo-plankton-associated bacterial abundance in space andin time, as well as among zooplankton species. Under-standing what factors contribute to this variability willbe an important task for researchers. Because zoo-plankton likely acquire bacteria via direct bacterialattachment and ingestion, zooplankton-associatedbacterial abundance could be dependent on ambientfree-living and particle-attached bacterial concentra-tions. Differences between zooplankton species maybe related to the zooplankton feeding strategy (e.g.non-selective versus selective feeders) and body size.Because both bacteria and zooplankton reside within aviscous environment, their encounter and interactionswill also be influenced by ambient flow. The use ofnew in situ flow visualization experiments (Dabiri et al.2005, Costello et al. 2008) and new mechanical modelsbased on hydrodynamic patterns might explain therelationship between flow, zooplankton, and microbialcommunity ecology. Simple and readily testable hypo-theses as such may unveil some general patterns inzooplankton–bacteria associations.
Past research on zooplankton–bacteria associationsis largely limited to crustacean zooplankton, and ex-tension of sampling effort to gelatinous zooplanktonand microzooplankton is much needed. Many investi-gators used whole animals or homogenates, and there-fore their measurements do not separate epibionts andgut microflora (Tables 1 & 2). Because the 2 communi-ties are exposed to very different environments, sepa-ration between the two, for example by microdissec-tion of the zooplankton gut (Peter & Sommaruga 2008),will be necessary for proper understanding of theirrespective physiology and ecology.
While previous work has tended to rely on culture-dependent methods and focused on a few pathogenicspecies (most notably Vibrio spp.), modern culture-independent molecular techniques have revealed verydiverse assemblages of bacteria associated withmarine and freshwater zooplankton, many of whosepathogenicity and other characteristics remain unex-plored. Techniques such as metagenome sequencing,bioinformatics, systems biology, and new statisticalapproaches should allow for detailed analyses ofchanges in bacterial community structure in relation tothe life cycle and behavior of the zooplankton host.Study of the metatranscriptome and proteome of thesecomplex bacterial communities will also help us under-stand their ecological roles on the hosts as well as inthe surrounding water, and when combined with sin-gle cell techniques such as Raman microscopy andNanoSims (secondary ion mass spectrometry), it willeven be possible to quantify microbial processes in andon live zooplankton. Microautoradiography in combi-nation with fluorescence in situ hybridization (MAR-FISH) can be used to more accurately measure specificsubstrate uptake and growth of bacteria associatedwith the different zooplankton body parts.
Although the presence of strict anaerobes inside thezooplankton gut has been known for some time (e.g.Proctor 1997), the physical-chemical characteristics ofa zooplankter’s gut remain poorly known. These envi-ronments may vary according to the feeding activitiesand physiology of the zooplankton and subsequentlyselect for different bacterial communities. The use ofpH- or oxygen-sensitive chemicals provides only acrude characterization of the gut environments (e.g.Pond et al. 1995). An alternative is to use microsen-sors to generate detailed profiles of the gut for pH,oxygen, nitrous oxides, sulfide, and other chemicalspecies, which has been done successfully with ter-restrial insects (e.g. Bignell & Anderson 1980). De-tailed characterization of the gut environment in com-bination with molecular analysis of the gut microflorawould allow researchers to study the selection andadaption of different bacteria in this unique micro-environment.
In addition to live zooplankton, other microenviron-ments such as zooplankton fecal pellets, molts, andcarcasses also warrant further study. Although bacter-ial activities on zooplankton fecal pellets have beenintensively studied in the past decades, they arelargely limited to copepod fecal pellets. Fecal pelletsproduced by other zooplankton groups have differentsizes and textures, but information on their bacterialactivities is scarce. The reported occurrence of strictanaerobes in zooplankton fecal pellets (e.g. Bianchi etal. 1992, Marty 1993) suggests that, similar to the gut,fecal pellets provide a semi-enclosed environment
272

Tang et al.: Zooplankton–bacteria interactions
where microbial activities proceed very differentlythan in the well oxygenated surface water. Microbialactivities in fecal pellets can contribute to water col-umn denitrification and methane production (Orem-land 1979, de Angelis & Lee 1994, Michotey & Bonin1997), but the internal oxygen environment of fecalpellets remains largely unknown, a gap that can befilled by microsensor profiling combined with ad-vanced molecular analysis.
