evolution of several cytoskeletal proteins of astrocytes in primary culture: effect of prenatal...
TRANSCRIPT

Neurochemical Research, VoL 16, No. 7, 1991, pp. 737-747
Evolution of Several Cytoskeletal Proteins of Astrocytes in Primary Culture: Effect of Prenatal Alcohol Exposure*
Rosana Sfiez 1, Maria Burgal 1, Jaime Renau-Piqueras z, Amparo Marqu6s 1, and Consuelo Guerri 1,3
(Accepted November 8, 1990)
In the present work we have analyzed, using immunoblotting and immunofluorescence techniques, the evolution of several cytoskeletal proteins during the development of astrocytes in primary culture. The effect of prenatal exposure to alcohol on these proteins was also evaluated. Micro- tubular protein c~-tubulin decreased approximately 47% from 4 to 7 days after which its content remained practically constant. Immunofluorescence studies showed also that the content of e~-tubulin was greater at day 4 of culture. This increase in fluorescence was coincident with the presence of globular particles which were found in interphase astrocytes and stained with both anti c~- and antf-[3-tubulin. These structures appeared only in proliferating cells. Glial fibrillary acidic protein (GFAP) and vimentin were analyzed as intermediate filament (IF) proteins. GFAP, in cytoskeletal preparations, increased regularly for 14 days followed by a decrease to day 21. In contrast, vimentin showed a progressive increase throughout the entire culture period. Fluorescence studies revealed some differences between the IF distribution patterns of GFAP and vimentin.
In astrocytes obtained from rats prenatally exposed to ethanol, decreases in the amounts of all the cytoskeletal proteins studied were found during the entire culture period. In these cells a striking disorganization of cytoskeleton was also observed. The alcohol-induced decrease of GFAP in cultured astrocytes was also found when this protein was studied in preparations from whole brain developed "in vivo".
KEY WORDS: Primary astrocytes; ethanol; glial fibrillary acidic protein; vimentin; tubulin.
INTRODUCTION
Prenatal exposure to ethanol during development induces a range of adverse effects in the offspring, the extreme of which is the fetal alcohol syndrome (FAS) (1). One of the most important features of this syndrome
1 Instituto Investigaciones Citol6gicas de la Caja de Ahorros de Val- encia (Centro Asociado del CSIC) Amadeo de Saboya 4, 46010- Valencia, Spain;
2 Centro de Investigaci6n Hospital "La Fe" 46009-Valencia, Spain. 3 To whom to address reprint requests: Consuelo Guerri, Instituto de
Investigaciones, Citol6gicas, Amadeo de Saboya, 4, 46010-Valen- cia, Spain. Tel: (6)3698500 FAX: (6)3601453.
* Special issue dedicated to Dr. Santiago Grisolia
737
is the apparent damage to the central nervous system (CNS) which is reflected in psychomotor and mental disabilities (1, 2) and, in fact, FAS is recognized as the leading known cause of mental retardation (3). Although neuronal alterations have been reported in both experi- mental animals (4) and in necropsies of children affected by FAS (5), the teratogenic mechanisms of ethanol on CNS development remain obscure. Since neuronal func- tion depends on the functional integrity of neuroglial cells (6) and because we have previously demonstrated that prenatal exposure to ethanol affects several mor- phological and functional characteristics of astrocytes (7, 8), we have postulated that ethanol-induced neuronal al- terations during development could be due to an initial
0364-3190/91/0700-0737506.50/0 �9 I991 Plenum Publishing Corporation

738 S~ez, B u r g a l , R e n a u - P i q u e r a s , M a r q u 6 s , and Guerri
damage by this toxic agent on proliferating and differ- entiating astrocytes (7, 9).
Astrocyte cytoskeleton, one of the most character- istic features of these cells, appears to perform important structural functions including the formation of a frame- work for migration of neurons during development. In- termediate filaments (IF), a component of cytoskeleton, give astrocytes stability (10) and in metabolic functions provide a support for attachment and transport of organ- elles (11). Astrocytes express several IF proteins includ- ing vimentin and glial fibrillary acidic protein (GFAP) (12). In addition, the structural function of IF appears to depend on the microtubules (MT) which are required for the formation and maintenance of processes in steI- late astrocytes (13, 14). However, no studies have been reported heretofore on the content and evolution of MT proteins in astrocytes in primary culture.
The GFAP is the major component in glial IF (15, 16) and has been used as a specific marker for glial cells, particularly in development of astrocytes both in vivo and in primary culture (10, 17, 18). In previous studies we have demonstrated that a reorganization of the anti- GFAP binding sites from clustered to random occurs during astrocyte differentiation (7). We also reported that prenatal exposure to ethanol de layed morpho log ica l as- t rocyte matura t ion , decreased the content of G F A P and al tered this reorganiza t ion of IF . Howeve r , whether pre- natal exposure to a lcohol (PEA) could alter other cyto- skelatal prote ins in as t rocytes in p r imary culture and whe ther the decrease observed in G F A P dur ing glial matura t ion " i n v i t r o " is also ref lected " i n v i v o " , re- mained to be clar i f ied.
The present s tudy has been under taken to analyze the content and dis t r ibut ion pat tern of the cytoskele ta l proteins a - tubu l in , G F A P and v iment in in as t rocytes during their deve lopment in p r imary cul ture , as wel l as to evaluate the effect of prenata l exposure to ethanol on these cytoskeletal proteins. In addition, the levels of G F A P and the effect of ethanol exposure dur ing brain devel- opment " i n v i v o " were also inves t igated.
E X P E R I M E N T A L P R O C E D U R E
Chemicals. Chemicals used were obtained from the following: Dulbecco's modified Eagle's medium from Gibco Lab. (Grand Island, N.Y.); fetal calf serum from Flow Lab. (UK).
Immunochemicals were obtained from: Anti-glial fibrillary acidic protein (GFAP, monoclonal), anti-vimentin (monoclonal, clone V-9), anti-neurofilament 68-KD (monoclonal), and anti-mouse IgG-FITC (fluorescein isothiocyanate) conjugate, were from Boehringer Chem- ical Co. (Germany); anti-~x tubulin (monoclonal, clone DM 1A), anti- tubulin (polyclonal), anti-GFAP (polyclonal), anti-rabbit IgG-TRITC
(tetramethylrhodamine isothiocyanate) conjugate, and anti-rabbit IgG- FITC conjugate; anti-mouse IgG-TRITC conjugate, and anti-mouse IgG peroxidase conjugate were from Sigma Chemical Co. (St. Louis, USA).
Animal Treatment. Adult virgin female Wistar rats with an initial body weight of 200-250 g were used. Females were fed before and during gestation with an ethanol or isocaloric control liquid diet (19).
After 4-5 weeks on the control or alcohol treatments, females in proestrus or initial estrus were placed overnight in cages with males of the same strain. Pregnant rats were housed in individual cages with their corresponding diet available, and a record was kept during ges- tation of the daily food and/or fluid intake and body weight.
We have previously reported the daily caloric intake of protein and ethanol throughout gestation by the dams (20). With this liquid alcoholic diet the blood ethanol levels in pregnant rats were approxi- mately 20-30 mmol/1. We have also reported that the rate of neonatal mortality of pups from alcohol-fed mothers is = 15% and that offspring from alcoholic rats were smaller than the corresponding pair-fed con- trol rats (20, 21, 22).
