xerocomus cisalpinus sp. nov., and the delimitation of species in the x. chrysenteron complex based...
TRANSCRIPT

Xerocomus cisalpinus sp. nov., and the delimitation of
species in the X. chrysenteron complex based on morphology
and rDNA-LSU sequences*
Ursula PEINTNER1#, Heidi LADURNER1 and Giampaolo SIMONINI2
1 Institute of Microbiology, University of Innsbruck, Technikerstrasse 25, 6020 Innsbruck, Austria.2Via Bellaria 8, 42100 Reggio Emilia, Italy.E-mail : [email protected]
Received 6 June 2002; accepted 31 March 2003.
Species delimitation is still controversial in the Xerocomus chrysenteron complex. We have therefore established
comprehensible and reliable species concepts based on statistical evaluation of morphological and ecological characters.We examined many collections from different geographical regions and different developmental stages withincollections. Quantitative micromorphological characters (basidiospores, pileipellis end cells) were measured in statistically
relevant numbers. The same material was used to generate 24 rDNA-LSU sequences, and the results of phylogeneticanalyses clearly confirmed our species concepts : spore size and ornamentation, length of the pileipellis end cells and‘pruinatus-hyphae’ are most valuable characters for the delimitation of species in this complex. Molecular data
demonstrated that the X. chrysenteron complex is a monophyletic group. All the examined species (X. chrysenteron,X. cisalpinus, X. pruinatus, X. ripariellus, X. dryophilus, X. fennicus, X. porosporus, and X. rubellus) represent independentlineages. The faintly striate spores, a key character characterising species of section Striatulispori, probably evolvedindependently. In addition, the ‘pruinatus-hyphae’ have multiple origins, and truncate spore apices are derived at least
twice. Xerocomus cisalpinus sp. nov. is characterised by striate spores, the presence of ‘pruinatus-hyphae’ and a pileipellisstrongly reminiscent of X. chrysenteron. For reasons of discussion, microscopical data are presented on Boletellusepiscopalis for the first time. Xerocomus fennicus (Boletellus) comb. nov. is proposed. We provide descriptions to all
included taxa. Our results once more demonstrate that reliably identified and characterised voucher collections are thebasic requirement for meaningful phylogenetic studies.
INTRODUCTION
Distinction of species in the genus Xerocomus hasalways been critical. Due to the high number of newlydescribed taxa (Bresinsky & Schwarzer 1969, Heine-mann, Rammeloo & Rullier 1988, Oolbekkink 1991,Klofac & Krisai-Greilhuber 1992) a reliable identifi-cation of species within this group of boletes became acomparison between various mycological schools andphilosophies. These mycological schools used empiri-cally weighted morphological characters based on per-sonal experience, and due to different weighting ofdifferent characters there was little agreement onbroadly acceptable species concepts. During our earlierstudies (Simonini 1996, 1998a, Ladurner 2001a, b), wedeveloped species concepts for taxa of the X. chry-senteron group based on a ‘fuzzy’ set of morphologicaland ecological characters. Subsequent to this, a small
bolete macroscopically strongly recalling X. chry-senteron was repeatedly collected in Italy during ourstudies of Xerocomus in southern Europe. Microscopi-cally, these collections exhibited intermediate charac-ters between X. pruinatus and X. chrysenteron andcalled into question our earlier species concepts. Thespores of these new collections were finely longi-tudinally striate and the typical, ‘pruinatus-hyphae’(Fig. 1), thick-walled, amyloid hyphae in the stipecontext (Ladurner & Poder 2000), were constantly ob-served. Both characters were not compatible with ourinterpretation of X. chrysenteron. In addition, thepileipellis structure of these collections did not matchthe species concept of X. pruinatus in Europe, whichwas broadly accepted only recently (Klofac & Krisai-Greilhuber 1992, Engel et al. 1996, Simonini 1998b).The striking combination of characters exhibited bythese new collections was not compatible with anyEuropean taxon with striate spores.
In order to verify our species concepts and to resolvethe puzzling question of whether or not this taxon was
* We dedicate this paper to Reinhold Poder on the occasion of his55th birthday.# Corresponding author.
Mycol. Res. 107 (6): 659–679 (June 2003). f The British Mycological Society 659
DOI: 10.1017/S0953756203007901 Printed in the United Kingdom.

a variety of X. pruinatus, to which it seemed closest, itstaxonomic position was investigated using moleculartechniques. Nuclear ribosomal DNA sequences of thelarge subunit (nLSU-rDNA) have proven to be mostuseful for molecular systematics of agarics and boletesat order, genus and species level (Binder & Fischer1998, Drehmel, Moncalvo & Vilgalys 1999, Moncalvoet al. 2000a, b, 2002, Grubisha et al. 2001, Humpertet al. 2001, Jarosch & Besl 2001, Binder & Hibbett2001, 2002). For this phylogenetic study we generatednumerous nLSU-rDNA sequences from the same col-lections that we had studied previously to establish ourmorphological and ecological species concepts.
Modern species delimitation in Xerocomus is now, inaddition to the classical macro-morphological charac-ters, based on measurable microscopical characterssuch as spore morphology and pileipellis structure.Currently, the X. chrysenteron group circumscribessix species, three of them (X. fennicus, X. pruinatus, X.ripariellus) with striate spores. For these species the newsection Striatulispori was recently erected (Redeuilh1998). We therefore also wanted to determine if sporestriation is a valuable, synapomorphic character for thedelimitation of groups within Xerocomus.
According to phylogenetic analyses of our moleculardata, the intermediate Italian collections represent anew, independent lineage within the X. chrysenteroncomplex. These collections can also be separatedmorphologically, we therefore describe Xerocomuscisalpinus sp. nov., and discuss its phylogenetic posi-tion. In addition, the distinguishing morphological andecological characters of taxa belonging to the X. chry-senteron complex are described and discussed.
MATERIALS AND METHODS
Macro- and microscopical characterisation
Comprehensible and reliable species concepts weredeveloped based on all available traditional methods(including statistics). Our concepts are based on theexamination of numerous collections from differentgeographical regions and on the study of differentdevelopmental stages within one collection. Quantitat-ive micromorphological characters (dimensions of theterminal pileipellis elements, spores, cystidia) weremeasured in statistically relevant numbers (n=31). Allcollections were carefully and consistently screened forqualitative characters such as the type of the pileipellis,pigmentation, spore ornamentation, microchemicalreactions, etc. These data were combined with therespective ecological data (associated trees, fruitingperiod, edaphic factors, etc.).
Colour descriptions of the basidiomes were usuallymade according to Kornerup & Wanscher (1978). Ad-ditionally, the colour code Seguy (1936) was used; inthis case, the letter ‘S’ precedes the colour codes.
The specimens were examined with standard micro-scopic techniques. Microscopic data were documentedby video prints produced with a Sony video cameraSASC-C350P and a Sony video printer UP 910. Sporesand hymenophoral elements were mounted in 3%KOH and measured on prints at 2900rmagnification.Pileipellis samples were studied in Congo red. Stipetissue was first treated with Melzer’s then washed in aconcentrated chlorale-hydrate solution for the exam-ination of the ‘pruinatus-hyphae’ (Ladurner & Poder2000). For statistical evaluation, 31 spores and 31terminal elements of the pileipellis were measured foreach collection. Measurements of spores and terminalelements of the pileipellis are given as (minimum)average¡standard deviation (maximum). The sporequotient (Q) is the ratio of spore length to width(Q=l/w). The spore volume is calculated as follows:V=4/3((l/2)*(w*w/2))*3.14/2.
For scanning electron microscopy (SEM), driedspecimens were rehydrated in 25% ammonia solutionfor 1 h, dehydrated in 70% aqueous ethanol for 1 h,fixed in pure 1,2-dimethoxy-methane for 1.5 h and thenimmersed in pure acetone for at least 2 h. After criticalpoint drying, samples were mounted on aluminiumporters and sputtered with gold. Micrographs wereprepared using a Zeiss DSM-950 SEM.
Fig. 1. ‘Pruinatus-hyphae’ are thick walled, amyloid hyphaein the stipe context as typically found in Xerocomus pruinatus.
‘Pruinatus-hyphae’ extend more or less abruptly to strikinglythick-walled, to 30 mm broad, irregularly formed hyphae ofvariable length and show a weak to strong amyloid reaction in
thewall dependingon their developmental stage.The inner sur-face has crater-like ornamentation. Nomarski. Bar=10 mm.
Xerocomus cisalpinus sp. nov. and the X. chrysenteron complex 660

