reconstructing euglenoid evolutionary relationships using three genes: nuclear ssu and lsu, and...
TRANSCRIPT

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Protist, Vol. 161, 603–619, October 2010http://www.elsevier.de/protisPublished online date 22 April 2010
ORIGINAL PAPER
Reconstructing Euglenoid EvolutionaryRelationships using Three Genes: Nuclear SSUand LSU, and Chloroplast SSU rDNA Sequencesand the Description of Euglenaria gen. nov.(Euglenophyta)
Eric W. Lintona,1, Anna Karnkowska-Ishikawac, Jong Im Kimd, Woongghi Shind,Mathew S. Bennettb, Jan Kwiatowskic,e, Bozena Zakrysc, and Richard E. Triemerb
aCentral Michigan University, Department of Biology, Mt. Pleasant, MI 48859, USAbMichigan State University, Department of Plant Biology, East Lansing, MI 48824, USAcUniversity of Warsaw, Department of Plant Systematics and Geography, Institute of Botany, Al. Ujazdowskie 4,00-478 Warsaw, Poland
dChungnam National University, Department of Biology, 220 Gung-Dong, Yuseong-Gu Daejeon 305-764,Republic of Korea
eUniversity of California, Department of Ecology and Evolutionary Biology, 453 Steinhaus Hall, Irvine,CA 92697, USA
Submitted September 24, 2009; Accepted February 28, 2010Monitoring Editor: Michael Melkonian
Using Maximum Likelihood and Bayesian analyses of three genes, nuclear SSU (nSSU) andLSU (nLSU) rDNA, and chloroplast SSU (cpSSU) rDNA, the relationships among 82 plastid-containingstrains of euglenophytes were clarified. The resulting tree split into two major clades: cladeone contained Euglena, Trachelomonas, Strombomonas, Colacium, Monomorphina, Cryptoglenaand Euglenaria; clade two contained Lepocinclis, Phacus and Discoplastis. The majority of themembers of Euglena were contained in clade A, but seven members were outside of thisclade. Euglena limnophila grouped with, and was thus transferred to Phacus. Euglena proxima wasa single taxon at the base of clade one and is unassociated with any subclade. Five members ofEuglena grouped together within clade one and were transferred into the newly erected genusEuglenaria. The monophyly of the remaining genera was supported by Bayesian and MaximumLikelihood analyses. Combining datasets resolved the relationships among ten genera ofphotosynthetic euglenoids.& 2010 Elsevier GmbH. All rights reserved.
Key words: Bayesian inference; euglenophyta; LSU; maximum likelihood; SSU.
Introduction
Euglenoids are a group of protozoans that mostcommonly have two flagella, one or both may beemergent. Members live in salt, brackish or fresh
1Corresponding author; fax þ1 11 989 772 3462.e-mail [email protected] (E.W. Linton).
& 2010 Elsevier GmbH. All rights reserved.doi:10.1016/j.protis.2010.02.002

Author's personal copy
water environments. Many are phagotrophic orosmotrophic, however, some euglenoids are alsoconsidered algae because they are photosyn-thetic, having acquired a chloroplast via second-ary endosymbiosis of a eukaryotic green alga(Gibbs 1978; Ishida et al. 1997; Leander 2004;Martin et al. 1992; Nozaki 2005). Our research isfocused on phylogenetic (evolutionary) relation-ships among the photosynthetic euglenoids with asingle emergent flagellum, while members of theoutgroup have two emergent flagella. Because oftheir long and intricate history, a brief review ofeuglenoid morphology and taxonomy is war-ranted. An in-depth review of euglenoid classifica-tion and molecular phylogenetics can be found inTriemer and Farmer (2007).
Since the genus Euglena was first described byEhrenberg (1830a, 1830b), their classification byvarious authors has largely been based onmorphological features such as: chloroplast typeand distribution, flagellar length, shape and dis-tribution of the storage product, and cell surfacefeatures. Therefore, an understanding of eugle-noid morphological features is necessary for theirproper classification into genera.
The unique surface feature of all euglenoids isthe pellicle, a series of interlocking protein stripssubtended by microtubules that lies below theplasma membrane. If the cell is rigid or capable ofa slight bending motion, the pellicle strips can bearranged either longitudinally or helically. How-ever, all cells capable of metaboly, a peristalticwriggling motion (Triemer et al. 2006), havehelically arranged pellicle strips. Underlying thepellicle strips of some euglenoids are membrane-bounded mucus secreting bodies known asmucocysts, sensu Mignot (1966). Mucocysts,when present, can be round or spindle-shaped,thus providing a diagnostic character for manyspecies.
An exceedingly useful phylogenetic character ofall photosynthetic euglenoids is the chloroplast.The chloroplast envelope consists of three mem-branes, indicating their secondary endosymbioticorigin, while the inner photosynthetic membranes(thylakoids) occur in stacks of three and lack thegrana stacks found in land plants. The chloroplastcan be absent, reduced to a colorless plastid insome secondary osmotrophic euglenoids, orcontain chlorophyll and take on a variety ofshapes, such as discoid, lenticular, lobed, sphe-rical, ribbon-like or stellate. While the samechloroplast shape can be found in a variety ofdifferent euglenoid species, the shape for anyparticular species is specific and in some cases
specific for the whole genus. The pyrenoid, aproteinaceous area within the chloroplast contain-ing Rubisco, is associated with the formation ofthe storage products. In euglenoids this product isparamylon a beta 1,3-glucan storage compound.This structure, like the chloroplast, is oftendiagnostic of a species or an entire genus ofeuglenoids. When present, the pyrenoid may becapped by paramylon on one side (haplopyre-noids), both sides (diplopyrenoid), or lacking aparamylon cap (naked). Additional morphologicalfeatures such as cell length, shape, degree ofcompression (flatness), presence and shape of thelorica (a protective extracellular matrix that sur-rounds cell) have also been used to define speciesand genera of euglenoids.
By 1849, taxonomists had erected six of theten photosynthetic euglenoid genera analyzedin this paper. The first six genera were Euglena(Ehrenberg 1830a, 1830b, 1838), Cryptoglena(Ehrenberg 1831), Trachelomonas and Colacium(Ehrenberg 1833), Phacus (Dujardin 1841) andLepocinclis (Perty 1849). The remaining fourgenera were separated from these original generabased on more detailed examinations of morpho-logical features or on significant molecular differ-ences. Using morphological differences Mono-morphina was split off from Lepocinclis byMereschkowsky (1877), but the description ofthe genus was not recognized in the Englishliterature until recently (Marin et al. 2003). Differ-ences in the structure of the lorica were used byDeflandre (1930) to erect Strombomonas from asubgroup of Trachelomonas. The use of moleculardata, predominantly nuclear small subunit rDNA(nSSU), and modern phylogenetic methods (Max-imum Likelihood and Bayesian analyses) usheredin a new era of euglenoid systematics.
In this new era, early molecular evidencenoted that some genera (Euglena, Phacus andLepocinclis) were not monophyletic (Linton et al.1999, 2000; Milanowski et al. 2001; Mullner et al.2001) which led to several taxonomic revisions. In2003, Marin et al. resurrected Monomorphina andmoved several former Phacus and a few Lepocin-clis species into the genus thus making bothgenera monophyletic. In addition, several Euglenaspecies that were nested with taxa from Lepocin-clis were moved into this genus. Strombomonaswas then dissolved back into Trachelomonas.More recently, Triemer et al. (2006) usingincreased taxon sampling and data from multiplegenes, re-established Trachelomonas and Strom-bomonas as independent genera and erected thenew genus Discoplastis to contain two species
E.W. Linton et al.604

