systematics and dna barcoding of free-living marine nematodes with emphasis on tropical desmodorids...
TRANSCRIPT

Nematology 16 (2014) 979-989 brill.com/nemy
Systematics and DNA barcoding of free-living marine nematodeswith emphasis on tropical desmodorids using nuclear SSU rDNA
and mitochondrial COI sequences
Maickel ARMENTEROS 1,∗, Ariadna ROJAS-CORZO 1, Alexei RUIZ-ABIERNO 1, Sofie DERYCKE 2,Thierry BACKELJAU 3,4 and Wilfrida DECRAEMER 3,5
1 Centro de Investigaciones Marinas, Universidad de La Habana, 16 # 114, CP 11300, Playa, Habana, Cuba2 Department of Biology, Marine Biology Section, Ghent University, Campus Sterre S8, B-9000 Ghent, Belgium
3 Royal Belgian Institute of Natural Sciences, Rue Vautier 21, B-1000 Brussels, Belgium4 Evolutionary Ecology Group, University of Antwerp, Groenenborgerlaan 171, B-2020 Antwerp, Belgium
5 Department of Biology, Nematology Research Unit, Ghent University, Ledeganckstraat 35, B-9000 Ghent, Belgium
Received: 15 April 2014; revised: 2 July 2014Accepted for publication: 2 July 2014; available online: 28 August 2014
Summary – The diversity and phylogenetic relationships of the Desmodoridae, a widespread tropical family of free-living marinenematodes, is hitherto poorly known both from molecular and taxonomic points of view. We performed a molecular phylogeneticanalysis of marine nematodes to: i) disentangle relationships among tropical desmodorid species; and ii) compare the performanceof the nuclear SSU rDNA and mitochondrial COI nucleotide sequences in 42 and 45 nominal species, respectively, to identifyspecies. We generated 27 new sequences of SSU rDNA belonging to five genera not previously sequenced, and 34 new sequencesof COI belonging to six genera and four families not previously sequenced. The SSU rDNA tree confirmed the Enoplida to be amonophyletic sister group to the Chromadorida. The family Comesomatidae is a sister group of the Xyalidae within the Monhysterida.Both DNA markers confirmed the congruence between the morphology- and molecular-based phylogenetic inferences for most of thefamilies. Desmodoridae was a monophyletic group, but the relationships within the family could not be recovered; the subfamiliesDesmodorinae and Spiriniinae were not monophyletic meanwhile the monophyly of Stilbonematinae was not fully supported due toa few specimens of questionable identity. COI performed better than SSU rDNA to disentangle relationships among closely relatedspecies and suggested the presence of cryptic diversity within Desmodoridae. COI is effective to explore cryptic diversity and barcodespecies within Nematoda, with a possible threshold of genetic distance of 5% between conspecific and interspecific sequences, butDNA barcoding is limited by the poor knowledge of the diversity and taxonomy of the group and the lack of a good reference databaseof vouchered COI sequences.
Keywords – 18S rDNA, Chromadorea, Enoplea, molecular, phylogeny, taxonomy.
Nematodes are one of the most diverse groups of meta-zoans (Zhang, 2011) and are probably the most abundantand ubiquitous (Margulis & Chapman, 2009). They playa key role in the functioning of marine ecosystems andhave been extensively studied from a fundamental (e.g.,diversity, phylogeny) and applied approach (e.g., biomon-itoring). As a result, the phylogenetic relationships withinNematoda have been intensively studied and a robust phy-logenetic framework exists for the phylum (Bert et al.,2011). Nevertheless, the taxonomy of many free-livingnematodes still remains largely unresolved and debated
∗ Corresponding author, e-mail: [email protected]
because of the regular occurrence of parallel evolution(Blaxter et al., 1998; van Megen et al., 2009) and con-served morphology (Holterman et al., 2008).
The first phylogenetic classification of free-living ne-matodes was established by Lorenzen (1994) using mor-phological features. It largely still stands for the intra-familial systematics. Relationships within the phylumhave been resolved by using the nuclear small subunit ofribosomal DNA (SSU rDNA) but these studies mostly in-cluded parasitic and soil nematodes (e.g., Aleshin et al.,1998; Blaxter et al., 1998). De Ley & Blaxter (2004) in-
© Koninklijke Brill NV, Leiden, 2014 DOI:10.1163/15685411-00002824

M. Armenteros et al.
troduced a new classification system based on the combi-nation of morphological and molecular features; impor-tantly, these authors translated the found clades into aLinnean classification. Meldal et al. (2007) improved thephylogeny of nematodes by increasing the number of ma-rine taxa in the analysis. The most inclusive phylogeneticstudy of nematodes currently available contains 1215 se-quences of SSU rDNA (van Megen et al., 2009). How-ever, marine taxa remain strongly under-represented (Bertet al., 2011). Moreover, the 46 sequences of marine taxaadded by Meldal et al. (2007) came from northwest Eu-ropean coastal waters, rendering the representation of ma-rine taxa from tropical regions hitherto scarce. Evidently,information on taxa from tropical areas may reveal novelinsights in the systematics and diversity of nematodes.