To our knowledge, bacterial colonization of crus-tacean zooplankton molts and carcasses has not beenstudied in detail using modern molecular techniquessuch as metagenomic and metatranscriptomic ap-proaches. This is surprising, considering the globalabundance of chitin and that chitin degradation byaquatic bacteria is essential for returning carbon andnitrogen to aquatic nutrient cycles (Kirchner 1995,Flintoft 2004). Zooplankton carcasses may be of partic-ular importance at times when other bacterial sub-strates are limited, e.g. during the clear water phasewhich follows the phytoplankton spring bloom in manytemperate lakes. Besides bacteria, aquatic fungi arealso capable of degrading chitin (Tang et al. 2006b,Wurzbacher et al. 2010), although their ecological rolein remineralizing zooplankton molts remains to beexplored. The combination of both molecular and eco-logical methods will provide a better understanding ofhow zooplankton molts and carcasses can affect bacte-rial community composition, microbial organic mattertransformation, and biogeochemical cycling in thepelagic zone.
Comparative studies of different carcass types arealso needed. For example, gelatinous zooplankton car-casses lack a chitinous covering, and presumablydecompose more readily than crustacean carcasses,yet the limited data available have suggested other-wise (Bickel & Tang in press). Bacterial decompositionof jellyfish is also of particular interest because of theperceived increase in occurrence of jellyfish ‘blooms’worldwide (Mills 2001, Purcell et al. 2007). Becausemost jellyfish species have few natural predators, theircarcasses at the termination of a bloom will likely havelarge impact on the water column and benthic micro-bial communities (Titelman et al. 2006, Tinta et al.2010).
The probiotic versus harmful effects of different bac-terial species on the zooplankton hosts have far-reach-ing implications for aquaculture practices as well aszooplankton ecophysiology and ecosystem health. Sofar, research on this topic is largely restricted to brineshrimp because of the relative ease with which com-mercially available encysted eggs may be disinfectedto produce bacteria-free animals for exposure experi-ments (e.g. Verschuere et al. 1999, 2000). Althoughcopepods and daphnids, the most representative of
metazoan zooplankton in aquatic environments, doproduce encysted resting eggs (e.g. Uye 1985, Alek-seev & Lampert 2001), these resting eggs are not easilyavailable, and the feasibility of using them to producebacteria-free animals has not been carefully evaluated.The difficulty in producing bacteria-free zooplanktoncultures will limit our ability to study how differentbacterial species or assemblages affect the host. Topartially circumvent this problem, comparative studiesof the physiology and ecology of the same zooplanktonspecies but colonized by different bacterial assem-blages may provide helpful insights.
CONCLUDING REMARKS
Increasing specialization in science has acceleratedthe advances of many research disciplines, but at thesame time has encouraged an increasingly fragmentedand incomplete understanding of the natural world.This is evident in the traditional separation of ‘micro-bial ecology’ and ‘zooplankton ecology.’ As a conse-quence, we may have missed a large portion of themicrobial world and hence are still far from fullyunderstanding biodiversity and functioning of theecosystem. Bridging these disciplines requires anunconventional way of thinking and approach toresearch, and recent studies on this topic have laid thenecessary foundation for it.
Our review shows that bacteria colonize and interactwith zooplankton in many ways with far-reachingimplications for microbial production, dispersal, di-versity maintenance, evolution, and biogeochemicalfluxes (Fig. 6). Nevertheless, available information isstill limited to only a few zooplankton groups and a fewenvironments, in particular copepods in coastal andestuarine waters. Because different zooplankton taxacan have very different life history traits that can influ-ence the form of zooplankton-bacteria interactions,extension of research efforts to other zooplankton taxais much needed. In the oligotrophic open ocean wherenutrients are limiting in the water column, direct asso-ciation with zooplankton may become more importantfor bacteria (cf. Grossart et al. 2009). Recent and con-tinuous methodological advances in microbiology andmicrobial ecology will allow researchers to study theseinteractions in unprecedented detail. As the complex-ity of zooplankton–bacteria interactions continues tounfold, new ecological concepts and models will beneeded to help researchers integrate the new informa-tion into our current understanding of the ecosystem.We hope that this article will encourage more collabo-ration between zooplankton ecologists and microbialecologists to advance this exciting and promisingresearch topic.
273

Aquat Microb Ecol 61: 261–277, 2010
Acknowledgements. K.W.T. received support from the U.S.National Science Foundation (award OCE-0814558). H.P.G.was supported by grants GR1540/11-2 and PA1655/1-1 fromthe German Science Foundation (DFG). We thank E. Machand C. Freund for assistance.