Author's personal copy
formerly assigned to Euglena. Our study uses 82taxa and an expanded dataset of three genes, twonuclear (nSSU and nLSU rDNA) and one plastid(cpSSU rDNA), to resolve relationships and defineten genera, nine previously described and onenewly erected genus Euglenaria, containing fiveformer members of Euglena.
Results
Phylogenetic Analysis
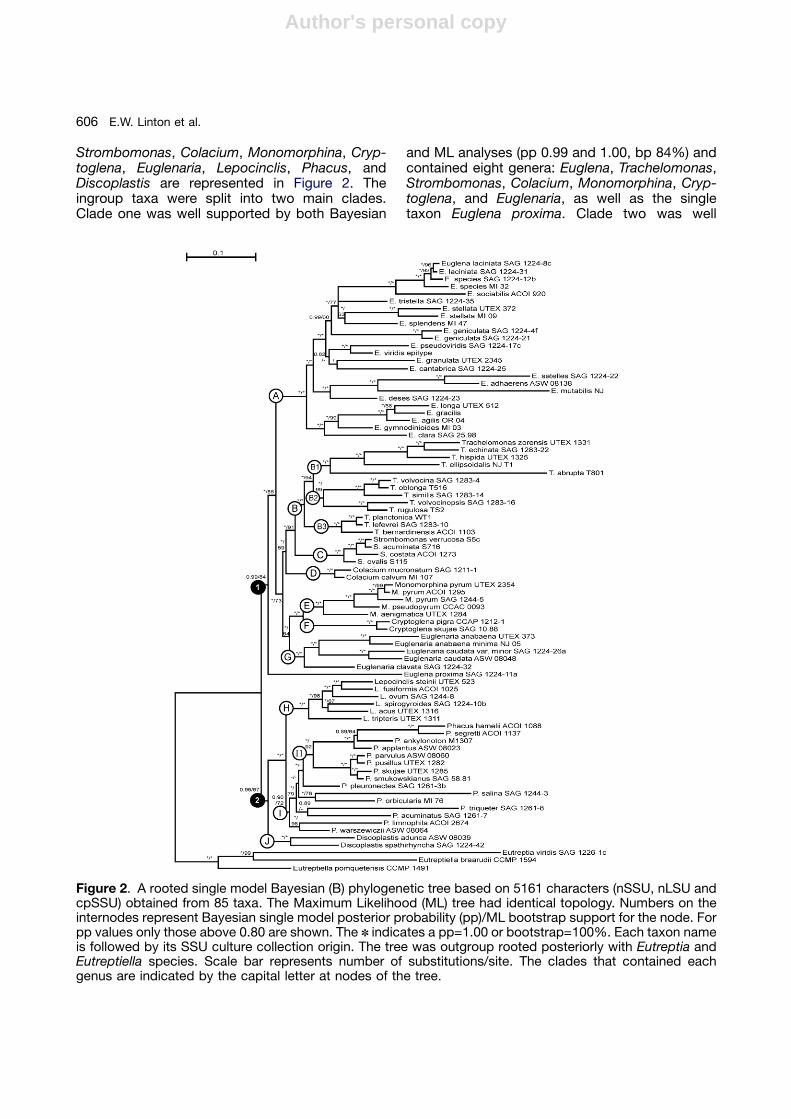
Eighty-two taxa representing 12 genera (10ingroup and two outgroup) were analyzed usinga combination of three genes. Two (nSSU andnLSU) were nuclear encoded and one (cpSSU)was encoded in the chloroplast or plastid (Euglenalonga) for a total of 5161 bases used in theanalysis (nSSU: 1740, nLSU: 2110 and cpSSU:1311). Of the 246 sequences used in this study 92were previously unpublished (nSSU: 12, nLSU: 22and cpSSU: 58). Bayesian (single and partitionmodel) and Maximum Likelihood (ML) analysesrecovered trees of identical topology (Fig. 2),except for the unresolved position of Euglenatristella, see below. The tree shows only singlemodel Bayesian support (pp) values then the MLsupport (bp) values. When the Bayesian supportvalues differ between the single and partitionmodels, the single model values are given in thetext first, followed by the partition model values.
The backbone of this multi-gene tree was wellsupported by both Bayesian posterior probabil-ities (pp) and ML bootstrap numbers (bp). Withinthe ten ingroup genera ten separate clades (A–J),as well as a sister relationship of the single taxonEuglena proxima within the clades (A–G), were allwell supported. Each genus formed a single majorclade except for Euglena that formed two distinctwell-supported clades (A and G) and the singletaxon Euglena proxima making Euglena a poly-phyletic genus (Fig. 2). Consequently, Euglenariawas erected to contain the taxa of clade G.
Analysis of each dataset individually (nSSU,nLSU and cpSSU) each resulted in less well-resolved and supported topologies (not shown)with separate clades for each genus, as in thecombined tree. The nSSU tree was the mostsimilar to the combined tree and differed only inplacing the Euglenaria clade as sister to theEuglena clade (0.92 pp). However, the nLSU andcpSSU each supported the separation of Eugle-naria from Euglena, as did a combined nSSU andnLSU dataset (not shown). The combined nSSU/nLSU dataset resulted in a well-supported (pp)tree of identical topology to the three gene treepresented in this study (Fig. 2).
Overview
The phylogenetic relationships among the fresh-water euglenoid genera Euglena, Trachelomonas,
Figure 1. (A) Alignment of the nSSU sequences between helix 18 and 180. Euglenaria caudata and Euglenaviridis (representing all other euglenoids) are indicated with the base C among all members of the new genusEuglenaria and the base A among all other euglenoids used in this analysis, with the exception of Eutreptiaviridis which has a G in this position (not shown). (B) Alignment of the nSSU sequences between helix 37 and38. Euglenaria caudata and Euglena viridis (representing the genus Euglena) are indicated with the base Tamong all members of the new genus Euglenaria and the base A or C (not shown) among all members of thegenus Euglena used in this analysis. The diagnostic bases are in bold. (C) Alignment of nSSU sequences ofstem 17 between Euglena proxima and Euglena viridis (representing all other euglenoids). Euglena proximahad a unique CGC\CCG sequence while most other euglenoids had a CAA\TTG sequence. The bases in thestem of helix 17 are in bold.
605Euglenoid Evolution using Three Genes
Author's personal copy
Strombomonas, Colacium, Monomorphina, Cryp-toglena, Euglenaria, Lepocinclis, Phacus, andDiscoplastis are represented in Figure 2. Theingroup taxa were split into two main clades.Clade one was well supported by both Bayesian
and ML analyses (pp 0.99 and 1.00, bp 84%) andcontained eight genera: Euglena, Trachelomonas,Strombomonas, Colacium, Monomorphina, Cryp-toglena, and Euglenaria, as well as the singletaxon Euglena proxima. Clade two was well
Figure 2. A rooted single model Bayesian (B) phylogenetic tree based on 5161 characters (nSSU, nLSU andcpSSU) obtained from 85 taxa. The Maximum Likelihood (ML) tree had identical topology. Numbers on theinternodes represent Bayesian single model posterior probability (pp)/ML bootstrap support for the node. Forpp values only those above 0.80 are shown. The n indicates a pp=1.00 or bootstrap=100%. Each taxon nameis followed by its SSU culture collection origin. The tree was outgroup rooted posteriorly with Eutreptia andEutreptiella species. Scale bar represents number of substitutions/site. The clades that contained eachgenus are indicated by the capital letter at nodes of the tree.
E.W. Linton et al.606

Author's personal copy
supported (pp 0.96 and 0.99) by Bayesian, butweakly supported (bp 67%) by ML analysis,contained three genera: Lepocinclis, Phacus, andDiscoplastis.
Clades A–G
Euglena: After the transfer of five taxa to thenew genus Euglenaria (clade G), Euglena remainsparaphyletic. Euglena proxima was well supportedas an independent lineage from the crownEuglena clade A by Bayesian and ML analyses(pp 0.99 and 1.00, bp 84%) on its own branch.Euglena proxima lacked the diagnostic base C ofEuglenaria and instead had the base A as found inall the remaining ingroup taxa. Still, E. proxima hada unique molecular signature in the nSSU at thebase of stem 17 CGC\CCG where all but eighttaxa on the tree had CAA\TTG (Fig. 1C). The eighttaxa (seven ingroup and one outgroup) hadvarious differences in this stem area, but nonewere like those found in E. proxima.
The crown clade A (pp 1.00, bp 100%) containedall of the remaining members of Euglena includingthe type species, Euglena viridis. Nearly all sisterrelationships within clade A were well supportedby Bayesian posterior probabilities (0.96–1.00,0.99–1.00) with the majority supported by 1.00.The exceptions were the position of E. tristella assister to E. splendens weakly supported by ML(51%) but well supported by single Bayesian (0.96),but weakly supported as sister to E. sociabilisby partition Bayesian (0.61). Therefore, its finalposition is represented as an unresolvedtrichotomy. The position of E. splendens as sisterto E. stellata was well supported by Bayesian butonly weakly supported by ML (pp 1.00 and 0.99, bp63%). Neither Bayesian nor ML fully supported theposition of E. cantabrica and E. granulata as sisters(pp 0.65 and 0.83, bpo50%) or their sisterrelationships to the E. viridis, E. pseudoviridisclade (pp 0.82 and 0.85, bpo50%).
Trachelomonas/Strombomonas/Colacium:These sister clades (B, C and D) were connectedby a short branch that was well supported byBayesian analysis (pp 1.00 and 0.99), but onlyweakly supported by ML analysis (bp 59%). Allmembers of these three clades produced profuseamounts of mucilage that was retained on the cellsurface as either a lorica (Trachelomonas andStrombomonas) or a stalk (Colacium).
Relationships among members of Trachelomo-nas, a specious genus that formed various sizeand shaped loricas with spines and/or pores, were
all well supported (pp 1.00, bp 94–100%) andcontained within the single clade B (pp 1.00, bp100%). Clade B divided into three well supportedsister clades B1 (pp 1.00, bp 100%) B2 (pp 1.00,bp 99%) and B3 (pp 1.00, bp 100%), whichdiffered based on the morphology of their loricas.
All Strombomonas were contained in clade C(pp 1.00, bp 100%) and all sister relationshipsamong species were well supported (pp 1.00, bp100%). The sister relationship between Strombo-monas (clade C) and Trachelomonas (clade B) waswell supported (pp 1.00, bp 91%) as well.Colacium contained six described species withtwo present in the analysis. These two membersformed a single clade D (pp 1.00, bp 100%).
Monomorphina/Cryptoglena: The relationshipbetween these two sister clades was well sup-ported by both Bayesian and ML analysis (pp 1.00,bp 100%). Five members of Monomorphina werecontained within clade E (pp 1.00, bp 100%) withstrongly supported sister relationships (pp 1.00,bp 99–100%). The two members of Cryptoglenawere contained within clade F (pp 1.00, bp 100%).
Euglenaria: This newly erected genus containedfive taxa previously in Euglena (clade A). Molecularanalysis has shown that these taxa formed astrongly supported (pp 1.00, bp 100%) clade (G)separate from other Euglena species in cladeA and the single taxon Euglena proxima. The sisterrelationships among the members of thisnew genus were equally well supported (pp 1.00,bp 100%). The relationship of Euglenaria (clade G)to Monomorphina (clade E) and Cryptoglena (cladeF) was well supported by Bayesian (pp 1.00), butweakly supported by ML (bp 64%) analyses.
Clades H–J
Lepocinclis: Lepocinclis formed a single wellsupported clade H (pp 1.00, bp 100%). The sisterrelationships among the six species within thisclade were all strongly supported (pp 1.00, bp100%), except for the sister relationship betweenL. spirogyroides and L. acus that had only weakML support (bp 67%). The sister relationshipbetween Lepocinclis (clade H) and Phacus(clade I) was well supported (pp 1.00, bp 100%).
Phacus: Clade I was well supported by the singleBayesian model (pp 0.96) but had the weakestsupport of all the major clades (pp 0.82, bp 72%)from partition Bayesian and ML analyses. This cladecontained 15 members of Phacus, which includedthe new member P. limnophila. The placement of
607Euglenoid Evolution using Three Genes

