tubular localization of silent calcium channels in crustacean skeletal muscle fibers
TRANSCRIPT

Tubular localization of silent calcium channels in crustacean skeletal muscle fibers
J. MONTERRUBIO�, G. ORTIZ, P. M. ORKAND and C. ZUAZAGA*Institute of Neurobiology, Department of Physiology, University of Puerto Rico, Medical Sciences Campus,201 Boulevard del Valle, San Juan, PR 00901, USA
Received 14 July 2001; accepted in revised form 28 March 2002
Abstract
Ca2þ-induced Ca2þ release (CICR) in the superficial abdominal flexor muscle of the crustacean Atya lanipes appearsto be mediated by a local control mechanism similar to that of vertebrate cardiac muscle, but with an unusually highgain. Thus, Ca2þ influx increases sufficiently the local concentration of Ca2þ in the immediate vicinity of thesarcoplasmic reticulum Ca2þ release channels to trigger the highly amplified release of Ca2þ required forcontraction, but is too low to generate a macroscopic inward current (i.e., the Ca2þ channels are silent). Todetermine the localization of the silent Ca2þ channels, the mechanical, electrophysiological and ultrastructuralproperties of the muscle were examined before and after formamide treatment, a procedure that produces thedisruption of transverse tubules of striated muscle. We found that tubular disruption decreased tension generationby about 90%; reduced inward current (measured as Vmax, the maximum rate of rise of Sr2þ action potentials) byabout 80%; and decreased membrane capacitance by about 77%. The results suggest that ca. 80% of the silent Ca2þ
channels are located in the tubular system. Thus, these studies provide further evidence to support the local controlmechanism of CICR in crustacean skeletal muscle.
Introduction
Electrical excitation in arthropod skeletal muscle rangesfrom low-amplitude graded responses to overshootingaction potentials, and is related to an increase in Ca2þ
conductance (crab: Fatt and Katz, 1953; Fatt andGinsborg, 1958; Mounier and Vassort, 1975; barnacle:Hagiwara and Naka, 1964; Keynes et al., 1973; crayfish:Hencek and Zachar, 1977; insect: Washio, 1972; Ash-croft and Stanfield, 1982; scorpion: Gilly and Scheuer,1984, 1993; isopod: Erxleben and Rathmayer, 1997).The role of the voltage-dependent Ca2þ channels presentin the surface/transverse (T)-tubule membranes is tocontrol the influx of Ca2þ required for tension genera-tion during excitation–contraction (E–C) coupling (Za-charova and Zachar, 1967; Gainer, 1968; Ashley andRidgway, 1970; Atwater et al., 1974; Caputo and DiPolo, 1978; Gilly and Scheuer, 1984). The influx of Ca2þ
alone, however, is insufficient for E–C coupling. Incrustacean skeletal muscle, as in vertebrate cardiacmuscle (Fabiato, 1983), Ca2þ influx triggers the releaseof larger amounts of Ca2þ from the sarcoplasmicreticulum (SR) to activate contraction, a mechanismknown as Ca2þ-induced Ca2þ release (CICR) (Ashleyand Lea, 1978; Caputo and Di Polo, 1978; Mounier and
Goblet, 1987; Vergara and Verdugo, 1988; Gyorke andPalade, 1992, 1993, 1994; Palade and Gyorke, 1993;Monterrubio et al., 2000).The superficial abdominal flexor muscle fibers of the
tropical freshwater shrimp Atya lanipes (Decapoda:Atyidae) are of the long sarcomere, tonic type (Bonillaet al., 1992). In contrast to other arthropod skeletalmuscle, these fibers are electrically inexcitable: they donot generate graded potentials or Ca2þ spikes whendepolarized (Zuazaga and del Castillo, 1985) and, undervoltage clamp conditions, inward Ca2þ currents inresponse to depolarizing voltage steps are unmeasurablysmall, even when all outward currents are suppressed(Lizardi et al., 1992; Monterrubio et al., 2000). How-ever, as in other crustacean muscles, Ca2þ influx is anabsolute requirement for contractile activation (Bonillaet al., 1992; Monterrubio et al., 2000). The voltage-dependent Ca2þ channels that mediate this influx can beunmasked, in the complete absence of extracellularCa2þ, using Sr2þ or Ba2þ as current carriers. We havecalled them ‘silent’ Ca2þ channels because, even whenthe driving force for Ca2þ is increased 10-fold, Ca2þ
current density remains unmeasurably low (Monterru-bio et al., 2000).To reconcile these observations we proposed that
CICR in this crustacean muscle is mediated by a localcontrol mechanism similar to that of vertebrate cardiacmuscle (see Gomez et al., 1997 and references therein)and, furthermore, that it operates with an unusuallyhigh gain. Thus, Ca2þ influx through the silent Ca2þ
channels is too low to generate a macroscopic inward
� Current address: Department of Biological Sciences, Faculty of
General Studies, University of Puerto Rico, Rıo Piedras Campus, Rıo
Piedras, PR 00931, USA
* To whom correspondence should be addressed: E-mail: czuaza-
Journal of Muscle Research and Cell Motility 23: 167–174, 2002. 167� 2002 Kluwer Academic Publishers. Printed in the Netherlands.

current, but increases sufficiently the local concentrationof Ca2þ in the immediate vicinity of the SR Ca2þ releasechannels to trigger the highly amplified release of Ca2þ
required for tension generation (Monterrubio et al.,2000). This local control proposal assumes that most ofthe silent Ca2þ channels are located in the T-tubulemembrane, in close proximity to the SR release channelsin the dyad.The present studies were carried out to determine the
localization of the silent Ca2þ channels in skeletalmuscle fibers of A. lanipes. The mechanical, electro-physiological, and ultrastructural properties of the fiberswere examined before and after formamide treatment, aprocedure that produces detubulation of frog skeletalmuscle fibers (del Castillo and Escalona de Motta, 1978;Argiro, 1981; Escalona de Motta et al., 1982; Putnam,1996) and isolated cardiac myocytes (Kawai et al.,1999). We found that the formamide procedure detu-bulated A. lanipes muscle fibers, and altered theirmechanical and electrophysiological properties in waysthat are compatible with a largely tubular localization ofthe silent Ca2þ channels. Preliminary results of this workhave previously been published (Monterrubio et al.,1999).