LITERATURE CITED
Alekseev V, Lampert W (2001) Maternal control of resting-egg production in Daphnia. Nature 414:899–901
Austin B, Allen DA (1982) Microbiology of laboratory-hatchedbrine shrimp (Artemia). Aquaculture 26:369–383
Azam F, Malfatti F (2007) Microbial structuring of marineecosystems. Nat Rev Microbiol 5:782–791
Barker J, Brown MRW (1994) Trojan horse of the microbialworld: protozoa and the survival of bacterial pathogens inthe environment. Microbiology 140:1253–1259
Bassler BL, Gibbons PJ, Yu C, Roseman S (1991) Chitin uti-lization by marine bacteria. Chemotaxis to chitin oligosac-charides by Vibrio furnissii. J Biol Chem 266:24268–24275
Beers JR (1966) Studies on the chemical composition of themajor zooplankton groups in the Sargasso Sea offBermuda. Limnol Oceanogr 11:520–528
Benner R (2002) Chemical composition and reactivity. In:Hansell DA, Carlson CA (eds) Biogeochemistry of marineDissolved Organic Matter. Academic Press, Amsterdam,p 59–90
Besiktepe S, Dam HG (2002) Coupling of ingestion and defe-cation as a function of diet in the calanoid copepod Acar-tia tonsa. Mar Ecol Prog Ser 229:151–164
Bianchi M, Marty D, Teysslé JL, Fowler SW (1992) Strictlyaerobic and anaerobic bacteria associated with sinkingparticulate matter and zooplankton fecal pellets. Mar EcolProg Ser 88:55–60
Bichai F, Payment P, Barbeau B (2008) Protection of water-borne pathogens by higher organisms in drinking water: areview. Can J Microbiol 54:509–524
Bickel SL, Tang KW (2010) Microbial decomposition of pro-teins and lipids in copepod versus rotifer carcasses. MarBiol 157:1613–1624
Bickel SL, Tang KW, Grossart HP (2009) Use of aniline blue todistinguish live and dead crustacean zooplankton compo-sition in freshwaters. Freshw Biol 54:971–981
Bignell DE, Anderson JM (1980) Determination of pH andoxygen status in the guts of lower and higher termites.J Insect Physiol 26:183–188
Billet DSM, Bett BJ, Jacobs CL, Rouse IP, Wigham BD (2006)Mass deposition of jellyfish in the deep Arabian Sea.Limnol Oceanogr 51:2077–2083
Braun ST, Proctor LM, Zani S, Mellon MT, Zehr JP (1999)Molecular evidence for zooplankton-associated nitrogen-fixing anaerobes based on amplification of the nifH gene.FEMS Microbiol Ecol 28:273–279
Bruland KW, Silver MW (1981) Sinking rates of fecal pelletsfrom gelatinous zooplankton (salps, pteropods, doliolids).Mar Biol 63:295–300
Calbet A (2001) Mesozooplankton grazing effect on primaryproduction: a global comparative analysis in marine eco-systems. Limnol Oceanogr 46:1824–1830
Carli A, Pane L, Casareto L, Bertone S, Pruzzo C (1993)Occurrence of Vibrio alginolyticus in Ligurian coast rock
274
High C-Sequestration
WithoutWith
Low C-Sequestration
Hotspots of organic matter
Bacterial colonization and protection sites
High nutrient and organic matter recycling
High CO2 fixation rates and biomass standing stock
Low CO2 fixation rates and biomass standing stock
Low nutrient and organic matter recycling
Fig. 6. Aquatic biogeochemical processes as influenced by zooplankton. Zooplankton have been shown to increase particulateorganic matter removal, nutrient regeneration, and vertical carbon flux. These processes will be modulated by bacteriaassociated with the zooplankton and fecal matter. Zooplankton and fecal matter also provide a microenvironment where
bacterial production, diversity maintenance, and evolution may proceed differently than in the surrounding water

Tang et al.: Zooplankton–bacteria interactions
pools (Tyrrhenian Sea, Italy) and its association with thecopepod Tigriopus fulvus. Appl Environ Microbiol 59:1960–1962
Carman KR (1994) Stimulation of marine free-living andepibiotic bacterial activity by copepod excretions. FEMSMicrobiol Ecol 14:255–262
