incorporating life histories and diet quality in stable isotope interpretations of crustacean...
TRANSCRIPT

Incorporating life histories and diet quality in stableisotope interpretations of crustacean zooplankton
MARC VENTURA* , † AND JORDI CATALAN*
*Limnology group (CSIC-UB), Centre for Advanced Studies of Blanes (CEAB), Spanish Research Council (CSIC), Girona,
Catalonia, Spain†National Environmental Research Institute, University of Aarhus, Silkeborg, Denmark
SUMMARY
1. Stable isotope studies have been extremely useful for improving general food web
descriptions due to their ability to simultaneously summarize complex trophic networks
and track the energy flow through them. However, when considering trophic relationships
involving only two or few species, application of general isotopic interpretations based on
average fractionation values may easily lead to misleading conclusions. In these cases a
more accurate consideration of the current processes involved in the isotopic fractionation
should be considered.
2. We investigated the trophic relationships of the crustacean zooplankton assemblage in
an alpine lake (Lake Redon, Pyrenees) by means of stable isotopes of carbon and nitrogen
and applied information on their life history and biochemical composition in the
interpretation.
3. The three species occurring in the lake had distinct isotopic signatures: the two copepod
species (the cyclopoid Cyclops abyssorum and the calanoid Diaptomus cyaneus) had higher
nitrogen isotopic composition than the cladoceran (Daphnia pulicaria), indicative of a higher
trophic position of the two copepods. Most intra-specific isotopic variability was
associated with growth, while the effect of metabolic turnover was negligible. The effects
of changes in the proportion of lipids was restricted to the adults of the two copepods.
4. Daphnia Juveniles showed ontogenetic shifts in their carbon, and nitrogen isotopic
composition. Cyclops copepodites only showed changes in carbon isotopic composition.
These isotopic shifts with changes in size were the result of: (i) the prevalence of
growth over metabolic turnover as the main factor for isotopic variability and (ii) feeding,
during the growth period, on isotopically depleted food in the case of Daphnia, and on
isotopically enriched food in the case of Cyclops.
5. The carbon isotopic variation in Cyclops juveniles could be explained by fitting an
isotopic growth model that considered that they fed entirely on Daphnia. However this was
not the case for nitrogen isotopic variability. Cyclops nitrogen isotopic composition
variation and the Cyclops to Daphnia nitrogen isotopic enrichment were closely correlated
to the quantity of Daphnia protein and to the dissimilarity in the essential amino acid
composition between the two species, which can be interpreted as an indication of
consumer nitrogen limitation.
Keywords: amino acids, carbon isotopes, fractionation, nitrogen isotopes, predator–prey
Correspondence: Marc Ventura, Limnology group (CSIC-UB), Centre for Advanced Studies of Blanes (CEAB), Spanish Research
Council (CSIC), Acces a la Cala Sant Francesc, 14. 17300-Blanes, Girona, Catalonia, Spain. E-mail: [email protected]
Freshwater Biology (2008) 53, 1453–1469 doi:10.1111/j.1365-2427.2008.01976.x
� 2008 The Authors, Journal compilation � 2008 Blackwell Publishing Ltd 1453

Introduction
Traditional food-web diagrams have been criticised as
being subjective constructs that are strongly biased by
our inability to observe the relevant taxonomic, spatial
and temporal variations in trophic interactions (Paine,
1988). The use of stable isotopes has improved food-
web descriptions due to their ability to simulta-
neously capture complex interactions and track
energy flow through ecological communities
(Peterson & Fry, 1987). The ratio of stable isotopes of
nitrogen (d15N) has been used to estimate trophic
positions because the d15N of a consumer is typically
enriched by 3–4& relative to its diet (Vander Zanden
& Rasmussen, 2001; Post, 2002; McCutchan et al.,
2003; Vanderklift & Ponsard, 2003). In contrast, the
ratio of carbon isotopes (d13C) changes only little as
carbon flows through food webs (Vander Zanden &
Rasmussen, 2001; Post, 2002; McCutchan et al., 2003;
Vanderklift & Ponsard, 2003).
Several assumptions are usually made when apply-
ing stable isotopes to field studies. These are based
on experimental studies and have been questioned
(Gannes, O’Brien & Martınez del Rio, 1997; Post, 2002).
For instance, the assumption of constancy in nitrogen
fractionation has frequently proved invalid (e.g. Webb,
Hedges & Simpson, 1998; Adams & Sterner, 2000;
Gaye-Siessegger et al., 2004), and the application of
constant average nitrogen fractionation values to
establish trophic positions should be applied with
caution and usually only when dealing with broad and
complex food webs with multiple trophic links and
many species (Post, 2002). In studies of trophic
relationships involving only two or few species,
potential factors of variability in isotopic fractionation
should be taken into account, including aspects of basic
biochemical composition, diet quality and life history.
The relative proportions of the main biochemical
compounds affect the isotopic composition of an
organism due to the distinct isotopic compositions of
proteins, lipids and chitin (Macko et al., 1990; Kling,
Fry & O’Brien, 1992). Changes in food quality can lead
to high nitrogen fractionation, equivalent to changes of
up to two trophic levels under the assumption of
constant 3–4& enrichment (Webb et al., 1998; Adams
& Sterner, 2000; Oelbermann & Scheu, 2002; Gaye-
Siessegger et al., 2004). The lack of a sufficient source of
amino acids in a given diet forces a higher nitro-
gen turnover in consumers and, therefore, a higher
isotopic discrimination (Hobson & Clark, 1992; Gan-
nes, Martınez del Rio & Koch, 1998). This deficiency
can either be due to the lack of total protein (Fantle
et al., 1999; Adams & Sterner, 2000; Gaye-Siessegger
et al., 2004) or to an amino acid imbalance between the
consumer and its diet (McClelland & Montoya, 2002).
Finally, depending on the isotopic turnover of each
organism, its isotopic signature will be related to
variable sources used throughout the life of the
organism (O’Reilly et al., 2002). Experimental studies
have shown that isotope turnover times may be linked
to metabolism, either related to maintenance or
growth (Hesslein, Hasllard & Ramlal, 1993; Ponsard
& Averbuch, 1999). The relative importance of both
isotopic incorporation routes differ between homeo-
therms and cold-water poikilotherms. For homeo-
therms Ponsard & Averbuch (1999) concluded that
turnover time was very quick due to a much higher
importance of maintenance metabolism over growth
incorporation. However, for poikilotherms living in
cold environments, growth has been shown to be of
much greater importance than maintenance metabo-
lism and the isotopic composition of poikilotherms
will therefore be a summary of their growing period
(Hesslein et al., 1993; Grey, 2000; Herzka & Holt, 2000).
Among freshwater zooplankton, copepods are
known for their trophic shift from the naupliar stage
to copepodites and adults, increasing their prey size
and switching feeding mode from grazing to hunting
(Dussart & Defaure, 1995; Santer, 1998). Copepod
predation on cladocerans can be a result of direct
predation of adults on small bodied Daphnia (Gliwicz
& Lampert, 1994; Gliwicz & Umana, 1994) or of
copepodite predation on the eggs by directly entering
into egg pouches of large bodied Daphnia species
(Gliwicz & Stibor, 1993; Hanazato & Dodson, 1995).
This latter predation mode is very relevant in alpine
lakes (Gliwicz & Boavida, 1996) and becomes a
mechanism leading to life cycle synchronization
(Gliwicz, Slusarczyk & Slusarczyk, 2001). Since cope-
pods have a relatively higher nitrogen and lower
phosphorus content than cladocerans (Ventura, 2006),
they are susceptible to nitrogen limitation. Therefore,
the study of copepods feeding on cladocerans is an
excellent case study for determining the extent to
which the isotopic composition of natural communi-
ties is affected by food quality changes.
Lake Redon, is an oligotrophic lake with
average microbial biomass ratios of 10 : 2 : 2 : 1 for
1454 M. Ventura and J. Catalan
� 2008 The Authors, Journal compilation � 2008 Blackwell Publishing Ltd, Freshwater Biology, 53, 1453–1469
phytoplankton : bacteria : heterotrophic nanoflage-
lates : ciliates during the ice-free period (Felip et al.,
1999). The lake zooplankton assemblage has only
three crustacean species, one cladoceran (Daphnia
pulicaria Forbes), one cyclopoid copepod (Cyclops
abyssorum Sars) and the calanoid copepod Diaptomus
cyaneus Gurney. Rotifers are usually less abundant
than crustaceans (the ratio between Daphnia and
rotifer biomass is approximately 10 : 1 during the
ice-free period; Camarero et al., 1999). Daphnia and
Cyclops are present in the plankton all year round,
while Diaptomus appears only during summer
(Ventura & Catalan, 2005). The feeding habits of the
first two species are well described, while those of the
calanoid copepod are unknown. D. pulicaria is a
generalist primary consumer (grazer), and C. abysso-
rum has been described as a secondary consumer
(carnivore) preying on Daphnia whenever it can
(Fryer, 1957; Schindler, 1971; Vandenbosch & Santer,
1993). It has been shown, however, that C. abyssorum
may survive on a strictly algal diet (Whitehouse &
Lewis, 1973; Hopp & Maier, 2005). Studies using
stable isotopes have shown C. abyssorum to exhibit a
more substantially enriched nitrogen isotopic ratio
compared to other copepods, such as Cyclops vicinus
Uljanin (Santer, Sommerwerk & Grey, 2006). In the
case of Lake Redon, as in other alpine lakes (Gliwicz &
Boavida, 1996; Gliwicz et al., 2001), it is to be expected
that the main diet of Cyclops will consist of Daphnia
due to the lower availability of other prey items.
The specific aims of this study were (i) to describe
the seasonal isotopic composition of the three species
and their trophic relationships; (ii) to estimate the
relative influence of lipids and chitin in the isotopic
variability of the three species; (iii) to determine
whether the isotopic composition of Cyclops could be
explained based on the isotopic composition of Daph-
nia and (iv) to examine whether the variability of the
isotopic distance between Cyclops and Daphnia, espe-
cially for nitrogen isotope, changed depending on the
protein quantity or quality of its putative food.