Author's personal copy
P. limnophila, was intriguing because of its spindle-shaped morphology, but was well supported (pp1.00, bp 96%) as a sister to P. warszewiczii (cellstriangular in apical view). Many of the branches inclade I were short with Bayesian and ML supportranging from no support (ppo0.90, bpo50%) to wellsupported (pp 1.00, bp 79%). The Phacus in clade I1were strongly supported (pp 1.00, bp 92%) as agroup, as were their sister relationships (pp 1.00, bp100%), except for P. ankylonoton (pp 0.89 and 0.84,bp 64%).
Discoplastis: This genus diverges prior to theLepocinclis and Phacus clades with strong Bayesian,but weak ML support (pp 0.96 and 0.99, bp 67%).The two members of this genus were well supported(pp 1.00, bp 100%) in clade J as sister taxa.
Taxonomic Revisions
Euglenaria Karnkowska, Linton et Kwiatowski,gen. nov.
Cellulae libere viventes, solitariae, cum unoflagello emergenti ubi natans, claviformes, fusi-formes vel cylindrico-fusiformes, metabolicae,angustatae postice, gradatim decrescentes incaudam acutam; chloroplasti parietales, lobati,pyrenoidum singulum continentes; pilei paramyla-cei utrinque chloroplasto insidentes; mucocystaenullae.Diagnosis
Cells free-living, solitary with one emergentflagellum when swimming; club-shaped, fusiformor cylindrically-fusiform, narrowing to the posteriorand tapering into a pointed tail-piece; metabolic,display euglenoid movement; parietal, lobedchloroplasts, each with a single pyrenoid accom-panied by bilateral paramylon caps located onboth sides of the chloroplast (diplopyrenoid);mucocysts absent.
Type species:Euglenaria caudata (Hubner) Karnkowska et
Linton, comb. nov.Basionym:Euglena caudata 1886. Hubner, E.F.W. Eugle-
naceen-Flora von Stralsund. Program Realgym-nasiums zu Stralsund. p. 13, Fig. 15.
Euglenaria caudata var. minor (Deflandre) Karn-kowska et Linton, comb. nov.
Basonym:Euglena caudata var. minor Deflandre, Bull. Soc.
Bot. de France. 71: 1119, Fig. 7, 1924.Euglenaria clavata (Skuja) Karnkowska et Linton,
comb. nov.
Basionym:Euglena clavata Skuja 1948. Taxonomie des
phytoplantons einigen Seen in Uppsala, Schwe-den. Symbolae Botanicae Upsaliensis 9(3): 189,190, pl. 22, Figs. 2–5.
Euglenaria anabaena (Mainx) Karnkowska etLinton, comb. nov.
Basionym:Euglena anabaena 1926. Mainx, F. Einige
neue Vertreter der Gattung Euglena Ehrenbergmit Unterstutzung der Gesellschaft zur Forderungdeutschen Wissenschaft, Kunst und Literaturin Bohmen. Arch. Protistenkd 54: 160-161,Fig. Dc.
Euglenaria anabaena var minima (Mainx)Karnkowska et Linton, comb. nov.
Basionym:Euglena anabaena var minima 1927. Mainx, F.
Beitrage zur Morphologie und Physiologie derEuglenen. I. Morphologische Beobachtung, Meth-ode, und Erfolge der Reinkultur. II. Untersuchunguber die Ernahrungs- und Reizphysiologie. Arch.Protistenkd 60:334, pl. 10, Figs. 3, 4.
The name, Euglenaria, was derived from thegenus Euglena and the Greek, ‘‘-aria’’ n. meaninglike. Thus, these taxa were morphologically indis-tinguishable from members of Euglena, but weregenetically distinct based on phylogeneticanalysis. Moreover, in the nSSU alignment allmembers of Euglenaria had a C base, four basesupstream of helix 180, where all other ingroupeuglenoids and the two Eutreptiella species havean A, while Eutreptia viridis has a G (Fig. 1A).Furthermore, in the nSSU alignment all membersof Euglenaria had a T, one base upstream of helix38, while all members of Euglena had either an Aor a C (Fig. 1B). The T base at this position wasfound in all other ingroup taxa except for Phacushamelii and P. segretti, which had the C base. Thebase C at this position was also found in all threeoutgroup taxa suggesting that C is the primitive(plesiomorphic) state. Together, the C baseupstream of helix 180 and the T base upstreamof helix 38 separate the Euglenaria from membersof Euglena and the other ingroup euglenoids.
PhacusThe genus Phacus was created and diagnosed
by Dujardin (1841) as rigid, free-swimming cellsthat were more or less flattened. Recently Marinet al. (2003) emended this diagnoses to moreaccurately define Phacus as a genus. However,due to Lepocinclis salina and Euglena limnophilabeing transferred into Phacus (see below), anemended diagnosis of the genus was againrequired.
E.W. Linton et al.608

Author's personal copy
Phacus Dujardin, Historie naturelle des Zoo-phytes-Infusoires. Paris. pp. 327, 334, 1841.Emend. Linton et Karnkowska
Phacus – Emended DiagnosisCells free-living, solitary with a single emergent
flagellum when swimming; usually laterally com-pressed (rarely not compressed or triangular in apicalview), spindle-shaped or ovoid; semi-rigid to rigid;numerous parietal disc-shaped chloroplasts of similarsize without pyrenoids; paramylon grains dimorphicin size, large ring-shaped, rod-shaped or discoidparamylon grains. Colorless forms known.
Phacus salina (Fritsch) Linton et Karnkowska,comb. nov.
Basionym:Lepocinclis salina 1914. Fritsch, F.E. Notes on
British Flagellates, I-IV. The New Phytologist, vol.13, Iss. 10. pg. 351, Fig. 3A, B.
Synonyms:Lepocinclis texta var. salina (Fritch) Popova,
Trudy Botanicheskogo Instituta Akademii NaukSSSR. 7: 301, pl. 11 Fig. 18, 1951; Euglena textavar. salina (Fritch) Popova, Euglenovyje vodorosli.Opredelitel prosnovodnych vodoroslej SSSR, 7.[Euglenophyta. The Handbook of FreshwaterAlgae]. pg. 159, Fig. 59, 1955.
Phacus limnophila (Lemmermann) Linton etKarnkowska, comb. nov.
Basionym:Euglena limnophila Lemmermann, Beitrage zur
Kenntnis der Planktonalgen. II. Beschreibung neuerFormen. Beihefte Botan. Centralbl. 76: pg 152 andLemmermann in Pascher’s Sussw. – Fl., 2: 130,Fig. 205, 1913.
Phacus was distinguished from Lepocinclisbased on phylogenetic analysis and moleculardifferences present in the nSSU alignmentbetween the two genera. The base T waspresent one base downstream of helix 15 andtwo bases downstream of helix 230 in all Lepocin-clis whereas all Phacus had the base C and A,respectively.
Discussion
Euglena
Past studies of euglenoid phylogenies have usedonly nuclear encoded genes. Most often this wasthe nuclear SSU rDNA (Busse and Preisfeld2002, 2003; Busse et al. 2003; Leander et al.2001; Linton et al. 1999, 2000; Marin et al. 2003;Preisfeld et al. 2000, 2001; Von der Heyden et al.2004) with one study (Talke and Preisfeld 2002)using the nuclear encoded flagellar PAR1 and
PAR2 genes, the results from which agreed withprevious nuclear SSU rDNA studies. The first non-nuclear gene study used chloroplast SSU rDNAsequence data from 17 photosynthetic species(Milanowski et al. 2001) and suggested polyphylyin Euglena, but the bootstrap support was weak.
More recent molecular studies have combined twogenes. The first by Milanowski et al. (2006) combinednuclear and chloroplast SSU rDNA sequence datafrom 35 ingroup and 2 outgroup taxa to conclude,among other things, that Euglena was a polyphyleticgenus with members present within four separateclades (A, BþCþDþ, E and G) of their tree. Clade Acontained most members of Euglena including thetype specimen (E. viridis), BþCþDþ containedEuglena anabaena, E contained E. proxima while Gcontained Euglena spathirhyncha and E. limnophila.The second study by Triemer et al. (2006) combinednuclear SSU and partial LSU rDNA sequence data toestablish a well resolved and supported phylogeny ofEuglenales (sensu Leedale). Their analysis of 84 taxasupported 9 ingroup genera and established the newgenus Discoplastis to contain D. spathirhyncha andD. adunca, two former members of Euglena. How-ever, their study did not include E. proxima or E.limnophila as in Milanowski et al. (2006), but it didinclude Euglena anabaena and three sister taxa thatformed an ‘‘anabaena’’ clade. This ‘‘anabaena’’ cladewas supported (pp 0.92, Fig. 1 Triemer et al. 2006) assister to the other members of Euglena. In summary,Discoplastis was erected to contain some formermembers of Euglena (Triemer et al. 2006) andestablish the monophyly of the genus. Unfortunately,Milanowski et al. (2006) showed that at least threeadditional members of Euglena (E. proxima, E.limnophila and the ‘‘anabaena’’ clade) would requireeither the erection of a new genus or the transfer toan existing genus to fully establish the monophyly ofEuglena. By combining a larger number of taxa andthree genes, this study has resolved two of the threeremaining taxonomic issues necessary to establishthe monophyly of Euglena.
This study has shown that other than mostmembers having a spindle shaped body, noderived (synapomorphic) morphological or mole-cular character capable of uniting members ofEuglena has yet been identified. However, someimportant characteristics of this genus wereidentified. The chloroplasts show more morpho-logical diversity than any other genus, rangingfrom absent to ribbon-like to lenticular to lobed ordiscoid with pyrenoids present in the chloroplastof some species but absent in others. Moreoverthe mucocysts, mucus secreting bodies below thepellicle strips, show great morphological diversity.
609Euglenoid Evolution using Three Genes