Materials and methods
Experiments were performed on fibers from the firstsegment of the superficial abdominal flexor muscle ofthe crustacean A. lanipes. They were dissected inphysiological saline solution (van Harreveld, 1936)having the following composition (mM): NaCl, 206;KCl, 5.4; CaCl2, 13.6; 3-(N-morpholino) propanesulf-onic acid (MOPS), 4.0; pH ¼ 7.4. Details of thepreparation and dissection have been previously pub-lished (Bonilla et al., 1992). Experiments were carriedout at room temperature (20–22�C).
Detubulation procedure
To detubulate muscle fibers, we soaked muscle fiberbundles in the physiological saline described above plus2 M formamide for 30 min, after which the fibers werere-exposed to the physiological saline. This procedurehas been reported to produce excitation–contractionuncoupling and detubulation of vertebrate skeletalmuscle fibers (del Castillo and Escalona de Motta,1978; Argiro, 1981; Escalona de Motta et al., 1982;Putnam, 1996) and cardiac muscle (Kawai et al., 1999).
Mechanical recordings
Small bundles of muscle fibers (6–8 fibers) were dissectedand allowed to rest for 30 min in physiological salinesolution. The fibers were transferred to the experimentalchamber and one end of the cuticle, to which the musclesare attached, was held in a hook fixed to the bottom ofthe chamber. The other end was hooked to the lever of a
Cambridge Series 400A isometric mechanoelectronictransducer, whose output was sampled by and stored ona personal computer for further analysis. Mechanicaltension was generated by extracellular electrical stimuliapplied to the bundle of fibers longitudinally via two fineplatinum wire electrodes connected to a Grass S11stimulator. Maximum tension in response to singlesuprathreshold stimuli was adjusted by stretching eachbundle of fibers studied.
Electrophysiological recordings
Current-clamp experiments were performed with twomicroelectrodes inserted in the middle of a muscle fiber.Depolarizing constant-current pulses were delivered viaa 2 M K-citrate filled microelectrode (4–7 MW), and thevoltage responses recorded with the second microelec-trode (3 M KCl; 20–25 MW), using an Axoclamp-2Aamplifier. A personal computer running pCLAMP 7software was used to trigger the internal command ofthe amplifier for injecting current pulses. Voltage signalswere differentiated using an RC circuit with a timeconstant of 10 ms. Membrane potentials and theirtime derivatives were sampled at 12-bit resolution every1–2 ms and stored on the computer for further analysis.Electrical activity was recorded in an isotonic Ca2þ-freesolution containing (mM): NaCl, 106.5; KCl, 5.4; SrCl2,80.0; MOPS, 4.0; EGTA, 5.0; pH ¼ 7.4. Because of itshigher permeability through calcium channels in thispreparation, Sr2þ generates large amplitude actionpotentials (Monterrubio et al., 2000).
Linear cable analysis
Intracellular recordings, described above, were used tomeasure the passive electrical constants of control andformamide-treated muscle fibers (Hodgkin and Rush-ton, 1946). Hyperpolarizing current pulses, adjusted toproduce steady-state voltage deflections, were injected,and the voltage responses measured at least threedifferent distances along the fiber length. The inter-electrode spacing was measured with a calibratedmicrometer in the eyepiece of the microscope.
Electron microscopy
Muscles used in the mechanical experiments weresubsequently fixed for electron microscopy. The mus-cles, pinned to the bottom of a custom-made Sylgard-lined plexiglass chamber to maintain resting length, wereimmersed in a fixative solution containing 1.0% glutar-aldehyde, 2.5% paraformaldehyde, 0.1% cacodylatebuffer, 0.05% calcium chloride and 3.0% lanthanumchloride at pH 7.5, and left overnight. The tissue wasrinsed in cacodylate buffer and placed in 3.0% lantha-num chloride for 2 h, cut in small pieces (1 mm), andplaced in 0.1% phosphate buffer at pH 7.4 for 10 min toprecipitate lanthanum salts. This lanthanum precipita-tion method was used to label parts of the tubular
168
system in communication with the extracellular space(Fahimi and Cotran, 1971). The material was post-fixedin 1.0% buffered osmium tetroxide and dehydrated inan ascending series of ethanol solutions with en blocuranyl acetate staining in 70% ethanol. Dehydrationwas continued, and the tissue was embedded in Med-cast/Araldite. Sections were cut on a Sorvall MT-2microtome, stained with uranyl acetate and lead citrate,and examined in a Philips EM 201 electron microscope.Control muscles were processed the same way.