Methods
Study site
Lake Redon (formerly Lake Redo) is an oligotrophic
glacial cirque lake located at 2240 m.a.s.l. in the central
Pyrenees (42�38¢N, 0�46¢E). It is relatively large (24 ha)
and one of the deepest lakes (73 m) in the Pyrenees. It is
dimictic, covered by ice during half of the year, usually
from mid-December until late May or the beginning of
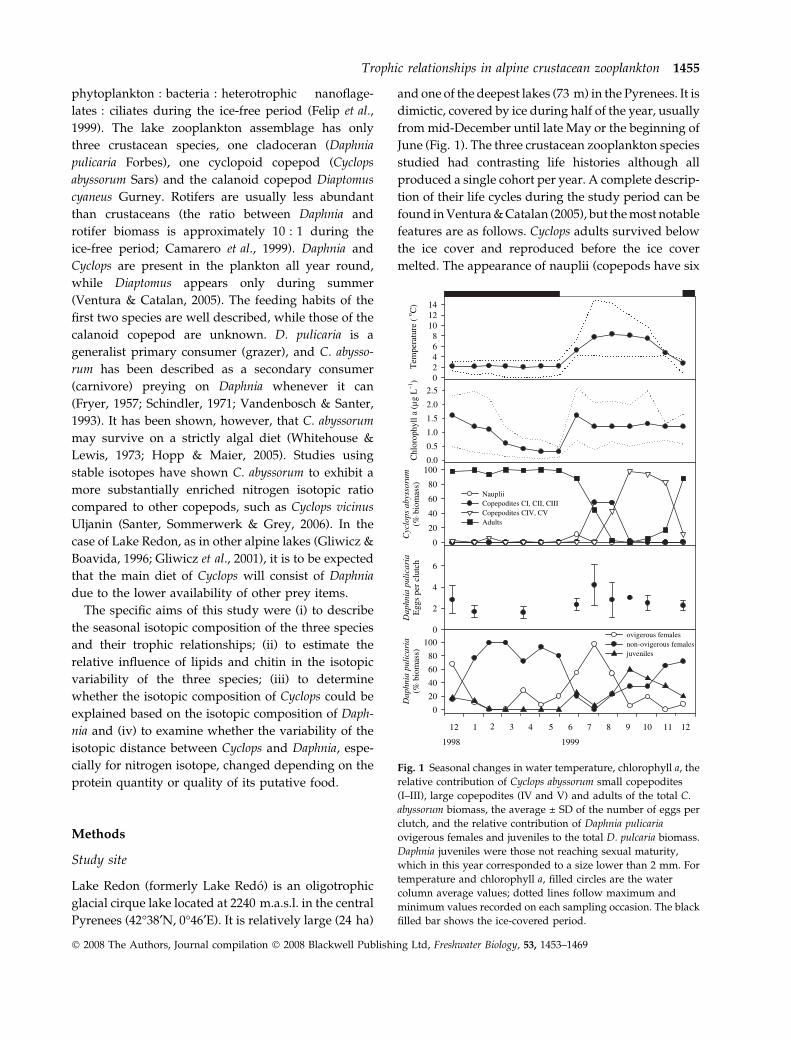
June (Fig. 1). The three crustacean zooplankton species
studied had contrasting life histories although all
produced a single cohort per year. A complete descrip-
tion of their life cycles during the study period can be
found in Ventura & Catalan (2005), but the most notable
features are as follows. Cyclops adults survived below
the ice cover and reproduced before the ice cover
melted. The appearance of nauplii (copepods have six
02468
101214
Tem
pera
ture
(o
)C
1999
paD
painh
uracil
aim oib
%(a
)ss
0
20
40
60
80
100ovigerous femalesnon-ovigerous femalesjuveniles
1998
Cyc
lrossyba
spoum
%(moib
)ssa
0
20
40
60
80
100
NaupliiCopepodites CI, CII, CIIICopepodites CIV, CVAdults
rolhC
ollyhp
(a
µL
g–1
)
0.0
0.5
1.0
1.5
2.0
2.5
12 1 2 3 4 5 6 7 8 9 10 11 12
hpaD
upain
racilai
tulcrep
sggE
hc
0
2
4
6
Fig. 1 Seasonal changes in water temperature, chlorophyll a, the
relative contribution of Cyclops abyssorum small copepodites
(I–III), large copepodites (IV and V) and adults of the total C.
abyssorum biomass, the average ± SD of the number of eggs per
clutch, and the relative contribution of Daphnia pulicaria
ovigerous females and juveniles to the total D. pulcaria biomass.
Daphnia juveniles were those not reaching sexual maturity,
which in this year corresponded to a size lower than 2 mm. For
temperature and chlorophyll a, filled circles are the water
column average values; dotted lines follow maximum and
minimum values recorded on each sampling occasion. The black
filled bar shows the ice-covered period.
Trophic relationships in alpine crustacean zooplankton 1455
� 2008 The Authors, Journal compilation � 2008 Blackwell Publishing Ltd, Freshwater Biology, 53, 1453–1469

naupliar, and five copepodite stages before adults
develop) coincided with the spring production maxi-
mum. Copepodites CI–CIII, the stages described to be
sufficiently small to enter a Daphnia egg pouch (Gliwicz
& Stibor, 1993; Gliwicz & Lampert, 1994; Gliwicz &
Boavida, 1996), dominated Cyclops biomass during July
and August (Fig. 1). Most Daphnia adult females also
survived below the ice cover, postponing reproduction
until the ice-free period (Fig. 1). Over-wintering
females (termed ‘first cohort females’ in our study)
were therefore clearly distinguishable from the juve-
niles born during the ice free period (termed ‘second
cohort females’). Due to their long life span these first
generation Daphnia females were all longer than
2.25 mm (Ventura & Catalan, 2005), the minimum size
allowing copepodites to enter their egg pouch (Gliwicz
& Lampert, 1994). The main reproductive period in
which most females had eggs occurred after the spring
overturn from June to August, with a peak in July when
all females carried eggs. In contrast, the appearance of
Daphnia juveniles did not occur until September (Fig. 1)
after the CIII copepodites had become CIV, suggesting
that this delay could have been due to copepodite
predation. A second indication of copepodite predation
on Daphnia eggs was the increased variability in the
number of eggs per female (Gliwicz & Lampert, 1994),
which was higher during the 2 months when the small
copepodites were present in the lake (Fig. 1). Further-
more, Daphnia females were discovered with copepo-
dites within their egg pouches in the July and August
samples. From September onwards, due to the larger
size of copepodites, predation on Daphnia was probably
restricted to direct predation on smaller Daphnia juve-
niles, which were found until November (Ventura &
Catalan, 2005). Coinciding with the end of the autumn
overturn and the start of the ice cover, few adult males
appeared and sporadic ephippia were produced.
Diaptomus was the only strictly diapausing species,
completing its life cycle within 3 months. In Lake
Redon, Diaptomus emergence also coincided with the
spring production maximum, and during their pres-
ence in the plankton of the lake they dominated the
zooplankton biomass (Ventura & Catalan, 2005).
Sample collection and preparation
The lake was sampled on 14 dates from December
1998, just after the lake was completely ice-covered,
until December 1999 when it was ice-covered again.
Samples were collected at the deepest point of the lake
either by drilling through the ice or from a platform
anchored all summer at the same spot. Zooplankton
samples were collected by vertical hauls from 65 m to
the surface with a 200 lm net. Sampled individuals
were kept alive and transported cold (4 �C) until they
were frozen ()20 �C) in the laboratory within a few
hours of collection. After thawing the samples, from
10 to several hundreds of individuals, depending on
the weight of each species and stage, were quickly
sorted under a dissecting microscope and were either
placed into pre-weighted tin capsules for stable
isotope analysis or in Teflon capsules for analysis of
total amino acids, chitin and lipids. At least three
combined sample replicates were analysed for each
species and stage for each sampling date, with the
exception of a few cases where sample material did
not suffice. Individuals were kept cold (<4 �C) during
the sorting process which lasted a few hours for all
samples. Dry weight (DW) was determined for all
samples after drying at 60 �C for 24 h and weighing
on a high precision microbalance (Ohaus Analytical
Plus, AP250D-0; Ohaus corporation, Florham park,
NJ, U.S.A.). Body lengths were measured in each
sample under an Olympus inverted microscope
(Daphnia from the upper edge of the head to the base
of tail spine; copepods from the anterior end of the
cephalothorax to the posterior end of the furca).
Chemical analyses
Dried samples for stable isotope analysis were packed
into tin capsules with vanadium pentaoxide as cata-
lyser to assure complete combustion. Samples were
analysed in a Delta C Finningan MAT mass spec-
trometer (Thermo Fisher Scientific, Inc., Waltham,
MA, U.S.A.) coupled online with a Carlo Erba CHNS
elemental analyzer (CE Instruments, Wigan, U.K.)
via a Finnigan conflo 2 interface. Specific stan-
dards were used for calibrating the isotopic signal:
sucrose (IAEA CH6), polyethilene (IAEA CH7) and
graphite (IAEA-USGS 24) for carbon, and ammonium
sulfate (IAEA-USGS 25, IAEA-N1 and IAEA-N2) and
potassium nitrate (IAEA-NO3) for nitrogen (Gonfian-
tini, 1978). Complete batches of all standards were run
at the beginning and at the end of each analytical
session, and IAEA CH6 and CH7, and IAEA-N1 and
IAEA-NO3 were run for every 12 samples to control
for linearity. Special care was taken with respect to
1456 M. Ventura and J. Catalan
� 2008 The Authors, Journal compilation � 2008 Blackwell Publishing Ltd, Freshwater Biology, 53, 1453–1469

weighing the samples and the standards to ensure
similar amplitudes. Results are reported using atmo-
spheric nitrogen and PeeDee belemnite carbonate as
references. Reproducibility was better than 0.1& and
0.3& for d13C and d15N respectively.
Protein content was measured as the sum of
individual amino acids. The amino acid analysis also
allowed for the combined determination of chitin,
since N-acetylglucosamine, the molecular constituent
of chitin, is an amino-sugar that after hydrolysis
appears in the aminogram as glucosamine. Total
amino acid samples were vacuum sealed and hydro-
lysed with HCl 6N at 112 �C for 16 h. An internal
norleucine standard was included in every sample
prior to hydrolysis to increase reproducibility. The
analysis was performed on a Biochrom20 (Amersham-
Pharmacia Biotech, Roosendaal, the Netherlands)
ion-exchange amino acid auto analyser following the
ninhidrine method of Spackman, Stein & Moore
(1958). A standard solution of 20 amino acids and
glucosamine was run for every 10 samples.
Total lipids were quantified gravimetrically.
Between 0.3 and 0.6 mg of dried zooplankton were
placed in a dichlormethane : methanol (2 : 1, v ⁄v)
solution (Folch, Lees & Sloane Stanley, 1957) and
sonicated for 30 min in an ultra-sound bath for lipid
extraction. The non-lipidic remains were then col-
lected on pre-weighted GF ⁄F Whatman filters, which
were dried and re-weighted. Lipid content was
determined by weight difference.
Biochemical composition influence on stable isotope
composition
To determine the possible sources of isotope variation,
we analysed the concentration of lipids and chitin for
each species. Lipids are known to have a more
depleted signature than whole-body carbon (DeNiro
& Epstein, 1977), while chitin has a very depleted
signature compared to whole-body nitrogen (Macko
et al., 1990). Isotopic ratios of the lipid and chitin free
fractions were calculated following:
d13CLF ¼ d13CSA�ðMLP � fLPÞ and
d15NCTF ¼ d15NSA�ðMCT � fCTÞ
where LF stands for lipid free fraction, SA refers to the
isotopic composition of the sample, MLP and MCT are
the mass fractions of lipids and chitin, and ƒLP and ƒCT
are the isotopic fractionation in lipid and chitin
compartments respectively. In this study we assumed
the fractionation in zooplankton lipid inhabiting a
similar temperature regime (ƒLP = )3; Kling et al.,
1992) and a fractionation in chitin of ƒCT = )9 (Macko
et al., 1990).
Dissimilarity index
Average Euclidean distance (Djk) was used to estimate
the dissimilarity between the amino acid composi-
tions of Cyclops and Daphnia.
Dij ¼ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiXn
i¼1
Xij � Xik
� �2
s
where Xij and Xik are the percentages of the amino
acid i of Daphnia (j) or Cyclops (k), and n is the number
of amino acids considered. The index was calculated
with all the amino acids or separately for the essential
amino acids (lysine, phenylalanine + tyrosine,
leucine, valine, threonine, isoleucine, histidine, cyste-
ine + methionine) and non-essential amino acids
(glutamic acid, aspartic acid, alanine, arginine, gly-
cine, proline and serine).
