rationale behind using valproic acid for non-hodgkin
TRANSCRIPT

7486
CONCLUSIONS: Based on its biomolecular mechanism of action, VPA appears to be a prom-ising initial treatment before initiating the stan-dard treatment in patients with NHL to overcome resistance.
Key Words:Epigenetics, Histone deacetylase inhibitors, Lym-
phoma, Non-Hodgkin lymphoma, Valproic acid.
Introduction
In 2021, 81,560 estimated cases of incidence and 20,720 estimated cases of mortality have been associated with Non-Hodgkin lymphoma (NHL), a hematological malignancy that is treatable but relatively incurable1. Some patients with NHL experience relapse and are resistant to existing standard therapies2-5. The estimated 5-year sur-vival rate for these patients was 73.2% from 2011 to 20171.
The primary concern regarding NHL treatment is achieving an optimal cure while improving the clinical outcome. Several studies6-9 have been conducted to resolve this concern. Epigenetics is one of the most investigated domains for devel-oping an optimal treatment for NHL because epi-genetic dysregulation has been suggested to play a role in lymphomagenesis10.
Several histone deacetylases (HDAC)-inhib-iting epigenetic drugs have been studied as po-
Abstract. – OBJECTIVE: Non-Hodgkin lym-phoma (NHL) is a hematological malignancy with a high rate of relapse and refractory cases. It is believed to be caused by resistance to stan-dard treatment modalities. Valproic acid (VPA), previously used as a broad-spectrum anticon-vulsant drug, has been proposed for NHL owing to its action of epigenetic modification by inhib-iting histone deacetylase. However, VPA studies on NHL are limited. This review describes the ra-tionale behind the use of VPA for NHL treatment, particularly focusing on its molecular mecha-nism of action.
MATERIALS AND METHODS: This is a narra-tive review. The literature search strategy for in-dexed Scopus articles was performed random-ly using PubMed and MEDLINE as the primary sources. No specific term was used.
RESULTS: Several mechanisms are respon-sible for NHL development. VPA can modulate these mechanisms via epigenetic and nonepi-genetic modifications. It may also have an im-pact on the proteins responsible for treatment resistance. The mechanisms of action of VPA in NHL are as follows: the induction of cell cy-cle arrest via the upregulation of cyclin-depen-dent protein kinase inhibitors; induction of Apo2 ligand or tumor necrosis factor-related apopto-sis-inducing ligand-induced apoptosis; inacti-vation of B-cell lymphoma 6; inhibition of Janus kinase/signal transducer and activator of tran-scription, phosphoinositide 3-kinase/Akt, and nuclear factor kappa B signaling pathways; up-regulation of tumor antigen as the primary target of immunotherapy; and strengthening of tumor immunosurveillance.
European Review for Medical and Pharmacological Sciences 2021; 25: 7486-7500
H. RIDWANSYAH1, I. WIJAYA2, M.H. BASHARI3, A.H.S. KARTAMIHARDJA4, B.S. HERNOWO5
1Graduate School of Biomedical Science, Faculty of Medicine, Universitas Padjadjaran, Bandung, Indonesia2Division of Hematology and Oncology, Department of Internal Medicine, Faculty of Medicine, Universitas Padjadjaran, Hasan Sadikin General Hospital, Bandung, Indonesia 3Division of Pharmacology and Therapy, Department of Basic Medical Sciences, Faculty of Medicine, Universitas Padjadjaran, Bandung, Indonesia 4Department of Nuclear Medicine and Molecular Imaging, Faculty of Medicine, Universitas Padjadjaran, Bandung, Indonesia 5Department of Anatomical Pathology, Faculty of Medicine, Universitas Padjadjaran, Bandung, Indonesia
Corresponding Author: Hastono Ridwansyah, MD; e-mail: [email protected]
Rationale behind using valproic acid for Non-Hodgkin lymphoma: a biomolecular perspective

Rationale behind using valproic acid for Non-Hodgkin lymphoma: a biomolecular perspective
7487
tential treatment options for NHL; these drugs may overcome epigenetic dysregulation. Some examples of HDAC inhibitors (HDACis) include romidepsin, vorinostat, chidamide, belinostat, valproic acid (VPA), and others11. HDACis can in-hibit HDACs that are overexpressed in lymphoma cells. These enzymes are important for regulat-ing certain protein expressions involved in cell growth, proliferation, and survival11.
VPA is more accessible and affordable than other HDACis because it is commonly used as a broad-spectrum anticonvulsant drug, mood stabiliz-er, and migraine prophylactic12. Drott et al13 reported that VPA in combination with rituximab, cyclophos-phamide, hydroxydaunorubicin, oncovin, and pred-nisone (R-CHOP) showed a positive outcome and tolerable safety level in patients with diffuse large B-cell lymphoma (DLBCL) in a phase 1 clinical tri-al13. This trial promises beneficial outcomes for DL-BCL. A deeper understanding of the biomolecular aspects of VPA for NHL will help understand the potential of this repurposed drug. This review de-scribes the possibility of using VPA for overcoming resistance to standard treatments in NHL.
Epigenetic Modification of Non-Hodgkin Lymphoma
Epigenetic alteration, which is partly involved in the progression of lymphoma, has become the focus of many scientists aiming to treat malignant lymphoma14. Epigenetic modification is a process through which gene expression is altered without any changes in the DNA sequence15. Three pro-cesses are involved in epigenetic modification: DNA methylation by DNA methyltransferases, histone modification by histone acetyltransferas-es (HATs) and HDACs, and translational repres-sion by noncoding RNA, including microRNA16.
Gene expression in epigenetic modification is dependent on chromatin conformations: heterochro-matin and euchromatin. Heterochromatin contains many inactive genes that are silenced, whereas eu-chromatin contains active genes that are accessible for binding by certain transcription factors16. Silenc-ing of tumor suppressor genes, DNA repair proteins, and cell cycle control proteins or enzymes plays an important role in lymphomagenesis17.
Highly condensed chromatin can repress tran-scription. This chromatin conformation is sup-ported by the methylation of DNA, primarily at the CpG island, by DNA methyltransferases (DNMT). The methylation of histone tails by the enhancer of zeste homolog 2 (EZH2), a catalyzer that adds a methyl group to a histone protein, and
deacetylation of histone tails by HDACs promote the heterochromatin topology. DNMT and EZH2 inhibitors can alter highly condensed chromatin into loose chromatin (euchromatin). By contrast, the transcriptionally permissive state of chroma-tin is endorsed by HDACis and HAT17.
Genes encoding HATs, such as CREBBP and EP300, undergo a loss-of-function mutation in 25% of patients with DLBCL and in 60% of patients with follicular lymphoma. This mutation leads to the silencing or repression of some proteins that are essential for inhibiting lymphoma progression. The loss or mutation of CREBBP or EP300 is an important biomarker for identifying patients with lymphoma that can receive HDACis18.
Role of HDACs in NHLHDACs are enzymes that catalyze the remov-
al of acetyl groups from the lysine and arginine residues of histones19. They can modify histone and several nonhistone proteins in some organ-elles. Certain proteins that function as apoptotic regulators, including p53 and B cell lymphoma 6 (Bcl-6), have been reported to be directly acetyl-ated by HDACs20,21. Aberrant HDAC activity can alter gene expression and some cellular pathways related to cell death, growth, or survival22.
There are four classes of HDACs in the human body, which are categorized based on their function and DNA sequence (Table I). HDACs can regulate certain proteins involved in tumor progression; thus, the presence or overexpression of some HDACs is associated with poor survival in NHL23,24.
Several HDACs may contribute to the inhibi-tion of cell death and progression of the lymphoma cell cycle25-27. HDACs also interfere with immu-nosurveillance, thereby playing an important role in the growth of lymphoma cells28. Furthermore, HDAC6, a class IIb enzyme, can impair CD20 ex-pression as a specific target for immunotherapy, such as that with rituximab; therefore, rituximab resistance may occur in lymphoma cells29.
HDACi for NHL TreatmentIn NHL, HDAC inhibition might be a target
for certain treatments18. HDACis are epigenetic drugs whose action is mediated by chromatin re-modeling via histone modification and by acetyl-ating the nonhistone proteins that impact tumor cell growth and survival. HDACis can alter the expression of several genes involved in lymphom-agenesis, thereby promoting tumor cell death, growth arrest, differentiation, and senescence as well as inhibiting angiogenesis22,30

H. Ridwansyah, I. Wijaya, M.H. Bashari, A.H.S. Kartamihardja, B.S. Hernowo
7488
Several HDACis have been approved by the US Food and Drug Administration (FDA) for treating hematological malignancies. HDACis that have been approved by the US FDA include vorinostat for cutaneous T-cell lymphoma (CTCL; 2006), ro-midepsin for CTCL (2009) and peripheral T-cell lymphoma (PTCL; 2011), belinostat for PTCL (2014), and panobinostat for multiple myeloma (2015)18,31-35. Each drug generally improves patient outcomes with a modest level of activity. When an HDACi is combined with another chemother-apeutic treatment, it demonstrates better effi-cacy but with a high rate of moderate-to-severe adverse events. Therefore, therapies combining various HDACis with other treatments should be broadly investigated in clinical trials to determine their efficacy and safety in NHL before approval.
Some HDACis that exhibit antitumor activity in lymphoma by inhibiting HDACs are shown in Table II36-57. Several HDACis have been approved by the FDA for T-cell lymphomas, although some are under investigation for B-cell lymphomas.
VPAVPA has been commonly used as a broad-spec-
trum anticonvulsant drug, mood stabilizer, and mi-graine prophylactic. It was first synthesized from Valeriana officinalis by Burton58. VPA is a short-
chain branched fatty acid administered orally or intravenously59. VPA has been reported to exert its epigenetic modulation effect via the inhibition of HDAC and help treat various malignancies12.