Statistical analyses
Statistics were carried out with the programSYSTAT10 (SPSS, Chicago, IL). 28 collections ofXerocomus chrysenteron, 18 collections of X. cisalpinus,and 25 collections of X. pruinatus were studied. Collec-tions were chosen in order to cover the widest possiblegeographic range of these taxa. Four biometrical vari-ables were considered: spore length (SPOLE) andwidth (SPOWI) as well as length and width of thepileipellis end cells (ENDLE, ENDWI). Mean values ofeach variable were calculated for each collection, re-spectively. Two sample t tests were performed with twosamples belonging to two different taxa in order to testthe null hypothesis that these biometrical variablesare not significantly different. Biometric variables canbe considered as useful to separate taxa when the nullhypothesis is rejected (P<0.005).
A discriminant analysis using complete estimation ofspore length, spore width, end cells length and end cellswidth has been performed. The aim of this analysis wasto firstly find a linear combination of the four variablesthat best discriminate among the three taxa, and sec-ondly to check if these three species groups could be
separated, and to thirdly find which variables are mostsignificant for distinction of taxa.
Molecular techniques
In order to establish the phylogenetic relationshipsof Xerocomus cisalpinus, nLSU-rDNA sequences wereproduced from the type material and three collec-tions from other localities. All together, 34 sequenceswere used for the phylogenetic analyses: 26 sequenceswere generated from the same material used to estab-lish our morphology- and ecology-based species con-cepts in the X. chrysenteron species complex. Eightadditional sequences were retrieved from GenBank.The material used is listed in Table 1, together with therespectiveGenBank accession numbers, collection num-bers, and information concerning the geographicalorigin and ecology.
DNA was isolated from dried herbarium materialfollowing standard protocols (Zolan & Pukkila 1986).Primers used for PCR amplification and sequencing ofthe nLSU-rDNA were LROR, LR3, LR5, LR3R(Moncalvo et al. 2000b). Amplifications were carriedout in a 25 ml reaction mix under standard conditions
Table 1. Material included in the phylogenetic analysis of the Xerocomus chrysenteron complex with the respective GenBank accession
numbers for rDNA-LSU sequences, the country of provenance and the potential host trees. When available, herbarium numbers are given
after the respective species epithet. Some sequences were retrieved from GenBank.
Species and herbarium number GenBank accession no. Country Potential host trees
X. chrysenteron IB19990951 AF514808 Austria Picea
X. chrysenteron IB19990952 AF514807 Austria Picea
X. chrysenteron IB20000405 AF514809 Austria Pinus, Picea
X. chrysenteron AF347103 Switzerland Mixed forest
X. ‘chrysenteron ’ AF071537 USA Unknown
X. ‘chrysenteron ’ AF050647 Germany Picea, Fagus
X. cisalpinus IB19900700 AF514814 Italy Quercus ilex
X. cisalpinus IB19980850 AF514815 Italy Juniperus, Fagus
X. cisalpinus IB20000700 AF514813 Italy Quercus suber
X. cisalpinus IB20000701 AF514812 Italy Quercus suber
X. dryophilus IB19990901 AF514823 Croatia Quercus ilex
X. dryophilus IB19991057 AF514822 Spain Pinus halepensis
X. fennicus Hel AF514820 Finland Betula, Picea
X. fennicus OULU AF514821 Finland Unknown
X. porosporus AF050645 Germany Unknown
X. porosporus IB19880304 AF514810 Spain Cedrus, Fraxinus
X. porosporus IB19990957 AF514811 Italy Quercus
X. pruinatus AF050645 Germany Quercus, Acer
X. pruinatus AF050644 Germany Mixed forest
X. pruinatus IB19920266 AF514824 Ukraine Fagus
X. pruinatus IB19950970 AF514826 Austria Fagus, Picea
X. pruinatus IB19961055 AF514825 Italy Quercus, Castanea
X. pruinatus IB19980366 AF514827 Austria Fagus, Picea
X. ‘pruinatus ’ AF402140 Switzerland Mixed forest
X. ripariellus GR21189 AF514817 France Unknown
X. ripariellus GR930920 AF514819 France Unknown
X. ripariellus IB19980360 AF514816 Spain Mixed hardwoods
X. ripariellus GR22465 AF514818 France Unknown
X. ‘rubellus ’ AF050649 Germany Mixed forest
X. rubellus GS1044 AF514830 Italy Quercus suber
X. rubellus GS961 AF514829 Italy Quercus cerris
X. rubellus IB19990917 AF514828 Croatia Quercus
X. subtomentosus AF139716 Germany Quercus
X. subtomentosus IB19980452 AF514831 Finland Pinus, Betula
X. subtomentosus IB19991000 AF514832 Italy Quercus pubescens
U. Peintner, H. Ladurner and G. Simonini 661
(Vilgalys &Hester 1990) in a PerkinElmer 9600 thermo-cycler (Perkin Elmer, Foster City, CA).
Sequencing was performed using fluorescent dyeterminator chemistries following the manufacturer’sinstructions (PerkinElmer) on automated sequencers(ABI 373A, ABI 377, PerkinElmer, Norwalk, CN). Se-quence chromatograms were compiled with Sequenchersoftware version 2.0 (Gene Codes, Ann Arbor, MI) orSequence Navigator (Perkin Elmer). Sequences weresubmitted to the GenBank under the accession nos.AF514807–AF514832; alignments are deposited inTreeBASE under study accession nos. S878 and matrixno. M1425.
Phylogenetic analyses
Phylogenetic analyses of 34 nLSU-rDNA sequencesbelonging to eight taxa of the Xerocomus chrysenteroncomplex were performed with PAUP* 4.0b8 (Swofford1998). Maximum parsimony was used to search foroptimal trees, employing the default PAUP* settings(i.e. mulpars=on, steepest descent not in effect, andmaxtrees=20000). Maximum parsimony was conduc-ted with a standard step matrix giving transitions twicethe weight of transversions. Analyses were carried outwith tree-bisection-connection (TBR) branch swap-ping, and with gaps treated as ‘missing’. Most parsi-monious trees (MPT) were found using 100 randomsequence addition replicates followed by a secondheuristic search using the trees from the previous searchas starting trees but applying a reconnection limit ofeight.
To evaluate branch robustness of trees generated byparsimony-based methods, bootstrap analyses (Fel-senstein 1985) were conducted using 500 replications,each consisting of ten heuristic searches with randomaddition sequences and TBR branch swapping.
The program Modeltest version 3.06 (Posada &Crandall 1998) was used to test the model of DNAsubstitution. The ML analysis under the GTR+Gmodel was performed with six substitution types anda user-specified substitution rate matrix, nucleotidefrequencies A=0.25450, C=0.21190, G=0.30560,T=0.22800, proportion of invariable sites=0.5965, rateheterogeneity following the discrete gamma approxi-mation with four categories and a shape parametera=0.7347. The heuristic search with TBR branchswapping used one MPT as starting tree. In addition,two hundred bootstrap replicates were run with ML(starting trees obtained via neighbour-joining, noTBR).
RESULTS
Statistical analyses of biometric variables
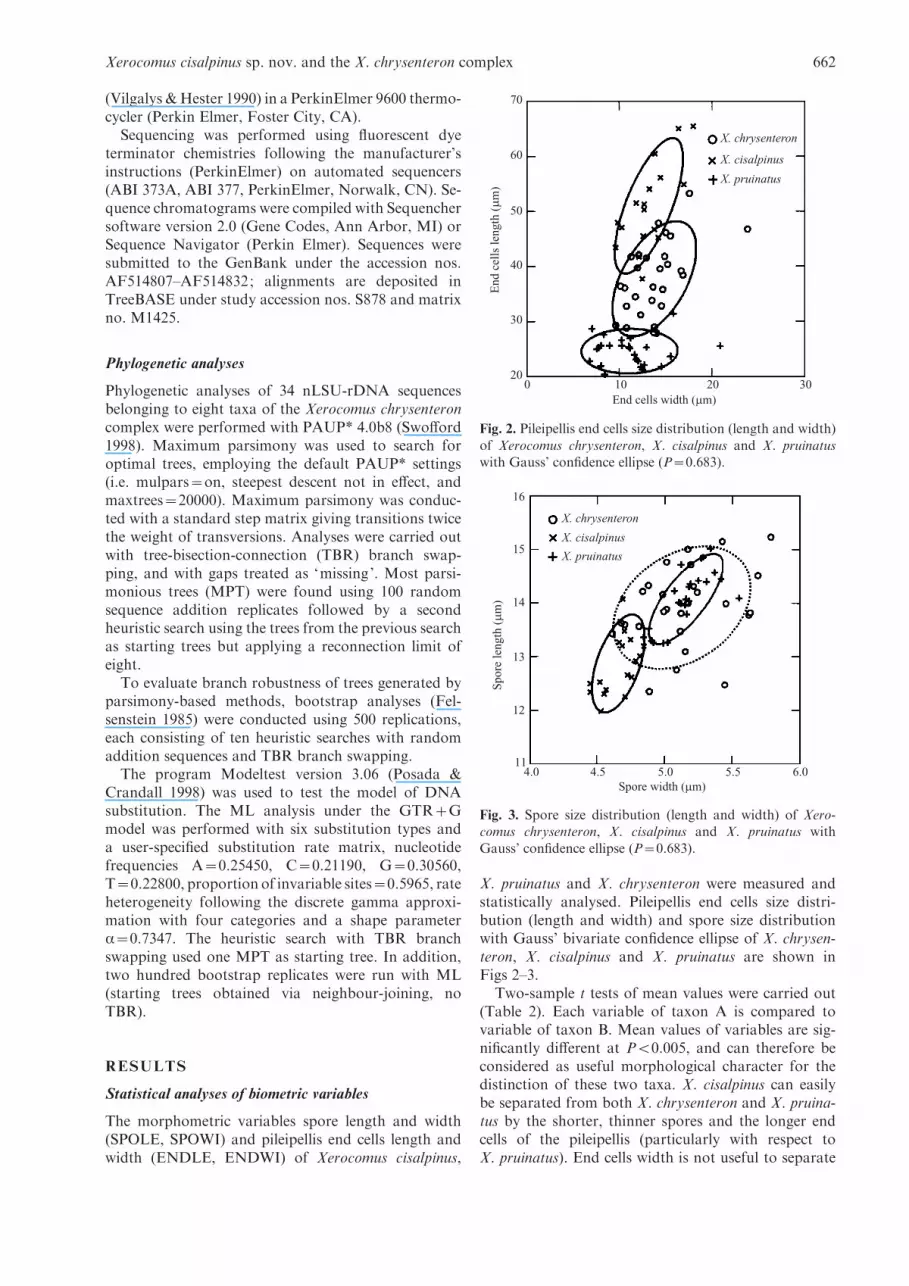
The morphometric variables spore length and width(SPOLE, SPOWI) and pileipellis end cells length andwidth (ENDLE, ENDWI) of Xerocomus cisalpinus,
X. pruinatus and X. chrysenteron were measured andstatistically analysed. Pileipellis end cells size distri-bution (length and width) and spore size distributionwith Gauss’ bivariate confidence ellipse of X. chrysen-teron, X. cisalpinus and X. pruinatus are shown inFigs 2–3.
Two-sample t tests of mean values were carried out(Table 2). Each variable of taxon A is compared tovariable of taxon B. Mean values of variables are sig-nificantly different at P<0.005, and can therefore beconsidered as useful morphological character for thedistinction of these two taxa. X. cisalpinus can easilybe separated from both X. chrysenteron and X. pruina-tus by the shorter, thinner spores and the longer endcells of the pileipellis (particularly with respect toX. pruinatus). End cells width is not useful to separate
70
60
50
40
30
200 10 20 30
End cells width (µm)
End
cel
ls le
ngth
(µm
)
X. pruinatus
X. cisalpinus
X. chrysenteron
Fig. 2. Pileipellis end cells size distribution (length and width)
of Xerocomus chrysenteron, X. cisalpinus and X. pruinatuswith Gauss’ confidence ellipse (P=0.683).
16
15
14
13
12
114.0
Spore width (µm)
Spo
re le
ngth
(µm
)
X. pruinatus
X. cisalpinus
X. chrysenteron
4.5 5.0 5.5 6.0
Fig. 3. Spore size distribution (length and width) of Xero-comus chrysenteron, X. cisalpinus and X. pruinatus with
Gauss’ confidence ellipse (P=0.683).
Xerocomus cisalpinus sp. nov. and the X. chrysenteron complex 662

X. cisalpinus from the two other taxa. X. chrysenteroncan be separated from X. pruinatus by end cell lengthand end cell width, but not by spore length or sporewidth. All three species can clearly be separated fromeach other by their end cell length.
The discriminant analysis also demonstrated thatend cell length is a significant variable: especially incombination with spore width, end cell length enablessignificant separations. Combinations of other mor-phometric characters gave poorly significant separ-ations.
The canonical variables are evaluated at the groupmeans. In the canonical variable plot (Fig. 4) the cen-troid for X. chrysenteron is at x=0.157, y=0.582, the
centroid for X. cisalpinus is at x=x3.341, y=x0.337,and the centroid of X. pruinatus is at x=2.230,y=x0.409. Equality of group means was tested withthe F statistics values. They are proportional to dis-tance measures of the centroids shown in the canonicalscores plot. The closer a case is to a particular group’slocation, the higher the probability that it belongs tothat group. Based on morphometric values, X. cisalpi-nus, X. chrysenteron and X. pruinatus can easily differ-entiated from each other: there is no overlap betweenX. cisalpinus and the others, and only a small overlapbetween X. chrysenteron and X. pruinatus. X. chry-senteron and X. pruinatus are the two most similar taxa(16.665).X. cisalpinus is more similar toX. chrysenteron(34.240) than to X. pruinatus (77.627).
In the classification matrix each case is classified intothe group where the value of its classification functionappears to be the highest. For X. chrysenteron, 23 casesare classified correctly (82%) and 5 cases are mis-classified (as X. pruinatus). For X. cisalpinus, 17 casesare classified correctly (94%) and 1 case is misclassified(as X. chrysenteron). For X. pruinatus, 24 cases areclassified correctly (96%) and 1 case is misclassified asX. chrysenteron.
The first canonical discriminant function (Table 3)is the linear combination of the variables that bestdiscriminate among groups, the second canonical dis-criminant function is orthogonal to the first and nextbest combination of variables. The first discriminant
Table 2. Morphometric variables of Xerocomus cisalpinus, X. pruinatus and X. chrysenteron are tested for their usefulness to distinguish
two taxa: two-sample t tests of mean values (MW) of the morphometric variables spore length (SPOLE), spore width (SPOWI), pileipellis
end cells length (ENDLE) and end cells width (ENDWI) were carried out. Each variable of taxon A is compared to variable of taxon B.
They are significantly different at P<0.005, and can therefore be considered as a useful morphological character for the distinction of
these two taxa.
Variable MW of taxon A MW of taxon B P-values
ENDLE X. cisalp. (51¡7.6 mm) X. pruin. (24.6¡2.6 mm) 0.000
ENDLE X. cisalp. (51¡7.6 mm) X. chrys. (37.7¡6.6 mm) 0.000
ENDLE X. chrys. (37.7¡6.6 mm) X. pruin. (24.6¡2.6 mm) 0.000
ENDWI X. cisalp. (13.2¡2.3 mm) X. pruin. (11.1¡3.3 mm) 0.024
ENDWI X. cisalp. (13.2¡2.3 mm) X. chrys. (13.8¡2.9 mm) 2.196
ENDWI X. chrys. (13.8¡2.9 mm) X. pruin. (11.1¡3.3 mm) 0.003
SPOLE X. cisalp. (12.9¡0.6 mm) X. pruin. (14.1¡0.5 mm) 0.000
SPOLE X. cisalp. (12.9¡0.6 mm) X. chrys. (13.9¡0.7 mm) 0.000
SPOLE X. chrys. (13.9¡0.7 mm) X. pruin. (14.1¡0.5 mm) 0.190
SPOWI X. cisalp. (4.7¡0.1 mm) X. pruin. (5.2¡0.2 mm) 0.000
SPOWI X. cisalp. (4.7¡0.1 mm) X. chrys. (5.1¡0.3 mm) 0.000
SPOWI X. chrys. (5.1¡0.3 mm) X. pruin. (5.2¡0.2 mm) 0.118
Table 3. Canonical discriminant functions (CDF) standardised by
within variances, eigenvalues and cumulative proportion of total
dispersion (Cum. prop. tot. disp.).
CDF 1 CDF 2
SPOLE 0.276 0.313
SPOWI 0.378 0.457
ENDLE x0.982 0.277
ENDWI 0.264 0.542
Eigenvalues 4.793 0.231
Cum. prop. tot. disp. 0.954 1.000
FACTOR 1
FAC
TO
R 2
X. pruinatus
X. cisalpinusX. chrysenteron
420–2–4–6–6
–4
–2
0
2
4
Fig. 4. Canonical score plot for Xerocomus cisalpinus, X.chrysenteron and X. pruinatus. The axes of this plot are thefirst two canonical discriminant functions, and the points are
the canonical variables scores. Confidence ellipses are centredaround the centroid of each group. X. cisalpinus, X. chry-senteron and X. pruinatus are strongly differentiated from
each other: there is no overlap between X. cisalpinus and theothers, and a small overlap only between X. chrysenteron andX. pruinatus.
U. Peintner, H. Ladurner and G. Simonini 663

