rotifer–crustacean interactions in a pseudokarstic lake: influence of hydrology
TRANSCRIPT

Rotifer–crustacean interactions in a pseudokarstic lake:influence of hydrology
Ulrike Obertegger Æ Andrea Borsato ÆGiovanna Flaim
Received: 14 December 2008 / Accepted: 5 August 2009
� Springer Science+Business Media B.V. 2009
Abstract Zooplankton abundance was related to
hydrological and environmental variables in a hydro-
logically dynamic lake fed by a pseudokarstic aquifer.
The study period (2002–2006) in Lake Tovel covered
different hydrological situations with water residence
time (WRT) having the lowest values in 2002 and the
highest values in 2003. WRT was negatively corre-
lated with silica concentrations and algal biovolume.
Furthermore, the biovolume of small algae was
highest in spring and summer, while large algae did
not show any pattern. In multivariate analysis, high
abundance of crustacean species in autumn and winter
was positively related to WRT and negatively to algal
biovolume, while high abundance of rotifer species in
spring and summer was negatively related to WRT
and positively to algal biovolume. With the exception
of Keratella cochlearis and Gastropus stylifer, rotifers
showed a pattern of crustacean avoidance, and three
groups were distinguished: (i) Ascomorpha ecaudis
and Polyarthra dolichoptera, (ii) Asplanchna prio-
donta and Synchaeta spp., and (iii) Filinia terminalis
and Keratella quadrata. These groups were associated
with different food sources and depths. We suggest
that WRT influenced the rotifer–crustacean relation-
ship by wash-out effects and competition for food
resources. The dynamics of single rotifer species were
attributable to specific feeding requirements and
adaptations. In summary, WRT determined the plat-
form for abiotic and biotic interactions that influenced
population dynamics of crustaceans and rotifers.
Keywords Water residence time �Zooplankton � Algal food � Competition �Depth preference
Introduction
Limnology has moved forward from treating lakes as
isolated basins to studying them as complex ecosys-
tems influenced by their catchments and surroundings.
Exterior factors such as solar irradiance, allochtho-
nous nutrient input, water inflow and anthropogenic
impacts affect the dynamics of organisms living
within an ecosystem (Wetzel 2001). Much is known
about the influence of several internal factors such as
temperature (Gillooly et al. 2002), oxygen (Mikschi
1989), food availability (Pourriot 1977), predation
(Brooks and Dodson 1965) and competition (Declerck
et al. 2003) on zooplankton dynamics. In the last
decade, however, many studies in reservoirs, ponds
and rivers have shown that different aspects of water
U. Obertegger (&) � G. Flaim
IASMA Research and Innovation Centre, Fondazione
Edmund Mach, Environment and Natural Resources Area,
Via E. Mach 2, 38010 San Michele all’Adige, TN, Italy
e-mail: [email protected]
A. Borsato
Museo Tridentino di Scienze Naturali, Via Calepina 14,
38100 Trento, TN, Italy
123
Aquat Ecol
DOI 10.1007/s10452-009-9285-0

movement (i.e. water-level fluctuations, length of wet
phase, water residence time (WRT) and flow rate)
explicitly or implicitly influence zooplankton abun-
dance and composition (e.g. rivers: Basu and Pick
1997; Thorp and Mantovani 2005; reservoirs: Naselli-
Flores and Barone 1997; Campbell et al. 1998; Hart,
2004; Geraldes and Boavida 2007; Mac Donagh et al.
2009; ponds: Girdner and Larson 1995; Brucet et al.
2005). In flood plains and apidly flushed lakes, lotic or
lentic conditions mediate rotifer or crustacean dom-
inance, and zooplankton dynamics are similar to that
in rivers (Walz and Welker 1998; Baranyi et al. 2002).
In shallow lakes, the influence of hydrology on
zooplankton has recently attracted research (de Souza
Cardoso and da Motta Marques 2009; Rennella and
Quiros 2006), while in deep lakes, WRT has seldom
been considered.
Lake (L.) Tovel is a deep (maximum depth 39 m)
lake fed by a pseudokarstic aquifer that surfaces in
several perilacustrine springs in the SW basin (Bor-
sato and Ferretti 2006). Karstic lakes originate by
limestone and/or gypsum dissolution; however, in the
case of L. Tovel, the depositions of a retreating
glacier account for the porous aquifer (Ferretti and
Borsato 2006). The quantification of water input by
karstic aquifers requires particular instrumentation
and long-term data. For this reason, the hydrology of
L. Tovel was subject to detailed studies of its aquifer
and basin (Borsato and Ferretti 2006; Ferretti and
Borsato 2006). Based on its dynamic and well studied
hydrology, this lake can function as a model for
ecosystem–hydrology interactions, evermore relevant
in a changing landscape.
We analysed a 5-year data set (2002–2006) to
address the following questions: (i) what are the
environmental factors that determine species interac-
tions?, and (ii) do all crustacean and rotifer species
exhibit the same relation with WRT and environ-
mental factors?