Results
Muscle contraction is irreversibly lost after T-tubuledisruption
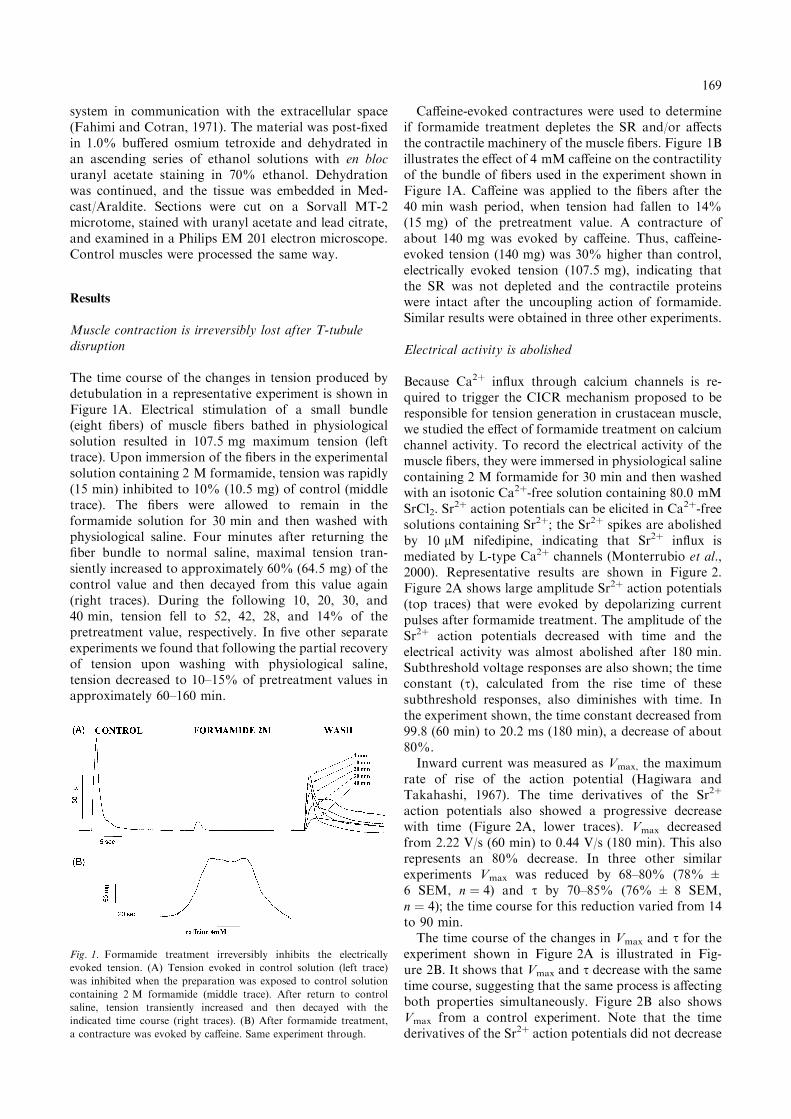
The time course of the changes in tension produced bydetubulation in a representative experiment is shown inFigure 1A. Electrical stimulation of a small bundle(eight fibers) of muscle fibers bathed in physiologicalsolution resulted in 107.5 mg maximum tension (lefttrace). Upon immersion of the fibers in the experimentalsolution containing 2 M formamide, tension was rapidly(15 min) inhibited to 10% (10.5 mg) of control (middletrace). The fibers were allowed to remain in theformamide solution for 30 min and then washed withphysiological saline. Four minutes after returning thefiber bundle to normal saline, maximal tension tran-siently increased to approximately 60% (64.5 mg) of thecontrol value and then decayed from this value again(right traces). During the following 10, 20, 30, and40 min, tension fell to 52, 42, 28, and 14% of thepretreatment value, respectively. In five other separateexperiments we found that following the partial recoveryof tension upon washing with physiological saline,tension decreased to 10–15% of pretreatment values inapproximately 60–160 min.
Caffeine-evoked contractures were used to determineif formamide treatment depletes the SR and/or affectsthe contractile machinery of the muscle fibers. Figure 1Billustrates the effect of 4 mM caffeine on the contractilityof the bundle of fibers used in the experiment shown inFigure 1A. Caffeine was applied to the fibers after the40 min wash period, when tension had fallen to 14%(15 mg) of the pretreatment value. A contracture ofabout 140 mg was evoked by caffeine. Thus, caffeine-evoked tension (140 mg) was 30% higher than control,electrically evoked tension (107.5 mg), indicating thatthe SR was not depleted and the contractile proteinswere intact after the uncoupling action of formamide.Similar results were obtained in three other experiments.
Electrical activity is abolished
Because Ca2þ influx through calcium channels is re-quired to trigger the CICR mechanism proposed to beresponsible for tension generation in crustacean muscle,we studied the effect of formamide treatment on calciumchannel activity. To record the electrical activity of themuscle fibers, they were immersed in physiological salinecontaining 2 M formamide for 30 min and then washedwith an isotonic Ca2þ-free solution containing 80.0 mMSrCl2. Sr
2þ action potentials can be elicited in Ca2þ-freesolutions containing Sr2þ; the Sr2þ spikes are abolishedby 10 lM nifedipine, indicating that Sr2þ influx ismediated by L-type Ca2þ channels (Monterrubio et al.,2000). Representative results are shown in Figure 2.Figure 2A shows large amplitude Sr2þ action potentials(top traces) that were evoked by depolarizing currentpulses after formamide treatment. The amplitude of theSr2þ action potentials decreased with time and theelectrical activity was almost abolished after 180 min.Subthreshold voltage responses are also shown; the timeconstant (s), calculated from the rise time of thesesubthreshold responses, also diminishes with time. Inthe experiment shown, the time constant decreased from99.8 (60 min) to 20.2 ms (180 min), a decrease of about80%.Inward current was measured as Vmax, the maximum
rate of rise of the action potential (Hagiwara andTakahashi, 1967). The time derivatives of the Sr2þ
action potentials also showed a progressive decreasewith time (Figure 2A, lower traces). Vmax decreasedfrom 2.22 V/s (60 min) to 0.44 V/s (180 min). This alsorepresents an 80% decrease. In three other similarexperiments Vmax was reduced by 68–80% (78% ±6 SEM, n ¼ 4) and s by 70–85% (76% ± 8 SEM,n ¼ 4); the time course for this reduction varied from 14to 90 min.The time course of the changes in Vmax and s for the
experiment shown in Figure 2A is illustrated in Fig-ure 2B. It shows that Vmax and s decrease with the sametime course, suggesting that the same process is affectingboth properties simultaneously. Figure 2B also showsVmax from a control experiment. Note that the timederivatives of the Sr2þ action potentials did not decrease
Fig. 1. Formamide treatment irreversibly inhibits the electrically
evoked tension. (A) Tension evoked in control solution (left trace)
was inhibited when the preparation was exposed to control solution
containing 2 M formamide (middle trace). After return to control
saline, tension transiently increased and then decayed with the
indicated time course (right traces). (B) After formamide treatment,
a contracture was evoked by caffeine. Same experiment through.