Carman KR, Dobbs FC (1997) Epibiotic microorganisms oncopepods and other marine crustaceans. Microsc Res Tech37:116–135
Caron DA, Madin LP, Cole JJ (1989) Composition and degra-dation of salp fecal pellets: implications for vertical flux inoceanic environments. J Mar Res 47:829–850
Carrias JF, Sime-Ngando T (2009) Bacteria, attached to sur-faces. In: Likens GE (ed) Encyclopedia of inland waters,Vol 3. Elsevier, Oxford, p 182–192
Cauchie HM (2002) Chitin production by arthropods in thehydrosphere. Hydrobiologia 470:63–96
Cherry RD, Higgo JJW, Fowler SW (1978) Zooplankton fecalpellets and element residence times in the ocean. Nature274:246–248
Colwell RR (1996) Global climate and infectious disease: thecholera paradigm. Science 274:2025–2031
Costello JH, Colin SP, Dabiri JO (2008) Medusan morpho-space: phylogentic constraints, biomechanical solutions,and ecological consequences. Invertebr Biol 127:265–290
Cottingham KL, Chiavelli DA, Taylor RK (2003) Environmen-tal microbe and human pathogen: the ecology and micro-biology of Vibrio cholerae. Front Ecol Environ 1:80–86
Cottrell MT, Kirchman DL (2003) Contribution of major bacte-rial groups to bacterial biomass production (thymidine andleucine incorporation) in the Delaware Estuary. LimnolOceanogr 48:168–178
Dabiri JO, Colin SP, Costello JH, Gharib M (2005) Flow pat-terns generated by oblate medusan jellyfish: field mea-surements and laboratory analyses. J Exp Biol 208:1257–1265
Dam HG, Peterson WT (1988) The effect of temperature onthe gut clearance rate constant of planktonic copepods.J Exp Mar Biol Ecol 123:1–14
Dattagupta S, Schaperdoth I, Montanari A, Mariani S, Kita N,Valley JW, Macalady JL (2009) A novel symbiosis betweenchemoautotrophic bacteria and a freshwater cave amphi-pod. ISME J 3:935–943
de Angelis M, Lee C (1994) Methane production during zoo-plankton grazing on marine phytoplankton. LimnolOceanogr 39:1298–1308
Delille D, Razouls S (1994) Community structures of hetero-trophic bacteria of copepod fecal pellets. J Plankton Res16:603–615
Diaz MR, Visscher PT, Taylor BF (1992) Metabolism ofdimethylsulfoniopropionate and glycine betaine by amarine bacterium. FEMS Microbiol Lett 96:61–66
Ducklow HW (1999) The bacterial component of the oceaniceuphotic zone. FEMS Microbiol Ecol 30:1–10
Dunne WM Jr (2002) Bacteria adhesion: Seen any goodbiofilms lately? Clin Microbiol Rev 15:155–166
Eilers H, Pernthaler J, Glöckner FO, Amann R (2000) Cultur-ability and in situ abundance of pelagic bacteria from theNorth Sea. Appl Environ Microbiol 66:3044–3051
Elliott DT, Tang KW (2009) Simple staining method for differ-entiating live and dead marine zooplankton in field sam-ples. Limnol Oceanogr Methods 7:585–594
Flintoft L (2004) Bacterial physiology: unlocking chitin degra-dation. Nat Rev Microbiol 2:90
Flood PR (1991) Architecture of and and water circulation andflow rate in the house of the planktonic tunicate Oiko-pleura labradoriensis. Mar Biol 111:95–111
Folt CL, Burns CW (1999) Biological drivers of zooplanktonpatchiness. Trends Ecol Evol 14:300–305
Fong W, Mann KH (1980) Role of gut flora in the transferof amino acids through a marine food chain. Can J FishAquat Sci 37:88–96
Friedland KD, Ahrenholz DW, Haas LW (2005) Viable gutpassage of cyanobacteria through the filter-feeding fishAtlantic menhaden, Brevoortia tyrannus. J Plankton Res27:715–718
Gardner WD (1977) Incomplete extraction of rapidly settlingparticles from water samples. Limnol Oceanogr 22: 764–768
Gooday GW (1990) The ecology of chitin degradation. AdvMicrob Ecol 11:387–430
Gowing MM, Silver MW (1983) Origins and microenviron-ments of bacteria mediating fecal pellet decomposition inthe sea. Mar Biol 73:7–16