Results
Intra-specific and gender differences
The three planktonic crustacean species inhabiting
Lake Redon differed in their mean stable isotope
composition of C and N (Table 1), the differences
being significant in one-way repeated measures-
ANOVAANOVA tests for d13C (F2,77 = 21.5, P < 0.001) and
d15N (F2,77 = 727.9, P < 0.001). To check for differences
between species pairs we conducted a Tukey’s post hoc
test. While the d13C signature of Cyclops was signif-
icantly different from Daphnia (P = 0.012) and Diapto-
mus (P = 0.001), there were non-significant differences
between Daphnia and Diaptomus (P = 0.212). The d15N
signature differed for all three species (P < 0.001).
Daphnia males only appeared occasionally and then
in very low densities, while copepod males and
females occurred in similar numbers when present
in the lake. We analysed males and females of the two
copepod species to determine whether there were
gender differences in isotopic signatures. Cyclops
males appeared only in January 1999, while Cyclops
Trophic relationships in alpine crustacean zooplankton 1457
� 2008 The Authors, Journal compilation � 2008 Blackwell Publishing Ltd, Freshwater Biology, 53, 1453–1469
females inhabited the lake from December 1998 to July
1999. As for Diaptomus, both sexes were present
during the same period. To avoid potential seasonal
differences in the diet of females, we compared the
isotopic composition of males and females of both
species when co-existing. Males and females of the
two species showed the same d15N values (Table 1),
one-way ANOVAANOVA showing no significant gender dif-
ferences either in Diaptomus (F1,10 = 1.99, P = 0.189) or
in Cyclops (F1,4 = 0.54, P = 0.502). Diaptomus presented
the same carbon isotopic values for both sexes
(F1,10 = 0.28, P = 0.611). However, there were gender
differences in the d13C values of Cyclops (F1,4 = 80.63,
P < 0.001), females being slightly more depleted
(Table 1).
Seasonal variability and trophic position as inferred
from stable isotope data
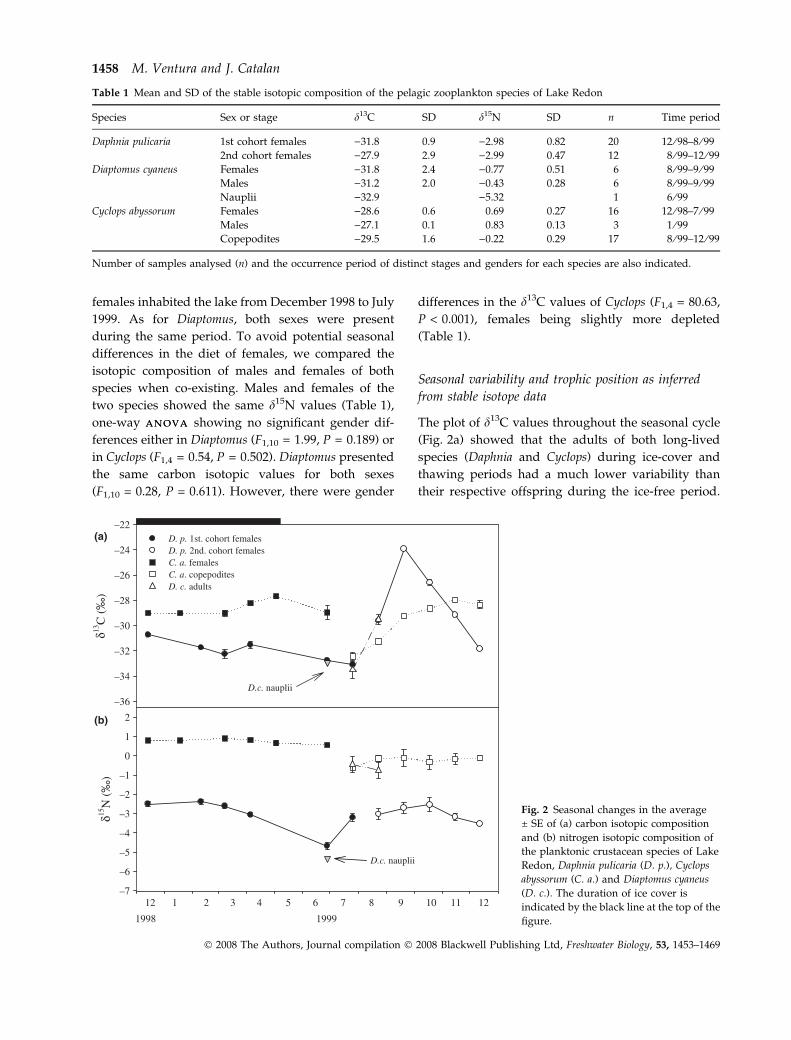
The plot of d13C values throughout the seasonal cycle
(Fig. 2a) showed that the adults of both long-lived
species (Daphnia and Cyclops) during ice-cover and
thawing periods had a much lower variability than
their respective offspring during the ice-free period.
Table 1 Mean and SD of the stable isotopic composition of the pelagic zooplankton species of Lake Redon
Species Sex or stage d13C SD d15N SD n Time period
Daphnia pulicaria 1st cohort females )31.8 0.9 )2.98 0.82 20 12 ⁄ 98–8 ⁄ 99
2nd cohort females )27.9 2.9 )2.99 0.47 12 8 ⁄ 99–12 ⁄ 99
Diaptomus cyaneus Females )31.8 2.4 )0.77 0.51 6 8 ⁄ 99–9 ⁄ 99
Males )31.2 2.0 )0.43 0.28 6 8 ⁄ 99–9 ⁄ 99
Nauplii )32.9 )5.32 1 6 ⁄ 99
Cyclops abyssorum Females )28.6 0.6 0.69 0.27 16 12 ⁄ 98–7 ⁄ 99
Males )27.1 0.1 0.83 0.13 3 1 ⁄ 99
Copepodites )29.5 1.6 )0.22 0.29 17 8 ⁄ 99–12 ⁄ 99
Number of samples analysed (n) and the occurrence period of distinct stages and genders for each species are also indicated.
D.c. n a uplii
δ 3 1
) ‰
(
C
–36
–34
–32
–30
–28
–26
–24
–22
δ 5 1
) ‰
(
N
–7
–6
–5
–4
–3
–2
–1
0
1
2
D.c. na uplii
(a)
(b)
1 2 1 2 3 4 5 6 7 8 9 1 0 1 1 1 2
1998 1999
D. p . 1st. coh o rt f e m a les D. p . 2nd . coho r t f e m a les C. a . females C. a . copep odites D. c . a dults
Fig. 2 Seasonal changes in the average
± SE of (a) carbon isotopic composition
and (b) nitrogen isotopic composition of
the planktonic crustacean species of Lake
Redon, Daphnia pulicaria (D. p.), Cyclops
abyssorum (C. a.) and Diaptomus cyaneus
(D. c.). The duration of ice cover is
indicated by the black line at the top of the
figure.
1458 M. Ventura and J. Catalan
� 2008 The Authors, Journal compilation � 2008 Blackwell Publishing Ltd, Freshwater Biology, 53, 1453–1469
The primary consumer Daphnia exhibited the highest
d13C variation. The adults of the calanoid copepod
Diaptomus changed their d13C from July to August
matching the d13C values of Daphnia. Diaptomus
nauplii also had an isotopic carbon signature similar
to that of Daphnia during their month of co-existence.
The seasonal changes in d15N of the different
species (Fig. 2b) showed a lower variability than
d13C, fluctuations in Daphnia being larger than in
Cyclops. The d15N values of Diaptomus adults were
similar to those of Cyclops copepodites and those of
Diaptomus nauplii were close to those of Daphnia
during the period they co-existed in the lake.
To describe the trophic positions of the two cope-
pod species of Lake Redon, we used the Daphnia
signature as a reference, since due to its unselective
feeding mode it constitutes a good estimate for the
base of the food chain (Vander Zanden & Rasmussen,
1999; Post, 2002; Matthews & Mazumder, 2003). The
average difference in isotopic nitrogen ratios (Dd15N)
between Cyclops and Daphnia was 3.5& (5.3–2.2&),
and 2.7& between Diaptomus and Daphnia (3.0–2.4&)
(Fig. 2b). Thus, Cyclops adults were at the top of the
zooplankton food chain and could potentially be
feeding on Daphnia. Diaptomus adults were almost
one trophic level above Daphnia, but were slightly
more depleted than Cyclops.
The Dd13C between Cyclops and Daphnia was 1.4&
on average but varied widely ()5.3& to +4.5&)
(Fig. 2a). The Dd13C between Diaptomus and Daphnia
was 0.3&. Carbon fractionation between consecutive
trophic levels is small (c. 0.4&: Post, 2002; Vanderklift
& Ponsard, 2003; McCutchan et al., 2003); therefore,
according to the Dd13C results, one would estimate
that Diaptomus could actually be feeding on Daphnia,
whereas Cyclops was not, contradicting what was
indicated by Dd15N.
Influence of biochemical composition on stable isotope
signatures
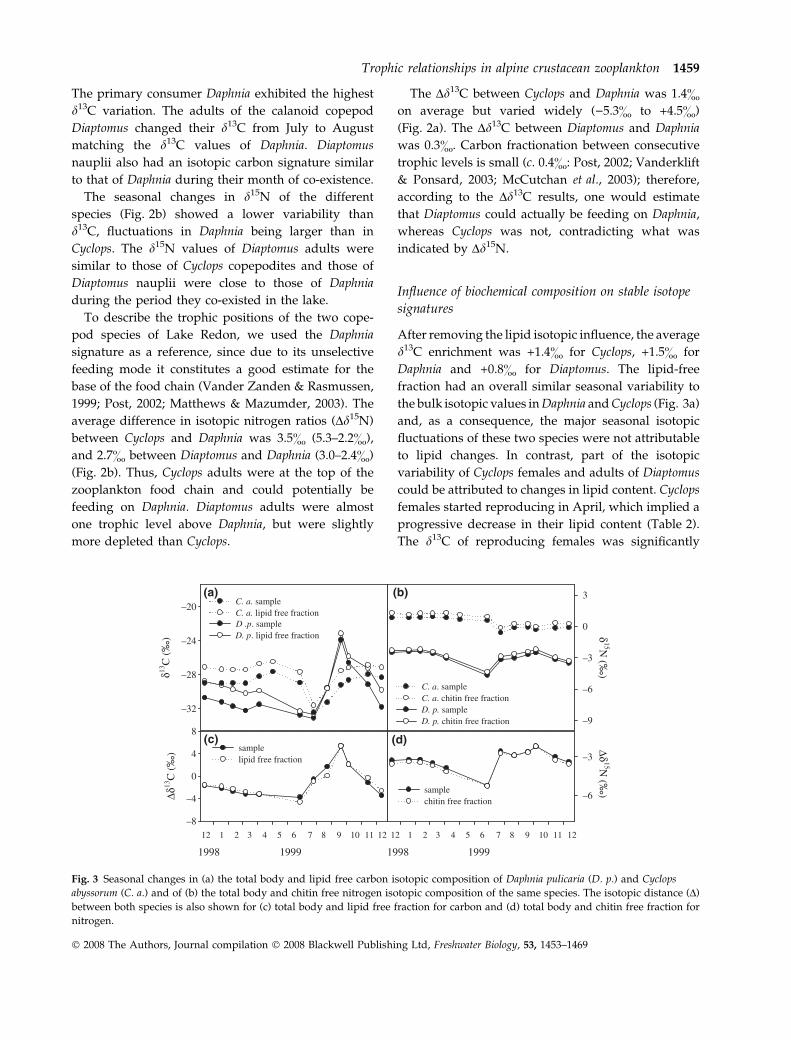
After removing the lipid isotopic influence, the average
d13C enrichment was +1.4& for Cyclops, +1.5& for
Daphnia and +0.8& for Diaptomus. The lipid-free
fraction had an overall similar seasonal variability to
the bulk isotopic values in Daphnia and Cyclops (Fig. 3a)
and, as a consequence, the major seasonal isotopic
fluctuations of these two species were not attributable
to lipid changes. In contrast, part of the isotopic
variability of Cyclops females and adults of Diaptomus
could be attributed to changes in lipid content. Cyclops
females started reproducing in April, which implied a
progressive decrease in their lipid content (Table 2).