169
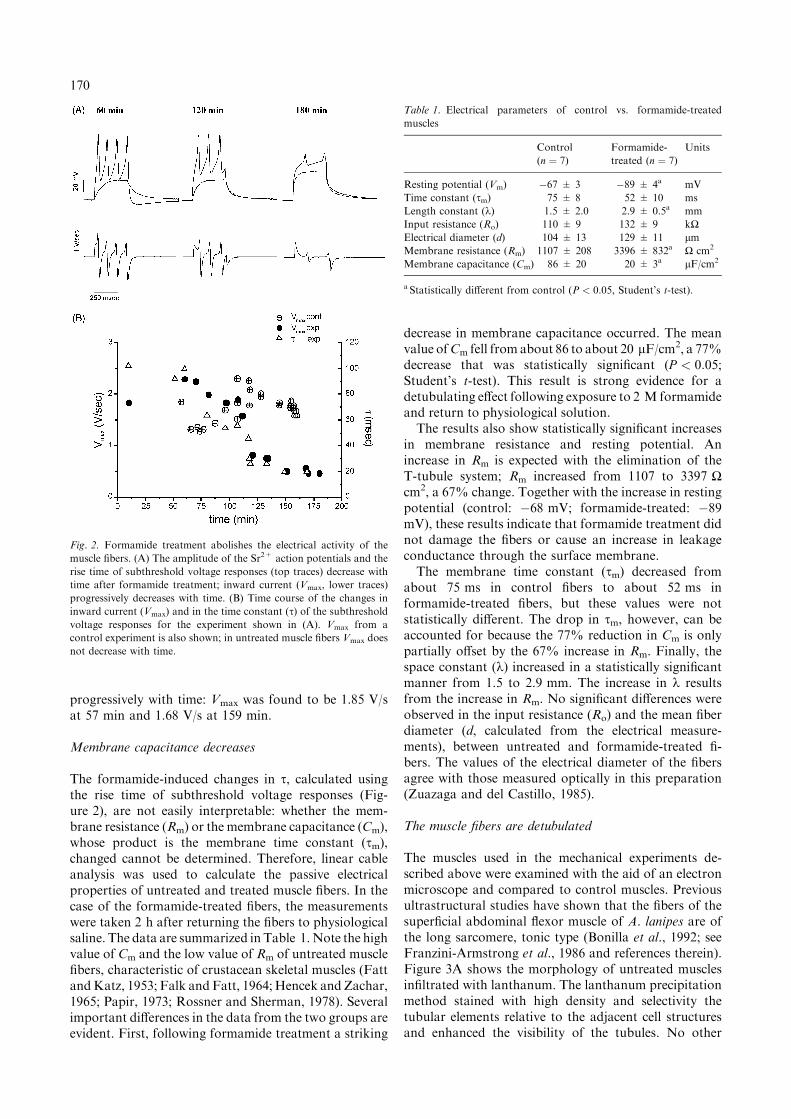
progressively with time: Vmax was found to be 1.85 V/sat 57 min and 1.68 V/s at 159 min.
Membrane capacitance decreases
The formamide-induced changes in s, calculated usingthe rise time of subthreshold voltage responses (Fig-ure 2), are not easily interpretable: whether the mem-brane resistance (Rm) or the membrane capacitance (Cm),whose product is the membrane time constant (sm),changed cannot be determined. Therefore, linear cableanalysis was used to calculate the passive electricalproperties of untreated and treated muscle fibers. In thecase of the formamide-treated fibers, the measurementswere taken 2 h after returning the fibers to physiologicalsaline. The data are summarized in Table 1.Note the highvalue of Cm and the low value of Rm of untreated musclefibers, characteristic of crustacean skeletal muscles (FattandKatz, 1953; Falk andFatt, 1964;Hencek andZachar,1965; Papir, 1973; Rossner and Sherman, 1978). Severalimportant differences in the data from the two groups areevident. First, following formamide treatment a striking
decrease in membrane capacitance occurred. The meanvalue ofCm fell from about 86 to about 20 lF/cm2, a 77%decrease that was statistically significant (P < 0:05;Student’s t-test). This result is strong evidence for adetubulating effect following exposure to 2 M formamideand return to physiological solution.The results also show statistically significant increases
in membrane resistance and resting potential. Anincrease in Rm is expected with the elimination of theT-tubule system; Rm increased from 1107 to 3397 Wcm2, a 67% change. Together with the increase in restingpotential (control: �68 mV; formamide-treated: �89mV), these results indicate that formamide treatment didnot damage the fibers or cause an increase in leakageconductance through the surface membrane.The membrane time constant (sm) decreased from
about 75 ms in control fibers to about 52 ms informamide-treated fibers, but these values were notstatistically different. The drop in sm, however, can beaccounted for because the 77% reduction in Cm is onlypartially offset by the 67% increase in Rm. Finally, thespace constant (k) increased in a statistically significantmanner from 1.5 to 2.9 mm. The increase in k resultsfrom the increase in Rm. No significant differences wereobserved in the input resistance (Ro) and the mean fiberdiameter (d, calculated from the electrical measure-ments), between untreated and formamide-treated fi-bers. The values of the electrical diameter of the fibersagree with those measured optically in this preparation(Zuazaga and del Castillo, 1985).