Materials and methods
Study site
L. Tovel (46�150 N, 10�570 E; area = 38 ha, volume
= 7.4 9 106 m3, maximum depth = 39 m, mean
depth = 19 m) is located 1,178 m above sea level
(a.s.l.) in the Adamello Brenta Natural Park—
Trentino, Italy (Fig. 1). The lake has a large and
deep (39 m) NE basin and a small and shallow (4 m)
SW basin and is frozen from December to April. Its
hydrology is extensively described in Borsato and
Ferretti (2006), Ferretti and Borsato (2006) and
Obertegger et al. (2007). Briefly, the S. Maria
Flavona tributary disappears 1 km above the lake in
the pseudokarstic aquifer (Pozzol) and reappears as
several perilacustrine springs in the SW basin which
account for the main water inflow (Fig. 1). The mean
daily water residence time (WRT) was calculated as
the ratio between the lake volume and the water
inflow based on mean daily data (24 measurements
per day); the temperature of the perilacustrine springs
in the SW basin was measured by thermo-sensors.
Sampling and sample processing
Samples were collected at biweekly (2002) or monthly
(2003, 2004, 2005 and 2006) intervals. However,
during winter the lake was often inaccessible. On each
sampling date, vertical profiles of water temperature
and dissolved oxygen (DO) saturation (Hydrolab
DS4a multiprobe) and Secchi disk readings were
taken. Zooplankton samples (total number for the
whole study period = 560) were taken with a 3-L
Kemmerer-like sampler at discrete depths (surface, 1,
2, 5, 10, 15, 20, 25, 30 and 35 m) in the NE basin of the
lake, filtered through a 10 lm plankton net, and fixed
with formalin (1% v/v final concentration). Phyto-
plankton samples (total number = 448) were taken in
Fig. 1 Part of the watershed area of Lake Tovel. Insert shows
geographical location
Aquat Ecol
123

conjunction to zooplankton samples and were also
collected at discrete depths (surface, 1, 2, 5, 10, 15, 20
and 25 m) in the euphotic zone (diffuse attenuation
coefficient of photosynthetically active radiation:
KdPAR = 0.19 m-1, Obertegger et al. 2008). Subsam-
ples were fixed with acid Lugol’s solution, and algae
were counted according to the Uthermohl technique
(1958). Algal biovolume was estimated from cell
dimensions. Furthermore, because zooplankton ability
to ingest algae may depend on cell size, algal
biovolume was separated by the greatest axial linear
dimension (GALD) into a small (B30 lm) and large
fraction ([30 lm) according to Naselli-Flores and
Barone (1997). For more detailed information see
Obertegger et al. (2007).
Data analysis
The depths 0, 1 and 2 m were not included in data
analyses because these layers are highly influenced
by ultraviolet radiation (Obertegger et al. 2008). We
consequently considered strata deeper than 2 m to
avoid confounding the strong impact of ultraviolet
radiation with other parameters influencing zooplank-
ton dynamics. Therefore, all statistical analyses were
based on the 5–25 m depths.
We used multivariate ordination methods to
investigate zooplankton dynamics dependent on
environmental variables. The decision about what
ordination to use was based on a Detrended Corre-
spondence Analysis according to the length of the
gradient measuring beta diversity of community
composition (Ter Braak and Smilauer 2002). Linear
multivariate techniques (i.e. Principal Components
Analysis—PCA and Redundancy Analysis—RDA)
were applied. The significance of eigenvalues of the
RDA was tested by Monte Carlo permutation tests
(n = 1,000) that were constrained to keep the tem-
poral sequence of data. Zooplankton species abun-
dance was log transformed to give equal importance
to each species. Environmental variables such as
WRT and algal biovolume were also log-trans-
formed, while the variables indicating astronomical
seasons (i.e. spring, summer, autumn and winter) and
depth were coded as dummy binary variables. In the
analysis, species data were centred to unit variance
and zero mean, and scaling was done with the focus
on species correlation. Additionally, the technique of
variance partitioning (Borcard et al. 1992) was used
to quantify the variation explained by explanatory
variables related to the first and second axis. Variance
partition is performed by a partial RDA: the amount
of variability explained by group variables Y (e.g.
variables related to the first axis) was determined
once the variability explained by the group variables
X (e.g. variables related to the second axis) was taken
into account. All statistical analyses were performed
using CANOCO 4.5 software (Ter Braak and Smil-
auer 2002).
Non-parametric correlations (Spearman) and mul-
tiple comparisons were performed in R (R Develop-
ment Core Team 2005) and were used to relate
environmental variables to each other.
Results
Chemical and physical properties of the lake
In the years investigated, L. Tovel showed weak
thermal stratification in summer. The temperature of
the perilacustrine springs was between 5 and 6�C
throughout the year (mean 5.5�C, standard deviation
(SD) 0.1�C) (Fig. 2). The lake’s surface water
Fig. 2 Upper panel: upper line—water residence time for the
study period, lower line—temperature of the perilacustrine
springs. Multi-annual seasonal mean biovolumes of small algae
(lower left panel) and large algae (lower right panel). Box plots
show percentiles (5th circle, 10th, 25th, 50th, 75th, 90th, and
95th circle)
Aquat Ecol
123

temperature was highest in summer 2003 (20.4�C).
Furthermore, water temperature was positively cor-
related with DO saturation and negatively correlated
with small and large algae (Table 1). Generally, DO
was high (DO at 25 m: mean 63%, SD 17%), and
anoxia was never a problem. Total phosphorus (mean
3.7 mg l-1, SD 1.7 mg l-1) was always low, while
inorganic nitrogen (mean 372 mg l-1, SD 42 mg l-1)
was always high.
