the digestive physiology of herbivorous, omnivorous and carnivorous crustacean larvae: a review
TRANSCRIPT

Aquaculture
ELSEVIER Aquaculture 155 (1997) 285-295
The digestive physiology of herbivorous, omnivorous and carnivorous crustacean larvae: a
review
D.A. Jones a, * , M. Kumlu b, L. Le Vay a, D.J. Fletcher a ’ School qf Ocmn Sciences. University of Wales, Bangor. Menai Bridge, Gwynedd, N. Wales, LLSY 5EY, UK
h School of Fisheries, Unirersitrsi ‘Cukurora, 01330, Bulculi. Admu, Turke?
Abstract
While most copepods are holoplanktonic, decapod larvae are meroplanktonic with a pelagic larval development ranging from days (Penaeidae) to weeks (most Palaemonidae, Palinuridae). Reproductive strategies result in either the early release of larvae in large numbers of small planktonic forms (Penaeidae) or smaller numbers of advanced larvae after parental incubation (Brachyura, Palaemonidae, Nephropidae, Palinuridae). Commercially-cultured decapod larvae exhibit a wide range of feeding strategies exploiting most of the trophic levels found within the planktonic ecosystem. Studies on these crustacean larvae demonstrate how their digestive physiol- ogy is adapted to different feeding strategies during larval development, and provide an insight into the design of appropriate artificial feeds for commercial culture. Comparative measurements of digestive enzyme levels reveal that trypsin-like protease appears to dominate in all larvae investigated. Highest levels occur in herbivorous penaeid and brachyuran larval stages. In contrast, carnivorous lobster and caridean larvae show low protease activity at first feeding and appear to rely upon high-energy digestible live prey for their nutrition. Ontogenetic changes in enzyme type, activity and content are displayed during the penaeid mysis and caridean, brachyuran late zoeal stages as larvae transfer to higher trophic levels. The range and extent of these changes are reviewed for the commercial larval groups of commercially important species. 0 1997 Elsevier Science B.V.
Ke.wvords: Review; Digestive enzymes; Crustacean larvae
* Corresponding author. Tel.: + 44-1248-382863; fax: + 44. I248 7 16367.
0044.8486/97/$17.00 0 1997 Elsevier Science B.V. All rights reserved
PI/ SOO44-8486(97)00 129-4
D.A. Jones et al./Aquaculture 155 (1997) 285-295
1. Introduction
Commercial crustacean culture encompasses a wide range of species, some used as live feeds (copepods, Artemia) others (penaeids, carideans) forming the mainstay of food production. Homarid, palinurid and brachyuran species are cultured for stock enhancement programmes, and there is increasing interest in carideans such as Lysmatu
and Stenopus in the marine ornamental trade. These species represent diverse crustacean groups which exhibit differences reflected in diverse larval forms (Table 1).
Groups such as the Branchiopoda ( Artemia), Copepoda (Tigriupus) and Penaeoidea (Penaeus) typically produce large numbers of eggs which hatch into a nauplius larva which often begins as a lecithotroph (Artemia, Penaeus) but becomes phytotrophic within days. In contrast, most decapods incubate eggs and release an advanced zoea larva which is usually zootrophic at first feeding. Time spent in the planktonic form varies greatly within groups and species (Table l), providing one of the greatest problems to the culturist. However, common to all of these differing larval forms is the need to extract nutrition from the primary and secondary production levels in the plankton.
The morphology of the gut of adult and larval crustaceans has been recently reviewed by Icely and Nott (1992) and by Jones et al. (in press), respectively. Functional larval morphology during ontogenetic development appears to be similar for most decapod larvae, with mouthparts used for food consumption while the proventriculus remains simple until the late larval stages. As most larvae are dependent upon enzymatic breakdown of ingested food, development of secretory hepatopancreatic tissue dictates the type of prey which can be consumed.