Against this background, we present a molecular sys-tematic analysis of marine desmodorid nematodes from atropical coral reef in the Caribbean Sea at Punta Francés,Cuba (Armenteros et al., 2012). The Desmodoridae Filip-jev, 1922 is one of the most speciose and diverse groups ofmarine nematodes (318 species after Hodda, 2011). How-ever, its systematics remains unresolved because of threereasons. First, Desmodoridae comprises many poorly de-fined taxa (e.g., Laxonema Cobb, 1933; MetadesmodoraSchuurmans Stekhoven, 1942) and only a few recentreviews have been published for speciose groups (e.g.,Desmodora de Man, 1889 by Verschelde et al. (1998)and Stilbonematinae by Tchesunov (2013)). Second, thereare conflicting morphological characters (Kampfer et al.,1998). Third, the group is still poorly documented byDNA sequence data, indicating a clear necessity to gener-ate sequence data from taxa that have hitherto been under-represented.
SSU rDNA is the most widely used DNA marker tostudy the systematics of nematodes because it is eas-ily amplified across the phylum and it has successfullyresolved several taxonomic species-level problems andrelationships (Rodrigues Da Silva et al., 2010). There-fore, we used SSU rDNA to explore certain problem-atic phylogenetic relationships within desmodorid nema-todes. In addition, we used the mitochondrial cytochromeoxidase c subunit 1 (COI) gene to resolve relationshipsamong closely related and/or cryptic nematode species(Derycke et al., 2005, 2010a) in a DNA barcoding context.
The objectives of this study are hence twofold: i) todisentangle phylogenetic relationships among tropicalfree-living marine desmodorids; and ii) to compare theability of SSU rDNA and COI to identify or differentiatespecies of marine nematodes.
Materials and methods
COLLECTION AND SORTING OF NEMATODES
Nematodes were collected in July 2010 in the SWregion of the Cuban Archipelago, Punta Francés Reef(21°36′29.68′′N, 83°10′34.40′′W) (Armenteros et al.,2012). Sediment samples were collected manually withplastic cores (2.6 cm diam. × 8 cm length) from sand flatsat 2-3 m depth with scarce vegetation and seagrass mead-ows of Thalassia testudinum.
Samples were sieved over a 45 μm mesh sieve andpreserved in DESS solution (Yoder et al., 2006) untilfurther processing. Nematodes were extracted from thesamples under a stereomicroscope and again stored inDESS solution.
DNA EXTRACTION AND SEQUENCING
Specimens were mounted, one by one, on temporarymicroscope preparations in a drop of distilled water. Theywere identified and photo-vouchered at magnifications of100×, 400× and 1000× using an Olympus BX41 lightmicroscope (LM) with interference contrast coupled toa Color View digital camera with the software OlympuscellˆD. Identifications were based on published taxonomicdescriptions compiled mostly in the NeMys database(available online at http://www.marinespecies.org).
Identified and photographed nematodes were subse-quently transferred to individual Eppendorf vials contain-ing worm lysis buffer consisting of 50 mM KCL, 10 MmTris, pH 8.3, 2.5 mM MgCl2, 0.45% NP 40 (TergitolSigma) and 0.45% Tween 20. Subsequently, they werefrozen at −20°C to break the cell walls. Afterwards, 1 μlof proteinase K (10 mg ml−1) was added and sampleswere incubated during 1 h at 65°C to digest the proteinsfollowed by another 10 min at 95°C to inactivate the pro-teinase K. Finally, the samples were centrifuged during1 min at 21 000 g. DNA extracts were stored at −80°C.
Fragments of the nuclear small subunit ribosomal DNA(SSU rDNA) and the mitochondrial cytochrome oxidase c
subunit 1 (COI) genes were PCR amplified. For SSUrDNA, we used the primer set G18S4 (f): 5′-GCT TGTCTC AAA GAT TAA GCC-3′ and 4R (r): 5′-GTA TCTGAT CGC CKT CGA WC-3′ which amplifies a fragmentof ca 1000 bp. For COI, we used the primer set JB3 (f):5′-TTT TTT GGG CAT CCT GAG GTT TAT-3′ and JB5(r): 5′-AGC ACC TAA ACT TAA AAC ATA ATG AAAATG-3′ which amplifies a fragment of 426 bp (Derycke etal., 2010a).
980 Nematology

Systematics of free-living marine nematodes
PCR amplifications were done in 25 μl of reactionvolume with the following mix: 1 μl of DNA template,2.5 μl of 10× PCR buffer, 2.5 μl of Coral Load 10×,2 μl of MgCl2 (25 mM), 0.5 μl deoxynucleotides (10 mMeach), 0.125 μl of primers (25 μM), and 0.125 μlof TopTaq DNA polymerase (5 U μl−1; Qiagen). PCRcycling conditions were: i) initial denaturation of 5 minat 95°C; and ii) 35 cycles (for COI) or 40 cycles (for SSUrDNA) of (94°C for 30 s, 54°C for 30 s, 72°C for 30 s),followed by a final extension of 10 min at 72°C.