function depends strongly on end cells length, and inthe second place on spore width. The first Eigenvalue(4.793) is high when compared to the second Eigen-value (0.231). This indicates that the first canonicaldiscriminant function captures most of the differencesamong the groups (cumulative proportion of total dis-persion is 95.4% with the first canonical discriminantfunction only). Thus, end cells length and spore widthare the most useful biometric features useful to separatethe three taxa in question.
Multivariate test statistics were applied to test thenull hypothesis of identity of the three groups X. chry-senteron, X. cisalpinus and X. pruinatus. The null hy-pothesis was clearly rejected for all three taxa (Table 4).
Phylogeny of the Xerocomus chrysenteron speciescomplex
Phylogenetic analyses were performed with PAUP*4.0b8 (Swofford 1998). 34 rDNA-LSU sequences ofnine Xerocomus spp. treated in this study were alignedmanually in the data editor of PAUP*. After exclusionof gaps and areas with ambiguous alignment, 975characters were analysed: 730 characters were constant,51 variable characters were parsimony-uninformativeand 94 characters were parsimony-informative.
Heuristic searches resulted in 20 000 most parsi-monious trees (MPT) with tree length of 234 steps, aconsistency index (CI) of 0.739, a retention index (RI)of 0.845 and a rescaled consistency index (RC) of 0.625.
The phylogenetic analysis with maximum likelihoodresulted in 11 ML trees (xln likelihood=2602.04007),one of them is presented in Fig. 5. The X. chrysenteronspecies complex can clearly be separated from the out-group X. subtomentosus (L. : Fr.) Quel. (BS 100%).Within the X. chrysenteron complex, X. rubellus se-quences fall into a distinct clade, which is sister tothe other seven included taxa X. chrysenteron, X. poro-sporus, X. cisalpinus, X. dryophilus, X. pruinatus,X. ripariellus andX. fennicus. Five of these seven species,namely X. porosporus, X. cisalpinus, X. dryophilus andthe species pair X. ripariellus/X. fennicus representindependent lineages with high bootstrap support (BS>70%). A clade of X. chrysenteron species is indicatedby tree topologies, but with low bootstrap support (BS53%). Basal relationships within the seven core speciesof the X. chrysenteron complex are not resolved, buttree topologies indicate X. chrysenteron as a basaltaxon, and an intermediate position of X. porosporus.X. ripariellus and X. fennicus are sister groups. In
addition, the other three lineages of X. cisalpinus,X. dryophilus and X. pruinatus are closely related andhave sister group relationships. The analysed sequencesof X. ripariellus include material of X. catalaunicus(IB19980360). The results from our phylogeneticanalysis confirm the synonymy of these two speciesepithets, as already shown in an earlier study based onmorphological characters (Ladurner et al. 2001).
The phylogenetic analyses confirm our species con-cepts, as all sequences belonging to one morphospeciesfell into the same clade. However, four sequences re-trieved from GenBank (X. chrysenteron AF050547,AF071537, X. pruinatus AF402140 and X. rubellusAF050649) clustered in different clades, demonstratingthe present taxonomic confusion around these taxa.The collections used for these four sequences are mis-identified. This further emphasises the urgent need ofbroadly accepted morphological species concepts.
TAXONOMY
Xerocomus cisalpinus Simonini, Ladurner & Peintner,sp. nov. (Figs 6, 14–17)
Etym. : Occurring south of the Alps.
Habitus X. chrysenteronis similis ; statura minuta; pileus35–80 mm diam, convexus, pallide ochraceogriseus, olivaceo
vel roseo colore suffusus, vulgo obscurior in maturitate, citominutis rimis ubique notatus; subpellis roseorubra saepe intercuticulae rimas conspicitur. Pori et tubuli flavi. Stipes 45–
80r4–9 mm, sursum flavus, ad basim ruber. Caro pallideflava, exalbescens, lente sed valde ad caeruleum coloremvarians ad stipitis basim. Sporae leviter striatae ut in X. prui-
nato, mediae, (10.5–) 13¡1 (16)r(4–) 4.5¡0.2 (–5.5) mm;Q=(2.18) 2.77¡0.18 (3.40); cuticulae partes plerumquecylindricae, supermembranam incrustatae,mediae, (13) 5¡17(123)r(4.5–) 13¡4 (–28.5) mm;Q=(1.51) 4.06¡1.50 (14.00).
Hyphae amyloideae ad stipitis basim ut in X. pruinato.Typus : Italia : Reggio Emilia : prope Villaminozzo in loco
dicto Resgadore, alt. 1100 m supra mare, in radura Fagus
sylvaticae nemore, 25 Sept. 1998, Franca Franceschetti(IB19980850 – holotypus).
Pileus 35–80 mm broad, fleshy, convex to pulvinateto almost plane, sometimes also slightly depressed atthe centre ; surface dry, tomentose, independently fromthe weather conditions always very soon areolate(cracking into minute scabs), the pink-reddish subpellisusually visible in the fissures, but exceptionally (oldbasidiomes, dry weather conditions) also only thewhitish flesh visible in the fissures ; very variable incolour : in very young basidiomes with uncrackedpileus cream-greyish, pale ochre-brown with an oliveshade (1/2D2/3, 3B/C2, 4/5B/C2/3, 5/6D2 to 5/6D4),often with a pale pink line at the extreme margin(6/7A2/3), sometimes with a dispersed rosey shade(7D3 to 6, 8E3/4, 8 to 11D2 to 4) or in wet weatherconditions even entirely fuchsia-pink, these tingesmore intense at the pileus margin (9/10A to C5 to 7);when cracked, the pileipellis scabs darker greyish-brown-olive (6/7E 4 to 6), flesh-coloured to rosey in the
Table 4. Multivariate test statistics. F is the ratio of variation
‘between’ the three groups Xerocomus chrysenteron, X. cisalpinus,
X. pruinatus and ‘within’ the three groups. D.F=degree of freedom.
Multivariate test F D.F P-tail
Wilks’ lambda=0.140 27.143 8,130 0.0000
Pillai’s trace=1.015 17.002 8,132 0.0000
Lawley-Hotelling trace=5.024 40.191 8,128 0.0000
Xerocomus cisalpinus sp. nov. and the X. chrysenteron complex 664

cracks (6/7A2 to 4, 8A2 to 5, 7 to 9A3 to 5, S199, 200);dull cream-whitish (3/4A2) cracks appear beneath thebrownish (6/7F5 to 7) pileipellis layer in dry weather.The pileipellis is often so minutely cracked that theintense pink colour of the subpellis beneath dominatesand the pileus colour seems pinkish red.
Tubes up to 5–10 mm long, separable, depressed tosinuate around the stipe, sometimes slightly decurrent ;initially bright yellow with olive shades (1/2A7/8, 3A/B7/8), later dull greenish yellow (2C/D7/8, 3/4C/D6),slowly bluing when bruised. Pores angular, uneven, inmature basidiomes 0.7–2 mm diam, concolorous to thetubes, slowly and only weakly bluing (S351, 352, 366,367) when bruised.
Stipe 45–80 mm long, 4–9 mm broad, cylindrical,sometimes with somewhat widened apex, usuallytapering towards the base but also weakly clavate orbulbose; vivid yellow at the apex (3A4 to 8), vivid redor dull red in the lower half (8–10A to C8, 8–11C/D7/8,S121, 136, 151, 166), more rarely red at the extremebase only or entirely yellow with some vague reddishstriae only; in older, overmature specimens the reddishtinges turn into a dull red (10/11E/F7/8, S101, 102) ; thestipe surface becomes dull ochre-brown (S336 to S338)when injured; sometimes the stipe appears orange dueto red dots or red striae in the yellow parts of the stipe;the stipe surface finely floccose or fibrillose, withoutreticulum; at the extreme base pale ochre-brown (S336to 338); basal mycelium white.
Context soft in the pileus, fibrillose and brittle inthe stipe; in the pileus pale yellow (3A3) soon fadingto dirty whitish when exposed (29/30A2), below thepileipellis showing a fine red line, particularly dis-tinct when the pileus shows no red shade, otherwise
inconspicuous; in the stipe vivid yellow (3A4/5) oryellow with dull reddish tinges that are concolorous tothe cortex (8 to 11C/D7/8), slowly but strongly bluing(S446 to 448) in the whole stipe or only in its lower partwhen cut, the red areas in the stipe base darkening, thestipe context at the extreme base dull ochre-brownish(S336 to 339); in the area of the stipe/cap connectionvivid yellow and only slightly turning green (2A/B7/8)or more or less pale aquamarine (S331 to 335) whencut; smell not distinctive, somewhat like wet iron; tastenot distinctive, slightly sour.
Spore deposit brownish with some olive tinges.Macrochemical reactions weak fleeting-amyloid re-
action in stipe context and hymenophoral trama of dryspecimen.
Basidiospores (10.5–)13¡1(16)r(4–)4.5¡0.5(–5.5) mm, Q=(2.2) 2.8¡0.2 (3.4), V=(92) 149¡37(465) mm3 (n=558), subfusiform, slender, with a weaklydeveloped but distinct supra-apicular depression, sporewall up to 0.5 mm thick, intensely honey-coloured andwith one to two guttules when mature, the spore sur-face finely longitudinally striate. Basidia 30–45r10–13 mm (n=15), inconspicuous, clavate, hyalineto yellowish, mainly 4-spored. Pleurocystidia scarce,50–90r7–15 mm (n=15), ventricose-fusiform, withhyaline to slightly yellowish content, cheilocystidiasimilar, scattered. Pileipellis a palisadoderm stronglyreminiscent of the pileipellis of X. chrysenteron, ter-minal elements mostly cylindrical and somewhattapering towards the apex, often papillate, also acorn-shaped to exceptionally subspherical, (13–)51¡17(–123)r(4.5–)13¡4(–28.5) mm, Q=(1.5) 4.1¡1.5(–14) in Congo red, n=(588), terminal and subter-minal elements moderately to heavily incrusted, only
Key to the species of the Xerocomus chrysenteron complex
1 Spores truncate . . . . . . . . . . . . . . . . . . 2
Spores not truncate . . . . . . . . . . . . . . . . . . 3
2(1) Spores smooth, basidiomes with dull colours, greyish brown, red tinges only weakly developed, often with a reddish ring zone at the stipe
apex, thermophil species associated with Quercus, Carpinus, Fagus, etc. . . . . . . . X. porosporus
Spores distinctly striate, young basidiomes vivid red, fading to olivaceous grey when older, with Betula, Alnus, etc., preferring nitrogen-
rich habitats . . . . . . . . . . . . . . . . . . X. fennicus
3(1) Spore quotient on average <2.5, mainly associated with deciduous trees . . . . . . . . . 4
Spore quotient on average >2.5, with both deciduous trees and conifers . . . . . . . . . 5
4(3) Spores on average less than 5.5 mm broad, honey-coloured; context in the base of the stipe with small red dots formed by a striking flame-
red pigment, or a thin red line delimiting the extreme base of the stipe; often associated with Tilia (but also with Quercus and other
deciduous trees and shrubs), preferring ‘disturbed’ sites, e.g. gardens, borders of trails, etc. . . . . . X. rubellus
Spores on average broader than 5.5 mm, dark brown to intense yellow-brown; stipe context intensely red coloured in the lower part
(reminding Boletus luridus), mainly under Quercus (and Pinus?) . . . . . . . . X. dryophilus
5(3) Spore surface smooth; usually associated with conifers or with Fagus in southern Europe, pileipellis a palisadoderm of moderately long,
cylindrical, parallel running hyphae, the terminal cells¡shortly cylindrical to broadly ovoid, acorn-shaped to cystidioid, exceptionally
subspherical . . . . . . . . . . . . . . . . . X. chrysenteron
Spore surface striate (striation difficult to see with light microscopy); pileipellis a palisadoderm of extremely variably shaped elements, the
terminal elements slender cylindrical to ¡broadly clavate to pear shaped, ovoid to spherical . . . . . . 6
6(5) Typical ‘pruinatus-hyphae’ lacking, associated with Alnus, Populus, Salix, etc., in humid, nitrogen rich places, context whitish to yellow,
spores on average less than 5 mm broad . . . . . . . . . . . . X. ripariellus
With ‘pruinatus-hyphae’ (thick-walled, amyloid hyphae in stipe context) . . . . . . . . . 7
7(6) Spores on average broader than 5 mm, associated withCastanea, Fagus, Picea, Pinus, etc., similar toX. chrysenteron, context bright yellow
and only weakly bluing when cut. Cap cuticle usually not areolate, few radial fissures start from the pileus margin. X. pruinatus
Spores on average less than 5 mm broad, associated withQuercus, Pinus, etc., similar to X. chrysenteron, context pale yellow, whitish when
cut, strongly bluing at least in the lower stipe. Cap cuticle always and rapidly areolate. . . . . . X. cisalpinus
U. Peintner, H. Ladurner and G. Simonini 665

