xvavedila zaragoza simposio anual de · la especificidad se determinó frente a adn de diferentes...
TRANSCRIPT

Simposio anual de
AVEDILA ZaragozAXV
18 y 19 de Noviembre de 2010
Asociación de veterinarios especialistas en diagnóstico laboratorial


Simposio anualXV
18 y 19 de Noviembre de 2010
zaragoza
Asociación de veterinarios especialistas en diagnóstico laboratorial


XV Simposio anual de AVEDILA
Comité organizador:
Mª Pilar Alvarez Marzo
Nicolás Abancens Tejero
Marta Fernández Baltanás
Mari Carmen Bernal Laborda
Juan José Alvarez Pardo
Juan Pedro Bueso Fran
Raúl Mainar Jaime
Marisa Pardo Izquierdo
Comité científico:
Ramón Juste Jordán
Concha Caballero Galván
Carmina Nogareda Burch
Gorka Aduriz Rekalde
Carlos Alvarez Pacios
Joseph Carrera Mauri
Pere Busquets Relats
Marta Eulalia García Sánchez
José Marín Sánchez Murillo
Lourdes Porquet Garanto


presentacion


9
presentacion
Estimados compañeros:
La Asociación de Veterinarios Especialistas en Diagnóstico Laboratorial AVEDILA ha elegido como sede, para su XV Simposio Anual, la ciudad de Zaragoza. Como miembros del Comité organizador, es para nosotros un inmenso honor y privilegio el poder recibir y acoger a congresistas de toda España, y esperamos cumplir dos objetivos que se repiten simposio tras simposio: aglutinar una gran número de profesionales preocupados en buscar y aplicar nuevas tecnologías diagnósticas mediante el intercam-bio de conocimiento científico en aspectos claves de la sanidad animal; y por otro lado poder disfrutar de la hospitalidad, gastronomía y cultura que ofrece la ciudad de Zaragoza al visitante.
Este año el Comité científico del XV Simposio ha preparado un programa de ponencias en el que se incluyen novedades de gran actualidad como puede ser el virus de la fiebre del Nilo Occidental y se revisan otros temas, que no por conocidos dejan de estar vigentes, como la Fiebre Q.
Agradeceros vuestra colaboración con la aportación de vuestros trabajos científicos, tanto en formato poster como de comunicación oral, y que nos brinda una gran oportunidad para poder compartir todo este conocimiento y experiencias con estos trabajos.
También agradecemos a todas aquellas instituciones y empresas que han patrocinado este evento y que con sus aportaciones tanto económicas como de todo tipo, han hecho posible su celebración.
Esperamos que la estancia entre nosotros y el contenido científico del simposio sean del agrado de todos.
Recibe un cordial saludo.
El Comité Organizador


ponencias


13
ponencias
CARACTERIZACIÓN DE CEPAS DE COXIELLA CIRCULANTES EN ESPAÑA
Isabel Jado1, Margarita Bolaños2, Ana L. García Pérez3, Cristina García-Amil1, Horacio Gil1, Raquel Escudero1, Pedro Anda1
1: Laboratorio de Espiroquetas y Patógenos Especiales, Instituto de Salud Carlos III2: Hospital Universitario Insular de Gran Canaria
3: Instituto Vasco de Investigación y Desarrollo Agrario (NEIKER Tecnalia)
IntroducciónLas manifestaciones clínicas de la infección aguda por Coxiella burnetii presentan claras diferencias entre áreas geográficas, produciendo desde un cuadro respiratorio que puede desembocar en neu-monía, hasta cuadros de fiebre de duración intermedia con mayor o menor afectación hepática. La evolución a cuadros crónicos como endocarditis, hepatitis granulomatosa, infección vascular y osteo-mielitis, entre otras, parece claramente relacionada con lesiones valvulares previas, en el caso de la endocarditis, no existiendo una hipótesis establecida para el resto de las manifestaciones crónicas.
Dada la dificultad de aislamiento de este agente, que requiere el uso de cultivos celulares e instalacio-nes de nivel 3 de bioseguridad, la disponibilidad global de cepas y, por tanto, de metodología de ca-racterización para el establecimiento de relaciones clínicas, es insuficiente. Con la reciente publicación de la secuencia completa del genoma de C. burnetii se ha descrito la existencia de diferentes grupos genómicos (GG), aunque dada la dificultad de la metodología empleada, su relación con las diferentes manifestaciones clínicas no ha sido evaluada. En este estudio presentamos un método molecular mul-tiplex para la determinación de los diferentes GG y su aplicación en cepas y muestras autóctonas.
Material y MétodosSe localizaron en el genoma 7 genes cuya ausencia determinaba la pertenencia de los aislados a cada uno de los GG descritos. Se diseñaron cebadores y sondas específicos para cada uno de los genes que, posteriormente, se multiplexaron en un único protocolo de amplificación. Los resultados se revelaron mediante hibridación en fase sólida con sondas específicas diseñadas para cada una de las 7 dianas. La especificidad se determinó frente a ADN de diferentes especies bacterianas cercanas (Anaplasma, Ehrlichia, Bartonella, Borrelia, Francisella, Rickettsia, Chlamydia, Legionella, Mycoplasma, Orientia, Leptospira, Brucella y Treponema), así como frente a ADN humano.
ResultadosLa sensibilidad para cada una de las dianas se situó entre 10 y 100 equivalentes de genoma y la especificidad resultó del 100% con las muestras ensayadas. Todos los aislados disponibles en la colección del CNM, así como muestras positivas de pacientes, fueron clasificados por este método, observándose ciertas asociaciones entre cuadro clínico y GG.

14
ponencias
Según los resultados obtenidos, se confirma una elevada variabilidad de cepas de C. burnetii que circulan en España. Por una parte, en el norte peninsular (País Vasco y Navarra) hemos detectado 4 GG. En Canarias, las cepas clínicas de origen agudo (que producen síndrome febril con afectación hepática) se agrupan con la cepa de referencia de origen crónico (Priscilla). Además, este resultado se encuentra apoyado por los datos de secuenciación del gen icd, coincidiendo por tanto con los estudios filogenéticos realizados previamente. La mayoría de las cepas/muestras procedentes de pa-cientes con enfermedad crónica (a excepción de 2 casos) se agrupan dentro del mismo GG, lo que coincide con lo descrito previamente en la literatura. Se trata de variantes de C. burnetii con un ge-noma reducido, lo que redundaría en un crecimiento más lento y, por tanto, en la estimulación de una respuesta inmune atenuada, lo que correlacionaría con su habilidad de producir cuadros crónicos. Un caso más extremo de este proceso se observó en algunas de las muestras procedentes de Canarias obtenidas a partir de pacientes y de Rattus norvegicus, que exhibieron una reducción aún mayor de sus genomas lo que llevó asociado un impedimento considerable para el cultivo celular, originando un reducido número de aislamientos. Por otra parte, tanto las cepas de referencia asociadas con enfermedad aguda, como una muestra respiratoria del norte del país mostraron todos los elementos genéticos analizados, agrupándose dentro del mismo GG.
ConclusionesAdemás de los valiosos datos sobre la circulación de los diferentes genotipos de Coxiella en nuestro país, destacar que se ha puesto a punto un método para la caracterización de la bacteria, tanto a partir de cepas como de muestras (clínicas y medioambientales), que va a permitir la realización de futuros estudios clínicos y epidemiológicos, así como la identificación y trazado de brotes.

15
ponencias
FIEBRE Q: ESTUDIOS DE CAMPO, DIAGNÓSTICO DE LABORATORIO Y CONTROL
Ana L. García-Pérez, Ianire Astobiza, Jesús F. Barandika, Ana Hurtado, Ramón A. JusteNEIKER-Instituto Vasco de Investigación y Desarrollo Agrario
Berreaga 1, 48160 Derio, Bizkaia
La fiebre Q en rumiantes domésticosLa fiebre Q es una zoonosis causada por la bacteria Coxiella burnetii, microorganismo de amplia distribución y que afecta a varias especies animales, incluyendo el hombre. Coxiella burnetii es muy persistente en el medio ambiente manteniéndose en la naturaleza a través de dos ciclos, un ciclo do-méstico del que forman parte los animales de granja y los animales de compañía (perros, gatos), y un ciclo silvestre en el que está implicada la fauna silvestre y las garrapatas.
En el ganado bovino la infección por C. burnetii origina problemas de infertilidad, metritis y mamitis. En el ganado ovino y caprino da lugar a abortos, endometritis, infertilidad, partos prematuros y un bajo peso de las crías, llegando a afectar hasta el 50% del rebaño. Los abortos por C. burnetii son tardíos, hacia el final de la gestación, y frecuentemente los animales presentan retención placentaria. La placentitis es el síntoma más característico. La placenta es más densa y puede contener grandes cantidades de exudado amarillento-marrón. Los animales no gestantes raramente muestran síntomas tras la primera infección, y aunque se sabe que la vía de infección más frecuente es la respiratoria, la coxiellosis en animales no causa alteraciones de tipo respiratorio, ni problemas hepáticos o cardiacos en infecciones de tipo crónico, como en la especie humana, sino que se localiza en el aparato repro-ductivo de la hembra, incluida la glándula mamaria, desde donde la infección se eliminará al medio ambiente una vez que los animales estén gestantes. La gestación parece ser un momento crítico para la reactivación de la infección con eliminación de numerosas bacterias con el feto, la placenta y fluidos amnióticos, tanto en el caso de que se produzca el aborto, como durante el parto normal (Woldehiwet, 2004; Astobiza y cols., 2010a). Además se produce la excreción de C. burnetii por la orina, heces, así como por el calostro y la leche. También se ha detectado en el semen, la lana, el pelo de los animales, estiércol, etc. (Maurin y Raoult, 1999).
EpidemiologíaTras el aborto o el parto, los periodos de eliminación de C. burnetii varían entre las especies animales, pero está descrito que las vacas eliminan la bacteria de forma prolongada durante varios meses y durante años sucesivos, por lo que representan un riesgo mayor de perpetuar la infección si se com-
16
ponencias
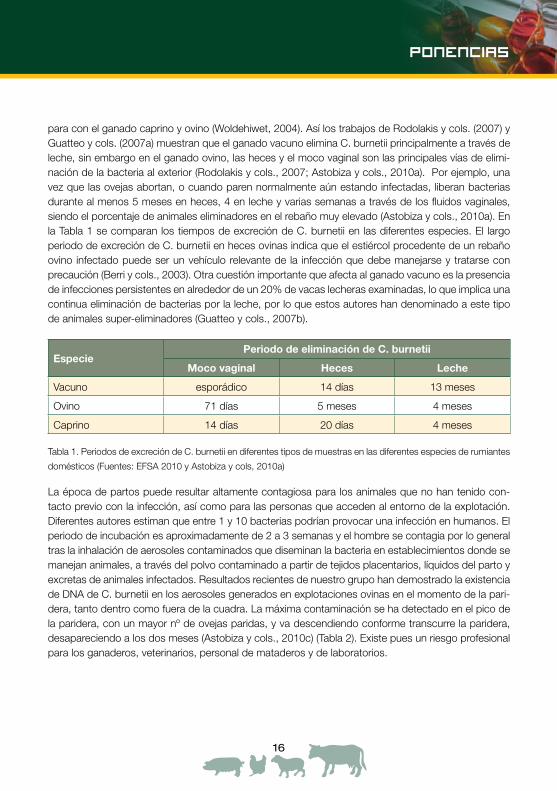
para con el ganado caprino y ovino (Woldehiwet, 2004). Así los trabajos de Rodolakis y cols. (2007) y Guatteo y cols. (2007a) muestran que el ganado vacuno elimina C. burnetii principalmente a través de leche, sin embargo en el ganado ovino, las heces y el moco vaginal son las principales vías de elimi-nación de la bacteria al exterior (Rodolakis y cols., 2007; Astobiza y cols., 2010a). Por ejemplo, una vez que las ovejas abortan, o cuando paren normalmente aún estando infectadas, liberan bacterias durante al menos 5 meses en heces, 4 en leche y varias semanas a través de los fluidos vaginales, siendo el porcentaje de animales eliminadores en el rebaño muy elevado (Astobiza y cols., 2010a). En la Tabla 1 se comparan los tiempos de excreción de C. burnetii en las diferentes especies. El largo periodo de excreción de C. burnetii en heces ovinas indica que el estiércol procedente de un rebaño ovino infectado puede ser un vehículo relevante de la infección que debe manejarse y tratarse con precaución (Berri y cols., 2003). Otra cuestión importante que afecta al ganado vacuno es la presencia de infecciones persistentes en alrededor de un 20% de vacas lecheras examinadas, lo que implica una continua eliminación de bacterias por la leche, por lo que estos autores han denominado a este tipo de animales super-eliminadores (Guatteo y cols., 2007b).
EspeciePeriodo de eliminación de C. burnetii
Moco vaginal Heces Leche
Vacuno esporádico 14 días 13 meses
Ovino 71 días 5 meses 4 meses
Caprino 14 días 20 días 4 meses
Tabla 1. Periodos de excreción de C. burnetii en diferentes tipos de muestras en las diferentes especies de rumiantes
domésticos (Fuentes: EFSA 2010 y Astobiza y cols, 2010a)
La época de partos puede resultar altamente contagiosa para los animales que no han tenido con-tacto previo con la infección, así como para las personas que acceden al entorno de la explotación. Diferentes autores estiman que entre 1 y 10 bacterias podrían provocar una infección en humanos. El periodo de incubación es aproximadamente de 2 a 3 semanas y el hombre se contagia por lo general tras la inhalación de aerosoles contaminados que diseminan la bacteria en establecimientos donde se manejan animales, a través del polvo contaminado a partir de tejidos placentarios, líquidos del parto y excretas de animales infectados. Resultados recientes de nuestro grupo han demostrado la existencia de DNA de C. burnetii en los aerosoles generados en explotaciones ovinas en el momento de la pari-dera, tanto dentro como fuera de la cuadra. La máxima contaminación se ha detectado en el pico de la paridera, con un mayor nº de ovejas paridas, y va descendiendo conforme transcurre la paridera, desapareciendo a los dos meses (Astobiza y cols., 2010c) (Tabla 2). Existe pues un riesgo profesional para los ganaderos, veterinarios, personal de mataderos y de laboratorios.

17
ponencias
Fecha muestreo Nº ovejas paridas
Nº Coxiella en aerosoles Nº ovejas eliminadoras de Coxiella al parto / analizadas
(%)Exterior cuadra
Interior cuadra
28/11/2008 92 1.69 3.47
Ovejas adultas: 61/91 (67%) Primalas: 0/8 (0%)
11/12/2008 32 Neg 1.72
22/12/2008 60 Neg 2.51
29/01/2009 29 Neg 2.56
02/03/2009 3 Neg Neg
02/04/2009 2 Neg Neg
Tabla 2. Contaminación medioambiental en el parto: carga bacteriana en los aerosoles del interior y del exterior de una instalación ovina (logaritmo Coxiella/filtro aire), y porcentaje de ovejas eliminadoras de DNA de C. burnetii en fluidos vaginales tras el parto.
Algunos estudios serológicos realizados en España han puesto de manifiesto una alta seroprevalencia en el ganado caprino de la Comunidad de Canarias y en el ganado vacuno de la Comunidad de Madrid (Pascual-Velasco, 1996; Rodríguez y cols., 2010), pero, en general, los estudios realizados en España hasta la fecha son escasos. En el País Vasco, en un estudio serológico reciente sobre la distribución de C. burnetii en especies de rumiantes en manejo extensivo se ha comprobado que la seropreva-lencia en las explotaciones ovinas es más alta (73.9%) que en ganado vacuno de carne (42.9%) o en caprino (45.5%), al igual que la seroprevalencia individual, más alta en ovino (12.3%) que en el resto de especies (8.3% en caprino, 6.6% en vacuno de carne) (Ruiz-Fons y cols., 2010). En el ganado vacuno lechero en régimen intensivo hemos encontrado un 48.2% de explotaciones con al menos un animal seropositivo, y una seroprevalencia individual media del 6.5% (García-Pérez y cols., 2010). Si bien las seroprevalencias halladas en el ganado caprino y vacuno son inferiores a las del ovino, a la vista de los resultados, estas especies también parecen tener relevancia en el ciclo doméstico de transmisión de fiebre Q al hombre (Tabla 3). Sin embargo, según un análisis de los brotes de fiebre Q en humanos a nivel mundial, se considera que el ganado ovino y caprino son la fuente principal de contagio para el hombre (Angelakis y Raoult, 2010; EFSA, 2010).
Especie Seroprevalencia explotaciones Seroprevalencia individual
Nº Nº Pos (%) Nº Nº Pos (%)
Vacuno leche 193 93 (48.2) 2.692 176 (6.5)
Vacuno carne 42 18 (42.9) 618 41 (6.6)
Ovino 46 34 (73.9) 1.298 160 (12.3)
Caprino 11 5 (45.5) 109 9 (8.3)
Tabla 3. Seroprevalencia de la fiebre Q en las especies de rumiantes domésticos en el País Vasco (Ruiz-Fons y cols.,
2010; y datos sin publicar)

18
ponencias
La infección por ingestión de leche y derivados lácteos contaminados es un factor menor en la trans-misión de C. burnetii, ya que las temperaturas de pasteurización de la leche y, probablemente, el proceso de curado de los quesos eliminan la bacteria. Por otra parte, se han observado casos en humanos en los que no se ha podido demostrar el contacto directo con animales; se sabe que las partículas aéreas que contienen los microorganismos pueden ser transportadas por el viento a gran distancia, incluso a más de 800 m y que permanecen viables en condiciones adversas durante largo tiempo, lo que explicaría la falta de antecedentes de contacto con animales en algunos pacientes. Coxiella burnetii es muy resistente en la naturaleza y puede sobrevivir durante varias semanas en zo-nas donde haya habido animales. Se ha llegado a aislar C. burnetii en suelos donde se habían retirado los animales 6 meses antes (Maurin y Raoult, 1999).
También se ha demostrado que la infección por C. burnetii está presente en otros animales domésti-cos (Lang, 1990; Pascual Velasco, 1996), si bien su importancia parece ser mucho menor. Es digno de mención un caso reciente en Soria en el que todos los miembros de una familia enfermaron por fiebre Q y se asoció con el parto de su mascota (una gata) (García de la Cruz y cols., 2010) Así mismo, la infección se ha detectado en numerosos animales silvestres cuya importancia parece radicar en su papel como reservorio, manteniendo y difundiendo la infección a través de las garrapatas. Se han involucrado diferentes especies debido a evidencias serológicas y moleculares. Así recientemente se ha detectado su presencia en ratones domésticos y ratones de campo en el entorno de las explota-ciones ovinas con antecedentes de abortos por fiebre Q (Barandika y cols., 2007) y en cérvidos del sur y norte de España (Ruiz-Fons y cols., 2008). Resultados recientes de nuestro grupo analizando por PCR muestras de bazo y/o pulmón también apuntan a otras especies como el jabalí, el corzo, la liebre europea y algunas especies de aves rapaces como posibles partícipes en el ciclo silvestre de C. burnetii, presentando prevalencias de entre el 4.3% y el 14.3%, dependiendo de la especie analizada (Astobiza y cols., 2010b).
Por otra parte, la fiebre Q está incluida en el grupo de enfermedades transmitidas por garrapatas, pero el papel de las garrapatas en la epidemiología de la fiebre Q es complejo y controvertido, y apa-rentemente, su mayor importancia podría ser como reservorio de la infección. En España, el primer aislamiento a partir de garrapatas, y asociado a los primeros casos de fiebre Q, fue a finales de los años 40 y principios de los 50, siendo las principales especies implicadas de los géneros Hyalomma y Rhipicephalus. Más recientemente con la aplicación de técnicas moleculares se ha observado la presencia de DNA de C. burnetii en diversas especies de garrapatas. Así, Toledo y cols. (2008) en la zona centro de España encontraron un 7% de garrapatas de la vegetación positivas, siendo las garrapatas del género Hyalomma las que mostraban una prevalencia más alta. En algunas zonas del Norte de España como el País Vasco, por el contrario, no parece que las garrapatas jueguen un papel importante ya que su presencia ha sido detectada a niveles mínimos (Barandika y cols., 2008), si bien es cierto que las garrapatas del género Hyalomma y otras especies que han resultado positivas en la zona centro no son frecuentes en el norte.