Author's personal copy
They may be absent or present, spherical orspindle in shape (Kosmala et al. 2009).
Because of the morphological diversity inEuglena, the species E. proxima (with chloroplaststhat are discoid and lack pyrenoids) cannot beseparated from Euglena (clade A) nor joined to theEuglenaria (clade G), because it lacks the diag-nostic nSSU bases (see diagnosis above). At thistime E. proxima must remain as a single unasso-ciated taxon until additional data (sequence and/or taxa) cause it to either clade with an existinggenus or other taxa clade with it allowing thediagnosis of a new genus.
Regrettably, despite the erection of Euglenaria(above) and the transfer of Euglena limnophila toPhacus the genus Euglena remains paraphyletic.Clade A contains the type Euglena viridis and thusretains the name Euglena.
Trachelomonas/Strombomonas/Colacium
A great deal of debate has occurred recently(Brosnan et al. 2003, 2005; Ciugulea et al. 2008;Marin et al. 2003; Moreira et al. 2001; Mullner et al.2001; Nudelman et al. 2003; Triemer et al. 2006)about the existence of Strombomonas (clade C)and whether to dissolve it back into Trachelomo-nas (clade B). While all of these molecular studiessupported the monophyly of the loricate forminggenera (Brosnan et al. 2003; Marin et al. 2003;Moreira et al. 2001; Mullner et al. 2001; Nudelmanet al. 2003) most failed to sustain Trachelomonasand Strombomonas as distinct genera (significantpp or bp support). All of these studies sufferedfrom limited taxa sampling, a weakness recog-nized by many of the authors (Brosnan et al. 2003;Moreira et al. 2001; Mullner et al. 2001; Nudelmanet al. 2003). Fortunately, the two most recentmolecular studies (Ciugulea et al. 2008; Triemeret al. 2006) used a larger number of taxa and twogenes (nuclear SSU/LSU rDNA), which showedsignificant support for maintaining these as dis-tinct genera.
The morphological diversity of the lorica hasbeen the chief diagnostic character essential forgenus and species identification within these twogenera. Our analysis has shown Trachelomonas tobe composed of three clades (B1, B2 and B3) thatcan be distinguished morphologically by the loricaand chloroplast. Species with spiny loricas, andlarge disc-shaped plastids with diplopyrenoids arecontained in clade B1, except for T. abrupta withinner projecting haplopyrenoids. Clade B2 con-tained species that are small and round with twoto five or more chloroplasts, five being the most
common, all with inner projecting haplopyrenoids.Clade B3 contained species that were oblong inshape with irregular surface ornamentation and upto ten chloroplasts, all with inner projectinghaplopyrenoids. Since all taxa in clades B2, B3and T. abrupta, the basal member of B1, containinner projecting haplopyrenoids; this would be theplesiomorphic state. These results are in agree-ment with the five Trachelomonas clades ofCiugulea et al. ((2008) A–E, Fig. 1).
The four members of Strombomonas formed asingle clade (C) that was well supported anddistinct from its sister Trachelomonas (clade B) inour study. The lorica lacks the distinctive spinesand pores associated with the loricas of Trache-lomonas. Although, our study contained only fourtaxa compared to the eight of Ciugulea et al.((2008) F–H, Fig. 1) with three taxa from their cladeF and one from their clade H (S. ovalis), our resultsare congruent with theirs. We expect future workinvolving molecular and morphological data toenable a much-needed revision of the currentTrachelomonas and Strombomonas taxonomybased on lorica morphology alone.
The two members of Colacium anchored theprofuse mucilage producing genera in our study.Although Colacium species do not produce loricasthey do produce copious amounts of mucilagefrom their anterior end. This mucilage is used forattachment to a substrate and subsequent stalk,and in some species, colony formation (Rosowskiand Kugrens 1973; Rosowski and Willey 1977).Moreover, like most Strombomonas and the basalmembers of Trachelomonas, all species of Cola-cium possess inner projecting haplopyrenoids.Our results are in agreement with recent molecularstudies (Ciugulea et al. 2008; Triemer et al. 2006)that support Colacium as the basal clade tocopious mucilage producing genera. Notably, thisis the first study to support the anchor position ofColacium by ML analysis, although the bootstrapsupport was weak (59%).
Monomorphina/Cryptoglena
Five members representing three species ofMonomorphina were used in our analyses. Allmembers of Monomorphina are diagnosed sensuKosmala et al. (2007) as being ovoid or slightlyoblate and rigid; a single parietal, sphericalchloroplast that is often perforated, 2–4 prominentlateral paramylon plates between the pellicle andthe chloroplast with M. pyrum as the epitype.Furthermore, based on molecular and morpholo-gical evidence Kosmala et al. (2007) synonymized
E.W. Linton et al.610

Author's personal copy
11 species of Monomorphina as M. pyrum anderected one new species M. pseudopyrum for theculture CCAC 0093 (originally misidentified as M.reeuwykiana by Marin et al. 2003). Our studysupports their changes and we have followed theirrecommendations; M. inconspicuus (ACOI 1295)and M. ovata (SAG 1244-5) are now considered asM. pyrum. Additionally, our study supports Mono-morphina (clade E) as being monophyletic andsister to Cryptoglena (clade F).
The members of Cryptoglena are similar toMonomorphina; cells ovoid, rigid with a singleparietal chloroplast and lateral, shield-shapedparamylon grains, pellicle lacking strip reduction.However, these genera have been shown inprevious molecular studies (Ciugulea et al. 2008;Kosmala et al. 2007; Marin et al. 2003; Milanowskiet al. 2006; Triemer et al. 2006) to be closelyrelated, but distinct genera. Additionally, based onthe amended diagnosis (Kosmala et al. 2007)Cryptoglena differ by lacking the strong pellicularridges found in Monomorphina, possessing amedian longitudinal furrow and having a chlor-oplast in the form of an open cylinder (C-shaped)lacking pyrenoids. These traits distinguish Cryp-toglena from Monomorphina. Our study supportsthe sister relationship between these genera andthe sister relationship to the newly erected genusEuglenaria.
Euglenaria
Members of this genus were formerly placed in thegenus Euglena and were morphologically indis-tinguishable from members of that genus. Thisposition was supported by molecular analyses,such as Marin et al. (2003) and Triemer et al. (2006)as well. However, molecular analysis byMilanowski et al. (2001, 2006) suggested thatEuglena anabaena was not a member of Euglena.But these studies had only one member of what isnow the Euglenaria clade (G) Euglena anabaena,so no reassignment of the taxon could have beenmade. Our study included five taxa in clade G,which has allowed an evaluation of both themorphological and molecular evidence. Morpho-logically uniting features of Euglenaria arethe parietal lobed chloroplast with diplopyrenoidsand absence of mucocysts. However, thesemorphological features do not differentiationEuglenaria from Euglena. Instead, the molecularnSSU characters are the diagnostic charactersthat separate Euglenaria from Euglena. While allEuglenaria have a C base upstream of helix 180
and a T base upstream of helix 38, all Euglena
have an A base upstream of helix 180 and a C or Abase upstream of helix 38. Furthermore, bothBayesian and ML analyses supported the separa-tion of these five taxa from Euglena and theirplacement as sister to the Cryptoglena andMonomorphina clades. Therefore, there is strongevidence and support for the erection of the newgenus Euglenaria and the transfer of five formermembers of Euglena into it.
Lepocinclis/Phacus
Six members of Lepocinclis and 15 members ofPhacus were included in our study. Cells are rigidor semi-ridged, sometimes exhibiting a slightbending or twisting motion, but none are meta-bolic as defined by Triemer et al. (2006). The cellscontain numerous small discoid chloroplast thatlack pyrenoids. This diagnosis could be appliedequally to both Phacus and Lepocinclis. The majordistinguishing characteristic was that all membersof Phacus were laterally compressed or flattened,while all members of Lepocinclis were notflattened in cross section. However, our studyhas resulted in the transfer of Lepocinclis salinaand Euglena limnophila into the genus Phacusrequiring the diagnosis to be emended to includecells that are spindle-shaped and ovoid. There-fore, being flattened is no longer a definingcharacteristic separating Phacus from Lepocin-clis.
What evidence is there for continuing thisseparation and not dissolving Lepocinclis (Perty1849) into Phacus (Dujardin 1841)? Neither this norany previous molecular phylogenetic study hasshown merging between these two genera; allhave shown them to be separate, well supportedclades. In addition, we have defined molecularsynapomorphies (shared derived) that distinguishthese genera from each other. In the nSSUalignments Lepocinclis species have the nucleo-tide T one base downstream of helix 15 and twobases downstream of helix 230. At these samepositions in Phacus the nucleotides C and A arepresent, respectively. We therefore recognizethese as two closely related sister clades andrecommend maintaining them as separate distinctgenera.
Discoplastis
Erected by Triemer et al. (2006), the two membersof Discoplastis contain numerous small discoidchloroplasts lacking pyrenoids and cells that arenot flattened, characteristics found in the closely
611Euglenoid Evolution using Three Genes