WRT showed a large variability for the study
period (median = 205 days, minimum = 23, maxi-
mum = 652) with 2002 having the lowest and 2003
the highest values (Fig. 2). Furthermore, WRT
showed the lowest values in spring (mean 127 days,
SD 45 days), and the highest values in winter (mean
414 days, SD 157 days) (multiple comparisons
(m.c.): P \ 0.01) and was positively correlated with
temperature and negatively correlated with small and
large algae and silica (Table 1).
Small algae were positively correlated with silica
(mean 0.72 mg l-1, SD 0.18 mg l-1) (Table 1).
Their biovolume was higher in spring and summer
than in autumn and winter (m.c.: P \ 0.01), and was
generally higher than that of large algae (m.c.:
P \ 0.001) (Fig. 2). In addition, the 5–25 m layer
was characterised by a homogenous distribution of
algal biovolume with no marked difference between
depths (m.c.: P [ 0.05). Secchi disk transparency
(mean 12.0 m, SD 3.6 m) was higher in autumn
compared to other seasons (m.c.: P \ 0.001).
Interaction between the zooplankton community
and environmental parameters
During the study period (2002–2006), rotifer diversity
was high with nine numerically important taxa in the
plankton: Ascomorpha ecaudis Perty, Asplanchna
priodonta Gosse, Filinia terminalis Plate, Gastropus
stylifer Imhof, Keratella cochlearis Gosse, Keratella
quadrata O.F.Muller, Synchaeta gr. stylata-pectinata
sensu Ruttner-Kolisko, Synchaeta gr. tremula-oblon-
ga sensu Ruttner-Kolisko, and Polyarthra dolichop-
tera Idelson. These species however, showed large
variations in temporal abundance (Table 2a). Overall,
rotifers numerically dominated the zooplankton
(median = 97%, minimum = 16%, maximum =
100% of rotifers with respect to total zooplankton
abundance). Cladocerans, especially Daphnia lon-
gispina O.F.Muller and Bosmina longirostris
O.F.Muller, made up most of the remaining zoo-
plankton and also showed large temporal variability
(Table 2b).
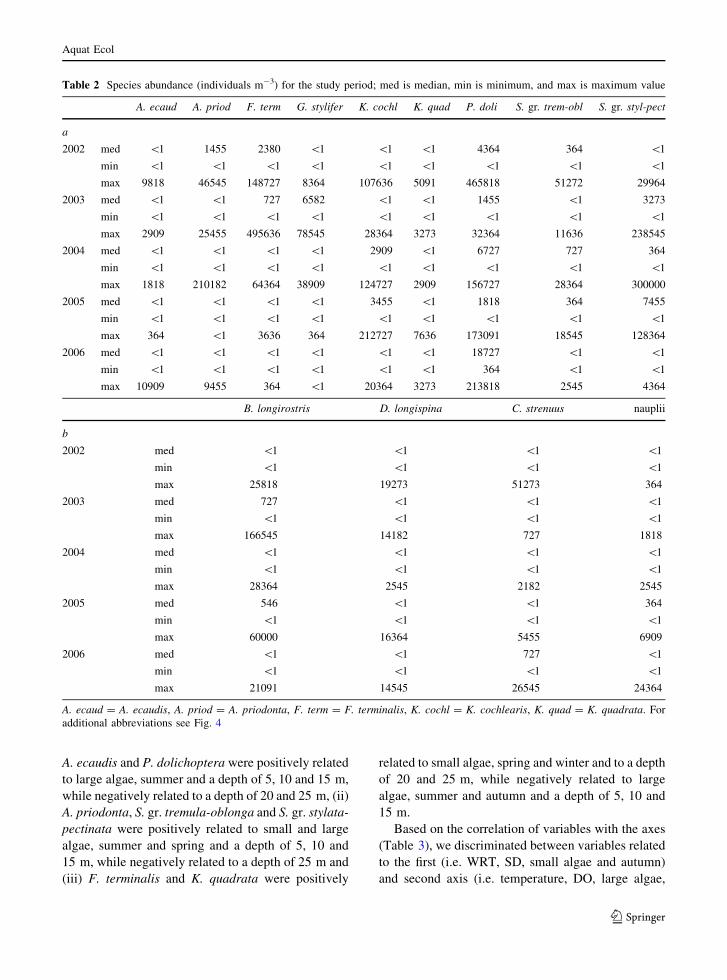
In a PCA based on abundance of zooplankton, the
first two axes accounted for 62.4% of species
variability. We could discriminate among rotifers,
cladocerans and nauplii and copepodites/adults of
Cyclops strenuus Fischer by their different relation to
the first two axes (Fig. 3). On the basis of this result,
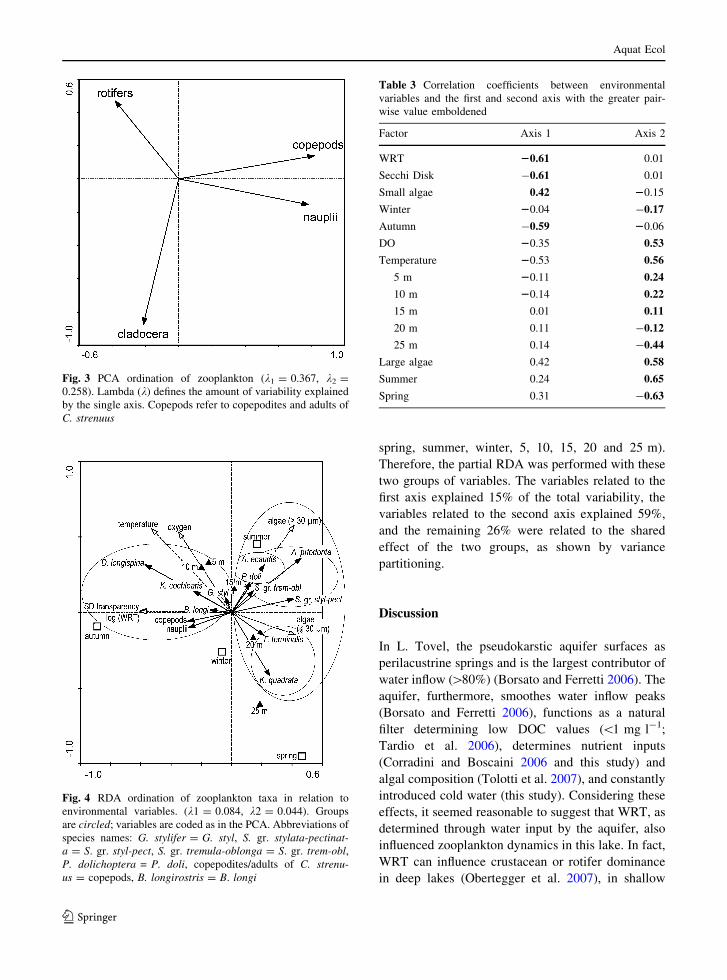
we further investigated species distribution by a RDA.
The RDA explained 22.9% of total species variability,
with the first two axes explaining 60.2% of species–
environment relation. The eigenvalues of the RDA
were statistically significant according to Monte Carlo
permutations (sum of all axes; P \ 0.001). In the
RDA, species showed specific relations with environ-
mental variables leading to a discrimination between
two main groups (Fig. 4). In the first group, copepods,
nauplii, B. longirostris, D. longispina, G. stylifer and
K. cochlearis were positively related to WRT, Secchi
disk transparency, temperature, DO, depth of 5 and
10 m and autumn, while they were negatively related
to small and large algae, depth of 20 and 25 m and
spring. In the second group, Ascomorpha ecaudis,
P. dolichoptera, Asplanchna priodonta, S. gr. tremu-