PCR products were loaded on 1.2% agarose gels,run in an electrophoresis chamber and visualised witha BioRad UV system. Amplifications were consideredsuccessful when a single band of the expected size wasobserved in the agarose gel. PCR products were purifiedusing the ExoSAP-IT kit following the instructions of themanufacturer (Affymetrix). Bidirectional sequencing wasperformed with an ABI 3130xl Genetic Analyzer usingthe BigDye v1.1 kit; precipitation and purification weredone following the instructions of the manufacturer usingformamide, ethanol and ethylenediaminetetraacetic acid.
DATA ANALYSES AND PHYLOGENETIC
RECONSTRUCTION
The chromatograms were used for the sequence qualitycontrol and sequences were edited by hand when neces-sary (i.e., gaps, double peaks). Forward and reverse se-quences were assembled with the software Chromas Litev2.01 (available online at www.technelysium.com.au) anda database of all the sequences was built in BioEdit v7.0.5(Hall, 1999).
The quality of the obtained sequences was individuallyassured by visual inspection and by a BLAST searchin GenBank; all the sequences had high identity toother nematode sequences. In addition, 21 desmodoridSSU rDNA sequences and 27 COI sequences of marinenematodes from GenBank were included in the analyses.Sequences were aligned using ClustalW in the softwareMEGA 5.10 (Tamura et al., 2011) with the defaultparameters. The ends of the obtained sequences weretrimmed (including primer sequences) and we deletedmanually the gaps, ambiguously aligned regions andidentical haplotypes. As COI is a protein coding gene,COI sequences were checked for stop-codons using theDrosophila mitochondrial DNA as genetic code in thesoftware DnaSP 5.10.1 (Librado & Rozas, 2009).
MEGA 5.10 was used for the selection of the best nu-cleotide substitution model using the Bayesian Informa-tion Criterion. Phylogenetic trees were inferred for SSU
rDNA and COI separately using the maximum likelihood(ML) method. Branch support was estimated by boot-strapping over 100 resamplings. Phylogenetic trees wererooted using the midpoint rooting technique as imple-mented in MEGA 5.10. New sequences have been up-loaded to the European Nucleotide Archive; with acces-sion numbers LK54702-LK54728 (for SSU rDNA) andLK054668-LK054701 (for COI). Pairwise p-distancesbetween haplotypes were calculated in MEGA 5.10 forSSU rDNA and COI; distances were classified in threeexclusive categories: conspecific, interspecific congenericand interspecific non-congeneric. The definition of thesecategories was based on the identity of the morphospeciesas resulted of the LM microscope observations. We re-frained from any nomenclatural change of these morphos-pecies even when they result in paraphyletic species/genusin the phylogenetic tree.
For the supra-familial classification, we follow De Ley& Blaxter (2004) and Hodda (2011); for the intra-familyclassification we follow Lorenzen (1994).
Results
SSU RDNA
We obtained 27 new SSU rDNA sequences of whichseveral belong to five genera (Cheironchus Cobb, 1917;Gomphionema Wieser & Hopper, 1966; Longicyatho-laimus Micoletzky, 1924; Steineria Micoletzky, 1922; andViscosia de Man, 1889) that have not been previouslysequenced. The final alignment comprised 51 sequencesof 586 bp of which 235 positions were parsimony infor-mative. The best nucleotide substitution model was theKimura 2-parameter with non-uniform evolutionary rates(K2 + G).
The ML tree recovered several clades of marine ne-matodes (Fig. 1). Enoplida was recovered as a clade butChromadorida could not be recovered as a well-supportedmonophyletic group. The order Monhysterida was recov-ered as a clade; the exception was Sphaerolaimus sp.NN004 which was not placed within the clade. The fam-ilies Comesomatidae and Xyalidae were placed as sistergroups within the order. The families Desmodoridae andOncholaimidae were also supported as clades. The generaLongicyatholaimus and Cheironchus were supported asmonophyletic taxa.
The Desmodoridae formed a well-supported clade,but each of the three subfamilies in our study (i.e.,Desmodorinae, Spiriniinae and Stilbonematinae) were not
Vol. 16(8), 2014 981

M. Armenteros et al.
Fig. 1. Maximum likelihood tree of marine nematode species based on 51 DNA sequences of SSU rDNA. The root of the tree is placedbased on midpoint rooting technique. Numbers above/below the branches indicate branch support based on 100 bootstrap replicates(values < 75% are not shown). Taxa above the family level are indicated by capital letters on the nodes (A: Chromadorida, B: Enoplida,C: Monhysterida). Families are indicated by square brackets. Sequences obtained in this study are coded as NN.
monophyletic (Fig. 2). Even at the genus level there wereseveral non-monophyletic taxa and/or without bootstrapsupport, such as Eubostrichus Greeff, 1869; Laxus Cobb,1894; Robbea Gerlach, 1956; Spirinia Gerlach, 1963; andStilbonema Cobb, 1920. A double checking of the identityof these species was made based on photo-vouchers inorder to confirm that no misidentified specimens existedin the tree.
Three sequences (Desmodora_pontica_AM234628.1,Spirinia_parasitifera_AY854217.1 and Spirinia_parasiti-fera_EF527426.1) from GenBank showed abnormallyhigh values of conspecific distances; we did not includethem in the descriptive statistics or in the histogram sincethere was no way to check for possible misidentification.Pairwise p-distances overlapped across the three cate-
gories: conspecific (mean = 0.022; range = 0.011-0.038),congeneric (mean = 0.054; range = 0.024-0.075) andnon-congeneric (mean = 0.12; range = 0.015-0.25). Thedistribution of the pairwise p-distances for SSU rDNA didnot show a clear gap between the three categories (i.e.,conspecific, congeneric and non-congeneric distances); infact, non-congeneric distances showed a bimodal distribu-tion (Fig. 3).