Pregnant control and alcoholic rats were routinely checked for births on day 21 of gestation. The parturition day was considered day 0 of the postnatal period (p.p.). After birth, the litters were culled to 8-10 pups per dam and maintained with their physiological mothers for the next 28 days. At 4, 7, 14, 21, and 28 days p.p., pups were killed by decapitation and the brains or cerebral cortex were imme- diately removed, frozen in liquid nitrogen, weighed and then stored at -70~ The same procedure was used for adult rat brain (65 days old).
Astrocyte Cultures. Primary cultures of astrocytes from 21-day- old fetuses were prepared from brain hemispheres as described pre- viously (7). Fetuses were obtained under sterile conditions from both control and chronic alcoholic rats. Ceils were plated on 35-ram Nunc tissue culture plastic dishes (1 x 10" cells/dish, 2 ml per dish). The medium was changed every 3 days. Possible contamination by neurons was assessed by immunoblotting and immunofluorescence using an anti-neurofilament 68-K.D.
Preparation of Astrocyte Cytoskeleton and Rat Brain Homoge- nates. Cytoskeletons from primary astrocytes were prepared according to the procedure of Chiu et al. (12), at several intervals after plating (4, 7, 14 and 21 days). The cytoskeleton pellet was dissolved in 0.25 ml of SDS-sample buffer or ES buffer (0.125M Tris-HC1 pH=6.8, 2% SDS, 2 mM EDTA, 2 mM PMSF, 5% (w/v) 2-mercaptoethanol [MSH], 10% glycerol and 0,025% bromophenol blue). Before adding MSH and bromophenol blue, aliquots were withdrawn to determine protein (23). Samples were then boiled for 5 min and stored at - 70~
When total cell proteins were assessed for a-tubulin immunode- tection, culture plates were rinsed two times with phosphate buffered saline (PBS). Monolayers were resuspended in 0.5 ml of ES buffer. Samples were boiled for 5 rain and stored at -70~
When brain homogenates were used, frozen control or alcohol- exposed brains (or cerebral cortex) were homogenized in 4 parts (w/v) of 0.125M Tris-HC1 pH=7.8 (containing 2% SDS, 2 mM PMSF, 2 mM EDTA) in a Potter-Elvehjem homogenizer at 4~ A portion of the homogenate (0.125 ml) was boiled in 1 ml of EB buffer for 5 minutes. The duration of the whole procedure was ~7 rain. Boiled samples were used in the same day for SDS-polyacrylamide gel elec- trophoresis and immunoblotting for GFAP. Protein concentration in tissue homogenates was estimated by the method of Lowry et al. (23) using bovine serum albumin as standard.
Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS- PAGE) and immunoblotting. After thawing, samples from the different culture periods were simultaneously heated to 100~ for 1 min and

Ethanol Alters Cytoskeleton of Astrocytes 739
proteins were separated in SDS-polyacrylamide slab gels following the discontinuous gel and buffer system of Laemmli (24) and employing 10% (w/v) polyacrylamide in the separation gel. Two sets of molecular weight standards (Sigma kits) were run per gel. Analytical gels were stained with Coomasie blue, destained and scanned in a LKB Ultrascan densitometer. Under these conditions no proteolysis was found. For immunoblotting, after electrophoresis proteins were transferred to ni- trocellulose paper (25) using a Semidray Electroblotter (Millipore) ap- paratus. The nitrocellulose paper was incubated for 2h at 25~ with monoclonal antibodies to a-tubulin (1:1000 dilution), GFAP (1:500 dilution) or vimentin (1:500 dilution) and then incubated for lh with anti-mouse IgG conjugate with peroxidase. After 5-10 min of color development, the nitrocellulose sheets were washed and photograph- ically reproduced on acetate films. The intensity of the bands was measured using a laser densitometer (Ultrascan, LKB).
Fluorescence Microscopy. Astrocyte monolayers growing on 16- ram glass coverslips were fixed, permeabilized and extracted for im- munofluorescence according to the procedure described by Bell et al.~ (26) with minor modifications. Astrocyte cytoskeletons were incubated simultaneously with anti-GFAP (i:10 dilution) and anti-vimentin (1:10 dilution) anti-a (1:100 dilution) or anti-13 (1:10 dilution) tubulin in- stead of the anti-vimentin. The incubation was carried out at 37~ during 60 rain. After washing several times in BSA-Tris-buffered sa- line, cells were incubated for 90 rain at 37~ with PBS containing: anti-mouse-IgG-TRITC and anti-rabbit-IgG-FITC for vimentin and GFAP respectively; and anti-mouse-IgG FITC and anti-rabbit-IgG-TRITC for tubulin and GFAP respectively. All the dilutions were 1/10.
Astrocytes with double labeling were examined in a Zeiss fluo- rescence microscope with vertical illuminator for incident light (Ex 450-490/FT 510/LP 520 for FITC and Ex 510-560/FT 580/LP 590 for TRITC). Micrographs were taken with the same optical conditions and using Kodak T-Max P3200 film which was developed according to manufacturer recommendations (Kodak T-Max developer, 11 rain at 21~
For the rapid measurement of intensity of green and red fluores- cence from individual cells, the following procedure was used. Anti- rabbit-IgG FITC and anti-mouse-IgG TRITC, at different dilutions, were spotted out onto Whatman-1 filter paper (10 ILl per spot) and the resultant spots were used to assess whether there was a direct corre- lation between fluorocrome concentration and the exposure time in the microscope, 3200 ASA. A constant fading time of 60 seconds was used before sample measurements. The fluorescence intensity from randomly selected individual ceils was determined using this proce- dure.
RESULTS
Methodological Considerations. In Figure 1 is shown the absorbance values obtained after measuring the in- tensity of the bands in the immunoblots versus the cell protein concentration. The intensity of the bands was directly related to the protein content and was linear over the concentrations tested as demonstrated by linear regression analysis. This was also found for vimentin and pure tubulin (data not shown).
Astrocyte Cultures. Under our conditions and as described previously (12), the cytoskeletal protein ac- counted for approximately 30% of total cell protein in
x l O e
~ 1 C
O
~ e �84
V- 4
5 w ~: 2
4 8 12 16 20 24 4 8 12 16 20 24 28
ASTROCYTE PROTEIN (,ug)
Fig. 1. Correlation between the GFAP or oL-tubulin content and astro- cyte protein concentration. The GFAP and a-tubulin content were determined by densitometric analysis of the immunoblots and results are expressed as relative absorbance units. Linear regression analysis indicates a strong correlation (r=0.992 for GFAP and r=0.982 for a-tubulin). Similar results were obtained for vimentin.
all the periods studied except at day 4 when the recovery was greater (=35-38%). In Figure 2 gel electrophoresis of Triton X-100 insoluble cytoskeleton protein shows three major bands identified as vimentin (tool wt 58,000), GFAP (tool wt 51,000) and actin (mol wt 43,000) which e.g. at 7 days represented 24%, 5%, and 25%, respec- tively, of the total cytoskeletal protein. Neither GFAP nor vimentin was found in the soluble fraction, as dem- onstrated by immunoblotting and Coomasie Blue stain- ing. In contrast, a band corresponding to actin was always observed (data not shown). The photograph of Figure 2 also shows that in 4-day cytoskeletal preparations there is a fourth band, located between vimentin and GFAP. This band was found to be a protein of a molecular weight of 54,000 and was identified as tubulin by im- munoblotting. At this point in the culture period, as de- termined by scanning densitometry, tubulin was estimated to be 8.5% of total cytoskeletal protein. Therefore the presence of o~-tubulin in the cytoskeletal protein at 4 days, may explain the increase in the yield of the Triton- insoluble fraction at this time.