exceptionally smooth. Stipitipellis consisting of looselyinterwoven, 3–5 mm broad hyphae with hyaline toyellowish content. A well-developed caulohymeniumwith spore-forming caulobasidia and caulocystidia ispresent in the upper half of the stipe. Stipe textureformed of parallel running, hyaline, 2–4 mm broadhyphae; in the lower half of the stipe the hyphae are
differentiated into a special hyphal type (‘pruinatus-hyphae’) in irregular intervals : inconspicuous, thin-walled and moderately broad hyphae extend more orless abruptly to strikingly thick-walled, up to 30 mmbroad, irregularly formed hyphae of variable length,which, depending on their individual developmentalstage, show a weak to strong amyloid reaction of the
X. chrysenteron IB19990952
X. chrysenteron AF347103
X. chrysenteron IB20000405
X. chrysenteron IB19990951
80
9399
93X. porosporus AF050645
X. porosporus IB19880304
X. porosporus IB19990957
73
78
X. cisalpinus IB20000701
X. cisalpinus IB20000700
X. cisalpinus IB19900700
X. cisalpinus IB19980850
X. chrysenteron AF071537
X. dryophilus IB19990901
X. dryophilus IB19991057
X. chrysenteron AF050647
X. pruinatus IB19920266
X. pruinatus IB19961055
X. pruinatus IB19950970
X. pruinatus IB19980366
X. pruinatus AF050644
X. ripariellus IB19980360
X. ripariellus GR21189
X. rubellus AF050649
X. ripariellus GR930920
X. ripariellus GR22465
X. fennicus H126
X. fennicus RJ126
100
98
6697
100
57
69
93
78 77
86
X. pruinatus AF402140
X. rubellus GS1044
X. rubellus GS961
X. rubellus IB1999091763100
100
100
100 X. subtomentosus IB19980452
X. subtomentosus IB19991000
X. subtomentosus AF139716
–0.005 substitutions/site
X. rubellus
X. ripariellus
X. fennicus
X. pruinatus
X. dryophilus
X. cisalpinus
X. chrysenteron
X. porosporus53
Fig. 5. One of 11 ML trees with xln likelihood of 2602.04007. MP bootstrap values are given above branches, ML bootstrapvalues >60% are given below branches. Branches not present in the ML strict consensus tree are grey. Branches present in
both strict consensus trees (MP and ML) are bold. The spores of a taxon are shown right of the respective clade. Xerocomussubtomentosus was used as outgroup.
Xerocomus cisalpinus sp. nov. and the X. chrysenteron complex 666

hyphal wall. The inner surface of these hyphae oftenshows a particular crater-like ornamented surface. Noclamp connections observed.
Microchemical reactions : with the exception of theamyloid reacting hyphae in the stipe, no particularreproducible macrochemical reactions were observed.
Habitat and distribution : Xerocomus cisalpinus wasrepeatedly collected in Italy, south of the Alps, inMediterranean areas: in the regions Lazio, EmiliaRomagna, Sardinia, Calabria; Trentino Alto Adige(South Tyrol) ; it fruits in the autumn and late autumnfrom sea level to 1150 m, usually scattered, in groups.X. cisalpinus is associated with broad-leaved trees,mainly Quercus spp. (Q. cerris, Q. robur, Q. pubescens,Q. frainetto, Q. suber, Q. ilex), but also with Fagus syl-vatica ; the occurrence under Pinus has to be confirmed.
Like many other mushroom species, Xerocomuscisalpinus is difficult to recognise in the field based onmacromorphological features alone. Because of itsobvious cracked cap cuticle, X. cisalpinus is somewhatsimilar to X. chrysenteron. However, X. cisalpinus caneasily be distinguished from both X. chrysenteron andX. pruinatus based on microscopic features : it has apileipellis structure consisting of substantial cylindricalhyphae similar to X. chrysenteron, but the faintly striatespores and amyloid ‘pruinatus- hyphae’ are typical forX. pruinatus. In addition, the spores of X. cisalpinus aresignificantly smaller than those of both X. chrysenteronand X. pruinatus.
No similar taxa could be found in an extensiveliterature screening of publications dealing withBoletus, Boletellus or Xerocomus. Only Boletellus epis-copalis could have some affinities with our collections :This bolete from Madagascar is described as small-sized, with a slender red stipe slightly inflated at thebase, the cap cuticle is dark brown, cracking in bigscabs and the spores are described as faintly ridged(Heim & Perreau 1963). We examined the typus ofBoletellus episcopalis (Fig. 18), which is conserved inethanol in PC(RH128). B. episcopalis is undoubtedlydifferent from X. cisalpinus : the spores of B. episcopalishave a pronouncedly striate surface, which is easilyvisible with a light microscope; the spores are (12–)13.5¡1 (15)r(5.5–)6¡0.5(–6.5) mm; Q=(2) 2.3¡0.1(2.6) ; V=(199) 252¡27 (322) mm3 (n=31), and thusclearly wider than spores of X. cisalpinus. ‘Pruinatus-hyphae’ are lacking. The pileipellis is in bad con-dition, but it appears more like a trichodermium thanlike a palisadoderm; the weakly incrusted pileipellisend cells are (26.5–)82¡39(–154.5)r(5.5–)10¡2.5(–15.5) mm; Q=(2) 8.7¡4.9 (22.1) (n=31) and muchmore elongated and usually tapering towards the tip(Fig. 18).
Additional collections examined : Italy : Sardegna : Nuoro,
Urzulei, Codula di Luna, alt. 50 m, with Quercus ilex, 2 Nov.1990, G. Simonini (IB19900700). Calabria : Cosenza, Acri,Cugnale di Falcone, alt. 1000 m, with Q. cerris, Pinus
calabrica, 8 Sept. 1995, C. Lavorato (IB19950800); ReggioCalabria, Caulonia, Monte Gremi, alt. 1050 m, with Q. ilex
and P. nigra, 29 Oct. 1995, G. Simonini (IB19950801); EmiliaRomagna, Reggio Emilia, Villaminozzo, La Magolese, alt.
1150 m, with Fagus sylvatica, 31 Aug. 1997, G. Simonini(IB19970995). Sardegna : Sassari, S. Teresa di Gallura, RenaMaiore, alt. 5 m, with P. pinea, Quercus sp., 29 Oct. 2000,G. Consiglio (IB20000702); Sassari, Tempio Pausania,
Aggius, alt. 500 m, with Q. suber, 3 Nov. 2000, G. Redeuilh(IB20000704, GS2310a). Lazio : Latina, Sabaudia, Selva delCirceo, alt. 17 m, with Q. suber, 29 Nov. 2001, A. Riva
(GS2405); loc. cit., with Quercus spp., 30 Nov. 2001, M.Manavella (GS 2400, GS2403); loc. cit., P. Signorello(GS2404); loc. cit., M. Manavella (GS2406); loc. cit., G. Si-
monini (GS2407, IB20010605) ; loc. cit., 1 Dec. 2001, G. Si-monini (GS2401); loc. cit., M. Manavella (GS2402). TrentinoAlto Adige : Bolzano, Naturns, Ladurnhof, alt. 800 m, with
Quercus pubescens, 16 May 1999, H. Ladurner (IB19991019).
Xerocomus chrysenteron (Bull.) Quel. 1888(Figs 7, 19–20)
Basionym: Boletus chrysenteron Bull., Hist. Champ. :328 (1791).
Pileus 30–120 (–200) mm broad, convex to pulvinateto almost hemispherical when young, more convex toapplanate when old, sometimes slightly depressed atthe centre, initially blackish-brown to chestnut-brown,exceptionally more beige or clay-coloured, often withan olive tinge, also honey-brown, towards the marginmore yellowish, in older basidiomes often fading fromthe centre to a pale clay-colour or greyish brown; thepileus margin often more olive-yellow or even reddishcoloured and then clearly contrasting to the rest of thepileus in older basidiomes (in dry periods already inyoung specimens). Pileus surface dry, young tomen-tose, becoming ¡velutinous-tomentose, never viscid,often cracking deeply into coarse clods, typically areo-late with the reddish subpellis visible in the fissures.
Tubes up to 11 mm long, slightly depressed, sinuatearound the stipe, in overmature basidiomes sometimeslike decurrent, initially pale yellow, then dull greenish-yellow, soon more yellowish-green. Pores large,0.7–1.5 mm diam, angular, uneven, concolorous to ormore olive-yellow, sometimes with rust-red spots,weakly to distinctly bluing when bruised.
Stipe 30–90r5–25 mm, cylindrical, solid, somewhattapering towards the base or also slightly widened.Surface dry, smooth to ingrown fibrillose, yellow at theapex, towards the base or completely or partially redcoloured by fine reddish scales or fibrils.
Context whitish-yellow to yellow in the pileus, belowthe pileipellis showing a fine, but distinct red line, moreintensely yellow in the stipe, below red coloured partsof the stipitipellis a pink to reddish coloration of thecontext might be observed. Context in the whole basi-diome ¡bluing when bruised. Basal mycelium whitishto pale yellow. Smell and taste inconspicuous to slightlysour. Colour of dried specimen inconspicuous.
Spore deposit brownish.Basidiospores (9–)14¡1(–17)r(4–)5¡0.5(–6.5) mm,
Q=(2.0) 2.7¡0.2 (3.5), V=(97) 194¡42 (349) mm3 in
U. Peintner, H. Ladurner and G. Simonini 667

Xerocomus cisalpinus sp. nov. and the X. chrysenteron complex 668

KOH 3% (n=930), subfusiform, with well-pronouncedsupra-apicular depression, moderately thick-walled(wall up to 0.5 mm thick), intensely honey-colouredand with one to two guttules when mature, smooth,inamyloid, not dextrinoid. Spores from fresh basi-diomes sometimes staining intensely dark greyish-bluewhen tested for amyloidity, but spores of the samebasidiomes react completely negative in dry condition.Basidia 30–40(–50)r10–15 mm (n=15), clavate, hya-line to yellowish in KOH 3%; mainly 4-spored.Pleurocystidia numerous, 50–100r10–18 mm (n=15),usually ventricose-fusiform with slender elongated neckand rounded tip in KOH 3%. Cheilocystidia similar,
numerous. Pileipellis a palisadoderm of cylindrical,mostly parallel, often heavily incrusted hyphae. Ter-minal elements versiform, cylindrical, cystidioid, bul-let-shaped, acorn-shaped, exceptionally subspherical,sometimes with elongated flexuous neck, irregular,(7.5–)37¡13.5(–104.5)r(5–)13.5¡4(–57.5) mm, Q=(0.8) 2.9¡1.0 (9.7) in Congo red (n=930), subterminalelements as broad as the terminal element, exception-ally also distinctly widened. Terminal elements smoothto heavily incrusted, the pigment often forming pro-nounced yellowish-brown scabs (KOH 3%). Stipiti-pellis a texture of loosely interwoven, richly ramified,mostly hyaline, 3–7 mm broad hyphae. A well-devel-oped caulohymenium of basidia, basidioles and caulo-cystidia covers the entire stipe; in the upper third ofthe stipe a continuous layer of caulohymenium is ob-served, towards the base it turns into ¡isolated bun-dles of caulohymenial elements. Stipe texture consist-ing of thin-walled, hyaline, 6–15 mm broad hyphae(KOH 3%). No clamp connections observed. Neithertrue amyloid, dextrinoid, nor other microchemicalreactions were observed in any part of the basidiomes.
Habitat and distribution : X. chrysenteron is wide-spread and common all over Europe, and usuallyoccurs under conifers. This taxon seems to prefer cli-matically moderate zones and undisturbed habitats.X. chrysenteron is only exceptionally found under de-ciduous trees, e.g. Fagus, but then preferably in colder,mountainous habitats in southern Europe. The mainfruiting period is July–September.
Collections examined. Austria : Tyrol : Innsbruck, Hunger-
burg, with Picea abies, 8 Aug. 1999, H. Ladurner(IB19990951); Afling, with Picea abies and Corylus avellana,13 Aug. 1999, R. Kuhnert-Finkernagel (IB19950800); Tulfes,
Fig. 14. Basidiomes of Xerocomus cisalpinus (holotype).
Bar=1 cm.
Fig. 15. Basidia, basidiospores, two cheilocystidia (left)and two pleurocystidia (right) of Xerocomus cisalpinus
(IB19980850 – holotype). Bar=10 mm.
Fig. 16. Pileipellis end cells and basidiospores of Xerocomuscisalpinus (IB19980850 – holotype). Bars=20 mm for pilei-
pellis, and 10 mm for basidiospores.
Figs 6–13. Basidiomes of the eight species of the X. chrysenteron species complex in their natural habitat. Fig. 6. Xerocomuscisalpinus (IB20010605). Fig. 7. X. chrysenteron (IB19970995). Fig. 8. X. dryophilus (GS2004). Fig. 9. X. fennicus (JV7150F);photograph by Jukka Vauras. Fig. 10. X. porosporus (GS2106). Fig. 11. X. pruinatus (GS2017). Fig. 12. X. ripariellus
(GR22541P – holotype p.p.) ; photograph by Guy Redeuilh. Fig. 13. X. rubellus (GS1894). Photographs where not otherwisestated by G.S.
U. Peintner, H. Ladurner and G. Simonini 669