COI
COI has been less studied than SSU rDNA in Nema-toda, with 2818 sequences belonging to 942 species vs15 415 sequences belonging to 3541 species, respectively(search in GenBank on 16 May 2014). So, with this study,
982 Nematology

Systematics of free-living marine nematodes
Fig. 2. A section of the maximum likelihood tree (SSU rDNA) in Figure 1 showing only the family Desmodoridae. Numbersabove/below the branches indicate branch support based on 100 bootstrap replicates (values < 75% are not shown). Subfamilymembership is indicated by colour: Stilbonematinae (red), Desmodorinae (blue) and Spiriniinae (green). Sequences obtained in thisstudy are coded as ND.
Vol. 16(8), 2014 983

M. Armenteros et al.
Fig. 3. Relative frequencies of pairwise p-distances for SSUrDNA (top) and COI (bottom) genetic markers.
we added 34 new COI sequences to this dataset; includ-ing six species of desmodorids not previously sequenced:Catanema exile, Desmodora pontica, Paradesmodora im-mersa, Robbea porosum, Stilbonema brevicolle and Za-lonema sp. The final alignment comprised 61 sequencesof 393 bp of which 291 positions were parsimony infor-mative. The nucleotide substitution model that best fittedthe data was the General Time Reversible model with non-uniform evolutionary rates and invariant sites (GTR +G + I).
The ML tree based on COI did not provide sup-port for the deep relationships in the phylogenetic back-bone of Nematoda (Fig. 4). Four families were recov-ered as well-supported clades: Xyalidae, Chromadoridae,Cyatholaimidae and Oncholaimidae. Thoracostomopsi-dae were non monophyletic because Calyptronema Mar-ion, 1870, Enoploides Ssaweljev, 1912 and TrileptiumCobb, 1933 were placed outside the clade. DichromadoraKreis, 1929 (Chromadoridae) was non-monophyletic. In
general, for the groups outside of Desmodoridae, therewas a good recovery of the relationships between speciesusing the COI marker. The family Desmodoridae couldnot be recovered as a well-supported clade.
Within Desmodoridae, the three subfamilies (i.e., Des-modorinae, Spiriniinae and Stilbonematinae) appearednon-monophyletic (Fig. 5). Specimens were clusteredconsistently within the genera Zalonema Cobb, 1920 andParadesmodora Schuurmans Stekhoven, 1942. However,Stilbonema brevicolle was unresolved because of lowbootstrap support. Divergent lineages within morpho-species were found for Robbea porosum.
The analysis of conspecific p-distances had a very widerange when specimens were classified based solely onmorphology (mean = 0.14; range = 0.0028-0.29). How-ever, we detected two lineages which are probably crypticspecies (R. porosum) and also detected one paraphyleticspecies (S. brevicolle) (Fig. 5). If specimens belongingto these species are removed from the pairwise conspe-cific group, the values were more consistent (mean =0.015; range = 0.0028-0.024). The congeneric distanceshad higher values (mean = 0.19; range = 0.14-0.28) andnon-congeneric (mean = 0.32; range = 0.15-0.47). Thehistogram of the COI pairwise p-distances with the re-moved values showed a clear gap between the conspecific(<2.5% genetic divergence) and the other two groups; thecongeneric distances had some overlapping with those ofthe non-congeneric ones (Fig. 3).
Discussion
The phylogenetic relationships based on SSU rDNAlargely agree with those of van Megen et al. (2009). Theposition of Comesomatidae has been uncertain (Holter-man et al., 2008), but our results support its membershipto the Monhysterida which is in agreement with Lorenzen(1994) and van Megen et al. (2009). The monhysteridsincluded in our study support the inclusion of Comeso-matidae and Xyalidae in a Monhysterida-dominated clade(5A sensu Holterman et al., 2008).