GFAP and Vimentin Development. GFAP and vi- mentin content in cytoskeleton preparations were deter- mined at several times during the primary culture of astrocytes obtained from brains of rat fetuses both con- trol and prenatally exposed to alcohol (PEA).
To analyze the evolution of GFAP and vimentin content, 7 Ixg of cytoskeletal protein were used per lane. This concentration is in the range of the protein concen- tration used in Figure 1. Therefore, changes in the in- tensity of GFAP-staining in Figure 3-A,B are due to differences in GFAP content. In control astrocytes, the GFAP content, (expressed as absorbance units/rag cell
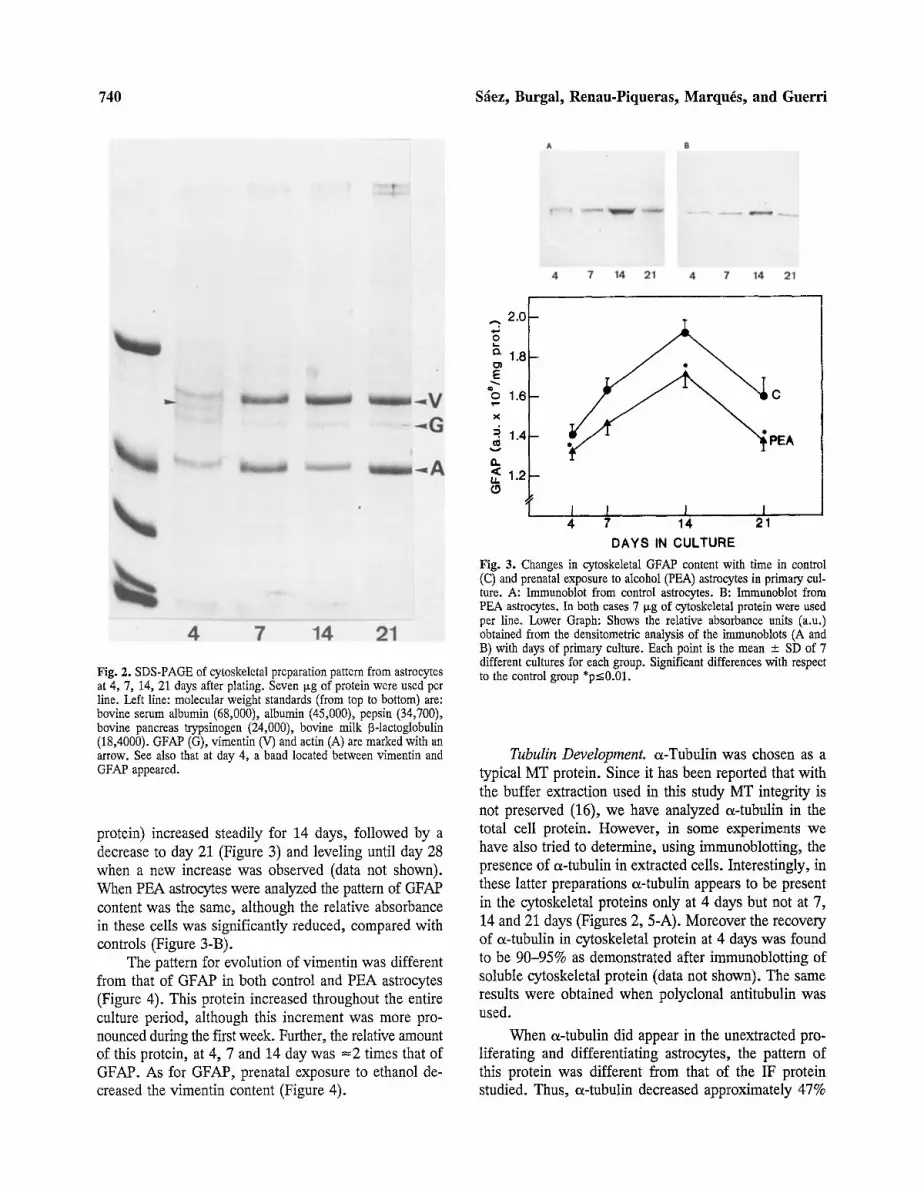
740 S~ez, Burgal, Renau-Piqueras, Marquis, and Guerri
A a
[ [
4 7 14 21 4 7 14 21
-.,V --G
4 7 Fig. 2. SDS-PAGE of cytoskeletal preparation pattern from astrocytes at 4, 7, 14, 21 days after plating. Seven ixg of protein were used per line. Left line: molecular weight standards (from top to bottom) are: bovine serum albumin (68,000), albumin (45,000), pepsin (34,700), bovine pancreas trypsinogen (24,000), bovine milk 13-1actoglobulin (18,4000). GFAP (G), vimentin (V) and actin (A) are marked with an arrow. See also that at day 4, a band located between vimentin and GFAP appeared.
protein) increased steadily for 14 days, followed by a decrease to day 21 (Figure 3) and leveling until day 28 when a new increase was observed (data not shown). When PEA astrocytes were analyzed the pattern of GFAP content was the same, although the relative absorbance in these cells was significantly reduced, compared with controls (Figure 3-B).
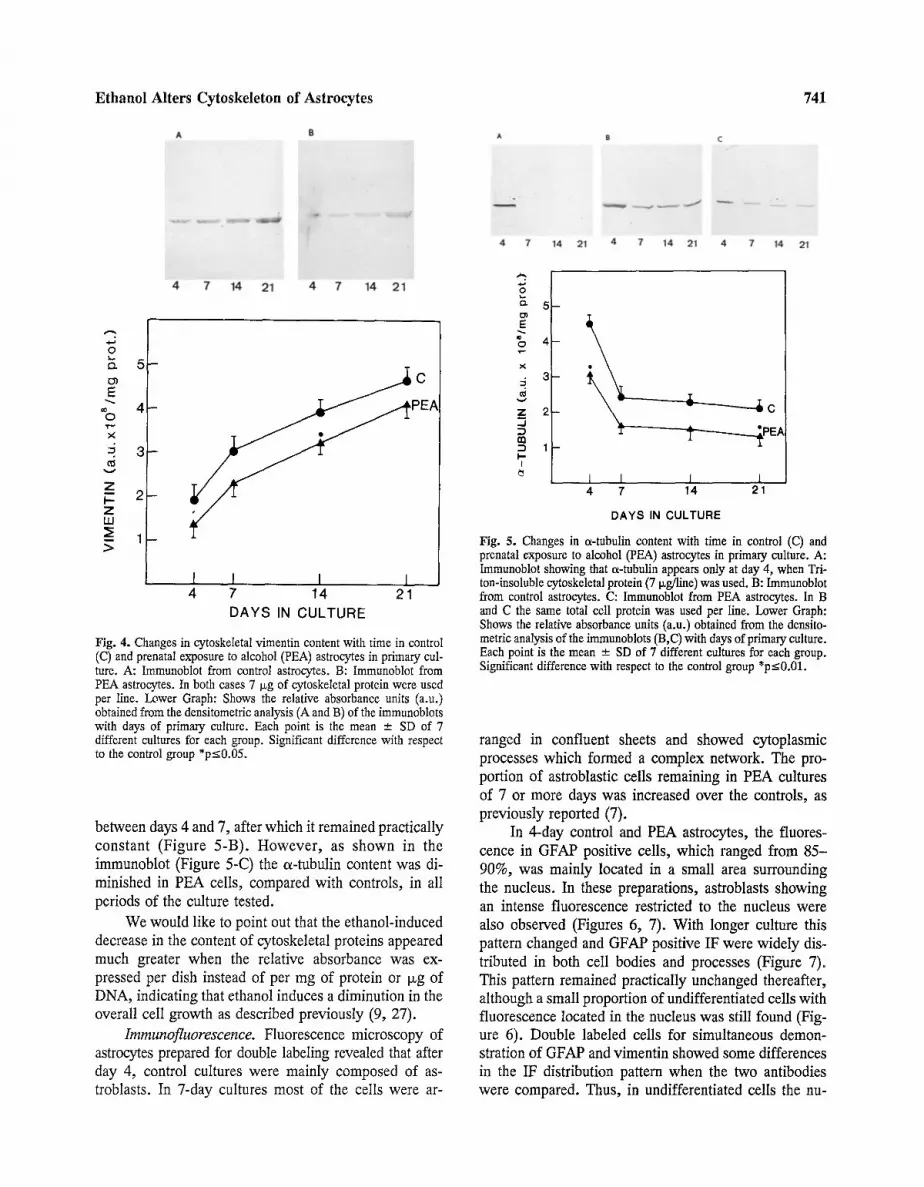
The pattern for evolution of vimentin was different from that of GFAP in both control and PEA astrocytes (Figure 4). This protein increased throughout the entire culture period, although this increment was more pro- nounced during the first week. Further, the relative amount of this protein, at 4, 7 and 14 day was =2 times that of GFAP. As for GFAP, prenatal exposure to ethanol de- creased the vimentin content (Figure 4).