Author's personal copy
related Lepocinclis and Phacus genera. However,Discoplastis is easily distinguished from the othertwo genera by being strongly metabolic, acharacteristic not found in either Lepocinclis orPhacus. Our phylogenetic analysis of the mole-cular differences among these genera supportsthis genus and its basal position to the Lepocinclisand Phacus genera.
Our study was the first euglenoid phylogeny touse three genes, two nuclear and one chloroplastto establish relationships among freshwaterphotosynthetic euglenoids. This study showedstrong support by both Bayesian and ML analysisfor 10 major ingroup clades with 10 correspondinggenera and the single unallied taxon Euglenaproxima. Our analysis resulted in the transfer oftwo taxa (Lepocinclis salina and Euglena limno-phila) to the genus Phacus and the concomitantemended diagnosis of Phacus to contain cells thatwere not flattened. This represents the firstchange in the defining morphological character-istic, cell flattened, of Phacus since Dujardinestablished it in 1841. The polyphyletic genusEuglena was moved closer towards monophyly bythe erection and transfer of five taxa to the newgenus Euglenaria along with the transfer ofEuglena limnophila to the genus Phacus. Unfortu-nately the unallied, but strongly supported, posi-tion of Euglena proxima remains and consequentlymaintains Euglena as a paraphyletic genus.
Methods
Cultures: Table 1 lists the 82 taxa with sources and GenBankaccession numbers used in this study. Alternative name(s) inculture collections or GenBank for the taxa used in this study,due to misidentification or taxonomic changes, are also givenin Table 1. All taxa were obtained from culture collections orwere collected from small ponds in New Jersey (NJ) andMichigan (MI), USA. Cultures NJ and MI used in this study,which are not available from public culture collections, will bemade available upon request. Ponds were sampled with aplankton net (mesh size, 20 mm) and euglenoid cells isolatedby a Pasteur capillary pipette. Cells were brought into unialgalculture and checked periodically for eukaryotic contaminants.All isolates were grown in modified AF-6 medium (Watanabeand Hiroki 1997) and/or soil–water medium (medium 3c,Schlosser 1994). Cultures were maintained at 20–22 1C underconditions of a 14:10 light:dark cycle with approximately30 mmol photons m�2 s�1 provided from cool whitefluorescent tubes.
Specimen identification: Identifications of cultures wereconfirmed microscopically when received using a ZeissAxioskop 2 Plus microscope (Carl Zeiss Inc., Hallbergmoos,Germany) or a Nikon Eclipse E-600 microscope (Nikon, Tokyo,Japan) both equipped with differential interference contrast(DIC) optics. Images were captured with an AxioCam HRC(Hallbergmoos) or Nikon DX-1200 photomicrographic system
attached to the microscope, respectively. A representativespecies of each clade in the context of their phylogeneticrelationships is shown in Figure 3.
DNA extraction, amplification, sequencing, andsequence alignment: DNA extraction, PCR amplificationprocedures, purification, sequencing, and sequence align-ment were conducted as described by Brosnan et al. (2003).Total genomic DNA was isolated from cultures with theDNeasy Tissue Kit (Qiagen Co., Valencia, CA, USA; Cat#69504) using the animal tissues protocol. The nSSU, nLSUand cpSSU rDNA sequences were amplified in 20-mLreactions. Amplification was performed using; GeneAmpPCR System 9700 (Perkin Elmer Co., Norwalk, CT, USA), MJResearch PTC-0200 DNA Engine Gradient Thermo Cycler (MJResearch Inc., Waltham, MA, USA), Mastercycler Personaland Mastercycler Gradient (Eppendorf AG, Hamburg, Ger-many), see Table 2 for PCR programs used. The PCRproducts were sized on 1% agarose gels and then purifiedusing the MiniElute Gel Extraction Kit (Qiagen Co.; Cat#28606) or QIAEXII Gel Extraction Kit (Qiagen Co.;Cat#20051) according to the manufacturer’s protocol. Thepurified template was sequenced with internal primers forconserved regions (nSSU, Linton et al. 2000 and Table 3;nLSU, Brosnan et al. 2003; cpSSU Table 3) using either an ABI377, 3700, 3730 or 3730xl dye terminator sequencer (AppliedBiosystems, Foster City, CA, USA). Sequences wereassembled into contigs by the SeqMan program from theLASERGENE package (DnaStar, Madison, WI, USA) or usingthe Contig Assembly Program (CAP2) in the Genetic DataEnvironment (GDE 2.2) program (Smith et al. 1994). Thesequences for the nSSU, nLSU and cpSSU rDNAs werealigned by eye using the secondary structure of Euglenagracilis Klebs as a guide (Kjer 1995; Wuyts et al. 2001) in GDE2.2. The conserved areas of the genes were readily alignableacross taxa and were used for phylogenetic analyses. Areasthat could not be unambiguously aligned were excluded fromanalyses. All three gene sequences used in this study werederived from the same culture when possible (Table 1). Whennot possible due to culturing or sequencing difficultiesalternative sources were used, the taxon identitieswere confirmed microscopically and by sequencecomparisons among the first 700 bases of the nSSUsequences.
Phylogenetic analysis: The three genes resulted in acombined dataset of 5161 bases: 1740 from nSSU rDNA,2110 from nLSU rDNA and 1311 from cpSSU rDNA foranalysis. All analyses were done from within MacGDE 2.3. TheBayesian Information Criterion (BIC) analysis using MODELT-EST 3.7 (Posada and Crandall 1998) was used to determinethe best model for each dataset (nSSU, nLSU and cpSSU) foruse in partition Bayesian analysis while a single model basedon the BIC was determined for the combined data for use inBayesian and Maximum Likelihood (ML) analysis. Trees wererooted using Eutreptia viridis, Eutreptiella braarudii andEutreptiella pomquetensis as outgroup taxa. The three genedata set and tree (Fig. 2) have been deposited in TreeBASE,study accession number is S2655 and the matrix accessionnumber is M5103.
Bayesian analyses: The analyses were performed in Mr.Bayes 3.1.2 (Ronquist and Huelsenbeck 2003) using a singlemodel for the data or with an individual model for each of thethree data partitions. Parameters for the single model were asfollows (GTRþIþG): Prset revmatpr=dirichlet (1.49, 3.86, 1.63,0.65, 5.78, 1.00) statefreqpr=dirichlet (0.23, 0.24, 0.29, 0.23)shapepr=exponential (0.79) pinvarpr=fixed (0.30). Parametersfor the partitioned datasets were as follows (GTRþIþG): nSSU
E.W. Linton et al.612