19
ponencias
Diagnóstico de laboratorioLa infección por C. burnetii se diagnostica confirmando la presencia del agente en muestras de fetos y placentas procedentes de brotes de abortos. La muestra de placenta es fundamental para el diag-nóstico laboratorial, ya que la infección suele focalizarse a nivel de placenta, donde la carga bacteriana es elevada y las lesiones son importantes. Una simple tinción de Stamp, es decir, la observación de bacterias de morfología compatible con Coxiella spp. en una impronta de placenta, proporciona infor-mación de gran valor de forma rápida y económica.
Las pruebas indirectas de detección de anticuerpos en sangre son las más ampliamente usadas. Se cuenta con métodos ELISA comerciales que tienen una alta sensibilidad y son fácilmente estanda-rizables, y sobre todo son los métodos de elección para la realización de estudios epidemiológicos (García-Pérez y cols., 2009; Ruiz-Fons y cols., 2010). En explotaciones con diagnóstico de fiebre Q se han hallado seroprevalencias en torno al 50% en ganado ovino (Astobiza y cols., 2010a), del 75% en ganado caprino (NEIKER, servicio de diagnóstico), y en torno al 50% en ganado vacuno (Ruiz-Fons y cols., 2009). Para la detección de infecciones recientes la Fijación de Complemento puede ser la técnica de elección, que goza de gran especificidad, aunque su sensibilidad es sensiblemente inferior al ELISA. Títulos de anticuerpos entre 1/10 y 1/40 se consideran infecciones latentes, y superiores a 1/40 indican que la infección está en fase evolutiva (OIE, 2008). En rumiantes domésticos existe un porcentaje de animales infectados que no seroconvierte (Rodolakis y cols., 2007; Rousset y cols., 2009), por lo que para el diagnóstico individual es necesario combinar técnicas serológicas y mole-culares (PCR).
Los métodos de aislamiento de C. burnetii precisan de instalaciones de nivel de seguridad 3, por lo que actualmente la técnica más usada para la detección e identificación de este agente es la PCR, que se basa en la amplificación del DNA específico de C. burnetii, técnica aplicable además a diversos tipos de muestras, tales como tejidos fetales (incluyendo placenta), hisopos vaginales de hembras abortadas, muestras de leche, heces, garrapatas, etc. La importancia de la incorporación de las téc-nicas moleculares al diagnóstico de la fiebre Q en muestras de abortos ovinos quedó reflejada en el estudio de Oporto y cols. (2006), en el que las técnicas convencionales aportaban una incidencia de C. burnetii como causa de aborto en el 3% de los rebaños estudiados, pero al incorporar las técnicas moleculares se pudo llegar a detectar DNA de C. burnetii en el 9% de rebaños, lo que supone preva-lencias similares a los abortos por Salmonella o Clhamydophila, en la zona de estudio.
Además, las técnicas serológicas y moleculares se pueden utilizar a gran escala para realizar estudios epidemiológicos y así conocer la distribución de C. burnetii en determinadas poblaciones animales. Las muestras de leche de tanque (LT) son muestras representativas de la explotación que permiten estimar si la infección puede estar presente en una explotación (Guatteo y cols., 2007b). Nuestra experiencia nos hace recomendar su uso, ya que mediante la investigación por ELISA y PCR de 154 muestras de leche de tanque ovina, conocemos la distribución y prevalencia de la fiebre Q en explo-taciones ovinas de la zona (García-Pérez y cols., 2009).

20
ponencias
Recientemente hemos realizado un estudio en 178 explotaciones de ganado bovino lechero en el País Vasco. En general se detectaron anticuerpos en el suero lácteo en el 66.9% de explotaciones. A la par que se tomaban muestras de LT, en cada explotación se obtuvieron unas 15 muestras de suero de animales de todas las edades. En las explotaciones cuya muestra de LT fue negativa a anticuerpos, la seroprevalencia media en los individuos analizados fue del 2.6% (3.5% en las vacas adultas). Las ex-plotaciones con LT positiva (+) mostraron una seroprevalencia media en los animales del 5.8% (11.7% en vacas), y finalmente la seroprevalencia media en animales de las explotaciones positivas (++) fue del 11.6% (22.9% en vacas). El nivel de anticuerpos en LT presentó una mejor correlación con el nivel de seroprevalencia de las hembras adultas, (R2=0.21, P<0.001), sin embargo la correlación no fue alta, lo que puede achacarse a la falta de seroconversión frente a Coxiella, mencionado anteriormente.
Cuando las muestras de LT de bovino lechero se analizaron mediante PCR se detectó DNA de C. burnetii en el 51.4% de las explotaciones. Las explotaciones con muestras de LT PCR positivo y ELISA positivo mostraron un nivel medio de seroprevalencia significativamente más elevado (9.1±1.1%) que las explotaciones con LT negativas a ambas técnicas (2.6±0.8%) (F=16.3, df=1, P<0.001).
La incorporación de técnicas de PCR a tiempo real nos han permitido cuantificar la carga bacteriana en las muestras de LT positivas por la PCR convencional, estando correlacionada significativamente con el nivel de anticuerpos en suero de LT (R2=0.31, P<0.0001).
Como conclusión, a la vista de estos resultados y de otros estudios realizados en ovino lechero (Gar-cía-Pérez y cols, 2009), se puede decir que C. burnetii está ampliamente distribuida en las explota-ciones ovinas y bovinas lecheras. El análisis de LT mediante ELISA tiene un gran valor para hacer una primera estimación de la presencia de C. burnetii en la explotación, ya que existe una correlación significativa con el nivel de anticuerpos y con la carga bacteriana, si bien debe de considerarse que las infecciones introducidas recientemente en la explotación pueden no ser detectadas en LT por ELISA debido a la baja prevalencia de animales infectados. Sin embargo, el ELISA aplicado a leches de tan-que es un método económico, y que puede ser aplicado en estudios epidemiológicos para conocer la situación en una determinada zona o región, o incluso para plantear programas de erradicación en ganado lechero, tal y como se han venido planteando los programas de erradicación de BVD en algunos países nórdicos.
Control de la fiebre QLas alternativas de control incluyen el tratamiento con antibióticos y la vacunación. Las tetraciclinas son el antibiótico de elección en el tratamiento de la fiebre Q, pero no es totalmente efectivo, pues tras el doble tratamiento con oxitetraciclina, los animales siguen siendo eliminadores de C. burnetii (Astobiza y cols., 2010b), aunque parece ser que los abortos cesan. Así, puesto que en los pequeños rumiantes esta infección puede cursar con un brote explosivo de abortos, la aplicación generaliza-da de preparados a base de este compuesto puede estar indicada para intentar paliar las posibles pérdidas económicas. En el ganado vacuno hay que hacer una cuidadosa evaluación de los datos clínico-epidemiológicos disponibles y de los resultados del laboratorio antes de aplicar este tipo de

21
ponencias
tratamientos, debido a las implicaciones que tiene en salud pública la aplicación de antibióticos en esta especie.
Por lo tanto, en los rumiantes domésticos, la manera más eficaz de controlar la infección sería la vacunación. Entre sus efectos beneficiosos se citan la disminución drástica de la tasa de animales excretores, una reducción significativa de la excreción de bacterias en la leche, en el moco vaginal y en las heces, así como una mayor protección frente al aborto. En estos momentos, no se dispone de una vacuna comercial registrada en España para rumiantes domésticos, pero existe una vacuna de fase I (COXEVAC, CEVA Salud Animal) comercializada excepcionalmente en algunos países y que ha dado unos excelentes resultados a nivel experimental (Arricau-Bouvery y cols., 2005). En explotacio-nes comerciales, nuestro grupo está evaluando la eficacia de la vacunación en la especie ovina, y si bien el primer año no se obtienen resultados significativos, al tercer año de vacunación no se detecta la eliminación de Coxiella en los animales recién paridos (Astobiza, datos sin publicar). En ganado va-cuno existe el estudio de Guatteo y cols. (2008) que ha ensayado la misma vacuna en explotaciones comerciales de ganado vacuno lechero infectadas. El resultado más relevante es el efecto protector de la vacunación en la recría, es decir, en el grupo de animales más jóvenes en el que la infección todavía no se ha extendido suficientemente. Los autores recomiendan vacunar a todo el efectivo en explotaciones no infectadas, y, en aquellas que lo están, prestar especial atención a la reposición. Otros estudios realizados en vacuno demuestran el efecto positivo de la vacunación a medio plazo, en un periodo de 4 años (EFSA, 2010).
Además, otras medidas generales para el control de la fiebre Q en rumiantes incluyen el mantenimien-to de unas condiciones higiénicas adecuadas en las explotaciones, sobre todo en el momento de los partos, con eliminación rápida de fetos y placentas, aislamiento de los animales afectados, evitando partos y la salida de los animales en el exterior de la cuadra. Además el uso de guantes y mascarillas por parte del personal de la explotación es imprescindible para reducir las posibilidades de contagio cuando se tocan productos derivados de los partos y/o abortos, así como realizar desinfecciones de las instalaciones con productos adecuados (Rodolakis y Cochonneau, 2009), y evitar la aplicación del estiércol como abono (Berri y cols, 2003). También es importante evitar el acceso a la explotación de visitas, y la utilización de calzas desechables cuando se permanezca en la cuadra y/o ambiente contaminado.
Como en el caso de otras infecciones, sería posible identificar a los animales portadores y eliminarlos del rebaño, siempre y cuando se combinen las técnicas de ELISA y PCR. El saneamiento tiene dos li-mitaciones: una que es caro porque requiere realizar controles continuos para evitar que se produzcan nuevas infecciones y otra que no se sabe si saneando la infección podría persistir en determinadas especies silvestres y domésticas sobre las que no se actúe.
El sector veterinario, los ganaderos y la Administración deben de ser conscientes de la existencia de esta zoonosis, que es un problema de Salud Pública. En Holanda, tras el grave brote de fiebre Q en humanos se han tomado una serie de medidas drásticas y dramáticas para los productores (EFSA,

22
ponencias
2010). En España, hasta hace no mucho tiempo la fiebre Q ha sido un problema que no se ha tenido en cuenta como causa de abortos y/o fallo reproductivo en rumiantes domésticos. Afortunadamente, hoy en día los productores son cada vez más conscientes de la necesidad de implementar medidas de bioseguridad que le protejan a sí mismo y a los demás contra diversas infecciones con implicaciones tanto sanitarias como productivas. Para reducir riesgos de entrada de C. burnetii en la explotación, se debe de transmitir una serie de recomendaciones a los ganaderos. La primera sería el restringir el acceso dentro de las instalaciones a personas ajenas a la explotación ya que ésta puede ser una de las vías de entrada de la infección. También hay que verificar la negatividad a anticuerpos (serología) y antí-geno (PCR) de C. burnetii en las nuevas compras de animales, pues la llegada de animales portadores es la vía de entrada más común. Finalmente, es necesario comprobar periódicamente la negatividad de la explotación a través de chequeos frecuentes en muestras de leche de tanque, si la explotación es de aptitud lechera, o mediante serologías periódicas de un porcentaje de animales elegidos al azar. Mientras los resultados sean negativos, bastará con mantener las medidas de bioseguridad. Si se de-tectase la aparición de fiebre Q, habría que realizar estudios individuales para identificar el punto y mo-mento de entrada y tratar con antibióticos de forma inmediata. A continuación se deberían solicitar los permisos necesarios para vacunar el efectivo total de animales, de forma que se dificulte el arraigo de nuevas infecciones, y monitorizar a la recría. Esta pauta de vacunación debería tener una periodicidad anual hasta que se demuestre la ausencia de infección. En todo caso, es muy importante el trabajo conjunto de ganaderos, veterinarios y autoridades sanitarias para la erradicación de esta zoonosis.
AgradecimientosAgradecemos la financiación proporcionada por INIA (RTA 2009-00017-00), y por el Departamento de Agricultura del Gobierno Vasco.
Referencias
Angelakis, E., Raoult, D. (2010). Q fever (Review). Vet. Microbiol, 140: 297-309.
Astobiza, I., Barandika, J.F., Hurtado, A., Juste, R.A., García-Pérez, A.L. (2010a). Kinetics of Coxiella burnetii excretion in a commercial dairy flock after treatment with Oxitetracycline. Vet. J., 184: 172-175.
Astobiza, I., Barral, M., Ruiz-Fons, F., Barandika, J.F., Gerrikagoitia, X., Hurtado, A., García-Pérez, A.L. (2010b). Investigation of Coxiella burnetii occurrence in wildlife and ticks. Vet. Microbiol (en prensa).
Astobiza, I, Barandika, JF, Ruiz-Fons, F., Hurtado, A., Povedano, I., Juste, R.A., García-Pérez, AL. (2010c). Coxiella burnetii shedding and environmental contamination at lambing in two highly naturally-infected dairy sheep flocks after vaccination. Res. Vet. Sci. (en revisión).
Arricau-Bovery, N., Soriau, A., Moutoussamy, A., Ladenise, K., Rodolakis, A., (2001). Etude de léx-crétion de Coxiella burnetii dans un modèle expérimental caprin et décontamination des lisiers par la

23
ponencias
cyanamide calcique. Rec. Rech. Rumin 8, 153-156.
Arricau-Bouvery, N., Souriau, A., Bodier, C., Dufour, P., Rousset, E., Rodolakis, A. (2005). Effect of vaccination with phase I and phase II Coxiella burnetii vaccines in pregnant goats. Vaccine 23, 4392-4402.
Barandika, J. F., Hurtado, A., Garcia-Esteban, C., Gil, H., Escudero, R., Barral, M., Jado, I., Juste, R. A., Anda, P., Garcia-Perez, A. L. (2007). Tick-borne zoonotic bacteria in wild and domestic small mammals in northern Spain. Appl.Environ.Microbiol. 73, 6166-6171.
Barandika JF, Hurtado A, García-Sanmartín J, Juste RA, Anda P, García-Pérez AL. (2008). Prevalence of tick-borne zoonotic bacteria in questing adult ticks from northern Spain. Vector Borne Zoonotic Dis. 8(6): 829-35.
Berri M, Rousset E, Champion JL, Arricau-Bouvery N, Russo P, Pepin M, Rodolakis A. (2003). Ovine manure used a a garden fertiliser as a suspected source of human Q fever. Vet. Rec. 153:269-70.
EFSA Panel on Animal Health and Welfare (AHAW); Scientific Opinion on Q Fever. EFSA Journal 2010; 8(5):1595. [114 pp.]. www.efsa.europa.eu
García de Cruz, S, Aldea-Mansilla, C, Nebreda, T Campos, A. Brote familiar de fiebre Q. Enferm Infecc Microbiol Clin .2010; 28(5):325–331.
García-Pérez, A.L., Astobiza, I., Barandika, J.F. Atxaerandio, R., Hurtado, A., Juste, R.A. (2009). Inves-tigation of Coxiella burnetii occurrence in dairy sheep flocks by bulk-tank milk analysis and antibody levels determination. J. Dairy Sci., 92, 1581-1584.
García-Pérez, AL, Ruiz-Fons, F., Astobiza, I., Barandika, J.F. Hurtado, A., Juste, R.A. (2010). Epidemio-logía y control de la Fiebre Q. XV Congreso Internacional de Medicina Bovina. Granada, Junio 2010.
Guatteo R, Beaudeau F, Joly A, Seegers H. (2007a). Coxiella burnetii shedding by dairy cows. Vet Res. 38: 849-60.
Guatteo, R., Beaudeau, F., Joly, A., Seegers, H. (2007b). Assessing the within-herd prevalence of Coxiella burnetii milk-shedder cows using a real-time PCR applied to bulk tank milk. Zoonoses Public Health, 54: 191-194.
Guatteo, R., Seegers, H., Joly, A., Beaudeau, F. (2008). Prevention of Coxiella burnetii shedding in infected dairy herds using a phase I C. burnetii inactivated vaccine. Vaccine 26, 4320-4328.
Lang GH. 1990. Coxiellosis in animals. Marrie TJ, ed. Q Fever. Boca Raton, CRC Press 1990; 23-48.
Maurin, M., Raoult, D. (1999). Q fever. Clin.Microbiol.Rev. 12, 518-553.
OIE, World Organization for Animal Health (2008). Q fever. In: Manual of diagnostic test and vaccines for terrestrial animals, 6th edition. Chapter 2.1.12. pp 292-303. http://www.oie.int

24
ponencias
Oporto, B., Barandika, J. F., Hurtado, A., Aduriz, G., Moreno, B., Garcia-Perez, A. L. (2006). Incidence of ovine abortion by Coxiella burnetii in northern Spain. Ann.N.Y.Acad.Sci. 1078, 498-501.
Pascual Velasco F, (1996). Fiebre Q. Junta de Castilla-León, Consejería de Sanidad y Bienestar Social.
Rodolakis, A., Berri, M., Hechard, C., Caudron, C., Souriau, A., Bodier, C. C., Blanchard, B., Camuset, P., Devillechaise, P., Natorp, J. C., Vadet, J. P., Arricau-Bouvery, N. (2007). Comparison of Coxiella burnetii shedding in milk of dairy bovine, caprine, and ovine herds. J.Dairy Sci. 90, 5352-5360.
Rodolakis, A., Cochonneau, D. (2009). Biology of Coxiella burnetii and host response to infection. Proceedings of Symposium “Q fever: An emerging disease”. European Buiatrics Forum, Marsella 2nd December 2009.
Rodriguez, NF, Carranza, C. Bolaños, M., Pérez-Arellano, JL, and C. Gutierrez (2010). Seroprevalence of Coxiella burnetii in domestic ruminants in Gran Canaria island, Spain. Transboundary and Emerging Diseases, 57: 66-67.
Rousset E, Berri M, Durand B, Dufour P, Prigent M, Delcroix T, Touratier A, Rodolakis A. (2009). Coxiella burnetii shedding routes and antibody response after outbreaks of Q fever-induced abortion in dairy goat herds. Appl Environ Microbiol. 75:428-33.
Ruiz-Fons, F., Rodriguez, O., Torina, A., Naranjo, V., Gortazar, C., de la Fuente J. (2008). Prevalence of Coxiella burnetti infection in wild and farmed ungulates. Vet Microbiol. 126, 282-286.
Ruiz-Fons, F., Astobiza, I., Barandika, J.F., Juste, R.A., Atxaerandio, R., García-Pérez, A.L. (2009). Estudio serológico de Coxiella burnetii en el ganado vacuno en sistemas de manejo extensivos e intensivos. Resultados preliminares. XIV congreso internacional de medicina bovina. A Coruña 6, 7 y 8 de mayo de 2009.
Ruiz-Fons, F., Astobiza, I., Barandika, J.F., Hurtado, A., Atxaerandio, R., Juste, R.A., García-Pérez, A.L. (2010) Serosurvey of Coxiella burnetii in domestic ruminants in semi-extensive grazing systems in northern Spain. BMC Vet. Res.6: 3.
Toledo A, Jado I, Olmeda AS, Casado-Nistal MA, Gil H, Escudero R, Anda P. (2008). Detection of Coxiella burnetii in Ticks Collected from Central Spain.Vector Borne Zoonotic Dis. 9:465-468.
Van den Brom, R., Vellema, P. (2009). Q fever outbreaks in small ruminants and people in the Nether-lands. Small Ruminant Res., 86: 74-79.
Woldehiwet Z. (2004). Q fever (coxiellosis): epidemiology and pathogenesis. Res Vet Sci. 77: 93-100.

25
ponencias
Q FEVER IN THE NETHERLANDS
Douwe Bakker and Hendrik-Jan RoestCentral Veterinary Institute of Wageningen UR
Lelystad, The [email protected]
IntroductionQ fever is a zoonosis caused by Coxiella burnetii. This is a Gram negative intracellular bacterium which is prevalent throughout the world, with the exception of New Zealand. Domestic animals are considered to be the main reservoir for Q fever in humans, although many other animal species can be infected with C. burnetii. Transmission from the animal source to humans is mainly accomplished through inhalation of contaminated aerosols. The main symptom of Q fever in goats and sheep is abortion. The main symptoms in cattle are reduced fertility and metritis, but abortion can also occur. Other animal species show no symptoms when infected. In humans, infection with C. burnetii remains asymptomatic in up to 60% of infected persons. After infection acute Q fever causes flu-like symptoms with fever, atypical pneumonia or hepatitis. In approximately 1 to 5% of the Q fever cases infection may progress into a chronic infection, often leading to endocarditis. Due to its zoonotic nature and low infectious dose, C. burnetii is a Bio Safety Level 3 organism and should be handled under appropriate laboratory conditions (2000/54/EU).
History of Q fever in the NetherlandsThe first three cases of human Q fever in the Netherlands were reported in 1956, although it is not clear if these cases were indigenous. Q fever became notifiable in 1975 and in the succeeding years it became clear that Q fever was present in the Netherlands mainly as an occupational disease. Between 1975 and 2006 on average 16 human cases were reported each year. Studies in domestic animals in the eighties showed Q fever was also present in cattle, sheep and goats, as well as in dogs and cats.
Onset of the outbreak of Q fever in the NetherlandsAbortions due to Q fever were reported in dairy goats for the first time in 2005. In this year, two abor-tion storms were reported. In 2006, six abortion storms in dairy goat herds and one in a dairy sheep herd were reported. All the dairy goat herds with abortions were located in the southern part of the Netherlands. In 2007, another seven dairy goat herds suffered abortions due to Q fever. In this year, the first human outbreak of Q fever in the Netherlands was reported, too, with in total 168 human cases. This outbreak was situated in the area with abortion storms due to Q fever in dairy goat herds. The abortion problems in dairy goats were considered to be the cause of the human Q fever due to the geographical overlap of cases, succession in time of the Q fever symptoms and the lack of other epidemiological sources.