A
L-
O'J
E % , f -
X
G.
r
2 . 0 B
1 . 8 -
1 . 6 -
1 . 4 -
1 . 2 -
oC
PEA
I I I I 4 7 14 21
DAYS IN CULTURE
Fig. 3. Changes in cytoskeletal GFAP content with time in control (C) and prenatal exposure to alcohol (PEA) astrocytes in primary cul- ture. A: Immunoblot from control astrocytes. B: Immunoblot from PEA astrocytes. In both cases 7 Ixg of cytoskeletal protein were used per line. Lower Graph: Shows the relative absorbance units (a.u.) obtained from the densitometric analysis of the immunoblots (A and B) with days of primary culture. Each point is the mean --- SD of 7 different cultures for each group. Significant differences with respect to the control group *p<_0.01.
Tubulin Development. a-Tubulin was chosen as a typical MT protein. Since it has been reported that with the buffer extraction used in this study MT integrity is not preserved (16), we have analyzed e~-tubulin in the total cell protein. However, in some experiments we have also tried to determine, using immunoblotting, the presence of a-tuhulin in extracted cells. Interestingly, in these latter preparations a-tubulin appears to be present in the cytoskeletal proteins only at 4 days but not at 7, 14 and 21 days (Figures 2, 5-A). Moreover the recovery of a-tubulin in cytoskeletal protein at 4 days was found to be 90-95% as demonstrated after immunoblotting of soluble cytoskeletal protein (data not shown). The same results were obtained when polyclonal antitubulin was used.
When c~-tubulin did appear in the unextracted pro- liferating and differentiating astrocytes, the pattern of this protein was different from that of the IF protein studied. Thus, a-tubulin decreased approximately 47%
Ethanol Alters Cytoskeleton of Astrocytes 741
A
~!!~ii~i !~i!~ii!i'~ii~i! i!i!! i~ ~ ~ i ~
4 7 14 21 4 7 14 21
k.. r'}
03 E
% x
5
v
Z 1 I-- Z LLI
>
5 -
4 -
3 -
2 -
1 -
I I I I 4 7 14 21
DAYS IN CULTURE
Fig. 4. Changes in cytoskeletal vimentin content with time in control (C) and prenatal exposure to alcohol (PEA) astrocytes in primary cul- ture. A: Immunoblot from control astrocytes. B: Immunoblot from PEA astrocytes. In both cases 7 Isg of cytoskeletal protein were used per line. Lower Graph: Shows the relative absorbance units (a.u.) obtained from the densitometric analysis (A and B) of the immunoblots with days of primary culture. Each point is the mean - SD of 7 different cultures for each group. Significant difference with respect to the control group *p<0.05.
between days 4 and 7, after which it remained practically constant (Figure 5-B). However, as shown in the immunoblot (Figure 5-C) the c~-tubulin content was di- minished in PEA cells, compared with controls, in all periods of the culture tested.
We would like to point out that the ethanol-induced decrease in the content of cytoskeletal proteins appeared much greater when the relative absorbance was ex- pressed per dish instead of per mg of protein or txg of DNA, indicating that ethanol induces a diminution in the overall cell growth as described previously (9, 27).
Immunofluorescence. Fluorescence microscopy of astrocytes prepared for double labeling revealed that after day 4, control cultures were mainly composed of as- troblasts. In 7-day cultures most of the cells were ar-
A B C
C
PEA
4 7 14 21 4 7 14 21 4 7 14 21
o. 5
E 2" o 4 ,r'*
X
Z 2 ,,.J :3
"~ 1 l - -
I
1 I I 1 4 7 14 21
DAYS IN CULTURE
Fig. 5. Changes in oL-tubulin content with time in control (C) and prenatal exposure to alcohol (PEA) astrocytes in primary culture. A: Immunoblot showing that cx-tubulin appears only at day 4, when Tri- ton-insoluble cytoskeletal protein (7 ~g/line) was used. B: Immunoblot from control astrocytes. C: Immunoblot from PEA astrocytes. In B and C the same total cell protein was used per line. Lower Graph: Shows the relative absorbance units (a.u.) obtained from the densito- metric analysis of the immunoblots (B,C) with days of primary culture. Each point is the mean __ SD of 7 different cultures for each group. Significant difference with respect to the control group *p_<0.01.
ranged in confluent sheets and showed cytoplasmic processes which formed a complex network. The pro- portion of astroblastic cells remaining in PEA cultures of 7 or more days was increased over the controls, as previously reported (7).