The d13C of reproducing females was significantly
δ Δ 3 1
) ‰
(
C
–8
–4
0
4
8
sam p le lipid f r ee f r action
δ 5 1
) ‰
(
N
–9
–6
–3
0
3
C. a . sam p le C. a . chitin f r ee f r action D. p. sam p le D. p. ch itin f r ee f r action
δ Δ
5 1
‰
( N
) –6
–3
sam p le chitin f r ee f r action
199 9
δ 3 1
) ‰
(
C
–32
–28
–24
–20
D . p . sam p le D. p . lipid f r ee f r actio n
C. a. sa m p l e C. a. lipid f r ee f r action
(a) (b)
(c) (d)
12 1 2 3 4 5 6 7 8 9 1 0 1 1 1 2
1 998 19 99
12 1 2 3 4 5 6 7 8 9 1 0 1 1 1 2
199 8
Fig. 3 Seasonal changes in (a) the total body and lipid free carbon isotopic composition of Daphnia pulicaria (D. p.) and Cyclops
abyssorum (C. a.) and of (b) the total body and chitin free nitrogen isotopic composition of the same species. The isotopic distance (D)
between both species is also shown for (c) total body and lipid free fraction for carbon and (d) total body and chitin free fraction for
nitrogen.
Trophic relationships in alpine crustacean zooplankton 1459
� 2008 The Authors, Journal compilation � 2008 Blackwell Publishing Ltd, Freshwater Biology, 53, 1453–1469
higher than that of non-reproducing females (Table 2),
but this difference disappeared after correcting for the
lipid lost during the reproductive period (Table 2). The
total lipid content of Diaptomus adults also decreased
from 45.5% of DW to only 5.5% from July to August
due to reproductive investment (Table 2). This
decrease explained 30% of the isotopic change. Non-
reproducing Daphnia females had higher d13C than
reproducing females in both the bulk and lipid-
corrected d13C (Table 2) and, therefore, carbon isotopic
differences between the two cohorts were not attribut-
able to the large difference in lipid content between
them. Lipid effects on Dd13C were almost negligible
between Daphnia and Cyclops (Fig. 3c) and between
Daphnia and Diaptomus (data not shown).
For all species, subtraction of the chitin fraction
resulted in a slight increase (0.3–0.5&) in d15N
(Table 2). The chitin composition was constant for
the three species, and correction for chitin changes
therefore had only a small effect on the seasonal
variability of the nitrogen isotope (Fig. 3b; Diaptomus
data not shown). There were non-significant differ-
ences between non-reproducing and reproducing
females of Cyclops and adults of Diaptomus. In
contrast, Daphnia first cohort non-reproducing females
had a higher d15N than reproducing females in both
bulk and chitin free composition (Table 2). Similarly,
Dd15N values among species remained unaltered for
the three species when corrected for chitin content
(Fig. 3d).
Ontogenetic isotopic shifts
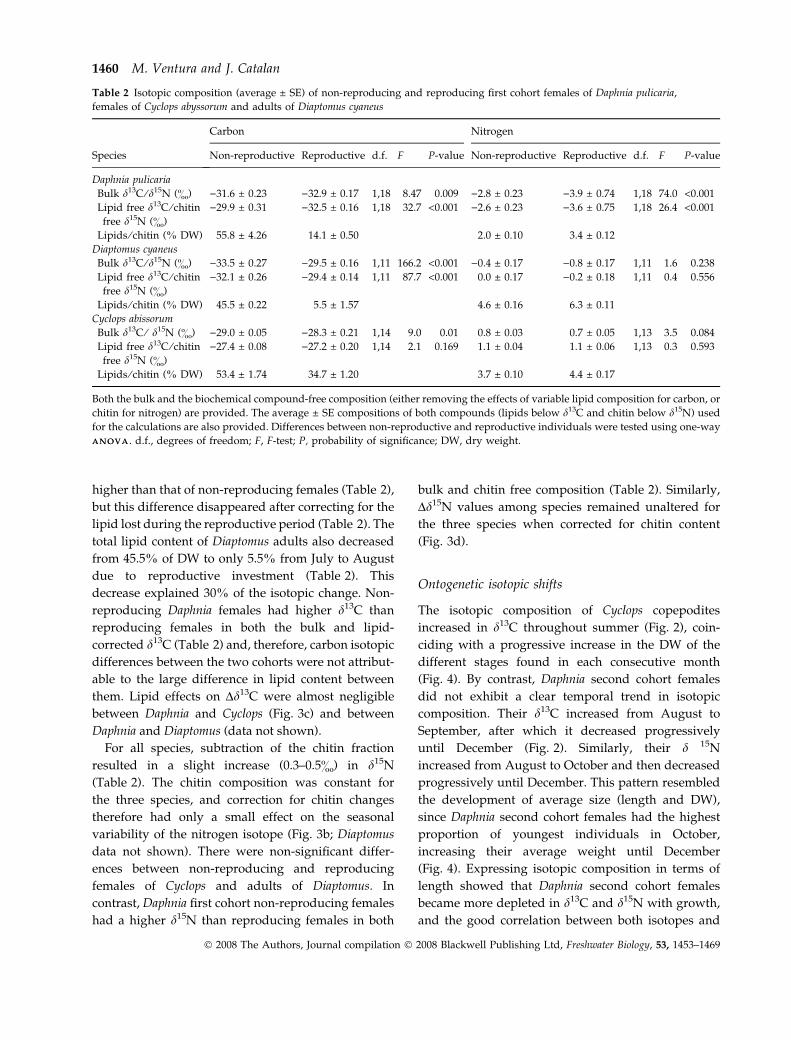
The isotopic composition of Cyclops copepodites
increased in d13C throughout summer (Fig. 2), coin-
ciding with a progressive increase in the DW of the
different stages found in each consecutive month
(Fig. 4). By contrast, Daphnia second cohort females
did not exhibit a clear temporal trend in isotopic
composition. Their d13C increased from August to
September, after which it decreased progressively
until December (Fig. 2). Similarly, their d 15N
increased from August to October and then decreased
progressively until December. This pattern resembled
the development of average size (length and DW),
since Daphnia second cohort females had the highest
proportion of youngest individuals in October,
increasing their average weight until December
(Fig. 4). Expressing isotopic composition in terms of
length showed that Daphnia second cohort females
became more depleted in d13C and d15N with growth,
and the good correlation between both isotopes and
Table 2 Isotopic composition (average ± SE) of non-reproducing and reproducing first cohort females of Daphnia pulicaria,
females of Cyclops abyssorum and adults of Diaptomus cyaneus
Species
Carbon Nitrogen
Non-reproductive Reproductive d.f. F P-value Non-reproductive Reproductive d.f. F P-value
Daphnia pulicaria
Bulk d13C ⁄d15N (&) )31.6 ± 0.23 )32.9 ± 0.17 1,18 8.47 0.009 )2.8 ± 0.23 )3.9 ± 0.74 1,18 74.0 <0.001
Lipid free d13C ⁄ chitin
free d15N (&)
)29.9 ± 0.31 )32.5 ± 0.16 1,18 32.7 <0.001 )2.6 ± 0.23 )3.6 ± 0.75 1,18 26.4 <0.001
Lipids ⁄ chitin (% DW) 55.8 ± 4.26 14.1 ± 0.50 2.0 ± 0.10 3.4 ± 0.12
Diaptomus cyaneus
Bulk d13C ⁄d15N (&) )33.5 ± 0.27 )29.5 ± 0.16 1,11 166.2 <0.001 )0.4 ± 0.17 )0.8 ± 0.17 1,11 1.6 0.238
Lipid free d13C ⁄ chitin
free d15N (&)
)32.1 ± 0.26 )29.4 ± 0.14 1,11 87.7 <0.001 0.0 ± 0.17 )0.2 ± 0.18 1,11 0.4 0.556
Lipids ⁄ chitin (% DW) 45.5 ± 0.22 5.5 ± 1.57 4.6 ± 0.16 6.3 ± 0.11
Cyclops abissorum
Bulk d13C ⁄ d15N (&) )29.0 ± 0.05 )28.3 ± 0.21 1,14 9.0 0.01 0.8 ± 0.03 0.7 ± 0.05 1,13 3.5 0.084
Lipid free d13C ⁄ chitin
free d15N (&)
)27.4 ± 0.08 )27.2 ± 0.20 1,14 2.1 0.169 1.1 ± 0.04 1.1 ± 0.06 1,13 0.3 0.593
Lipids ⁄ chitin (% DW) 53.4 ± 1.74 34.7 ± 1.20 3.7 ± 0.10 4.4 ± 0.17
Both the bulk and the biochemical compound-free composition (either removing the effects of variable lipid composition for carbon, or
chitin for nitrogen) are provided. The average ± SE compositions of both compounds (lipids below d13C and chitin below d15N) used
for the calculations are also provided. Differences between non-reproductive and reproductive individuals were tested using one-way
A N O V AA N O V A. d.f., degrees of freedom; F, F-test; P, probability of significance; DW, dry weight.