Hasental, with Pinus, Picea, 28 July 2000, H. Ladurner(IB20000405). – Italy : Bozen : Partschins, Greithof, with
Picea, H. Ladurner (IB19970898); for additional materialsee Ladurner (2001a). – Switzerland : Zurich : with Picea abiesand Fagus sylvatica, 20 Aug. 1985, C. Lavorato (GS1541).
Xerocomus dryophilus (Thiers) Singer 1986 (Figs 8, 21)
Basionym: Boletus dryophilus Thiers, Calif. Mush. : 82(1975).
Pileus 40–100 mm broad, young convex to pulvinate,with age often more flattened, then often also with anirregularly bent margin; young brownish-red to red,sometimes the red cap covered by an olive-brownishtomentum, with maturity at least in certain cases darkred, in other cases the red tinges fade and the brown-olive colour predominates. Young basidiomes oftenwith a rosy hue caused by the pale tomentum; some-times the still involute margin appears entirely rose-red; pileus surface dry to slightly viscid, in youngbasidiomes strongly tomentose, in older ones oftennearly glabrous, occasionally wrinkled or cracked intofine to coarse scabs.
Tubes to 10 mm long, weakly depressed to narrowlyadnexed around the stipe, in old basidiomes some-times appearing decurrent, olive-yellow,¡bluing wheninjured. Pores large, 0.5–1 mm diam, angular, uneven,concolorous with the tubes, bluing when bruised.
Stipe 50–80r10–20 mm, solid, cylindrical or api-cally slightly enlarged. Stipe surface dry to somewhatviscid, smooth. Stipe apex yellow, towards the basereddish, the red tinges more pronounced in older basi-diomes. The red coloured parts of the stipe darkeningon pressure.
Context in the stipe concolorous with its surface, in-tensely red in the lower half of the stipe (reminiscent ofB. luridus) and turning – with the exception of the stipebase – everywhere blue when cut. Context in the pileusyellow, weakly reddish under the pileipellis, onlyweakly bluing. Basal mycelium yellowish-white. Tasteand smell not significant. Colour of exsiccata not sig-nificant.
Spore deposit brown.Basidiospores (11–)13¡1(–17.5)r(5–)6¡0.5
(–7) mm, Q=(1.7) 2.2¡0.2 (3), V=(151) 252¡46(414) mm3 in KOH 3% (n=465), elliptical, with amostly inconspicuous supra-apicular depression, mod-erately to distinctly thick-walled (wall up to 0.9 mmthick), mature intensely honey-coloured to brownwith one to two guttules, smooth, not amyloid, notdextrinoid. Basidia 35–45(–50)r11–14 mm (n=15),clavate, hyaline to yellowish in KOH 3%; mainly4-spored. Pleurocystidia numerous, 60–85r10–15 mm(n=15), somewhat irregular fusoid-ventricose withrounded apex, thin-walled, hyaline to yellowish inKOH 3%. Cheilocystidia similar, numerous. Pileipellisa trichoderm consisting of long, slender, partially inter-woven, occasionally also branched, septate hyphae.Terminal elements cylindrical, slender, elongated, alsoovoid or cystidiod, sometimes branched at the apex ordiverticulate, usually tapering, but also with roundedends, sometimes anastomosing, (8.5–)54.5¡25(–194.5)r(5–)9¡3.5(–14) mm, Q=(1.3) 6.2¡3.4 (23.1) inCongo red (n=465), often smooth, otherwise finelyincrusted, sometimes also showing a granular yellowish
Fig. 17. Scanning electron microscopy (SEM) pictures of
basidiospores of Xerocomus cisalpinus (IB19980850 – holo-type) with finely longitudinally striate spore surface.Bar=2 mm.
Fig. 18. Pileipellis end cells and basidiospores of Boletellusepiscopalis (holotype). Bar=20 mm for pileipellis, and 10 mmfor basidiospores.
Xerocomus cisalpinus sp. nov. and the X. chrysenteron complex 670

19 20
21 22
23 24
25 26
27 28
Figs 19–28. Pileipellis and basidiospores of species of the Xerocomus chrysenteron group. Fig. 19. X. chrysenteron (GS1541).Fig. 20. X. chrysenteron (IB19970898). Fig. 21. X. dryophilus (GS1941). Fig. 22. X. fennicus (Typus). Fig. 23. X. porosporus(IB1988304). Fig. 24. X. ripariellus (GR21189). Fig. 25. X. pruinatus (IB19991024). Fig. 26. X. pruinatus (IB19950970).
Fig. 27. X. rubellus (GS961). Fig. 28. X. rubellus (IB19990917). Bar=20 mm for pileipellis, and 10 mm for basidiospores.
U. Peintner, H. Ladurner and G. Simonini 671

content in KOH 3%. Hyphae of the lower pileipellismoderately to distinctly incrusted, subterminal el-ements only exceptionally widened. Stipitipellis a loosetexture of slender, loosely interwoven, abundantlyramified, hyaline hyphae of 3–5 mm diam with slightlygranulose content. Bundles of clavate, hyaline elementsoccur over the entire length of the stipe. Stipe texturereminding X. subtomentosus, consisting of thin- tothick-walled (wall to 1.5 mm thick) 6–10 mm broad hy-phae, the hyphal content in KOH 3% partly finely-granulose. Without clamp connections. No particularmicrochemical reactions observed in any parts of thebasidiomes.
Habitat and distribution : X. dryophilus probably oc-curs in most thermophilic mixed deciduous forests witha preference for Quercus spp. It is known from North-and Central America and from Europe (Mediterraneanarea), where it fruits solitarily, to gregarious in de-ciduous forests. In Europe X. dryophilus was foundunder Q. ilex, Q. pubescens, Q. robur and Fagus silva-tica. On Mallorca the species was collected also in thedune zone under Pinus halepensis. The identity ofAmerican and European material was confirmed byHarry Thiers based on morphological characters(Simonini 1994).
Collections examined. Croatia : Loscinji, airport, with Pis-tacia lentiscus, Erica arborea, Quercus ilex, Juniperus oxyce-
drus, 6 Oct. 1999, Anonymous (IB19990901). – Spain : Alcoleadel Pinar, 5 Sept. 1999, Anonymous (IB19991057); for ad-ditional material see Ladurner (2001a). – Italy : Reggio Emi-
lia : near Villaminozzo, Calizzo, with Q. pubescens, 21 Sept.1999, G. Simonini (GS1941); Emilia Romagna, Quat-trocastella, Parco di Roncolo, alt. 320 m, with Q. pubescens, 3Oct. 2000, G. Simonini (GS2004). –Mexico : Baja California,
with Quercus agrifolia, 13 Feb.1993, A. Montecchi, det.G. Simonini (GS 0933); Canon de las Animas, with Q. agri-folia, 28 Jan. 1998, G. Moreno (IB19980827); for additional
material see Simonini (1994).
Xerocomus fennicus (Harmaja) Ladurner & Simonini,comb. nov. (Figs 9, 22)
Basionym: Boletellus fennicus Harmaja, Karstenia39(2) : 37 (1999).
Pileus 10–70 mm broad, convex to plane, initiallybright red, when older the colour changes to brownfrom the centre, but some red tinges usually remainpersistent, particularly at the pileus margin. Pileussurface dry, ¡finely tomentose, often finely crackingfrom the margin towards the centre and showing theyellowish context in the fissures.
Tubes 2–8 mm long, adnate to depressed around thestipe, bright yellow, intensely bluing when injured.Pores usually rather large, roundish to angular, con-colorous with the tubes, intensely bluing when bruised.
Stipe 20–70r4–15 mm, solid, cylindrical, concolor-ous with the pileus, the apex usually paler or yellowish,often ¡brown in old basidiomes, finely floccose orfibrillose.
Context yellowish white, strongly bluing. Taste andsmell not significant. Basal mycelium pale, whitish.Colour of dried specimen not significant.
Spore deposit brown.Macrochemical reactions exsiccata with a weak
‘fleeting-amyloid’ reaction in stipe context and hy-menophoral trama.
Spores (10–)12.5¡1(–15.5)r(4–)5¡0.5(–6.5) mm,Q=(2) 2.6¡0.2 (3.4), V=(85) 152¡27 (278) mm3 inKOH 3% (n=2635), some slender and nearly cylin-drical, others elliptic to broadly subfusiform, rathervariable in shape, size and differentiation of the sporeapex, mature spores usually with a distinctly truncateapex, but, in some specimens only few truncate sporesare observed. Depending on their shape, the sporesshow a distinct supra-apicular depression; they areusually moderately thick-walled (wall to 0.6 mm thick),mainly honey-coloured and mono- to biguttulate whenmature, distinctly longitudinally striate (striation evenstronger developed than in X. ripariellus), inamyloid,not dextrinoid. Basidia 30–45r9–12 (n=15), clavate,hyaline to yellowish in KOH 3%, mainly 4-spored.Pleurocystidia scarce, 35–80r10–18 mm (n=15), ven-tricose-fusiform, often tapering at the apex or withelongated neck, most hyaline, rarely with brownishcontent in KOH 3%. Cheilocystidia similar, scattered.Pileipellis a physalo-palisadoderm consisting of mod-erately long, cylindrical, septate hyphae. Terminal el-ements versiform, rather variable in shape and size, theshape of the terminal elements ranging from cylindricalwith rounded or tapering apex to bullet-shaped,broadly pear-shaped, ovoid, elliptical, subspherical oralmost spherical, sometimes with elongated outgrows,(10–)32¡11(–77)r(4–)15.5¡5(–43) mm, Q=(0.8)2.2¡0.8 (7.8) in Congo red (n=685), the penultimateelement only exceptionally widened. Pileipellis elementssmooth to heavily incrusted, in some collections withlarge, refractive plaques (KOH 3%) not reacting withCongo red. Stipitipellis consisting of slender, looselyinterwoven, mainly hyaline, 3–5 mm broad hyphae withslightly granulose content. In the upper half of the stipea closed layer of well-developed caulohymenium con-sists of bundles of caulocystidia and spore-formingcaulobasidia. Stipe texture in most collections in-conspicuous, formed of parallel running, ¡hyaline,3–10 mm broad hyphae. Without clamp connections. Inseven out of 94 collections of X. fennicus amyloid‘pruinatus-hyphae’ were found in the stipe context ;no other particular microchemical reactions wereobserved.
Habitat and distribution : Up to now Xerocomuxfennicus is known only from Finland (hemiboreal andsouthern boreal zone) and Austria (one collection nearSalzburg), possibly also from Belgium (Schreiner 2000).The fungus occurs in rich deciduous or mixed forests,also in alder thickets on lakeshores and seashores,pastures, parks, yards, and meadows, favouringnitrogen-rich soils, (nearly) always associated withBetula, often accompanied by Alnus sp. and Urtica.
Xerocomus cisalpinus sp. nov. and the X. chrysenteron complex 672