la-oblonga, S. gr. stylata-pectinata, K. quadrata and
F. terminalis were negatively related to WRT, Secchi
disk transparency, and autumn. Additionally within
the latter group, three subgroups could be distin-
guished that showed this general pattern but had
different trends regarding other parameters: (i)
Table 1 Non-parametric correlation of environmental vari-
ables: the upper section gives significance (*P \ 0.05, **
P \ 0.01, *** P \ 0.001), the lower section gives correlation
(sections separated by the shaded areas); missing values refer
to non-significant correlations; DO is dissolved oxygen, temp is
temperature, B30 lm refers to small algae, [30 lm refers to
large algae, and SiO2 is silica
DO temp ≤ 30 µm
> 30 µm
SiO2 WRT
DO *** ***
temp 0.65 *** ** *** * ≤ 30 µm -0.17 *** *** ***
> 30 µm -0.24 0.19 * SiO2 -0.58 -0.49 0.25 ** WRT 0.32 -0.50 -0.29 -0.45
Aquat Ecol
123
A. ecaudis and P. dolichoptera were positively related
to large algae, summer and a depth of 5, 10 and 15 m,
while negatively related to a depth of 20 and 25 m, (ii)
A. priodonta, S. gr. tremula-oblonga and S. gr. stylata-
pectinata were positively related to small and large
algae, summer and spring and a depth of 5, 10 and
15 m, while negatively related to a depth of 25 m and
(iii) F. terminalis and K. quadrata were positively
related to small algae, spring and winter and to a depth
of 20 and 25 m, while negatively related to large
algae, summer and autumn and a depth of 5, 10 and
15 m.
Based on the correlation of variables with the axes
(Table 3), we discriminated between variables related
to the first (i.e. WRT, SD, small algae and autumn)
and second axis (i.e. temperature, DO, large algae,
Table 2 Species abundance (individuals m-3) for the study period; med is median, min is minimum, and max is maximum value
A. ecaud A. priod F. term G. stylifer K. cochl K. quad P. doli S. gr. trem-obl S. gr. styl-pect
a
2002 med \1 1455 2380 \1 \1 \1 4364 364 \1
min \1 \1 \1 \1 \1 \1 \1 \1 \1
max 9818 46545 148727 8364 107636 5091 465818 51272 29964
2003 med \1 \1 727 6582 \1 \1 1455 \1 3273
min \1 \1 \1 \1 \1 \1 \1 \1 \1
max 2909 25455 495636 78545 28364 3273 32364 11636 238545
2004 med \1 \1 \1 \1 2909 \1 6727 727 364
min \1 \1 \1 \1 \1 \1 \1 \1 \1
max 1818 210182 64364 38909 124727 2909 156727 28364 300000
2005 med \1 \1 \1 \1 3455 \1 1818 364 7455
min \1 \1 \1 \1 \1 \1 \1 \1 \1
max 364 \1 3636 364 212727 7636 173091 18545 128364
2006 med \1 \1 \1 \1 \1 \1 18727 \1 \1
min \1 \1 \1 \1 \1 \1 364 \1 \1
max 10909 9455 364 \1 20364 3273 213818 2545 4364
B. longirostris D. longispina C. strenuus nauplii
b
2002 med \1 \1 \1 \1
min \1 \1 \1 \1
max 25818 19273 51273 364
2003 med 727 \1 \1 \1
min \1 \1 \1 \1
max 166545 14182 727 1818
2004 med \1 \1 \1 \1
min \1 \1 \1 \1
max 28364 2545 2182 2545
2005 med 546 \1 \1 364
min \1 \1 \1 \1
max 60000 16364 5455 6909
2006 med \1 \1 727 \1
min \1 \1 \1 \1
max 21091 14545 26545 24364
A. ecaud = A. ecaudis, A. priod = A. priodonta, F. term = F. terminalis, K. cochl = K. cochlearis, K. quad = K. quadrata. For
additional abbreviations see Fig. 4
Aquat Ecol
123
spring, summer, winter, 5, 10, 15, 20 and 25 m).