1460 M. Ventura and J. Catalan
� 2008 The Authors, Journal compilation � 2008 Blackwell Publishing Ltd, Freshwater Biology, 53, 1453–1469
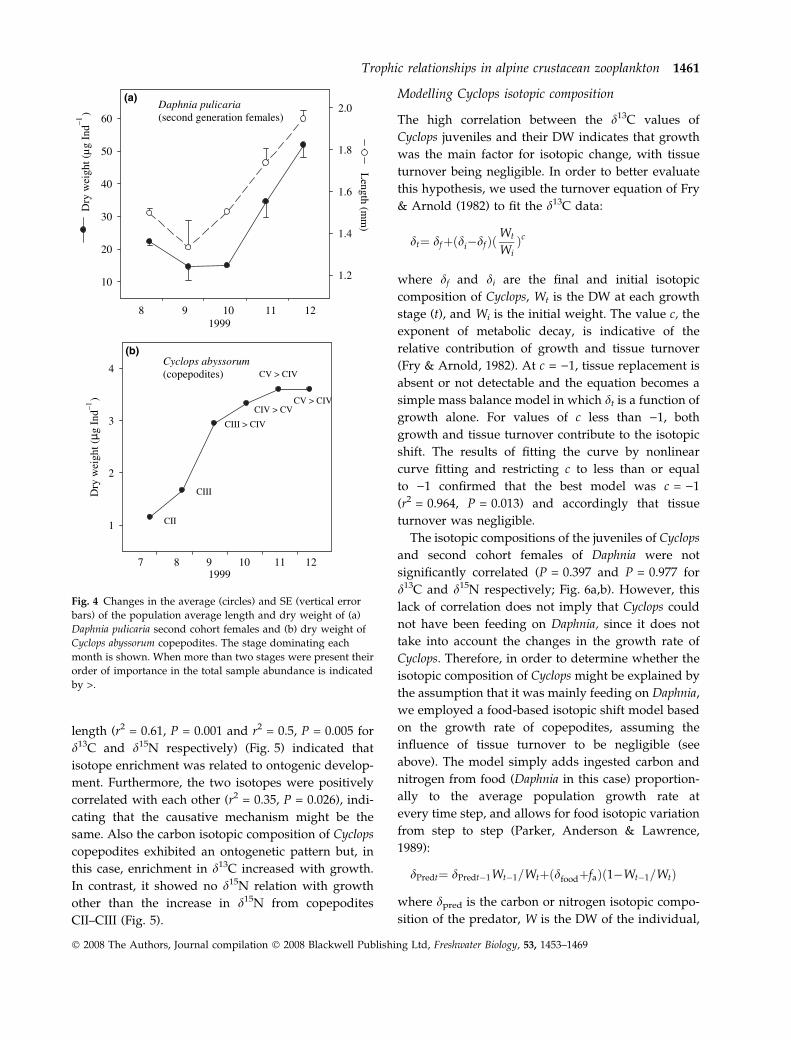
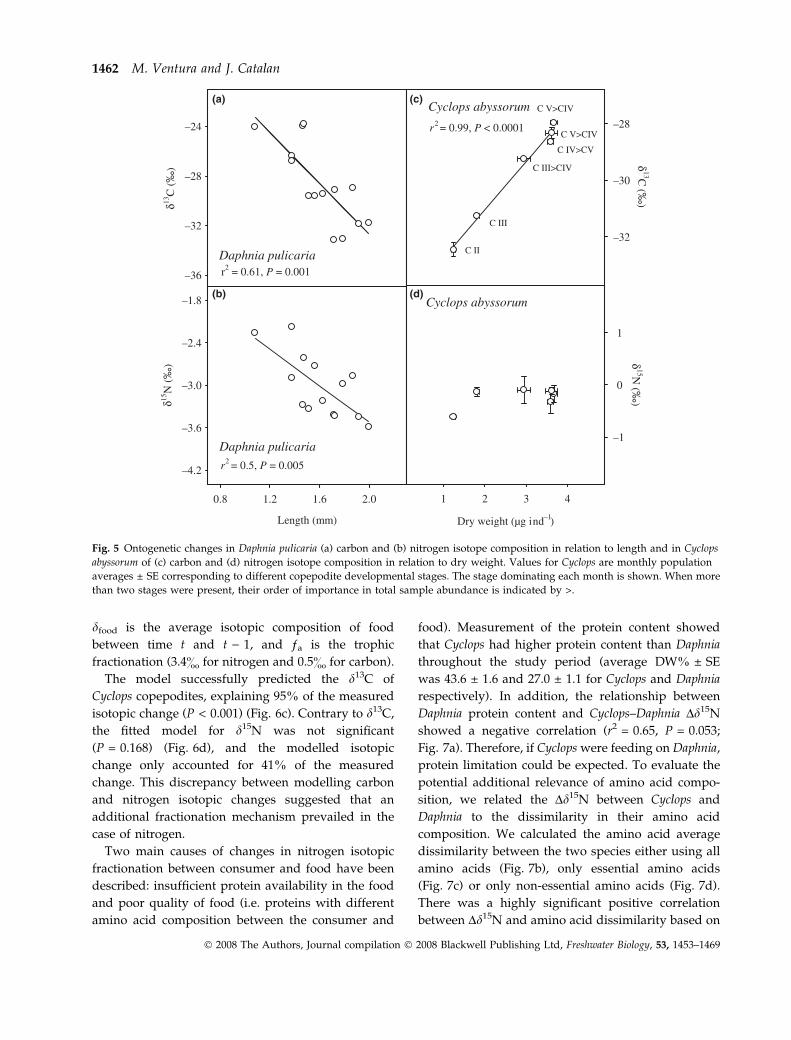
length (r2 = 0.61, P = 0.001 and r2 = 0.5, P = 0.005 for
d13C and d15N respectively) (Fig. 5) indicated that
isotope enrichment was related to ontogenic develop-
ment. Furthermore, the two isotopes were positively
correlated with each other (r2 = 0.35, P = 0.026), indi-
cating that the causative mechanism might be the
same. Also the carbon isotopic composition of Cyclops
copepodites exhibited an ontogenetic pattern but, in
this case, enrichment in d13C increased with growth.
In contrast, it showed no d15N relation with growth
other than the increase in d15N from copepodites
CII–CIII (Fig. 5).
Modelling Cyclops isotopic composition
The high correlation between the d13C values of
Cyclops juveniles and their DW indicates that growth
was the main factor for isotopic change, with tissue
turnover being negligible. In order to better evaluate
this hypothesis, we used the turnover equation of Fry
& Arnold (1982) to fit the d13C data:
dt¼ dfþðdi�dfÞðWt
WiÞc
where df and di are the final and initial isotopic
composition of Cyclops, Wt is the DW at each growth
stage (t), and Wi is the initial weight. The value c, the
exponent of metabolic decay, is indicative of the
relative contribution of growth and tissue turnover
(Fry & Arnold, 1982). At c = )1, tissue replacement is
absent or not detectable and the equation becomes a
simple mass balance model in which dt is a function of
growth alone. For values of c less than )1, both
growth and tissue turnover contribute to the isotopic
shift. The results of fitting the curve by nonlinear
curve fitting and restricting c to less than or equal
to )1 confirmed that the best model was c = )1
(r2 = 0.964, P = 0.013) and accordingly that tissue
turnover was negligible.
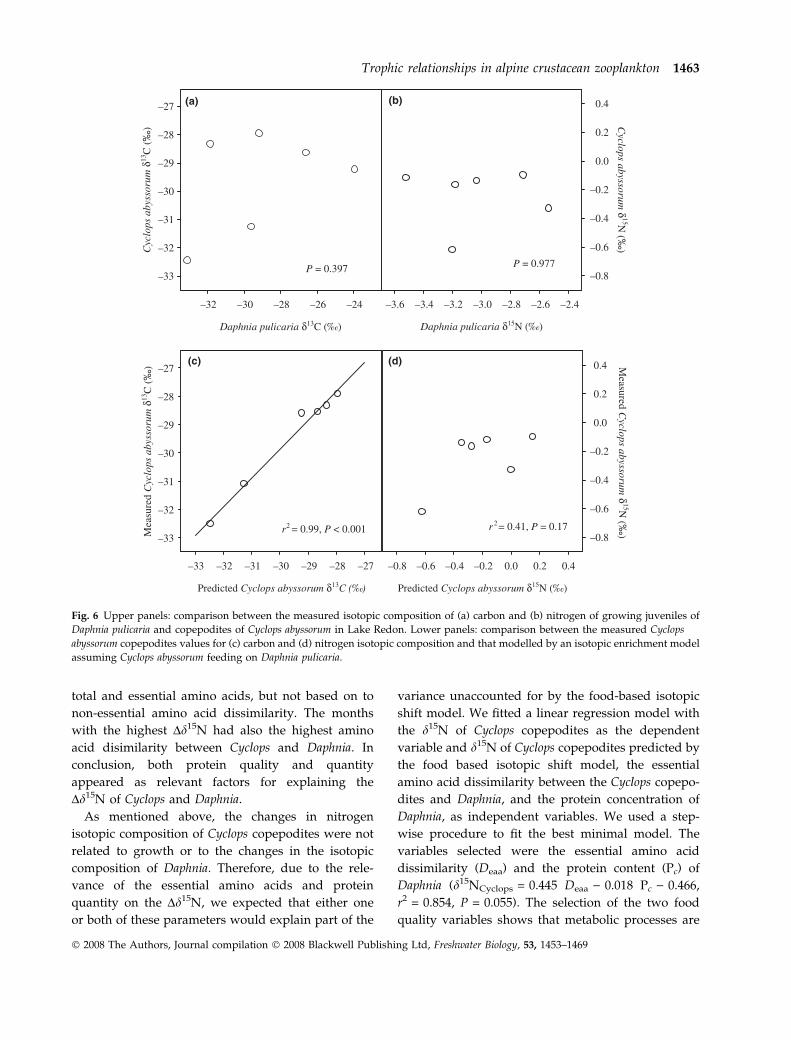
The isotopic compositions of the juveniles of Cyclops
and second cohort females of Daphnia were not
significantly correlated (P = 0.397 and P = 0.977 for
d13C and d15N respectively; Fig. 6a,b). However, this
lack of correlation does not imply that Cyclops could
not have been feeding on Daphnia, since it does not
take into account the changes in the growth rate of
Cyclops. Therefore, in order to determine whether the
isotopic composition of Cyclops might be explained by
the assumption that it was mainly feeding on Daphnia,
we employed a food-based isotopic shift model based
on the growth rate of copepodites, assuming the
influence of tissue turnover to be negligible (see
above). The model simply adds ingested carbon and
nitrogen from food (Daphnia in this case) proportion-
ally to the average population growth rate at
every time step, and allows for food isotopic variation
from step to step (Parker, Anderson & Lawrence,
1989):
dPredt¼ dPredt�1Wt�1=WtþðdfoodþfaÞð1�Wt�1=WtÞ
where dpred is the carbon or nitrogen isotopic compo-
sition of the predator, W is the DW of the individual,
10
20
30
40
50
60
1.2
1.4
1.6
1.8
2.0
Cyclops abyssorum(copepodites)
yrD
(thgiew
µnI
gd–1
)
1
2
3
4
CII
CIII
CIII > CIV
CIV > CV
CV > CIV
CV > CIV
Daphnia pulicaria(second generation females)
yrD
µ(thgiew
dnIg
–1)
Length
()
mm
8 9 10 11 121999
8 9 10 11 121999
7
(a)
(b)
Fig. 4 Changes in the average (circles) and SE (vertical error
bars) of the population average length and dry weight of (a)
Daphnia pulicaria second cohort females and (b) dry weight of
Cyclops abyssorum copepodites. The stage dominating each
month is shown. When more than two stages were present their
order of importance in the total sample abundance is indicated
by >.
Trophic relationships in alpine crustacean zooplankton 1461
� 2008 The Authors, Journal compilation � 2008 Blackwell Publishing Ltd, Freshwater Biology, 53, 1453–1469
dfood is the average isotopic composition of food
between time t and t ) 1, and ƒa is the trophic
fractionation (3.4& for nitrogen and 0.5& for carbon).
The model successfully predicted the d13C of
Cyclops copepodites, explaining 95% of the measured
isotopic change (P < 0.001) (Fig. 6c). Contrary to d13C,
the fitted model for d15N was not significant
(P = 0.168) (Fig. 6d), and the modelled isotopic
change only accounted for 41% of the measured
change. This discrepancy between modelling carbon
and nitrogen isotopic changes suggested that an
additional fractionation mechanism prevailed in the
case of nitrogen.