26
ponencias
The 2007 outbreak of human Q fever was at that time considered to be a single event, like outbreaks in other countries, but in 2008 the outbreak continued with in total 1000 human cases. The start of the increase of human cases in 2008 triggered measures to prevent shedding of C. burnetii in dairy goats and dairy sheep and to prevent transmission to humans.
Response to the Q fever outbreak in the NetherlandsThe first response to the continued Q fever outbreak was mandatory notification of abortion rates over 5% in dairy goat and dairy sheep farms. This passive surveillance showed the hot spots of massive excretion of C. burnetii. In 2008, seven dairy goat farms and one dairy sheep farm were notified, and in 2009 six dairy goat farms. Additional measures were executed, such as how to handle manure, visitor bans, vermin control and the active removal of placentas from the stables, to prevent the transmission of C. burnetii from the animal source to humans. To prevent shedding of C. burnetii in dairy goats and dairy sheep, a voluntary vaccination program was started in a specified area in 2008. The vaccine of choice was the commercially available phase I vaccine. Because this vaccine was not registered in the Netherlands, the Dutch Ministry of Agriculture granted exemption. In 2009, the vaccination area was extended and the vaccination was made mandatory, but due to a lack of vaccine not all dairy goats and dairy sheep could be vaccinated in time, before breeding. In 2009, an active surveillance system was introduced via bulk tank milk testing to find all possible risk farms for human Q fever. As a result, the number of positive farms increased to about 90.
In 2009, the total number of human cases rose to 2355 and the government wanted to take all possi-ble measures to stop the increase of human cases in 2010, resulting in the culling of all pregnant goats and sheep on Q fever-positive farms and a breeding ban for non-pregnant goats.
Current Q fever situation in the NetherlandsTo date, 357 human Q fever cases were registered in 2010 and no peak could be observed like in the previous years. This is a significant decrease compared to 2009. Ninety-three dairy goat and dairy sheep farms are positive in the bulk tank milk sample.
What could be the cause of the Dutch Q fever outbreak?Dairy goats and dairy sheep are considered to be the source of the human outbreak on the basis of epidemiological investigations. This is confirmed by a small study on the genotypes of C. burnetii present in dairy goats and humans. Transmission from dairy goats and dairy sheep to humans could have been facilitated by the warm weather in the springs of 2007, 2008 and 2009. In addition, no other sources could be identified to date. But why did C. burnetii cause abortion storms in the dairy goat population? Till 2005, Q fever in dairy goats was not diagnosed nor were abortion storms seen. So what happened? One of the explanations is the strong increase of the dairy goat population. After the introduction of the milk quotation system for dairy cattle, the dairy goat industry in the Netherlands ex-panded. After the outbreaks of Classical Swine Fever in 1997 and Foot and Mouth Disease in 2001, the dairy goat population has doubled in the past ten years from nearly 100.000 in 2000 to over 230.000 in 2009. Most of these goats were held in the southern part of the Netherlands and in this population

27
ponencias
C. burnetii could be spread. Another additional explanation could be the emergence of a more virulent strain of C. burnetii that could easily spread in the dairy goat population, but also infect humans. Re-search on the virulence of the strain is ongoing. Hopefully, future research will give the answer to why the Dutch Q fever outbreak started.
Some results of ongoing researchA lot of research has been initiated in the human field, the veterinary field and in collaboration. For example Q fever strains were isolated for the first time in the Netherlands. Till 2009 there were no cul-ture facilities nor experience available. In collaboration with the Bundeswehr Institute for Microbiology, a culture method of C. burnetii was implemented at the Central Veterinary Institute of Wageningen UR. Culture is performed in a BSL 3 facility on Buffalo Green Monkey cells. Isolation from placenta material is relatively easy to perform. Strains are now used for goat experiments on the pathogenesis of C. bur-netii in goats and sequence and resistance studies are planned.
Epidemiological investigations showed that living within two kilometres from a goat farm was associa-ted with a higher risk of Q fever compared to living more than five kilometres from a farm. Genotyping of C. burnetii in clinical samples from goats revealed that one genotype is predominantly present in all dairy goats farms in the south of the country. The C. burnetii with this genotype is expected to be transmitted to humans, but no robust data from humans are available yet.
ConclusionQ fever problems surprised the Netherlands despite its presence in the years before. A close collabo-ration between the human field and the veterinary field on the level of researchers as well as on the level of policy makers was needed to battle the disease. To date, it is not clear why Q fever became a problem and hopefully, future research will give that answer.
References1. Angelakis E, Raoult D. Q Fever. Vet Microbiol. 2010 Jan 27;140(3-4):297-309.
2. Arricau-Bouvery N, Rodolakis A. Is Q fever an emerging or re-emerging zoonosis? Vet Res. 2005 May-Jun;36(3):327-49.
3. Arricau-Bouvery N, Souriau A, Bodier C, Dufour P, Rousset E, Rodolakis A. Effect of vaccina-tion with phase I and phase II Coxiella burnetii vaccines in pregnant goats. Vaccine. 2005 Aug 15;23(35):4392-402.
4. Karagiannis I, Schimmer B, Van Lier A, Timen A, Schneeberger P, Van Rotterdam B, et al. In-vestigation of a Q fever outbreak in a rural area of The Netherlands. Epidemiol Infect. 2009 Sep;137(9):1283-94.
5. Klaassen CH, Nabuurs-Franssen MH, Tilburg JJ, Hamans MA, Horrevorts AM. Multigenotype Q fever outbreak, the Netherlands. Emerging infectious diseases. 2009 Apr;15(4):613-4.

28
ponencias
6. Raoult D, Marrie TJ, Mege JL. Natural history and pathophysiology of Q fever. Lancet Infect Dis. 2005 Apr;5(4):219-26.
7. Roest HIJ, Tilburg JJHC, Van der Hoek W, Vellema P, Van Zijderveld FG, Klaassen CH, et al. The Q fever epidemic in the Netherlands: history, onset, response and reflection. Epidemiology and Infection. 2010; Accepted.
8. Roest HIJ, Ruuls RC, Buijs R, Spierenburg M, Vellema P, Dercksen D, et al. Geografical distribution of MLVA types of Coxiella burnetii isolates from goats in the Netherlands. Medical Corps Internatio-nal Forum. [abstract]. 2009;4(supplement to MCIF 4/2009):13.
9. Schimmer B, Ter Schegget R, Wegdam M, Zuchner L, de Bruin A, Schneeberger PM, et al. The use of a geographic information system to identify a dairy goat farm as the most likely source of an urban Q-fever outbreak. BMC infectious diseases. 2010;10:69.
10. Sidi-Boumedine, K1., Rousset, E1., Henning, K2., Ziller, M2., Niemczuck, K3., Roest, H. I. J4. and Thiéry R. Scientific report. Development of harmonised schemes for the monitoring and repor-ting of Q fever in animals in the European Union. 2010. http://www.efsa.europa.eu/en/scdocs/doc/48e.pdf
11. Simon More, Jan Arend Stegeman, Joerg Hartung, Annie Rodolakis, Hendrik-Jan Roest, Piet Ve-llema Richard Thiery, Heinrich Neubauer, Wim van der Hoek, Howard Needham, Katharina Staerk, Bart Goossens, Jane Richardson, Ana Afonso,Milen Georgiev. Scientific opinion on Q fever, EFSA panel on Animal Health and Welfare, European Food Safety Authority (EFSA) Error: No se encuen-tra la fuente de referencia. http://www.efsa.europa.eu/en/scdocs/doc/1595.pdf

29
ponencias
FIEBRE Q EN HOLANDA
Douwe Bakker and Hendrik-Jan RoestCentral Veterinary Institute of Wageningen UR
Lelystad, The [email protected]
Introducción
La Fiebre Q es una zoonosis causada por Coxiella burnetii, una bacteria intracelular Gram negativa
prevalente en todo el mundo, excepto en Nueva Zelanda. Los animales domésticos se consideran el
principal reservorio de Fiebre Q para el hombre, aunque muchos otros animales pueden infectarse
con C. burnetii. La transmisión de los animales al hombre ocurre principalmente a través de aerosoles.
En cabras y ovejas el principal síntoma de la Fiebre Q es el aborto. En vacas es fertilidad reducida y
metritis, aunque el aborto también puede ocurrir. Otras especies animales no muestran sintomatología
clínica tras la infección. En las personas la infección es asintomática en el 60% de los casos. En una
infección aguda la Fiebre Q produce síntomas similares a una gripe, con fiebre, neumonía atípica o
hepatitis. La infección puede progresar a una infección crónica que frecuentemente conduce a en-
docarditis en el 1 al 5% de los casos. Debido a su naturaleza zoonótica y a la baja dosis de infección
requerida, C. burnetii es un organismo que se debe manejar bajo condiciones de bioseguridad de
nivel 3 (2000/54/EU).
Historia de la Fiebre Q en HolandaEn Holanda los primeros tres casos de Fiebre Q en personas se registraron en 1956, aunque no está claro si estos casos eran autóctonos. La Fiebre Q se convirtió en Enfermedad de Declaración Obli-gatoria en 1975 y en los siguientes años fue evidente que estaba presente en el país, principalmente como enfermedad profesional. Entre 1975 y 2006 se han registrado una media de 16 casos humanos cada año. En los años 80 algunos estudios sobre Fiebre Q en animales domésticos evidenciaron que esta infección estaba también presente en vacas, ovejas y cabras, así como en perros y gatos.
Inicio del brote de Fiebre Q en HolandaEn 2005 se registraron los primeros abortos por Fiebre Q. En ese año se registraron 2 tormentas de abortos. En 2006 se registraron 6 tormentas de abortos en cabras y 1 en ovejas de leche. Todos los rebaños de cabras con abortos estaban localizados en el Sur de Holanda. En 2007, otros 7 rebaños de cabras sufrieron abortos por Fiebre Q. En este año también se registró el primer caso en humanos con un total de 168 casos registrados. Este brote humano se localizó en el área donde se situaban los rebaños de cabras afectados. Los problemas abortivos en las cabras fueron considerados la causa

30
ponencias
de Fiebre Q en humanos debido al solapamiento geográfico de los casos, la sucesión temporal de los síntomas de Fiebre Q y la ausencia de otras fuentes de infección.
El brote en humanos ocurrido en 2007 se consideró un evento aislado, como los ocurridos en otros países, pero en 2008 el brote continuó con un total de 1.000 casos. El incremento de casos humanos en ese año desencadenó el inicio de medidas para prevenir la eliminación de C. burnetii en cabras y ovejas y prevenir la transmisión a las personas.
Respuesta al brote de Fiebre Q en HolandaLa primera respuesta al brote continuado de Fiebre Q fue la obligatoriedad de notificar porcentajes de abortos superiores al 5% en rebaños de cabras y ovejas. Esta vigilancia pasiva mostró los puntos calientes de excreción de C. burnetii. En 2008, se notificaron brotes en 7 rebaños de cabras y 1 de ovejas y en 2009 en 6 rebaños de cabras. Se ejecutaron medidas adicionales tales como la implemen-tación de manejos adecuados del estiércol, prohibición de visitas, control de vermes y eliminación de placentas de los establos para prevenir la transmisión de C. burnetii al hombre. En 2008, con el fin de prevenir la eliminación de C. burnetii en los rebaños de cabras y ovejas se inició un programa voluntario de vacunación en un área determinada. La vacuna elegida fue una vacuna comercial fase I. Dado que esta vacuna no estaba registrada en Holanda, el Ministerio de Agricultura holandés tuvo que hacer una excepción. En 2009, se extendió el área de vacunación y la vacunación pasó a ser obligatoria, pero debido a la falta de vacuna no todos los rebaños de cabras y ovejas pudieron ser vacunados a tiempo, es decir, antes de la época de cubrición. En 2009, se inició un sistema de vigilancia activa vía muestreo en tanque de leche. Como consecuencia el número de granjas positivas se incrementó a unas 90.
En 2009, el número de casos humanos se incrementó a 2.355, y el gobierno quiso tomar todas las medidas posibles para detener el incremento de casos humanos en 2010. Se sacrificaron todas las cabras y ovejas gestantes procedentes de rebaños positivos a Fiebre Q y se prohibió la cubrición de hembras vacías en esos rebaños.
Situación actual de la Fiebre Q en HolandaHasta la fecha, se han registrado 357 casos de Fiebre Q en humanos en 2010 y no se ha podido observar ningún pico como ocurrió en años anteriores. Esto supone una significativa reducción com-parado con el año 2009. Noventa y tres rebaños de cabras y ovejas son positivas a partir de muestras de tanque de leche.
¿Cuál podría ser la causa del brote holandés de Fiebre Q?Las cabras y ovejas de leche se consideran la fuente del brote humano de acuerdo con las investiga-ciones epidemiológicas. Esto se confirmó con un estudio sobre los genotipos de C. burneti aislados de cabras y personas. La transmisión de cabras y ovejas a personas podría haberse visto facilitada por el tiempo cálido de las primaveras de 2007, 2008 y 2009. No se ha podido identificar ninguna otra fuente de infección hasta el momento. Pero, ¿por qué C. burnetii causó tormentas de abortos en la población de cabras lecheras? Hasta 2005 la Fiebre Q en cabras no se diagnosticaba y tampoco se observaban tormentas de abortos. Entonces, ¿qué ocurrió? Una de las explicaciones es el gran incremento de la

31
ponencias
población de cabras lecheras. Tras la introducción del sistema de cuotas lecheras en ganado bovino, la industria de leche de cabra en Holanda se expandió. En los últimos 10 años, tras los brotes de Peste Porcina Clásica en 1997 y Fiebre Aftosa en 2001, la población de cabras se ha doblado pasando de cerca de 100.000 en el año 2000 a más de 230.000 en 2009. La mayoría de estos animales estaban localizados en el Sur de Holanda y es en esta población donde C. burnetii pudo diseminarse. Otra ex-plicación adicional sería la emergencia de una cepa más virulenta de C. burnetii que podría diseminar-se fácilmente en la población de cabras, y además infectar a las personas. Las investigaciones sobre la virulencia de la cepa están en marcha. Esperamos que las próximas investigaciones nos permitan dar con la respuesta sobre por qué se inició el brote holandés de Fiebre Q.
Algunos resultados de la investigación en marchaSe han iniciado un gran número de investigaciones tanto en medicina humana como en medicina veterinaria, así como de colaboración entre ambas. Por ejemplo, por primera vez se han aislado cepas de Fiebre Q en Holanda. Hasta 2009 no había ni instalaciones para el cultivo de este organismo ni se disponía de la experiencia necesaria. En colaboración con el Instituto de Microbiología de Bundes-wehr, se desarrolló un método de cultivo de C. burnetii en el Instituto Central de Veterinaria de Wage-ningen. El cultivo se lleva a cabo en instalaciones de bioseguridad de nivel 3 sobre células de Mono Verde Búfalo. El aislamiento a partir de material placentario es relativamente fácil de realizar. Las cepas se utilizan ahora para el desarrollo de experimentos sobre patogénesis de C. burnetii en cabras y hay planeados nuevos estudios sobre secuencias y resistencias.
Las investigaciones epidemiológicas han mostrado que vivir a menos de 2 kilómetros de una granja de cabras se asoció con un mayor riesgo de Fiebre Q comparado con vivir a más de 5 kilómetros de una granja caprina. El genotipado de C. burnetii a partir de muestras clínicas de cabras reveló que existe un genotipo predominante en todas las granjas caprinas del Sur del país. Se espera que este sea el genotipo que ha infectado a las personas, pero todavía no existen datos suficientemente fiables para confirmarlo.
ConclusiónLos problemas con la Fiebre Q sorprendieron a Holanda a pesar de que esta enfermedad está presen-te desde hace años. Para combatirla fue necesaria una estrecha colaboración entre medicina humana y veterinaria tanto a nivel de investigación como de legislación. Todavía no está claro por qué la Fiebre Q llegó a ser un problema y esperamos que futuras investigaciones den una respuesta.
Referencias1. Angelakis E, Raoult D. Q Fever. Vet Microbiol. 2010 Jan 27;140(3-4):297-309.
2. Arricau-Bouvery N, Rodolakis A. Is Q fever an emerging or re-emerging zoonosis? Vet Res. 2005 May-Jun;36(3):327-49.
3. Arricau-Bouvery N, Souriau A, Bodier C, Dufour P, Rousset E, Rodolakis A. Effect of vaccina-tion with phase I and phase II Coxiella burnetii vaccines in pregnant goats. Vaccine. 2005 Aug 15;23(35):4392-402.

32
ponencias
4. Karagiannis I, Schimmer B, Van Lier A, Timen A, Schneeberger P, Van Rotterdam B, et al. In-vestigation of a Q fever outbreak in a rural area of The Netherlands. Epidemiol Infect. 2009 Sep;137(9):1283-94.
5. Klaassen CH, Nabuurs-Franssen MH, Tilburg JJ, Hamans MA, Horrevorts AM. Multigenotype Q fever outbreak, the Netherlands. Emerging infectious diseases. 2009 Apr;15(4):613-4.
6. Raoult D, Marrie TJ, Mege JL. Natural history and pathophysiology of Q fever. Lancet Infect Dis. 2005 Apr;5(4):219-26.
7. Roest HIJ, Tilburg JJHC, Van der Hoek W, Vellema P, Van Zijderveld FG, Klaassen CH, et al. The Q fever epidemic in the Netherlands: history, onset, response and reflection. Epidemiology and Infection. 2010; Accepted.
8. Roest HIJ, Ruuls RC, Buijs R, Spierenburg M, Vellema P, Dercksen D, et al. Geografical distribution of MLVA types of Coxiella burnetii isolates from goats in the Netherlands. Medical Corps Internatio-nal Forum. [abstract]. 2009;4(supplement to MCIF 4/2009):13.
9. Schimmer B, Ter Schegget R, Wegdam M, Zuchner L, de Bruin A, Schneeberger PM, et al. The use of a geographic information system to identify a dairy goat farm as the most likely source of an urban Q-fever outbreak. BMC infectious diseases. 2010;10:69.
10. Sidi-Boumedine, K1., Rousset, E1., Henning, K2., Ziller, M2., Niemczuck, K3., Roest, H. I. J4. and Thiéry R. Scientific report. Development of harmonised schemes for the monitoring and re-porting of Q fever in animals in the European Union. 2010. http://www.efsa.europa.eu/en/scdocs/doc/48e.pdf
11. Simon More, Jan Arend Stegeman, Joerg Hartung, Annie Rodolakis, Hendrik-Jan Roest, Piet Ve-llema Richard Thiery, Heinrich Neubauer, Wim van der Hoek, Howard Needham, Katharina Staerk, Bart Goossens, Jane Richardson, Ana Afonso,Milen Georgiev. Scientific opinion on Q fever, EFSA panel on Animal Health and Welfare, European Food Safety Authority (EFSA) Error: No se encuen-tra la fuente de referencia. http://www.efsa.europa.eu/en/scdocs/doc/1595.pdf

33
ponencias
LA PESTE DE LOS PEQUEÑOS RUMIANTES Y LA FIEBRE/ENCEFALITIS POR VIRUS WEST NILE: DIAGNOSTICO ACTUAL Y PERSPECTIVAS DE
FUTURO
Miguel Ángel Jiménez ClaveroCentro de Investigación en Sanidad Animal (CISA)-INIA, Ctra Algete-El Casar s/n, 28130, Valdeolmos (Madrid).
ResumenLa Oficina Internacional de Sanidad Animal (OIE) define “enfermedad emergente” como una nueva infección que resulta de la evolución o cambio de un agente patogénico ya existente, una infección conocida que se disemina a una nueva área geográfica o población, o una enfermedad o agente pa-togénico previamente no reconocido, diagnosticado por primera vez. Las enfermedades emergentes, además de una constante amenaza para sanidad animal y para la salud pública (pues muchas de ellas tienen carácter zoonótico), representan un formidable reto para el diagnóstico de laboratorio. Ello se debe a que los patógenos emergentes son un fenómeno en esencia inesperado, de difícil previsión. En determinados casos este diagnóstico se hace especialmente difícil, pues el patógeno emergente es un agente nuevo, no descrito, y para el cual no se dispone de técnicas para su detección e iden-tificación. El laboratorio de diagnóstico especializado en enfermedades emergentes y transfronterizas debe convivir con cierta incertidumbre. Para mitigarla, debe conocer la situación epidemiológica en re-lación con las patologías emergentes más acuciantes en su entorno, y actualizar frecuentemente esta información. Pero debe mirar también más allá, porque en un mundo globalizado los patógenos tienen grandes oportunidades para invadir un nuevo territorio. Dos casos recientes servirán como ejemplo de lo que supone una incursión de una enfermedad emergente en nuestro entorno: la peste de los pequeños rumiantes, que desató una grave epizootia en Marruecos en 2008, representando un inmi-nente riesgo de entrada en España, que afortunadamente no se ha llegado a materializar, y la fiebre/encefalitis por virus West Nile (o Nilo Occidental) que recientemente ha causado –por vez primera en nuestro país- brotes de enfermedad en caballos y humanos en Andalucía. De igual modo, estos dos ejemplos servirán para revisar las actuaciones que es necesario implementar en esta materia en cuan-to al diagnóstico de laboratorio, y como se están abordando desde nuestro laboratorio en el Centro de Investigación en Sanidad Animal (CISA), el cual cuenta con una larga experiencia en investigación y desarrollo de métodos de diagnóstico de enfermedades emergentes y transfronterizas.