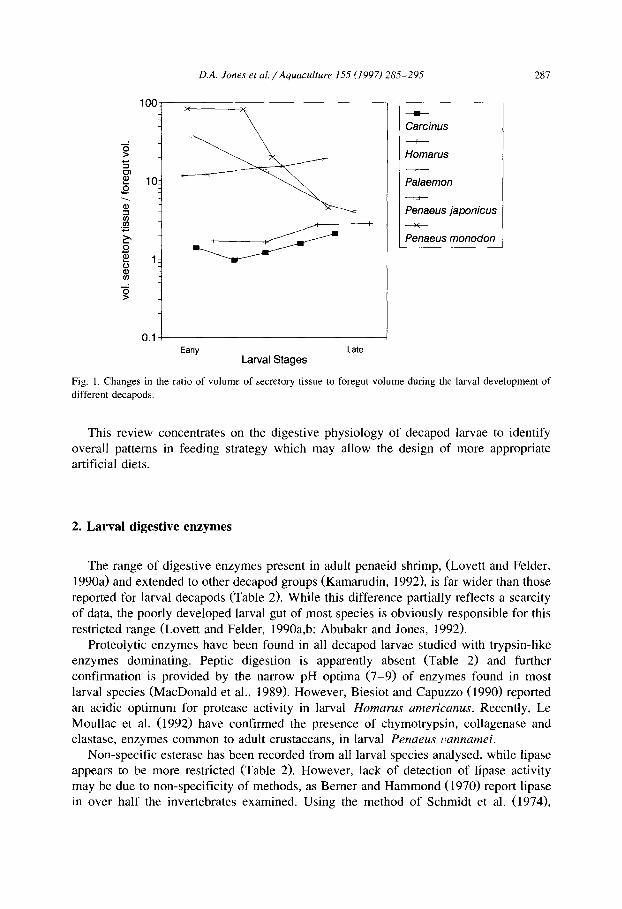
In Fig. 1 the ratio of enzyme secretory tissue to foregut volume for different species of decapod larvae and stages is presented (data from Deru, 1990; Abubakr, 1991). The presence of anterior midgut diverticulae (AMD) gives protozoea and early mysis larvae of penaeids an advantage that is lost during the later mysis and early postlarval stages (Lovett and Felder, 1990a) when these diverticulae regress and hepatopancreas/foregut volumes approach ratios seen in other decapod groups (Fig. 1).
Table I The number, duration and trophic level of larval stages for cultured crustacean groups. H: herbivorous, 0:
omnivorous; C: carnivorous. f: few; m: most
Group Stages Duration Trophic
Nauplius/ Protozoea Zoea Mysis Megalopa (days) level
metanauplius
Branchiopoda 11 0 0 0 11-15 H-O Copepoda 5-6 5 0 0 10-30 H-,O,C
Penaeoidea 5-6 (8) 3 2-5 Postlarva 9-15 H-tO,C
Brachyura 0 3-6 1 15-30 0, c
Caridea 0 (0-4)(f) 5-9(m)-20(f) 1 (2-3) 14-150 0, c
Palinuroidea 0 6-15 1 90-330 c
Nephropoidea 0 3-4 1 15-45 c
D.A. Jones et al. /Aquaculture 155 (1997) 285-295 287
Carcinus
Homarus
Palaemon
Penaeus japonicus
0.1’ Early Late
Larval Stages
Fig. I, Changes in the ratio of volume of secretory tissue to foregut volume during the larval development of
different decapods.
This review concentrates on the digestive physiology of decapod larvae to identify overall patterns in feeding strategy which may allow the design of more appropriate artificial diets.
2. Larval digestive enzymes
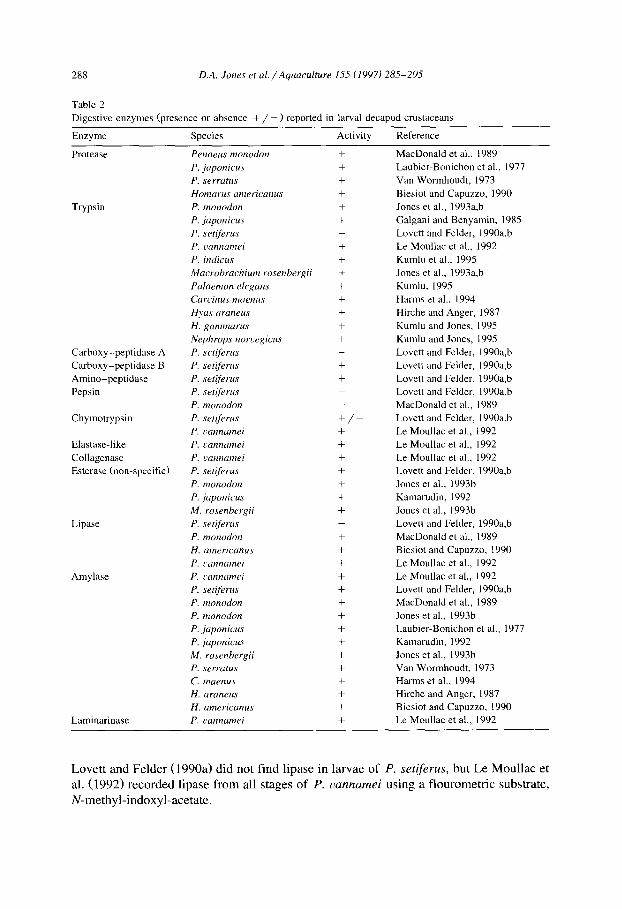
The range of digestive enzymes present in adult penaeid shrimp, (Lovett and Felder, 1990a) and extended to other decapod groups (Kamarudin, 1992), is far wider than those reported for larval decapods (Table 2). While this difference partially reflects a scarcity of data, the poorly developed larval gut of most species is obviously responsible for this restricted range (Lovett and Felder, 1990a,b; Abubakr and Jones, 1992).