Collections examined : Finland : Varsinais-Suomi : Nauvocommune, Berghamn, Boskar, with Betula spp., Alnus gluti-
nosa, Picea abies, 3 Sep. 1992, J. Vauras JV7150F. Etela-Hame (Tavastia australis) : Ylojarvi, Soppeenmaki, park,beneath ash (Fraxinus) trees, five fruit bodies, 5 Aug. 1993,R. Jarvenpaa (OULUF46399).Uusimaa (Nylandia) : Helsinki,
Myllypuro-Puotinharju, herb-rich mixed forest (Betula,Picea, Populus tremula, Prunus padus, Salix caprea, Sorbus,Aegopodium, Filipendula, Rubus idaeus, Urtica dioica), on
detritus on soft humus, 19 Aug. 1988, R. Saarenoksa (H). ;Lammi, Pappila, biological Station of the University ofHelsinki, in park under Betula, 9 Sept. 1968, H. Harmaja
(H – holotype of Boletallus fennicus).
Xerocomus porosporus Imler, Bull. Soc. mycol. Fr. 74 :97 (1958). (Figs 10, 23)
Pileus 20–60(–139) mmbroad, young pulvinate, slightlyconvex, then applanate, sometimes irregularly bent,olive-ochre, olive-brown, sometimes nearly blackish-brown when young, paler when old, also with greyishtinges, dull. Pileus margin slightly paler when young,often more greyish. Pileus surface dry, ¡finely tomen-tose, often finely to distinctly areolate and showing thepale yellowish to whitish context in the fissures.
Tubes 13–20 mm long, adnate to adnexed to thestipe, initially pale yellow, later yellowish-green to dullolive-green, bluing when injured. Pores rather narrow,0.2–0.5 mm in diameter, roundish to angular, con-colorous to the tubes or somewhat rust-red in olderbasidiomes, bluing when bruised.
Stipe 30–80(–110)r4–20 mm, solid, cylindrical,sometimes with widened apex, usually tapering towardsthe base, concolorous with the pileus, in old basidiomeseven darker, only at the apex with a narrow yellow(-ish)zone, downwards dull greyish-brown, olive-brown orblackish brown, often with a narrow reddish zone inthe upper part or with inconspicuous reddish patchesin the lower part, finely floccose or fibrillose (in olderbasidiomes).
Context whitish to cream-coloured in the pileus, inthe stipe also pale yellow, there sometimes with reddishtinges, brownish to dark brown in the stipe base, bluingthroughout the whole basidiomes. Taste and smell in-conspicuous. Basal mycelium whitish to greyish-white.Colour of exsiccata inconspicuous.
Spore deposit brownish.Macrochemical reactions not observed.Basidiospores (11–)14¡1(–16.5)r(5–)6¡0.5
(–6.5) mm, Q=(1.8) 2.5¡0.3 (3.1), V=(144) 241¡40(355) mm3 in KOH 3% (n=217), elliptical to subfusi-form with a distinctly truncate apex in at least 80% ofmature spores, the spore apex thick-walled at the twoapical angles and often markedly thin-walled betweenthese angles, with no distinct germ pore but apicallyonly moderately thick-walled (wall up to 0.7 mm thick),with distinct supra-apicular depression, intenselyhoney-coloured to yellow-brown, and with one totwo guttules when mature, smooth, inamyloid, notdextrinoid. Basidia 30–40r9–13 (n=15), with 3–5 mm
long sterigmata inconspicuous, clavate, hyaline toyellowish in KOH 3% mainly 4-spored. Pleurocystidiascarce, 30–90r9–16 mm (n=15), slender, mainlyventricose-fusiform, hyaline, rarely with brownish con-tent in KOH 3%. Cheilocystidia similar. Pileipellis aphysalo-palisadoderm consisting of cylindrical, mostlyparallel running hyphae. Terminal elements versiform,cylindrical, cystidioid, acorn-shaped, bullet-shaped, ex-ceptionally subspherical, irregular, (8–)33¡11.5(–68)r(4.5–)7.5¡2(–15) mm, Q=(0.8) 2.6¡1.2 (7.6) inCongo red (n=217). Pileipellis elements smooth toheavily incrusted (KOH 3%). Stipitipellis consisting ofslender, loosely interwoven, mainly hyaline, 3–5 mmbroad hyphae with slightly granulose content. In theupper half of stipe a closed layer of well-developedcaulohymenium consists of partly spore-bearing caulo-basidia (basidioles) and caulocystidia . Stipe textureinconspicuous, formed of parallel, ¡hyaline, 2–4 mmbroad hyphae. Without clamp connections. No par-ticular microchemical reactions observed. Neither trueamyloid nor dextrinoid reactions observed in any partsof the basidiomes.
Habitat and distribution : Xerocomus porosporus iswidespread but rather rare in Europe with a preferencefor climatically favoured regions. It is found from Mayto November under deciduous trees (Fagus, Carpinusbetulus, Quercus cerris, Q. petraea, Q. robur, Q. suber,Betula, Crataegus, Rubus) and in mixed forests, excep-tionally also under Pinus and Picea.
Collections examined : Spain : Montseny, park of the Castel
Montseny, with Fraxinus, Cedrus, Sorbus aucuparia, 18 Oct.1988, R. Poder (IB1988304). – Italy : South Tyrol : Naturns,with Quercus pubescens, Juniperus communis and Fraxinus
ornus, 28 Aug. 1999, H. Ladurner (IB1999957); EmiliaRomagna, Villaminozzo, Pian Vallese, alt. 1350 m, withFagus sylvatica, 18 Jul. 1999, G. Simonini (GS2106). For
additional material see Ladurner (2001a).
Xerocomus pruinatus (Fr.) Quel. 1888(Figs 1, 11, 25–26)
Basionym: Boletus pruinatus Fr., in Fries & Hok,Boleti : 9 (1835).
Pileus 30–100 (–150) mm broad, young convex, pul-vinate, later applanate to flattened or depressed, fleshy,initially usually blackish brown, deep purple-brown,deep brown, later more dark brown to purple, excep-tionally purple to blood-red. In old basidiomes thebrown colours may change into dull olive tinges. Pileusmargin lighter coloured, whitish, yellowish, apricot torusty. Pileus surface dry, when young alveolate-rugoseand fugaceous pruinose, later mat and sometimesalmost smooth. Pileipellis rarely cracking and if so,then the cracking usually begins at the pileus margin asfine fissures ; subsequently the whole pileipellis may alsobreak into coarse clods.
Tubes long, even longer than the diam. of thepileus context, reaching more than 10 mm, younglemon-yellow, bright yellow, chrome-yellow, then
U. Peintner, H. Ladurner and G. Simonini 673

greenish-yellow, adnexed with a tooth, usually weaklybluing when bruised. Pores concolorous with the tubes,small to medium sized, only exceptionally wider than1 mm, roundish to isodiametrically angular, ageingwithout green tinges, turning to rust-brown when in-jured.
Stipe 30–80(–120)r10–30(–40) mm, cylindrical toventricose-fusiform, solid, sturdy, weakly fibrillose,young bright yellow, minutely floccose, the initiallyyellow scales turning red in older basidiomes or aftercollecting, so that the stipe, with exception of theyellow apex, appears ¡entirely reddish. The stipe baseoften turns brownish when bruised.
Context firm, ¡purely yellow, in older specimensometimes more ochraceous, exceptionally almostwhite, then often also reddish tinges are observed in thestipe context ; inconstantly weakly bluing. Smell andtaste banal, somewhat acidulous.
Fresh spore deposit mustard-yellow, dry with olivetinges.
Macrochemical reactions the context of freshmaterial turns olive-green with Melzer.
Basidiospores (9–)14¡1(–17)r(4–)5¡0.5(–6.5) mm,Q=(2.0) 2.7¡0.2 (3.6), V=(68) 195¡38 (340) mm3
(n=806) in KOH 3%, subfusiform, with well-pro-nounced supra-apicular depression, with slightlythickened walls (wall to 0.5 mm thick), intensely honeycoloured and with one to two guttules when mature,spore surface finely longitudinally striate, inamyloid,not dextrinoid. Basidia 30–45r9.5–15 (n=15), clavate,with hyaline to yellowish content in KOH 3%, mainly4-spored. Pleurocystidia scattered, 50–95r10–16 mm,ventricose-fusiform, with hyaline to yellowish contentin KOH 3% (n=15). Cheilocystidia scarce, similar topleurocystidia. Pileipellis a palisadoderm formed ofrather variable, versiform elements, the shape of theterminal elements ranging from small cylindrical tospherical. The slender and short cylindrical terminalelements often show an apical widening, reminiscentof a drumstick (this apical widening of the pileipelliselements is typical of X. pruinatus), the subterminal el-ement usually wider than the terminal one; sometimesa slender, cylindrical terminal element is followed by aspherical subterminal element of multiple diameterdownwards or even chains of one to three spherical el-ements are observed, their diameter increases down-wards. The size of the terminal pileipellis elements isalso highly variability : (8–)25.5¡9(–67)r(3.5–)11¡5(–47.5) mm, Q=(0.5–)2.5¡1 (7) mm in Congo red(n=806). Depending on the developmental stage of thebasidiomes and the collection, two principal pileipellistypes can roughly be distinguished: pileipellis type1: terminal elements rather slender, ¡cylindrical todrumstick-shaped, smooth to weakly incrusted, thesmooth ones usually with an intracellular brown pig-ment, the subterminal cell often widened and as shortor even shorter than the terminal element and normallywithout intracellular pigmentation, the whole pileipelliswith weakly to moderately developed incrustations.
Pileipellis type 2 terminal elements broad, often bullet-shaped to subspherical, only exceptionally slender andcylindrical, in the latter case often followed by aroundish penultimate element, often with chains of twoto three roundish elements, but the terminal elementalso in this case narrower than the subterminal cell,intracellular pigment only in the appendage-like nar-row terminal cells, the subspherical and bullet-shapedelements normally moderately to heavily incrusted, thewhole pileipellis in type 2 with much stronger devel-oped incrustations than in type 1. Transitional stagesbetween these two extremes are frequently found. Sti-pitipellis consisting of slender, loosely interwoven,mainly hyaline, 3–5 mm broad hyphae; well-developedcaulohymenium on the entire length of the stipe with anearly closed layer of caulobasidia and caulocystidia.Stipe texture formed of parallel running, ¡hyaline,2–4 mm broad hyphae, which in the lower half ofthe stipe differentiate in irregular intervals into the‘pruinatus-hyphae’ : inconspicuous, thin-walled andmoderately broad hyphae extend more or less abruptlyto strikingly thick-walled, to 30 mm broad, irregularlyformed hyphae of variable length, which, dependingon their individual developmental stage, show a weakto strong amyloid reaction. The inner wall of thesehyphae often shows a particular, uneven surface ofcrater-like elements (Fig. 1). In all textures thrombo-plere hyphae occur in thrombomorph or meromorphstate. Apart from the amyloid reacting hyphae in thestipe context no particular microchemical reactionswere observed. In a few collections, spores reacted witha slight greyish hue when mounted in Melzer’s and thebasidia reacted dextrinoid. No metachromatic reactionwas observed in spores or hyphae.
Habitat and distribution : Xerocomus pruinatus iswidespread in Europe, it was collected in Austria, theformer Czechoslovakia, Denmark, England, France,Germany, Hungary, Italy, Poland, Sweden, Spain, andSwitzerland; it is probably often misinterpreted as X.chrysenteron. The main fruiting period of X. pruinatusis autumn (September to November), being only ex-ceptionally collected during summer. Basidiomes fruitunder conifers (Abies, Picea, Pinus) as well as underdeciduous trees (Acer, Carpinus, Castanea, Fagus,Fraxinus, Quercus, etc.), in open grassy sides, on trailborders or in forests. No preferences for particular soiltypes have been observed.
Collections examined : Austria : Carintia : Knappenberg,with Fagus and Picea, 8 Oct. 1995, M. Kirchmair & R. Poder(IB19950970). Karnten : Knappenberg, with Picea betweenmosses, 8 Oct. 1998, R. Poder (IB19980366). – Italy : Parma :
Stabielle, with Quercus and Castanea, 2 Oct. 1996, R. Poder &H. Ladurner : (IB19961955). Liguria : Savona, Bardineto, alt.750 m, with Fagus sylvatica and Castanea sativa, 10 Oct. 1998,
G. Simonini (GS2017). – Ukraine : Kiev, Angarskii, Prereval,road from Alutscha to Simferopol, 6 Oct. 1992,M. M. Moser(IB19920066). – Spain : Cantabria : malataja, Quercus pyre-
naica, 24 Oct. 1999, A. Munoz & R. Luis (IB19991024). Foradditional material see Ladurner (2001a).
Xerocomus cisalpinus sp. nov. and the X. chrysenteron complex 674

Xerocomus ripariellus Redeuilh, Docums. Mycol.26(104) : 30 (1997) (Figs 12, 24)
Boletellus catalaunicus Poder, G. Moreno, Rocabruna& Tabares, Mycotaxon 62 : 232–234 (1997).
Pileus 30–70 mm broad, young convex, then applanate,initially ¡red, blood-red, cherry-red, sometimes vina-ceous, but also more brownish-red, fading to clay,greyish-brown or dust-coloured from the centre. Pileusmargin brighter, whitish, yellowish, long time involute.Pileus surface uneven, gibbose, ¡velvety, pruinose.
Tubes young nearly white, then lemon yellow, turn-ing greenish yellow, ¡bluing when bruised. Poresconcolorous, irregular, noticeable wide when old,¡bluing when bruised.
Stipe 30–60(–90)r10–20(–25) mm, ¡smooth, sub-cylindrical to slightly fusiform, slightly tapering at thebase, usually rather robust, the apex concolorous withthe pores, downwards ¡reddish floccose as in X.chrysenteron, sometimes longitudinally striate, in olderbasidiomes with haematoma-like spots of variousnumber, the stipe base turning dull brown on pressure.
Context initially almost whitish to light lemon-yellow, sometimes also bright yellow, more brownishbeneath the pileipellis and the stipitipellis, ¡intenselyochraceous in the stipe base, in the lower half of thestipe often vinaceous to violet, elsewhere whitish toyellow, often strongly bluing in the middle part of thestipe. Taste and smell banal. Basal mycelium whitish toyellowish. Colour of dried specimens not significant.
Spore deposit brownish with some olive tinges.Macrochemical reactions all collections but one (IB
19940617) showed a distinct ‘fleeting-amyloid’ reactionin stipe context and hymenophoral trama; this excep-tion might be explained by the bad preservation of thiscollection. No other macrochemical reactions observed.
Basidiospores (10.5–)13.5¡1(–17)r(4–)4.5¡0.5(–5.5) mm, Q=(2.2) 2.9¡0.2 (3.6), V=(85) 152¡25(228) mm3 in KOH 3% (n=713), slender, subfusiform,with weakly developed but distinct supra-apicular de-pression, with slightly thickened walls (wall to 0.5 mmthick), intensely honey-coloured and with one to twoguttules when mature, the surface finely longitudinallystriate (striation more pronounced than in X. pruina-tus), inamyloid and not dextrinoid. Basidia 30–45r9.5–14 mm (n=15), clavate, with hyaline to yellowish tointensely yellow content in KOH 3%; mainly 4-spored.Pleurocystidia scarce, 50–90r8–16 mm (n=15), ventri-cose-fusiform, with hyaline to yellowish content inKOH 3%. Cheilocystidia scattered, similar to thepleurocystidia. Pileipellis a physalo-palisadoderm ofextremely variably shaped elements, usually a palisadeof versiform elements is observed but in some basi-diomes the pileipellis appears nearly epitheloid, theterminal elements are slender cylindrical to broadlyclavate to pear shaped, ovoid, spherical, with or with-out elongated terminal outgrow. Single spherical el-ements or chains of two to three widened, ¡spherical
terminal elements usually turn abruptly into slendercylindrical hyphae. In some basidiomes ‘brushes’ ofwidened terminal elements sit on a common, reversepear-shaped subterminal cell, forming ‘floriform’ struc-tures ; apart from the shape the dimensions of the ter-minal pileipellis elements also vary strongly within asingle specimen as well as between different basidiomesor different collections : (7–)29.5¡10(–63.5)r(4.5–)16.5¡5.5(–46) mm, Q=(0.6) 2¡0.9 (7.4) mm in Congored (n=1023), terminal and subterminal elements finelyto heavily incrusted; in some basidiomes the terminalelements are nearly smooth and hyaline in KOH 3%, inother basidiomes (often of the same collection) heavyincrustations peel off the terminal and subterminalelements forming coarse scabs in KOH 3%. Sphericaland subspherical terminal elements of larger diameterusually show much heavier incrustations than slendercylindrical ones. In some basidiomes large, refractive,not congophile plaques are observed (KOH 3%).Stipitipellis consisting of loosely interwoven, 3–5 mmbroad hyphae with hyaline to yellowish content, thehyphal ends mainly rounded, sometimes slightlywidened. In the upper half of stipe a well-developedcaulohymenium is observed. Stipe context formed ofparallel, densely packed, 4–8 mm broad, thin-walledhyphae with hyaline to yellowish, sometimes granularcontent in KOH 3%. No clamp connections observed.No particular microchemical reactions observed.
Habitat and distribution : Xerocomus ripariellus ismostly found in humid habitats, at lake shores or atriver banks, in dunes, but also in (humid) gardens,parks, etc., associated with deciduous trees (especiallywith Populus, Alnus and Salix).
Collections examined : France : Yvelines, 78, Etang d’Or,beside the water, alt. 50 m, with Salix, 29 Sept. 1995,Redeuilh, (GR22541P – holotype p. p.) ; Paris, Foret d’Orient,
20 Sept. 1993, Mahieu (GR93092); Paris, Nouvelle station,17 Sept. 1995, Mahieu (GR22465) ; sine loc., 5 Oct. 1994,Anonymous (GR21189). – Spain : Catalonia : Riells de Mon-
tseny, among litter on sandy soil in mixed hardwoods, 16Sept. 1998, A. Rocabruna (IB19980360, as Boletellus catalau-nicus). For additional material see Ladurner (2001a).
Xerocomus rubellus Quel., Ass. Fr. Avanc. Sci. 5 : 620(1895). (Figs 13, 27–28)
Pileus 40–100(–150) mm broad, young nearly hemi-spherical, then applanate, also slightly depressed atthe centre, sometimes irregularly bent, of extremelyvariable colours, blood-red, cherry-red, orange-red,rose-red, pink, ochraceous-pink to ochraceous-red,incarnadine, ochraceous, yellowish-brown, brown,grey-brown, olive-grey, grey, dark brown, black-brown,typically fading with age. Pileus margin somewhatpaler, initially involute, then plane or also revolute,acute. Pileus surface dry, ¡finely tomentose, oftencracking especially at the centre, showing the yellow oralso red context in the fissures and then resemblingX. chrysenteron.
U. Peintner, H. Ladurner and G. Simonini 675