Two main causes of changes in nitrogen isotopic
fractionation between consumer and food have been
described: insufficient protein availability in the food
and poor quality of food (i.e. proteins with different
amino acid composition between the consumer and
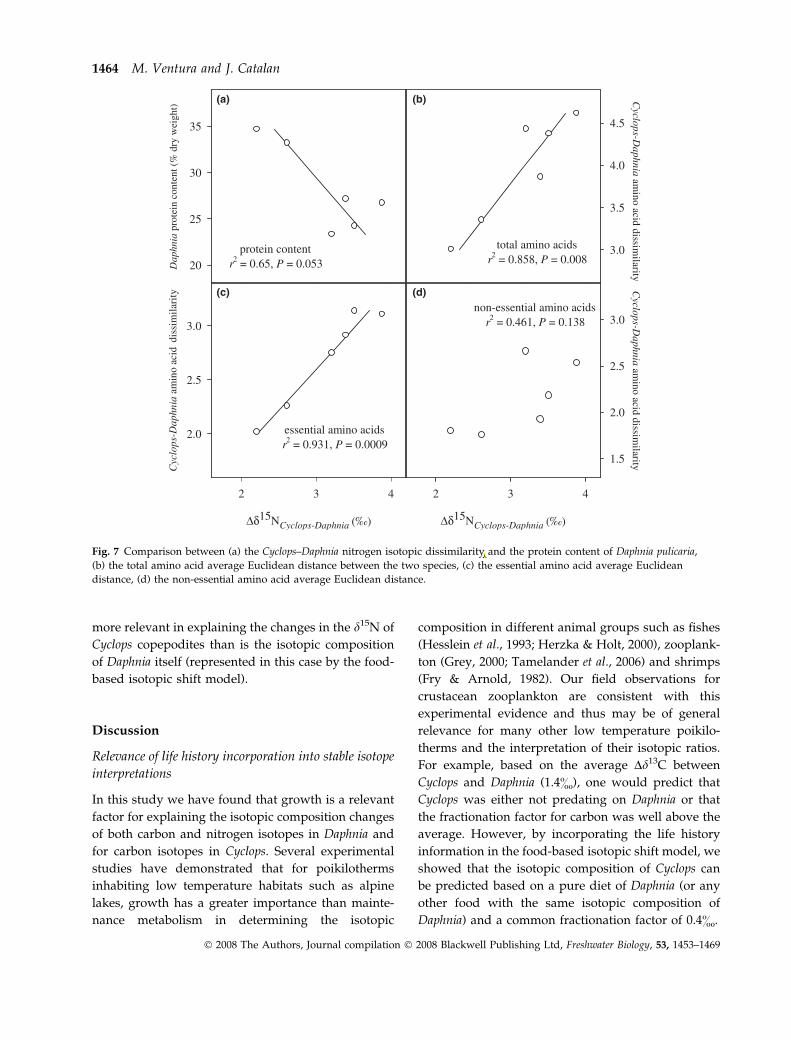
food). Measurement of the protein content showed
that Cyclops had higher protein content than Daphnia
throughout the study period (average DW% ± SE
was 43.6 ± 1.6 and 27.0 ± 1.1 for Cyclops and Daphnia
respectively). In addition, the relationship between
Daphnia protein content and Cyclops–Daphnia Dd15N
showed a negative correlation (r2 = 0.65, P = 0.053;
Fig. 7a). Therefore, if Cyclops were feeding on Daphnia,
protein limitation could be expected. To evaluate the
potential additional relevance of amino acid compo-
sition, we related the Dd15N between Cyclops and
Daphnia to the dissimilarity in their amino acid
composition. We calculated the amino acid average
dissimilarity between the two species either using all
amino acids (Fig. 7b), only essential amino acids
(Fig. 7c) or only non-essential amino acids (Fig. 7d).
There was a highly significant positive correlation
between Dd15N and amino acid dissimilarity based on
δ 3 1
) ‰
(
C
–36
–32
–28
–24
Length (m m )
0 . 8 1 . 2 1 . 6 2 . 0
δ 5 1
( N
‰
)
–4.2
–3.6
–3.0
–2.4
–1.8
Daphnia pulicaria
Daphnia pulicaria
r 2 = 0.61, P = 0.00 1
r 2 = 0. 5, P = 0.00 5
δ 3 1
( C
‰
)
–32
–30
–28
Dry weight (µg i n d –1 )
1 2 3 4
δ 5 1
( N
‰
)
–1
0
1
Cyclops abyssorum
Cyclops abyssorum
r2 = 0.99, P < 0.0001
(a) (c)
(b) (d)
C I I
C II I
C I I I > CI V
C IV >CV
C V > C I V
C V > CIV
Fig. 5 Ontogenetic changes in Daphnia pulicaria (a) carbon and (b) nitrogen isotope composition in relation to length and in Cyclops
abyssorum of (c) carbon and (d) nitrogen isotope composition in relation to dry weight. Values for Cyclops are monthly population
averages ± SE corresponding to different copepodite developmental stages. The stage dominating each month is shown. When more
than two stages were present, their order of importance in total sample abundance is indicated by >.
1462 M. Ventura and J. Catalan
� 2008 The Authors, Journal compilation � 2008 Blackwell Publishing Ltd, Freshwater Biology, 53, 1453–1469
total and essential amino acids, but not based on to
non-essential amino acid dissimilarity. The months
with the highest Dd15N had also the highest amino
acid disimilarity between Cyclops and Daphnia. In
conclusion, both protein quality and quantity
appeared as relevant factors for explaining the
Dd15N of Cyclops and Daphnia.
As mentioned above, the changes in nitrogen
isotopic composition of Cyclops copepodites were not
related to growth or to the changes in the isotopic
composition of Daphnia. Therefore, due to the rele-
vance of the essential amino acids and protein
quantity on the Dd15N, we expected that either one
or both of these parameters would explain part of the
variance unaccounted for by the food-based isotopic
shift model. We fitted a linear regression model with
the d15N of Cyclops copepodites as the dependent
variable and d15N of Cyclops copepodites predicted by
the food based isotopic shift model, the essential
amino acid dissimilarity between the Cyclops copepo-
dites and Daphnia, and the protein concentration of
Daphnia, as independent variables. We used a step-
wise procedure to fit the best minimal model. The
variables selected were the essential amino acid
dissimilarity (Deaa) and the protein content (Pc) of
Daphnia (d15NCyclops = 0.445 Deaa ) 0.018 Pc ) 0.466,
r2 = 0.854, P = 0.055). The selection of the two food
quality variables shows that metabolic processes are
r 2 = 0. 99, P < 0.001
Pr edi c ted Cycl ops abyssorum δ 13 C (‰ )
–33 –32 –31 –30 –29 –28 –27
M
d e r u s a e C
y c l
m
u r o s s y b a s p o
δ 3 1
) ‰
(
C
r 2 = 0. 41, P = 0.17
P r ed icted C y clops abyssorum δ 15 N (‰ )
–0.8 –0.6 –0.4 –0.2 0.0 0.2 0.4
d e
r u
s a
e M
C
y m
u
r o
s s
y b
a s
p o
l c
δ51
(N
‰)
D a phni a pul ic aria δ 13 C (‰)
–32 –30 –28 –26 –24
C y c
l m
u r o s s y b a
s p o δ
3 1 )
‰
( C
–33
–32
–31
–30
–29
–28
–27
–33
–32
–31
–30
–29
–28
–27
Daph nia p u li cari a δ 15 N ( ‰ )
–3.6 –3.4 –3.2 –3.0 –2.8 –2.6 –2.4
l c
y C
u
r o
s s
y b
a s
p o
m δ 5
1 (
N
‰ )
–0.8
–0.6
–0.4
–0.2
0.0
0.2
0.4
–0.8
–0.6
–0.4
–0.2
0.0
0.2
0.4
P = 0. 397 P = 0.977
(a) (b)
(c) (d)
Fig. 6 Upper panels: comparison between the measured isotopic composition of (a) carbon and (b) nitrogen of growing juveniles of
Daphnia pulicaria and copepodites of Cyclops abyssorum in Lake Redon. Lower panels: comparison between the measured Cyclops
abyssorum copepodites values for (c) carbon and (d) nitrogen isotopic composition and that modelled by an isotopic enrichment model
assuming Cyclops abyssorum feeding on Daphnia pulicaria.
Trophic relationships in alpine crustacean zooplankton 1463
� 2008 The Authors, Journal compilation � 2008 Blackwell Publishing Ltd, Freshwater Biology, 53, 1453–1469
more relevant in explaining the changes in the d15N of
Cyclops copepodites than is the isotopic composition
of Daphnia itself (represented in this case by the food-
based isotopic shift model).
Discussion
Relevance of life history incorporation into stable isotope
interpretations
In this study we have found that growth is a relevant
factor for explaining the isotopic composition changes
of both carbon and nitrogen isotopes in Daphnia and
for carbon isotopes in Cyclops. Several experimental
studies have demonstrated that for poikilotherms
inhabiting low temperature habitats such as alpine
lakes, growth has a greater importance than mainte-
nance metabolism in determining the isotopic
composition in different animal groups such as fishes
(Hesslein et al., 1993; Herzka & Holt, 2000), zooplank-
ton (Grey, 2000; Tamelander et al., 2006) and shrimps
(Fry & Arnold, 1982). Our field observations for
crustacean zooplankton are consistent with this
experimental evidence and thus may be of general
relevance for many other low temperature poikilo-
therms and the interpretation of their isotopic ratios.
For example, based on the average Dd13C between
Cyclops and Daphnia (1.4&), one would predict that
Cyclops was either not predating on Daphnia or that
the fractionation factor for carbon was well above the
average. However, by incorporating the life history
information in the food-based isotopic shift model, we
showed that the isotopic composition of Cyclops can
be predicted based on a pure diet of Daphnia (or any
other food with the same isotopic composition of
Daphnia) and a common fractionation factor of 0.4&.
3. 0
3. 5
4. 0
4. 5
Dap
hnia
pro
tein
con
tent
(%
dry
wei
ght)
20
25
30
35
Δδ 15 N Cyclops-D a phnia (‰)
2 3 4
aD-spolcy
Cnhp
iama
indi
ssim
ilari
tydica
o
2.0
2.5
3.0
Δδ 15 N Cyclops-D ap hnia (‰)
2 3 4
dissimilarity
polc
yC
hpa
D-s
ain
maic
ao n
id
dissimilarity
polc
yC
hpa
D-s
ain
madi
caon
i
1. 5
2. 0
2. 5
3. 0
to tal amin o acids r 2 = 0. 85 8, P = 0.008
essen t ial am ino acids r 2 = 0.93 1, P = 0.0009
n on-essential am ino aci d s r 2 = 0.46 1, P = 0.138
pro t ein con t ent r 2 = 0 . 65, P = 0.053
(a) (b)
(c) (d)
Fig. 7 Comparison between (a) the Cyclops–Daphnia nitrogen isotopic dissimilarity and the protein content of Daphnia pulicaria,
(b) the total amino acid average Euclidean distance between the two species, (c) the essential amino acid average Euclidean
distance, (d) the non-essential amino acid average Euclidean distance.
1464 M. Ventura and J. Catalan
� 2008 The Authors, Journal compilation � 2008 Blackwell Publishing Ltd, Freshwater Biology, 53, 1453–1469

Ontogenetic isotopic changes
Within zooplankton several ecological factors may
cause intra-specific isotopic variation independently
of food isotopic change, such as ontogenetic diet
changes or gender differences in food utilization. For
example, many copepods increase the size of their
prey from early naupliar stages to copepodites and
adults (Dussart & Defaure, 1995), and thus an isotopic
shift during ontogeny can be expected. In this study,
Diaptomus nauplii in June had a similar isotopic
signature to that of Daphnia (Table 1 and Fig. 2),
which agrees with the fact they both graze on small
planktonic organisms. In July, the dominant Diapto-
mus stage was adult and no nauplii were found. In
July, Diaptomus adults had the same isotopic compo-
sition as Cyclops copepodite CII, and both were 2.65&
in d15N above the Daphnia of July. Assuming that
nauplii in July would have had a signature similiar to
that of Daphnia, we predict that both Diaptomus and
Cyclops copepodite CII were almost one trophic level
above, and that in the case of Cyclops the shift from
primary consumer to secondary consumer occurs
during the first copepodite stage. This is probably
related to the major structural change that copepods
face from nauplius to copepodite CI. This result
agrees with what has been found in experimental
(Gliwicz & Stibor, 1993; Hanazato & Dodson, 1995)
and field studies (Gliwicz & Boavida, 1996; Gliwicz
et al., 2001). The progressive ontogenetic isotopic
enrichment observed for Cyclops copepodites was
therefore not due to a trophic shift but resulted from
the conjunction of two aspects: (i) predominance of
growth as the explanatory factor of body isotopic
change (reduced or negligible metabolic turnover)
and (ii) feeding on isotopically isotopic enriched food
during the main growth periods. This enriched food
was actually Daphnia, which was isotopically enriched
compared to Cyclops during the period in which
Cyclops were predominantly growing (Fig 2a), and
showed that copepodite predation can be an ecolog-
ically relevant but generally neglected cause of Daph-
nia population growth reduction.