Proteolytic enzymes have been found in all decapod larvae studied with trypsin-like enzymes dominating. Peptic digestion is apparently absent (Table 2) and further confirmation is provided by the narrow pH optima (7-9) of enzymes found in most larval species (MacDonald et al., 1989). However, Biesiot and Capuzzo (1990) reported an acidic optimum for protease activity in larval Homarus americanus. Recently, Le Moullac et al. (1992) have confirmed the presence of chymotrypsin, collagenase and elastase, enzymes common to adult crustaceans, in larval Penaeus uannamei.
Non-specific esterase has been recorded from all larval species analysed, while lipase appears to be more restricted (Table 2). However, lack of detection of lipase activity may be due to non-specificity of methods, as Bemer and Hammond (1970) report lipase in over half the invertebrates examined. Using the method of Schmidt et al. (1974),
288
Table 2
D.A. Jones et al./Aquaculture 155 (19971285-295
Digestive enzvmes (aresence or absence + / - 1 reuorted in larval decanod crustaceans
Enzyme Species Activity Reference
Protease
Trypsin
Carboxy-peptidase A
Carboxy-peptidase B
Amino-peptidase
Pepsin
Chymotrypsin
Elastase-like
Collagenase
Esterase (non-specific)
Lipase
Amylase
Laminarinase
Penaeus monodon
P. japonicus
P. serratus
Homarus americanus
P. monodon
P. japonicus
P. setiferus
P. wnnamei
P. indicus
Macrohrachium rosenbergii
Palaemon elegans
Carcinus maenus
Hws araneus
H. gammarus
Nephrops nowegicus
P. setiferus
P. setiferu
P. setiferus
P. setiferus
P. monodon
P. setifeiws
P. L,annamei
P. c,annamei
P. cannamei
P. .setiferu.s
P. monodon
P. japonicus
M. rosenhergii
P. setiferus
P. monodon
H. americanus
P. cannamei
P. cannamei
P. setiferus
P. monodon
P. monodon
P. japonicus
P. japonicus
M. rosenbergii
P. serratus
C. maenus
H. crraneus
H. americanus
P. cannamei
_
+ + + + + + + + + + + + + + + + + + _ -
+/ + + + + + + + _ + + + + + + + + + + + + + + t
MacDonald et al., 1989
Laubier-Bonichon et al., 1977
Van Wormhoudt, 1973
Biesiot and Capuzzo, 1990
Jones et al., 1993a,b
Galgani and Benyamin, 1985
Lovett and Felder, 199Oa.b
Le Moullac et al., 1992
Kumlu et al., 1995
Jones et al., 1993a,b
Kumlu, 1995
Harms et al.. 1994
Hirche and Anger, 1987
Kumlu and Jones, 1995
Kumlu and Jones, 1995
Lovett and Felder, 1990a,b
Lovett and Felder, 1990a,b
Lovett and Felder, 1990a,b
Lovett and Felder, 1990a,b
MacDonald et al., 1989
Lovett and Felder, I990a,b
Le Moullac et al., 1992
Le Moullac et al., 1992
Le Moullac et al., 1992
Lovett and Felder, l990a,b
Jones et al., 1993b
Kamarudin, 1992
Jones et al., l993b
Lovett and Felder, 1990a,b
MacDonald et al., 1989
Biesiot and Capurao, 1990
Le Moullac et al., 1992
Le Moullac et al., 1992 Lovett and Felder, 1990a,b
MacDonald et al., 1989
Jones et al., 1993b
Laubier-Bonichon et al., 1977
Kamarudin, 1992
Jones et al., 1993b
Van Wormhoudt, 1973
Harms et al., 1994 Hirche and Anger, 1987
Biesiot and Capuzzo, 1990
Le Moullac et al., 1992
Lovett and Felder (1990a) did not find lipase in larvae of P. setiferus, but Le Moullac et al. (1992) recorded lipase from all stages of P. uannamei using a flourometric substrate, N-methyl-indoxyl-acetate.

D.A. Jones et al./Aqtmculture 155 (19971285-295 289
2.1. Estimation of digestice enzymes
Detection and measurement of the activity levels of larval enzymes has proved difficult due to the small amounts of material usually available and the non-specificity of substrates (see above and Lovett and Felder (1990a) for review). In addition, a wide range of units have been used to express activity, including activity relative to hepatopancreas weight, hepatopancreas protein, soluble or whole body protein and whole body wet or dry weight. Total enzyme activity (units/larva> provides valuable data concerning the digestive capacity of the whole animal and is useful in evaluating the contribution made by live feeds (Munilla-Moran et al., 1990). However, activity must be related to protein content or body weight when comparing larvae of different sizes or species. Excision of the gut is impossible in most larval crustaceans and use of whole tissue homogenates is the preferred method for sample preparation. Although this method introduces the potential for inhibition of activity of the enzyme being assayed, trypsin content in P. juponicus larvae measured by radioimmunoassay appears to correlate well with activity measured with a specific substrate (Galgani and Benyamin, 1985). Lovett and Felder (1990a) have demonstrated that most trypsin. esterase and amylase activity emanates from the gut rather than non-gut tissue, further justifying the use of whole larval homogenates. These investigators also found that. apart from the nauplius and first protozoea stages, little ontogenetic change in soluble protein levels occurred in P. setiferus. This observation allows comparison between measurements in
terms of activity mg-’ dry weight. The use of different substrates, temperatures and buffers and the expression of
digestive enzyme activity in different units confounds comparison of data from different laboratories. Furthermore, fluctuations in enzyme activity relative to daily metabolic rhythms, stage of moult (Harms et al., 1991) and season (Biesiot and Capuzzo, 1990) may be expected. Direct comparison of activities of different enzymes within one species must also be conducted cautiously, because different assay methods may be used and activities expressed in units for which the relative physiological significance is not clear.