Gram L, Grossart HP, Schlingloff A, Kiørboe T (2002) Possiblequorum sensing in marine snow bacteria: production ofacylated homoserine lactones by Roseobacter strains iso-lated from marine snow. Appl Environ Microbiol 68:4111–4116
Gries T, Güde H (1999) Estimates of the nonconsumptive mor-tality of mesozooplankton by measurement of sedimenta-tion losses. Limnol Oceanogr 44:459–465
Grossart HP (2010) Ecological consequences of bacterio-plankton lifestyles: changes in concepts are needed. Envi-ron Microbiol, doi:10.1111/j.1758-2229.2010.00179.x
Grossart HP, Simon M (1998) Bacterial colonization andmicrobial decomposition of limnetic organic aggregates(lake snow). Aquat Microb Ecol 15:127–140
Grossart HP, Simon M, Logan BE (1997) Formation of macro-scopic organic aggregates (lake snow) in a large lake:the significance of transparent exopolymer particles,phytoplankton, and zooplankton. Limnol Oceanogr 42:1651–1659
Grossart HP, Dziallas C, Tang KW (2009) Bacterial diversityassociated with freshwater zooplankton. Environ Micro-biol Rep 1:50–55
Grossart HP, Dziallas C, Leunert F, Tang KW (2010) Bacteriadispersal by hitchhiking on zooplankton. Proc Natl AcadSci USA, doi:10.1073/pnas.1000668107
Hansen B, Bech G (1996) Bacteria associated with a marineplanktonic copepod in culture. I. Bacterial genera in sea-water, body surface, intestines and fecal pellets andsuccession during fecal pellet degradation. J Plankton Res18:257–273
Hansson LJ, Norrman B (1995) Release of dissolved organiccarbon (DOC) by the scyphozoan jellyfish Aurelia auritaand its potential influence on the production of plankticbacteria. Mar Biol 121:527–532
Harding GCH (1973) Decomposition of marine copepods.Limnol Oceanogr 18:670–673
Harris JM (1993) The presence, nature, and role of gutmicroflora in aquatic invertebrates: a synthesis. MicrobEcol 25:195–231
Heidelberg JF, Heidelberg KB, Colwell RR (2002) Bacteria ofthe ?-subclass Proteobacteria associated with zooplankton inChesapeake Bay. Appl Environ Microbiol 68:5498–5507
Holland R, Hergenrader R (1981) Bacterial epibionts of diap-tomid copepods. Trans Am Microsc Soc 100:56–65
Honjo S, Roman MR (1978) Marine copepod fecal pellets: pro-duction, preservation, and sedimentation. J Mar Res 36:45–57
Hülsmann S, Voigt H (2002) Life history of Daphnia galeata ina hypertrophic reservoir and consequences of non-con-sumptive mortality for the initiation of a midsummerdecline. Freshw Biol 47:2313–2324
275

Aquat Microb Ecol 61: 261–277, 2010
Hülsmann S, Weiler W (2000) Adult, not juvenile mortality asa major reason for the midsummer decline of a Daphniapopulation. J Plankton Res 22:151–068
Huq A, Small EB, West PA, Huq MI, Rahman R, Colwell RR(1983) Ecological relationships between Vibrio choleraeand planktonic crustacean copepods. Appl Environ Micro-biol 45:275–283
Itoi S, Okamura T, Koyama Y, Sugita H (2006) Chitinolyticbacteria in the intestinal tract of Japanese coastal fishes.Can J Microbiol 52:1158–1163
Jacobsen T, Azam F (1984) Role of bacteria in copepod fecalpellet decomposition: colonization, growth rates and min-eralization. Bull Mar Sci 35:495–502
Jerde CW, Lasker R (1966) Moulting of euphausiid shrimp:shipboard observation. Limnol Oceanogr 11:120–124
Jeuniaux C (1971) Hemolymph-Arthropoda. In: Florkin M,Scheer BT (eds) Chemical zoology, Vol VI. AcademicPress, New York, NY, p 63–118
Jeuniaux C (1982) La chitine dans le regne animal. Bull SocZool Fr 107:363–386
Keller MD, Korjeff-Bellows W (1996) Physiological aspects ofthe production of dimethylsulfoniopropionate (DMSP) bymarine phytoplankton. In: Kiene RP, Visscher PT, KellerMD, Kirst GO (eds) Biological and environmental chem-istry of DMSP and related sulfonium compounds. PlenumPress, New York, NY, p 131–142