Author's personal copy
Ta
ble
1.
Taxa
used
with
culture
co
llectio
nand
GenB
ank
accessio
nco
des.
Culture
for
seq
uencin
ga
GenB
ank
accessio
nnum
ber
Taxo
nnS
SU
nLS
Ucp
SS
UnS
SU
nLS
Ucp
SS
U
Co
laciu
mcalv
um
Ste
inM
I107
MI
107
MI
107
EF
999907
EF
999910
EU
624035
C.
mucro
natu
mB
ourr
elly
et
Chad
efa
ud
SA
G1211-1
(C.
sid
ero
pus)
SA
G1211-1
(C.
sid
ero
pus)
SA
G1211-1
(C.
sid
ero
pus)
AJ532441
EF
999906
EU
221482
Cry
pto
gle
na
pig
raE
hre
nb
erg
CC
AP
1212/1
CC
AP
1212/1
CC
AP
1212/1
AJ532437
DQ
140101
EU
221483
C.
skuja
eM
arin
et
Melk
onia
nS
AG
10.8
8(P
hacus
ag
ilis)
SA
G10.8
8(P
hacus
ag
ilis)
SA
G10.8
8(P
hacus
ag
ilis)
AY
014998
AY
130236
EU
221484
Dis
co
pla
stis
ad
unca
(Schill
er)
Trie
mer
AS
W08039
(Eug
lena
cf.
ad
unca)
AS
W08039
(Eug
lena
cf.
ad
unca)
AS
W08095
(Eug
lena
cf.
ad
unca)
AJ532453
DQ
140102
EU
750704
D.
spath
irhyn
cha
(Skuja
)Tr
iem
er
SA
G1224-4
2(E
ug
lena
spath
irhyn
cha)
SA
G1224-4
2(E
ug
lena
spath
irhyn
cha)
SA
G1224-4
2(E
ug
lena
spath
irhyn
cha)
AJ532454
DQ
140100
AY
626060
Eug
lena
ad
haere
ns
Matv
ienko
AS
W08138
AS
W08138
AS
W08138
EU
750713
FJ486278
EU
750706
E.
ag
ilis
Cart
er
OR
04
OR
04
OR
04
AF
115279
FJ377538
EU
624033
E.
canta
brica
Pring
sheim
SA
G1224-2
5S
AG
1224-2
5S
AG
1224-2
5A
J532412
AY
523020
AY
626047
E.
cla
raS
kuja
SA
G25.9
8S
AG
25.9
8S
AG
25.9
8A
J532423
DQ
140114
EU
750707
E.
dese
sE
hre
nb
erg
SA
G1224-2
3(E
.lim
no
phila
)S
AG
1224-2
3(E
.lim
no
phila
)S
AG
1224-2
3(E
.lim
no
phila
)A
J532407
DQ
140122
EU
750709
E.
genic
ula
taD
uja
rdin
SA
G1224-4
fS
AG
1224-4
fS
AG
1224-4
fA
Y070248
EU
624022
AY
070252
E.
genic
ula
taD
uja
rdin
SA
G1224-2
1(E
.vi
rid
is)
SA
G1224-2
1(E
.vi
rid
is)
SA
G1224-4
bA
Y523034
AY
523027
AF
289241
E.
gra
cili
sK
leb
sS
AG
1224-5
/15
UC
LA
variety
SA
G1224-5
/15
M12677
X53361
X12890
E.
gra
nula
ta(K
leb
s)
Schm
itz
UT
EX
2345
(E.
sang
uin
ea)
UT
EX
2345
(E.
sang
uin
ea)
UT
EX
2345
(E.
sang
uin
ea)
AJ532422
DQ
140099
EU
370511
E.
gym
no
din
ioid
es
Zakry
sM
I03
MI
03
MI
03
DQ
140148
DQ
140105
EU
221486
E.
lacin
iata
Pring
sheim
SA
G1224-8
c(E
.g
ranula
ta)
SA
G1224-8
c(E
.g
ranula
ta)
SA
G1224-8
c(E
.g
ranula
ta)
AJ532421
AY
523021
EU
221488
E.
lacin
iata
Pring
sheim
SA
G1224-3
1S
AG
1224-3
1S
AG
1224-3
1A
J532420
AY
523022
EU
221487
E.
long
a(P
ring
sheim
)M
arin
et
Melk
onia
nU
TE
X512
(Ast
asi
alo
ng
a)
UT
EX
512
(Ast
asi
alo
ng
a)
CC
AP
1204/1
7a
(Ast
asi
alo
ng
a)
AF
112871
AY
130223
AJ294725
E.
muta
bili
sS
chm
itz
NJ
NJ
Sand
yA
Y523038
DQ
140124
EU
221489
E.
pro
xim
aD
ang
eard
SA
G1224-1
1a
SA
G1224-1
1a
SA
G1224-1
1a
EU
624027
EU
624017
FJ374877
E.
pse
ud
ovi
rid
isC
had
efa
ud
SA
G1224-1
7c
(E.
virid
is)
SA
G1224-1
7c
(E.
virid
is)
SA
G1224-1
7c
(E.
virid
is)
AY
523037
DQ
140125
EU
370498
E.
sate
lles
Bra
sla
vska-S
pecto
rova
SA
G1224-2
2(E
.d
ese
s)S
AG
1224-2
2(E
.d
ese
s)S
AG
1224-2
2(E
.d
ese
s)A
J532406
EU
624016
EU
373477
E.
socia
bili
sD
ang
eard
AC
OI
920
AC
OI
920
AC
OI
920
EU
750715
EU
624024
EU
750710
E.
sp
ecie
sM
I32
MI
32
MI
32
DQ
140158
DQ
140123
EU
221490
E.
sp
ecie
sS
AG
1224-1
2b
(E.
socia
bili
s)S
AG
1224-1
2b
(E.
socia
bili
s)S
AG
1224-1
2b
(E.
socia
bili
s)D
Q140149
DQ
140106
EU
221491
E.
sple
nd
ens
Dang
eard
MI
47
MI
47
MI
47
DQ
140150
DQ
140107
EU
221492
E.
stella
taM
ain
xM
I09
MI
09
MI
09
AY
523032
AY
523024
EU
221493
E.
stella
taM
ain
xU
TE
X372
UT
EX
372
SA
G1224-1
4A
F150936
AY
130229
AF
289244
E.
tris
tella
Chu
NJ
NJ
SA
G1224-3
5D
Q140151
DQ
140109
AF
289246
613Euglenoid Evolution using Three Genes
Author's personal copy
Tab
le1.
(co
ntinued
)
Culture
for
seq
uencin
ga
GenB
ank
accessio
nnum
ber
Taxo
nnS
SU
nLS
Ucp
SS
UnS
SU
nLS
Ucp
SS
U
E.
virid
isep
ityp
eE
hre
nb
erg
AT
CC
PR
A110
AT
CC
PR
A110
AT
CC
PR
A110
AY
523036
AY
523031
EU
221494
Eug
lenaria
anab
aena
(Main
x)
Karn
ko
wska
et
Lin
ton
UT
EX
373
UT
EX
373
SA
G1224-1
5b
AF
242548
DQ
140115
AF
289240
E.
anab
aena
var.
min
ima
(Main
x)
Karn
ko
wska
et
Lin
ton
NJ
05
NJ
05
NJ
05
DQ
140155
DQ
140119
EU
221485
E.
caud
ata
(Hub
ner)
Karn
ko
wska
et
Lin
ton
AS
W08048
AS
W08048
AS
W08048
EU
750714
EU
624010
EU
750698
E.
caud
ata
var.
min
or
(Defland
re)
Karn
ko
wska
et
Lin
ton
SA
G1224-2
6a
(E.
caud
ata
)S
AG
1224-2
6a
(E.
caud
ata
)S
AG
1224-2
6a
(E.
caud
ata
)D
Q140147
DQ
140103
FJ374877
E.
cla
vata
(Skuja
)K
arn
ko
wska
et
Lin
ton
SA
G1224-3
2(E
ug
lena
po
lym
orp
ha)
SA
G1224-3
2(E
ug
lena
po
lym
orp
ha)
NJ
CP
AJ532436
DQ
140104
EU
750708
Eutr
ep
tia
virid
isP
ert
yS
AG
1226-1
cS
AG
1226-1
cS
AG
1226-1
cA
J532395
DQ
140108
AF
289247
Eutr
ep
tiella
bra
aru
dii
Thro
nd
sen
CC
MP
1594
CC
MP
1594
CC
MP
1594
DQ
249879
EU
624026
DQ
249873
Eutr
ep
tiella
po
mq
uete
nsi
s(M
cLachla
n,
Seg
uelet
Fritz
)M
arin
et
Melk
onia
nC
CM
P1491
CC
MP
1491
CC
MP
1491
AJ532398
EU
624012
EU
750699
Lep
ocin
clis
acus
var.
majo
r(M
ulle
r)M
arin
et
Melk
onia
nU
TE
X1316
(Eug
lena
acus
var.
majo
r)U
TE
X1316
(Eug
lena
acus
var.
majo
r)
SA
G1224-1
b(E
ug
lena
acus)
AF
152104
AY
130226
EU
221495
L.
fusi
form
is(C
art
er)
Lem
merm
ann
AC
OI
1025
AC
OI
1025
AC
OI
1025
AY
935697
EU
624011
AF
289249
L.
ovu
m(E
hre
nb
erg
)M
inkevic
hS
AG
1244-8
SA
G1244-8
SA
G1244-8
AF
110419
AY
130235
AY
221726
L.
spiro
gyr
oid
es
Marin
et
Melk
onia
nS
AG
1224-1
0b
(Eug
lena
oxy
uris)
SA
G1224-1
0b
(Eug
lena
oxy
uris)
SA
G1224-1
0b
(Eug
lena
oxy
uris)
AJ532464
AY
130814
AF
289243
(Eug
lena
spiro
gyr
a)
L.
stein
iiLem
merm
ann
UT
EX
523
(L.
buets
chlii
)U
TE
X523
(L.
buets
chlii
)U
TE
X523
(L.
buets
chlii
)A
F096993
AY
130815
EU
221496
L.
trip
teris
(Duja
rdin
)M
arin
et
Melk
onia
nU
TE
X1311
(Eug
lena
trip
teris)
UT
EX
1311
(Eug
lena
trip
teris)
SA
G1224-1
6A
F445459
AY
130230
EU
221497
Mo
no
mo
rphin
aaenig
matica
(Dre
zep
ols
ki)
Nud
elm
an
et
Trie
mer
UT
EX
1284
(Phacus
meg
alo
psi
s)U
TE
X1284
(Phacus
meg
alo
psi
s)
CC
AP
1261/9
(Phacus
aenig
maticus)
AF
190814
DQ
140117
AY
626054
(Mo
no
mo
rphin
ast
riata
)M
.p
seud
op
yrum
Ko
sm
ala
et
al.
CC
AC
0093
(M.
reeuw
ykia
na)
CC
AC
0093
(M.
reeuw
ykia
na)
CC
AC
0093
(M.
reeuw
ykia
na)
AJ532433
EF
999909
EU
750700
M.
pyr
um
(Ehre
nb
erg
)M
arin
et
Melk
onia
nS
AG
1244-5
(Lep
ocin
clis
ova
ta)
SA
G1244-5
(Lep
ocin
clis
ova
ta)
SA
G1244-5
(Lep
ocin
clis
ova
ta)
AF
061338
AY
130234
EU
221500
M.
pyr
um
(Ehre
nb
erg
)M
arin
et
Melk
onia
nA
CO
I1295
(Phacus
inco
nsp
icuus)
AC
OI
1295
(Phacus
inco
nsp
icuus)
AC
OI
1295
(Phacus
inco
nsp
icuus)
DQ
140129
DQ
140111
EU
221499
M.
pyr
um
(Ehre
nb
erg
)M
arin
et
Melk
onia
nU
TE
X2354
(Phacus
pyr
um
)U
TE
X2354
(Phacus
pyr
um
)U
TE
X2354
(Phacus
pyr
um
)A
F112874
AY
130238
EU
624034
Phacus
acum
inatu
sS
tokes
SA
G1261-7
(P.
bra
chyk
entr
on)
UT
EX
1317
(P.
bra
chyk
entr
on)
SA
G1261-7
(P.
bra
chyk
entr
on)
AJ532481
AY
130820
EU
221501
P.
anky
lono
ton
Po
chm
ann
CC
AC
0043
(P.
ranula
)C
CA
C0043
(P.
ranula
)C
CA
C0043
(P.
ranula
)A
J532484
EU
624020
EU
624036
P.
ap
pla
natu
sP
ochm
ann
AS
W08023
AS
W08023
AS
W08023
EU
624031
EU
624015
EU
750701
E.W. Linton et al.614