The bioavailability of VPA is >80% after oral ad-ministration, and its peak blood concentration is ob-served within 2 h59. Several pathways are employed in VPA metabolism in the human body, including glucuronidation, β-oxidation, and cytochrome P450 (CYP)-mediated oxidation pathways. Oxidation in-volving cytochrome P450 is employed by only 10% VPA metabolism pathways60-62. The half-life of VPA varies significantly from 9 to 18 h59.
As an antiseizure drug, an appropriate VPA dose for common use is initially 15 mg/kg, with slow titration to the therapeutic dose. Dosages of 25-30 mg/kg/day and even 60 mg/kg/day may be required for some patients. In plasma, the thera-peutic levels of VPA should be 350-700 µM for continuous VPA treatment in patients with epi-lepsy59. VPA dose for NHL treatment is relatively different from that used as an antiseizure, which is explained further in next section.
Mechanism of Action of VPAThe mechanism of action of VPA for NHL is
shown in Figure 1. Several signaling pathways are affected by VPA. It functions through HDAC
Table I. Histone deacetylase classification and location.
Class Name Location
Class I HDAC1, HDAC2, HDAC3, HDAC8 NucleusClass IIa HDAC4. HDAC5, HDAC7, HDAC9 Nucleus and cytoplasmClass IIb HDAC6, HDAC10 CytoplasmClass III SIRT 1-7 Nucleus and cytoplasmClass IV HDAC11 Nucleus and cytoplasm
Abbreviations: HDAC: histone deacetylase; SIRT: Sirtuin.
Table II. HDACis in clinical trials for various hematological malignancies.
HDACi HDAC target Trial phase
Vorinostat19,31,36–38 Class I, II, and IV Marketed CTCL, phase II FL, MZL, MCL, HLRomidepsin17,32,33,39 Class I, II, and III Marketed CTCL, PTCLBelinostat40–43 Class I, II, and IV Marketed PTCL, phase II B-cell lymphomaPanobinostat34,44–47 Class I–IV Phase III MM, phase II CTCL, DLBCLMocetinostat48–50 Class I and IV Phase II DLBCL, FL, HLValproic acid12,13 Class I, II (except HDAC8 and 9) Phase II DLBCLChidamide51,52 HDAC 1, 2, 3, 10 Phase I/II B-NHL (ongoing), phase II PTCLAbexinostat53 Class I–IV Phase II CLL, B-NHL, T-NHLQuisinostat54,55 Class I and II Phase II CTCLEntinost at56,57 HDAC 1, 2, 3, 11 Phase II HL, leukemia

Rationale behind using valproic acid for Non-Hodgkin lymphoma: a biomolecular perspective
7489
inhibition-mediated histone 3 lysine 9 acetyla-tion63, nonhistone modification, and direct protein acetylation or dephosphorylation.
VPA Induces Cell-Cycle Arrest and Apoptosis by Upregulating p21 and p27
The survival of tumor cells depends on the cell cycle control system. The cell cycle is regulated by cyclin-dependent protein kinases (Cdks). Cyclins in a particular phase of the cell cycle can activate Cdks and induce cell cycle entry progression. Cdk inhibitor proteins (CKIs) comprising p27, p21, and p16 can limit cell cycle progression by suppressing G1/S-Cdk and S-Cdk activities after DNA dam-age, leading to the cessation of the cell cycle64.
Newbold et al65 reported that p21 (CDKN1A) does not contribute to cell cycle arrest or apoptosis. How-ever, high expression of p27 (CDKN1B) has been reported to induce apoptosis following G1 cell cycle arrest in a mouse model of B-cell lymphoma. This elevated expression occurred at the translational rather than at the transcriptional level65. A previous
study66 indicated that an HDACi (romidepsin) upreg-ulated p21 and promoted cell cycle arrest in mantle cell lymphoma (MCL). However, the data from this in vitro study were inconsistent. An in vivo study65 clarified these results by demonstrating that p21Waf1 is not essential in lymphomagenesis, although the aforementioned HDACi strongly influences it. Fi-nally, p27 is considered crucial because it induces apoptosis and cell cycle arrest in B-cell lymphoma through an epigenetic mechanism65.
Cyclin D1, which is essential for cell survival regulation, binds to Cdk4 and Cdk6 and promotes the cell cycle. Most patients with MCL exhibit high expression of cyclin D1; however, MCL de-velopment cannot adequately occur via a single genetic aberration67; it requires another oncogenic aberration, such as disruptions in the ataxia telan-giectasia mutated and p5368.
A subtype of NHL, MCL, is found in indolent variants with low p21 expression. By contrast, low p27 expression is found in aggressive vari-ants69. The implication of low expression of both
Figure 1. Proposed mechanism of VPA action in NHL. Abbreviations: Bcl-6, B-cell lymphoma 6; CD20: cluster of differen-tiation 20; C-FLIP: cellular FLICE-inhibitory protein; HDAC, histone deacetylase; IKKβ, inhibitor of NFκB kinase β; IRAK-1, interleukin-1 receptor-associated kinase-1; MHCII, major histocompatibility complex class II; NHL, non-Hodgkin lymphoma; NFκB, nuclear factor kappa B; PTEN, phosphatase and tensin homolog; PD1/PD-L1, programmed cell death protein 1/pro-grammed death-ligand 1; VPA, valproic acid.

H. Ridwansyah, I. Wijaya, M.H. Bashari, A.H.S. Kartamihardja, B.S. Hernowo
7490
CKIs is the progression of the cell cycle and pro-liferation of B-cell lymphoma. Some studies26,70 have demonstrated that the low expression of some CKIs, such as p21 and p57, is associated with HDAC1 and HDAC2 and that the low ex-pression of p27 is associated with a high expres-sion of HDAC3 in activated B-cell (ABC) DLB-CL. Eliminating HDAC1 and HDAC2 activities can normalize the expression of these CKIs.
Some studies71,72 investigating the effect of VPA as an HDACi on tumor cells suggest that VPA can inhibit tumor cell proliferation and sur-vival by upregulating the expression of p21Waf1/
Cip1 and p27Kip1. The increase in p21 expression is induced through the acetylation of the histone in the p21 promoter region. The enhancement of p21 expression can block retinoblastoma (Rb) phos-phorylation, thereby increasing the number of Rb/E2F complex activity. Reduction in free E2F can prevent DNA replication73.
VPA Promotes Apo2-L/Tumor Necrosis Factor (TNF)-Related Apoptosis-Inducing Ligand (TRAIL)-Induced Apoptosis by Downregulating Cellular FLICE-Inhibitory Protein
Apo2-L/TRAIL is a member of the TNF gene superfamily; it can induce apoptosis by bind-ing to the death receptors (DR) on the plasma membrane. As the primary receptors of Apo2-L/TRAIL, DR4 and DR5 have cytoplasmic death domains that transmit the apoptosis signal74,75.
DR4 and DR5 are well known as TRAIL-R1 and TRAIL-R2, respectively. Both mediate apop-tosis and are highly expressed in primary human B cells and B-cell lymphoma76-78. Five essential factors increase the sensitivity of this pathway: decoy receptors, cellular FLICE-inhibitory pro-tein (c-FLIP, an inhibitor of apoptosis), proteins, interferons, and nuclear factor kappa B (NFκB)74.
c-FLIP expression has been detected in 46.7% of patients with DLBCL79. c-FLIP, an antiapoptot-ic protein, inhibits the extrinsic apoptosis signal-ing pathway by blocking DR signaling, thereby disturbing caspase-8 activation at the death-in-ducing signaling complex80,81. The dysregulation of extrinsic apoptosis due to c-FLIP overexpres-sion contributes to tumor progression and sensi-tivity to chemotherapeutic drugs in PTCL25.
The suppression of c-FLIP can sensitize tu-mor cells to TRAIL-mediated apoptosis82. VPA, an inhibitor of HDAC, can modulate extrinsic apoptosis via certain mechanisms in various cancer cells25,83,84. Some studies25,83,84 have re-
ported that inhibiting HDAC1 and HDAC4 by VPA downregulates c-FLIP expression. When c-FLIP is downregulated, DR4 and DR5 are more sensitive to proapoptotic ligands.
VPA Inhibits the Janus Kinase (JAK)/Signal Transducers and Activators of Transcription (STAT) Pathway Through Epigenetic and Nonepigenetic Mechanisms
Depending on the signal, tissue, and cellular context, JAK/STAT signaling pathways mediate multiple cellular responses, including cell division, differentiation, migration, death, and cell surviv-al85. JAK proteins and phosphorylated STAT are activated after the binding of cytokines or growth factors to B-cell receptors. This leads to STAT dimerization, which then translocates into the nu-cleus where it activates various effector genes to express Bcl2, myc, cyclin D1, and VEGF86.
DLBCL, an aggressive type of B-cell lymphoma, deregulates the JAK/STAT pathway and drives tu-mor cells toward prolonged survival87. The suppres-sor of cytokine signaling (SOCS) proteins regulates cell survival by inhibiting JAKs. The loss-of-func-tion mutation of SOCS1 functions as a tumor sup-pressor gene (TSG) and induces tumor cell progres-sion. A group of patients with this mutation showed shorter overall survival (OS)88. Based on these find-ings, SOCS1 was studied as a potential target for the treatment of B-cell lymphoma89-91.
Unlike SOCS1, SOCS3 does not function as a TSG in T-cell lymphoma92. Although the role of epigenetic mechanisms at the level of SOCS expression in lymphoma cells remains unclear, Kim et al93 reported that HDACis modulate the expression of SOCS1 and SOCS3 in cervical can-cer cells and that histone acetylation by VPA acti-vates TSG SOCS1 and SOCS3 in colon carcinoma cells. Tumor cells subsequently undergo apopto-sis94. However, the role of epigenetic mechanism of VPA action in modulating SOCS expression in lymphoma remains unclear.