Our results confirm the monophyly of the Desmodor-idae (Meldal et al., 2007, 2009). Conversely, the ma-rine desmodorid subfamilies Desmodorinae and Spirini-inae were non-monophyletic in agreement with previousstudies (Holterman et al., 2008; van Megen et al., 2009;Armenteros et al., 2014). Consistently with our findings,no defining synapomorphy has been reported for thesetwo subfamilies. Stilbonematinae have been reported tobe monophyletic (Kampfer et al., 1998) and indeed most
984 Nematology
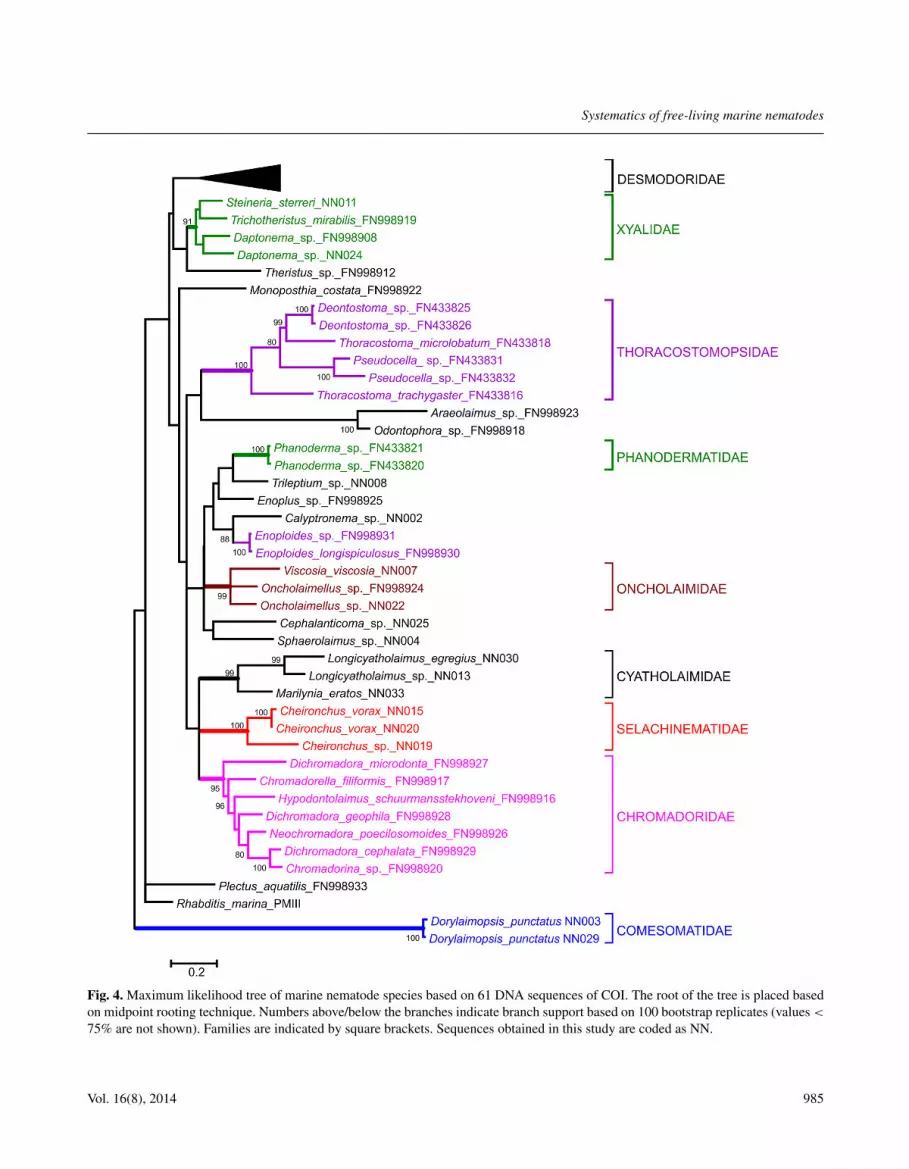
Systematics of free-living marine nematodes
Fig. 4. Maximum likelihood tree of marine nematode species based on 61 DNA sequences of COI. The root of the tree is placed basedon midpoint rooting technique. Numbers above/below the branches indicate branch support based on 100 bootstrap replicates (values <
75% are not shown). Families are indicated by square brackets. Sequences obtained in this study are coded as NN.
Vol. 16(8), 2014 985

M. Armenteros et al.
Fig. 5. A section of the maximum likelihood tree (COI) in Figure 4 showing only the family Desmodoridae. Numbers above/below thebranches indicate branch support based on 100 bootstrap replicates (values < 75% are not shown). Subfamily membership is indicatedby colour: Stilbonematinae (red), Desmodorinae (blue) and Spiriniinae (green). Sequences obtained in this study are coded as ND.
of the stilbonematin species in our analysis clustered to-gether in both trees, thereby supporting this hypothesis.However, there were some specimens placed out of thecluster that we consider questionable because of: i) theconsiderable difficulties to identify some specimens be-cause the coiled habitus; and ii) there are some apparentlyanomalous DNA sequences. Actually, the inclusion ofmost of the SSU rDNA sequences from GenBank by Ar-menteros et al. (2014) notably weakened the phylogeneticsignal of Stilbonematinae and as result they concludedthat the subfamily was non-monophyletic. However, Stil-bonematinae have been characterised by two synapomor-phies: glandular sensory organs (Bauer-Nevelsick et al.,1995) and ectosymbiotic chemoautotrophic bacteria onthe cuticle (Polz et al., 1992). Thus, we support the hy-pothesis of monophyly of Stilbonematinae based on ourmolecular evidence and consider the existence of possiblemisidentification of specimens. Finally, future studies to
disentangle the systematics within Desmodoridae shouldinclude other species and genera.