Therefore, the partial RDA was performed with these
two groups of variables. The variables related to the
first axis explained 15% of the total variability, the
variables related to the second axis explained 59%,
and the remaining 26% were related to the shared
effect of the two groups, as shown by variance
partitioning.
Discussion
In L. Tovel, the pseudokarstic aquifer surfaces as
perilacustrine springs and is the largest contributor of
water inflow ([80%) (Borsato and Ferretti 2006). The
aquifer, furthermore, smoothes water inflow peaks
(Borsato and Ferretti 2006), functions as a natural
filter determining low DOC values (\1 mg l-1;
Tardio et al. 2006), determines nutrient inputs
(Corradini and Boscaini 2006 and this study) and
algal composition (Tolotti et al. 2007), and constantly
introduced cold water (this study). Considering these
effects, it seemed reasonable to suggest that WRT, as
determined through water input by the aquifer, also
influenced zooplankton dynamics in this lake. In fact,
WRT can influence crustacean or rotifer dominance
in deep lakes (Obertegger et al. 2007), in shallow
Fig. 3 PCA ordination of zooplankton (k1 = 0.367, k2 =
0.258). Lambda (k) defines the amount of variability explained
by the single axis. Copepods refer to copepodites and adults of
C. strenuus
Fig. 4 RDA ordination of zooplankton taxa in relation to
environmental variables. (k1 = 0.084, k2 = 0.044). Groups
are circled; variables are coded as in the PCA. Abbreviations of
species names: G. stylifer = G. styl, S. gr. stylata-pectinat-a = S. gr. styl-pect, S. gr. tremula-oblonga = S. gr. trem-obl,P. dolichoptera = P. doli, copepodites/adults of C. strenu-us = copepods, B. longirostris = B. longi
Table 3 Correlation coefficients between environmental
variables and the first and second axis with the greater pair-
wise value emboldened
Factor Axis 1 Axis 2
WRT 20.61 0.01
Secchi Disk -0.61 0.01
Small algae 0.42 20.15
Winter 20.04 -0.17
Autumn -0.59 20.06
DO 20.35 0.53
Temperature 20.53 0.56
5 m 20.11 0.24
10 m 20.14 0.22
15 m 0.01 0.11
20 m 0.11 -0.12
25 m 0.14 -0.44
Large algae 0.42 0.58
Summer 0.24 0.65
Spring 0.31 -0.63
Aquat Ecol
123

lakes (Rennella and Quiros 2006), and in reservoirs
(Geraldes and Boavida 2007).
Our analysis indicated that rotifers, cladocerans
and copepods tended to exclude each other. In fact, in
temperate lakes zooplankton usually show a temporal
succession with rotifers followed by crustaceans
(Sommer et al. 1986). In addition to temporal
distribution, spatial distribution also has to be con-
sidered. Zooplankton position themselves in the
water column to maximise growth but several
parameters result in trade-offs between survival and
reproduction. We found that rotifers were separated
from crustaceans based on two gradients: the first
gradient corresponded to WRT, Secchi disk transpar-
ency and small algae and was related to autumn,
while the second gradient corresponded to depth
distribution, temperature, DO and large algae and was
related to spring, summer and winter.
The relation of zooplankton with WRT can be
explained by wash-out effects (e.g. Campbell et al.
1998; Walz and Welker 1998; Baranyi et al. 2002;
Rennella and Quiros 2006). Rotifers’ shorter gener-
ation time gives them a decisive advantage over
crustaceans by permitting a faster recovery from
wash-out (Baranyi et al. 2002). Furthermore, juvenile
stages have reduced swimming capacities as shown
for copepods (Maar et al. 2003) and for Daphnia pulex
(Dodson and Ramcharan 1991). In our analysis, all
life stages of copepods were positively related to
WRT, and it seemed reasonable to suggest that the
effect of WRT on adults was indirectly mediated by
the lack of juveniles, more susceptible to wash-out
effects than adults. Furthermore, highest abundance of
crustaceans occurred in autumn and winter in contrast
to spring and summer. These seasons had sufficiently
long periods with high WRT (WRT [ 193 for
23 days) to favour crustacean abundance as indicated
by Obertegger et al. (2007). However, even if
cladocerans and copepods were present in other
seasons, they had their highest abundance in autumn
and winter, confirming the positive relationship
between WRT and crustacean abundance.