The muscle fibers are detubulated
The muscles used in the mechanical experiments de-scribed above were examined with the aid of an electronmicroscope and compared to control muscles. Previousultrastructural studies have shown that the fibers of thesuperficial abdominal flexor muscle of A. lanipes are ofthe long sarcomere, tonic type (Bonilla et al., 1992; seeFranzini-Armstrong et al., 1986 and references therein).Figure 3A shows the morphology of untreated musclesinfiltrated with lanthanum. The lanthanum precipitationmethod stained with high density and selectivity thetubular elements relative to the adjacent cell structuresand enhanced the visibility of the tubules. No other
Fig. 2. Formamide treatment abolishes the electrical activity of the
muscle fibers. (A) The amplitude of the Sr2+ action potentials and the
rise time of subthreshold voltage responses (top traces) decrease with
time after formamide treatment; inward current (Vmax, lower traces)
progressively decreases with time. (B) Time course of the changes in
inward current (Vmax) and in the time constant (s) of the subthresholdvoltage responses for the experiment shown in (A). Vmax from a
control experiment is also shown; in untreated muscle fibers Vmax does
not decrease with time.
Table 1. Electrical parameters of control vs. formamide-treated
muscles
Control
(n ¼ 7)
Formamide-
treated (n ¼ 7)
Units
Resting potential (Vm) �67 ± 3 �89 ± 4a mV
Time constant (sm) 75 ± 8 52 ± 10 ms
Length constant (k) 1.5 ± 2.0 2.9 ± 0.5a mm
Input resistance (Ro) 110 ± 9 132 ± 9 kWElectrical diameter (d) 104 ± 13 129 ± 11 lmMembrane resistance (Rm) 1107 ± 208 3396 ± 832a W cm2
Membrane capacitance (Cm) 86 ± 20 20 ± 3a lF/cm2
a Statistically different from control (P < 0.05, Student’s t-test).
170
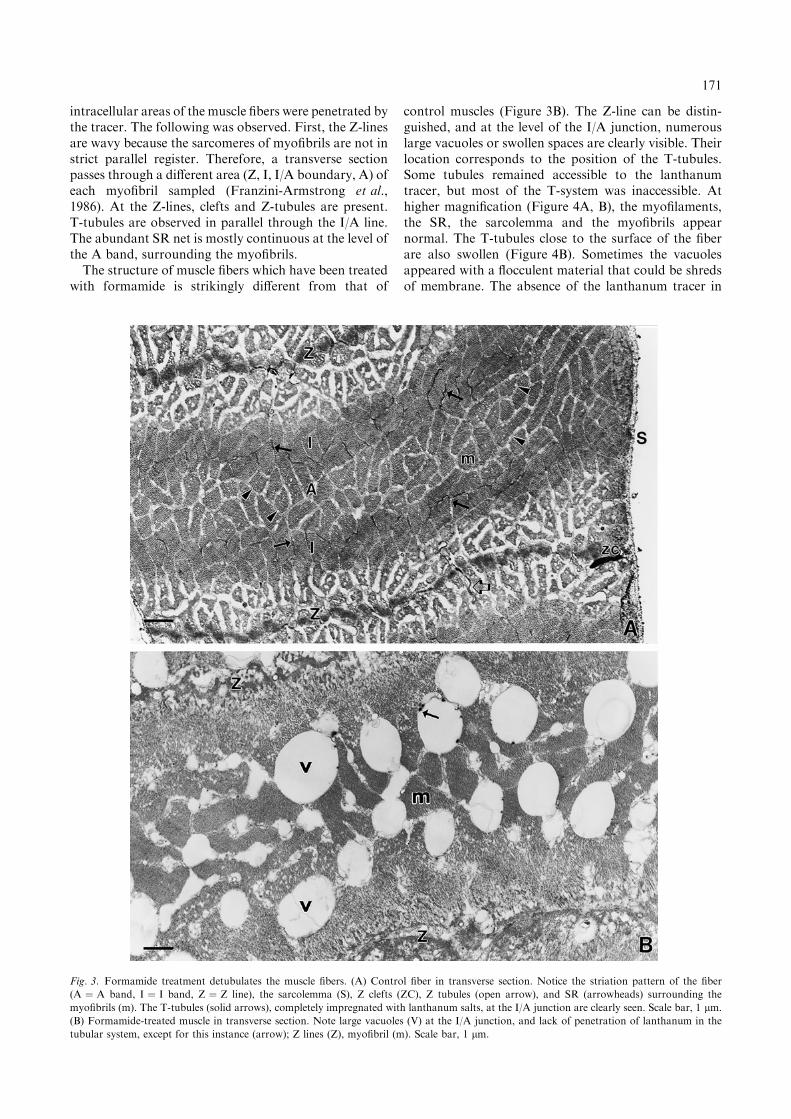
intracellular areas of the muscle fibers were penetrated bythe tracer. The following was observed. First, the Z-linesare wavy because the sarcomeres of myofibrils are not instrict parallel register. Therefore, a transverse sectionpasses through a different area (Z, I, I/A boundary, A) ofeach myofibril sampled (Franzini-Armstrong et al.,1986). At the Z-lines, clefts and Z-tubules are present.T-tubules are observed in parallel through the I/A line.The abundant SR net is mostly continuous at the level ofthe A band, surrounding the myofibrils.The structure of muscle fibers which have been treated
with formamide is strikingly different from that of
control muscles (Figure 3B). The Z-line can be distin-guished, and at the level of the I/A junction, numerouslarge vacuoles or swollen spaces are clearly visible. Theirlocation corresponds to the position of the T-tubules.Some tubules remained accessible to the lanthanumtracer, but most of the T-system was inaccessible. Athigher magnification (Figure 4A, B), the myofilaments,the SR, the sarcolemma and the myofibrils appearnormal. The T-tubules close to the surface of the fiberare also swollen (Figure 4B). Sometimes the vacuolesappeared with a flocculent material that could be shredsof membrane. The absence of the lanthanum tracer in
Fig. 3. Formamide treatment detubulates the muscle fibers. (A) Control fiber in transverse section. Notice the striation pattern of the fiber
(A ¼ A band, I ¼ I band, Z ¼ Z line), the sarcolemma (S), Z clefts (ZC), Z tubules (open arrow), and SR (arrowheads) surrounding the
myofibrils (m). The T-tubules (solid arrows), completely impregnated with lanthanum salts, at the I/A junction are clearly seen. Scale bar, 1 lm.