35
ponencias
DIAGNÓSTICO DE LA BESNOITIOSIS BOVINA
Juan Antonio Castillo*, José Miguel Marcén, Adriana EstebanDpto de Patología Animal. Facultad de Veterinaria. Universidad de Zaragoza*.
IntroducciónLa Besnoitiosis bovina (anasarca, elefantiasis o elephant skin disease) es una enfermedad parasitaria producida por un protozoo esporozoario, Besnoitia besnoiti. Afecta principalmente a los bóvidos, aunque se ha descrito en otras especies de rumiantes (ovino, caprino, ciervos, antílopes, corzo etc). Desde el punto de vista clínico se caracteriza por una fase febril inicial seguida de una fase crónica caracterizada por lesiones cutáneas, con engrosamiento, dermatosis, edemas y un estado de ema-ciación marcado. Los machos normalmente desarrollan infertilidad permanente.
Su primera descripción se hizo en el Sur Francia (Besnoit y Robin, 1912) y desde entonces se ha ido conociendo su extensión por todo el mundo. En nuestro país la primera cita se realizó en el País Vasco en 1990 y desde entonces se ha ido extendiendo por amplias zonas geográficas de la mitad norte de la península. Si a esto añadimos su alta morbilidad (altos descensos productivos) podemos decir que se trata de una enfermedad emergente de primera magnitud en el ganado bovino en España y en Europa.
Patogenia, clínica y lesionesLos animales comienzan a desarrollar signos clínicos aproximadamente a las dos semanas post-infección (p.i), sin embargo hay animales en los cuales el periodo de incubación se puede alargar hasta los dos meses.
La aparición de la enfermedad en Europa y Sudáfrica es estacional y tiene una clara relación con los periodos de actividad trófica de los vectores potenciales. Así en Francia aparece fundamentalmente en los meses de verano, julio-septiembre, teniendo un máximo en el mes de agosto. Esta situación es muy similar a la que se ha observado en nuestro país.
La besnoitiosis de los bóvidos puede ser leve o grave pero generalmente no mortal.
La forma grave se caracteriza por fiebre, pérdida del apetito, apatía, enrojecimiento de la piel y el hocico, hinchazón de la piel de cara y el cuello, inflamación de los testículos y fotofobia. Al cabo de unos cuantos días, la hinchazón de la piel de las partes superiores del cuerpo se va desplazando a las partes inferiores, como papada, pecho y prepucio (machos). Durante este periodo (fase aguda), el

36
ponencias
cual suele durar aproximadamente 3 semanas, los animales adquieren un mal estado general debido a la progresiva pérdida de apetito. La fase aguda es el reflejo de la multiplicación rápida del parásito durante el estadio de taquizoíto y la invasión del tejido conectivo y posterior inicio de la formación de los quistes intracelulares.
Esta fase aguda es seguida de una fase crónica, la cual se caracteriza por un engrosamiento y en-durecimiento de la piel, con un destacado plegamiento de la misma (signo típico de piel de elefante). En la formas leves de la enfermedad la fase aguda suele pasar desapercibida o ser confundida con otras afecciones.
La fase crónica de la enfermedad tiene su origen en la formación y asentamiento en su localización de-finitiva del parásito formando los quistes en el tejido conjuntivo. Los signos clínicos se van sucediendo conforme a este esquema: pérdida de pelo, alopecias, sobre todo en las zonas cutáneas con mayor carga parasitaria, e incluso desprendimiento de trozos de piel esclerosada. Es frecuente la aparición de grietas profundas en carne viva en los pliegues, que a menudo están infectadas por gérmenes oportunistas o larvas de moscas.
Los ganglios linfáticos suelen estar hipertróficos. Los movimientos son dolorosos y progresivamente mas difíciles para el animal. En casos graves suele aparecer una descarga nasal amarillenta que forma costras y puede llegar a obstruir las fosas nasales.
Los animales permanecen apáticos y la falta de apetito lleva aparejado un proceso de emaciación más a menos acusado. Los machos invariablemente llegan a ser estériles (casi siempre de forma perma-nente) y uno o los dos testículos se tornan secos y disminuye su tamaño.
Varias semanas después de la fase febril inicial se comienzan a ver a simple vista pequeños quistes blanquecinos macroscópicos en la parte interior de los parpados y en otras mucosas explorables. Un gran número de los animales infectados no suelen presentar manifestaciones clínicas y la aparición de estos quistes en los párpados y conjuntiva ocular suele ser el único signo de la enfermedad.
DiagnósticoEl diagnóstico de las enfermedades parasitarias, de forma general, sigue el siguiente orden de desarrollo:
a) Diagnóstico Clínicob) Diagnóstico Etiológicoc) Diagnóstico Anatomo-Patológicod) Diagnóstico Epidemiológico
El diagnóstico en vivo generalmente se basa en la aparición de signos clínicos típicos de la fase cróni-ca, fundamentalmente: engrosamiento de la piel, esclerodermia, aparición de quistes blanquecinos en la conjuntiva ocular y palpebral y también en la mucosa nasal y vulvar (se aprecia bien por palpación suave). (Fig. a y b)

37
ponencias
La fase aguda suele pasar desapercibida ya que puede confundirse con otras muchas patologías
Fig a.- Esclerodermia generalizada Fig b.-Quistes macroscópicos en esclerótica
En casos de sospecha o para confirmar se debe recurrir a la biopsia de piel (sobre todo de la zona superior y laterales del cuello o zona del perineal) y su observación en fresco o por medio de técnicas histológicas. (Fig. c, d, e y f)
Fig c. Corte en fresco con quistes de Besnoitia Fig d.- Corte histológico con quistes de Besnoitia
Fig e. Bradizoitos libres de Besnoitia Fig f. Quiste de Besnoitia

38
ponencias
El diagnóstico serológico da buenos resultados. Entre las técnicas de elección se encuentra la IFI, utilizando como antígeno taquizoítos obtenidos de cultivos celulares (HeLa o MARCK 145) o de jer-bos. El punto de corte seguro se puede establecer en la dilución 1/100 para los positivos. Por debajo de esta dilución es difícil deducir la infección y además puede haber reacciones cruzadas con otros Apicomplexa. La correlación entre el titulo 1/100 y la aparición de signos clínicos o biopsia suele ser muy alta. También están disponibles técnicas inmunoenzimáticas (ELISA) con muy alta sensibilidad y especificidad, como la desarrollada por el grupo SALUVET de la Facultad de Veterinaria de Madrid. La concordancia entre ambas pruebas es muy alta y su elección depende del tipo de uso que se vaya a hacer.
Fig g. IFI positiva de Besnoitia
Por último, se pueden utilizar técnicas de detección de DNA (PCR’s). Están descritas y en marcha dos tipos de metodologías, una cualitativa Nested (anidada) PCR y otra cuantitativa RT-PCR (Real Time) las cuales dan muy buenos resultados en lo que se refiere a sensibilidad y especificidad.
Un buen número de animales presentan infecciones subclínicas o asintomáticas y sin embargo son seropositivos.

39
ponencias
DÍPTEROS VECTORES DE ENFERMEDADES. BIOLOGÍA E IDENTIFICACIÓN
Javier LucientesDepartamento de Patología AnimalFacultad de Veterinaria de Zaragoza
Vector, según la OMS, es aquel artrópodo hematófago responsable de la transmisión activa de un agente patógeno. Son muchos los artrópodos que han colonizado el cuerpo de otros animales, sobre todo superiores, para hacer de él su hábitat permanente o su fuente de alimentación. Y sin lugar a dudas la sangre es un alimento que aporta muchos nutrientes de calidad llegando a ser imprescindible para la supervivencia de numerosas especies.
La Clase Insecta o Hexapoda posee abundantes ejemplos de estos animales que se han adaptado a la vida parásita, y dentro de ellos es tal vez el Orden Díptera uno de los más importantes por las repercusiones que tienen los hábitos hematófagos de algunos géneros que les ha adaptado para poder transmitir enfermedades tan importantes como la Malaria, el Dengue, las Trypanosomiasis, las Leishmaniosis, etc.
A nivel de España son nueve las Familias de dípteros en las que encontramos especies que se ali-mentan de sangre: Culicidae, Psychodidae (Phlebotominae), Simuliidae, Ceratopogonidae, Tabani-dae, Muscidae, Hippoboscidae, Nycteribiidae y Streblidae. Las seis primeras son las más conocidas porque son más abundantes y afectan sobre todo al hombre y a los animales domésticos. La Familia Hippoboscidae incluye las denominadas moscas de las plumas pero también tiene especies que parasitan a mamíferos como el melófago de la oveja o la mosca de las caballerías. Las Familias Nyc-teribiidae y Streblidae son parásitas de murciélagos.
No todos los géneros de las Familias Muscidae y Tabanidae tienen costumbres hematófagas quedan-do relegados, sobre todo en los Muscidos, a unos pocos géneros como Stomoxys o mosca de los establos. En las demás Familias en casi todas las especies las hembras tienen hábitos hematófagos que son necesarios para la maduración de los ovocitos y la puesta de huevos.
Esta necesidad de acceder a la sangre de sus hospedadores las ha obligado a adaptar su aparatos bucales modificándolos para esta función. Modificaciones que nos van a ayudar en su diferenciación de Familias vecinas con otros tipos de alimentación.
Vamos a profundizar en las especies de tamaño más pequeño que son además las más abundantes, y que resultan más difíciles de identificar. Los Culicidae o Mosquitos verdaderos poseen en estado adul-

40
ponencias
to un aparato bucal muy diferenciado. Las piezas bucales, o proboscis, son alargadas conformando un labio y un estilete que les permite perforar la piel y acceder directamente a los vasos sanguíneos para succionar sangre. Son insectos denominados solenófagos. Su tamaño es en general entre 7 y 15 mm de longitud. El cuerpo es esbelto con largas patas y estrechas alas dobladas en tijera sobre el dorso en reposo. Tienen el cuerpo y las alas recubiertos de escamas y sedas importantes para la clasificación a nivel de especie.
Esta Familia es muy numerosa y presentan costumbres tróficas muy variadas. La gran mayoría son crepusculares y nocturnos, pero muchas especies, sobre todo del género Aedes pican incluso por el día.
Las larvas y las pupas se encuentran siempre en aguas estancadas o con ligera corriente. Cada es-pecie suele tener habitats de cría muy característicos que pueden ir desde aguas semipermanentes a permanentes, aguas limpias, contaminadas, muchas especies admiten aguas salitrosas y algunas incluso muy saladas. Los podemos encontrar en hábitats naturales como lagunas, charcas, orillas de ríos, huecos con agua en árboles, etc. O artificiales como depósitos, canales, cultivos, neumáticos, recipientes de vidrio o cerámica, etc. Las larvas se diferencian muy fácilmente pues son alargadas con aspecto vermiforme y están continuamente nadando, subiendo y bajando desde la superficie donde tienen que ascender a respirar aire. Tienen los últimos segmentos muy modificados presentando un sifón respiratorio característico. Se alimentan de microorganismos y restos orgánicos cogiéndolos di-rectamente con la boca que presenta un aparato bucal de tipo masticador. Después de cuatro mudas se transforman en pupa, también acuáticas con una forma característica pues poseen un cefalotórax muy desarrollado y un abdomen aplanado y adaptado para nadar. En esta fase no se alimentan pero son muy móviles para huir de posibles predadores.
Los Simuliidae es otra familia muy importante sobre todo en nuestro país por las molestias que origi-nan. Los adultos tienen un cuerpo fuerte, recuerdan a moscas muy pequeñas y como tienen en gene-ral colores oscuros reciben el nombre de Moscas negras. Su tamaño oscila entre 3 y 5 mm. Las patas son cortas, las alas cruzadas en el dorso y un aparato bucal corto pues está compuesto por piezas pequeñas, pero fuertes y con los bordes dentados o afilados lo que les permite cortar la piel y el tejido subcutáneo formando pequeñas heridas que acumulan sangre de las que se alimentan directamente. Son insectos telmófagos. Su picadura es muy dolorosa y su saliva origina reacciones alérgicas muy severas en personas sensibles. Su actividad es únicamente diurna no volando por la noche.
Las larvas son muy características pues las vamos a encontrar siempre en aguas corrientes y fijadas por su parte posterior a un sustrato como piedras, vegetales (algas, carrizos,…), plásticos, etc. Estas larvas tienen un aspecto vermiforme con una longitud la última fase larvaria de entre 10 y 12 mm. con el final del abdomen más voluminoso. Resalta de su morfología la presencia de dos premandíbulas muy desarrolladas en forma de abanico, que le sirven para filtrar el agua y retener en ellas las partículas alimenticias. La pupa también es muy característica y comparte el mismo hábitat que las larvas. Las larvas ya maduras tejen, con una seda que desprenden, una especie de capullo en forma de zapatilla

41
ponencias
en sentido contrario a la corriente, donde se introduce y realiza la muda a pupa. Esta no se mueve permanece dentro del capullo protegida de posibles desplazamientos. Tienen unos filamentos respira-torios que sobresalen del tórax de la pupa que son muy característicos y de importancia taxonómica.
Los Ceratopogónidos son los más pequeños de este grupo pues miden entre algo menos de 2 y hasta 4 mm de longitud. En castellano se denominan Jejenes y sobre todo en algunas zonas llegan a ser una plaga muy molesta por sus picaduras. Tienen unas patas muy pequeñas, las alas las pliegan sobre el dorso en reposo y cuando se posan en la piel para picar son realmente difíciles de ver. Tienen un apa-rato bucal de tipo cortador chupador por lo que las picaduras pueden ser dolorosas. La mayoría de las especies tienen las alas con manchas distribuidas de forma irregular siendo este patrón de dibujo un elemento de gran valor taxonómico. Son crepusculares y nocturnos.
Los hábitats larvarios son muy diversos dependiendo del género incluso de la especie. Necesita sus-tratos con materia orgánica y humedad, y éstos pueden ser desde limos de zonas encharcadas, barros o materia orgánica en descomposición. Las larvas de aspecto vermiforme de hasta 5 mm de longitud, son muy agresivas y tienen un amplio espectro de alimentación llegando a ser caníbales.
El último grupo que vamos a ver es el de los Flebotomos. Son dípteros de la Familia Psychodidae y subfamilia Phlebotominae. Son insectos muy característicos en estado adulto. Miden entre 3 y 5 mm de longitud. Son de color marrón claro a beige y tienen patas muy largas. La cabeza es de implanta-ción ligeramente inferior lo que les proporciona un aspecto giboso. Tienen un aparato bucal de tipo cortador chupador con estiletes cortos y fuertes con los bordes dentados. Son telmófagos.
Lo más característico de este grupo es que tiene largas alas de forma lanceolada, con nervios para-lelos que cuando están en reposo las dejan abiertas en forma de “V” por lo que son muy fáciles de identificar. Son insectos nocturnos y crepusculares.
Las larvas de aspecto vermiforme también son terrestres. No se encuentran en zonas encharcadas aunque requieren un cierto grado de humedad. Tienen un tamaño de hasta 6 o 7 mm y se alimentan de materia orgánica. Se localizan en una amplia variedad de hábitats siempre que tengan alimento disponible como restos vegetales, heces de herbívoros, etc.


43
ponencias
EL DIAGNOSTICO DE ENFERMEDADES E INFECCIONES EMERGENTES EN EL CERDO
Joaquim Segalés
Centre de Recerca en Sanitat Animal (CReSA), Fundació UAB-IRTA, Esfera UAB and Departament de Sanitat i Anatomia Animals, Edifici V, Universitat Autònoma de Barcelona, 08193 Bellaterra (Barce-
lona); E-mail: [email protected]
IntroducciónEstudiando la literatura existente, uno se da cuenta de que existen muchas definiciones potenciales sobre lo que son las enfermedades infecciosas emergentes. En una de estas definiciones se conside-ran aquellas cuya incidencia ha incrementado en los últimos 20 años y que amenazan con continuar incrementando a corto plazo. La historia natural de las nuevas infecciones es relativamente simple desde un punto de vista genérico (Figura 1). No obstante, y especialmente en un ámbito de enferme-dad clínica evidente, se asume que convivir con un escenario enzoótico puede ser peor que la erradi-cación de la misma; incluyendo la amenaza de lo que significa el riesgo de re-infección. Esta situación es especialmente importante cuando se está lidiando con infecciones y enfermedades que causan una pérdida de producción o competitividad, dado que ello supondría unas desventajas económicas muy significativas, no solo para productores sino también para países o regiones.
Figura 1. Historia natural de las nuevas infecciones/enfermedades.
Existencia previa en el ambiente
(¿en la misma especie?) (¿en otras especies?)
Aparición súbita (usualmente epizoótica)
Extensión a la población susceptible
Fase enzoótica
Vacunación/Tratamiento Erradicación
Control (mantenimiento
enzoótico)
Eliminación Población susceptible

44
ponencias
Enfermedades/infecciones emergentes en cerdosDentro de la terminología “enfermedades infecciosas emergentes” existen distintas sub-clasificaciones potenciales (Fauci et al., 2005):
• enfermedades emergentes nuevas (aquellas que no han sido reconocidas anteriormente).• enfermedades re-emergentes o resurgentes (aquellas que pueden haber sido reconocidas desde
hace décadas o siglos, pero que han vuelto en un formato clínico-epidemiológico distinto o en una localización distinta).
• enfermedades emergentes deliberadas (aquellas que han sido introducidas intencionadamente).
En condiciones normales, y hablando de enfermedades del cerdo, la última categoría no debería tener una consideración seria. No obstante, entre la primera categoría de enfermedades emergentes si tendríamos algunas enfermedades significativas, incluyendo el síndrome respiratorio y reproductivos porcino (PRRS), el cuál emergió sobre finales de los años 80, o el síndrome multisistémico de adelga-zamiento post-destete o circovirosis porcina, que apareció a finales de los 90. Finalmente, dentro de lo que consideramos enfermedades re-emergentes, deberíamos incluir a las pestes porcinas (clásica y Africana) o hasta enfermedades como la de Glässer.
Las enfermedades citadas en el párrafo anterior se consideran de gran preocupación por parte de los productores de cerdos, pero también existe un número muy importante de infecciones, básicamente víricas, que se consideran emergentes en la especie porcina, y que mayoritariamente provocan infec-ciones sub-clínicas. Entre ellas se destaca:
Virus de la Hepatitis E (HEV) – familia HepeviridaeTorque teno sus virus (TTSuV) – familia AnelloviridaeTorovirus porcino (PoTV) – familia CoronaviridaeCalicivirus entéricos porcinos (norovirus y sapovirus) – familia CaliciviridaeBocavirus porcino – familia ParvoviridaeTeschovirus porcinos – familia PicornaviridaeSapelovirus – familia PicornaviridaeAdenovirus porcino – familia Adenoviridae¿Otros?
El momento preciso de introducción de estas infecciones emergentes en la cabaña porcina es habi-tualmente desconocido, a no ser que esta introducción represente una enfermedad clínico-patológi-camente distinta a lo previamente existente y sea causante de pérdidas económicas importantes. De hecho, los estudios retrospectivos realizados hasta el momento han mostrado que la mayor parte de enfermedades o infecciones consideradas emergentes en el fondo no eran tales, sino que el agente infeccioso causal se ha mantenido como desconocido por un período de tiempo más o menos largo. Un buen ejemplo sería el de circovirus porcino tipo 2 (PCV2), el agente etiológico esencial de la cir-covirosis porcina. La asociación de este virus con la enfermedad se establece en 1998 (Hamel et al., 1998), pero los estudios retrospectivos realizados indican la existencia de infección con este virus ya