Crustaceans’ dominance in autumn and winter also
coincided with a decrease of algal food. This decrease
could be related to different factors such as zoo-
plankton grazing or nutrient decline. Grazing by
rotifers can result in a reduction of algae (Herzig
1987; Gosselain et al. 1998). However, Tolotti et al.
(2007) note the low influence of zooplankton grazing
on diatom abundance in L. Tovel. Furthermore,
crustaceans and rotifers can affect algal abundance
and composition not only directly through grazing but
also indirectly through nutrient regeneration (Carillo
et al. 1990; Urabe 1993). We suggest that in autumn
factors other than grazing were responsible for algal
decline while nutrient regeneration might be impor-
tant in spring and summer when rotifers and algal
food were abundant.
The concentration of nutrients is another important
factor for algal composition and abundance. In
L. Tovel, the negative correlation of WRT with
algae and silica indicated that increasing WRT might
result in a decline of silica and consequently in a
decline of diatoms because of reduced silica input
and concomitant algal consumption.
In periods of high WRT, crustaceans might
compensate the reduced food sources by their higher
filtration capacity compared to rotifers (Herzig 1987).
However, even if rotifers also could be favoured by
high WRT, only few species were associated with
crustaceans. Crustacean-rotifer interactions are deter-
mined by predation (Williamson 1993), exploitative
competition for food of similar size, and mechanical
interference (Burns and Gilbert 1986). These inter-
actions often result in a marked seasonality with
suppression of rotifers by crustaceans. Based on our
analysis, the rotifers G. stylifer and K. cochlearis
were surprisingly associated with crustaceans, and we
suggest that only specific autecological traits might
faciliate this coexistence. While illoricate G. stylifer
might not possess any evident morphological
defence, a lorica with spines might plausibly reduce
predation on K. cochlearis. In fact, copepods such as
Mesocyclops edax have to break the lorica that
hinders ingestion (Williamson 1993). Even if mor-
phological defenses are a necessary condition to
maintain high rotifer abundance in the presence of
abundant mesozooplankton, these characteristics are
not sufficient (Yoshida et al. 2003). Apart from their
resistance to predation, the compensatory effects of
reproduction should also be considered. The short
development time of rotifers with respect to crusta-
ceans is of general importance in maintaining a stable
population (Walz 1995). This characteristic might
allow for fast recovery from predation losses even if
dependent on temperature and food supply. In
addition, competition for food under reduced
resources might be generally important. Keratella
Aquat Ecol
123

cochlearis is adapted to low food concentrations
(Walz 1995), while G. stylifer preferentially feeds on
dinoflagellates (Nogrady et al. 1993) that are abun-
dant in L. Tovel (Flaim et al. 2003). Among
dinoflagellates Gymnodinium uberrimum Kofoid and
Swezy was the most abundant species with which
G. stylifer showed a positive correlation (r = 0.22,
P \ 0.01). Additionally, population dynamics of this
species are positively related to water temperature
(Herzig 1987; Spoljar et al. 2005). In fact, G. stylifer
had its highest abundance in the hot summer and
autumn of 2003. In summary, K. cochlearis and
G. stylifer might compensate scarce food sources and
population losses caused through crustacean preda-
tion by (i) low resource competition, (ii) specific
feeding strategies, (iii) short generation time and (iv)
having the advantage of being the only rotifer species
exploiting resources in coexistence with crustaceans.
Apart from food, temperature and oxygen are other
important parameters for the spatial and temporal
variation in rotifer abundance (Mikschi 1989). Tem-
perature per se determines development rates (Gillo-
oly et al. 2002), and DO saturation is crucial for
direct uptake and respiration rates in rotifers. How-
ever, Armengol et al. (1998) relate the importance of
temperature and oxygen to other associated factors
such as food availability and competition. In L. Tovel,
most species seemed to prefer the 5–15 m layer that
was characterised by high DO values and abundant
algal food; deeper layers were inhabited by species
mainly feeding on bacteria. We suggest that oxygen
supply was not critical and species were adapted to
the low water temperatures at depths lower than 5 m
(summer mean at 5 m 10�C, SD 2.9�C). However in
L. Tovel, some species migrate to the surface layer at
night to take advantage of warmer temperatures
(Obertegger et al. 2008).
Within the main rotifer group, three subgroups
could be distinguished. In the first group, A. ecaudis
feeds on large food cells and dinoflagellates, and
P. dolichoptera feeds on diatoms, crysomonads and
cryptomonads (Pourriot 1977). In our study, both
species were closely related to large algae; we
suggest that these rotifers could exploit food sources
inaccessible to others, and therefore probably had a
competitive advantage. In the second group, Syncha-
eta species feed on small and on large food particles
and Asplanchna is omniphagous (Pourriot 1977).
These species were among the largest rotifers in
L. Tovel, indicating a high food demand according
to the size-efficiency theory of Stemberger and
Gilbert (1987). In the third group, K. quadrata and
F. terminalis are adapted to cold temperatures and
feed on small algae, flagellates, bacteria and detritus
(Nogrady et al. 1993). These characteristics may have
favoured their occurrence in the deeper layers and
simultaneously offered the possibility to avoid com-
petition with crustaceans and other rotifer species.
In summary, we suggest that rotifers generally
responded in the same way to WRT and competition
with crustaceans, and the differentiation among
rotifers was mainly related to specific food require-
ments and adaptations.