Several desmodorid SSU rDNA sequences from Gen-Bank were apparently anomalously placed in the treein the initial performed phylogenetic reconstruction(e.g., Robbea sp. JN968260.1 and Spirinia parasitiferaJN968278.1). The original publications did not provideenough detail to check the species identification of thesesequences and therefore were not included in the tree.The vouchering of sequenced nematode specimens withimages (photos or videos), as suggested by De Ley etal. (2005) is essential for the interpretation and subse-quent validation of phylogenetic analyses. For instance,the relatively high value of the p-distance (0.20) betweenspecimens NN019 and NN020 of Cheironchus spp. sug-gested that different species may be involved. Furtherscrutiny of photo-vouchers from several specimens indi-cated that there were indeed differences in the numbers of
986 Nematology

Systematics of free-living marine nematodes
pre-cloacal supplements (12 vs 13) and numbers of turnsand size of the amphidial fovea (5.5 vs 3.5 turns). We la-belled a posteriori NN019 as Cheironchus sp. and it maybe a new species within the genus.
The taxa that were morphologically identified andwhich had not been previously sequenced (e.g., Steineria,Zalonema) were placed consistently within clades repre-senting the families. This result supports the high congru-ence between the morphology- and DNA-based systemat-ics at family-level of marine nematodes (van Megen et al.,2009; Derycke et al., 2010a).
SSU rDNA has been useful to barcode nematodegroups with high nucleotide substitution rates (e.g., ani-mal- and plant-parasite species) (Holterman et al., 2006),although it is considerably less informative for discrimi-nating closely related marine nematode taxa (see Bhaduryet al., 2006). The use of the I3-M11 partition of the COIgene, as suggested by Derycke et al. (2010a), clearlyperforms much better than SSU rDNA in discriminatingspecies of genera such as Cheironchus, Dichromadora,Enoploides and Thoracostoma Marion, 1870. The locus isrelatively easy to amplify using JB3-JB5 primers and weobtained a 73% of amplification success without attempt-ing to improve the protocols of Derycke et al. (2010a),even if these authors had a higher success: 87.8%.
Our study supports the usefulness of COI for prospect-ing nematode species and we obtained evidence of cryp-tic diversity that could be further explored based on thevoucher photographs. We found divergent genetic lin-eages within the morphospecies R. porosum and S. bre-vicolle which are flagged as putative cryptic species. Forsome taxa, a posteriori analysis based on photo vouch-ers revealed morphological divergence (e.g., Cheironchus,Desmodora, Paradesmodora), but for other taxa we couldnot morphologically differentiate nominal species whichwere divergent in the tree. Evidence of cryptic diversityhas been published for several marine species, namely:Litoditis marina (Derycke et al., 2005), Thoracostomatrachygaster (Derycke et al., 2010b; Silva de Oliveiraet al., 2012), Halomonhystera disjuncta (Derycke et al.,2007) and Terschellingia longicaudata (Bhadury et al.,2008).
The existence of substantial cryptic diversity in nema-todes may be explained by a combination of: i) conservedmorphology (Holterman et al., 2008); ii) fast nucleotidesubstitution rates both in mitochondrial and nuclear DNA(Blouin et al., 1998; Holterman et al., 2006; Gissi etal., 2008); and iii) restricted dispersal and gene flow atlarge geographical scales (Derycke et al., 2013). The lat-
ter point is less relevant in our study given the small areain which the samples were taken (i.e., <10 m distanceamong cores). However, there is also the possibility ofspeciation in other regions and secondary contact of thesympatric species.
COI separated clearly conspecific from interspecificsequences (i.e., barcoding gap). All conspecific geneticdistances were <3% of genetic divergence, whereas allinterspecific distances (within and among genera) were>14%. Therefore, the threshold value of 5% of geneticdivergence suggested by Derycke et al. (2010a) also holdsin our study. COI seems promising for DNA barcodingof nematodes, which is in agreement with previousstudies (e.g., Derycke et al., 2010a; Siddall et al., 2012).However, the main limitation for the implementationof COI barcoding for marine nematodes is the poorknowledge of the diversity and taxonomy of the group(Bucklin et al., 2011) and the lack of a good referencedatabase of vouchered COI sequences.
In summary, our main conclusions are: i) the mono-phyly of Enoplida is re-affirmed, with Chromadorida asa sister group; ii) the family Comesomatidae is a sistergroup of Xyalidae within Monhysterida; iii) both DNAmarkers confirm the congruence between morphology-and molecular-based phylogenetic inferences for mostof the families included in our study, although non-congruence occurs at intra-familial level (i.e., crypticspecies); iv) Desmodoridae is a monophyletic group, butthe relationships within the family could not be recovered;the subfamilies Desmodorinae and Spiriniinae were non-monophyletic meanwhile the monophyly of Stilbonemati-nae was not fully supported due to a few specimens ofwhich the identification could be questioned; and v) cur-rently, COI is effective to explore cryptic diversity andbarcode species within Nematoda, providing a good ref-erence database is available.
Acknowledgements
Arantza Elejalde, Annelien Rigaux and Karin Breugel-mans are acknowledged for their help with the laboratorywork. We also thank Harald Gruber-Vodicka for allow-ing us the use of Eubostrichus fertilis sequences. Thisresearch was partially funded by a 2010 Post-Doctoralfellowship from Belgian Science Policy to M.A. Fieldexpeditions were funded in the framework of WallaceaOperation Cuba 2009 and 2010. The authors thank twoanonymous reviewers whose comments improved themanuscript.