VPA directly inhibits STAT3 through dephos-phorylation, thereby inactivating JAK/STAT sig-naling, which reduces tumor cell proliferation and induces apoptosis95,96. Ni et al97 reported that the inhibition of HDAC3 by VPA might suppress STAT3 in natural killer cells, suggesting that histone and nonhistone protein modifications by VPA regulate the JAK/STAT pathway in some cancers. However, further studies are warranted to completely elucidate VPA action through JAK/STAT pathway modulation in lymphoma.

Rationale behind using valproic acid for Non-Hodgkin lymphoma: a biomolecular perspective
7491
VPA Inhibits Akt/mammalian Target of Rapamycin (mTOR) in the Phosphoinositide 3-Kinase (PI3K)/Akt/mTOR Signaling Pathway
The PI3K/Akt signaling pathway is essential for regulating the cell cycle and is directly asso-ciated with cell survival and proliferation in lym-phoma98. It is also associated with tumorigenesis in human cancer stem cells99. Multiple aberrations of several genes associated with PI3K/Akt signal-ing drive cells toward malignancy. The mTOR is a downstream protein of this pathway that can reg-ulate cell growth and death. Gene amplification and mutation of mTOR contribute to malignancy in various cancers100.
Phosphatase and tensin homolog (PTEN), a pro-tein that can inhibit PI3K and Akt activation, ceases the PI3K/Akt signaling pathway98. The loss-of-func-tion gene mutation of PTEN plays a vital role in the oncogenic activation of B-cell lymphoma. The hy-peractivity of Akt signaling is associated with poor survival in patients with DLBCL101,102. The role of HDACs regarding the low expression of PTEN is epigenetically unclear in lymphoma. However, in another type of solid tumor, such as gastric cancer cells, HDAC1/2 inhibition leads to an increased PTEN expression and promotes apoptosis and auto-phagy. Therefore, PTEN expression might be partly downregulated by HDAC1/2 presence103.
The overexpression of some HDACs occurs in various cancer cells, including hematological ma-lignancy (such as lymphoma)51. HDAC1 inhibition by VPA can increase PTEN expression, thereby influencing Akt to follow the Akt/mTOR signal-ing pathway. Finally, VPA can induce cell death via autophagy and apoptosis, which are associat-ed with Akt and mTOR protein activities103,104.
The Akt/mTOR pathway is associated with the JAK/STAT signaling pathway. mTOR can promote the dimer form of STAT to activate on-cogenic or prosurvival transcription in the nucle-us86. Based on this explanation, VPA directly in-hibits cell proliferation and limits cell survival by disrupting several signaling pathways.
VPA Inactivates Bcl-6 and Upregulates p53p53 protein plays a vital role in response to cel-
lular DNA damage induced by various factors. It drives cellular apoptosis through intrinsic and ex-trinsic mechanisms. It can also upregulate p21 via cell cycle arrest because it acts as a potent inhib-itor of Cdks 1, 2, 3, and 4. Many NHL subtypes have low expressions of p53 and p21, which leads to a reduction in DNA damage response, preven-
tion of apoptosis, and continuation of cell cycle progression10.
Bcl-6 is an oncogene involved in lymphoma-genesis, primarily in germinal center B-cell DL-BCL. It requires the formation of complexes with certain HDACs to prevent the transcription acti-vation of p53, leading to the expansion of germi-nal center B cells. Bcl-6 is activated via interac-tion with HDACs 4, 5, 7, and 9 as corepressors27. By contrast, Bcl-6 inactivation results in an in-crease in p53 and apoptosis in DLBCL105.
VPA plays a vital role in inhibiting most of the class I and II HDACs and disrupting Bcl-6 activ-ities. Without HDACs, Bcl-6 becomes inactive. This condition leads to an increased expression of p53 and p21. Thus, tumor cells may undergo apoptosis96,106.
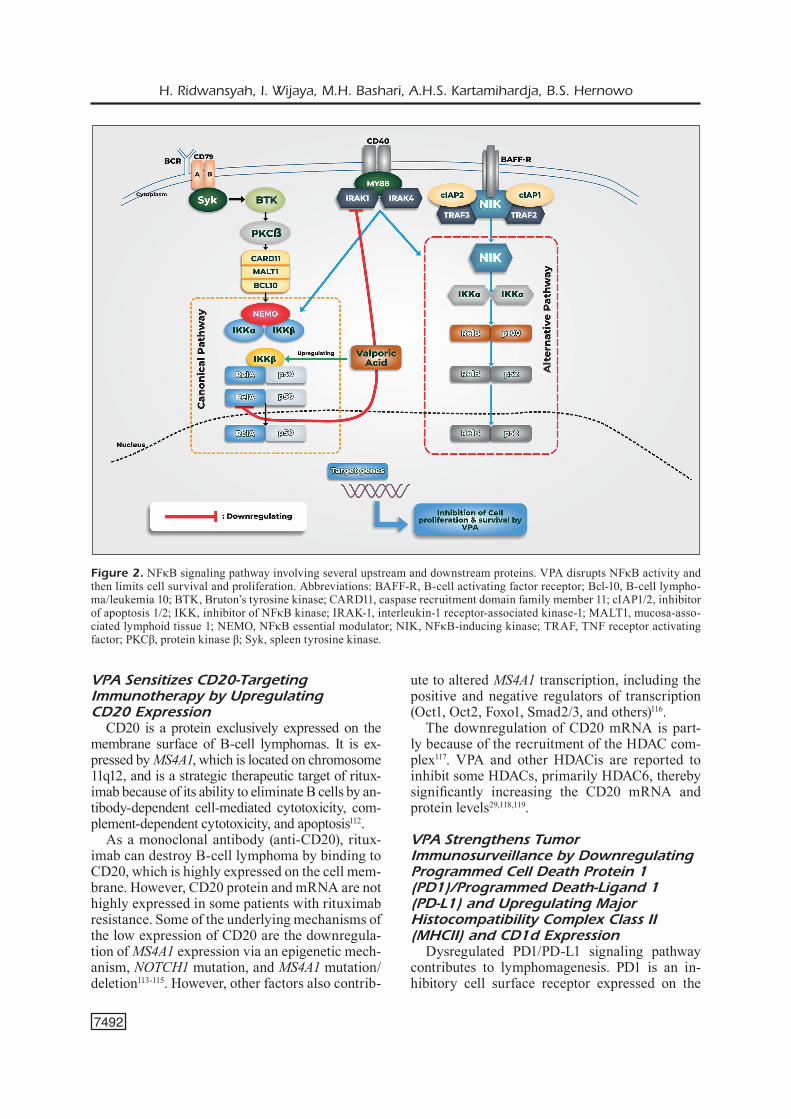
VPA Inhibits the NFκB Signaling Pathway
The NFκB signaling pathway is often employed for cell survival. This pathway is of two types: classic and alternative. The canonical pathway of NFκB is initiated by the induction of B-cell re-ceptor and CARD11/MALT1/Bcl-10 complex, which then phosphorylate the inhibitor of NFκB kinase (IKK). This process activates the inhibitor of NFκB alpha. Heterodimer RelA/p50 then trans-locates into the nucleus and is responsible for cell survival, proliferation, inflammation, and innate immunity86. NFκB-inducing kinase (NIK) initiates the alternative pathway of NFκB via the activation of BAFF-R, MY88, and IKK. NIK-induced RelB/p50 and RelB/p52 transcription factors then pro-mote lymphoid organogenesis, adaptive immunity, anti-inflammation, and B-cell maturation107.
The persistent activation of NFκB signaling ap-pears in ABC-DLBCL and is responsible for cell survival108. This activation occurs because of a gain-of-function mutation in the genes of various proteins involved in the NFκB pathway, including CD79A, CD79B, CARD11, MYD88, Bcl-10, MALT1, and REL; this constitutively activates the downstream signal for cell proliferation and survival107,109.
VPA can attenuate NFκB activity by inhibit-ing the upstream and downstream mediators of the NFκB signaling pathway (Figure 2). It can also reduce the expression of interleukin-1 recep-tor-associated kinase-1 as an upstream mediator of NFκB activation. Studies110,111 have shown that VPA downregulates RelA (p65) but upregulates IκBa via nonhistone modifications. The mecha-nism of this action, particularly in another NHL subtype, should be confirmed in future studies.
H. Ridwansyah, I. Wijaya, M.H. Bashari, A.H.S. Kartamihardja, B.S. Hernowo
7492
VPA Sensitizes CD20-Targeting Immunotherapy by Upregulating CD20 Expression
CD20 is a protein exclusively expressed on the membrane surface of B-cell lymphomas. It is ex-pressed by MS4A1, which is located on chromosome 11q12, and is a strategic therapeutic target of ritux-imab because of its ability to eliminate B cells by an-tibody-dependent cell-mediated cytotoxicity, com-plement-dependent cytotoxicity, and apoptosis112.
As a monoclonal antibody (anti-CD20), ritux-imab can destroy B-cell lymphoma by binding to CD20, which is highly expressed on the cell mem-brane. However, CD20 protein and mRNA are not highly expressed in some patients with rituximab resistance. Some of the underlying mechanisms of the low expression of CD20 are the downregula-tion of MS4A1 expression via an epigenetic mech-anism, NOTCH1 mutation, and MS4A1 mutation/deletion113-115. However, other factors also contrib-
ute to altered MS4A1 transcription, including the positive and negative regulators of transcription (Oct1, Oct2, Foxo1, Smad2/3, and others)116.
The downregulation of CD20 mRNA is part-ly because of the recruitment of the HDAC com-plex117. VPA and other HDACis are reported to inhibit some HDACs, primarily HDAC6, thereby significantly increasing the CD20 mRNA and protein levels29,118,119.