In 4-day control and PEA astrocytes, the fluores- cence in GFAP positive ceils, which ranged from 85- 90%, was mainly located in a small area surrounding the nucleus. In these preparations, astroblasts showing an intense fluorescence restricted to the nucleus were also observed (Figures 6, 7). With longer culture this pattern changed and GFAP positive IF were widely dis- tributed in both cell bodies and processes (Figure 7). This pattern remained practically unchanged thereafter, although a small proportion of undifferentiated ceils with fluorescence located in the nucleus was still found (Fig- ure 6). Double labeled cells for simultaneous demon- stration of GFAP and vimentin showed some differences in the IF distribution pattern when the two antibodies were compared. Thus, in undifferentiated cells the nu-
742 Sfiez, Burgal, Renau-Piqueras, Marquis, and Guerri
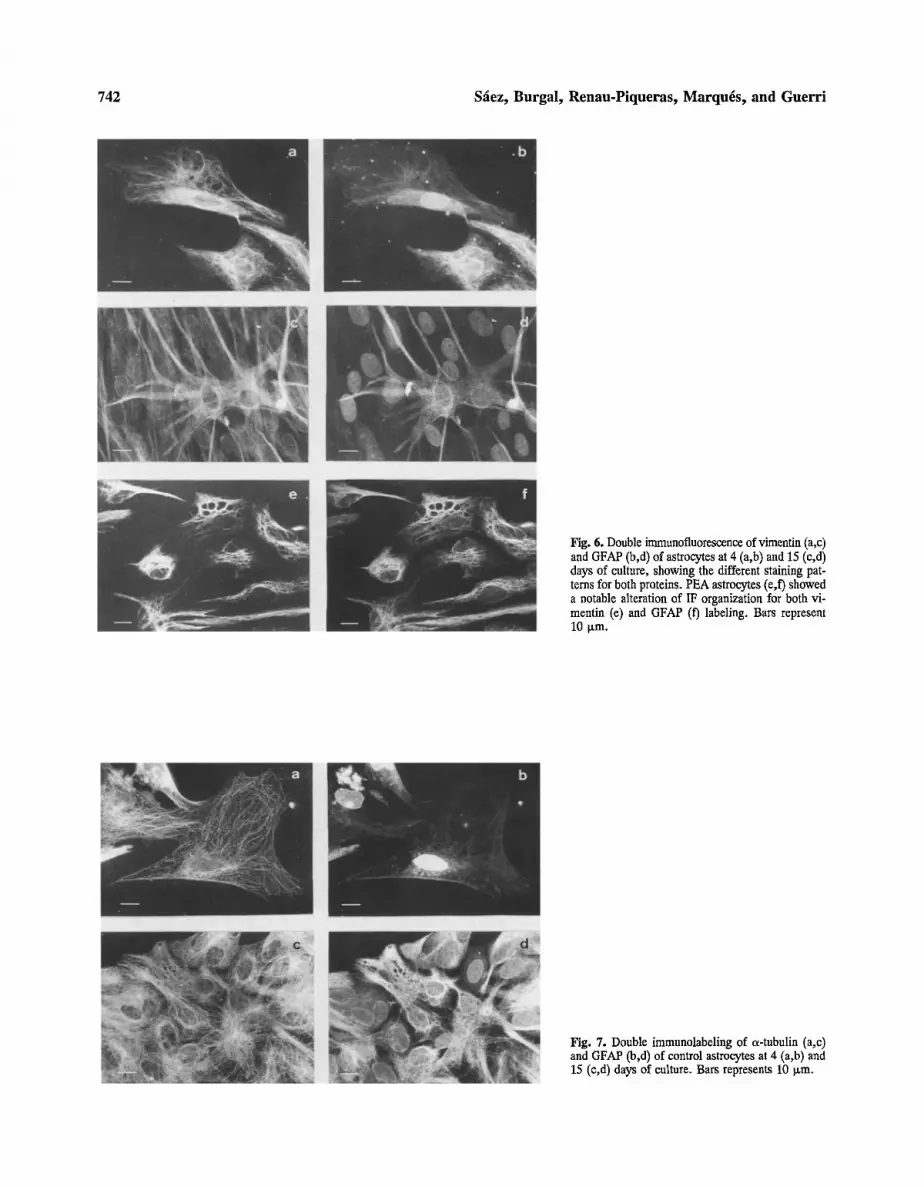
Fig. 6. Double immunofluorescence of vimentin (a,c) and GFAP (b,d) of astrocytes at 4 (a,b) and 15 (c,d) days of culture, showing the different staining pat- terns for both proteins. PEA astrocytes (e,f) showed a notable alteration of IF organization for both vi- mentin (e) and GFAP (f) labeling. Bars represent 10 Ixm.
Fig. 7. Double immunolabeling of a-tubulin (a,c) and GFAP (b,d) of control astrocytes at 4 (a,b) and 15 (e,d) days of culture. Bars represents 10 Ixm.

Ethanol Alters Cytoskeleton of Astrocytes 743
cleus was labeled for GFAP but not for vimentin, except for the nuclear envelope. These cells also contained a number of IF which showed fluorescence only for vi- mentin (Figure 6). In some intermediate and mature cells there were a number of IF showing only GFAP expres- sion (Figure 6).
On the other hand when 4-day control and PEA cultures were labeled with e~- or [3-tubulin, cells in dif- ferent phases of mitosis were frequently found showing an intense fluorescence located in MT of the spindle (Figure 8). These elements were, however, unstained when anti-GFAP or anti-vimentin were used in double- labeling experiments (Figure 8). We have further ob- served, in interphase cells, that the fluorescence pattern for both tubulins was clearly different from that of IF (Figures 7, 8) at all the culture times studied.
An interesting finding was the presence, in most interphase astrocytes of 4 and 5 days, of cytoplasmic globular particles staining intensely positive for both tub- ulins (Fig. 8). These structures, located mainly in the perinuclear region, were not stained with anti-GFAP in double fluorescence preparations (Figure 8), when this antibody and anti-~ or anti-[3-tubulin were used simul- taneously. The presence of these particles coincided with a poor development of MT and, conversely, when cells showed a well developed MT system, these structures were absent (Figures 7, 8). Thus, in astrocytes of 7 days, showing a relatively developed cytoskeleton, few glob- ular particles were found, and there were none in dif- ferentiated astrocytes with a well-developed MT system
(Figure 7). These globular particles were found in both control and PEA ceils.
Quantitative analysis during astrocyte development of the content of GFAP, vimentin and a-tubulin showed a pattern similar to that found using immunoblotting.
PEA astrocytes showed, in all the culture periods studied, a decrease in the content of the different cyto- skeletal proteins analyzed, compared with controls, as well as important alterations in the organization of both IF and MT (Figure 6).
Immunoblotting of GFAP During Brain Develop- ment. To determine whether the effect of prenatal ex- posure to ethanol on glial development which we observed in vitro was also reflected "in vivo", we have deter- mined this protein in developing whole rat brain and the results are presented as a percentage of that found in adult control brain. The protein loading for immuno- blotting was arranged so that the blots corresponded quantitatively to different amounts of GFAP.