2.2. Ratio of difSerent enzymes within larcae
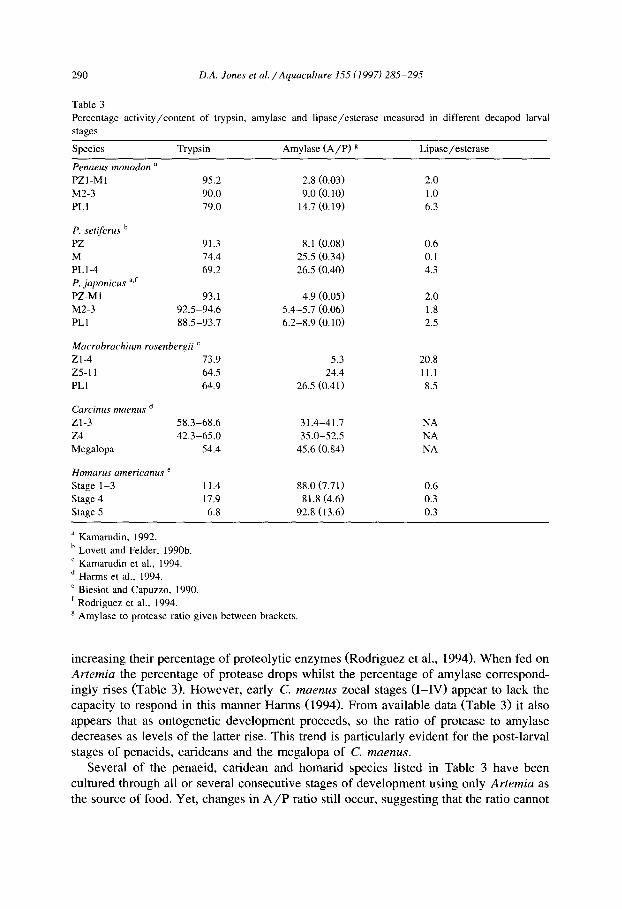
Recognizing the previously stated limitations, the percent occurrence of trypsin, amylase and lipase/esterase measured within different larval species and stages in several studies is presented in Table 3. Without exception those species which princi- pally consume a diet of microalgae contain the highest percentage of proteolytic enzyme. All penaeid phytotrophic larval stages, while differing slightly among species show an apparent predominance of trypsin content amongst digestive enzymes mea- sured. In contrast, for carnivorous larvae such as H. americanus a higher ratio of amylase to protease (A/P ratio) is apparent. Early stage Mucrobruchium rosenbergii and Carcinus maenms, which also rely upon Artemia for successful larval development, maintain intermediate levels of trypsin.
In response to the exclusive feeding of algae, larvae of the same stage within individual species, such as P. japonicus mysis and C. maenus zoea IV, respond by
290 D.A. Jones et al./Aquaculture 155 (1997) 285-295
Table 3 Percentage activity/content of trypsin, amylase and lipase/esterase measured in different decapod larval
stages
Species Trypsin Amylase (A/P) g Lipase/esterase
Penaeus monodon a
PZI-Ml
M2-3
PLI
P. setijerus b
PZ
M
PLl-4
P. japonicus a,f
PZ-M 1
M2-3
PLI
95.2 2.8 (0.03)
90.0 9.0 (0.10) 79.0 14.7 (0.19)
91.3 8.1 (0.08) 0.6
14.4 25.5 (0.34) 0.1 69.2 26.5 (0.40) 4.3
93.1 4.9 (0.05) 2.0
92.5-94.6 5.4-5.7 (0.06) 1.8
88.5-93.7 6.2-8.9 (0.10) 2.5
Macrobrachium rosenbergii ’
21-4 73.9
25-l 1 64.5
PLI 64.9
Carcinus maenus d
21-3 58.3-68.6
24 42.3-65.0
Megalopa 54.4
Homarus americanus e
Stage l-3 11.4
Stage 4 17.9
Stage 5 6.8
5.3 20.8
24.4 11.1
26.5 (0.41) 8.5
31.4-41.7 NA
35.0-52.5 NA
45.6 (0.84) NA
88.0 (7.71) 0.6 81.8 (4.6) 0.3
92.8 (13.6) 0.3
2.0
1.0
6.3
a Kamarudin, 1992.
b Lovett and Felder, 1990b.