Author's personal copy
P.
ham
elii
Allo
rge
et
Lef� e
vre
AC
OI
1088
AC
OI
1088
AC
OI
1088
DQ
397673
EU
624019
EU
221502
P.
limno
phila
(Lem
merm
ann)
Lin
ton
et
Karn
ko
wska
AC
OI
1026
(Eug
lena
limno
phila
)A
CO
I1026
(Eug
lena
limno
phila
)
AC
OI
1026
(Eug
lena
limno
phila
)
DQ
249877
EU
624025
AY
626056
P.
orb
icula
ris
Hub
ner
AIC
B502
MI
76
AIC
B502
AY
935698
EU
624018
AY
626057
P.
parv
ulu
sK
leb
sA
SW
08060
AS
W08060
AC
OI
1093
(P.
pusi
llus)
AF
283314
DQ
140127
EU
221503
P.
ple
uro
necte
s(M
ulle
r)D
uja
rdin
SA
G1261-3
bS
AG
1261-3
bS
AG
1261-3
bA
J532475
AY
130824
AF
289251
P.
pusi
llus
Lem
merm
ann
UT
EX
1282
UT
EX
1282
UT
EX
1282
AF
190815
AY
130237
EU
750702
P.
salin
a(F
rits
ch)
Lin
ton
et
Karn
ko
wska
SA
G1244-3
(Lep
ocin
clis
salin
a)
SA
G1244-3
(Lep
ocin
clis
salin
a)
SA
G1244-3
(Lep
ocin
clis
salin
a)
EU
624028
EU
624023
AY
221727
P.
seg
retiiA
llorg
eet
Lef� e
vre
AC
OI
1337
AC
OI
1337
AC
OI
1337
EU
624030
EU
624014
EU
221504
P.
skuja
eS
kvo
rtzo
vU
TE
X1285
(P.
caud
ata
)U
TE
X1285
(P.
caud
ata
)U
TE
X1285
(P.
caud
ata
)A
F181968
AY
130823
EU
750711
P.
smulk
ow
skia
nus
(Zakry
s)
Kusb
er
SA
G58.8
1(P
.si
mili
s)S
AG
58.8
1(P
.si
mili
s)A
CO
I1226
(P.
sim
ilis)
AJ532467
AY
130239
AY
626059
P.
triq
uete
r(E
hre
nb
erg
)D
uja
rdin
SA
G1261-8
SA
G1261-8
SA
G1261-8
AJ532485
EU
624013
EU
221505
P.
wars
zew
iczi
iD
reze
po
lski
AS
W08064
AS
W08064
AS
W08064
EU
624032
EU
624021
EU
750703
Str
om
bo
mo
nas
acum
inata
(Schm
ard
a)
Defland
reN
JS
716
NJ
S716
SA
G1280-1
(T.
co
nsp
ers
a)
EU
624029
AY
359914
EU
221506
S.
co
stata
Defland
reA
CO
I1273
AC
OI
1273
AC
OI
2992
DQ
140152
AY
359915
AF
289253
S.
ova
lis(P
layfa
ir)
Defland
reN
JS
115
NJ
S115
NJ
S115
DQ
140133
AY
359919
EU
750712
S.
verr
uco
sa(v
on
Dad
ay)
Defland
reN
JS
5c
NJ
S5c
AC
OI
2476
(S.
acum
inata
)E
F999896
AY
359911
AY
626051
Trachelo
mo
nas
ab
rup
ta(S
wirenko
)D
efland
reN
JT
801
NJ
T801
NJ
T801
DQ
140134
AY
359941
EU
221507
T.b
ern
ard
inensi
sV
ischer
AC
OI
1103
AC
OI
1103
AC
OI
1103
EF
999908
AY
359950
EU
221509
T.echin
ata
Sin
gh
SA
G1283-2
2S
AG
1283-2
2S
AG
1283-2
2A
Y015001
AY
130242
EU
221510
T.elli
pso
idalis
Sin
gh
NJ
ST
1N
JS
T1
NJ
ST
1D
Q140135
AY
359935
EU
221511
T.his
pid
a(P
ert
y)
Defland
reU
TE
X1325
(T.o
blo
ng
avar.
puncta
ta)
UT
EX
1326
UT
EX
539
(T.
his
pid
ava
r.co
ronata
)
AF
445462
AY
130817
EU
221513
T.le
fevr
eiD
efland
reS
AG
1283-1
0S
AG
1283-1
0S
AG
1283-1
0D
Q140136
AY
359949
EU
221514
T.o
blo
ng
aLem
merm
an
NJ
T516
NJ
T516
NJ
T516
DQ
140137
AY
359947
EU
221515
T.p
lancto
nic
aS
wirenko
NJ
WT
1N
JW
T1
NJ
WT
1D
Q140138
AY
359954
EU
221516
T.ru
gulo
saS
tein
NJ
TS
2N
JT
S2
NJ
TS
2D
Q140140
AY
359942
EU
221517
T.si
mili
sS
tokes
SA
G1283-1
4S
AG
1283-1
4S
AG
1283-1
4D
Q140142
AY
359948
AY
626055
T.vo
lvo
cin
aE
hre
nb
erg
SA
G1283-4
UT
EX
1327
UT
EX
1327
AJ532451
AY
359953
EU
221519
T.vo
lvo
cin
op
sis
Svirenko
SA
G1283-1
6S
AG
1283-1
6S
AG
1283-1
6D
Q140144
AY
359944
EU
221520
T.zo
rensi
sLef� e
vre
UT
EX
1331
UT
EX
1331
UT
EX
1331
DQ
140145
AY
359952
EU
221521
New
or
up
date
dseq
uences
are
ind
icate
din
bo
ldty
pe.
aS
train
sla
bele
d,A
CO
I,C
oim
bra
Co
llectio
no
fA
lgae,C
oim
bra
,P
ort
ug
al;
AS
W,A
lgenkultur-
Sam
mlu
ng
an
der
Univ
ers
itat
Wie
n,V
ienna,A
ustr
ia;
AT
CC
,A
merican
Typ
eC
ulture
co
llectio
n,
Manassa,
VA
,U
SA
;C
CA
C,
Culture
co
llectio
no
fA
lgae
at
the
Univ
ers
ity
of
Co
log
ne,G
erm
any;
CC
AP,
Culture
co
llectio
no
fA
lgae
and
Pro
tozo
a,
Sco
tland
;C
CM
P,C
ulture
co
llectio
no
fM
arine
Phyto
pla
nkto
n,
US
A;
MI,
Mic
hig
an
iso
late
Trie
mer
lab
,U
SA
;N
J,
New
Jers
ey
iso
late
Trie
mer
lab
,U
SA
;S
AG
,S
am
mlu
ng
vo
nA
lgenkulture
nP
flanze
np
hysio
log
isches
Institu
td
er
Univ
ers
itat
Go
ttin
gen,
Germ
any;
UT
EX
Culture
Cente
ro
fA
lgae,
Austin,
TX
,U
SA
.
615Euglenoid Evolution using Three Genes

Author's personal copy
Figure 3. A rooted Bayesian phylogenetic tree, as in Figure 2 but with each of the major clades collapsed(indicated by a capital letter) and a representative taxon from that clade pictured (not to scale). Names ofpictured taxa from left to right; Euglena viridis, Trachelomonas hispida, Strombomonas verrucosa, Colaciummucronatum, Monomorphina pyrum, Cryptoglena skujae, Euglenaria anabaena, Euglena proxima, Lepocinclisspirogyroides, Phacus orbicularis, Discoplastis spathirhyncha. The tree was outgroup rooted posteriorly withEutreptia and Eutreptiella species (not shown).
Table 2. PCR reaction parameters.
General PCR parameters96 1C 2 min, 35 cycles (95 1C 30 s, 37 1C – 56 1C 30 s, 72 1C 1 min) 72 1C 6 min, and a 4 1C hold
PCR parameters for cpSSUF/cpSSUR and cpSSU647F/cpSSUR95 1C 5 min, 37 cycles (95 1C 1 min, 44 1C 2 min, 72 1C 2 min – 7 cycles and 95 1C 30 s, 54 1C 30 s, 72 1C1 min – 30 cycles) 72 1C 10 min, and a 7 1C hold
PCR parameters for cpSSUF/cpSSU854R951 C 5 min, 37 cycles (951 C 1 min, 481 C 2 min, 721 C 2 min – 7 cycles and 951 C 30 s, 561 C 30 s, 721 C1 min – 30 cycles) 721 C 10 min, and a 71 C hold
PCR parameters for nSSUF/nSSU557R95 1C 5 min, 39 cycles (95 1C 1 min, 48 1C 90 s, 72 1C 1 min – 7 cycles and 95 1C 30 s, 56 1C 30 s, 72 1C 45 s– 32 cycles) 72 1C 10 min, and a 7 1C hold
PCR parameters for nSSU382F/nSSU1263R95 1C 5 min, 39 cycles (95 1C 1 min, 40 1C 90 s, 72 1C 90 s – 7 cycles and 95 1C 30 s, 49 1C 30 s, 72 1C 90 s –32 cycles) 72 1C 10 min, and a 7 1C hold
PCR parameters for nSSU1154F/nSSUR95 1C 5 min, 37 cycles (95 1C 1 min, 54 1C 1 min, 72 1C 45 s – 7 cycles and 95 1C 30 s, 59 1C 30 s, 72 1C 45 s– 32 cycles) 72 1C 10 min, and a 7 1C hold
E.W. Linton et al.616

Author's personal copy
revmatpr=dirichlet (1.37, 3.16, 1.45, 0.59, 4.53, 1.00) state-freqpr=dirichlet (0.21, 0.29, 0.28, 0.22) shapepr=exponential(0.73) pinvarpr=fixed (0.28); nLSU revmatpr=dirichlet (1.70,3.69, 1.80, 0.62, 6.23, 1.00) statefreqpr=dirichlet (0.21, 0.26,0.33, 0.21) shapepr=exponential (0.82) pinvarpr=fixed (0.33);cpSSU revmatpr=dirichlet (1.00, 5.18, 1.00, 1.00, 8.85, 1.00)statefreqp=dirichlet (0.33, 0.13, 0.20, 0.34) shapepr=expo-nential (0.59) pinvarp r=fixed (0.24). The analysis used fourMarkov chains (4,000,000 generations per chain), with treessaved every 100 generations discarding the first 8,000 trees. Amajority-rule consensus tree was created from the remaining32,001 trees. Convergence among these trees was confirmedvia the sump command.
Maximum likelihood analyses: The analysis was per-formed with PAUPn 4.0b10 for Macintosh OS X (Swofford 2002)using the following parameters (GTRþIþG): Base=(0.22750.2439 0.2943) Nst=6 Rmat=(1.4872 3.8611 1.6340 0.64815.7788) Rates=gamma Shape=0.7907 Pinvar=0.3016Shape=0.7907 NCat=4 RepRate=Mean Initbrlen=Rogers STart-vals=ParsApprox RECon=Marginal. The single ML tree wasobtained by a heuristic search using random stepwise additionwith 5 replicates, TBR branch swapping and MULTREES on.The 100 ML bootstrap repetitions were obtained by a Heuristicsearch using Random Stepwise addition with two replicates.The 100 non-parametric bootstraps were done as foursimultaneous independent searches of 25 bootstraps, eachrun on a separate CPU. The trees were aggregated andweighted according to the number of trees found in eachbootstrap replicate, so that all replicates had equal weight, a50% majority rule consensus bootstrap tree was obtained.
Acknowledgements
The authors wish to acknowledge the financialsupport provided by the National Science Foun-dation PEET program (Partnership for EnhancedExpertise in Taxonomy, grant no. DEB 4-21348)and by the Ministry of Science and HigherEducation grant no. N303 010 32/0552. We thankProf. Tomasz Majewski, Warsaw, Poland, forproviding a Latin diagnose for Euglenaria gen. nov.
References
Brosnan S, Brown PJ, Farmer MA, Triemer RE (2005)Morphological separation of the euglenoid generaTrachelomonas and Strombomonas based on loricadevelopment and posterior strip reduction. J Phycol 41:590–605
Brosnan S, Shin W, Kjer KM, Triemer RE (2003) Phylogeny ofthe photosynthetic euglenophytes inferred from the nuclear SSUand partial LSU rDNA. Int J Syst Evol Microbiol 53:1175–1186
Busse I, Preisfeld A (2002) Phylogenetic position of Rhyncho-pus sp. and Diplonema ambulator as indicated by analyses ofeuglenozoan small subunit ribosomal DNA. Gene 284:83–91
Busse I, Preisfeld A (2003) Systematics of primary osmo-trophic euglenids: a molecular approach to the phylogeny ofDistigma and Astasia (Euglenozoa). Int J Syst Evol Microbiol53:617–624
Busse I, Patterson DJ, Preisfeld A (2003) Phylogeny ofphagotrophic euglenids (‘‘Euglenozoa’’): a molecularapproach based on culture material and environmentalsamples. J Phycol 39:828–836
Ciugulea I, Nudelman MA, Brosnan S, Triemer RE (2008)Phylogeny of the euglenoid loricate genera Trachelomonasand Strombomonas (Euglenophyta) inferred from nuclear SSUand LSU rDNA. J Phycol 44:406–418
Dujardin F (1841) In: Histoire naturelle des Zoophytes.Infusoires Roret, Paris
Ehrenberg CG (1830a) Neue Beobachtungen uber blutartigeErscheinungen in Agypten, Arabien und Sibirien, nebst einerUbersicht und Kritik der fruher bekannten. Pogg Annalen derPhysik Chemie 18:477–514
Ehrenberg CG (1830b) Beitrage zur Kenntnis der Organisa-tion der Infusorien und ihrer geographischen Verbreitung,besonders in Sibirien. Abh Konigl Akad Wissensch Berlin:1–88
Ehrenberg CG (1831) Uber die Entwickelung und Lebens-dauer der Infusionsthiere; nebst ferneren Beitragen zu einerVergleichung ihrer organischen Systeme. Abh Konigl AkadWissensch Berlin 1832:1–154
Table 3. New primers used in this study.
Primer Sequence (5’–3’)
cpSSU-F TTG ATC CTG GCT CAG GAT GAA CGC TcpSSU-647F ATT TCC AGT GTA GCG GTGcpSSU-854R AGG CGG GAC ACT TAA CGC GTTcpSSU-R CAA GGA GGT GAT CCA GCC GCA CCT TnSSU-F CAG TGG GTC TGT GAA TGG CTC CnSSU-557R TTA CCG CAG CTG CTG GCnSSU-570F GTG CCA GCA GCT GCG GTnSSU-1141F GAA ACT TAA AGG AAT TGnSSU-1154F GGA ATT GAC GGA ATG GCA CCnSSU-1263R GAG CGG CCA TGC ACC ACnSSU-R CGA CGG GCG GTG TGT ACA AGT
F=Forward Primer, R=Reverse Primer.
617Euglenoid Evolution using Three Genes