45
ponencias
por 1962 en Europa (Jacobsen et al., 2008). Es más, la enfermedad asociada a este virus, la circovi-rosis porcina, también pasó desapercibida durante varios años, ya que estos estudios retrospectivos muestran casos más o menos esporádicos que se correspondían de forma inequívoca a los criterios diagnósticos de la enfermedad desde 1985 (Jacobsen et al., 2008). De forma similar, la infección por el HEV, descrito inicialmente en 1997 (Meng et al., 1997), se ha detectado posteriormente de forma retrospectiva al menos desde 1985 (Casas et al., 2009). Y otro ejemplo más, el de TTSuV, que se describió inicialmente en 1999 (Leary et al., 1999) y que retrospectivamente ha sido detectado en 1985 (Segalés et al., 2009). Un factor común en la mayoría de estas infecciones es que la primera evidencia de infección coincide en general con el primer año de estudio retrospectivo, lo que significa que estos agentes se encontraban ya diseminados en la cabaña porcina mundial con anterioridad. De hecho, la falta de sueros de cerdo realmente antiguos impide conocer con precisión la fecha de introducción de estos agentes víricos en la cabaña porcina. Quizás la excepción a esta regla sería el virus del PRRS. El PRRS se describe como una enfermedad significativa en el cerdo a mediados de los 80 en Norte-América y a finales de 1990 en Europa (Zimmerman et al., 2006). Desde el punto de vista retrospectivo, sí se podría actualmente tener una idea de la introducción del virus del PRRS en el cerdo, dado que los primeros sueros positivos a la detección de anticuerpos fueron detectados en 1979 en Norte-América (Carman et al., 1995) y 1988 en Europa (Ohlinger, 1992), fechas relativamente cercanas a la emergencia del PRRS como una enfermedad clínica importante. Esta situación con el virus del PRRS también ha facilitado un número importante de especulaciones sobre el origen del virus, siendo probablemente la más aceptada aquella que lo relaciona con un virus mutante del virus de la lactato-deshidrogenasa elevada del ratón (arterivirus) que presumiblemente realizó un salto inter-especie hacia el jabalí y sub-siguientemente al cerdo doméstico (Plagemann et al., 2003). Por tanto, vistos los datos retrospectivos aportados por la mayoría de enfermedades/infecciones que conside-ramos actualmente como emergentes, esta última terminología es, probablemente, una simplificación no demasiado realista. Al fin y al cabo, y ello aplicaría a la mayoría de enfermedades e infecciones de todas las especies a nivel mundial: “una determinada infección o enfermedad existe en un granja, región o país dependiendo del entusiasmo con que se busque” (citado por el Dr. C. Pijoan, University of Minnesota, Estados Unidos, 1998).
¿Cómo diagnosticar estas enfermedades/infecciones emergentes?Desde el punto de vista de trabajo diagnóstico, actualmente parece que si no se trabaja con técnicas de alta tecnología como PCR cuantitativa o microarrays, uno está anticuado. Es cierto que estas nuevas tecnologías han supuesto una revolución en el trabajo diagnóstico y de investigación a nivel mundial. No obstante, técnicas como la histopatología por ejemplo, continúan siendo metodologías extremadamente útiles que deben ser utilizadas e interpretadas en el contexto adecuado. De hecho, la mayor parte de enfermedades del cerdo que causan lesiones macro y/o microscópicas evidentes tienden a ser de origen infeccioso, con lo que las técnicas de biología molecular han mejorado aspec-tos importantes como la sensibilidad y especificidad de cara a la detección de agentes causales, así como la caracterización de los mismos.

46
ponencias
Es interesante remarcar que los científicos que inicialmente trabajaron y describieron dos de las enfer-medades emergentes más significativas del cerdo, PRRS y circovirosis porcina, eran patólogos. Las primeras descripciones del PRRS como enfermedad nueva incluyeron la descripción de una lesión microscópica básica consistente: neumonía intersticial (Zimmerman et al., 2006), mientras que la pre-sentación como nueva entidad de la circovirosis porcina incluía la depleción linfocitaria conjuntamente con inflamación granulomatosa de los órganos linfoides (Clark, 1997). Por tanto, el papel de metodo-logías clásicas como la anatomía patológica se puede considerar aún como una pieza fundamental en el diagnóstico y caracterización de enfermedades emergentes, sea en cerdos u otras especies. No obstante, estas técnicas clásicas difícilmente permiten la descripción de nuevas entidades que, en general, se presentan como infecciones sub-clínicas (tal como HEV, TTSuV y tantas otras). Ello también hace que nos preguntemos cuán importantes sean estas infecciones que no causan enfer-medad clínica. Sería un error pensar que si estas nuevas infecciones se manifiestan aparentemente de forma exclusiva como sub-clínicas, entonces no merecen nuestra atención. Cabe destacar que la participación de estos nuevos agentes puede potencialmente tener una actividad sinergística con otros patógenos en el contexto de enfermedades multifactoriales. PCV2 ya fue un ejemplo de ello a fi-nales de los 90 (Segalés et al., 2005), y otros agentes como retrovirus porcinos (endógenos/exógenos) (Tucker et al., 2006) o TTSuV2 (Aramouni et al., 2010) se han asociado a situaciones de baja sanidad y enfermedades asociadas a PCV2, respectivamente.
ReferenciasAramouni, M., Segalés, J., Sibila, M., Martin-Valls, G.E., Nieto, D., Kekarainen, T., 2010. Torque teno sus virus 1 and 2 viral loads in 1 postweaning multisystemic wasting syndrome (PMWS) and porcine dermatitis and nephropathy syndrome (PDNS) affected pigs. Enviado para publicación.
Carman, S., Sanford, S.E., Dea, S., 1995. Assessment of seropositivity to porcine reproductive and respiratory syndrome (PRRS) virus in swine herds in Ontario -1978 to 1982. Can. Vet. J. 36, 776-777.
Casas, M., Pujols, J., Rosell, R., de Deus, N., Peralta, B., Pina, S., Casal, J., Martín, M., 2009. Re-trospective serological study on hepatitis E infection in pigs from 1985 to 1997 in Spain. Vet Microbiol. 135, 248-252.
Clark, E.G., 1997. Post-weaning multisystemic wasting syndrome. Proc. Annu Meet Am Assoc Swine Pract, pp.499-501.
Fauci, A.S., 2005. Emerging and reemerging diseases: the perpetual challenge. Acad. Med. 80: 1079-1085.
Hamel, A.L., Lin, LL., Nayar, G.P.S., 1998. Nucleotide sequence of porcine circovirus associated with postweaning multisystemic wasting syndrome in pigs. J. Virol. 72, 5262-5267.

47
ponencias
Jacobsen, B., Krueger, L., Seeliger, F., Bruegmann, M., Segalés, J., Baumgaertner, W., 2009. Retros-pective study on the occurrence of porcine circovirus 2 infection and associated entities in Northern Germany. Vet Microbiol. 138, 27-33.
Leary, T.P., Erker, J.C., Chalmers, M.L., Desai, S.M., Mushahwar, I.K., 1999. Improved detection sys-tems for TT virus reveal high prevalence in humans, non-human primates and farm animals. J. Gen. Virol. 80, 2115-2120.
Meng, X.J., Purcell, R,H., Halbur, P.G., Lehman, J.R., Webb, D.M., Tsareva, T.S., Haynes, J.S., Thac-ker, B.J., Emerson, S.U., 1997. A novel virus in swine is closely related to the human hepatitis E virus. Proc. Natl. Acad. Sci. USA 94, 9860-9865.
Ohlinger, V., 1992. PRRS/Blue ear disease/SIRS monthly newsletter. Meredith M, Ed. Pig Disease Information Centre, Cambridge.
Plagemann, P.G., 2003. Porcine reproductive and respiratory syndrome virus: origin hypothesis. Emerg Infect Dis. 9(8):903-908.
Segalés, J., Allan, G.M., Domingo, M., 2005. Porcine circovirus diseases. Anim. Health Res. Rev. 6: 119-142.
Segalés, J., Martínez-Guinó, L., Cortey, M., Navarro, N., Huerta, E., Sibila, M., Pujols, J., Kekarainen, T., 2009. Retrospective study on swine Torque teno virus genogroups 1 and 2 infection from 1985 to 2005 in Spain. Vet Microbiol. 134(3-4): 199-207
Tucker, A.W., Mellencamp, M.M., Donadeu, M., Scobie, L., 2006. Retroviremia in commercial pigs and its preliminary association with poor health. J Clin Microbiol. 44(10): 3846-3847.
Zimmerman, J., Benfield, D.A., Murtaugh, M.P., Osorio, F., Stevenson, G.W., Torremorell, M., 2006. Porcine reproductive and respiratory syndrome virus (porcine arterivirus). In: Diseases of Swine. Blac-kwell Sciences. Pp.387-417.


49
ponencias
RECOGIDA DE MUESTRAS PARA ANÁLISIS LABORATORIAL EN UN MEDIO EXÓTICO.
EXPEDICIÓN ESPAÑOLA A LA ANTÁRTIDA 2010.
García Peña FJ1, García Párraga D2, Ortega Mora LM3, Rengifo C3, Pedraza S3
1 LCV de Algete (Madrid). Ministerio de Medio Ambiente, Medio Rural y Marino.2 L´Oceanogràfic. Ciudad de las Artes y las Ciencias de Valencia.
3 Grupo SALUVET. Dpto de Sanidad Animal. Facultad de Veterinaria de Madrid.
IntroducciónLa Antártida ha sido hasta hace relativamente poco tiempo un continente aislado. Geográficamente, el Océano Austral constituye un ecosistema cerrado delimitado por la Convergencia Antártica, actuando ésta como una barrera biológica que en general ha impedido el paso de aves y mamíferos. Este aisla-miento ha hecho por un lado que los animales antárticos pueden haberse visto libres de enfermedades comunes en otras zonas y por otro, que se desconozcan las enfermedades propias de estas poblacio-nes. En los últimos años, el interés por la Antártida se ha visto reflejado en un incremento tanto de las investigaciones que se llevan a cabo como, más marcadamente aún, en el turismo en la región (Figura 1). Este incremento de la actividad humana puede estar influyendo directamente en las poblaciones animales que habitan en ella tanto a nivel de la dinámica de poblaciones como de introducción de enfermedades y riesgo de diseminación tanto de las enfermedades endémicas como exóticas.
FIGURA 1: Evolución del turismo en la Antártida 1992-2007.

50
ponencias
En la Campaña Antártica Española 2009-2010 se desarrollaron 22 proyectos de investigación que implicaron aproximadamente a 112 investigadores y 32 instituciones. La mayor parte de ellos están relacionados con estudios de geofísica, fisiología y biología, siendo nuestro proyecto “Estado sanitario de fócidos y otáridos en Peninsula Antartica: impacto de la actividad humana e influencia del gradiente latitudinal” el primero plenamente veterinario hasta donde nosotros conocemos.
Este proyecto continúa proyectos anteriores en los que se inició una línea de investigación sobre el estado sanitario de las focas y leones marinos antárticos, ya que estos animales constituyen unos excelentes bioindicadores del impacto que puede estar provocando la actividad humana en la Antár-tida (Figura 2). En concreto, se pretende evaluar el estado sanitario de los mismos en Isla Decepción y otras zonas de las Islas Shetland del Sur y Península Antártica. La determinación de la presencia y frecuencia de ciertos patógenos permitirá comprobar su estado sanitario y sentará las bases para el seguimiento o monitorización del mismo a largo plazo. Asimismo, el estudio comparativo de pobla-ciones de animales procedentes de zonas con distinto nivel de actividad humana permitirá evaluar el potencial impacto de ésta tanto a nivel de dinámica de poblaciones como sanitario.
FIGURA 2: actividad turística en Fumarolas (Isla Decepción).
Especies de elecciónLos mamíferos marinos son considerados buenos bioindicadores de los cambios ambientales a medio y largo plazo debido a que muchas especies son longevas y se encuentran en la cúspide de la cadena trófica. Según se reconoce en el informe final del Comité para la Protección del Medio Ambiente (XXVI Reunión Consultiva del Tratado Antártico) reunido en Madrid del 9 al 20 de junio del 2003, es escasa la información que se tiene sobre las enfermedades propias de los animales antárticos y además: “...hay una gran necesidad de realizar estudios complementarios que permitan conocer la situación sanitaria de otras especies y poblaciones de animales que habitan el ecosistema antártico, con la finalidad de contribuir al conocimiento científico y orientar la creación de medidas de administración de los recur-sos vivos marinos de la Antártida”, insistiendo además en que “...es fundamental destinar esfuerzos

51
ponencias
para estimar la situación sanitaria de las poblaciones naturales de vertebrados superiores de la An-tártida, mediante su seguimiento periódico, como también, dimensionar su efecto sobre la dinámica poblacional de las especies afectadas y realizar estudios epidemiológicos de las enfermedades que los afectan. Tales estudios, podrán servir como un indicador complementario de la acción humana en ese ecosistema y mejorar las medidas de prevención de la potencial contaminación biológica desde o hacia la Antártida (propias de las actividades domésticas del hombre en la Antártida),...”
Entre los mamíferos marinos presentes en el continente antártico y su entorno destacan el lobo fino antártico (Arctocephalus gazella), la foca de Weddell (Leptonychotes weddelli) y el elefante marino austral (Mirunga leonina) (Figura 3) de los cuales hay escasa información a pesar de constituir consti-tuyen unos excelentes bioindicadores del impacto negativo que puede estar provocando la actividad humana en la Antártida. Otros mamíferos de interés son la foca cangrejera (Lobodon carcinophagus) y la foca leopardo (Hydrurga leptonyx), de los cuales se han recogido un escaso número de muestras de heces en anteriores campañas, pero son especies no gregarias y que prefieren permanecer sobre bloques de hielo, lo cual impide su captura.
Lobo marino antártico Arctocephalus gazella
Elefante marino del sur Mirounga leonina
Foca de Weddell Leptonychotes weddelli
FIGURA 3: especies estudiadas para lavalora-ción de estado sanitario e impacto humano.

52
ponencias
Preparativos precampaña
La preparación de la campaña antártica es una tarea ardua y que comienza meses antes del traslado
de los investigadores a la Antártida.
En primer lugar, el personal seleccionado debe pasar un reconocimiento médico previo en el mes de
julio o agosto.
Todo el material que se va a utilizar en la campaña incluyendo ropa, material de captura, material para
reanimación y anestesia, hisopos… debe ser listado y enviarse a Cartagena a principios de octubre
para su embarque en el buque BIO Las Palmas.
Asimismo, hay que enviar al Comité Polar el plan de campaña indicando el número de investigadores
que van a trasladarse, zonas de muestreo, fechas aproximadas de llegada y tiempo de permanencia
en cada zona, número aproximado de animales que se van a capturar y número aproximado de heces
que se van a recoger. También hay que solicitar los correspondientes permisos:
- Solicitud de toma de muestras e intromisión perjudicial.
- Solicitud de entrada en zonas ASPA o zonas de especial protección.
Elección de zonas de muestreo
Como ya se ha indicado en los objetivos del proyecto, se elegirán zonas con impacto humano alto
(zonas de turismo y zonas con bases) y zonas con baja o nula presencia humana con la finalidad de
comparar los resultados que se obtengan en ambas zonas y poder de esta forma estimar el impacto
humano sobre las poblaciones de mamíferos marinos.
En la Tabla 1 se recogen las zonas seleccionadas y sus características. Sin embargo, no siempre es
posible el muestreo en todas las zonas solicitadas, ya que las necesidades de las Bases españolas y
de otros proyectos puede dar lugar a cambios de ruta y calendario del BIO Las Palmas. Así, en la cam-
paña 2009-2010 no fue posible la toma de muestras en Cabo Shirref (Tabla 1) debido a que quedaba
fuera de la ruta que el barco había establecido.

53
ponencias
Tabla 1
Zona Localización Impacto humano Animales
Isla Decepción 63°00’S; 60°40’W
Alto (base antártica y
turismo)
Foca de Weddell Lobo antártico
Foca cangrejera Foca leopardo
Caleta Potter, Isla Rey Jorge 62°15’S;58°37’W
Alto (base antártica)
Elefante marino
Punta Hannah, Isla Livingston 62°39’S; 60°36’W
Medio-alto (turismo)
Lobo antártico Elefante marino
Península Byers, Isla Livingston 62°34’S; 61°13’W
Bajo (área protegida)
Elefante marino
Cabo Shirreff, Isla Livingston 62º47’S; 60º27’W
Bajo(área protegida)
Foca de Weddell Lobo antártico
Foca cangrejera Foca leopardo
Isla Avian 67°º46’S; 68°43’W
Bajo (área protegida)
Foca de Weddell Lobo antárticoElefante marino
En la figura 4 se indican las distintas zonas de muestreo solicitadas en el transepto a lo largo de Penín-sula Antárticas. En la figura 5 se muestra la localización de la Base Gabriel de Castilla y las áreas dónde se concentran las diferentes especies animales objeto de muestreo en la Isla Decepción.
Figura 4

54
ponencias
Figura 5
Sin embargo, todo el plan de campaña está sujeto a modificaciones, debido principalmente a las siguientes causas:
- Climatología: las condiciones atmosféricas son muy variables y pueden impedir la salida a las zonas de muestreo. El acceso a las zona siempre se lleva a cabo en Zodiac y con velocidades de viento por encima de 15 nudos no se recomienda la navegación.
- Necesidades de otros proyectos: los diferentes proyectos tienen su plan de campaña y pueden por ejemplo darse casos en los que hayan solicitado desembarcar en islas donde no hay mamíferos marinos (por ejemplo en Isla Yalour) o donde las condiciones del terreno hacen imposible la captura de los mismos.
- Accesibilidad y zonas de fondeo adecuadas.- Personal disponible: para la captura y toma de muestras de lobo marino son necesarias un mínimo
de 5-6 personas y para elefantes y foca de Weddell, entre 8-9 personas. Esto hace que sea nece-saria la colaboración de la tripulación del barco BIO Las Palmas o del personal de la Base Gabriel de Castilla, colaboración que siempre nos han prestado.
Protocolo de toma de muestras
1- Obtención de muestras de heces.
Se recogen heces de suelo para examen parasitológico y bacteriológico principalmente. En esta cam-paña nos hemos ayudado de una cámara termográfica para evaluar la frescura de las heces y tomar muestras de las heces más frescas.

55
ponencias
Para realizar el análisis parasitológico se recogía un mínimo de 25 g de heces. Para el cultivo bacteria-no se tomaban muestras con hisopos, los cuales se introducían en diferentes medios de transporte:
- Medio de transporte Cary-Blair.
- Medio de transporte Amies con carbón activado.
- Medio de criopreservación FBP con carbón activado.
En la campaña 2009-2010 se tomaron un total de 113 muestras de heces con la siguiente distribución:
Elefante marino Foca Weddell Lobo marino
Península Byers 18 - -
Punta Hanna 18 - -
Isla Ronger - - 1
Isla Avian 18 - 7
Isla Rey Jorge. Jubany 15 - 4
Isla Decepción - 3 29
2- Muestras de animales capturados
Una vez que el animal ha sido atrapado y está bien sujeto se procede a la extracción de sangre, ob-tención de hisopos, colocación del crotal, morfometría y toma de muestras de pelo.
Para la extracción de sangre se procede a la desinfección de la zona, colocación del compresor y dependiendo de la especie animal se obtendrá:
- En lobos marinos: se extienden las aletas caudales y se realiza la punción en alguna de las venas interdigitales (Figura 1).
- En elefantes marinos y focas de Weddell: se extienden las aletas caudales y se realiza la punción entre el tercer y cuarto dedo hasta localizar el seno o red venosa plantar (Figura 2).
Seguidamente se realiza la obtención de hisopos rectales, genitales, nasales, oculares y orales. Para su conservación se introducen en los medios de transporte ya citados anteriormente. Además, se introducía un hisopo en PBS estéril para realizar análisis virológicos.
Las muestras se mantienen a -20º C hasta su cultivo. Uno de los principales problemas en relación con el aislamiento bacteriano es el tiempo que transcurre entre la obtención de las muestras (enero / fe-brero) y su cultivo (mayo/junio). En este sentido, los resultados obtenidos con los hisopos mantenidos en el medio de transporte FBP con carbón activado han sido sorprendentes, aislándose una elevada proporción de Campylobacter spp y Mycoplasma spp a partir de los mismos.