VPA Strengthens Tumor Immunosurveillance by DownregulatingProgrammed Cell Death Protein 1 (PD1)/Programmed Death-Ligand 1(PD-L1) and Upregulating Major Histocompatibility Complex Class II (MHCII) and CD1d Expression
Dysregulated PD1/PD-L1 signaling pathway contributes to lymphomagenesis. PD1 is an in-hibitory cell surface receptor expressed on the
Figure 2. NFκB signaling pathway involving several upstream and downstream proteins. VPA disrupts NFκB activity and then limits cell survival and proliferation. Abbreviations: BAFF-R, B-cell activating factor receptor; Bcl-10, B-cell lympho-ma/leukemia 10; BTK, Bruton’s tyrosine kinase; CARD11, caspase recruitment domain family member 11; cIAP1/2, inhibitor of apoptosis 1/2; IKK, inhibitor of NFκB kinase; IRAK-1, interleukin-1 receptor-associated kinase-1; MALT1, mucosa-asso-ciated lymphoid tissue 1; NEMO, NFκB essential modulator; NIK, NFκB-inducing kinase; TRAF, TNF receptor activating factor; PKCβ, protein kinase β; Syk, spleen tyrosine kinase.

Rationale behind using valproic acid for Non-Hodgkin lymphoma: a biomolecular perspective
7493
membrane of the effector T cell, whereas PD-L1 is expressed on antigen-presenting cells (APCs) and overexpressed in several lymphoma cells, in-cluding Hodgkin lymphoma, ABC-DLBCL, and primary mediastinal B-cell lymphoma. When PD-L1 is overexpressed in lymphoma cells, it can cause T cell exhaustion and immune escape86. High HDAC6 expression has been detected in the B cells of patients with chronic lymphocytic leukemia and DLBCL. The selective inhibition of HDAC6 can reduce the inhibitory molecule PD1/PD-L128. VPA can attenuate the immunosuppres-sive function of APC through some mechanisms, including interleukin 4 receptor-α/arginase axis, PD-L1, toll-like receptor 4 signaling pathway, and Rb1 derepression, thereby maintaining tumor im-munosurveillance120.
CD1d is an antigen-presenting molecule at the surface membrane of B cells that can bind spe-cifically to the T-cell receptor on natural killer cells. HDAC2 binds to the CD1d promoter, which dysregulates the presentation of the tumor antigen by CD1d. HDAC2 inhibition by several HDACis can increase CD1d-mediated antigen presentation at the surface membrane of B cells in MCL121,122. VPA having inhibitory activity against HDAC2 is suggested to increase tumor immunosurveillance via this mechanism.
Major histocompatibility complex class II (MHCII) is a protein on the cell membrane that presents antigens to CD4+ T cells to initiate an-tigen-specific immune responses. It is constitu-tively expressed on dendritic cells, B cells, mac-rophages, and activated T cells. Therefore, it has an important function in tumor immunosurveil-lance. MHC class II transactivator (CIITA) has been reported to be a key modulator of MHCII transcription. The lack of MHCII partly contrib-utes to the poor survival of patients with DLBCL. The inhibition of class I HDAC, primarily via the inhibition of HDAC1, can significantly increase the expression of CIITA and MHCII123. Therefore, VPA can strengthen immunosurveillance primar-ily via HDAC1 inhibition.
Clinical Trials of VPA for NHLA case report124 demonstrated that VPA led to
a complete response in a 64-year-old woman with stage IV refractory germinal center DLBCL. This previously untreated patient received R-CHOP as the first-line treatment, but after several months, she suffered recurrence with disease progression. She then received R-ESHAP for two cycles but showed no response. Thereafter, she received
R-ICE and salvage (taxol, topotecan, and ritux-imab) treatments but again showed no response. Approximately 1 year later, she received 250-mg VPA thrice a day for pain relief and as an an-ti-lymphoma agent. She showed a complete treat-ment response after 6 months of VPA administra-tion, with no adverse events124.
In their VALFRID study, Drott et al13 report-ed that VPA use with R-CHOP resulted in better OS in patients with DLBCL, in which 96.8% of 33 patients showed 2-year OS with tolerable side effects13. The study also demonstrated significant improvement in the 2-year progression-free sur-vival rate, which was 84.7% among the patients. While administering VPA to treat patients with DLBCL, the maximum tolerated dose (MTD) of 60 mg/kg/day for 3 days should be considered on days 1–3, which results in the plasma VPA con-centration of 600-1200 µM13.
Various adverse events associated with VPA administration have been reported. Munster et al125 reported that the administration of 120 mg/kg/day VPA might lead to somnolence, confu-sion, hearing loss, and other neurovestibular dis-orders. However, the previously reported serious adverse events were not noted in the VALFRID study13 because the MTD of VPA was only 60 mg/kg/day. However, the dosage did result in hearing disorders; thus, VPA requires close moni-toring for adverse events. Other side effects relat-ed to VPA administration were tolerable13; how-ever, this phase I clinical trial13 included only 33 patients with DLBCL. Therefore, these findings should be validated in a subsequent phase trial us-ing an appropriate study design with a larger sam-ple size to confirm the efficacy and safety of VPA.
A study evaluating the efficacy and safety of valproate involving 44 patients with CTCL was conducted in combination with hydralazine as a type of DNA methyltransferase inhibitor. The to-tal dose of valproate was 30 mg/kg t.i.d. daily in constitutive 28-day cycles in a phase II clinical trial126. The trial concluded that the combination of valproate and hydralazine is safe and effective in CTCL, although it cannot be compared with vorinostat and romidepsin in terms of efficacy. However, this drug combination should be ex-tensively investigated by including more patients with CTCL126.
Future Direction
Various subtypes of NHL are resistant to some of the available therapies, including chemothera-py, immunotherapy, and radiotherapy. VPA has

H. Ridwansyah, I. Wijaya, M.H. Bashari, A.H.S. Kartamihardja, B.S. Hernowo
7494
the potential to modulate several proteins asso-ciated with resistance to standard therapeutic modalities. The mechanism of resistance varies based on the type of therapy.
Several signaling pathways, such as PI3K/Akt, NFκB, and JAK/STAT3, are responsible for resis-tance to R-CHOP in NHL127. Based on previous-ly described mechanisms, particular proteins in each pathway may be regulated by VPA through epigenetic mechanisms.
Upon activation via phosphorylation, STAT3 contributes to resistance to radiotherapy in B-cell lymphoma128,129. STAT3 inactivation or dephos-phorylation induced by VPA may increase the sensitivity of the JAK/STAT3 signaling pathway to radiotherapy.
Low CD20 expression is one of the underlying mechanisms of rituximab resistance. Rituximab can function only by binding to the specific pro-tein CD20117,130. VPA can upregulate CD20 ex-pression in B-cell lymphoma through epigenetic modification.
NHL can be resistant to several PD1 inhibi-tors. Drug resistance in NHL is caused by the low expression of MHCI and MHCII. MHCs are as-sociated with a strong response to PD1 blockade induced by PD1 inhibitors131. VPA can upregu-late the expression of MHCI and MHCII, thereby strengthening tumor immunosurveillance.
Based on the biomolecular mechanism of ac-tion, the use of VPA with other standard treatments should be further evaluated in well-designed trials. This would clarify whether VPA can significantly optimize the efficacy of standard therapy in clini-cal settings. Furthermore, this will help overcome resistance to immunotherapy, chemotherapy, and radiotherapy in patients with NHL.
Conclusions
The use of VPA for NHL treatment is a topic of interest. The rationale for using VPA to treat NHL can be explained by its mechanism of ac-tion. VPA generally functions by regulating the HDAC–target protein axis, as explained previous-ly. The inhibition of HDACs by VPA results in the alteration of multiple signaling pathways that can inhibit lymphomagenesis by modulating several proteins. Moreover, proteins in several signaling pathways also contribute to the resistance of NHL to several standard treatment modalities. VPA ap-pears to be a promising initial treatment before the administration of the standard therapy in pa-
tients with NHL. However, further trials should be broadly conducted to establish VPA use for NHL treatment in clinical practice.
Authors’ ContributionsH.R. and I.W. conceptualized the study; H.R. wrote first original draft; I.W., M.H.B., B.S.H., and A.H.S.K. reviewed the draft and revised it substantially; B.S.H. acted as the supervisor of the team and performed the final revision of the manuscript. All authors have read and agreed to the published version of the manuscript.
AcknowledgmentsThis work was supported by Program Magister Dok-tor Menuju Sarjana Unggul (PMDSU) Kementerian Riset dan Teknologi dan Pendidikan Tinggi Republik Indonesia (No: 2928/UN6.3.1/LT/2020) and Academic Leadership Grant from Universitas Padjadjaran.
Conflicts of InterestThe authors declare no conflicts of interest.
References
1) National Cancer Institute. Cancer Stat Facts: Non-hodgkin lymphoma [Internet]. 2021 [cited 2021 Sept 11]. Available from: https://seer.cancer.gov/statfacts/html/nhl.html.
2) Salles G, Barrett M, Foà R, Maurer J, O’Brien S, Valente N, Wenger M, Maloney DG. Rituximab in B-cell hematologic malignancies: a review of 20 years of clinical experience. Adv Ther 2017; 34: 2232-2273.
3) Crump M, Neelapu SS, Farooq U, Van Den Neste E, Kuruvilla J, Westin J, Link BK, Hay A, Cerhan JR, Zhu L, Boussetta S, Feng L, Maurer MJ, Na-vale L, Wiezorek J, Go WY. Outcomes in refrac-tory diffuse large B-cell lymphoma: results from the international SCHOLAR-1 study. Blood 2017; 130: 1800-1808.
4) Wu JQ, Song YP, Su LP, Zhang MZ, Li W, Hu Y, Zhang XH, Gao YH, Niu ZX, Feng R, Wang W, Peng JW, Li XL, Ouyang XN, Wu CP, Zhang WJ, Zeng Y, Xiao Z, Liang YM, Zhuang YZ, Wang JS, Sun ZM, Bai H, Cui TJ, Feng JF. Three-year fol-low-up on the safety and effectiveness of rituxi-mab plus chemotherapy as first-line treatment of diffuse large B-cell lymphoma and follicular lym-phoma in real-world clinical settings in china: a prospective, multicenter, noninterventional study. Chin Med J 2018; 131: 1767-1775.