Author's personal copy
Ehrenberg CG (1833) Dritter Beitrag zur Erkenntnis grosserOrganisation in der Richtung des kleinsten Raumes. AbhKonigl Akad Wissensch Berlin:145–336
Ehrenberg CG (1838) Die Infusionsthierchen als vollkommeneOrganismen. Ein Blick in das tiefere organische Leben derNatur. Nebst einem Atlas von vierundsechzig colorirtenKupfertafeln, gezeichnet vom Verfasser. Leipzig Verlag vonLeopold Voss.
Fritsch FE (1914) Notes on British Flagellates, I-IV. NewPhytol vol 13 (Iss 10): 341–351
Gibbs S (1978) The chloroplast of Euglena may haveevolved from symbiotic green algae. Can J Bot 56:2883–2889
Hubner EFW (1886) Euglenaceen-Flora von Stralsund. Pro-gram Realgymnasiums zu Stralsund, pp 1–20
Ishida K, Cao Y, Hasegawa M, Okada N, Hara Y (1997) Theorigin of chlorarachniophyte plastids, as inferred from phylo-genetic comparisons of amino acid sequences of EF-Tu. J MolEvol 45:682–687
Kjer KM (1995) Use of rRNA secondary structure inphylogenetic studies to identify homologous positions: anexample of alignment and data presentation from the frogs.Mol Phylogenet Evol 4:314–330
Kosmala S, Karnkowska-Ishikawa A, Milanowski R,Kwiatowski J, Zakrys B (2009) Phylogeny and systematicsof species from the genus Euglena (Euglenaceae) with axial,stellate chloroplasts based on morphological and moleculardata – new taxa, emended diagnoses and epitypifications. JPhycol 45:464–481
Kosmala S, Milanowski R, Brzoska K, Pekala M, Kwia-towski J, Zakrys B (2007) Phylogeny and systematics of thegenus Monomorphina (Euglenaceae) based on morphologicaland molecular data. J Phycol 43:171–185
Leander BS, Triemer RE, Farmer MA (2001) Characterevolution in heterotrophic euglenoids. Eur J Protistol 37:337–356
Leander B (2004) Did trypanosomatid parasites have photo-synthetic ancestors? Trends Microbiol 12:251–258
Linton E, Hittner D, Lewandowski CL, Auld T, Triemer RE(1999) A molecular study of euglenoid phylogeny using smallsubunit rDNA. J Eukaryot Microbiol 46:217–223
Linton EW, Nudelman MA, Conforti V, Triemer RE (2000) Amolecular analysis of the euglenophytes using SSU rDNA. JPhycol 36:740–746
Mainx F (1926) Einige neue Vertreter der Gattung EuglenaEhrenberg mit Unterstutzung der Gesellschaft zur Forderungdeutschen Wissenschaft. Kunst und Literatur in Bohmen. ArchProtistenkd 54:150–160
Mainx F (1927) Beitrage zur Morphologie und Physiologie derEuglenen. I. Morphologische Beobachtung, Methode, undErfolge der Reinkultur. II. Untersuchung uber die Ernahrungs-und Reizphysiologie. Arch Protistenkd 60:305–414
Marin B, Palm A, Klingberg M, Melkonian M (2003)Phylogeny and taxonomic revision of plastid-containingEuglenophytes based on SSU rDNA sequence comparisonsand synapomorphic signatures in the SSU rRNA secondarystructure. Protist 154:99–145
Martin W, Sommerville CC, Loiseaux de Goer S (1992)Molecular phylogenies of plastid origins and algal evolution. JMol Evol 35:385–403
Mereschkowsky KS (1877) Etjudy nad prostejsimi zivotnymisevera Rossii. Trudy S-Peterburgsk Obshch Estestvoisp 8:1–299
Milanowski R, Zakrys B, Kwiatowski J (2001) Phylogeneticanalysis of chloroplast small-subunit rRNA genes of the genusEuglena Ehrenberg. Int J Syst Evol Microbiol 51:773–781
Milanowski R, Kosmala S, Zakrys B, Kwiatowski J (2006)Phylogeny of photosynthetic euglenophytes based on com-bined chloroplast and cytoplasmic SSU rDNA sequenceanalysis. J Phycol 42:721–730
Mignot JP (1966) Structure et ultrastructure de quelqueseuglenomonadines. Protistologia 2:51–140
Moreira D, Lopez-Garcia P, Rodriguez-Valera F (2001) Newinsights into the phylogenetic position of diplonemids: GþCcontent bias, differences of evolutionary rate and anew environmental sequence. Int J Syst Evol Microbiol51:2211–2219
Mullner AN, Angler DG, Samuel R, Linton EW, Triemer RE(2001) Phylogenetic analysis of phagotrophic, phototrophic,and osmotrophic euglenoids by using the nuclear 18S rDNAsequence. Int J Syst Evol Microbiol 51:783–791
Nozaki H (2005) A new scenario of plastid evolution: plastidprimary endosymbiosis before the divergence of the ‘‘Plan-tae,’’ emended. J Plant Res 118:247–255
Nudelman MA, Rossi MS, Conforti V, Triemer RE (2003)Phylogeny of the Euglenophyceae based on small subunit rDNAsequences: taxonomic implications. J Phycol 39:226–235
Perty M (1849) Uber verticale Verbreitung mikroskopischerLebensformen. Mitt Naturforsch Ges Bern:17–45
Posada D, Crandall KA (1998) Modeltest: testing the modelof DNA substitution. Bioinformatics 14:817–818
Preisfeld A, Berger S, Busse I, Liller S, Ruppel HG (2000)Phylogenetic analyses of various euglenoid taxa (Euglenozoa)based on 18S rDNA sequence data. J Phycol 36:220–226
Preisfeld A, Busse I, Klingberg M, Talke S, Ruppel HG(2001) Phylogenetic position and inter-relationships of theosmotrophic euglenids based on SSU rDNA data, withemphasis on the Rhabdomonadales (Euglenozoa). Int J SystEvol Microbiol 51:751–758
Ronquist F, Huelsenbeck JP (2003) MRBAYES 3: Bayesianphylogenetic inference under mixed models. Bioinformatics19:1572–1574
Rosowski JR, Kugrens P (1973) Observations on theeuglenoid Colacium with special reference to the formationand morphology of attachment material. J Phycol 9:370–383
Rosowski JR, Willey RL (1977) Development of mucilaginoussurfaces in euglenoids. I. Stalk morphology of Colaciummucronatum. J Phycol 13:16–21
Schlosser UG (1994) SAG Sammlung von Algenkulturen atthe University of Gottingen. Catalogue of strains of 1994. BotActa 107:113–187
Skuja H (1948) Taxonomie des Phytoplanktons Einiger Seenin Uppland Schweden. Symb Bot Ups 9:1–399
E.W. Linton et al.618

Author's personal copy
Smith SW, Overbeek R, Woese CR, Gilbert W, Gillevet PM(1994) The genetic data environment: an expandable GUI formultiple sequence analysis. Comput Appl Biosci 10:671–675
Swofford DL (2002) In: PAUPn: Phylogenetic Analysis UsingParsimony (nand Other Methods). Version 4.0b10. SinauerAssociates, Sunderland, MA
Talke S, Preisfeld A (2002) Molecular evolution of eugle-nozoan paraxonemal rod genes par1 and par2 coincides withphylogenetic reconstruction based on small subunit rDNAdata. J Phycol 38:995–1003
Triemer R, Linton E, Shin W, Nudelman M, Brosnan S,Monfils A, Bennett M (2006) Phylogeny of the Euglenalesbased upon combined SSU and LSU rDNA sequencecomparisons and description of Discoplastis gen. nov.(Euglenophyta). J Phycol 42:731–740
Triemer RE, Farmer MA (2007) A Decade of EuglenoidMolecular Phylogenetics. In Brodie J, Lewis J (eds) Unravelingthe Algae: The Past, Present and Future of Algal Systematics.Systematics Association Series. CRC Press, Boca Raton, FL,pp 315–329
Von der Heyden S, Chao EE, Vickerman K, Cavalier-SmithT (2004) Ribosomal RNA phylogeny of bodonid and diplone-mid flagellates and the evolution of Euglenozoa. J EukaryotMicrobiol 51:402–416
Watanabe MM, Hiroki M (1997) In: NIES – Collection List ofStrains, 5th ed. National Institute for Environmental Studies,Tsukuba, 127 pp
Wuyts J, De Rijk P, Van de Peer Y, Winkelman T,DeWachter R (2001) The European large subunit ribosomalRNA database. Nucleic Acids Res 29:175–177
619Euglenoid Evolution using Three Genes