While the influence of WRT on plankton dynamics
in shallow lakes is known (de Souza Cardoso and da
Motta Marques 2009; Rennella and Quiros 2006), the
effects of hydrology in deeper lakes can remain
hidden when adequate hydrological parameters are
not used (Spoljar et al. 2005). By using the appro-
priate hydrological variable (i.e. WRT), our study
underlined the importance of hydrology for rotifer
species dynamics in a deep lake. We suggest that
WRT directly determined wash-out effects of zoo-
plankton and patterns of algal food, and consequently
also indirectly determined zooplankton dynamics.
Therefore, we hypothesised that WRT was the
starting point for abiotic and biotic factors. This
coupling of abiotic and biotic processes is in line with
the ‘‘multiple driving force hypothesis’’ of Pinel-
Alloul (1995). In fact, environmental variables shared
effects on species distribution, and therefore interre-
lation among parameters could not be neglected. In
the present study, specific species related to WRT in
different ways, and experimental studies are required
to quantify the influence of WRT on single species. In
addition, the unexplained variability of species
abundance could be determined by other factors such
as fish predation (Brooks and Dodson 1965), parasit-
ism (Nogrady et al. 1993), crowding (Stelzer and
Snell 2006), or intra-species competition (Declerck
et al. 2003) that also influence zooplankton dynamics.
In conclusion, the pseudokarstic catchment mark-
edly influenced the plankton of its associated lentic
ecosystem, and the dynamics of crustaceans and
rotifers were based on WRT and competition for
algal food.
Aquat Ecol
123

Acknowledgements This study was partially supported by
ECOPLAN and CERCA Research Grants (Province of Trento,
Italy), and by research activity funded by FEM-IASMA. We
thank Flavio Corradini for providing chemical data, Gino
Leonardi and Vigilio Pinamonti for help with sampling, and
Damaso Calliari and Federica Fiammingo for phytoplankton
counting.
References
Armengol X, Esparcia A, Miracle RM (1998) Rotifer vertical
distribution in a strongly stratified lake: a multivariate
analysis. Hydrobiologia 387/388:161–170
Baranyi C, Hein T, Holarek C, Keckeis S, Schiemer F (2002)
Zooplankton biomass and community structure in a
Danube River floodplain system: effects of hydrology.
Freshw Biol 47:473–482
Basu BK, Pick FR (1997) Phytoplankton and zooplankton
development in a lowland, temperate river. J Plankton Res
19:237–253
Borcard D, Legendre P, Drapeau P (1992) Partialling out the
spatial component of ecological variation. Ecology
73:1045–1055
Borsato A, Ferretti P (2006) Hydrological monitoring of Lake
Tovel and its catchment [in Italian]. Studi Trent Sci Nat,
Acta Biol 81:205–223
Brooks JL, Dodson SI (1965) Predation, body size, and com-
position of plankton. Science 150:28–35
Brucet S, Boix D, Lopez-Flores R, Badosa A, Moreno-Amich
R, Quintana X (2005) Zooplankton structure and
dynamics in permanent and temporary Mediterranean salt
marshes: taxon-based and size-based approaches. Arch
Hydrobiol 162:535–555
Burns CW, Gilbert JJ (1986) Effects of daphnid size and
density on interference between Daphnia and Keratellacochlearis. Limnol Oceanogr 31:848–858
Campbell CE, Knoechel R, Copeman D (1998) Evaluation of
factors related to increased zooplankton biomass and
altered species composition following impoundment of a
Newfoundland reservoir. Can J Fish Aquat Sci 55:
230–238
Carillo P, Criz-Pizarro L, Sanchez-Castillo P (1990) Analysis
of phytoplankton-zooplankton relationships in an oligo-
trophic lake under natural and manipulated conditions.
Hydrobiologia 200/201:49–58
Corradini F, Boscaini A (2006) Physical and chemical char-
acteristics of Lake Tovel (central Alps) [in Italian]. Studi
Trent Sci Nat, Acta Biol 81:307–326
Declerck S, Geenens V, Podoor N, Conde-Porcuna JM,
DeMeester L (2003) Intraspecific density dependence in
the dynamics of zooplankton under hypertrophic condi-
tions. Can J Fish Aquat Sci 60:919–928
Dodson S, Ramcharan C (1991) Size-specific swimming
behavior of Daphnia pulex. J Plankton Res 13:1367–1379
de Souza Cardoso L, da Motta Marques D (2009) Hydrody-
namics-driven plankton community in a shallow lake.
Aquat Ecol 43:73–84
Flaim G, Rott E, Corradini F, Toller G, Borghi B (2003) Long-
term trends in species composition and diurnal migration
of dinoflagellates in Lake Tovel (Trentino, Italy). Hyd-
robiologia 502:357–366
Ferretti P, Borsato A (2006) Hydrogeological study of Lake
Tovel and its valley [in Italian]. Studi Trent Sci Nat, Acta
Biol 81:189–204
Geraldes AM, Boavida MJ (2007) Zooplankton assemblages in
two reservoirs: one subjected to accentuated water level
fluctuations, the other with more stable water levels.