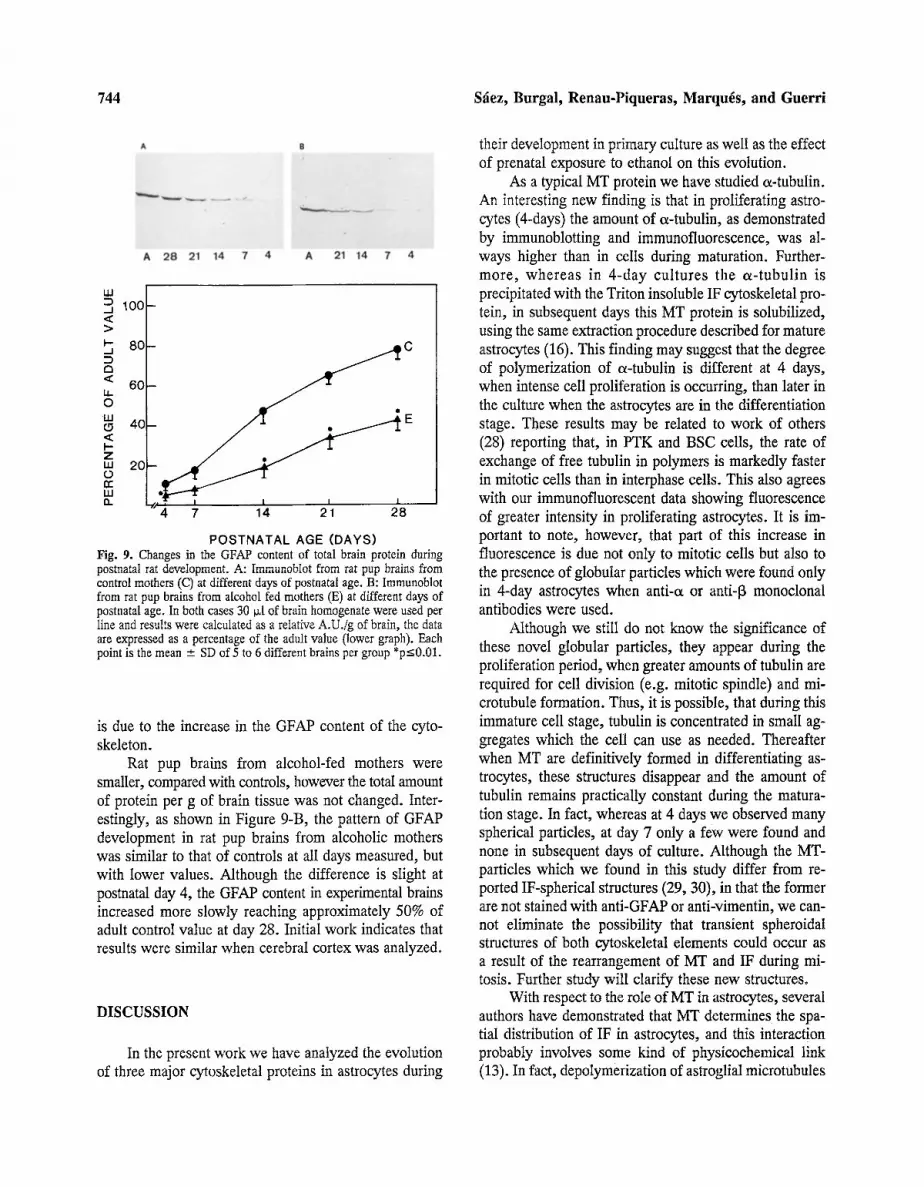
As shown in Figure 9-A, GFAP content in whole control rat pup brains, expressed as relative absorbance per gram wet weight, shows a progressive increase from 4 to 28 days, reaching approximately 80% of adult val- ues by day 28. Proteolytic degradation products of GFAP, were not observed under our experimental conditions.
It is important to note that, since it has been dem- onstrated that not more than 2% of the total GFAP was in the soluble fraction and this percentage remains con- stant during brain development (18), our results indicate that the increase in total GFAP during brain development
Fig. 8. Immunofluorescence showing the presence of spherical particles stained for a-tubulin (a) but not for GFAP (b) from 4 day astrocytes. In (c) and (d) are shown the staining patterns of mitotic cells labeled for tubulin (c) and GFAP (d). Bars rep- resents 10 ~m.
744 S~ez, Burgal, Renau-Piqueras, Marquis, and Guerri
!i;: ii?!i !i~
A 28 21 14 7 4 A 21 14 7 4
�9 ~ 100 > ~, 80
< 60
4C
,,z, 2c
~
i �9 �9 E
4 7 14 21 28
POSTNATAL AGE (DAYS) Fig. 9. Changes in the GFAP content of total brain protein during postnatal rat deveIopment. A: Immunoblot from rat pup brains from control mothers (C) at different days of postnatal age. B: Immunoblot from rat pup brains from a[cohol fed mothers (E) at different days of postnatal age. In both cases 30 ~[ of brain homogenate were used per iine and results were calculated as a reIative A.U./g of brain, the data are expressed as a percentage of the adult value (lower graph). Each point is the mean - SD of 5 to 6 different brains per group *p<0.01.
is due to the increase in the GFAP content of the cyto- skeleton.
Rat pup brains from alcohol-fed mothers were smaller, compared with controls, however the total amount of protein per g of brain tissue was not changed. Inter- estingly, as shown in Figure 9-B, the pattern of GFAP development in rat pup brains from alcoholic mothers was similar to that of controls at all days measured, but with lower values. Although the difference is slight at postnatal day 4, the GFAP content in experimental brains increased more slowly reaching approximately 50% of adult control value at day 28. Initial work indicates that results were similar when cerebral cortex was analyzed.
DISCUSSION
In the present work we have analyzed the evolution of three major cytoskeletal proteins in astrocytes during
their development in primary culture as well as the effect of prenatal exposure to ethanol on this evolution.
As a typical MT protein we have studied a-tubulin. An interesting new finding is that in proliferating astro- cytes (4-days) the amount of a-tubulin, as demonstrated by immunoblotting and immunofluorescence, was al- ways higher than in cells during maturation. Further- more, whereas in 4-day cultures the a-tubulin is precipitated with the Triton insoluble IF cytoskeletal pro- tein, in subsequent days this MT protein is sohbilized, using the same extraction procedure described for mature astrocytes (16). This finding may suggest that the degree of polymerization of c~-tubulin is different at 4 days, when intense cell proliferation is occurring, than later in the culture when the astrocytes are in the differentiation stage. These results may be related to work of others (28) reporting that, in PTK and BSC cells, the rate of exchange of free tubulin in polymers is markedly faster in mitotic cells than in interphase cells. This also agrees with our immunofluorescent data showing fluorescence of greater intensity in proliferating astrocytes. It is im- portant to note, however, that part of this increase in fluorescence is due not only to mitotic cells but also to the presence of globular particles which were found only in 4-day astrocytes when anti-~ or anti-[3 monoclonal antibodies were used.
Although we still do not know the significance of these novel globular particles, they appear during the proliferation period, when greater amounts of tubulin are required for cell division (e.g. mitotic spindle) and mi- crotubule formation. Thus, it is possible, that during this immature cell stage, tubulin is concentrated in small ag- gregates which the cell can use as needed. Thereafter when MT are definitively formed in differentiating as- trocytes, these structures disappear and the amount of tubulin remains practically constant during the matura- tion stage. In fact, whereas at 4 days we observed many spherical particles, at day 7 only a few were found and none in subsequent days of culture. Although the MT- particles which we found in this study differ from re- ported IF-spherical structures (29, 30), in that the former are not stained with anti-GFAP or anti-vimentin, we can- not eliminate the possibility that transient spheroidal structures of both cytoskeletal elements could occur as a result of the rearrangement of MT and IF during mi- tosis. Further study will clarify these new structures.
With respect to the roIe of MT in astrocytes, several authors have demonstrated that MT determines the spa- tial distribution of IF in astrocytes, and this interaction probably involves some kind of physicochemical link (13). In fact, depolymerization of astroglial microtubules

Ethanol Alters Cytoskeleton of Astrocytes 745
by colchicine provokes a simultaneous disassembly of the gliofilaments (13, 14).
Our results also demonstrate that in both primary culture and during postnatal brain development "in vivo", GFAP increases as a result of astroglia differentiation, which implies an increase in the number of gliofilaments as well as their organization with other components of the cytoskeleton such as MT. These data corroborate other "in vivo" results showing that the concentration of GFAP increases steadily throughout development of the animal (18, 31, 32, 33).