Keyhani NO, Roseman S (1999) Physiological aspects of chitincatabolism in marine bacteria. Biochim Biophys Acta 1473:108–122
King CH, Sanders RW, Shotts EB Jr, Porter KG (1991) Differ-ential survival of bacteria ingested by zooplankton from astratified eutrophic lake. Limnol Oceanogr 36:829–845
Kiørboe T, Grossart HP, Ploug H, Tang K (2002) Mechanismsand rates of bacterial colonization of sinking aggregates.Appl Environ Microbiol 68:3996–4006
Kirchman DL, White J (1999) Hydrolysis and mineralization ofchitin in the Delaware Estuary. Aquat Microb Ecol 18:187–196
Kirchner M (1995) Microbial colonization of copepod bodysurfaces and chitin degradation in the sea. Helgol Meere-sunters 49:201–212
Kirn TJ, Jude BA, Taylor RK (2005) A colonization factor linksVibrio cholerae environmental survival and human infec-tion. Nature 438:863–866
Kobari T, Ikeda T (2001) Ontogenetic vertical migration andlife cycle of Neocalanus plumchrus (Crustacea: Cope-poda) in the Oyashio region, with notes on regional varia-tions in body sizes. J Plankton Res 23:287–302
Komar PD, Morse AP, Small LF (1981) An analysis of sinkingrates of natural copepod and euphausiid fecal pellets.Limnol Oceanogr 26:172–180
Koropatnick TA, Engle JT, Apicella MA, Stabb EV, GoldmanWE, McFall-Ngai MJ (2004) Microbial factor-mediateddevelopment in a host-bacterial mutualism. Science 306:1186–1188
Kragh T, Sondergaard M, Borch NH (2006) The effect of zoo-plankton on the dynamics and molecular composition ofcarbohydrates during an experimental algal bloom. J Lim-nol 65:52–58
Lampert W (1989) The adaptive significance of diel verticalmigration of zooplankton. Funct Ecol 3:21–27
Lampitt RS, Noji T, von Bodungen B (1990) What happens tozooplankton faecal pellets? Implications for material flux.Mar Biol 104:15–23
Lawrence JR, Caldwell DE (1987) Behavior of bacterial streampopulations within the hydrodynamic boundary layersof surface microenvironments. Microb Ecol 14: 15–27
Lebrato M, Jones DOB (2009) Mass deposition event of Pyro-soma atlanticum carcasses off Ivory Coast (West Africa).Limnol Oceanogr 54:1197–1209
Ledyard KM, Dacey JWH (1996) Microbial cycling of DMSPand DMS in coastal and oligotrophic seawater. LimnolOceanogr 41:33–40
Marty DG (1993) Methanogenic bacteria in seawater. LimnolOceanogr 38:452–456
Meibom KL, Li XB, Nielsen AT, Wu CY, Roseman S, School-nik GK (2004) The Vibrio cholera chitin utilization pro-gram. Proc Natl Acad Sci USA 101:2524–2529
Michotey V, Bonin P (1997) Evidence for anaerobic bacterialprocesses in the water column: denitrification and dissim-ilatory nitrate ammonification in the northwesternMediterranean Sea. Mar Ecol Prog Ser 160:47–56
Mills CE (2001) Jellyfish blooms: Are populations increasingglobally in response to changing ocean conditions?Hydrobiologia 451:55–68
Møller EF (2005) Sloppy feeding in marine copepods: prey-size-dependent production of dissolved organic carbon.J Plankton Res 17:27–35
Møller EF, Thor P, Nielsen TG (2003) Production of DOC byCalanus finmarchicus, C. glacialis and C. hyperboreusthrough sloppy feeding and leakage from fecal pellets.Mar Ecol Prog Ser 262:185–191
Møller EF, Riemann L, Søndergaard M (2007) Bacteria associ-ated with copepods: abundance, activity and communitycomposition. Aquat Microb Ecol 47:99–106
Nagasawa S, Nemoto T (1988) Presence of bacteria in guts ofmarine crustaceans and on their fecal pellets. J PlanktonRes 10:559–564
Olsen AI, Olsen Y, Attarmadal Y, Chrisie K, Birkbeck TH,Skjermo J, Vadstein O (2000) Effects of short termfeeding of microalgae on the bacterial flora associatedwith juvenile Artemia franciscana. Aquaculture190:11–25