Aquat Ecol 41:273–284
Gillooly JE, Charnov EL, West GB, Savage VM, Brown JH
(2002) Effects of size and temperature on development
time. Nature 417:70–73
Girdner SF, Larson GL (1995) Effects of hydrology on zoo-
plankton communities in high-mountain ponds, Mount
Rainer National Park, USA. J Plankton Res 17:1731–1755
Gosselain V, Viroux L, Descy JP (1998) Can a community of
small-bodied grazers control phytoplankton in rivers?
Freshw Biol 39:9–24
Hart RC (2004) Cladoceran periodicity patterns in relation to
selected environmental factors in two cascading warm-
water reservoirs over a decade. Hydrobiologia 526:
99–117
Herzig A (1987) The analysis of planktonic rotifer populations:
a plea for long-term investigations. Hydrobiologia
147:163–180
Maar M, Nielsen TG, Stips A, Visser AW (2003) Microscale
distribution of zooplankton in relation to turbulent diffu-
sion. Limnol Oceanogr 48:1312–1325
Mac Donagh ME, Casco MA, Claps MC (2009) Plankton
relationships under small water level fluctuations in a
subtropical reservoir. Aquat Ecol 43:371–381
Mikschi E (1989) Rotifer distribution in relation to temperature
and oxygen content. Hydrobiologia 186(187):209–214
Naselli-Flores L, Barone R (1997) Importance of water-level
fluctuations on population dynamics of cladocerans in a
hypertrophic reservoir (Lake Arancio, south-west Sicily,
Italy). Hydrobiologia 360:223–232
Nogrady TR, Wallace L, Snell TW (1993) Rotifera vol 1:
Biology, ecology and systematics. In: Dumont HJF (ed)
Guides to the identification of the microinvertebrates of
the continental waters of the world 4. SPB Academic
Publishing, The Hague
Obertegger U, Flaim G, Braioni MG, Sommaruga R, Corradini
F, Borsato A (2007) Water residence time as a driving
force of zooplankton structure and succession. Aquat Sci
69:575–583
Obertegger U, Flaim G, Sommaruga R (2008) Multifactorial
nature of rotifer water layer preferences in an oligotrophic
lake. J Plankton Res 30:633–643
Pinel-Alloul B (1995) Spatial heterogeneity as a multiscale
characteristics of zooplankton community. Hydrobiologia
300/301:17–42
Pourriot R (1977) Food and feeding habits of rotifers. Arch
Hydrobiol 8:243–260
R Development Core Team (2005) R: a language and environ-
ment for statistical computing. R Foundation for Statistical
Computing, Vienna (available at: http://www.r-project.
org/)
Rennella AM, Quiros R (2006) The effects of hydrology on
plankton biomass in shallow lakes of the Pampa Plain.
Hydrobiologia 556:181–191
Aquat Ecol
123

Sommer U, Gliwicz ZM, Lampert W, Duncan A (1986) The
PEG-model of seasonal succession of planktonic events in
freshwaters. Arch Hydrobiol 106:433–471
Spoljar M, Habdija I, Primc-Habdija B, Sipos L (2005) Impact
of environmental variables and food availability on rotifer
assemblage in the karstic Barrage Lake Visovac (Krka
River Croatia). Internat Rev Hydrobiol 90:555–579
Stelzer CP, Snell TW (2006) Specificity of the crowding
response in the Brachionus plicatilis species complex.
Limnol Oceanogr 51:125–130
Stemberger RS, Gilbert JJ (1987) Rotifer threshold food con-
centrations and the size-efficiency hypothesis. Ecology
68:181–187
Tardio M, Spitale D, Ghetti F, Bagnoli C, Nannicini L, Seritti
A, Corradini F, Cantonati M (2006) Selected results from
enclosure experiments obtained in Lake Tovel during the
SALTO-WP6 Project. Studi Trent Sci Nat, Acta Biol
81:397–411
Ter Braak CJF, Smilauer P (2002) CANOCO reference manual
and CanoDraw for Windows user’s guide: Software for
Canonical Community Ordination (version 4.5). Micro-
computer Power (Ithaca, NY, USA).
Thorp J, Mantovani S (2005) Zooplankton of turbid and
hydrologically dynamic prairie rivers. Freshw Biol 50:
1474–1491
Tolotti M, Corradini F, Boscaini A, Calliari D (2007) Weather-
driven ecology of planktonic diatoms in Lake Tovel
(Trentino, Italy). Hydrobiologia 578:147–156
Urabe J (1993) N and P cycling coupled by grazers’activity:
food quality and nutrient release by zooplankton. Ecology
4:2337–2350
Uthermohl H (1958) Zur Vervollkommung der quantitativen
Phytoplankton-Methodik. Verh Int Ver Limnol 9:1–38
Walz N (1995) Rotifer population in plankton communities:
energetics and life history strategies. Experientia 51:
437–453
Walz N, Welker M (1998) Plankton development in a rapidly
flushed lake in the River Spree system (Neuendorfer See,
Northeast Germany). J Plankton Res 11:2071–2087
Wetzel R (2001) Limnology: lake and river ecosystems. Aca-
demic Press, San Diego
Williamson CE (1993) Linking predation risk models with
behavioral mechanisms: identifying population bottle-
necks. Ecology 74:320–331
Yoshida T, Urabe J, Elser JJ (2003) Assessment of ‘top-down’
and ‘bottom-up’ forces as determinants of rotifer distri-
bution among lakes in Ontario, Canada. Ecol Res 18:
639–650
Aquat Ecol
123