56
ponencias
Figura 1 (Tomada de Rommel, 2000) Figura 2 (Tomada de Rommel, 2000)
El marcaje de los animales se realiza con crotales de plástico de diferente color según el año de campa-ña, con un número de identificación y una dirección de correo de contacto. La colocación es diferente según la especie animal, en la aleta dorsal derecha en el caso de otáridos y en la aleta caudal derecha en el caso de pinnípedos (Figura 3).
Figura 3: zona de colocación del crotal en otáridos y pinnípedos.
El pelo se toma con unas pinzas a lo largo de todo el dorso del animal. Finalmente, se mide la longitud del animal desde la punta del hocico hasta la punta de la cola.
En la campaña 2009-2010 se capturaron un total de 80 animales con la siguiente distribución por especies y zonas:

57
ponencias
Elefante marino Foca Weddell Lobo marino
Isla Ronger - 1 -
Isla Avian 2 4 14
Isla Decepción 1 14 44
Finalmente, también se tomaron muestras de la fosa séptica de la Base Gabriel de Castilla y muestras de agua de diferentes puntos de bahía Foster.
Agradecimientos
Proyectos: CGL2004-22025-E, CGL2005-25073-E, CTM2008-00570.
Margarita Yela (MICINN)
Miki Ojeda (MTU)
Sonia Ramos (CPE)
BIO LAS PALMAS Yago Ceballos González-Llanos
BAE GABRIEL DE CASTILLAJosé Gonzálvez Valles (Pepe)
OXOID ESPAÑATania Fernández Sastre


comunicaciones orales


61
comunicaciones orales
EVALUACIÓN DE LA INTERFERENCIA DE LA VACUNACIÓN FRENTE A PARATUBERCULOSIS EN EL DIAGNÓSTICO DE TUBERCULOSIS EN
CABRAS INFECTADAS EXPERIMENTALMENTE CON MYCOBACTERIUM CAPRAE
1Pérez de Val, Bernat*; 1Martín M.; 1López-Soria S.; 1Nofrarias M.; 2Garrido J.M.; 1Solanes D.; 2Juste R.A.; 1Domingo M.
1Centre de Recerca en Sanitat Animal (CReSA), UAB-IRTA, Campus de la Universitat Autònoma de Barcelona, 08193 Bellaterra, Barcelona, Catalunya. 2Departamento de Sanidad Animal, NEIKER-
Tecnalia, 48160 Derio, Bizkaia, Euskadi.
A diferencia de lo que sucede en el ganado bovino, en el que la campaña de erradicación de tubercu-losis en España impone la prohibición del uso de vacunas micobacterianas, la vacunación frente a pa-ratuberculosis está permitida en el caprino pese a que sus efectos en el diagnóstico de la tuberculosis no han sido todavía evaluados. Para estudiar dichos efectos se diseñó el presente estudio en el que de 20 cabras, 10 fueron vacunadas frente a paratuberculosis con 2,5 mg/ml del producto Silirum® (CZV, Porriño, España) y 10 mantenidas como grupo control no vacunado. A las 14 semanas post-vacunación, todas las cabras fueron inoculadas con M. caprae. En la prueba del Interferón-γ (Bovigam, Prionics, Suiza) con antígenos PPD aviar y PPD bovina se observó una respuesta claramente aviar en el grupo vacunado a partir de la semana 4 post-vacunación salvo un 10% y un 60% de positivos bovinos en las semanas 12 y 14, respectivamente; las reacciones en la semana 16 volvieron a ser de tipo aviar (80%) o negativas (20%). El desafío con M. caprae cambió radicalmente la respuesta de ambos grupos de manera que en la semana 18, el 90% de los animales vacunados y el 100% de los controles, y en las sucesivas semanas hasta el final del seguimiento, el 100% de ambos grupos mos-traron reacciones inequívocamente bovinas. Además, las reacciones cruzadas con PPD bovina que se observaron en el grupo vacunado se corrigieron en el 100% de los casos utilizando antígenos es-pecíficos de complejo M. tuberculosis como ESAT-6/CFP-10 y Rv3615c (VLA, Reino Unido). Por otro lado, el 90% de los animales fueron positivos a la prueba de intradermotuberculinización comparada realizada 12 semanas post-infección. Con respecto a las serologías, el 50% y 70% de los animales fueron considerados seropositivos a paratuberculosis (Paratub.Serum-S, Institut Pourquier, Francia) a las 14 semanas post-vacunación y post-infección respectivamente, mientras que todos los anima-les fueron considerados seropositivos a M. caprae por ELISA utilizando el antígeno MPB83 (Lionex, Alemania). Los resultados patológicos y bacteriológicos no mostraron diferencias estadísticamente significativas entre ambos grupos aunque se apreció una ligera tendencia a un efecto de protección cruzada en el grupo vacunado.

62
comunicaciones orales
PREVALENCIA DE CRYPTOSPORIDIUM Y GIARDIA EN BOVINOS DE LA PROVINCIA DE ÁLAVA
Guillermo A. Cardonaa, Pilar Goñib, Larraitz Arriolac, Hélène Carabind, Guy Robinsone, Juan C. Fernández-Crespof, Antonio Clavelb, Rachel M. Chalmerse, David Carmenag
aLaboratorio Pecuario de Eskalmendi, Diputación Foral de Álava, Ctra. de Azua 4, 01520 Vitoria-Gasteiz, España
bDepartmento de Microbiología, Medicina Preventiva y Salud Pública, Facultad de Medicina, Universi-dad de Zaragoza, Domingo Miral s/n, 50009 Zaragoza, España
cUnidad de Epidemiología, Departamento de Salud Pública de Guipúzcoa, Gobierno Vaco, Av. Nava-rra 4, 2013 San Sebastián, España
dDepartment of Biostatistics and Epidemiology, College of Public Health, Oklahoma University Health Sciences Center, 801 Northeast 13th Street, Room 303, Oklahoma City, OK 73104, USA
eUK Cryptosporidium Reference Unit, Public Health Wales, Microbiology ABM, Swansea, Singleton Hospital, Swansea SA2 8QA, UK
fSubdirección de Salud Pública de Álava, Departamento de Sanidad, Gobierno Vasco, Avda. Santia-go 11, 01002 Vitoria-Gasteiz, España
gMRC Clinical Sciences Centre, Cellular Stress Group, Faculty of Medicine, Imperial College, Ham-mersmith Hospital Campus, Du Cane Road, London W12 0NN, UK
ResumenLos protozoarios Cryptosporidium y Giardia son enteropatógenos que pueden infectar una gran varie-dad de vertebrados. Estos microorganismos son una de las principales causas de diarrea en especies pecuarias, generando considerables pérdidas económicas. Presentamos por primera vez datos sobre la prevalencia de estos protozoos en bovinos de la provincia de Álava (País Vasco). La presencia de Cryptosporidium y Giardia fue detectada en muestras de heces analizadas mediante inmunoensayos comerciales: ELISA para la detección de coproantígenos e inmunocromatografía. Se obtuvieron un total de 227 muestras de heces procedentes de 22 explotaciones ganaderas entre Octubre de 2008 y Mayo de 2009. Dependiendo de la técnica usada, la presencia de Cryptosporidium y Giardia fue detectada en el 3,1–7,5% y 1,8–5,3% de las muestras analizadas, respectivamente. En el 62% de las explotaciones estudiadas se hallaron animales infectados con Cryptosporidium, y en el 43% con Giardia. Las muestras positivas a Cryptosporidium y Giardia por ELISA/inmunocromatografía fueron re-analizadas mediante microscopia convencional o de inmunofluorescencia y posterior caracteriza-ción molecular mediante PCR usando como marcadores los genes que codifican para la subunidad pequeña del ARN ribosómico (Cryptosporidium) o la enzima triosa fosfato isomerasa (Giardia). Ningu-no de los aislados pudo ser amplificado con éxito, por lo que se desconoce el potencial zoonótico de estos hallazgos. Nuestros resultados demuestran que la cryptosporidiosis/giardiosis son enfermeda-des presentes en el ganado bovino de la provincia de Álava, aunque con unas prevalencias inferiores a las reportadas en poblaciones bovinas de similares características en otras regiones españolas.
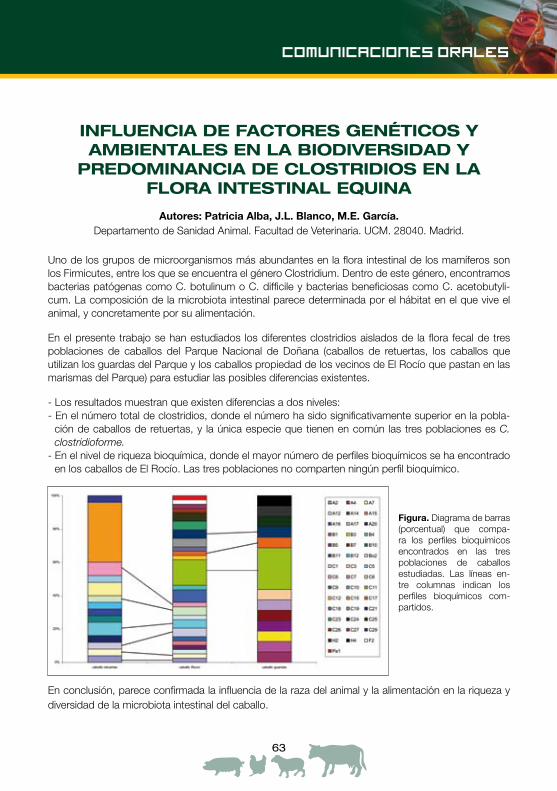
63
comunicaciones orales
INFLUENCIA DE FACTORES GENÉTICOS YAMBIENTALES EN LA BIODIVERSIDAD Y
PREDOMINANCIA DE CLOSTRIDIOS EN LA FLORA INTESTINAL EQUINA
Autores: Patricia Alba, J.L. Blanco, M.E. García.Departamento de Sanidad Animal. Facultad de Veterinaria. UCM. 28040. Madrid.
Uno de los grupos de microorganismos más abundantes en la flora intestinal de los mamíferos son los Firmicutes, entre los que se encuentra el género Clostridium. Dentro de este género, encontramos bacterias patógenas como C. botulinum o C. difficile y bacterias beneficiosas como C. acetobutyli-cum. La composición de la microbiota intestinal parece determinada por el hábitat en el que vive el animal, y concretamente por su alimentación.
En el presente trabajo se han estudiados los diferentes clostridios aislados de la flora fecal de tres poblaciones de caballos del Parque Nacional de Doñana (caballos de retuertas, los caballos que utilizan los guardas del Parque y los caballos propiedad de los vecinos de El Rocío que pastan en las marismas del Parque) para estudiar las posibles diferencias existentes.
- Los resultados muestran que existen diferencias a dos niveles:- En el número total de clostridios, donde el número ha sido significativamente superior en la pobla-
ción de caballos de retuertas, y la única especie que tienen en común las tres poblaciones es C. clostridioforme.
- En el nivel de riqueza bioquímica, donde el mayor número de perfiles bioquímicos se ha encontrado en los caballos de El Rocío. Las tres poblaciones no comparten ningún perfil bioquímico.
En conclusión, parece confirmada la influencia de la raza del animal y la alimentación en la riqueza y diversidad de la microbiota intestinal del caballo.
Figura. Diagrama de barras (porcentual) que compa-ra los perfiles bioquímicos encontrados en las tres poblaciones de caballos estudiadas. Las líneas en-tre columnas indican los perfiles bioquímicos com-partidos.

64
comunicaciones orales
DESARROLLO DE UN ELISA DE DOBLE RECONOCIMIENTO PARA LA DETECCIÓN DE
ANTICUERPOS ESPECÍFICOS DEL VIRUS DE LA FIEBRE DEL VALLE DEL RIFT
Rebollo Belén1., Venteo A1., Boshra, H.2, Martín-Folgar R.2, Brun A.2 y Sanz A1. 1. INGENASA 2. CISA- INIA
El virus del la fiebre del valle del Rift (RVFV) pertenece al género Phlebovirus de la familia Bunyavi-ridae. Es el causante de una enfermedad emergente que se transmite por numerosas especies de mosquitos y causa graves epidemias en rumiantes y el hombre en África y la península Arábiga. Una vacuna efectiva y un diagnóstico serológico eficaz son objetivos prioritarios para el control de esta enfermedad, considerada en USA como potencial agente bioterrorista. INGENASA en colaboración con el CISA-INIA ha desarrollado un ELISA de Doble Reconocimiento (DR) capaz de detectar anti-cuerpos específicos del RVFV. El ELISA DR se basa en el antigenado de la placa con la proteína de la nucleocápsida viral (N) expresada en E.coli y el uso de la misma proteína conjugada con peroxidasa como revelador. Es por tanto un test apto para cualquier especie y tipo de anticuerpo (IgG, IgM, IgA, etc.). Para valorar la sensibilidad del ensayo se analizaron ovejas inmunizadas experimentalmente con plásmidos de expresión que llevan insertado el gen de la proteína N bajo el promotor del CMV. Poste-riormente, todos los animales recibieron un desafío de virus atenuado y se analizaron serológicamente. El ensayo fue capaz de detectar como positivos tanto los animales vacunados como los infectados. El análisis ROC de los resultados obtenidos con 490 ovejas y 94 vacas de zonas libres de la enfermedad indicó un 99.5% de especificidad. Actualmente se están analizando sueros procedentes de diferentes orígenes para confirmar la sensibilidad analítica y la utilidad de este ensayo para diagnóstico y vigilan-cia epidemiológica de la fiebre del Valle del Rift.

65
comunicaciones orales
DIAGNÓSTICO SEROLÓGICO DE SALMONELLA SPP.: CONSIDERACIONES ANTE SU USO COMO
HERRAMIENTA DE CLASIFICACIÓN EN FUTUROS PROGRAMAS DE CONTROL
EN PORCINO
Vico, Juan Pablo; Mainar-Jaime R.C. Centro de Investigación y Tecnología Agroalimentaria,
Gobierno de Aragón. Avda. Montañana, 930. 50059 Zaragoza.
Uno de los principales objetivos de la UE en materia de Salud Pública es el control de la salmonelosis animal. La serología, a partir de suero o jugo muscular, se perfila como método de elección para la implementación de programas de control de salmonelosis porcina. Sin embargo, el rendimiento diag-nóstico de las mismas no ha sido suficientemente evaluado en animales de campo. En un primer estu-dio se compararon tres ELISAS comerciales (Herdcheck® Swine Salmonella, Salmotype® Pig Screen, y PrioCHECK® Salmonella) sobre 837 animales pertenecientes a 34 rebaños. Se observó una gran variabilidad en cuanto al rendimiento diagnóstico, lo que resultó en que una misma explotación pudiera ser clasificada de forma diferente respecto a su nivel de seroprevalencia en función del ELISA utilizado. En otro estudio se evaluó la concordancia entre resultados de densidad óptica (DO) a partir de suero y jugo muscular con un ELISA comercial (Herdcheck® Swine Salmonella) en dos poblaciones porcinas con diferentes seroprevalencias (51 hembras adultas y 41 cerdos en engorde). Los resultados medios de DO de suero sanguíneo fueron significativamente más altos que los obtenidos de jugo muscular (38.5 vs. 27.9; P<0.001), siendo la diferencia media entre ellos significativamente distinta de cero (P<0.0001). El coeficiente de correlación entre los resultados de ambas matrices, aunque significativo, fue bajo (r=0.53). Estos resultados evidencian la importancia de la elección tanto del ELISA como de la matriz (jugo muscular ó suero sanguíneo) a la hora de implementar programas de control para reducir la prevalencia de Salmonella spp. en cerdos.

66
comunicaciones orales
DESARROLLO DE UN ELISA DE BLOQUEO PARA LA DETECCIÓN DE ANTICUERPOS ESPECÍFICOS
DEL VIRUS DEL NILO OCCIDENTAL
Venteo Angel1., Rebollo B1., Sotelo, E.2, Llorente, F.2, Gallardo, C.2, Figuerola, J.3, Jiménez-Clavero MA. 2, Camuñas A. 1, Rodríguez MJ1., Sarraseca J1. y Sanz A1.
1. INGENASA. 2 CISA INIA. 3EBD-CSIC.
El virus del Nilo occidental (VNO) es un patógeno zoonótico emergente transmitido por artrópodos perteneciente a la familia Flaviviridae, género Flavivirus, con un amplio rango de hospedadores dife-rentes (aves, caballos y humanos). INGENASA en colaboración con el CISA-INIA ha desarrollado un diagnóstico basado en un ELISA de bloqueo capaz de detectar anticuerpos específicos del VNO en las diferentes especies susceptibles. El ensayo utiliza un anticuerpo monoclonal específico de la gli-coproteína E de la envuelta viral, que neutraliza la infección por WNV in vitro. La evaluación del test se realizó teniendo como referencia la técnica de seroneutralización (VNT), gold estándar para esta pato-logía. Utilizando sueros experimentales se demostró la precocidad del ensayo al detectar anticuerpos a día 3 post-infección. Además se analizaron 146 sueros de aves silvestres y 91 sueros de caballo de campo obteniendo una especificidad del 81,2% y el 96,5% respectivamente, y un 100% de sensibili-dad con respecto a la VNT. En suero de 8 ratones inoculados con otro flavivirus (Usutu), el ELISA fue negativo. En sueros de 5 aves silvestres infectadas con virus Usutu y positivos para VNT solo 1 de ellas fue positiva en el ELISA. Este ensayo, además de su alta sensibilidad, especificidad y amplio rango de uso, tiene la ventaja respecto a otros ELISAs comerciales de requerir una mínima cantidad de muestra (10 µl) por ensayo. Por su rapidez y sencillez en relación a la VNT puede ser un método serológico de criba útil para diagnóstico y vigilancia epidemiológica de esta enfermedad.

67
comunicaciones orales
ESTUDIO DE LA BIODIVERSIDAD DE LA FLORA FECAL DEL GANSO COMÚN: INFLUENCIA DE LA MIGRACIÓN
Autores: José Luis Blanco1, P. Alba1, J. Figuerola2, M.E. García1.1. Departamento de Sanidad Animal. Facultad de Veterinaria. UCM. 28040. Madrid.
2. Estación Biológica de Doñana. 41013 Sevilla.
El ganso común (Anser anser) es una especie de ave migratoria que pasa los inviernos en el sur de Europa. El estudio de su flora fecal, y los factores que la condicionan, es clave para determinar su capacidad como animal portador de enfermedades infecciosas. Por ello, se ha estudiado la sucesión ecológica que se produce en el ecosistema intestinal comparando los microorganismos que encon-tramos en la flora intestinal de una población de gansos, al llegar al Parque Nacional de Doñana y tras dos meses de invernada allí.
Los resultados indican que:- No hay diferencias significativas en el número total de bacilos grampositivos, cocos grampositivos y
bacilos gramnegativos aislados en uno y otro grupo. - Se observan diferencias en las especies concretas aisladas.- En el segundo grupo, el que ha pasado dos meses en el Parque, se aprecia una disminución de
diversidad a nivel de género y un aumento a nivel de especies en la familia Enterobacteriaceae.- La abundancia es significativamente mayor en el segundo grupo. - La extrapolación matemática del número total de aislados indica que el número real de microorganis-
mos en el ecosistema intestinal es de uno o dos órdenes de magnitud mayor que el hallado.- Se han encontrado especies de microorganismos no descritas en la actualidad.
Figura. Representación gráfica del porcen-taje de microorganismos diferentes aislados. En la imagen A se muestran los aislados en el conjunto del primer grupo de gansos, deno-minado A. En la imagen B, los resultados del grupo de gansos B. En ambas figuras, el azul representa el porcentaje de bacilos gramne-gativos, el granate el de cocos grampositivos y el amarillo el de bacilos grampositivos.
Estos resultados reflejan la influencia del hábitat del animal, y de los alimentos que encuentra en cada biotopo, en la riqueza y abundancia del ecosistema intestinal.