(B) Formamide-treated muscle in transverse section. Note large vacuoles (V) at the I/A junction, and lack of penetration of lanthanum in the
tubular system, except for this instance (arrow); Z lines (Z), myofibril (m). Scale bar, 1 lm.
171
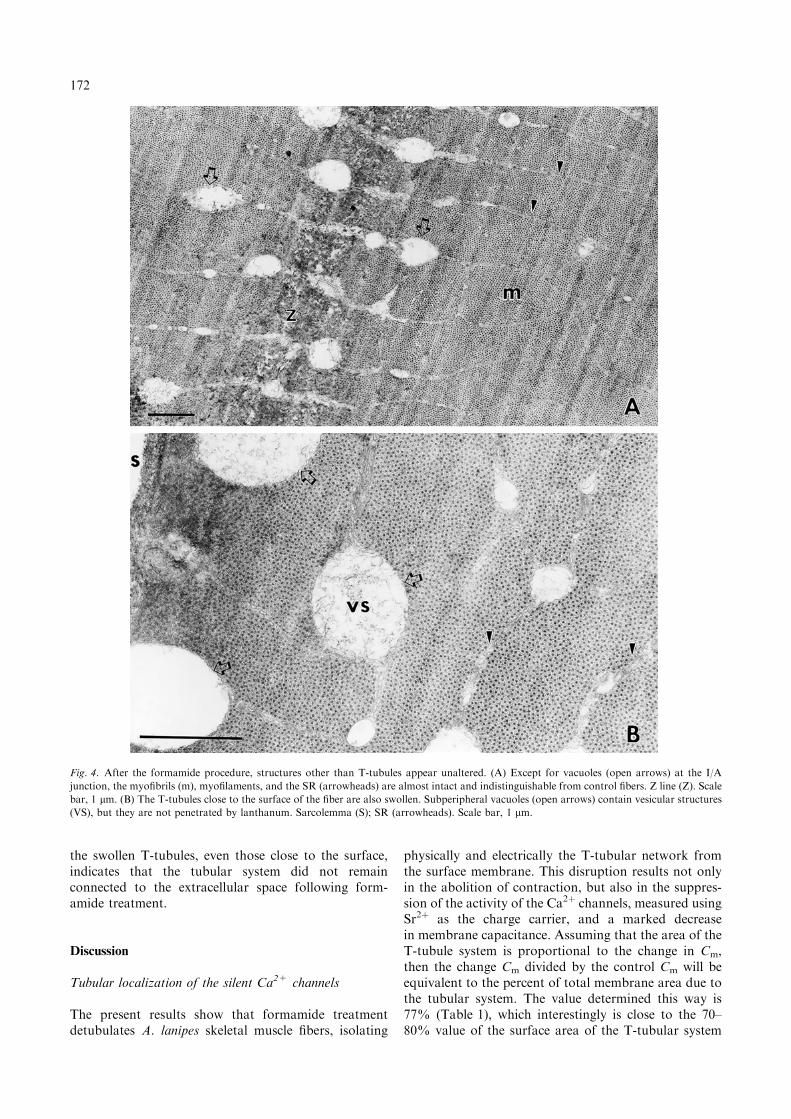
the swollen T-tubules, even those close to the surface,indicates that the tubular system did not remainconnected to the extracellular space following form-amide treatment.
Discussion
Tubular localization of the silent Ca2+ channels
The present results show that formamide treatmentdetubulates A. lanipes skeletal muscle fibers, isolating
physically and electrically the T-tubular network fromthe surface membrane. This disruption results not onlyin the abolition of contraction, but also in the suppres-sion of the activity of the Ca2þ channels, measured usingSr2þ as the charge carrier, and a marked decreasein membrane capacitance. Assuming that the area of theT-tubule system is proportional to the change in Cm,then the change Cm divided by the control Cm will beequivalent to the percent of total membrane area due tothe tubular system. The value determined this way is77% (Table 1), which interestingly is close to the 70–80% value of the surface area of the T-tubular system
Fig. 4. After the formamide procedure, structures other than T-tubules appear unaltered. (A) Except for vacuoles (open arrows) at the I/A
junction, the myofibrils (m), myofilaments, and the SR (arrowheads) are almost intact and indistinguishable from control fibers. Z line (Z). Scale
bar, 1 lm. (B) The T-tubules close to the surface of the fiber are also swollen. Subperipheral vacuoles (open arrows) contain vesicular structures
(VS), but they are not penetrated by lanthanum. Sarcolemma (S); SR (arrowheads). Scale bar, 1 lm.