5) Bellei M, Foss FM, Shustov AR, Horwitz SM, Mar-cheselli L, Kim WS, Cabrera ME, Dlouhy I, Na-gler A, Advani RH, Pesce EA, Ko YH, Martinez

Rationale behind using valproic acid for Non-Hodgkin lymphoma: a biomolecular perspective
7495
V, Montoto S, Ciattone C, Moskowitz A, Spina M, Biasoli I, Manni M, Federico M. The outcome of peripheral T-cell lymphoma patients failing first-li-ne therapy: a report from the prospective, Inter-national T-Cell Project. Haematologica 2018; 103: 1191-1197.
6) Perksy DO, Li H, Rimsza LM, Barr PM, Popplewell LL, Bane CL, Gehr AV, LeBlanc M, Fisher RI, Smi-th SM, Friedberg JW. A phase I/II trial of vorino-stat (SAHA) in combination with rituximab-CHOP in patients with newly diagnosed advanced stage diffuse large B cell lymphoma (DLBCL): SWOG S0806. Am J Hematol 2018; 93: 486-493
7) Martin P, Bartlett NL, Chavez JC, Reagan JL, Smith SM, LaCasce AS, Jones JA, Drew J, Wu C, Mulvey E, Revuelta MV, Cerchietti L, Leonard JP. A phase I, open-label, multicenter trial of oral azacitidine (cc-486) plus R-CHOP in patients with high-risk, previously untreated diffuse lar-ge B cell lymphoma, grade 3B follicular lympho-ma or transformed lymphoma. Blood 2021; blo-od.2021011679
8) Vitale C, Strati P. CAR T-cell therapy for B-cell non-Hodgkin lymphoma and chronic lymphocytic leukemia: clinical trials and real-world experien-ces. Front Oncol 2020; 10: 849
9) Wang N, Meng Y, Wu Y, He J, Liu F. Efficacy and safety of chimeric antigen receptor T cell immu-notherapy in B-cell non-Hodgkin lymphoma: a sy-stematic review and meta-analysis. Immunothe-rapy 2021; 13: 345-357
10) Shaffer AL, Young RM, Staudt LM. Pathogenesis of human B cell lymphomas. Annu Rev Immunol 2012; 30: 565-610.
11) Chen IC, Sethy B, Liou JP. Recent update of HDAC inhibitors in lymphoma. Front Cell Dev Biol 2020; 8: 576391.
12) Chateauvieux S, Morceau F, Dicato M, Diederich M. Molecular and therapeutic potential and toxi-city of valproic acid. J Biomed Biotechnol 2010; 2010: 479364.
13) Drott K, Hagberg H, Papworth K, Relander T, Jer-keman M. Valproate in combination with rituximab and CHOP as first-line therapy in diffuse large B-cell lymphoma (VALFRID). Blood Adv 2018; 2: 1386-1392.
14) Ribeiro ML, Reyes-Garau D, Armengol M, Fer-nández-Serrano M, Roué G. Recent advances in the targeting of epigenetic regulators in B-cell non-Hodgkin lymphoma. Front Genet 2019; 10: 986.
15) Lakshmaiah KC, Jacob LA, Lokanatha D, Sal-danha SC. Epigenetic therapy of cancer with histone deacetylase inhibitors. J Can Res Ther 2014; 10: 469-478.
16) Booth S, Collins G. Epigenetic targeting in lym-phoma. Br J Haematol 2021; 192: 50-61.
17) Lue JK, Amengual JE, O’Connor OA. Epigenetics and lymphoma: can we use epigenetics to prime or reset chemoresistant lymphoma programs? Curr Oncol Rep 2015; 17: 40.
18) Sermer D, Pasqualucci L, Wendel H-G, Melnick A, Younes A. Emerging epigenetic-modulating therapies in lymphoma. Nat Rev Clin Oncol 2019; 16: 494-507.
19) Markozashvili D, Ribrag V, Vassetzky YS. Histo-ne deacetylase inhibitors and epigenetic regula-tion in lymphoid malignancies. Invest New Drugs 2015; 33: 1280-1291.
20) Yang H, Green MR. Epigenetic programing of B-cell lymphoma by BCL6 and its genetic deregu-lation. Front Cell Dev Biol 2019; 7: 272.
21) Reed SM, Quelle DE. P53 acetylation: regulation and consequences. Cancers 2015; 7: 30-69.
22) West AC, Johnstone RW. New and emerging HDAC inhibitors for cancer treatment. J Clin In-vest 2014; 124: 30-39.
23) Lee SH, Yoo C, Im S, Jung J-H, Choi HJ, Yoo J. Expression of histone deacetylases in diffuse lar-ge B-cell lymphoma and its clinical significance. Int J Med Sci 2014; 11: 994-1000.
24) Min SK, Koh YH, Park Y, Kim HJ, Seo J, Park H-R, Cho SJ, Kim IS. Expression of HAT1 and HDAC1, 2, 3 in diffuse large B-cell lymphomas, peripheral T-cell lymphomas, and NK/T-cell lymphomas. Ko-rean J Pathol 2012; 46: 142-150.
25) Zheng Z, Cheng S, Wu W, Wang L, Zhao Y, Shen Y, Janin A, Zhao W-L. c-FLIP is involved in tumor progression of peripheral T-cell lymphoma and targeted by histone deacetylase inhibitors. J He-matol Oncol 2014; 7: 88.
26) Yamaguchi T, Cubizolles F, Zhang Y, Reichert N, Kohler H, Seiser C, Matthias P. Histone deacetyla-ses 1 and 2 act in concert to promote the G1-to-S progression. Gene Dev 2010; 24: 455-469.
27) Lemercier C, Brocard M-P, Puvion-Dutilleul F, Kao H-Y, Albagli O, Khochbin S. Class II histone dea-cetylases are directly recruited by BCL6 transcriptio-nal repressor. J Biol Chem 2002; 277: 22045-22052.
28) Powers JJ, Maharaj KK, Sahakian E, Xing L, Pe-rezVillarroel P, Knox T, Quayle S, Jones SS, Vil-lagra A, Sotomayor EM, Pinilla-Ibarz J. Histone deacetylase 6 (HDAC6) as a regulator of immune check-point molecules in chronic lymphocytic leu-kemia (CLL). Blood 2014; 124: 3311.
29) Bobrowicz M, Dwojak M, Pyrzynska B, Stachura J, Muchowicz A, Berthel E, Venezia-Dalla N, Ko-zikowski M, Siernicka M, Miazek N, Zapala, P, Do-magala A, Bojarczuk K, Malenda A, Barankiewicz J, Graczyk-Jarzynka A, Zagozdzon A, Gabrysiak M, Diaz JJ, Karp M, Lech-Maranda E, Firczuk M, Giannopoulos K, Efremov DG, Laurenti L, Baa-tout D, Frenzel L, Malinowska A, Slabicki M, Zenz T, Zerrouqi A, Golab J, Winiarska M. HDAC6 inhi-bition upregulates CD20 levels and increases the efficacy of anti-CD20 monoclonal antibodies. Blo-od 2017; 130: 1628-1638.
30) Ghasemi S. Cancer’s epigenetic drugs: where are they in the cancer medicines? Pharmacogenomi-cs J 2020; 20: 367-379.
31) Olsen EA, Kim YH, Kuzel TM, Pacheco TR, Foss FM, Parker S, Frankel SR, Chen C, Ricker JL, Ar-

H. Ridwansyah, I. Wijaya, M.H. Bashari, A.H.S. Kartamihardja, B.S. Hernowo
7496
duino JM, Duvic M. Phase IIB multicenter trial of vorinostat in patients with persistent, progressive, or treatment refractory cutaneous T-cell lympho-ma. JCO 2007; 25: 3109-3115.
32) Whittaker SJ, Demierre M-F, Kim EJ, Rook AH, Lerner A, Duvic M, Scarisbrick J, Reddy S, Robak T, Becker JC, Samtsov A, McCulloch W, Kim YH. Final results from a multicenter, international, pi-votal study of romidepsin in refractory cutaneous T-cell lymphoma. JCO 2010; 28: 4485-4491.
33) Coiffier B, Pro B, Prince HM, Foss F, Sokol L, Greenwood M, Caballero D, Borchmann P, Mor-schhauser F, Wilhelm M, Pinter-Brown L, Padma-nabhan S, Shustov A, Nichols J, Carroll S, Balser J, Balser B, Horwitz S. Results from a pivotal, open-label, phase II study of romidepsin in relap-sed or refractory peripheral T-cell lymphoma after prior systemic therapy. JCO 2012; 30: 631-636.
34) San-Miguel JF, Hungria VTM, Yoon SS, Beksac M, Dimopoulos MA, Elghandour A, Jedrzejczak WW, Günther A, Nakorn TN, Siritanaratkul N, Corradini P, Chuncharunee S, Lee J-J, Schlos-sman RL, Shelekhova T, Kwee Y, Daryl T, Num-benjapon, T Cavenagh JD, Hou J, LeBlanc R, Nahi H, Qiu L, Salwender H, Pulini S, Moreau P, Warzocha K, White D, Bladé J, Chen W, De la Rubia J, Gimsing P, Lonial S, Kaufman JL, Ocio EM, Veskovski L, Sohn SK, Wang MC, Lee JH, Einsele H, Sopala M, Corrado C, Bengoudifa B-R, Binlich F, Richardson PG. Panobinostat plus bor-tezomib and dexamethasone versus placebo plus bortezomib and dexamethasone in patients with relapsed or relapsed and refractory multiple mye-loma: a multicentre, randomised, double-blind phase 3 trial. Lancet Oncol 2014; 15: 1195-1206.
35) O’Connor OA, Horwitz S, Masszi T, Van Hoof A, Brown P, Doorduijn J, Hess G, Jurczak W, Kno-blauch P, Chawla S, Bhat G, Choi MR, Walewski J, Savage K, Foss F, Allen LF, Shustov A. Belinostat in patients with relapsed or refractory peripheral T-cell lymphoma: results of the pivotal phase II BE-LIEF (CLN-19) Study. JCO 2015; 33: 2492-2499.