’ Kamarudin et al., 1994.
d Harms et al., 1994.
’ Biesiot and Capuzzo, 1990.
’ Rodriguez et al., 1994.
g Amylase to protease ratio given between brackets.
increasing their percentage of proteolytic enzymes (Rodriguez et al., 1994). When fed on Artemia the percentage of protease drops whilst the percentage of amylase correspond- ingly rises (Table 3). However, early C. maenu~ zoeal stages (I-IV) appear to lack the capacity to respond in this manner Harms (1994). From available data (Table 3) it also appears that as ontogenetic development proceeds, so the ratio of protease to amylase decreases as levels of the latter rise. This trend is particularly evident for the post-larval stages of penaeids, carideans and the megalopa of C. maenus.
Several of the penaeid, caridean and homarid species listed in Table 3 have been cultured through all or several consecutive stages of development using only Artemia as the source of food. Yet, changes in A/P ratio still occur, suggesting that the ratio cannot
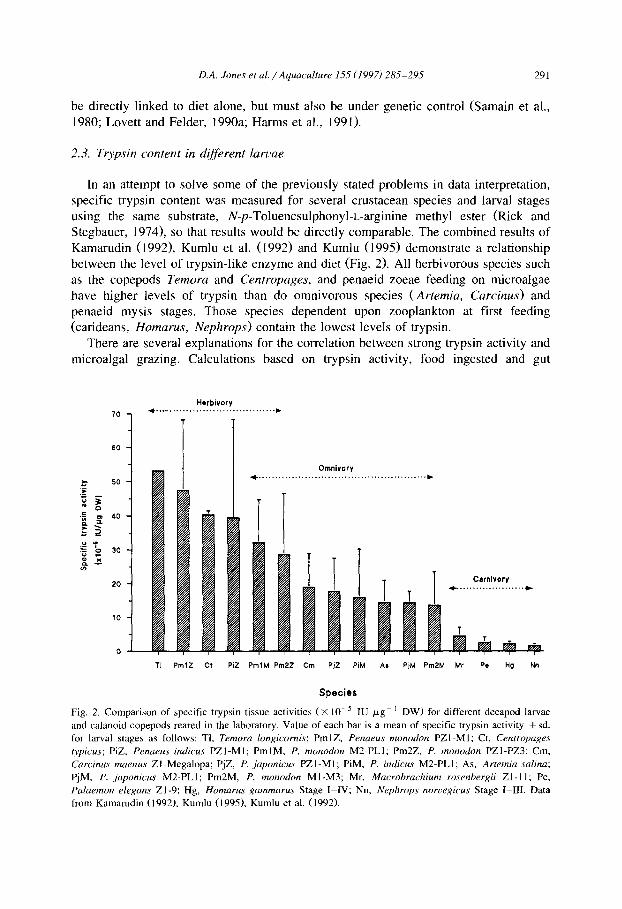
D.A. Jones et al./Aquaculture 155 (1997) 285-295 291
be directly linked to diet alone, but must also be under genetic control (&main et al., 1980; Lovett and Felder, 1990a; Harms et al., 1991).
2.3. Trypsin content in different larvae
In an attempt to solve some of the previously stated problems in data interpretation, specific trypsin content was measured for several crustacean species and larval stages using the same substrate, N-p-Toluenesulphonyl-L-arginine methyl ester (Rick and Stegbauer, 1974), so that results would be directly comparable. The combined results of Kamarudin (1992), Kumlu et al. (1992) and Kumlu (1995) demonstrate a relationship between the level of trypsin-like enzyme and diet (Fig. 2). All herbivorous species such as the copepods Temora and Centropages, and penaeid zoeae feeding on microalgae have higher levels of trypsin than do omnivorous species (Artemia, Curcinus) and penaeid mysis stages. Those species dependent upon zooplankton at first feeding (carideans, Homarus, Nephrops) contain the lowest levels of trypsin.
There are several explanations for the correlation between strong trypsin activity and microalgal grazing. Calculations based on trypsin activity, food ingested and gut
0
Herbivory *..._........,........................
Omnivorv *..,.....,................,.........................~
TI Pm12 Ct Pi2 PmlM Pm22 Cm PjZ PM As PJM Pm2M Mr Pa Hp Nn
Species
Fig. 2. Comparison of specific trypsin tissue activities (X 10-j IU pg- ’ DW) for different decapod larvae
and calanoid copepods reared in the laboratory. Value of each bar is a mean of specific trypsin activity + sd.
for larval stages as follows: Tl, Temora longicornis; PmlZ, Penaeus monodon PZl-Ml; Ct, Centropages
typicus; PiZ, Penaeus indicus PZl-Ml; PmlM, P. monodon M2-PLl; Pm2Z, P. monodon PZl-PZ3; Cm,
Carcinus maenus Zl-Megalopa; PjZ, P. japonicus PZl-MI; PiM, P. indicus MZ-PLl; As, Artemia salina;
PjM, P. japonicus M2-PLl; Pm2M, P. monodon Ml-M3; Mr. Macrohrachium rosenbergii 21-11; Pe,
Palaemon elegans Zl-9; Hg, Homarus gammarus Stage I-IV; Nn, Nephrops norvegicus Stage I-III. Data from Kamarudin (1992). Kumlu (1995). Kumlu et al. (1992).