Tubes to 12 mm long, adnate to slightly decurrent,initially lemon-yellow, later greenish-yellow, slightlybluing when injured. Pores large, wider than 1 mm,roundish to angular, concolorous with the tubes, inmature basidiomes with orange-rusty tinges, onlyweakly bluing when bruised.
Stipe 25–70(–120)r4–35 mm, solid, versiform, oftenslender, cylindrical, also stout, bulbose to ventricose-subfusiform, always tapering towards the base andslightly rooting, ornamented with darker brownish orreddish scales or fibrils on ochraceous ground, colourof the stipe surface similar to the pileus colour.
Context soft, pale yellow, in base of the stipe with atypical flame-red pigment in form of small dots or a thinred line. Context only weakly bluing and exceptionallyalso reddening in the cap, the reddish discolorationfading on drying. Taste and smell inconspicuous. Basalmycelium whitish-yellow. Colour of dried specimensnot significant.
Spore deposit brown with olive tinges.Macrochemical reactions weak ‘fleeting-amyloid’
reaction in stipe context and exceptionally also in thehymenophoral trama of exsiccata, but most collectionsshow a strong dextrinoid reaction in all textures ; nofurther macrochemical reactions observed.
Basidiospores (9–)12¡1(–15)r(3.5–)5¡0.5(–6.5) mm, Q=(1.8–)2.3¡0.2(–3.3), V=(76) 169¡35(–355) mm3 in KOH 3% (n=1767), broadly elliptical tobroadly fusiform, with weak to distinct supra-apiculardepression, honey-coloured and with one to two gut-tules when mature, smooth, inamyloid and notdextrinoid. Basidia 30–45r9–13 mm (n=15), incon-spicuous, clavate, hyaline to yellowish in KOH 3%mainly 4-spored. Pleurocystidia scattered, 40–60r8–13 mm (n=15), slender, ventricose-fusiform, oftenwith elongated neck, most hyaline, rarely with yellow-ish content in KOH 3%. Cheilocystidia similar, numer-ous, often with yellowish content in KOH 3%. Pilei-pellis a physalo-palisadoderm consisting of moderatelylong, cylindrical, septate hyphae. Terminal cells versi-form, acorn-shaped, bullet-shaped, subspherical,broadly cylindrical with rounded apex to cystidioid,sometimes also with elongated neck, but also slender,cylindrical, slightly tapering towards the tip (7.5–)32¡11(–93.5)r(5.5–)12¡4(–41) mm, Q=(0.8) 2.8¡1.0 (9.4) in Congo red (n=1798); subterminal elementssometimes slightly widened. Pileipellis elements smoothto distinctly incrusted by a fine granular, yellowishpigment, exceptionally also heavily incrusted elementsare observed (KOH 3%). Stipitipellis consisting ofslender, loosely interwoven, mainly hyaline, 3–5 mmbroad hyphae. In the upper third of the stipe bundles ofcaulohymenium consist of clavate, hyaline to yellowcoloured basidia, basidioles and broadly ventricose-fusiform cystidia with an often extremely elongatedneck (to 30 mm long). Stipe texture not significant,formed of parallel running, ¡hyaline, 2–4 mm broadhyphae; scattered to scarce flame-red crystals (hyphalexcretions) dissolving in KOH 3% are found in the
lower stipe context. Without clamp connections. Noparticular microchemical reactions observed in anypart of the basidiomes.
Habitat and distribution : Xerocomus rubellus is wide-spread in Europe. Brown forms are more frequentlyfound in central, northern and western Europe than thetypical red forms. X. rubellus fruits between July andNovember under deciduous trees and shrubs (e.g.Quercus spp., Tilia sp., Corylus avellana), often inurban areas, on lawn, on disturbed road sides, also inopen grass land, but only exceptionally in open, un-disturbed woods. This species is reported from Austria,Croatia, England, Finland, France, Germany, theNetherlands, Spain, but occurs probably in all Euro-pean countries with moderate to warm climate.
Collections examined : Croatia : Veli Loscinji, with Quercusilex and Olea europea, 8 Oct. 1999, H. Ladurner
(IB19990917). – Italy : Reggio Emilia : Viale Ramezzini, withQuercus cerris and Tilia, 5 Sept. 1993, G. Simonini (GS961);Grosseto, Orbetello, ‘Le Piane’, with Quercus suber, 30 Sept.
1993, G. Simonini (GS1044); Emilia Romagna, Viano, Pul-piano, alt. 520 m, with Quercus cerris, 7 Jun. 1998, G. Simo-nini (GS1894). For additional material see Ladurner (2001a).
DISCUSSION
Nomenclatural notes
Some modern authors (e.g. Hansen & Knudsen 1992)have rejected the name Xerocomus chrysenteron due toconsiderable nomenclatural confusion and ambiguousinterpretations. Bulliard’s (1791) original description ofBoletus chrysenteron seems to mix characters of bothX. chrysenteron and the ‘brown aspect ’ of X. rubellus(i.e. X. communis (Bull.) Bon 1985). Moreover, Bulliardhimself synonymised B. chrysenteron with B. cupreusSchaeff. 1774. However, we regard the name X. chry-senteron as a name in current use, which should beconserved (Redeuilh & Simonini 1994).
The old name Boletus subtomentosus var. pascuusPers. 1825 (syn. Xerocomus pascuus (Pers.) Gilb. 1931)is a nomen dubium, which should not be further applied.The brief macroscopical description given by Persoon(1825) (‘pileo subtomentoso, poris flesuosis crassis,carne immutabili, stipite brevi squamuloso toto cocci-neo’) fits with many Xerocomus species, making it im-possible to attribute Persoon’s description to any taxonas it is currently circumscribed. Krombholz’s (1831–46)description and pictures of B. pascuus seem to agreewell with the modern concept of X. chrysenteron (darkbrown cap, yellow stem, purple red minutely squamu-lose at the base), but the cap cuticle was not reported ascracking. Corda’s (1837–54) description and picture ofB. pascuus (dark brown, velvety, cracked at margin cap,purple red stem basis internally and outwardly) agreesperfectly with the modern concept of X. chrysenteron.Also the habitat on pastures supports the interpret-ation of B. pascuus as X. chrysenteron.
Xerocomus cisalpinus sp. nov. and the X. chrysenteron complex 676

Klofac & Krisai-Greilhuber (1992) and Lannoy &Estades (2001) used the name X. pruinatus var. pascuus(Pers.) Klofac & Krisai-Greilhuber for a taxon witha smooth cap (not wrinkle-folded as is typical forX. pruinatus), and orange colours on cap and stipe.We interpret X. pruinatus var. pascuus in the sense ofKlofac & Krisai-Greilhuber (1992) as a synonym ofX. pruinatus : the range of X. pruinatus end cells alsocircumscribes taxa attributed to the nomen dubiumX. pruinatus var. pascuus (Ladurner 2001a, b).
In addition, the name X. rubellus is embedded in aconsiderable amount of nomenclatural confusion. Thelack of type material and the varying interpretations(Krombholz 1836, Quelet 1895, Singer 1945, Snell &Dick 1958, Binyamini & Avizohar-Hershenzon 1973,Klofac & Krisai-Greilhuber 1992, Simonini 1998a)make this taxon in the original sense uninterpretable.However, we consider this species epithet as a namein current use and so we will continue to apply it totaxa as delimited above. Epiypification of X. rubellusmight contribute to a badly needed unification of theX. rubellus species concept.
Phylogeny of the species complex
Our morphology-based species concepts were con-firmed by the results of the molecular phylogeneticanalyses based on the same material. Thus, sequencedata of the rDNA-LSU are useful for the delimitationof groups in the Boletales, as already shown in earlierstudies (Fischer et al. 1997, Binder 1999, Jarosch & Besl2001, Binder & Bresinsky 2002). Basal relationships arenot fully resolved, but several studies using rDNA se-quences have shown that basal branches are often dif-ficult to resolve based on LSU phylogenies only (Binder1999,Moncalvo et al. 2000a, b, 2002,Wagner & Fischer2001, Thomas et al. 2002).
Our phylogenetic analyses prove that the X. chry-senteron group is a distinct evolutionary lineage, whichcan clearly be separated from other boletes by sequencedata and by morphological characters. Binder (1999)proposed the provisional generic name ‘Paraxero-comus ’ for this species complex based on a fewsequences as well as morphological and chemotaxo-nomical characters. However, as long as relationshipsof Boletales at the genus level are not satisfactorilyunderstood, especially between Boletus, Xerocomus,Boletellus and Pseudoboletus, we do not recommendnomenclatural changes at generic level.
One striking result is the isolated, basal position ofX. rubellus within the X. chrysenteron complex. Likemost other species of this group, X. rubellus is a macro-scopically very variable species. Brown forms areeasily confused with X. chrysenteron (from which theydiffer by the lower spore quotient) and red forms areoften confused with X. dryophilus (which differs bybroader spores). Red forms of X. pruinatus, X. ripar-iellus and X. fennicus differ by their striate spores, andthe lack of a flame red pigment in the base of the stipe.
The well-supported, basal position of X. rubellusindicates that smooth spores are a synapomorphiccharacter: this implies that both, spore striation andspores with a truncate apex (sometimes remindinga germ pore), are secondarily derived within theX. chrysenteron complex.
Although basal relationships are not fully resolved,tree topologies indicate that taxa with striate spores donot form a monophyletic group. Thus, the sectionStriatulisporae does probably not represent natural re-lationships. The slight spore striation, which is oftenhard to discern with a light microscope, appears to be aderived character. It can be speculated, that spore stri-ation evolved independently in several lineages of thisspecies complex. In X. fennicus the spore striation ismost obvious. Therefore, this species was originallyplaced in Boletellus. Our phylogenetic analyses dem-onstrate that X. fennicus belongs to the X. chrysenteronspecies complex, where it is closely related to X. ripar-iellus. Therefore we proposed a recombination of thisspecies epithet into Xerocomus.
Truncate spores as found in X. fennicus and X. poro-sporus are secondarily derived characters with twoindependent origins. Also ‘pruinatus-hyphae’ havemultiple origins as they are found in X. pruinatus,X. cisalpinus and sporadically also in X. fennicus.
All species within the X. chrysenteron complex canclearly be distinguished based on LSU sequences.However, sequences retrieved from GenBank often donot match with the respective clade. Our analyses showthat ‘X. rubellus ’ AF050649 is a misidentified X. ripar-iellus, and that ‘X. chrysenteron ’ AF050647 mightbe a misinterpreted X. pruinatus. The American ‘X.chrysenteron ’ AF071537 could be conspecific withX. cisalpinus indicating that this new taxon might alsooccur in California. ‘X. pruinatus ’ AF402140 could notbe assigned to any clade, but it could be within themorphological and molecular range of X. chrysenteron.
In summary, sequence data are a valuable and prom-ising tool for Boletales systematics. But the utilisationof material that is macro- and micro-morphologicallywell characterised is an indispensable prerequisite forreliable results in molecular systematics.
Delimiting micromorphological characters
Micromorphological characters as well as the ecologyare often neglected in species concepts of boletes. Thedetermination and delimitation of taxa was, up tonowadays, based mainly on macroscopical characters,which are often very variable and therefore misleading.All attempts in molecular systematics to address ques-tions on species level are doomed to fail, when thematerial used is not reliably identified. Bolete speciesconcepts must be placed on a solid base and includeconcrete, micromorphological characters such as sporesize and shape and characters of the pileipellis struc-ture. As we have shown, features of the spores and ofthe pileipellis are particularly reliable characters for
U. Peintner, H. Ladurner and G. Simonini 677