Oremland RS (1979) Methanogenic activity in plankton sam-ples and fish intestines: a mechanism for in situ methano-genesis in oceanic surface waters. Limnol Oceanogr24:1136–1141
Parsons TR, Takahashi M, Hargrave B (1977) Biologicaloceanographic processes, 2nd edn. Pergamon Press,Oxford
Pathak SP, Gopal K (2005) Occurrence of antibiotic and metalresistance in bacteria from organs of river fish. EnvironRes 98:100–103
Peduzzi P, Herndl GJ (1992) Zooplankton activity fueling themicrobial loop: differential growth response of bacteriafrom oligotrophic and eutrophic waters. Limnol Oceanogr37:1087–1092
Peter H, Sommaruga R (2008) An evaluation of methods tostudy the gut bacterial community composition of fresh-water zooplankton. J Plankton Res 30:997–1006
Pitt KA, Welsh DT, Condon RH (2009) Influence of jellyfishblooms on carbon, nitrogen and phosphorus cycling andplankton production. Hydrobiologia 616:133–149
Plante CJ, Shriver AG (1998) Patterns of differential digestionof bacteria in deposit feeders: a test of resource partition-ing. Mar Ecol Prog Ser 163:253–258
Plante CJ, Wilde SB (2001) Bacterial recolonization of deposit-feeder egesta: in situ regrowth or immigration? LimnolOceanogr 46:1171–1181
Pond DW, Harris RP, Brownlee C (1995) A microinjectiontechnique using a pH-sensitive dye to determine the gutpH of Calanus helgolandicus. Mar Biol 123:75–79
Poulicek M, Gaill F, Goffinet G (1998) Chitin biodegradationin marine environments. ACS Symp Ser 707:163–210
276

Tang et al.: Zooplankton–bacteria interactions
Proctor LM (1997) Nitrogen-fixing, photosynthetic, anaerobicbacteria associated with pelagic copepods. Aquat MicrobEcol 12:105–113
Pruzzo C, Crippa A, Bertone S, Pane L, Carli A (1996) Attach-ment of Vibrio alginolyticus to chitin mediated by chitin-binding proteins. Microbiology 142:2181–2186
Pruzzo C, Vezzulli L, Colwell RR (2008) Global impact of Vib-rio cholerae interactions with chitin. Environ Microbiol10:1400–1410
Purcell JE, Uye S, Lo WT (2007) Anthropogenic causes ofjellyfish blooms and their direct consequences for humans:a review. Mar Ecol Prog Ser 350:153–174
Ramaiah N, Hill RT, Chun J, Ravel J, Matte MH, Straube WL,Colwell RR (2000) Use of a chiA probe for detection ofchitinase genes in bacteria from the Chesapeake Bay.FEMS Microbiol Ecol 34:63–71
Rico-Mora R, Voltolina D (1995) Effects of bacterial isolates fromSkeletonema costatus cultures on the survival of Artemiafranciscana nauplii. J Invertebr Pathol 66: 203–204
Riemann L, Azam F (2002) Widespread N-acetyl-D-glu-cosamine uptake among pelagic marine bacteria and itsecological implications. Appl Environ Microbiol 68:5554–5562
Riemann L, Titelman J, Båmstedt U (2006) Links between jel-lyfish and microbes in a jellyfish dominated fjord. MarEcol Prog Ser 325:29–42
Schuett C, Doepke H (2009) Endobiotic bacteria and theirpathogenic potential in cnidarian tentacles. Helgol MarRes, doi 10.1007/s10152-009-0179-2
Selmi G (2001) Ectosymbiotic bacteria on ciliated cells of arotifer. Tissue Cell 33:258–261
Simon M, Grossart HP, Schweitzer B, Ploug H (2002) Micro-bial ecology of organic aggregates in aquatic ecosystems.Aquat Microb Ecol 28:175–211
Sochard MR, Wilson DF, Austin B, Colwell RR (1979) Bacteriaassociated with the surface and gut of marine copepods.Appl Environ Microbiol 37:750–759
Stief P, Poulsen M, Nielsen LP, Brix H, Schramm A (2009)Nitrous oxide emission by aquatic macrofauna. Proc NatlAcad Sci USA 106:4296–4300
Tamplin ML, Gauzens AL, Huq A, Sack D, Colwell RR (1990)Attachment of Vibrio cholera serogroup O1 to zooplank-ton and phytoplankton of Bangladesh waters. Appl Envi-ron Microbiol 56:1977–1980
Tang KW (2005) Copepods as microbial hotspots in the ocean:effects of host feeding activities on attached bacteria.Aquat Microb Ecol 38:31–40