In addition, the increasing concentration of GFAP in cultures with maximum at day 14, is in accordance with previous studies (17, 34). It has been suggested that this peak corresponds to the burst of astroglial dif- ferentiation at the time of myelination (34). Further, it has recently been reported that GFAP-mRNA also fol- lowed this biphasic evolution (35)~
In contrast to GFAP, information about the content of vimentin as well as on the possible co-expression of this protein with GFAP have been reported only during brain development "in vivo" (32, 36). Under the culture conditions used here, the content of vimentin increased throughout the culture period, with the pattern differing somewhat from that of GFAP. Although it has been pos- tulated that vimentin is the major protein of IF in im- mature glia and that the expression "'in vitro" of this protein may be due to an alteration in cell maturation (32), we have shown that astrocytes differentiate after 10 days in primary culture, (7, 8, 9, 27). Such increases correlate with the "in vivo" results obtained in astro- cytes isolated from rat brain (8). Moreover, vimentin positive astrocytes in 14-day, or longer cultures showed in double fluorescence experiments, that the shape, IF distribution pattern and GFAP staining correspond to mature astrocytes. Relative to the time during develop- ment when vimentin appears, it has been reported that in mammalian embryos, the expression of this protein generally precedes that of GFAP (36). Indeed, at 4 days we have found more vimentin than GFAP and many immature cells showed IF expressing only vimentin. Also it has been reported that the expression of GFAP seems to coincide with the onset of myelination (32). In addi- tion, in glioma and differentiating astrocytes GFAP and vimentin are frequently found to be co-expressed (37, 38, 39) forming heteropolymers (38). This raises the question whether GFAP molecules expressed during de- velopment are incorporated into pre-existing vimentin filaments or whether they form independent homopoly- mers some of which may later interact, forming heter- opolymers while others are maintained as GFAP or
vimentin homopolymers at the end of astrocyte matu- ration. Thus, it has been demonstrated in cultured human glioma cells that although in a large proportion of the ceils these elements are organized in heteropolymer fil- aments, some long homopolymers which stained as either GFAP or vimentin were also found (38, 39). Our results agree with these observations since we have found var- iations in GFAP and vimentin fluorescence staining in some cells.
Another important result from this study is that ex- posure to ethanol during brain development alters the cytoskeleton structure of astrocytes, also affecting gli- ogenesis "in vivo". This is supported by the following: (a) The levels of both IF cytoskeletal proteins GFAP and vimentin decrease during the proliferation and differen- tiation of astrocytes in primary culture. The decrease in GFAP was not due to an increase in degradation prod- ucts, since neither of these products were detected in either control or in PEA astrocytes. In addition, we have previously demonstrated that PEA astrocytes fail to de- velop processes or acquire a filamentous IF distribution pattern (7). (b) The levels of astrocyte cx-tubulin were diminished in PEA astrocytes compared with control cells. (c) GFAP levels in brains of rats exposed to alcohol pre- and postnatally during gliogenesis "in vivo", decrease as much as to ~-35% by day 28 of the postnatal period, compared with controls.
In view of these results the question arises, what is the mechanism(s) by which ethanol is able to alter the astrocyte cytoskeleton and thereby glial development? It is known that both acute and chronic ethanol intake alter cytoskeletal elements including IF and MT from adult liver (40, 41). It has been postulated that the MT alter- ations may be due to the binding of acetaldehyde with liver tubulin, decreasing the amount of polymerized tub- ulin and therefore the number of microtubules (40, 42). Since we have demonstrated that fetus brains are ex- posed "in utero" to both alcohol and acetaldehyde (8), this mechanism could also be implicated in the effect on tubulin observed in astrocytes and therefore on IF. Fur- thermore, since many astrocytes are derived from radial glia, established very early in development (43, 44) and since possibly the decision determining the levels of GFAP is also made during the "in titero" stage, alcohol-in- duced changes in nutrients and growth factors during this stage could (45, 46) alter the normal GFAP expression during glial development. In fact, GFAP levels in astro- cytes in culture seem to be modulated by certain hor- mones and factors which may act via cyclic-AMP- dependent systems (47).
Finally, whatever mechanism accounts for the

746 Sfiez, Burgal, Renau-Piqueras, Marqu6s, and Guerri
ethanol-induced alterations in the cytoskeleton of astro- gliaI celIs, damage to astrocytes becomes very important in understanding neuronal response to ethanol exposure when one considers how closely neuronal maintenance is related to astrocyte function. Thus, it is known that the cytoskeleton of astrocytes creates a framework which constitutes the scaffolding that guides neuronal migra- tion and differentiation (6, 43, 48). Ethanol induced glial alterations could therefore be one of the mechanisms underlying the CNS dysfunctions observed in children with FAS and in animals prenatally exposed to alcohol.
ACKNOWLEDGMENTS
We thank M. March and I. Bosc~ for their expert technical as- sistance. This research was supported by DGICYT (PB87-0505) and FISss 90/0897. C.G. and J.R-P. are members of the K.U.M.C.-I.I.C. Exchange Program.
REFERENCES
1. Streissguth, A. P., and Martin, J. C. 1983. Prenatal effects of alcohol abuse in humans and laboratory animals. Vol. 7, Pages 539-589, in Kissin, B., Begleiter, H. (eds.), The pathogenesis of alcoholism, Plenum Press, New York.
2. Streissguth, A. P., Bookstein, F. L., Sampson, P. D., and Barr, H. M. 1989. Neurobehavioral effects of prenatal alcohol: Part III. PLS analyses of neuropsychologic test. Neurotoxicol. and Teratol. 11:493-507.
3. Abel, E. L., and Sokol, R. J. 1986. Fetal alcohol syndrome is now leading cause of mental retardation. Lancet 2:1222.
4. Miller, M. W. 1986. Effects of alcohol on the generation and migration of cerebral cortical neurons. Science 233:1308-1311.
5. Clarren, S. K., Alvord, E. C. Jr., Sumi, S. M., Streissguth, A. P., and Smith, D. W. 1978. Brain malformations related to pre- natal exposure to ethanol. J. Peds. 92:64-67.
6. Bradford, H. F. 1986. Glial cells: Mechanical and functional sup- porting cells of the nervous system. Pages 62-117, in Bradford, H. F. (ed.), Chemical Neurobiology, W. H. Freeman and Com- pany, New York.
7. Renan-Piqueras, J., Zaragozfi, R., De Paz, P., Bfiguena-Cervel- Iera, R., Mejias, L., and Guerri, C. 1989. Effects of prolonged ethanol exposure on the glial fibrillary acidic protein-containing intermediate filaments of astrocytes in primary culture: A quan- titative immunofluorescence and immunogold electron micro- scopic study. J. Histochem. Cytochem. 37:229-240.
8. Guerrri, C., Marquds, A., Sancho-Tello, M., and Renau-Pi- queras, J. 1989. Effect of prenatal exposure to alcohol on mem- brane-bound enzymes during astrocyte development in vivo and in primary culture. Int. J. Dev. Biol. 33:239-244.
9. Guerri, C., S~ez, R., Sancho-Tello, M., Martin de Aguilera, E. and Renau-Piqueras, J. 1990. Ethanol alters astrocyte develop- ment: A study of critical periods using primary cultures. Neuro- chem. Res. 15:559-565.
10. Trimmer, P. A., Reier, P. J., Tae H.-O., and Eng, F. E. 1982. An ultrastructural and immunocytochemical study of astrocyfic differentiation in vitro. Changes in the composition and distribu- tion of the cellular cytoskeleton. J. Neuroimmunoi. 2:235-260.
11. Goldman, R. D., Goldman, A. E., Green, K. J., Jones, J. C. R., Jones, S. M., and Yang, H. Y. 1986. Intermediate filament net-
works: Organization and possible functions of a diverse group of cytoskeletal elements. J. Cell Sei. 5:69-97.
12. Chin, F. -C., Norton, W. T., and Fields, K. L. 1981. The cy- toskeleton of primary astrocytes in culture contains actin, glial fibriUary protein, vimentin. J. Neurochem. 37:147-155.