292 D.A. Jones et al./Aquaculture 155 (1997) 285-295
residence time demonstrate that a balance is achieved for larvae at each trophic level (Jones et al., 1993b; Jones, in press). Algal food tends to have a variable biochemical composition that is dependant on temporal and environmental factors. Carbon-nitrogen (C:N) ratios in rapidly dividing phytoplankton are typically between 6: 1 - 10: 1, but can approach the upper limit (17: 1) considered to be suitable to support healthy animal growth when cells are senescent (see review by Parsons et al., 1977). Moreover, the absolute protein content of phytoplankton, particularly silicaceous diatoms, may be lower than that of zooplankton. Crustacean larvae require adequate dietary protein to maintain a tissue C:N ratio of approximately 4-5:l during development (Anger, 1988; Le Vay et al., 1993), as well as to provide an energy source. Most herbivorous feeders have high ingestion rates and short gut retention times, resulting in high rates of turnover (Kurmaly et al., 1989). Thus, for penaeid protozoea, an intake of up to eight times larval body weight per day is processed (Jones et al., in press), a strategy of high enzyme activity extracting sufficient protein to support metabolism and growth.
For species feeding above the first trophic level, ingested food consists of a protein content and C:N ratio similar to larval tissue and extraction of adequate protein can be achieved with lower trypsin activity. Carnivorous larvae such as Homarus and Nephrops
(Fig. 2) compensate for low trypsin levels by greatly extending food retention times (up to 16 h) and rigorous selection of high energy, highly digestible prey (Kurmaly et al., 1990). Accordingly, the A/P ratio is dictated by trypsin activity requirements which are high in herbivorous but low in carnivorous larvae. Energetic loss through metabolism for penaeid zoea is only 3% of assimilated energy as compared to 34% for H. americanus
stage I larvae (Jones et al., in press) demonstrating the advantage promoted by this high turnover, herbivorous strategy.
2.4. Enzyme response to diet
While factors such as genetic control, gut morphology and trophic level act to constrain enzymatic response to diet, abundant evidence indicates that diet may be used to manipulate enzyme activity, at least during late larval stages. To date, successful use of artificial diets for decapod larvae is restricted to penaeid larvae and late stage carideans such as Macrobruchium (Jones et al., 1993a).
Supplementation of artificial feeds with algal extracts has been demonstrated to enhance tryptic activity in penaeid protozoea and to promote increased growth and survival (Kumlu and Jones, in press). These responses resulted from enhanced digestion of the artificial feed. First feeding larvae of other crustacean groups do not respond similarly because without well developed AMD, the capacity to raise enzyme activity levels is absent. An exception is found in the Artemia nauplius stages which possess a complex of proteolytic enzymes (Osuna et al., 1977) that may explain their ability to adapt to artificial feeds.
Greater flexibility in enzymatic response is seen for later stage penaeid, caridean and brachyuran larvae where there are clear changes in levels of enzymatic activity in response to diets of different composition. When artificial diet is fed to late stage M. rosenbergii zoeae (Kumlu and Jones, 19951, and to penaeid mysis larvae (Jones et al., 1993a; Le Vay et al., 1993; Kumlu and Jones, in press) trypsin activity, higher than that

D.A. Jones et al./Aqunculture 155 (1997) 285-295 293
measured in Artemia-fed controls, is invoked. Similar elevation of trypsin activity has been observed in mysis stage P. japonicus (Rodriguez et al., 1994) and the megalopa of Hyas areneus (Harms et al., 1991) when algae are substituted for Artemia.
As gut retention times are longer in all late stage larvae and midgut gland volume increases, lower levels of trypsin activity in Artemia-fed larvae are likely due to the highly digestible nature of this prey (i.e. high substrate availability, and nutrients readily available for assimilation). Thus increase in trypsin activity when other diets are substituted may be interpreted as a response to substrate scarcity or indigestibility (Harris et al., 1986). If this relationship is correct, measurement of tryptic activity in late stage larvae and post-larvae provides an assay for diet digestibility.