68
comunicaciones orales
AISLAMIENTO E IDENTIFICACIÓN DE BESNOITIA BESNOITI EN CULTIVOS CELULARES A PARTIR
DE MUESTRAS CLÍNICAS1Aleida Villa Espinosa; 1Ana Fernández Ros y 2Gema Álvarez García
1EXOPOL Autovacunas y Diagnóstico. Pol. Río Gállego, 50840 San Mateo de Gállego Zaragoza, España. Tel 976 69 45 25, [email protected]
2Facultad de Veterinaria (UCM) Dpto. Sanidad animal. Grupo Saluvet Pabellón central, planta- 3 Avda. Puerta de Hierro s/n 28040 Madrid.
ResumenBesnoitia besnoiti, un protozoo de la familia Sarcocystidae es el agente etiológico de la Besnoitiosis una enfermedad emergente en Europa propagada por tábanos y moscas. Los hospedadores interme-diarios son bóvidos y otros rumiantes a los que infecta provocándoles anorexia, edemas, anasarca, esterilidad, ataxias, alopecias, engrosamiento de la piel y deterioro del bienestar animal.
El muestreo se realizó en una explotación de ganado bovino Charolais de la provincia de Salamanca. Algunos sementales con graves lesiones clínicas fueron sacrificados y los pulmones, ojos y piel se emplearon para el aislamiento e identificación del parásito.
Los estudios realizados incluyeron: observación microscópica directa de los quistes en fresco, cultivo in vitro en las líneas celulares MDBK; MA-104; VERO y COE; identificación de los taquizoitos en cultivo mediante, tinción de PAS, Inmunofluorescencia indirecta y PCR, así como la evaluación serológica de animales sintomáticos y asintomáticos dentro del foco.
Todas las líneas celulares empleadas fueron eficientes para la multiplicación de Besnoitias obtenién-dose ocho aislamientos.
Hemos podido establecer los protocolos de diagnóstico in vitro para Besnoitia besnoiti y elaborar medios de diagnóstico a partir de cystozoitos y taquizoitos de cepas autóctonas.

69
comunicaciones orales
ESTUDIO DE LA EVOLUCIÓN DE ANTICUERPOS DE COXIELLA BURNETII EN UN REBAÑO DE
VACAS LECHERAS DE ALTA PRODUCCIÓN EN EL NORESTE DE ESPAÑA DURANTE UN AÑO.
Tutusaus Joan, Garcia-Ispierto I, López-Gatius F, Nogareda,C. Departament de Producció animal. ETSEA. Universitat de Lleida
Coxiella burnetii, agente causante de Fiebre Q, es una zoonosis distribuida mundialmente. Esta bacte-ria puede producir desórdenes reproductivos tales como abortos, muerte fetal, crías débiles, mastitis, metritis e infertilidad en el ganado vacuno lechero. El diagnóstico serológico por ELISA se considera una de las pruebas de elección para el estudio seroepidemiológico. El objetivo de este trabajo fue evaluar los anticuerpos de C. burnetii en 478 vacas lecheras de alta producción. El análisis serológico se realizó mediante un kit comercial de ELISA indirecto LSIVET. Una muestra se consideró negativa, positiva 1, 2, 3 o 4 según si el título era ≤ 40, de 40 a ≤ 100, de 100 a ≤ 200, de 200 a ≤ 300 y > 300, respectivamente. La prevalencia de anticuerpos fue del 51% en 2009 y 49,4% en 2010. El 65% de los animales se mantuvieron estables en el mismo grupo, el 10,5% disminuyeron y el 24,5% aumen-taron durante el periodo de estudio. El 4,8% de los animales seropositivizaron, mientras que un 6,5% seronegativizaron. El análisis de la leche de tanque por RT-PCR en los dos años estudiados confirmó una fuerte excreción bacteriana (>10.000 bacterias/ml). Los resultados obtenidos son similares a otros publicados, sugiriendo la amplia distribución de la infección y la estabilidad de los anticuerpos.

70
comunicaciones orales
VALORACIÓN DE TRES ELISAS DE DETECCIÓN DE ANTICUERPOS EN SUERO
FRENTE A M.HYOPNEUMONIAE.
Rebordosa-Trigueros Xavier, Porquet-Garanto LourdesLaboratorios Hipra, S.A 17170 Amer, Girona, Spain.
Mycoplasma hyopneumoniae es un microorganismo patógeno que presenta una elevada incidencia económica en la producción porcina a nivel mundial. Se asocian con la infección por M.hyopneumoniae desde cuadros bronco-neumónicos suaves hasta neumonías severas. Estas últimas se asocian nor-malmente con infecciones secundarias por virus o bacterias. Es así M.hyopneumoniae una de las piezas importantes del denominado Complejo Respiratorio Porcino (CRP). Por ello, es muy importante el poder disponer de sistemas adecuados de seguimiento y control de la circulación de este microor-ganismo en la explotaciones porcinas. Uno de los sistemas habituales de control de la infección por M.hyopneumoniae es la serología. Cualquier prueba serológica frente a M. hyopneumoniae debe tener en cuenta que la sero-conversión asociada a la infección por M.hyopneumoniae es lenta, comparada con la que se observa en otros patógenos habituales del cerdo. Resulta además conveniente que estas pruebas detecten la sero-conversión asociada con la vacunación y que, así mismo, posibiliten el segui-miento de la circulación del microorganismo después de la vacunación. En el presente trabajo hemos comparado tres ELISAs desarrollados para la detección de anticuerpos frente a M.hyopneumoniae en suero porcino. Para ello se ha partido de un conjunto de muestras procedentes de animales libres de M.hyopneumoniae que posteriormente se han vacunado utilizando diferentes vacunas comerciales y finalmente se han infectado de forma experimental. Se han analizado y comparado las prestaciones de cada uno de estos ELISAs frente a los tres tipos de muestras generadas en este estudio; sueros libres o negativos, sueros vacunados y sueros vacunados/infectados. Los resultados han mostrado diferencias en las prestaciones de cada uno de estos productos que creemos que se deben tener en cuenta a la hora de interpretar los resultados serológicos.

71
comunicaciones orales
PRIMERA DETECCIÓN DE BABESIA OCCULTANS EN GARRAPATAS DEL
GÉNERO HYALOMMA PROCEDENTES DE TÚNEZ
Amaia Ros-García1, M’Ghirbi Y.2, Bouattour A. 2, Juste R.A.1, Hurtado A. 1
1 Department of Animal Health, NEIKER - Instituto Vasco de Investigación y Desarrollo Agrario, Berreaga 1, 48160 Derio, Bizkaia, España
2 Service d’Entomologie Médicale, Institut Pasteur de Tunis, 1002 Tunis, TúnezE-mail : [email protected]; [email protected]
Los hemoparásitos de los géneros Theileria y Babesia son agentes patógenos transmitidos por ga-rrapatas que causan importantes pérdidas económicas. El control efectivo de estas infecciones re-quiere técnicas de diagnóstico rápidas, sensibles y específicas que permitan detectar e identificar las distintas especies que infectan a los animales y las garrapatas. Una técnica que cumple estos requisitos es la hibridación por “Reverse Line Blot” (RLB), consistente en la hibridación de los pro-ductos PCR con sondas específicas de especie, género y grupo, inmovilizadas en una membrana de nylon y dispuestas a modo de macroarray. Dentro de un estudio sobre las piroplasmosis en Túnez se recogieron muestras de sangre y garrapatas no alimentadas para su análisis por RLB. En 3 ga-rrapatas Hyalomma marginatum se obtuvo una intensa señal de hibridación con la sonda de grupo que identifica a todas las especies de piroplasmas pero no se observó hibridación con ninguna de las sondas especie-específicas indicando la presencia de una nueva especie. Tras clonar y secuenciar el gen 18S rRNA completo se identificó como Babesia occultans, una babesia bovina de escaso poder patógeno descrita en África Subsahariana. Este estudio constituye la primera descripción de B. occultans en Túnez y demuestra la utilidad de la RLB para el estudio de la prevalencia y diversidad de piroplasmas presentes en una determinada región. Como parte de este estudio se ha diseñado además una nueva sonda específica para B. occultans que hemos incorporado al panel de 24 son-das disponibles en nuestro laboratorio para la detección e identificación de piroplasmas.

72
comunicaciones orales
ENSAYO COLABORATIVO DE UN PROTOCOLO DE ELISA DE TUBERCULOSIS BOVINA EN
SUIDOS
Juste RA1, Boadella M2, Garrido F3, Domínguez L4, Garrido JM1, Gortázar C2, de Juan L4
1 NEIKER-Tecnalia. Derio. Bizkaia 2 IREC. Ciudad Real
3 Laboratorio Central de Sanidad Animal de Santa Fe. Santa Fe. Granada4 VISAVET. Madrid
El desarrollo de nuevas técnicas de diagnóstico laboratorial requiere, además de su validación inicial, una evaluación de su eficacia en las distintas condiciones de trabajo de cada laboratorio. La tubercu-losis bovina es un problema importante en el UE en el que algunas especies silvestres parecen jugar un papel que hace que su erradicación de los sistemas de producción bovina más extensivos se vea muy dificultado. Las pruebas serológicas no se han considerado tradicionalmente suficientemente eficaces para ser aplicadas en el diagnóstico de esta infección. Sin embargo, recientes estudios en jabalí han demostrado que en esta especie tienen una elevada eficacia. La verificación de que este método tiene la suficiente robustez para su aplicación extensiva requería su aplicación en diferentes laboratorios. Para ello se ha realizado un ensayo colaborativo en el que han participado 4 laboratorios y en el que se han utilizado 25 sueros de jabalíes infectados de forma natural y 21 negativos cuyo estatus se determinó mediante técnicas microbiológicas y patológicas. El test utilizado ha sido el TB-ELISA-VK (Vacunek, Derio, Bizkaia). Los resultados indican que la prueba utilizada tiene unos niveles de concordancia (kappa) excelentes que fueron desde el 0,7310 al 0,9560. La sensibilidad y el valor predictivo negativo fueron del 100% en todos los laboratorios y repeticiones. La robustez del test se confirmó por los resultados de uno de los laboratorios que, aun no validando el control positivo, no sufrió una disminución de los valores alcanzados para los parámetros señalados.

73
comunicaciones orales
VALIDATION OF COMMERCIAL ELISA KITS FOR DETECTION OF ANTI-BTV ANTIBODIES IN NATURALLY-INFECTED AND VACCINATED
ANIMALS
Loic Comtet, Michael Mockey, Philippe Pourquier1 IDVET, 167 rue Mehdi Ben Barka, 34070 Montpellier, France ([email protected])
2 IDVET, Montpellier, France
The IDVET range of Bluetongue ELISA kits are used worldwide and in particular in Northern Europe, for detection of anti-BTV antibodies in naturally-infected and vaccinated animals.
The poster will present the IDVET BTV product line, including:
- indirect milk ELISA- Early Detection serum ELISA- One-Step serum ELISA and- Classical competitive VP7 serum ELISA.
DVET’s latest innovations using other antigens for BTV sero-diagnotics will also be presented.


Posters


77
Posters
DISTRIBUCIÓN GEOGRÁFICA DE CULICOIDES (DÍPTERA: CERATOPOGONIDAE) EN LA
COMUNIDAD VALENCIANA.
A. Tamarit, B. Cortell, C. Mollá, ML. Roche, C.Caballero. Unidad de Análisis en Sanidad Animal. Consellería de Agricultura, Pesca y Alimentación. Generalitat Valenciana.
Resumen: Durante el 2009 se realizó en nuestro Laboratorio la identificación entomológica de la fauna de dípte-ros del género Culicoides de nuestra Comunidad, incluida dentro del Programa Nacional de Lengua Azul.
Durante este periodo se analizaron un total de 364 muestras. En cada muestra se realizó el recuento total de Culicoides y se procedió a la identificación de los mismos utilizando claves taxonómicas ba-sadas en su patrón alar.
Del total de Culicoides presentes en las muestras, se identificó el complejo Culicoides obsoletus en el 50.4% y el complejo Culicoides pulicaris en el 35%. Las otras especies identificadas (14.6%), se distribuyeron de la siguiente forma: C. circumscriptus 5.5%, C. univittatus 5%, C. parroti 1.1%, C. fes-tivipennis 0.7%, C. maritimus 0.5%, C. nubeculosus 0.03% y Culicoides spp. con un 1.6%. La especie C. parroti sólo llegó a identificarse en la provincia de Castellón. La distribución geográfica de estos complejos fue distinta según la provincia. El complejo C. obsoletus fue predominante en la provincia de Castellón con un 66%, mientras el complejo C. pulicaris predominó en Alicante (97,5%). Los datos para la provincia de Valencia fueron 9.4% para el complejo C. obsoletus y 75% para el complejo C. pulicaris. En el complejo C. pulicaris la especie mayoritaria fue C. newsteadi. De esta forma se observa la predilección en la distribución del complejo C. obsoletus por la zona norte de nuestra comunidad.
Agradecimientos: Nuestro más sincero agradecimiento al Dr. D. Javier Lucientes y a la Dra. Dª. Rosa Estrada por su labor de asesoramiento y apoyo.

78
Posters
DETECCIÓN DE PROTEÍNA PRION PATOLÓGICA EN PLACENTAS DE OVINO COMO
MÉTODO DIAGNÓSTICO DE SCRAPIE1Garza Mª Carmen, 1Acín C., 1Marín B., Bolea R., Monzón E., 1Badiola J.J., 2Monleón E.
1Dpto. Patología animal. Universidad de Zaragoza [email protected]. Producción Animal. Universitat de Lleida.
La detección de la proteína prion patológica (PrPsc) en tejido linfoide obtenido mediante biopsias es el único método de diagnóstico in vivo de la enfermedad de scrapie. Sin embargo, esta técnica presenta una baja sensibilidad debido principalmente a la gran variabilidad en la implicación de dicho tejido en la patogenia del scrapie. La acumulación de la PrPsc en placentas procedentes de animales infectados, incluso en fases preclínicas de la enfermedad, hace que este tejido también pueda ser utilizado para el diagnóstico in vivo. El objetivo del presente estudio es la valoración de la placenta como muestra para el diagnóstico in vivo del scrapie, utilizando técnicas de diagnóstico rápido.
Se procesaron un total de 50 placentas procedentes de animales enfermos de scrapie diagnosticados mediante biopsia de tejido linfoide. Las muestras se analizaron mediante la técnica de inmunohistoquí-mica y las técnicas rápidas modificadas de Western Blot de Prionics y los ELISA de IDEXX y Bio-Rad. Los resultados mostraron que la técnica de diagnóstico más rápida y sensible es el ELISA de IDEXX. Sólo cuando los fetos presentan genotipo resistente, la sensibilidad del diagnóstico es menor que con las biopsias de tejido linfoide.
Además, se analizaron todos los cotiledones de dos placentas (73 y 49) de animales infectados de scrapie que no presentaban signos clínicos de la enfermedad, detectándose PrPsc en todos, mos-trando por tanto una distribución amplia y homogénea de la PrPsc en este tejido.

79
Posters
PREVALENCIA DE MICOTOXINAS ANALIZADAS EN MATERIAS PRIMAS RECIBIDAS EN NUESTRO
LABORATORIO DURANTE EL AÑO 2008
Análisis realizados por ELISA y HPLCA.C. Letosa Almalé, B.Bringola Casas, L.Espada Boix
Aflatoxina B1 Pocas muestras positivas, valores muy bajos en general.MAIZ: materia prima con más muestras positivas y con los valores más elevados pero se encuentran dentro de los niveles permitidos por legislación en todos los casos excepto en una muestra (33.95 ppb).
Ocratoxina A Poca presencia en general en todas las materias primas, alguna muestra positiva des-tacable maíz (228.28 ppb), cebada (89.42 ppb) y salvado de trigo (35 ppb), siempre por debajo del valor recomendado.
Deoxinivalenol Presencia en todas las muestras siempre con valores dentro de lo recomendado.MAIZ: Amplio porcentaje de las muestras son positivas con valores diversos hasta 4.000 ppb.CEBADA, HARINA DE SOJA, la mayoría de las muestras positivas, con menores valores de contami-nación que en maíz, un máximo de 176.92 ppb cebada, 413.36 ppb harina de soja.TRIGO, y SALVADO DE TRIGO: Muchas muestras son positivas, destacan valores como 4.150 ppb en trigo y 1.600 ppb en salvado de trigo.
Fumonisinas Los valores obtenidos en materias que no sean maíz no son significativos.MAÍZ: La mayoría de las muestras son positivas hasta un máximo de 10.000 ppb, dos muestras con concentraciones aún mayores, con 28.000 ppb y mm22 con 32.400 ppb. Valores por debajo de lo recomendados.
Zearalenona En general todas las materias primas con baja prevalencia, valores por debajo de lo recomendado.MAÍZ y HARINA DE SOJA: Pocos positivos, menores de 180 ppb, destaca un maíz con 2.400 ppb.CEBADA, TRIGO, SALVADO DE TRIGO: Pocos valores positivos, menores de 60 ppb.

80
Posters
MICOTOXINAS EN PIENSOS ANALIZADOS EN NUESTRO LABORATORIO DURANTE EL AÑO
2008
Análisis realizados por ELISAA.C. Letosa Almalé, B.Bringola Casas, L.Espada Boix
Los datos utilizados para el presente estudio corresponden a analíticas de micotoxinas llevadas a cabo en muestras de piensos durante 2008. Las muestras problema corresponden a muestras remitidas al laboratorio de diagnóstico de INZAR, S.L., bien como parte de muestreos para control de calidad de piensos y materias primas, bien como muestras sospechosas de una posible contaminación por micotoxinas con o sin sintomatología en los animales.
A partir de los resultados obtenidos podemos destacar las siguientes conclusiones:
Aflatoxina B1: No se observan diferencias por especie. Todos los valores dentro de los niveles legisla-dos. El 58% del total de muestras de pienso tienen valores menores al límite de detección (1 ppb).
Ocratoxina A: No se observan diferencias por especie. Todos los valores dentro de los niveles legisla-dos. El 65% del total de muestras de pienso tienen valores menores al límite de detección (2.5 ppb).
Deoxinivalenol: No se observan diferencias por especie. El 92% del total de muestras de pienso tie-nen valores mayores al límite de detección (18.5 ppb). Tres muestras de pienso de cerdos con valores por encima de los recomendados.
Fumonisinas: No se observan diferencias por especie. El 98% de las muestras tiene valores mayores al límite de detección. Todas las muestras con valores dentro de lo legislado.
Zearalenona: No se observan diferencias por especie. Cuatro muestras de pienso de cerdos (lecho-nes) por encima de los valores recomendados.

81
Posters
PREVALENCIA DE CAMPYLOBACTER SPP. EN BROILERS A SACRIFICIO DESDE 1998 A 2009
Biarnés Suñé, Mar1*, Blanco, A.1, Camprubí, Q.1, Canals, N.1 y Porta, R.1
1Centro de Sanidad Avícola de Cataluña y Aragón (CESAC), Reus, Españ[email protected]
ABSTRACTCampylobacter spp, y principalmente Campylobacter jejuni, es el agente zoonótico más aislado de las toxoinfeciones alimentarias en humana en la UE. Entre otras, se ha descrito como causa de Cam-pilobacteriosis el consumo o manipulación de carne de pollo cruda contaminada. Las especies de Campylobacter más aisladas en avicultura son: C. jejuni, C. coli y C. lari.
En el CESAC, Centro de Sanidad Avícola de Cataluña y Aragón, se realiza los análisis establecidos en los programas de control sanitario de todas las explotaciones avícolas de Cataluña y Aragón, para el control de las enfermedades más importantes en avicultura.
El objetivo de este trabajo fue conocer la prevalencia en las explotaciones de Cataluña de Campylo-bacter spp en pollos de engorde a la edad de sacrificio. El intervalo del estudio fue desde 1998 al 2009 y se analizaron un total de 1.290 lotes de explotaciones escogidas aleatoriamente. La técnica utilizada fue el aislamiento e identificación de Campylobacter spp a partir de muestras de contenido intestinal.
Los años 2003, 2004 y 2005, fueron los más representativos en muestreo, con más de 150 lotes de pollos analizados por año. El porcentaje de positivos fue en todos los años superior a un 68%. La me-dia de los porcentajes de positivos de los 3 años con más muestreo fue 79,4% y la de todos los años de 86,66%. De los 12 años del estudio, sólo en 1998, 2000, 2003 y 2007, el porcentaje de aislados de C. jejuni, fue superior al resto de especies, concretamente un 57,14%, 49,5%, 46,2% y 43%.