36) Bubna AK. Vorinostat-An overview. Indian J Der-matol 2015; 60: 419-423.
37) Ogura M, Ando K, Suzuki T, Ishizawa K, Oh SY, Itoh K, Yamamoto K, Au WY, Tien HF, Matsuno Y, Terauchi T, Yamamoto K, Mori M, Tanaka Y, Shimamoto T, Tobinai K, Kim WS. A multicentre phase II study of vorinostat in patients with relap-sed or refractory indolent B‐cell non‐Hodgkin lym-phoma and mantle cell lymphoma. Br J Haematol 2014; 165: 768-776.
38) Kirschbaum MH, Goldman BH, Zain JM, Cook JR, Rimsza LM, Forman SJ, Fisher RI. A phase 2 study of vorinostat for treatment of relapsed or refractory Hodgkin lymphoma: Southwest Oncology Group Study S0517. Leuk Lymphoma 2012; 53: 259-262.
39) Grant C, Rahman F, Piekarz R, Peer C, Frye R, Robey RW, Gardner ER, Figg WD, Bates SE. Ro-midepsin: a new therapy for cutaneous T-cell lym-phoma and a potential therapy for solid tumors. Expert Rev Anticanc Ther 2010; 10: 997-1008.
40) Poole RM. Belinostat: First Global Approval. Dru-gs 2014; 74: 1543-1554.
41) Sawas A, Radeski D, O’Connor OA. Belinostat in patients with refractory or relapsed peripheral T-cell lymphoma: a perspective review. Ther Adv Hematol 2015; 6: 202-208.
42) Reimer P. New developments in the treatment of peripheral T-cell lymphoma - role of Belinostat. Cancer Manag Res 2015; 7: 145-151.
43) Puvvada SD, Li H, Rimsza LM, Bernstein SH, Fi-sher RI, LeBlanc M, Schmelz M, Glinsmann-Gib-son B, Miller TP, Maddox AM, Friedberg JW, Smi-th SM, Persky DO. A phase II study of belinostat (PXD101) in relapsed and refractory aggressive B-cell lymphomas: SWOG S0520. Leuk Lympho-ma 2016; 57: 2359-2369.
44) Eleutherakis-Papaiakovou E, Kanellias N, Kastri-tis E, Gavriatopoulou M, Terpos E, Dimopoulos MA. Efficacy of panobinostat for the treatment of multiple myeloma. J Oncol 2020; 2020: 1-11.
45) Moore D. Panobinostat (Farydak): A novel option for the treatment of relapsed or relapsed and re-fractory multiple myeloma. P T 2016; 41: 296-300.
46) Duvic M, Dummer R, Becker JC, Poulalhon N, Or-tiz Romero P, Grazia Bernengo M, Lebbé C, As-saf C, Squier M, Williams D, Marshood M, Tai F, Prince HM. Panobinostat activity in both bexaro-tene-exposed and -naïve patients with refractory cutaneous T-cell lymphoma: results of a phase II trial. Eur J Cancer 2013; 49: 386-394.
47) Zaja F, Salvi F, Rossi M, Sabattini E, Evangelista A, Ciccone G, Angelucci E, Gaidano G, Zanni M, Ladetto M, Chiappella A, Vitolo U, Zinzani PL, Ca-lifano C, Tucci A, Patti C, Pileri SA, Lenti V, Picca-luga PP, Cavallo F, Volpetti S, Perali G, Assouline S, Mann KK, Morin R, Alcaide M, Bushell K, Fanin R, Levis A. Single-agent panobinostat for relap-sed/refractory diffuse large B-cell lymphoma: cli-nical outcome and correlation with genomic data. A phase 2 study of the Fondazione Italiana Linfo-mi. Leuk Lymphoma 2018; 59: 2904-2910.
48) Batlevi CL, Crump M, Andreadis C, Rizzieri D, As-souline SE, Fox S, Van der Jagt RHC, Copeland A, Potvin D, Chao R, Younes A. A phase 2 study of mocetinostat, a histone deacetylase inhibitor, in relapsed or refractory lymphoma. Br J Haema-tol 2017; 178: 434-441.
49) Younes A, Oki Y, Bociek RG, Kuruvilla J, Fanale M, Neelapu S, Copeland A, Buglio D< Galal A, Besterman J, Li Z, Drouin M, Patterson T, Ward MR, Paulus JK, Ji Y, Medeiros LJ, Martell RE. Mo-cetinostat for relapsed classical Hodgkin’s lym-phoma: an open-label, single-arm, phase 2 trial. Lancet Oncol 2011; 12: 1222-1228.
50) Fournel M, Bonfils C, Hou Y, Yan PT, Trachy-Bour-get MC, Kalita A, Liu J, Lu AH, Zhou NZ, Robert MF, Gillespie J, Wang JJ, Ste-Croix H, Rahil J, Lefebvre S, Moradei O, Delorme D, MacLeod AR, Besterman JM, Li Z. MGCD0103, a novel isotype-selective histone deacetylase inhibitor, has broad spectrum antitumor activity in vitro and in vivo. Mol Cancer Ther 2008; 7: 759-768.

Rationale behind using valproic acid for Non-Hodgkin lymphoma: a biomolecular perspective
7497
51) Guan XW, Wang HQ, Ban WW, Chang Z, Chen HZ, Jia L, Liu FT. Novel HDAC inhibitor chidamide synergizes with rituximab to inhibit diffuse large B-cell lymphoma tumour growth by upregulating CD20. Cell Death Dis 2020; 11: 20.
52) Shi Y, Dong M, Hong X, Zhang W, Feng J, Zhu J, Yu L, Ke X, Huang H, Shen Z, Fan Y, Li W, Zhao X, Qi J, Huang H, Zhou D, Ning Z, Lu X. Resul-ts from a multicenter, open-label, pivotal phase II study of chidamide in relapsed or refractory pe-ripheral T-cell lymphoma. Ann Oncol 2015; 26: 1766-1771.
53) Ribrag V, Kim WS, Bouabdallah R, Lim ST, Coif-fier B, Illes A, Lemieux B, Dyer MJS, Offner F, Fel-loussi Z, Kloos I, Luan Y, Vezan R, Graef T, Mor-schhauser F. Safety and efficacy of abexinostat, a pan-histone deacetylase inhibitor, in non-Hodgkin lymphoma and chronic lymphocytic leukemia: re-sults of a phase II study. Haematologica 2017; 102: 903-909.
54) Child F, Ortiz‐Romero PL, Alvarez R, Bagot M, Stadler R, Weichenthal M, Alves R, Quaglino P, Beylot-Barry M, Cowan R, Geskin LJ, Pérez-Ferr-iols A, Hellemans P, Elsayed Y, Phelps C, For-slund A, Kamida M, Zinzani PL. Phase II multicen-tre trial of oral quisinostat, a histone deacetylase inhibitor, in patients with previously treated stage IB – IVA mycosis fungoides/Sézary syndrome. Br J Dermatol 2016; 175: 80-88.
55) Arts J, King P, Mariën A, Floren W, Beliën A, Jans-sen L, Pilatte I, Roux B, Decrane L, Gilissen R, Hi-ckson I, Vreys V, Cox E, Bol K, Talloen W, Goris I, Andries L, Du Jardin M, Janicot M, Page M, Van Emelen K, Angibaud P. JNJ-26481585, a novel “second-generation” oral histone deacetylase inhi-bitor, shows broad-spectrum preclinical antitumo-ral activity. Clin Cancer Res 2009; 15: 6841-6851.
56) Batlevi CL, Kasamon Y, Bociek RG, Lee P, Gore L, Copeland A, Sorensen R, Ordentlich P, Cruick-shank S, Kunkel L, Buglio D, Hernandez-Ilizalit F, Younes A. ENGAGE- 501: phase II study of entino-stat (SNDX-275) in relapsed and refractory Hodgkin lymphoma. Haematologica 2016; 101: 968-975.
57) Prebet T, Sun Z, Ketterling RP, Zeidan A, Green-berg P, Herman J, Juckett M, Smith MR, Malick L, Paietta E, Czader M, Figueroa M, Gabrilove J, Erba HP, Tallman MS, Litzow M, Gore SD. Azaciti-dine with or without Entinostat for the treatment of therapy-related myeloid neoplasm: further resul-ts of the E1905 North American Leukemia Inter-group study. Br J Haematol 2016; 172: 384-391.
58) Burton BS. On the propyl derivatives and decom-position products of ethylacetoacetate. American Chemical Journal 1882; 3: 385-395
59) Porter RJ, Rogawski MA. Basic & clinical phar-macology. 14th ed. Katzung BG, editor. New York: McGraw Hill Lange, 2018.
60) Ghodke-Puranik Y, Thorn CF, Lamba JK, Leeder JS, Song W, Birnbaum AK, Altman RB, Klein TE. Valproic acid pathway: pharmacokinetics and pharmacodynamics. Pharmacogenet Genom 2013; 23: 236-241.
61) Argikar UA, Remmel RP. Effect of aging on glucu-ronidation of valproic acid in human liver microso-mes and the role of UDP-Glucuronosyltransfera-se UGT1A4, UGT1A8, and UGT1A10. Drug Metab Dispos 2009; 37: 229-236.
62) Tan L, Yu JT, Sun YP, Ou JR, Song JH, Yu Y. The influence of cytochrome oxidase CYP2A6, CYP2B6, and CYP2C9 polymorphisms on the plasma concentrations of valproic acid in epileptic patients. Clinical Neurol Neurosur 2010; 112: 320-323.
63) Damm J, Gordon S, Ehinger M, Jerkeman M, Gullberg U, Hultquist A, Drott K. Pharmacologi-cally relevant doses of valproate upregulate CD20 expression in three diffuse large B-cell lymphoma patients in vivo. Exp Hematol Oncol 2015; 4: 4.