Vol. 16(8), 2014 987

M. Armenteros et al.
References
Aleshin, V.V., Kedrova, O.S., Milyutina, I.A., Vladychenskaya,N.S. & Petrov, N.B. (1998). Relationships among nematodesbased on the analysis of 18S rRNA gene sequences: molec-ular evidence for monophyly of chromadorian and secernen-tian nematodes. Russian Journal of Nematology 6, 175-184.
Armenteros, M., Ruiz-Abierno, A., Sosa, Y. & Pérez-García,J.A. (2012). Habitat heterogeneity effects on macro- andmeiofauna (especially nematodes) in Punta Francés coralreef (SW Cuban Archipelago). Revista de InvestigacionesMarinas. Universidad de La Habana 32, 50-61.
Armenteros, M., Ruiz-Abierno, A. & Decraemer, W. (2014).Taxonomy of Stilbonematinae (Nematoda: Desmodoridae):description of two new and three known species and phylo-genetic relationships within the family. Zoological Journal ofthe Linnean Society 171, 1-21.
Bauer-Nebelsick, M., Blumer, M., Urbancik, W. & Ott, J.A.(1995). The glandular sensory organ of Desmodoridae (Ne-matoda) – ultrastructure and phylogenetic implications. In-vertebrate Biology 114, 211-219.
Bert, W., Karssen, G. & Helder, J. (2011). Phylogeny and evo-lution of nematodes. In: Jones, J., Gheysen, G. & Fenoll, C.(Eds). Genomics and molecular genetics of plant-nematodeinteractions. Berlin, Germany, Springer.
Bhadury, P., Austen, M.C., Bilton, D.T., Lambshead, P.J.D.,Rogers, A.D. & Smerdon, G.R. (2006). Development andevaluation of a DNA-barcoding approach for the rapid iden-tification of nematodes. Marine Ecology Progress Series 320,1-9.
Bhadury, P., Austen, M.C., Bilton, D.T., Lambshead, P.J.D.,Rogers, A.D. & Smerdon, G.R. (2008). Evaluation of com-bined morphological and molecular techniques for marine ne-matode (Terschellingia spp.) identification. Marine Biology154, 509-518.
Blaxter, M.L., De Ley, P., Garey, J.R., Liuk, L.X., Scheldeman,P., Vierstraete, A., Vanfleteren, J.R., Mackey, L.Y., Dorris,M., Frissei, L.M. et al. (1998). A molecular evolutionaryframework for the phylum Nematoda. Nature 392, 71-75.
Blouin, M.S., Yowell, C.A., Courtney, C.H. & Dame, J.B.(1998). Substitution bias, rapid saturation, and the use ofmtDNA for nematode systematics. Molecular Biology andEvolution 15, 1719-1727.
Bucklin, A., Steinke, D. & Blanco-Bercial, L. (2011). DNAbarcoding of marine Metazoa. Annual Review of MarineScience 3, 471-508.
De Ley, P. & Blaxter, M. (2004). A new system for Nematoda:combining morphological characters with molecular trees,and translating clades into ranks and taxa. In: Cook, R. &Hunt, D. J. (Eds). Proceedings of the Fourth InternationalCongress of Nematology, 8-13 June 2002, Tenerife, Spain.Nematology Monographs and Perspectives 2, pp. 633-653.Leiden, The Netherlands, Brill.
De Ley, P., De Ley, I.T., Morris, K., Abebe, E., Mundo-Ocampo,M., Yoder, M., Heras, J., Waumann, D., Rocha-Olivares, A.,Burr, A.H.J. et al. (2005). An integrated approach to fastand informative morphological vouchering of nematodes forapplications in molecular barcoding. Philosophical Transac-tions of the Royal Society of London. Series B: Biological Sci-ences 360, 1945-1958.
Derycke, S., Remerie, T., Vierstraete, A., Backeljau, T., Van-fleteren, J., Vincx, M. & Moens, T. (2005). Mitochon-drial DNA variation and cryptic speciation within the free-living marine nematode Pellioditis marina. Marine EcologyProgress Series 300, 91-103.
Derycke, S., Backeljau, T., Vlaeminck, C., Vierstraete, A., Van-fleteren, J., Vincx, M. & Moens, T. (2007). Spatiotemporalanalysis of population genetic structure in Geomonhysteradisjuncta (Nematoda, Monhysteridae) reveals high levels ofmolecular diversity. Marine Biology 151, 1799-1812.
Derycke, S., Vanaverbeke, J., Rigaux, A., Backeljau, T. &Moens, T. (2010a). Exploring the use of cytochrome oxidasec subunit 1 (COI) for DNA barcoding of free-living marinenematodes. PloS One 5, e13716.
Derycke, S., De Ley, P., Tandingan De Ley, I., Holovachov, O.,Rigaux, A. & Moens, T. (2010b). Linking DNA sequencesto morphology: cryptic diversity and population geneticstructure in the marine nematode Thoracostoma trachygaster(Nematoda, Leptosomatidae). Zoologica Scripta 39, 276-289.
Derycke, S., Backeljau, T. & Moens, T. (2013). Dispersaland gene flow in free-living marine nematodes. Frontiers inZoology 10, 1-12.
Gissi, C., Iannelli, F. & Pesole, G. (2008). Evolution of the mito-chondrial genome of Metazoa as exemplified by comparisonof congeneric species. Heredity 101, 301-320.