Tang KW, Dam HG, Visscher PT, Fenn TD (1999) Dimethylsul-foniopropionate (DMSP) in marine copepods and its relationwith diets and salinity. Mar Ecol Prog Ser 179: 71–79
Tang KW, Visscher PT, Dam HG (2001) DMSP-consumingbacteria associated with the calanoid copepod Acartiatonsa (Dana). J Exp Mar Biol Ecol 256:185–198
Tang KW, Freund CS, Schweitzer CL (2006a) Occurrence ofcopepod carcasses in the lower Chesapeake Bay and theirdecomposition by ambient microbes. Estuar Coast ShelfSci 68:499–508
Tang KW, Hutalle KML, Grossart HP (2006b) Microbial abun-dance, composition and enzymatic activity during de-composition of copepod carcasses. Aquat Microb Ecol 45:219–227
Tang KW, Bickel SL, Dziallas C, Grossart HP (2009a) Micro-bial activities accompanying decomposition of cladoceranand copepod carcasses under different environmentalconditions. Aquat Microb Ecol 57:89–100
Tang K, Dziallas C, Hutalle-Schmelzer K, Grossart HP (2009b)Effects of food on bacterial community composition associ-ated with the copepod Acartia tonsa Dana. Biol Lett5:549–553
Thor P, Dam HG, Rogers DR (2003) Fate of organic carbonreleased from decomposing copepod fecal pellets in rela-tion to bacterial production and ectoenzyme activity.Aquat Microb Ecol 33:279–288
Tilly K, Elias AF, Errett J, Fischer E, Iyer R, Schwartz I, BonoJL, Rosa P (2001) Genetics and regulation of chitobioseutilization in Borrelia burgdorferi. J Bacteriol 183:5544–5553
Tinta T, Malej A, Kos M, Turk V (2010) Degradation of theAdriatic medusa Aurelia sp. by ambient bacteria. Hydro-biologia 645:179–191
Titelman J, Riemann L, Sørnes TA, Nilsen T, Griekspoor P,Båmstedt U (2006) Turnover of dead jellyfish: stimulationand retardation of microbial activity. Mar Ecol Prog Ser325:43–58
Turley CM, Mackie PJ (1995) Bacteria and cyanobacterial fluxto the deep NE Atlantic on sedimenting particles. Deep-Sea Res I 42:1453–1474
Turner JT (1979) Microbial attachment to copepod fecal pel-lets and its possible significance. Trans Am Microsc Soc98:131–135
Uye S (1985) Resting egg production as a life history strategyof marine planktonic copepods. Bull Mar Sci 37:440–449
Verschuere L, Rombaut G, Huys G, Dhont J, Sorgeloos P, Ver-straet W (1999) Microbial control of the culture of Artemiajuveniles through preemptive colonization by selectedbacterial strains. Appl Environ Microbiol 65:2527–2533
Verschuere L, Heang H, Criel G, Sorgeloos P, Verstraete W(2000) Selected bacterial strains protect Artemia spp. fromthe pathogenic effects of Vibrio proteolyticus CW8T2.Appl Environ Microbiol 66:1139–1146
Visscher PT, Diaz MR, Taylor BF (1992) Enumeration of bac-teria which cleave or demethylate dimethylsulfoniopropi-onate in the Caribbean Sea. Mar Ecol Prog Ser 89:293–296
Visser AW, Jónasdóttir SH (1999) Lipids, buoyancy and theseasonal vertical migration of Calanus finmarchicus. FishOceanogr 8:100–106
Wainwright PF, Mann KH (1982) Effect of antimicrobial sub-stances on the ability of the mysid shrimp Mysis stenolepisto digest cellulose. Mar Ecol Prog Ser 7:309–313
Weiss P, Schweitzer B, Amann R, Simon M (1996) Identifica-tion in situ and dynamics of bacteria on limnetic organicaggregates (lake snow). Appl Environ Microbiol 62:1998–2005
West EJ, Welsh DT, Pitt KA (2009) Influence of decomposingjellyfish on the sediment oxygen demand and nutrientdynamics. Hydrobiologia 616:151–160
Wurzbacher C, Bärlocher F, Grossart HP (2010) Fungi in lakeecosystems. Aquat Microb Ecol 59:125–149
Yu C, Lee AM, Bassler BL, Roseman S (1991) Chitin utilizationby marine bacteria — a physiological function for bacterialadhesion to immobilized carbohydrates. J Biol Chem 266:24260–24267
277
Submitted: February 15, 2010; Accepted: April 26, 2010 Proofs received from author(s): June 20, 2010