Accordingly, the lack of tryptic enzyme reponse to artificial diet seen in early stage caridean, penaeid and other crustacean larvae and more importantly post-larval (PL l- 14)
penaeids (Ribeiro and Jones, 1995) shows the lack of flexibility during these stages. The relatively high secretory tissue/foregut volume ratio in first-feeding penaeid larvae (Fig. l), coupled with rapid gut evacuation rates, allow enough of the more readily assimi- lated components of the feed to be extracted to support survival and growth. However, regression of the AMD in early postlarval stages lowers the capacity for enzyme response, so that digestion of artificial diet is less effective. This change is compounded by longer gut retention times, which although suited for highly efficient assimilation of readily digestible foods, makes little impact upon less digestible artificial feeds. Simi- larly, for those omnivorous and carnivorous larvae (Fig. 2) which lack the physiological capacity to raise enzyme levels in response to artificial diets, use of live food remains the only method of culture.
References
Abubakr, M.A., 199 I. Studies on the functional morphology of the decapod larval gut in relation to diet. PhD
Thesis, University of Wales, Bangor, UK, 230 pp.
Abubakr, M.A., Jones, D.A.. 1992. Functional morphology and ultrastructure of the anterior mid-gut
diverticulae of Penaru.s monodon (Fabricius, 1798) larvae. Crustaceana 62, 142-158.
Anger, K.. 1988. Growth and elemental (C, N, H) composition in Innchus dwsettensis (Decapoda: Majidae) larvae reared in the laboratory. Mar. Biol. 99, 255-260.
Berner, D.L., Hammond. E.G., 1970. Phylogeny of lipase specificity. Lipids 5, 558-562.
Biesiot, P.M., Capuzzo, J.M., 1990. Changes in the digestive enzyme activities during early development of
the American lobster Homarus umericanus Milne Edwards. J. Exp. Mar. Biol. Ecol. 136, 107-122.
Deru, J., 1990. Studies on the development and nutrition of the caridean prawn Mrrcrohrcrchium rosenbergi;
(De Man) (Crustacea: Decapoda). PhD Thesis, University of Wales, Bangor, UK, 306 pp.
Galgani, F.G., Benyamin. Y.. 1985. Radioimmunoassay of shrimp trypsin: application to the larval develop-
ment of Pencreus,juponicus Bate, 1888. J. Exp. Mar. Biol. Ecol. 87, 145-151.
Harms, J.A., Anger, K.. Klaus, S.. Seeger, B., 1991. Nutritional effects on ingestion rate, digestive activity, growth, and biochemical composition of Hyus trraneus L. (Decapoda: Majidae). J. Exp. Mar. Biol. Ecol. 145. 233-265.
Harms, J.. Meyer-Harms, B.. Dawirs. R.R., Anger, K., 1994. Growth and physiology of Cnrcinus marnus
(Decapoda. Portunidae) larvae in the field and in laboratory experiments. Mar. Ecol. Prog. Ser. 108, 107-I 18.
Harris. R.P.. Samain, J.F., Moal, J., Martin-Jezequel, V., Poulet. S.A., 1986. Effects of algal diet on digestive activity in Cc&nu.s helgolandicus. Mar. Biol. 90, 353.

294 D.A. Jones et al./Aquaculture 155 (19971285-295
Hirche, H.J., Anger, K., 1987. Digestive enzyme activities during larval development of Hyas areneus
(Decapoda: Majidae). Comp. Biochem. Physiol. 87B, 297-302.
Rely, J., Nott, J., 1992. Digestion and absorption: digestive system and associated organs. In: Harrison, F.W.,
Humes, A.G. (Eds.), Microscopic Anatomy of Invertebrates, Vol. 10, Decapod Crustacea, Wiley-L&,
New York, pp. 147-201.
Jones, D.A., Larval microparticulate diets. Rev. Fish. Sci., in press.
Jones, D.A., Kamarudin, MS., Le Vay, L., 1993a. The potential for replacement of live feeds in larval culture.
J. World Aquacult. Sot. 24, 199-210.
Jones, D.A., Le Vay, L., Kamarudin, M.S., 1993b. Feeding and nutritional requirements of penaeid shrimp
larvae. In: Caldtron, J., Sandoval, V. (Eds.), Memorias Primer Congress0 Ecuatoriano de Acuicultura, San
Pedro de Manglaralto, Ecuador, CENAIM, pp. 45-52.
Jones, D.A., Yule, A., Holland, D.L., Larval nutrition. In: Conklin, D., D’Abramo, L. (Eds.), Crustacean
Nutrition, World Aquaculture Society, Baton Rouge, LA, in press.
Kamarudin, M.S., 1992. Studies on the digestive physiology of crustacean larvae. PhD Thesis, University of Wales, Bangor, UK, 190 pp.
Kamarudin, MS., Jones, D.A., Le Vay, L., Abidin, A.Z., 1994. Ontogenetic change in digestive enzyme
activity during larval development of Macrobrachium rosenbergii. Aquaculture 123, 323-333.
Kumlu, M., 1995. Physiology of decapod crustacean larvae with special reference to diet. PhD Thesis,
University of Wales, Bangor, UK, 256 pp.
Kumlu, M., Sarihan, E., Tekelioglu, N., 1992. Trypsin activity in larvae of Penaeus monodon Fabricius 1789
(Crustacea, Decapoda, Penaeidae) in relation to their diet. Israeli J. Aquacult. Barnidgeh 44, 103-l IO.