64) Alberts B. Molecular biology of the cell. Sixth edi-tion. New York, NY: Garland Science, Taylor and Francis Group, 2015.
65) Newbold A, Salmon JM, Martin BP, Stanley K, Johnstone RW. The role of p21waf1/cip1 and p27Kip1 in HDACi-mediated tumor cell death and cell cycle arrest in the Eμ-myc model of B-cell lymphoma. Oncogene 2014; 33: 5415-5423.
66) Scotto L, Serrano XJ, Zullo K, Kinahan C, Deng C, Sawas A, Bates S, O’Connor OA. ATM inhibi-tion overcomes resistance to histone deacetylase inhibitor due to p21 induction and cell cycle arrest. Oncotarget 2020; 11: 3432-3442.
67) Cassaday RD, Goy A, Advani S, Chawla P, Na-chankar R, Gandhi M, Gopal AK. A phase II, sin-gle-arm, open-label, multicenter study to evaluate the efficacy and safety of P276-00, a cyclin-de-pendent kinase inhibitor, in patients with relapsed or refractory mantle cell lymphoma. Clin Lympho-ma Myeloma Leuk 2015; 15: 392-397.
68) Müller A, Zang C, Chumduri C, Dörken B, Daniel PT, Scholz CW. Concurrent inhibition of PI3K and mTORC1/mTORC2 overcomes resistance to ra-pamycin induced apoptosis by down-regulation of Mcl-1 in mantle cell lymphoma: PI3K/AKT/mTOR Signaling in Mantle Cell Lymphoma. Int J Cancer 2013; 133: 1813-1824.
69) Zaidi ARS, Dresman S, Burt C, Rule S, McCallum L. Molecular signatures for CCN1, p21 and p27 in progressive mantle cell lymphoma. J Cell Com-mun Signal 2019; 13: 421-434.
70) Zhou Z, Fang Q, Ma D, Zhe N, Ren M, Cheng B, Li P, Liu P, Lin X, Tang S, Hu X, Liao Y, Zhang Y, Lu T, Wang J. Silencing heme oxygenase-1 increa-ses the sensitivity of ABC-DLBCL cells to histone deacetylase inhibitor in vitro and in vivo. Oncotar-get 2017; 8: 78480-78495.
71) Sanaei M, Kavoosi F. Effect of valproic acid on the class I histone deacetylase 1, 2 and 3, tumor suppressor genes p21WAF1/CIP1 and p53, and intrinsic mitochondrial apoptotic pathway, pro- (Bax, Bak, Bim) and anti- (Bcl-2, Bcl-xL, and Mcl-1) apoptotic genes expression, cell viability, and apoptosis induction in hepatocellular carcinoma HepG2 cell line. Asian Pac J Cancer Prev 2021; 22: 89-95.

H. Ridwansyah, I. Wijaya, M.H. Bashari, A.H.S. Kartamihardja, B.S. Hernowo
7498
72) Sanaei M, Kavoosi F. Effect of 5-aza-2’-deoxycyti-dine in comparison to valproic acid and trichosta-tin a on histone deacetylase 1, dna methyltran-sferase 1, and cip/kip family (p21, p27, and p57) genes expression, cell growth inhibition, and apoptosis induction in colon cancer sw480 cell line. Adv Biomed Res 2019; 8: 52.
73) Zhang Z, Hao C, Wang L, Liu P, Zhao L, Zhu C, Tian X. Inhibition of leukemic cells by valproic acid, an HDAC inhibitor, in xenograft tumors. On-coTargets Ther 2013; 6: 733-740.
74) de Miguel D, Lemke J, Anel A, Walczak H, Marti-nez-Lostao L. Onto better TRAILs for cancer tre-atment. Cell death differ 2016; 23: 733-747.
75) Lemke J, Karstedt SV, Zinngrebe J, Walczak H. Getting TRAIL back on track for cancer therapy. Cell Death Differ 2014; 21: 1350-1364.
76) Staniek J, Lorenzetti R, Heller B, Janowska I, Sch-neider P, Unger S, Warnatz K, Seidl M, Venhoff N, Thiel J, Smulski CR, Rizzi M. TRAIL-R1 and TRAIL-R2 mediate TRAIL-dependent apoptosis in activated primary human B lymphocytes. Front Immunol 2019; 10: 951.
77) Daniel D, Yang B, Lawrence DA, Totpal K, Balter I, Lee WP, Gogineni A, Cole MJ, Yee SF, Ross S, Ashkenazi A. Cooperation of the proapoptotic receptor agonist rhApo2L/TRAIL with the CD20 antibody rituximab against non-Hodgkin lympho-ma xenografts. Blood 2007; 110: 4037-4046.
78) Cillessen SAGM, Meijer CJLM, Ossenkoppele GJ, Castricum KCM, Westra AH, Niesten P, Muris JJF, Nijdam HF, van der Hem KG, Flens M, Hooijberg E, Oudejans JJ. Human soluble TRAIL/Apo2L indu-ces apoptosis in a subpopulation of chemotherapy refractory nodal diffuse large B-cell lymphomas, determined by a highly sensitive in vitro apoptosis assay. Br J Haematol 2006; 134: 283-293.
79) Markovic O, Marisavljevic D, Cemerikic-Martino-vic V, Filipovic B, Radovanović S, Zdravković M, Stanisavljevic D, Mihaljevic B. c-FLIP does not correlate with response to immunochemotherapy treatment and outcome of patients with nodal diffuse large B-cell lymphoma. Biomed Pharma-cother 2013; 67: 445-449.
80) Goldar S, Khaniani MS, Derakhshan SM, Bara-daran B. Molecular mechanisms of apoptosis and roles in cancer development and treatment. Asian Pac J Cancer Prev 2015; 16: 2129-2144.
81) Kretz AL, Trauzold A, Hillenbrand A, Knippschild U, Henne-Bruns D, von Karstedt S, Lemke J. TRAILblazing strategies for cancer treatment. Cancers 2019; 11: 456.
82) Shirley S, Micheau O. Targeting c-FLIP in cancer. Cancer Lett 2013; 332: 141-150
83) Lee BS, Kim YS, Kim HJ, Kim DH, Won HR, Kim YS, Kim CH. HDAC4 degradation by combined TRAIL and valproic acid treatment induces apop-totic cell death of TRAIL-resistant head and neck cancer cells. Sci Rep 2018; 8: 12520.
84) Bangert A, Cristofanon S, Eckhardt I, Abhari BA, Kolodziej S, Häcker S, Vellanki SHK, Lausen J,
Debatin K-M, Fulda S. Histone deacetylase inhibi-tors sensitize glioblastoma cells to TRAIL-induced apoptosis by c-myc-mediated downregulation of cFLIP. Oncogene 2012; 31: 4677-4688.
85) Harrison DA. The JAK/STAT pathway. Cold Spring Harb Perspect Biol 2012; 4: a011205.
86) Sun R-F, Yu Q-Q, Young KH. Critically dysregu-lated signaling pathways and clinical utility of the pathway biomarkers in lymphoid malignancies. Chronic Dis Transl Med 2018; 4: 29-44.
87) Camicia R, Winkler HC, Hassa PO. Novel drug targets for personalized precision medicine in relapsed/refractory diffuse large B-cell lympho-ma: a comprehensive review. Mol Cancer 2015; 14: 207.
88) Mellert K, Martin M, Lennerz JK, Lüdeke M, Stai-ger AM, Kreuz M, Löffler M, Schmitz N, Trümper L, Feller AC, Hartmann S, Hansmann M-L, Klap-per W, Stein H, Rosenwald A, Ott G, Ziepert M, Möller P. The impact of SOCS1 mutations in dif-fuse large B‐cell lymphoma. Br J Haematol 2019; 187: 627-637.
89) Ilangumaran S, Rottapel R. Regulation of cytoki-ne receptor signaling by SOCS1. Immunol Rev 2003; 192: 196-211.
90) Durham GA, Williams JJL, Nasim MT, Palmer TM. Targeting SOCS Proteins to Control JAK-STAT Si-gnalling in Disease. Trends Pharmacol Sci 2019; 40: 298-308.
91) Weniger M, Melzner I, Menz C, Wegener S, Bucur A, Dorsch K, Mattfeldt T, Barth TFE. Mutations of the tumor suppressor gene SOCS-1 in classical Hodgkin lymphoma are frequent and associated with nuclear phospho-STAT5 accumulation. On-cogene 2006; 25: 2679-2684.
92) Brender C, Lovato P, Sommer VH, Woetmann A, Mathiesen A-M, Geisler C, Wasik M, Ødum N. Constitutive SOCS-3 expression protects T-cell lymphoma against growth inhibition by IFNα. Leu-kemia 2005; 19: 209-213.
93) Kim M-H, Kim M-S, Kim W, Kang MA, Cacalano NA, Kang S-B, Shin YJ, Jeong J-H, Scott RJ. Sup-pressor of Cytokine Signaling (SOCS) Genes Are Silenced by DNA Hypermethylation and Histone Deacetylation and Regulate Response to Ra-diotherapy in Cervical Cancer Cells. PLoS One 2015; 10: e0123133.
94) Sanaei M, Kavoosi F, Behjoo H. Effect of valproic acid and zebularine on SOCS-1 and SOCS-3 gene expression in colon carcinoma SW48 cell line. Exp Oncology 2020; 42: 183-187.
95) Zhu S, Denman CJ, Lee DA. Valproic acid selecti-vely inhibits STAT3 phosphorylation. Blood 2009; 22: 1720.
96) Cotto M, Cabanillas F, Tirado M, García MV, Pa-checo E. Epigenetic therapy of lymphoma using histone deacetylase inhibitors. Clin Transl Oncol 2010; 12: 401-409.
97) Ni L, Wang L, Yao C, Ni Z, Liu F, Gong C, Zhu X, Yan X, Watowich SS, Lee DA, Zhu S. The hi-stone deacetylase inhibitor valproic acid inhibits

Rationale behind using valproic acid for Non-Hodgkin lymphoma: a biomolecular perspective
7499
NKG2D expression in natural killer cells through suppression of STAT3 and HDAC3. Sci Rep 2017; 7: 45266.