species differentiation in the Xerocomus chrysenteroncomplex. But spore dimensions are often very variable,as far as minima and maxima are concerned. There-fore, a statistically significant number of spores mustbe measured for a reliable identification. Truncatespores and spore striation are good delimiting charac-ters, but spore striation is often difficult to detect witha light microscope. The degree of striation alsodepends on the maturity of the spores: young sporesare more or less smooth, while mature spores are faintlystriate. The ornamentation is faintest in X. pruinatusand X. cisalpinus, and increasingly pronounced inX. ripariellus and X. fennicus.
A reliable identification of species in the X. chry-senteron complex is also possible based on the structureof the pileipellis and the dimensions of its end cells. Inparticular, the length of the pileipellis end cells is agood delimiting character, although infraspecific vari-ability is always present to a certain degree. ‘Pruinatus-hyphae’ are currently known only from three species,and therefore are a good character for their identifi-cation. Thus, an accurate identification of specieswithin the X. chrysenteron group can easily be achievedwith the combination of the three morphologicalcharacters spores, pileipellis and ‘pruinatus-hyphae’.In such way reliably identified and characterisedmaterial is the key to the often puzzling phylogeny ofboletes. The combination of classical and molecularmethods can then form a solid base for a more naturalsystem of mushrooms that is urgently needed.
ACKNOWLEDGEMENTS
We are grateful to our friends Daniele and Massimo Antonini,
Antonella Bartocchi, Marco Contu, Roberto Fontenla, Franca Fran-
ceschetti, Carmine Lavorato, Mauro Manavella, Eugenio, Giulio e
Lanfranco Mariotti, Guy Redeuilh, Alfredo Riva, Giancarlo Sassi,
Pietro Signorello for having supported our study with a large amount
of collections from all over the Mediterranean area. We thank Guy
Redeuilh and Jukka Vauras for permission to publish their photo-
graphs of Xerocomus ripariellus and X. fennicus, respectively. Special
thanks go to Giovanni Consiglio for the Latin diagnosis, to Esteri
Ohenoja for loans of material and translation of Finnish specimen
labels, and to Bart Buyck for loaning us the type of Boletellus
episcopalis.
REFERENCES
Binder, M. (1999) Zur molekularen Systematik der Boletales: Boleti-
nae und Sclerodermatinae subordo nov. Dissertation, University of
Regensburg.
Binder, M. & Fischer, M. (1998) [‘1997’] Molekularbiologische
Charakterisierung der Gattungen Boletellus und Xerocomus:
Xerocomus pruinatus und verwandte Arten. Bollettino del Gruppo
Micologico Bresadola 40 : 79–90.
Binder, M. & Bresinsky, A. (2002) Derivation of a polymorphic lin-
eage of Gasteromycetes from boletoid ancestors. Mycologia 94 :
85–98.
Binder, M. & Hibbett, D. S. (2001) Evolutionary switches between
brown rotting saprophytes and the ectomycorrhizal symbionts in
the Boletales. Inoculum 52 : 21.
Binder, M. & Hibbett, D. S. (2002) Higher-level phylogenetic re-
lationships of homobasidiomycetes (mushroom-forming fungi)
inferred from four rDNA regions. Molecular Phylogenetics and
Evolution 22 : 76–90.
Binyamini, N. & Avizohar-Hershenzon, Z. (1973) Boletaceae of
Israel. II. Gyroporus, Xerocomus and Boletus. Transactions of the
Bristish Mycological Society 60 : 99–103.
Bresinsky, A. & Schwarzer, G. (1969) Mikroskopische Analyse der
Hutdeckschichten einiger Agaricales, Boletales und Russulales.
Zeitschrift fur Pilzkunde 35 : 263–293.
Bulliard, J. B. F. (1791)Histoire des Champiguous de la France. Paris.
Corda, A. C. J. (1837–54) Icones Fungorum hucusque cognitorum.
6 vols. J. G. Calve, Prague.
Drehmel, D., Moncalvo, J. M. & Vilgalys, R. (1999) Molecular
phylogeny of Amanita based on large-subunit ribosomal DNA
sequences: implications for taxonomy and character evolution.
Mycologia 91 : 610–618.
Engel, H., Dermek, A., Klofac, W. & Ludwig, E. (1996) Schmier- und
Filzrohrlinge s.l. in Europa. Verlag Heinz Engel, Weidhausen.
Felsenstein, J. (1985) Confidence limits on phylogenies: An approach
using the bootstrap. Evolution 39 : 783–791.
Fischer, M., Jarosch, M., Binder, M. & Besl, H. (1997) On the sys-
tematics of the Boletales: Suillus and related genera. Zeitschrift
Fuer Mykologie 63 : 173–188.
Grubisha, L. C., Trappe, J. M., Molina, R. & Spatafora, J. W. (2001)
Biology of the ectomycorrhizal genus Rhizopogon. V. Phylogentic
relationships in the Boletales: inferred from LSU rDNA sequences.
Mycologia 93 : 82–89.
Hansen, L. & Knudsen, H. (1992) Nordic Macromycetes. Vol. 2.
Nordsvamp, Copenhagen.
Heim, R. & Perreau, J. (1963) Le genre Boletellus a Madagascar et en
Nouvelle-Caledonie. Revue de Mycologie 28 : 191–193.
Heinemann, P., Rammeloo, J. A. & Rullier, E. (1988) L’ornementa-
tion sporale des Xerocomaceae spores dites lisses. Bulletin du Jardin
Botanique Nationale de Belgique 58 : 513–534.
Humpert, A. J., Muench, E. L., Giachini, A. J., Castellano, M. A.
& Spatafora, J. W. (2001) Molecular phylogenetics of Ramaria
and related genera: evidence from nuclear large subunit and
mitochondrial small subunit rDNA sequences. Mycologia 93 :
465–477.
Jarosch, M. & Besl, H. (2001) Leucogyrophana, a polyphyletic
genus of the order Boletales (Basidiomycetes). Plant Biology 3 :
443–448.
Klofac, W. & Krisai-Greilhuber, I. (1992) Xerocomus chrysenteron
und ahnlich aussehende Rohrlinge. Osterreichische Zeitschrift fur
Pilzkunde 1 : 19–59.
Kornerup, A. & Wanscher, J. H. (1978) Methuen Handbook of
Colour. 3rd edn. Methuen, London.
Krombholz, J. V. (1831–46) Naturgetreue Abbildungen und Beschrei-
bungen der essbaren, schadlichen und verdachtigen Schwamme.
10 vols. J. G. Calve, Prague.
Ladurner, H. (2001a) The Xerocomoideae of Europe. PhD thesis,
University Innsbruck.
Ladurner, H. (2001b) La variabilita microscopica di Xerocomus
pruinatus e la sua delimitazione da specie simili. Micologia e Vege-
tazione Mediterranea 16 : 46–54.
Ladurner, H. & Poder, R. (2000) A new hyphal type found in
Xerocomus pruinatus. Osterreichische Zeitschrift fur Pilzkunde 9 :
11–15.
Ladurner, H., Poder, R., Rocabruna, A. & Tabares, M. (2001)
Boletellus catalaunicus Poder, Moreno, Rocabruna et Tabares : a
synonym of Xerocomus ripariellus Redeuilh. Revista Catalana de
Micologia 23 : 121–125.
Lannoy, G. & Estades, A. (2001) Les Bolets. Flore Mycologique
d’Europe, Amiens.
Moncalvo, J. M., Drehmel, D. & Vilgalys, R. (2000a) Variation in
modes and rates of evolution in nuclear and mitochondrial ribo-
somal DNA in the mushroom genus Amanita (Agaricales, Basi-
diomycota): phylogenetic implications. Molecular Phylogenetics
and Evolution 16 : 48–63.
Moncalvo, J. M., Lutzoni, F. M., Rehner, S. A., Johnson, J. &
Vilgalys, R. (2000b) Phylogenetic relationships of agaric fungi
Xerocomus cisalpinus sp. nov. and the X. chrysenteron complex 678

based on nuclear large subunit ribosomal DNA sequences. Sys-
tematic Biology 49 : 278–305.
Moncalvo, J. M., Vilgalys, R., Redhead, S. A., Johnson, J., James,
T. Y., Aime, M. C., Hofstetter, V., Verduin, S. J. W., Larsson, E.,
Baroni, T. J., Thorn, R. G., Jacobsson, S., Clemencon, H. &
Miller, O. K. jr (2002) One hundred and seventeen clades of
euagarics. Molecular Phylogenetics and Evolution 23 : 357–400.
Oolbekkink, G. T. (1991) The taxonomic value of the ornamentation
in the Xerocomus group of Boletus. Persoonia 14 : 245–273.
Persoon, C. H. (1825) Mycologia Europaea, vol. 2. J. J. Palmi,
Erlangen.
Posada, D. & Crandall, K. A. (1998) Modeltest: testing the model of
DNA substitution. Bioinformatics 14 : 817–818.
Quelet, L. (1895) Quelques especes critiques ou nouvelles de al flore
mycologique de la France. Compts Render d’ Association francais
de Avancement du Science 20.
Redeuilh, G. (1997) Xerocomus ripariellus Redeuilh sp. nov. Docu-
ments Mycologiques 26 : 30–31.
Redeuilh, G. (1998) Une nouvelle section dans le genre Xerocomus
(Boletaceae). Xerocomus sectio Striatulispori sect. nov. Documents
Mycologiques 28 : 73–74.
Redeuilh, G. & Simonini, G. (1994) Comitato per la unificazione dei
nomi dei boleti Europei; Schede Xerocomus. Atti delle II Giornate
Europee di Micologia Mediterranea C.E.M.M. (A.E.). Nuoro.
Schreiner, J. (2000) Xerocomus ripariellus fur Deutschland nachge-
wiesen. Zeitschrift fur Mykologie 66 : 151–160.
Seguy, E. (1936) Code Universel des Couleurs. Seguy, Paris.
Simonini, G. (1994) Boletus dryophilus Thiers, specie nuova oer
Europa. Rivista di Micologia 37 : 205–219.
Simonini, G. (1996) Delimitazione di alcuni Xerocomus, con l’ausilio
dell’esame della struttura cuticolare e delle spore. Atti delle Gior-
nate C.E.M.M. 2 : 45–72.
Simonini, G. (1998a) Xerocomus chrysenteron e Xerocomus rubellus :
delimitazione e casi di simulazione. Micologia e Vegetazione
Mediterranea 13 : 69–89.
Simonini, G. (1998b) Fungi non delineati VI: Qualche specie rara o
poco conosciuta della Famiglia Boletaceae. Libreria Mykoflora,
Alassio.
Singer, R. (1945) New Boletaceae from Florida (a preliminary com-
munication). Mycologia 37 : 797–799.
Snell, W. H. & Dick, E. A. (1958) Notes on Boletes. X. A few
miscellaneous discussions and a new subspecies. Mycologia 50 :
57–65.
Swofford, D. L. (1998) PAUP*4.00: phylogenetic analysis using par-
simony (and other methods). Version beta version 4.0d64. Sinauer
Associates, Sunderland, MA.
Thomas, K. A., Peintner, U., Moser, M. & Manimohan, P. (2002)
Anamika, a new mycorrhizal genus of Cortinariaceae from India
and its phylogenetic position based on ITS and LSU sequences.
Mycological Research 108 : 245–251.
Vilgalys, R. & Hester, M. (1990) Rapid genetic identification
and mapping of enzymatically amplified ribosomal DNA from
several Cryptococcus species. Journal of Bacteriology 172 :
4238–4246.
Wagner, T. & Fischer, M. (2001) Natural groups and a revised system
for the European poroid Hymenochaetales (Basidiomycota) sup-
ported by nLSU rDNA sequence data. Mycological Research 105 :
773–782.
Zolan, M. E. & Pukkila, P. J. (1986) Inheritance of DNAmethylation
in Coprinus cinereus. Molecular and Cellular Biology 6 : 195–200.
Corresponding Editor: S. A. Redhead
U. Peintner, H. Ladurner and G. Simonini 679