82
Posters
ESTUDIO DE LOS NIVELES DE ANTICUERPOS EN MUESTRA DE LECHE DE TANQUE EN
BROTES DE BVD
Ignacio Arnaiz Seco1, Carnero MI 1, Calavia P 1, Eiras C 1, Diéguez FJ 2
1Laboratorio de Sanidade e Produción Animal de Galicia. Consellería do Medio Rural. 2Departamento de Producción Animal. Facultade de Veterinaria. Universidade de Santiago de Compostela.
La diarrea vírica bovina (BVD) es una enfermedad vírica ampliamente difundida. En la aparición de brotes juegan un papel fundamental los animales persistentemente infectados (PI), producto de la infección en los primeros meses de gestación de sus madres con el virus de la BVD. Su presencia en la explotación, ya sea por nacimiento o incorporación, produce una rápida difusión del virus que se traduce en una fuerte respuesta de anticuerpos en el resto de animales del rebaño. El muestreo periódico de leche de tanque para la analítica de anticuerpos frente a la BVD ha demostrado ser una valiosa herramienta para la detección y seguimiento de brotes en las explotaciones bovinas lecheras. El objetivo de este estudio es determinar los diferentes perfiles de anticuerpos en muestras de leche de tanque, en función de las causas que provocaron su aparición.
Se estudiaron 50 explotaciones con un brote de BVD comprobado por la detección de al menos un animal PI. El comportamiento en la respuesta de anticuerpos en las muestras de leche de tanque obtenidas durante los 12 meses anteriores y posteriores a la aparición de uno o más animales PI varió significativamente según el brote se produjera por incorporación de los animales, nacimiento en la explotación o presencia durante varios años de los mismos.

83
Posters
CARACTERIZACIÓN DE CEPAS DE ESCHERICHIA COLI PRODUCTORAS DE
BETA-LACTAMASAS (BLEEs) DE ESPECTRO EXTENDIDO EN GANADO VACUNO DE GALICIA
Gómez Gómez Verónica1,2, Méndez A1, Mamani R2, López C2, Lamas A1, Ruíz de Valbuena JA1, Blanco J2, Mora A2.
Laboratorio de Sanidade e Produción animal de Galicia. Consellería do Medio Rural. Xunta de Galicia (1) e Departamento de Microbioloxía e Parasitoloxía, USC (2).
Las beta-lactamasas de espectro extendido (BLEEs) se definen como enzimas capaces de hidrolizar las cefalosporinas de amplio espectro y los monobactamas, pero no las cefamicinas o los carbapene-mas. Actualmente existe una gran preocupación por el incremento observado de cepas de Escheri-chia coli portadoras de BLEEs tanto en pacientes humanos como animales.
En el presente estudio caracterizamos los tipos de BLEEs y las resistencias antimicrobianas asociadas producidas por 38 cepas de Escherichia coli aisladas de casos clínicos (mamitis y diarreas) en ganado bovino de Galicia. Las muestras fueron obtenidas en el Laboratorio de Sanidad y Producción Animal de Galicia (LASAPAGA) entre los años 2003-2009.
Determinamos que 28 de las 38 cepas de E. coli estudiadas eran productoras de BLEE tipo CTX-M-14 y 10 cepas del tipo CTX-M-32. Además encontramos una asociación significativa entre la producción de CTX-M-14 y la resistencia a la gentamicina, y entre la producción de CTX-M-32 y las resistencias al aztreonam y ceftazidima.
En este estudio hemos descrito, por primera vez en España, el aislamiento en ganado vacuno de ce-pas de Escherichia coli productoras de CTX-M-14 y, por primera vez a nivel mundial, de productoras de CTX-M-32.

84
Posters
OPTIMIZACIÓN DE UN MÉTODO PARA REALIZAR PCR A PARTIR DE MUESTRAS CONGELADAS DE CEPAS DEL GÉNERO
CAMPYLOBACTER EN MEDIOS SEMISÓLIDOS.
Carbonero Martínez, Alfonso, Torralbo, A., Borge, C., García-Bocanegra, I., Arenas, A., Arenas- Montes, A., Gómez-López, J.C., Perea, A.
Departamento de Sanidad Animal. Universidad de Córdoba.
En el marco de un trabajo etio-epidemiológico acerca del género Campylobacter en perros, se aislaron casi cien cepas que fueron clasificadas dentro de este género por sus características fenotípicas, tras lo que fueron congeladas a -60 ºC para su almacenamiento en un medio semisólido que aumenta la tolerancia al oxígeno. En un siguiente paso, se procedió a realizar una PCR multiplex para la identifica-ción a nivel de género y especie, pasando las colonias previamente a agar sangre como se describe en distintos protocolos, y aislándose a continuación el ADN mediante el método de adición de cloroformo y posterior calentamiento – congelación. Sin embargo, cerca de un 20% de las cepas no pudieron volver a aislarse en los medios de cultivo tras su congelación. En consecuencia, se decidió intentar el aislamiento de ADN directamente a partir del medio de congelación, lográndose la identificación con éxito en el 100% de las ocasiones.
Con este nuevo protocolo que se propone no sólo se logra no incrementar el número de cepas re-cuperadas tras la congelación, sino que también se reduce sustancialmente el tiempo invertido, ya que se ahorran las 48 horas precisas para el crecimiento de las cepas en agar. Así mismo, supone un ahorro económico, al eliminarse el coste de los medios y de los reactivos precisos para proporcionar las condiciones de microaerofilia requeridas para el crecimiento de las especies de este género.

85
Posters
EVALUACIÓN DE DIFERENTES PRUEBAS SEROLÓGICAS PARA EL DIAGNÓSTICO DE LA BRUCELOSIS DE LOS PEQUEÑOS RUMIANTES EN ANIMALES NATURALMENTE INFECTADOS.
Ignacio García-Bocanegra1*; J.J. Pérez2; A. Arenas1; A. Perea1; A. Pacios3; A. Carbonero1; C. Borge1; M. A. González2
1Departamento de Sanidad Animal. Universidad de Córdoba.2Laboratorio de Producción y Sanidad Animal de Sevilla. Junta de Andalucía.
3Servicio de Sanidad Animal. Consejería de Agricultura y Pesca. Junta de Andalucía.
La brucelosis es una enfermedad de distribución mundial causada por bacterias del género Brucella, microorganismos capaces de infectar a humanos y a diferentes especies de animales domésticos y silvestres. El aislamiento microbiológico de Brucella spp. es considerado la técnica “gold standard” para el diagnóstico de la enfermedad. Así mismo, el test de rosa de bengala (TRB) y el test de fijación del complemento (TFC) son las únicas técnicas serológicas de referencia empleadas en el diagnóstico de la brucelosis en pequeños rumiantes en los programas de lucha (Council Directive 91/68/EEC, OIE, 2004).
En el presente trabajo se evaluó la sensibilidad (SE) y especificidad (ES) del TRB y cuatro test inmu-noenzimáticos comerciales (dos ELISA indirectos, un ELISA de bloqueo y un ELISA de competición) para el diagnóstico de B. melitensis en pequeños rumiantes.
Para el estudio se emplearon 81 sueros controles negativos, obtenidos de una región considerada oficialmente indemne de brucelosis (OIE, 2009) y 80 muestras controles positivos procedentes de animales naturalmente infectados previamente confirmados mediante cultivo y CFT. El punto de corte (cut-off) óptimo se estableció como la suma de los valores máximos de SE y ES obtenidos a partir del área bajo la curva. Los valores de concordancia, kappa (k), para cada test fueron igualmente calculados.
Empleando los cut-off propuestos por los fabricantes, todas las técnicas presentaron valores de SE y ES por encima del 95%, con la excepción de un ELISA indirecto. El ajuste de los valores de cut-off per-mitió optimizar los valores intrínsecos de los diferentes ELISAs en función de las diferentes condiciones epidemiológicas. Con excepción de un ELISA indirecto (k = 0.76), los ELISA comerciales presentaron una elevada concordancia (k ≥ 0.95). Los resultados obtenidos indican que las técnicas ELISA podrían ser empleadas de forma ventajosa en el diagnóstico oficial de la brucelosis en pequeños rumiantes.

86
Posters
CONCENTRACIONES DE HEMOGLOBINA, HIERRO Y FERRITINA EN YEGUAS PURA RAZA
ESPAÑOLA DURANTE EL CICLO ESTRAL
Katy Satué Ambrojo, Antonio Calvo Departamento de Medicina y Cirugía Animal. Universidad CEU-Cardenal Herrera (Valencia)
Durante el ciclo estral en diversas especies como la mujer y la perra, se producen modificaciones substanciales en el volumen sanguíneo, relacionadas con pérdidas de sangre. Dichos cambios se aso-cian a su vez con variaciones en los marcadores del estado férrico. El objetivo de la presente investi-gación ha sido analizar las concentraciones de hemoglobina (HB), hierro (Fe) y ferritina (Ferr) en yeguas PRE durante el ciclo estral. Se tomaron muestras de sangre venosa a 20 yeguas PRE reproductoras sanas durante ambas fases folicular (FF) y luteínica (FL) del ciclo estral. Las concentraciones de HB se determinaron con un analizador semiautomático Sysmex F-820. Las determinaciones de Fe fueron realizadas mediante espectrofotometría (espectrofotómetro Metrolab 2300 Plus V3®). Los niveles de Ferr se analizaron mediante turbidimetría utilizando reactivos de Spinreact®, siendo el coeficiente de variación intraensayo del 5.0%. El análisis de variación no mostró diferencias estadísticamente signi-ficativas en las concentraciones de HB (FF: 12,4±1,6; FL: 12,4±1,3 g / dl), Fe (FF: 164,3 ± 32,3; FL: 183.7 ± 47.5 mg / dl) y Ferr (FF: 161,4 ± 77,2; FL: 156.7 ± 77.9 mg / dl) entre ambas fases del ciclo estral. A nivel diagnóstico, el análisis de los niveles de hemoglobina, hierro y ferritina en la yegua PRE, no justifica el muestreo o la toma de decisiones clínicas en momentos concretos del ciclo estral.

87
Posters
DESARROLLO Y VALIDACIÓN DE UN ELISA PARA LA DETECCIÓN DE ANTICUERPOS
FRENTE A MYCOBACTERIUM BOVIS EN CIERVO. ESTUDIO COMPARADO DE DIFERENTES
PREPARACIONES ANTIGÉNICAS.
M. Barral1, O. Aurtenetxe2, N. Berano3, C. Gortázar4, M. Singh5, JM Garrido1, RA Juste1
1- NEIKER-Tecnalia. Derio (Bizkaia) [email protected]. 2- Hospital de Cruces. Barakaldo (Bizkaia).
3- Fraisoro laboratorio agroambiental Zizurkil (Gipuzkoa). 4- IREC. Ciudad Real.
5- LIONEX GmbH. Germany.
La tuberculosis causada por Mycobacterium bovis (bTB) es una enfermedad de distribución mundial que afecta a las personas, al ganado doméstico y a las especies silvestres. El diagnóstico de la bTB en las especies silvestres se hace habitualmente mediante el estudio post mortem de muestras de tejido, sin embargo cada vez con más frecuencia es necesario llevar a cabo el diagnóstico in vivo. En este trabajo se desarrolla un ELISA indirecto para la detección de anticuerpos frente a bTB en ciervos. Se emplearon como controles sueros de 4 ciervos mantenidos en cautividad que se inmunizaron con M. bovis, M. paratuberculsosis y M. avium inactivados. Se evaluaron los antígenos PPDb, ESAT 6, MPB70, MPB83 y CFP10 de forma individual o en grupos y la reacción se optimizó empleando diferentes diluciones de suero y de conjugado, buscando las combinación es que ofrecían los mejores resultados en los sueros control. El ELISA puesto a punto se validó con 120 sueros obtenidos de ciervos en los que se había confirmado la presencia o ausencia de bTB mediante cultivo y estudio lesional. Se obtuvo una sensibilidad del 76,19% y una especificidad del 86,87% para el punto de corte seleccionado, usando una combinación de MPB70, MPB83 y CFP10, dilución 1/200 de suero y 0,05 µg/ml de Proteina G. El ELISA desarrollado podría tener un importante potencial en el diagnóstico in vivo de la tuberculosis en ciervos tanto si se usa sólo como en combinación con otras técnicas que cuantifican la inmunidad celular.

88
Posters
APLICACIÓN DE TÉCNICAS DE LABORATORIO VETERINARIAS A LA INVESTIGACIÓN
BIO-MÉDICA: CARACTERIZACIÓN DE UNA NUEVA VARIANTE DE ENCEFALOPATÍA
ESPONGIFORME HUMANA
Rodríguez-Martínez AB1, Garrido JM1, Zarranz JJ2, Arteagoitia JM3, M de Pancorbo M4, Atarés B5, Bilbao MJ6, Ferrer I7, Juste RA1
1 Departamento de Sanidad Animal, Neiker-Tecnalia, Berreaga 1, 48160 Derio, Bizkaia2 Servicio de Neurologia, Hospital de Cruces, Plaza Cruces-gurutzeta 12, 48902 Barakaldo, Bizkaia3 Departamento de Sanidad y Consumo, Gobierno Vasco, San Sebastian-Donostia Kalea 1, 01010
Vitoria-Gasteiz, Alava4 Departamento deZoologia y Biologia Celular, Paseo Universidad 7, Universidad del País Vasco,
01006 Vitoria-Gasteiz, Alava5 Servicio de Anatomía Patológica, Hospital de Txagorritxu, José Achótegui s/n, 01009 Vitoria-
Gasteiz, Alava6 Servicio de Neurologia, Hospital de Mendaro, Mendarozabal s/n, 20850 Mendaro, Guipúzcoa
7 Institut de Neuropatologia, Servei Anatomia Patològica, IDIBELL-Hospital Universitari de Bellvitge, Carrer Feixa Llarga s/n, 08907 Hospitalet de Llobregat, Barcelona
El diagnóstico de las EE. EE. TT implica la detección y caracterización de su agente causal, el prión o proteína priónica patógena (PrPSc). En las formas animales, el diagnóstico se basa en la detección de PrPSc mediante tests de pruebas rápidas, mientras que en las formas humanas el diagnóstico se realiza en base a criterios clínicos, neuropatológicos y moleculares. Los protocolos utilizados para el tipado de la proteína patógena incluyen el tratamiento con proteasas, cuya función es destruir total-mente la proteína priónica celular normal, e hidrolizar parcialmente la PrPSc que presenta un núcleo resistente a degradación denominado PrP27-30. Algunas enfermedades, como el Scrapie Atípico o la recientemente descrita Prionopatía Sensible a Proteasas, tienen la característica de presentar una proteína patógena con una mayor sensibilidad a degradación con proteasas. En estos casos, el tratamiento con PK implica la destrucción de una elevada fracción de PrPSc, suponiendo incluso la ausencia de señal tras el análisis mediante inmunoblot, que en consecuencia, puede conducir a un incorrecto diagnóstico. En este contexto, el uso de herramientas diagnósticas basadas en la captura de PrPSc sin necesidad de tratamiento con PK, así como en el empleo de condiciones de digestión con PK menos agresivas podría proporcionar mayor sensibilidad de detección y tipado de la proteína patógena. Por tanto, el objetivo de ese trabajo es exponer un ejemplo del uso de herramientas diag-nósticas con estas condiciones utilizadas en el diagnóstico de EE.EE.TT animales aplicadas al análisis del primer caso de Prionopatía Sensible a Proteasas 129MV descrito en España.



indice


93
indice
Presentación:• Presentación ........................................................................................................................... 7
Ponencias:• Caracterización de cepas de Coxiella circulantes en España. Dr. Pedro Anda (Instituto Carlos III. Madrid) ............................................................................ 13
• Fiebre Q: Estudios de campo, diagnóstico de laboratorio y control. Dr. Ana L. García-Perez (Neiker, Bilbao) .................................................................................. 15
• Fiebre Q en Holanda.(Inglés/Español) Dr. Douwe Bakker (Central Veterinary Institute, Lelystad, Holanda) .......................................... 25
• La peste de los pequeños rumiantes y la fiebre/encefalitis por virus West Nile: diagnóstico actual y perspectivas de futuro. Dr. Miguel Angel Jiménez Clavero (CISA, Valdeolomos, Madrid) .............................................. 33
• Técnicas de diagnóstico de Besnoitia en el laboratorio. Dr. José Antonio Castillo (Facultad de Veterinaria de Zaragoza)............................................... 35
• Dípteros vectores de enfermedades: biología e identificación. Dr. Javier Lucientes (Facultad de Veterinaria de Zaragoza) ...................................................... 39
• Diagnóstico de enfermedades emergentes en porcino Dr. Joaquín Segalés (CRESA, Barcelona) ................................................................................ 43
• Recogida de muestras para análisis laboratorial en medio exótico: Expedición española a la Antártida 2010. Dr. F.García Peña (LCV, Algete) ............................................................................................... 49
Comunicaciones orales:• Evaluación de la interferencia de la vacunación a paratuberculosis en el diagnóstico de tuberculosis en cabras infectadas experimentalmente con Mycobacterium caprae. ........... 61
• Prevalencia de Crypstosporidium y Giardia en bovinos de la provincia de Alava ...................... 62
• Influencia de factores genéticos y ambientales en la biodiversidad y predominancia de Clostridios en la flora intestinal equina................................................................................ 63
• Desarrollo de un ELISA de doble reconocimiento para la detección de anticuerpos específicos del virus de la Fiebre del Valle del Rift. ............................................... 64
• Diagnóstico serológico de Salmonella spp.: consideraciones antes de su uso como herramienta de clasificación en futuros programas de control en porcino. ..................... 65
• Desarrollo de un ELISA de bloqueo para la detección de anticuerpos específicos del virus del Nilo Occidental. ................................................................................................... 66
• Estudio de la biodiversidad de la flora fecal del ganso común: influencia de la migración. ....... 67

94
indice
• Aislamiento e identificación de Besnoitia besnoiti en cultivos celulares a partir de muestras clínicas. .................................................................................................................. 68
• Estudio de la evolución de anticuerpos de Coxiella burnetti en un rebaño de vacas lecheras de alta producción en el noreste de España durante un año. .......................... 69
• Valoración de tres ELISA´s de detección de anticuerpos en suero frente a M.hyopneumoniae. .................................................................................................... 70
• Primera detección de Babesia occultans en garrapatas del género Hyalomma procedentes de Tunez. ......................................................................................... 71
• Ensayo colaborativo de un protocolo de ELISA de tuberculosis bovina en suidos. .................. 72
• Validation of commercial ELISA kits for detection of anti-BTV antibodies in naturally-infected and vaccinated animals ............................................................................... 73
Posters:• Distribución geográfica de Culicoides (Diptera: Ceratopogonidae) en la Comunidad Valenciana. .................................................................................................. 77
• Detección de proteina prión patológica en placentas de ovino como método de diagnóstico de Scrapie. ........................................................................................ 78
• Prevalencia de micotoxinas en materias primas. Datos zona de Aragón 2008. Análisis realizados por ELISA y HPLC ..................................................................................... 79
• Micotoxinas en piensos analizados en nuestro laboratorio durante año 2008. Análisis realizados por ELISA. ................................................................................................. 80
• Prevalencia de Campylobacter spp. en broilers a sacrificio desde 1988 a 2009. ............................................................................................................... 81
• Estudio de los niveles de anticuerpos en muestra de leche de tanque en brotes de BVD. ...................................................................................................... 82
• Caracterización de cepas Escherichia Coli productoras de beta-lactamasas (BLEEs) de espectro extendido en ganado vacuno de Galicia. ................................................ 83
• Optimización de un método para realizar PCR a partir de muestras congeladas de cepas del género Campylobacter en medios semisólidos. ................................................. 84
• Evaluación de diferentes pruebas serológicas para el diagnóstico de la brucelosis de los pequeños rumiantes en animales naturalmente infectados. ................. 85
• Concentraciones de hemoglobina, hierro y ferritina en yeguas pura raza española durante el ciclo estral. .............................................................................................. 86
• Desarrollo y validación de un ELISA para la detección de anticuerpos frente a Mycobacterium bovis en ciervos. Estudio comparado de diferentes preparaciones antigénicas. ...................... 87
• Aplicación de técnicas de laboratorio veterinarias a la investigación bio-médica: Caracterización de una nueva variante de encefalopatía espongiforme humana. .................... 88

Patrocinadores
Colaboradores
Organizadores
Consejo Generalde Colegios Veterinarios
XV SIMPOSIO AVEDILA