13. Ciesielski-Treska, J., Bader, M. -F., and Aunis, D. 1982. Mi- crotubular organization in flat epitheloid and stellate process-bear- ing astrocytes in culture. Neurochem. Res. 7:275-286.
14. Goetschy, J. -F., Ulrich, G., Aunis, D., and Ciesielski-Treska, J. 1986. The organization and solubility properties of intermediate filaments and microtubules of cortical astrocytes in culture. J. Neurocytol. 15:375-387.
15. Eng, L. F., Vanderhaeghen, J. J., Bignami, A., and Gerstl, B. 1971. An acidic protein isolated from fibrous astrocytes. Brain Res. 28:351.
16. Chiu, F. -C., and Goldman, J. E. 1984. Synthesis and turnover of cytoskeletal proteins in cultured astmcytes. J. Neurochem. 42:166-174.
17. Senseubrenner, M., Devilliers, G., Bock, E., and Porte, A. 1980. Biochemical and ultrastructural studies of cultured rat astmglial cells. Effect of brain extract and dibutyril cyclic AMP on glial fibrillary acidic protein and glial filaments. Differentiation. 17:51- 61.
18. Malloeh, G. D. A., Clark, J. B., and Burnet, F. R. 1987. Glial fibrillary acidic protein in the cytoskeletal and soluble protein fractions of the developing rat brain. J. Neuroehem. 48:299-306.
19. Lieber, C. S., and DeCarli, L. M. 1976. Animal models of ethanol dependence and liver injury in rats and baboons. Fed. Proc. 35:1232- 1236.
20. Sanchis, R., Sancho-Tello, M., and Guerri, C. 1986. The effects of chronic alcohol consumption on pregnant rats and their off- spring. Alcohol and Alcoholism. 21:295-305.
21. Guerri, C., Esquifino, A., Sanchis, R., and Grisolia, S. 1984. Growth, enzymes and hormonal changes in offspring of alcohol- fed rats. Pages 85-102, in Mechanisms of alcohol damage in utero, Ciba Foundation Symposium 105, Pitman, London.
22. Guerri, C., and Grisolfa, S. 1982. Effects of prenatal and postnatal exposure of rats to alcohol: changes in (Na*-K+)ATPase. Phar- macol. Biochem. and Behav. 17:927-932.
23. Lowry, O. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J. 1951. Protein measurement with Folin phenol reagent. J. Biol. Chem. 193:265-275.
24. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly of the head of the bacteriophage "i"4. Nature. 227:680-- 685.
25. Towbin H., Staehelin T., and Gordon J. 1979. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets. Procedure and some applications. Froc. Natl. Acad. Sci. 76, 4350- 4354.
26. Bell, P. B., Rundquints, I., Suensson, I., and Collins, U. P. 1987. Use of cytofluorometry to evaluate binding of antibodies to the cytoskeleton of cultured ceils. J. Histochem. Cytochem. 35:1381- 1388.
27. Renau-Fiqueras, J., Saneho-Tello, M., Zaragoz~i, R., and Guerri, C. 1988. Effect of ethanol on the development of astrocytes in primary culture. Advances in Biosciences. 71:269-273.
28. Saxton, W. M., and Mclntosh, R. 1984. Tubulin dynamics in cultured mammalian ceils. J. Cell. Biol. 99:2175-2186.
29. Franke, W., Schmid, E., and Grund, C. 1984. Intermediate fil- ament proteins in nonfilamentous structures: Transient disintegra- tion and inclusion of subunit proteins in granular aggregates. Cell. 30:103-113.
30. Horwitz, B., Kupfer, H., Eshhar, A., and Geiger, B. 1981. Re- organization of arrays of prekeratin filaments during mitosis. Exp. Cell Res. 134:281-286.
31. Bignami, A., and Dahl, D. 1974. Astrocyte specific protein and neuroglial differentiation. An immunofluorescence study with an- tibodies to the glial fibrillary acidic protein. J. Comp. Neurol. 153:27-38.

E t h a n o l A l t e r s C y t o s k e l e t o n of As t rocy te s 747
32. Dahl, D. 1981. Vimentin-GFAP protein transition in rat neuroglia cytoskeleton occurs at the time of myelination. J. Neurosci. Res. 6:741-748.
33. Weir, M. D., Patel, A. J., Hunt, A., and Thomas, D. G. T. 1984. Developmental changes in the amount of glial fibrillary acidic protein in three regions of the rat brain. Develop. Brain. Res. 15:147-154.
34. Bock, E., Moller, M., Nissen, C., and Sensenbrenner, M. 1977. Glial fibrillary acidic protein in primary astroglial cell cultures derived from newborn rat brain. FEBS Lett. 83:207-211.
35. Tardy, M., Fages, C., Riol, H., LePrince, G., Rataboul, P., Charriere-Bertrand, C., and Nufiez, J. 1989. Developmental expression of the glial fibrillary acidic protein mRNA in the cen- tral nervous system and cultured astrocytes. J. Neurochem. 52:162- 167.
36. Schnitzer, J., Werner, W. F., and Shachner, M. 1981. Immu- nocytochemical demonstration of vimentin in astrocytes and epen- dymal cells of developing and adult mouse nervous system. J. Cell Biol. 90:435-447.
37. Osborn, M., Ludwig-Festl, M., Weber, K., Bignami, A., Dahl, D., and Bayrenther, K. 1981. Expression of glial and vimentin type intermediate filaments in cultures derived from human glial material. Differentiation. 19:161-167.
38. Quilan, R. A., and Franke, W. W. 1983. Molecular interactions in intermediate-sized filaments revealed by chemical cross-link- ing. Eur. J. Biochem. 132:477-484.
39. Paetman, A. 1988. Glial fibrillary acidic protein, vimentin and
fibronectin in primary cultures of human glioma and fetal brain. Acta Neuropathol. 75:448--455.
40. Matsuda, Y., Baraona, E., Salaspuro, M., and Lieber, C. S. 1979. Effects of ethanol on liver microtubules and Golgi appa- ratus. Lab. Invest. 41:455--463.
41. French, S. W., Katsuma, Y., Ray, M. B., and Swierenga, S. H. H. 1987. Cytoskeletal pathology induced by ethanol. Ann. N.Y. Acad. Sci. 492:262-275.
42. Tuma, D. J., and Sorrell, M. F. 1987. Functional consequences of acetaldehyde binding to proteins. Alcohol and Alcoholism. 1:61- 66.
43. Rakic, P. 1988. Specification of cerebral cortical areas. Science. 241:170-176o
44. McKay, R. D. G. 1989. The origins of cellular diversity in the mammalian central nervous system. Cell. 58:815--821.
45. Henderson, G. I., Patwardhn, R. V., Hoynnpa, A. M., and Scheuker, S. 1981. Fetal alcohol s3/ndrome: Overview of patho- genesis. Neurobehav. Toxicol. Teratol. 3:73-80.
46. Henderson, G. I., Heitman, D. W., and Scheuker, S. 1987. Ef- fects of ethanol on rat placental and fetal hepatocyte transport of amino acids. Ann. N.Y. Acad. Sci. 492:224-232.
47. Morrison, R. S., DeVellis, J., Lee, Y. L., Bradshaw, R. A., and Eng, L. F. 1985. Hormones and growth factors induce the syn- thesis of glial fibrillary acidic protein in rat brain astrocytes. J. Neuros. Res. 14:167-176.
48. Hatten, M. E., and Liem, R. K. H. 1981. Astroglial cells provide a template for the positioning of developing cerebellar neurons in vitro. J. Cell. Biol. 90:622-630.