Similarly to Cyclops d13C, changes in C and N
isotopes in Daphnia were related to changes in size.
There was a prevalence of growth effect combined
with feeding on isotopically depleted food during the
periods of growth. Other recent studies have found
similar Daphnia size-based isotopic variation in both
d13C (Matthews & Mazumder, 2006) and d15N
(Matthews & Mazumder, 2007). In the case of d13C,
these authors showed that the size-based isotope
variations in Daphnia were related to differential size-
based habitat selection in vertical gradients in the d13C
of particulate organic matter. In particular, they
showed that the epilimnion was enriched compared
to the hypolimnion, which corresponded with a
predominantly allochthonous origin of the epilimnetic
carbon and a predominantly autochthonous origin of
the hypolimnetic carbon. In our study site, the d13C of
the dominant vegetation of the Lake Redon catchment
was )24.9& and that of epilimnetic seston )25.4 ±
0.2& (Catalan et al., 2004), a value very similar to
those of the smaller stages of Daphnia ()24.1&),
whereas the hypolimnetic seston of Lake Redon had
an average isotopic composition of )28.5 ± 0.6&
below the thermocline (Catalan et al., 2004). Since
seston can be found to be enriched by about 3& with
respect to zooplankton in low phosphorous lakes
(del Giorgio & France, 1996), zooplankton in Lake
Redon feeding below the thermocline could have a
d13C of )31.5&, a value similar to those found for the
largest Daphnia in our study. Some species of Daphnia
(including D. pulicaria, the species inhabiting
Lake Redon) increase their filter size with ontogeny,
and increase the mean size of captured particles
(Lampert, 1974; Brendelberger, 1991). Ontogenetic
isotopic changes in these species are therefore to be
expected if the different size particles have distinct
isotopic compositions. A possible explanation for the
observed isotopic changes could be a diet change with
increasing phytoplankton contribution. In contrast to
d13C, the sestonic d15N was similar (c. )2&) between
epilimnion and hypolimnion in Lake Redon (Catalan
et al., 2004) and thus it could not explain the size-
based isotopic change in Daphnia. A similar pattern
has been described by Matthews & Mazumder (2007),
who found that seston had lower variability than
Daphnia. The origin of such variability is still an open
question.
Changes in adult isotopic composition
Changes with reproduction differed among the three
study species. Cyclops had the same d15N or d13C
when corrected for changes in lipid and chitin
content. Diaptomus adults also had the same d15N
but their d13C decreased in reproducing adults. This
Trophic relationships in alpine crustacean zooplankton 1465
� 2008 The Authors, Journal compilation � 2008 Blackwell Publishing Ltd, Freshwater Biology, 53, 1453–1469

decrease was only partially explained by a strong
decrease in lipid content. The lack of complete
explanation could be due to the use of an erroneous
fractionation factor for lipids. The value used in this
study ()3&) was chosen since it was obtained for
zooplankton living in similar conditions (Kling et al.,
1992). However, a more recent study focussing on
different zooplankton groups has found values rang-
ing from )4.4& to )8.2& (Smyntek et al., 2007), which
could explain almost all the difference in the isotopic
composition change observed in Diaptomus with
reproduction.
Contrasting with the two copepods, reproducing
Daphnia first cohort females increased their d13C and
d15N compared with over-wintering females. Changes
in biochemical composition were not responsible for
this change, since lipid correction increased the
difference in d13C from 1.3& to 2.6&, while chitin
correction had no effect on d15N. Similar results have
been found experimentally by Grey (2000) and
Tamelander et al. (2006). Their studies demonstrated
that adult copepods (the freshwater copepod Cyclops
sp. in the first study and the marine Calanus glacialis in
the second) did not equilibrate their body isotopic
composition with the composition of the experimental
food. Grey (2000) also found that Daphnia rapidly
equilibrated its body content with the signature of
experimental food and concluded that this was very
likely related to differences in the moulting charac-
teristics of copepods and cladocerans. While cope-
pods stop moulting once they are adults, cladocerans
continue moulting with the production of each egg
clutch (Tessier et al., 1983). Daphnia females of Lake
Redon did not start reproduction until May (Ventura
& Catalan, 2005). Reproducing Daphnia females (June
and July) were therefore more depleted in d13C or
d15N than non-reproducing females. Furthermore,
non-reproducing females had a more constant isoto-
pic composition (Fig. 2). Therefore, it is very likely
that only reproducing Daphnia females reflect the
variability occurring in their resources. In contrast,
non-reproducing Daphnia females and the isotopic
composition of Cyclops adults did not necessarily
reflect the composition of their diet, but rather a
‘summary’ of diet through their ontogeny.
Copepods show a sexual size dimorphism, which is
more pronounced in cyclopoid than in calanoid
copepods (Gilbert & Williamson, 1983). Two main
hypotheses have been suggested to explain the
adaptive evolution of this dimorphism: sexual selec-
tion and ecological causation (Shine, 1989). If the
second hypothesis is true, one would expect that
trophic gender differences would be more pro-
nounced in cyclopoid copepods, which should be
reflected in differences in their isotopic composition.
The almost identical isotopic composition of males
and females of both copepod species suggests that
there is no difference in the feeding of males and
females, at least, until their last stage. In fact, copepod
males and females have an almost identical morphol-
ogy during their ontogeny, and it is not until they
moult from copepodite V to adults that their mor-
phology distinctively differs. The question of whether
males and females feed on similar resources in the last
stage remains a subject for future studies.
Variability in Cyclops d15N enrichment
Cyclops d15N variability was not related to growth, but
to the protein concentration of Daphnia and the
essential amino acid dissimilarity between the two
species. Experimental studies have also reported a
lower protein content of the diet (e.g. in crabs by
Fantle et al., 1999; Daphnia by Adams & Sterner, 2000
and fish by Gaye-Siessegger et al., 2004) or an amino
acid imbalance between the consumer and its diet
(e.g. in birds by Hobson & Clark, 1992) that substan-
tially increases the tissue-diet isotopic fractionation.
The proposed mechanism for an increase in tissue-
diet fractionation with lower protein content is related
to a lower growth rate of animals and a progressive
increase in d15N of the animal through isotopic
selective excretion of ammonia (e.g. in crabs Fantle
et al., 1999 and fish Trueman, Mcgill & Guyard, 2005).
Unlike the effects of gross protein availability, the
effects of a changing amino acid composition on the
tissue-diet fractionation are much less studied. Some
of the amino acids are known to be essential for
animals because they are unable to synthesize the
carbon skeleton of the amino acids. This explains the
preservation of the d13C signature of the essential
amino acids from the diet, while the non-essential
amino acids undergo intense turnover and thus
fractionation (Fantle et al., 1999). This explanation for
d13C cannot be applied to d15N, since amino acids
derived from protein breakdown are deaminated to
keto acids, a portion of which is reaminated and
reincorporated into body tissue. Each transamination
1466 M. Ventura and J. Catalan
� 2008 The Authors, Journal compilation � 2008 Blackwell Publishing Ltd, Freshwater Biology, 53, 1453–1469

results in nitrogen isotope discrimination, with
excreted nitrogen being depleted in 15N and the
retained amino acids being correspondingly enriched
(Scrimgeour et al., 1995). The specific mechanisms
driving changes in the d15N of individual amino acid
are not well described. A comparison of the individ-
ual amino acid d15N of a marine rotifer and those with
the d15N values of its food showed that only seven of
the 13 amino acids analysed had a clear d15N
fractionation (McClelland & Montoya, 2002), half of
them being essential. Therefore, the physiological
mechanisms behind the relationship between essential
amino acid dissimilarity and the Cyclops–Daphnia
Dd15N found in this study require further investiga-
tion. However, a possible explanation may be an
increased amino acid metabolism in diets deficient in
essential amino acids, including remobilization of
stored amino acids; the same not being true in the case
of high deficiency in non-essential amino acids.
Finally, the lack of correlation between the Cyclops
d15N and growth and its positive correlation with the
protein and amino acid dissimilarity between Daphnia
and Cyclops may be interpreted as evidence of
nitrogen or amino acid limitation of Cyclops growth.
In a parallel study, we found that Cyclops contained
almost half of the phosphorus content that was
present in Daphnia during the whole seasonal cycle
(Ventura & Catalan, 2005). Therefore, it seems
unlikely that copepods would suffer from phospho-
rous limitation when feeding on Daphnia. This is
probably true for most carnivorous cyclopoid cope-
pods feeding on cladocerans, since all cyclopoids have
a lower phosphorus content than cladocerans
(Ventura, 2006). Similarly, Daphnia had a higher
phosphorus content than seston (Ventura & Catalan,
2005), suggesting that its growth might be limited by
phosphorous availability, which would also explain
why both d13C and d15N showed a similar relationship
with Daphnia growth (Fig. 5a,b).
In conclusion, our results show that in field studies
dealing with a community of a small number of
species, stable isotope variation can only be used to
study diet changes and nutrient limitations when life
histories are properly considered.
Acknowledgments
We thank LL. Camarero, B. Claramunt, A. Miro, P.
Renom, T. Buchaca and G. Cots for their assistance
with the field work, and R. Franco for laboratory
assistance. The total amino acid analysis and the
stable isotope C and N analyses were performed at the
Serveis Cientıfico Tecnics at the University of Barce-
lona, whose assistance is kindly acknowledged. A.M.
Poulsen assisted with linguistic ⁄editorial corrections.
We thank T. Buchaca, T. Larsen, the editor and
anonymous referees for very helpful comments on
the manuscript. The research was partially supported
by the European Commission projects MOLAR
(ENV4-CT95-0007) and EUROLIMPACS (GOCE-
CT-2003-505540) and the Spanish MEC project
TRAZAS (CGL2004-02989). MV was partially sup-
ported by a Marie Curie post-doctoral grant (MEIF-
CT-2005-010554) and a Juan de la Cierva contract.
References
Adams T.S. & Sterner R.W. (2000) The effect of dietary
nitrogen content on trophic level 15N enrichment.
Limnology and Oceanography, 45, 601–607.
Brendelberger H. (1991) Filter mesh size of cladocerans
predicts retention efficiency for bacteria. Limnology and
Oceanography, 36, 884–894.
Camarero L., Felip M., Ventura M., Bartumeus F. &
Catalan J. (1999) The relative importance of the
planktonic food web in the carbon cycle of an oligo-
trophic mountain lake in a poorly vegetated catchment
(Redo, Pyrenees). Journal of Limnology, 58, 203–212.
Catalan J., Ventura M., Vives I. & Grimalt J.O. (2004)
The roles of food and water in the bioaccumulation
of organochlorine compounds in high mountain lake
fish. Environmental Science & Technology, 38, 4269–
4275.
DeNiro M.J. & Epstein S. (1977) Mechanism of carbon
isotope fractionation associated with lipid synthesis.
Science, 197, 261–263.
Dussart B.H. & Defaure D. (1995) Copepoda: Introduction
to Copepoda. SPB Academic Publishing, Amsterdam.