172

determined for long-sarcomere tonic fibers of thecrayfish by stereological methods (Rossner and Sher-man, 1978: Uhrik et al., 1980). Because the formamideinduced decrease in Cm correlates well with the 80%decrease in inward current (Vmax, Figure 2), the datasuggest that the Ca2þ channels are uniformly distributedin the surface/T-tubule membranes. Furthermore, be-cause the surface area of the T-tubules accounts for 77%of the total membrane area, the results suggest that closeto 80% of the Ca2þ channels are located in the tubules.These are the channels responsible for increasing thelocal concentration of Ca2þ in the vicinity of the SRCa2þ release channels, that triggers the highly amplifiedrelease of Ca2þ required for tension generation. Due tothe large diameter of the muscle fibers in this prepa-ration (Zuazaga and del Castillo, 1985; this work,Table 1), Ca2þ channels in the surface membrane aretoo far away from the SR to participate in CICR. Thesechannels, although not involved in E–C coupling,contribute to the electrogenicity of the muscle fibers,and are responsible for the low-amplitude responsesobserved in detubulated fibers using Sr2þ as the currentcarrier (Figure 2A).
Comparison to previous work
The present study extends previous work in two ways.First, our results demonstrate that formamide cansuccessfully be used to detubulate crustacean musclefibers. The effects of formamide treatment on themechanical, electrophysiological and ultrastructuralproperties of A. lanipes muscle fibers are very similarto those of glycerol, the solute most commonly usedto produce an osmotic shock that disrupts the cou-pling between the T-tubules and the surface mem-brane of crustacean muscles (Fujino et al., 1972; Papir,1973; Castel and Papir, 1975; Zacharova and Uhrik,1978). We used glycerol in an earlier series of experi-ments (data not shown), but formamide was preferedbecause of its speed of action. As reported by others, wefound that uncoupling of excitation from contractiontakes 1 h exposure to glycerol (400 mM) and then90 min to 2 h re-exposure to physiological saline (Za-charova and Uhrik, 1978). Probably because of thegreater membrane permeability of formamide (Argiro,1981; Kawai et al., 1999), this procedure requires lesstime and is more reliable than the usual glycerol shockmethod.Second, the observation that detubulation suppresses
the activity of the Ca2þ channels provides directevidence for the tubular site of the coupling of Ca2þ
entry across the membrane to Ca2þ release from the SRin crustacean muscle fibers. Previous detubulationstudies were designed to examine changes in tensiongeneration, passive electrical constants, and ultrastruc-ture of the fibers, and have not addressed this issue(Fujino et al., 1972; Papir, 1973; Castel and Papir, 1975;Zacharova and Uhrik, 1978).
Implications for E–C coupling
The main conclusion of the present results is that theCa2þ channels in the superficial abdominal flexor musclefibers of A. lanipes are located mainly in the T-tubularsystem. Because under physiological conditions thesefibers are electrically inexcitable (Zuazaga and delCastillo, 1985; Lizardi et al., 1992; Monterrubio et al.,2000), we use the term silent Ca2þ channels to describethem. The tubular localization of the silent Ca2þ
channels ensures their proximity to the SR Ca2þ releasechannels in the dyad. These results provide furtherevidence to support the local control mechanism ofCICR in crustacean muscle as proposed (Gyorke andPalade, 1992, 1993; Monterrubio et al., 2000).
Acknowledgements
We are grateful to Drs L.F. Santana and A.I. Selverstonfor a careful reading of the manuscript and usefulsuggestions. This work was supported by NationalInstitutes of Health grants NS-07464, RR-03051, GM-08224, and National Science Foundation grant HRD-9353130.
References
Argiro V (1981) Excitation–contraction uncoupling of striated muscle
fibres by formamide treatment: evidence of detubulation. J Muscle
Res Cell Motil 2: 283–294.
Ashcroft FM and Stanfield PR (1982) Calcium and potassium currents
in muscle fibres of an insect (Carausius morosus). J Physiol 323: 93–
115.
Ashley CC and Lea TJ (1978) Calcium fluxes in single muscle fibers
measured with a glass scintillator probe. J Physiol 282: 307–331.
Ashley CC and Ridgway EB (1970) On the relationships between
membrane potential, calcium transient and tension in single
barnacle muscle fibers. J Physiol 209: 105–130.
Atwater I, Rojas E and Vergara J (1974) Calcium influxes and tension
in perfused single barnacle muscle fibers under potential control.
J Physiol 243: 523–551.
Bonilla M, Garcıa MC, Orkand PM and Zuazaga C (1992) Ultra-
structural and mechanical properties of electrically inexcitable
skeletal muscle fibers of the crustacean Atya lanipes. Tissue and
Cell 24: 525–535.
Caputo C and Di Polo R (1978) Contractile phenomena in voltage
clamped barnacle muscle. J Gen Physiol 71: 467–488.
Castel M and Papir D (1975) The ultrastructure of normal and glycerol
treated muscle in the ghost crab, Ocypode cursor. Cell Tiss Res 159:
369–378.
del Castillo J and Escalona de Motta G (1978) A new method for
excitation–contraction uncoupling in frog skeletal muscle. J Cell
Biol 78: 782–784.
Erxleben C and Rathmayer W (1997) A dihydropyridine-sensitive
voltage-dependent calcium channel in the sarcolemmal membrane
of crustacean muscle. J Gen Physiol 109: 313–326.
Escalona de Motta G, Smith DS, Cayer M and del Castillo J (1982)
Mechanism of the excitation–contraction uncoupling of frog
skeletal muscle by formamide. Biol Bull 163: 276–286.
Fabiato A (1983) Calcium induced release of calcium from the cardiac
sarcoplasmic reticulum. Am J Physiol 245: C1–C14.
Fahimi HD and Cotran RS (1971) Permeability studies in heat-
induced injury of skeletal muscle using lanthanum as a fine
structural tracer. Am J Pathol 62: 143–157.
173

Falk G and Fatt P (1964) Linear electrical properties of striated muscle
fibers observed with intracellular electrodes. Proc Roy Soc Ser B
160: 69–123.
Fatt P and Ginsborg BL (1958) The ionic requirements for the
production of action potentials in crustacean muscle fibers.
J Physiol 142: 516–543.