98) Westin JR. Status of PI3K/Akt/mTOR pathway inhibitors in lymphoma. Cl Lymph Myelom Leuk 2014; 14: 335-342.
99) Xia P, Xu X-Y. PI3K/Akt/mTOR signaling pathway in cancer stem cells: from basic research to clini-cal application. Am J Cancer Res 2015; 5: 1602-1609.
100) Murugan AK. mTOR: Role in cancer, metastasis and drug resistance. Semin Cancer Biol 2019; 59: 92-111.
101) Wang X, Cao X, Sun R, Tang C, Tzankov A, Zhang J, Manyam GC, Xiao M, Miao Y, Jabbar K, Tan X, Pang Y, Visco C, Xie Y, Dybkaer K, Chiu A, Ora-zi A, Zu Y, Bhagat G, Richards KL, Hsi ED, Choi WWL, van Krieken JH, Huh J, Ponzoni M, Ferreri AJM, Møller MB, Parsons BM, Winter JN, Piris MA, Li S, Miranda RN, Medeiros LJ, Li Y, Xu-Mo-nette ZY, Young KH. Clinical significance of PTEN deletion, mutation, and loss of PTEN expression in de novo diffuse large b\B-cell lymphoma. Neo-plasia 2018; 20: 574-593.
102) Wang J, Xu-Monette ZY, Jabbar KJ, Shen Q, Manyam GC, Tzankov A, Visco C, Wang J, Mon-tes-Moreno S, Dybkær K, Tam W, Bhagat G, Hsi ED, van Krieken JH, Ponzoni M, Ferreri AJM, Wang S, Møller MB, Piris MA, Medeiros LJ, Li Y, Pham LV, Young KH. AKT hyperactivation and the potential of AKT-targeted therapy in diffuse large B-cell lympho-ma. Am J Pathol 2017; 187: 1700-1716.
103) Sun J, Piao J, Li N, Yang Y, Kim K, Lin Z. Val-proic acid targets HDAC1/2 and HDAC1/PTEN/Akt signalling to inhibit cell proliferation via the induction of autophagy in gastric cancer. FEBS J 2020; 287: 2118-2133.
104) Xia Q, Zheng Y, Jiang W, Huang Z, Wang M, Ro-driguez R, Jin X. Valproic acid induces autophagy by suppressing the Akt/mTOR pathway in human prostate cancer cells. Oncol Lett 2016; 12: 1826-1832.
105) Cardenas MG, Oswald E, Yu W, Xue F, MackKe-rell AD Jr, Melnick AM. The expanding role of the BCL6 oncoprotein as a cancer therapeutic target. Clin Cancer Res 2017; 23: 885-893.
106) Kim EM, Jung CH, Kim J, Hwang SG, Park JK, Um HD. The p53/p21 complex regulates cancer cell invasion and apoptosis by targeting Bcl-2 fa-mily proteins. Cancer Res 2017; 77: 3092-3100.
107) Nagel D, Vincendeau M, Eitelhuber AC, Krapp-mann D. Mechanisms and consequences of con-stitutive NF-κB activation in B-cell lymphoid mali-gnancies. Oncogene 2014; 33: 5655-5665.
108) Davis RE, Brown KD, Siebenlist U, Staudt LM. Con-stitutive nuclear factor κB activity is required for survival of activated B-cell like diffuse large B cell lymphoma cells. J Exp Med 2001; 194: 1861-1874.
109) Kennedy R, Klein U. Aberrant Activation of NF-κB Signalling in Aggressive Lymphoid Malignancies. Cell 2018; 7: 189.
110) Schwertheim S, Worm K, Schmid K, Sheu-Gra-bellus S-Y. Valproic acid downregulates NF-κB p50 activity and IRAK-1 in a progressive thyroid carcinoma cell line. Horm Metab Res 2014; 46: 181-186.
111) Liu Z, Jing Q, Wang Y, Li Y, Mi F, Xiang C, Fu R. The short-term effect of histone deacetylase inhi-bitors, chidamide and valproic acid, on the NF-κB pathway in multiple myeloma cells. Int J Mol Med 2018; 43: 285-293.
112) Bonavida B. Postulated mechanisms of resistan-ce of B-NHL to rituximab treatment regimes: stra-tegies to overcome resistance. Crit Rev Oncog 2014; 41: 667-677.
113) Tomita A. Genetic and epigenetic modulation of CD20 expression in B-cell malignancies: mole-cular mechanisms and significance to rituximab resistance. JCEH 2016; 56: 89-99.
114) Nakamaki T, Fukuchi K, Nakashima H, Ariizumi H, Maeda T, Saito B, Yanagisawa K, Tomoyasu S, Homma M, Shiozawa E, Ota H. CD20 gene dele-tion causes a CD20 negative relapse in diffuse lar-ge B cell lymphoma. Eur J Haematol 2012; 89: 6.
115) Pozzo F, Bittolo T, Arruga F, Bulian P, Macor P, Tissino E, Gizdic B, Rossi FM, Bomben R, Zuc-chetto A, Benedetti D, Degan M, D’Arena G, Chia-renza A, Zaja F, Pozzato G, Rossi D, Gaidano G, Del Poeta G, Deaglio S, Gattei V, Dal Bo M. NOTCH1 mutations associate with low CD20 level in chronic lymphocytic leukemia: evidence for a NOTCH1 mutation-driven epigenetic dysregula-tion. Leukemia 2016; 30: 182-189.
116) Pavlasova G, Mraz M. The regulation and fun-ction of CD20: an “enigma” of B-cell biology and targeted therapy. Haematologica 2020; 105: 1494-1506.
117) Sugimoto T, Tomita A, Hiraga J, Shimada K, Kiyoi H, Kinoshita T, Naoe T. Escape mechanisms from antibody therapy to lymphoma cells: Downregula-tion of CD20 mRNA by recruitment of the HDAC complex and not by DNA methylation. Biochem Bioph Res Co 2009; 390: 48-53.
118) Shimizu R, Kikuchi J, Wada T, Ozawa K, Kano Y, Furukawa Y. HDAC inhibitors augment cyto-toxic activity of rituximab by upregulating CD20 expression on lymphoma cells. Leukemia 2010; 24: 1760-1768.
119) Scialdone A, Hasni MS, Damm JK, Lennartsson A, Gullberg U, Drott K. The HDAC inhibitor valpro-ate induces a bivalent status of the CD20 promo-ter in CLL patients suggesting distinct epigenetic regulation of CD20 expression in CLL in vivo. On-cotarget 2017; 8: 37409-37422.
120) Xie Z, Ago Y, Okada N, Tachibana M. Valproic acid attenuates immunosuppressive function of myeloid-derived suppressor cells. J Pharmacol Sci 2018; 137: 359-365.
121) Tiper IV, Webb TJ. Histone deacetylase inhibitors enhance CD1d-dependent NKT cell responses to lymphoma. Cancer Immunol Immunother 2016; 65: 1411-1421.

H. Ridwansyah, I. Wijaya, M.H. Bashari, A.H.S. Kartamihardja, B.S. Hernowo
7500
122) Wang X, Waschke BC, Woolaver RA, Chen SMY, Chen Z, Wang JH. HDAC inhibitors overcome im-munotherapy resistance in B-cell lymphoma. Pro-tein Cell 2020; 11: 472-482.
123) Cycon KA, Mulvaney K, Rimsza LM, Persky D, Murphy SP. Histone deacetylase inhibitors activa-te CIITA and MHC class II antigen expression in diffuse large B-cell lymphoma. Immunology 2013; 140: 259-272.
124) Zain J, Rotter A, Weiss L, Forman S, Kirschbaum MH. Valproic acid monotherapy leads to CR in a patient with refractory diffuse large B cell lympho-ma. Leuk Lymphoma 2007; 48: 1216-1218.
125) Munster P, Marchion D, Bicaku E, Lacevic M, Kim J, Centeno B, Daud A, Neuger A, Minton S, Sul-livan D. Clinical and Biological Effects of Valproic Acid as a Histone Deacetylase Inhibitor on Tumor and Surrogate Tissues: Phase I/II Trial of Valproic acid and Epirubicin/FEC. Clin Cancer Res 2009; 15: 2488-2496.
126) Espinoza-Zamora JR, Labardini-Méndez J, Sosa-E-spinoza A, López-González C, Vieyra-García M, Candelaria M, Lozano-Zavaleta V, Toledano-Cue-vas D, Zapata-Canto N, Cervera E, Dueñas-Gonz-
áles A. Efficacy of hydralazine and valproate in cu-taneous T-cell lymphoma, a phase II study. Expert Opin Investig Drugs 2017; 26: 481-487.
127) Wang L, Li LR. R-CHOP resistance in diffuse large B-cell lymphoma: biological and molecular mechanisms. Chin Med J 2021; 134: 253-260.
128) Spitzner M, Ebner R, Wolff HA, Ghadimi BM, Wienands J, Grade M. STAT3: A Novel Molecular Mediator of Resistance to Chemoradiotherapy. Cancers 2014; 26: 1986-2011.
129) Otero DC, Poli V, David M, Rickert RC. Cutting edge: inherent and acquired resistance to radia-tion-induced apoptosis in B cells: a pivotal role for STAT3. J Immunol 2006; 177: 6593-6597.
130) Zou L, Song G, Gu S, Kong L, Sun S, Yang L, Cho WC. Mechanism and treatment of rituximab resistance in diffuse large B-cell lymphoma. Curr Cancer Drug Targets 2019; 19: 681-687.
131) Wang X, Waschke BC, Woolaver RA, Chen Z, Zhang G, Piscopio AD, Liu X, Wang JH. Histo-ne-deacetylase inhibition sensitizes PD1 blocka-de-resistant B-cell lymphomas. Cancer Immunol Res 2019; 7: 1318-1331.