Hall, T.T. (1999). BioEdit: a user-friendly biological se-quence alignment editor and analysis program for Windows95/98/NT. Nucleic Acids Symposium Series 41, 95-98.
Hodda, M. (2011). Phylum Nematoda Cobb 1932. In: Zhang,Z.-Q. (Ed.). Animal biodiversity: an outline of higher-levelclassification and survey of taxonomic richness. Zootaxa,3148, pp. 63-95.
Holterman, M., van der Wurff, A., van den Elsen, S., van Megen,H., Bongers, T., Holovachov, O., Bakker, J. & Helder, J.(2006). Phylum-wide analysis of SSU rDNA reveals deepphylogenetic relationships among nematodes and acceleratedevolution toward crown clades. Molecular Biology and Evo-lution 23, 1792-1800.
Holterman, M., Holovachov, O., van den Elsen, S., van Megen,H., Bongers, T., Bakker, J. & Helder, J. (2008). Small subunitribosomal DNA-based phylogeny of basal Chromadoria (Ne-matoda) suggests that transitions from marine to terrestrialhabitats (and vice versa) require relatively simple adaptations.Molecular Phylogenetics and Evolution 48, 758-763.
Kampfer, S., Sturmbauer, C. & Ott, J.A. (1998). Phylogeneticanalysis of rDNA sequences from adenophorean nematodes
988 Nematology

Systematics of free-living marine nematodes
and implications for the Adenophorea-Secernentea contro-versy. Invertebrate Biology 117, 26-36.
Librado, P. & Rozas, J. (2009). DnaSP v5: a software for com-prehensive analysis of DNA polymorphism data. Bioinfor-matics 25, 1451-1452.
Lorenzen, S. (1994). The phylogenetic systematics of freelivingnematodes. London, UK, The Ray Society.
Margulis, L. & Chapman, M.J. (2009). Kingdoms and domains:an illustrated guide to the phyla of life on Earth. San Diego,CA, USA, Academic Press.
Meldal, B.H.M., Debenham, N.J., De Ley, P., Tandingan De Ley,I., Vanfleteren, J.R., Vierstraete, A.R., Bert, W., Borgonie, G.,Moens, T., Tyler, P.A. et al. (2007). An improved molecularphylogeny of the Nematoda with special emphasis on marinetaxa. Molecular Phylogenetics and Evolution 42, 622-636.
Polz, M., Felbeck, H., Novak, R., Nebelsick, M. & Ott, J.A.(1992). Chemoautotrophic, sulfur-oxydizing symbiotic bac-teria on marine nematodes: morphological and biochemicalcharacterization. Microbial Ecology 24, 313-329.
Rodrigues Da Silva, N.R., Da Silva, M.C., Fonseca-Genevois,V.G., Esteves, A.M., De Ley, P., Decraemer, W., Rieger,T.T. & Dos Santos Correia, M.T. (2010). Marine nematodetaxonomy in the age of DNA: the present and future ofmolecular tools to assess their biodiversity. Nematology 12,661-672.
Siddall, M.E., Kvist, S., Phillips, A. & Oceguera-Figuero, A.(2012). DNA barcoding of parasitic nematodes: is it Kosher?Journal of Parasitology 98, 692-694.
Silva De Oliveira, D.A., Decraemer, W., Holovachov, O., Burr,J., Tandingan De Ley, I., De Ley, P., Moens, T. & Derycke,
S. (2012). An integrative approach to characterize crypticspecies in the Thoracostoma trachygaster Hope, 1967 com-plex (Nematoda: Leptosomatidae). Zoological Journal of theLinnean Society 164, 18-35.
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M. &Kumar, S. (2011). MEGA5: molecular evolutionary geneticsanalysis using maximum likelihood, evolutionary distance,and maximum parsimony methods. Molecular Biology andEvolution 28, 2731-2739.
Tchesunov, A.V. (2013). Marine free-living nematodes of thesubfamily Stilbonematinae (Nematoda, Desmodoridae): taxo-nomic review with descriptions of a few species from the NhaTrang Bay, Central Vietnam. Meiofauna Marina 20, 71-94.
van Megen, H., van den Elsen, S., Holterman, M., Karssen,G., Mooyman, P., Bongers, T., Holovachov, O., Bakker, J. &Helder, J. (2009). A phylogenetic tree of nematodes basedon about 1200 full-length small subunit ribosomal DNAsequences. Nematology 11, 927-950.
Verschelde, D., Gourbault, N.E. & Vincx, M. (1998). Revisionof Desmodora with description of new Desmodorids (Nema-toda) from Hydrothermal Vents of the Pacific. Journal of theMarine Biological Association of the United Kingdom 78, 75-112.
Yoder, M., De Ley, I.T., King, I.W., Mundo-Ocampo, M.,Mann, J., Blaxter, M., Poiras, L. & De Ley, P. (2006).DESS: a versatile solution for preserving morphology andextractable DNA of nematodes. Nematology 8, 367-376.
Zhang, Z.-Q. (2011). Animal biodiversity: an introduction tohigher-level classification and taxonomic richness. Zootaxa3148, 7-12.
Vol. 16(8), 2014 989