Fantle M.S., Dittel A.I., Schwalm S.M., Epifiano C.E. &
Fogel M.L. (1999) A food web analysis of the juvenile
blue crab, Callinectes sapidus, using stable isotopes in
whole animals and individual amino acids. Oecologia,
120, 416–426.
Felip M., Bartumeus F., Halac S. & Catalan J. (1999)
Microbial plankton assemblages, composition and
biomass, during two ice-free periods in a deep high
mountain lake (Estany Redo, Pyrenees). Journal of
Limnology, 58, 193–202.
Folch J., Lees M. & Sloane Stanley G.H. (1957) A simple
method for the isolation and purification of total lipids
Trophic relationships in alpine crustacean zooplankton 1467
� 2008 The Authors, Journal compilation � 2008 Blackwell Publishing Ltd, Freshwater Biology, 53, 1453–1469

from animal tissues. Journal of Biological Chemistry, 226,
497–509.
Fry B. & Arnold C. (1982) Rapid 13C ⁄ 12C turnover during
growth of brown shrimp (Panaeus aztecus). Oecologia,
54, 200–204.
Fryer G. (1957) The feeding mechanism of some fresh-
water cyclopoid copepods. Proceedings of the Zoological
Society of London, 129, 1–25.
Gannes L.Z., O’Brien D.M. & Martınez del Rio C. (1997)
Stable isotopes in animal ecology: assumptions, cave-
ats, and a call for more laboratory experiments.
Ecology, 78, 1271–1276.
Gannes L.Z., Martınez del Rio C. & Koch P. (1998)
Natural abundance variations in stable isotopes and
their potential uses in animal physiological ecology.
Comparative Biochemistry and Physiology, 119A, 725–737.
Gaye-Siessegger J., Focken U., Abel H. & Becker K. (2004)
Individual protein balance strongly influences d15N
and d13C values in Nile tilapia, Oreochromis niloticus.
Naturwissenschaften, 91, 90–93.
Gilbert J.J. & Williamson C.E. (1983) Sexual dimorphism
in zooplankton. Annual Review of Ecology and System-
atics, 14, 1–33.
del Giorgio P.A. & France R.L. (1996) Ecosystem-specific
patterns in the relationship between zooplankton and
POM or microplankton 13C. Limnology and Oceanogra-
phy, 41, 359–365.
Gliwicz Z.M. & Boavida M.J. (1996) Clutch size and body
size at first reproduction in Daphnia pulicaria at
different levels of food and predation. Journal of
Plankton Research, 18, 881–894.
Gliwicz Z.M. & Lampert W. (1994) Clutch-size variability
in Daphnia: body-size related effects of egg predation
by cyclopoid copepods. Limnology and Oceanography,
39, 479–485.
Gliwicz Z.M. & Stibor H. (1993) Egg predation by
copepods in Daphnia brood cavities. Oecologia, 95,
295–298.
Gliwicz Z.M. & Umana G. (1994) Cladoceran body size
and vulnerability to copepod predation. Limnology and
Oceanography, 39, 419–424.
Gliwicz Z.M., Slusarczyk A. & Slusarczyk M. (2001) Life
history synchronization in a long-lifespan single-
cohort Daphnia population in a fishless alpine lake.
Oecologia, 128, 368–378.
Gonfiantini R. (1978) Standards for stable isotope
measurements in natural compounds. Nature, 271,
534–536.
Grey J. (2000) Trophic fractionation and the effects of diet
switch on the carbon stable isotopic ‘signatures’ of
pelagic consumers. Verhandlungen der Internationale
Vereinigung fur Theoretische und Angewandte Limnologie,
27, 3187–3191.
Hanazato T. & Dodson S.I. (1995) Morphological
defenses of Daphnia against Copepod predation on
eggs. Archiv fur Hydrobiologie, 133, 49–59.
Herzka S.Z. & Holt G.J. (2000) Changes in isotopic
composition of red drum (Sciaenops ocellatus) larvae in
response to dietary shifts: potential applications to
settlement studies. Canadian Journal of Fisheries and
Aquatic Sciences, 57, 137–147.
Hesslein R.H., Hasllard K.A. & Ramlal P. (1993) Replace-
ment of sulphur, carbon and nitrogen in tissue of
growing broad whitefish (Coregonus nasus) in response
to a change in diet traced by d34S, d13C and d15N.
Canadian Journal of Fisheries and Aquatic Sciences, 50,
2071–2076.
Hobson K.A. & Clark R.G. (1992) Assessing avian diets
using stable isotopes. 2. Factors influencing diet-tissue
fractionation. Condor, 94, 189–197.
Hopp U. & Maier G. (2005) Implication of the feeding
limb morphology for herbivorous feeding in some
freshwater cyclopoid copepods. Freshwater Biology, 50,
742–747.
Kling G.W., Fry B. & O’Brien W.J. (1992) Stable isotopes
and planktonic trophic structure in arctic lakes. Ecol-
ogy, 73, 561–566.
Lampert W. (1974) A method for determining food
selection by zooplankton. Limnology and Oceanography,
19, 995–998.
Macko S.A., Helleur R., Hartley G. & Jackman P. (1990)
Diagenesis of organic matter: a study using stable
isotopes of individual carbohydrates. Organic Geochem-
istry, 16, 1129–1137.
Matthews B. & Mazumder A. (2003) Compositional and
interlake variability of zooplankton affect baseline
stable isotope signatures. Limnology and Oceanography,
48, 1977–1987.
Matthews B. & Mazumder A. (2006) Habitat specializa-
tion and the exploitation of allochthonous carbon by
zooplankton. Ecology, 87, 2800–2812.
Matthews B. & Mazumder A. (2007) Distinguishing
trophic variation from seasonal and size-based isotopic
(d15N) variation of zooplankton. Canadian Journal of
Fisheries and Aquatic Sciences, 64, 74–83.
McClelland J.W. & Montoya J.P. (2002) Trophic relation-
ships and the nitrogen isotopic composition of amino
acids in plankton. Ecology, 83, 2173–2180.
McCutchan J.H. Jr, Lewis W.J. Jr, Kendall C. & McGrath
C.C. (2003) Variation in trophic shift for stable isotope
ratios of carbon, nitrogen, and sulphur. Oikos, 102, 378–
390.
O’Reilly C.M., Hecky R.E., Cohen A.S. & Plisnier P.-D.
(2002) Interpreting stable isotopes in food webs:
recognizing the role of time averaging at different
trophic levels. Limnology and Oceanography, 47, 306–309.
1468 M. Ventura and J. Catalan
� 2008 The Authors, Journal compilation � 2008 Blackwell Publishing Ltd, Freshwater Biology, 53, 1453–1469

Oelbermann K. & Scheu S. (2002) Stable isotope enrich-
ment (d15N and d13C) in a generalist predator (Pardosa
lugubris, Araneae: Lycosidae): effects of prey quality.
Oecologia, 130, 337–344.
Paine R.T. (1988) Food webs – road maps of interactions
or grist for theoretical development. Ecology, 69, 1648–
1654.
Parker P.L., Anderson R.K. & Lawrence A. (1989) A d13C
and d15N tracer study of nutrition in aquaculture:
Penaeus vannamei in a pond growth system. In: Stable
Isotopes in Ecological Research, (Eds P.W. Rundel, J.R.
Ehleringer & K.A. Nagy) pp. 288–303, Springer-Verlag,
New York.
Peterson B.J. & Fry B. (1987) Stable isotopes in ecosystem
studies. Annual Review of Ecology and Systematics, 18,
293–320.
Ponsard S. & Averbuch P. (1999) Should growing and
adult animals fed on the same diet show different d15N
values? Rapid Communications in Mass Spectrometry, 13,
1305–1310.
Post D.M. (2002) Using stable isotopes to estimate trophic
position: models, methods, and assumptions. Ecology,
83, 703–718.
Santer B. (1998) Life cycle strategies of free-living
copepods in fresh waters. Journal of Marine Systems,
15, 327–336.
Santer B., Sommerwerk N. & Grey J. (2006) Food niches
of cyclopoid copepods in eutrophic Plubsee deter-
mined by stable isotope analysis. Archiv fur Hydrobiol-
ogie, 167, 301–316.
Schindler J.E. (1971) Food quality and zooplankton
nutrition. Journal of Animal Ecology, 40, 589–595.
Scrimgeour C.M., Gordon S.C., Handley L.L. &
Woodford J.A.T. (1995) Trophic levels and anomalous
delta-N-15 of insects on Raspberry (Rubus idaeus L).
Isotopes in Environmental and Health Studies, 31, 107–115.
Shine R. (1989) Ecological causes of the evolution of
sexual dimorphism: a review of the evidence. Quarterly
Review of Biology, 64, 419–441.
Smyntek P.M., Teece M.A., Schulz K.L. & Thackeray S.J.
(2007) A standard protocol for stable isotope analysis
of zooplankton in aquatic food web research using
mass balance correction models. Limnology and Ocean-
ography, 52, 2135–2146.
Spackman D.H., Stein W.H. & Moore S. (1958) Automatic
recording apparatus for use in the chromatography of
amino acids. Analytical Chemistry, 30, 1190–1206.
Tamelander T., Soreide J.E., Hop H. & Carroll M.L.
(2006) Fractionation of stable isotopes in the Arctic
marine copepod Calanus glacialis: effects on the isoto-
pic composition of marine particulate organic matter.
Journal of Experimental Marine Biology and Ecology, 333,
231–240.
Tessier A.J., Henry L.L., Goulden C.E. & Durand M.W.
(1983) Starvation in Daphnia: energy reserves and
reproductive allocation. Limnology and Oceanography,
28, 667–676.
Trueman C.N., Mcgill R.A.R. & Guyard P.H. (2005) The
effect of growth rate on tissue-diet isotopic spacing in
rapidly growing animals. An experimental study with
Atlantic salmon (Salmo salar). Rapid Communications in
Mass Spectrometry, 19, 3239–3247.
Vandenbosch F. & Santer B. (1993) Cannibalism in
Cyclops abyssorum. Oikos, 67, 19–28.
Vander Zanden M.J. & Rasmussen J.B. (1999) Primary
consumer d13C and d15N and the trophic position of
aquatic consumers. Ecology, 80, 1395–1404.
Vander Zanden M.J. & Rasmussen J.B. (2001) Variation in
d15N and d13C trophic fractionation: implications for
aquatic food web studies. Limnology and Oceanography,
46, 2061–2066.
Vanderklift M.A. & Ponsard S. (2003) Sources of varia-
tion in consumer-diet 15N enrichment: a meta-analysis.
Oecologia, 136, 169–182.
Ventura M. (2006) Linking biochemical and elemental
composition of freshwater and marine crustacean
zooplankton. Marine Ecology – Progress Series, 327,
233–246.
Ventura M. & Catalan J. (2005) Reproduction as one of
the main causes of temporal variability in the elemen-
tal composition of zooplankton. Limnology and Ocean-
ography, 50, 2043–2056.
Webb S.C., Hedges R.E.M. & Simpson S.J. (1998) Diet
quality influences the d13C and d15N of locusts and
their biochemical components. Journal of Experimental
Biology, 201, 2903–2911.
Whitehouse J.W. & Lewis B.G. (1973) The effect of diet
and density on development, size and egg production
in Cyclops abyssorum. Crustaceana, 25, 225–236.
(Manuscript accepted 23 January 2008)
Trophic relationships in alpine crustacean zooplankton 1469
� 2008 The Authors, Journal compilation � 2008 Blackwell Publishing Ltd, Freshwater Biology, 53, 1453–1469