Kumlu, M., Jones, D.A., 1995. Feeding and digestion in the caridean shrimp larvae of Palaemon elegans
Rathke and Macrobrachium rosenbergii De Man (Crustacea: Palaemonidae) on live and artificial diets.
Aquacult. Nutr. 1, 3-12.
Kumlu, M., Jones, D.A. The effect of live and artificial diets on growth, survival and trypsin activity in larvae
of Penaeus indicus H. Milne Edwards (Penaeidae: Crustacea). J. World Aquacult. Sot., in press. Kumlu et al., 1995.
Kurmaly. K., Yule, A.B., Jones. D.A., 1989. An energy budget for the larvae of Penaeus mono&n
(Fabricius). Aquacult. 81, 13-25.
Kurmaly, K., Jones, D.A., Yule, A.B., 1990. Acceptability and digestion of diets fed to larval stages of
Homarus gammarus and the role of dietary conditioning behaviour. Mar. Biol. 106, 18 1- 190.
Laubier-Bonichon, A., Van Wormhoudt, A., Sellos, D., 1977. Croissance larvaire contrlee de Panaeus
japonicus Bate, enzymes digestives et changement de regimes alimentaires. Actes Colloques CNEXO 4,
131-145.
Le Moullac, G., Roy, P., Van Wormhoudt, A., 1992. Effects of trophic prophylactic factors on some digestive
enzymatic activities of Penaeus cannamei larvae. In: Caldtron, J., Sandoval, V. (Eds.), Memorias Primer Congress0 Ecuatoriano de Acuicultura, San Pedro de Manglaralto, Ecuador, CENAIM, pp. 8 l-86.
Le Vay, L., Rodriguez, A., Kamarudin, M.S., Jones, D.A., 1993. Influence of live and artificial diets on tissue
composition and trypsin activity in Penaeus japonicus larvae. Aquaculture 118, 287-297.
Lovett, D.L., Felder, D.L., 1990a. Ontogenetic change in digestive enzyme activity of larval and postlarval
white shrimp Penaeus set$erus (Crustacea Decapoda, Penaeidae). Biol. Bull. 178, 144-159.
Lovett, D.L., Felder, D.L., 199Ob. Ontogenetic changes in enzyme distribution and midgut function in
developmental stages of Penaeus setijerus (Crustacea Decapoda, Penaeidae). Biol. Bull. 178, 160-174. MacDonald, N.L., Stark, J.R., Keith, M., 1989. Digestion and nutrition of the prawn Penaeus monodon. J.
World Aquacult. Sot. 20, 53A.
Munilla-Moran, R., Stark, J.R., Barbour, A., 1990. The role of exogenous enzymes in digestion in cultured turbot larvae Scophthalmus maximus L. Aquaculture 88, 337-350.
Osuna, C., Asuncion, O., Sillero, A., Gunther Sillero, M.A., Sebastian, J., 1977. Induction of multiple
proteases during the early larval development of Artemia salina. Develop. Biol. 61, 94-103. Parsons, T.R., Takahashi, M., Hargrave, B., 1977. Biological Oceanographic Processes, 2nd edn., Pergamon,
Oxford, 332 pp.
Ribeiro, F.A.L.T., Jones, D.A., 1995. Growth and trypsin activity in the Indian white prawn Penaeus indicus
postlarvae. Aquaculture 95, 148, Abstract 275.

D.A. Jones et al./Aquaculture 155 (19971285-295 295
Rick, W., Stegbauer, H.P., 1974. cY-Amylase. In: Bergmeyer, H.U. (Ed.), Methods of Enzymatic Analysis,
Vol. 2. Academic Press, New York, pp. 885, 889.
Rodriguez, A., Le Vay, L., Mourente, G., Jones, D.A., 1994. Biochemical composition and digestive enzyme
activity in larvae and postlarvae of Penaeus japonicus during herbivorous and carnivorous feeding. Mar.
Biol. 118, 45-51.
Samain, J.F., Meal, J., Daniel, J.Y., Le Coz, J.R., Jezequel, M., 1980. The digestive enzymes amylase and
trypsin during the development of Artemia: effect of food conditions. In: Persoone, G., Sorgeloos, P.,
Roels, O., Jasper& E. (Ed.), The Brine Shrimp Artemiu 2, Physiology, Biochemistry, Molecular Biology,
Universa Press, Wetteren, pp. 239-255.
Schmidt, F.H.H., Stork, K., von Dahl, K., 1974. Lipase. In: Bergmeyer, H.U. (Ed.), Methods of Enzymatic
Analysis, Academic Press, New York, pp. 819-823.
Van Wormhoudt, A., 1973. Variations des prottases, des amylases et des protCines soluble au curs du
dCveloppement larvaire chez Paluemon serratus. Mar. Biol. 19, 245-248.