Fatt P and Katz B (1953) The electrical properties of crustacean muscle
fibers. J Physiol 120: 171–204.
Franzini-Armstrong C, Eastwood AB and Peachey LD (1986) Shape
and disposition of clefts, tubules, and sarcoplasmic reticulum in
long and short sarcomere fibers of crab and crayfish. Cell Tiss Res
244: 9–19.Fujino M, Yamaguchi T and Fujino S (1972) ‘Glycerol effect’ in
various kinds of muscle cell. Jpn J Physiol 22: 477–489.
Gainer H (1968) The role of calcium in excitation–contraction
coupling of lobster muscle. J Gen Physiol 52: 88–110.
Gilly WF and Scheuer T (1984) Contractile activation in scorpion
striated muscle fibers. J Gen Physiol 84: 321–345.
Gilly WF and Scheuer T (1993) Voltage-dependent calcium and
potassium conductances in striated muscle fibers from the scorpi-
on, Centuroides sculpturatus. J Membr Biol 134: 155–167.
Gomez AM, Valdivia HH, Cheng H, Lederer MR, Santana LF,
Cannell MB, Mc Cune SA, Altschuld RA and Lederer WJ (1997)
Defective excitation–contraction coupling in experimental cardiac
hypertrophy and heart failure. Science 276: 800–806.Gyorke S and Palade P (1992) Calcium-induced calcium release in
crayfish skeletal muscle. J Physiol 457: 195–210.
Gyorke S and Palade P (1993) Role of local Ca2þ domains in
activation of Ca2þ-induced Ca2þ release in crayfish muscle. Am J
Physiol 33: C1505–C1512.
Gyorke S and Palade P (1994) Ca2þ-dependent negative control mech-
anism for Ca2þ release in crayfish muscle. J Physiol 476: 315–
322.
Hagiwara S and Naka K-I (1964) The initiation of spike potential in
barnacle muscle fibers under low intracellular Ca2þ. J Gen Physiol
48: 141–162.Hagiwara S and Takahashi K (1967) Surface density of calcium ion
and calcium spikes in the barnacle muscle fiber membrane. J Gen
Physiol 50: 583–601.
Hencek M and Zachar J (1965) The electrical constants of single
muscle fibers of the crayfish (Astacus fluviatilis). Physiol Bohemo-
slov 14: 297–311.
Hencek M and Zachar J (1977) Calcium currents and conductances in
the muscle membrane of the crayfish. J Physiol 268: 51–57.
Hodgkin AL and Rushton WAH (1946) The electrical constants of a
crustacean nerve fibre. Proc Roy Soc Ser B 133: 444–479.
Kawai M, Hussain M and Orchard CH (1999) Excitation–contraction
coupling in rat ventricular myocytes after formamide-induced
detubulation. Am J Physiol 277: H603–H609.
Keynes RD, Rojas E, Taylor RE and Vergara J (1973) Calcium and
potassium systems of a giant barnacle muscle fiber under mem-
brane potential control. J Physiol 229: 409–455.
Lizardi L, Garcıa MC, Sanchez JA and Zuazaga C (1992) Sulfhydryl
alkylating agents induce calcium current in skeletal muscle fibers of
a crustacean (Atya lanipes). J Membr Biol 129: 167–178.
Monterrubio J, Lizardi L and Zuazaga C (2000) Silent calcium
channels in skeletal muscle fibers of the crustacean Atya lanipes.
J Membr Biol 173: 9–17.
Monterrubio J, Ortiz G, Orkand PM and Zuazaga C (1999) Tubular
localization of Ca2þ channels in crustacean skeletal muscle fibers.
Biophys J 76: A298.
Mounier Y and Goblet C (1987) The role of different calcium sources
in the excitation–contraction coupling in crab muscle fibres. Can
J Physiol Pharmacol 65: 667–671.
Mounier Y and Vassort G (1975) Initial and delayed membrane
currents in crab muscle fiber under voltage-clamp conditions.
J Physiol 251: 589–608.
Palade P and Gyorke S (1993) Excitation–contraction coupling in
crustacea: do studies on these primitive creatures offer insights
about EC coupling more generally? J Muscle Res Cell Motil 14:
283–287.
Papir D (1973) The effect of glycerol treatment on crab muscle fibres.
J Physiol 230: 313–330.
Putnam RW (1996) Intracellular pH regulation in detubulated frog
skeletal muscle fibers. Am J Physiol 271: C1358–C1366.Rossner KL and Sherman RG (1978) Invaginated membrane in
crustacean tonic muscle fibers: estimates of membrane capacitance.
Am J Physiol 235: C220–C226.
Uhrik B, Novotova M and Zachar J (1980) A quantitative estimation
of components in crayfish muscle fibres by stereological methods.
Pflugers Arch 387: 281–286.
van Harreveld A (1936) A physiological solution for freshwater
crustaceans. Proc Soc Exp Biol Med 34: 428–432.
Vergara J and Verdugo P (1988) Calcium transients in voltage clamped
giant barnacle muscle fibers. Biophys J 53: 647a.
Washio H (1972) The ionic requirements for the initiation of
action potentials in insect muscle fibers. J Gen Physiol 59: 121–
134.
Zacharova D and Uhrik B (1978) Kinetics of functional and
morphological changes during decoupling and recoupling induced
by glycerol in isolated muscle fibres of the crayfish. Cell Tiss Res
192: 167–185.
Zacharova D and Zachar J (1967) The effect of external calcium ions
on the excitation–contraction coupling in single muscle fibers of
the crayfish. Physiol Bohemoslov 16: 191–207.
Zuazaga C and del Castillo J (1985) Generation of calcium action
potentials in crustacean muscle fibers following exposure to
sulfhydryl reagents. Comp Biochem Physiol 82C: 409–416.
174