poly(l-lactide) microfilaments enhance peripheral nerve regeneration across extended nerve lesions
TRANSCRIPT

Poly(L-Lactide) Microfilaments EnhancePeripheral Nerve Regeneration AcrossExtended Nerve Lesions
Teri-T B. Ngo,1 Paula J. Waggoner,2 Andres A. Romero,2 Kevin D. Nelson,2
Robert C. Eberhart,2 and George M. Smith1*1Department of Physiology, Spinal Cord and Brain Injury Research Center, University of Kentucky, Albert B.Chandler Medical Center, Lexington, Kentucky2Joint Program in Biomedical Engineering, University of Texas Southwestern Medical Center, Dallas, Texas,and University of Texas at Arlington, Arlington, Texas
After injury, axonal regeneration occurs across shortgaps in the peripheral nervous system, but regenerationacross larger gaps remains a challenge. To improve re-generation across extended nerve defects, we have fab-ricated novel microfilaments with the capability for drugrelease to support cellular migration and guide axonalgrowth across a lesion. In this study, we examine thenerve repair parameters of non-loaded filaments. To ex-amine the influence of packing density on nerve repair,wet-spun poly(L-Lactide) (PLLA) microfilaments werebundled at densities of 3.75, 7.5, 15, and 30% to bridgea 1.0-cm gap lesion in the rat sciatic nerve. After 10weeks, nerve cable formation increased significantly inthe filament bundled groups when compared to empty-tube controls. At lower packing densities, the number ofmyelinated axons was more than twice that of controls orthe highest packing density. In a consecutive experi-ment, PLLA bundles with lower filament-packing densitywere examined for nerve repair across 1.4- and 1.8-cmgaps. After 10 weeks, the number of successful regen-erated nerves receiving filaments was more than twicethat of controls. In addition, nerve cable areas for controlgroups were significantly less than those observed forfilament groups. Axonal growth across 1.4- and 1.8-cmgaps was more consistent for the filament groups thanfor controls. These initial results demonstrate that PLLAmicrofilaments enhance nerve repair and regenerationacross large nerve defects, even in the absence of drugrelease. Ongoing studies are examining nerve regenera-tion using microfilaments designed to release neurotro-phins or cyclic AMP. © 2003 Wiley-Liss, Inc.
Key words: nerve guidance channels; entubulation;PLLA; bioresorbable polymers
Axonal regeneration can occur after peripheral ner-vous system (PNS) injury when the injury gap is short andthe nerve ends can be anastomosed. When the gap is toolong and anastomosis of the nerve ends is difficult, axonalregeneration and eventual innervation of the target organis unlikely. Under these conditions, several repair materialshave been proposed to increase the prospects of nerve
regeneration, including autologous nerve grafts (Lundborget al., 1982a; Hafiz et al., 1985; Evans et al., 1995),allografts (Dubuisson, 1997), xenografts, muscles (Norriset al., 1988; Gattuso et al., 1989; DeFranzo et al., 1994),and blood vessels (Chiu et al., 1982; Foidart-Dessalle et al.,1997). Although these materials enhance regeneration,there are numerous disadvantages, such as limited avail-ability, a requisite additional surgical step, size mismatch-ing (Rodriguez et al., 1999), a need for immunosuppres-sion (allografts and xenografts) (Bain et al., 1988), and aloss of nerve function at the donor site (autologous grafts).
An alternative to using natural components is toemploy entubulation techniques to bridge the nerve gap.Synthetic as well as natural materials have been shaped intotubes such that the two nerve stumps could be insertedinto the ends of the tube, thereby allowing the nerve toregenerate across a well-defined gap. These tubes, typicallyreferred to as nerve guides or nerve guidance channels, actto sequester chemotrophic and chemotactic moleculeswhile reducing the formation of scar tissue around thereforming nerve. Tubular nerve guides have been con-structed from a variety of materials, including siliconerubber (Lundborg et al., 1982b; Williams et al., 1983;Gibson and Daniloff, 1989; Danielsen et al., 1993; Dahlinet al., 1995), collagen (Archibald et al., 1991; Li et al,1992; Chamberlain et al., 1998), and bioresorbable poly-mer tubes made of poly(L-lactide) acid (PLLA) (Seckel etal., 1984; da Silva et al., 1985), poly(glycolic acid; PGA)(Dellon and Mackinnon, 1988; Mackinnon and Dellon,1990; Keeley et al., 1991), polyglactin (Molander et al.,1983), or blends of these components (den Dunnen et al.,1995, 1996, 1997; Aldini et al., 1996; Rodriguez et al.,1999). The latter group has the advantage of degrading
Contract grant sponsor: NINDS; Contract grant number: R01NS40592.
*Correspondence to: Dr. George M. Smith, Department of Physiology MS508, University of Kentucky, Albert B. Chandler Medical Center, Lexing-ton, KY 40536-0298. E-mail: [email protected]
Received 29 August 2002; Revised 27 November 2002; Accepted 10December 2002
Journal of Neuroscience Research 72:227–238 (2003)
© 2003 Wiley-Liss, Inc.

over time in vivo (Leenslag et al., 1987; Pistner et al.,1993), thus avoiding the need for a second surgery toremove scaffolding material. In most instances, nerve re-generation within hollow tubes is limited to approxi-mately 1-cm gaps in the rat sciatic nerve (Lundborg et al.,1982b; Seckel et al., 1984).
To expand the regenerative potential of the entubu-lation method, tubes have been preloaded with fibrin(Williams and Varon, 1985; Williams, 1987), trophic fac-tors (Rich et al., 1989), Schwann cells (Guenard et al.,1992; Levi et al., 1997), and extracellular matrix mole-cules, such as collagen (Wells et al., 1997; Labrador et al.,1998), fibronectin (Bailey et al., 1993), and laminin (Mad-ison et al., 1985, 1988; Tong et al., 1994). It is thoughtthat these materials provide a substrate that enhances mi-gration of nonneuronal cells (e.g., fibroblasts, endothelialcells, and Schwann cells), which act to form a cell- andmatrix-containing cable between the separated stumps(Labrador et al., 1998). The formation of this cellular cableprovides the bridging material for axons to extend acrossthe gap. Potentially, axonal regeneration could be furtherenhanced if the bridging structure was designed to directand organize the longitudinal formation of the nerve ca-ble. Lundborg et al. (1997) has demonstrated that bridgingfilaments made from resorbable suture greatly enhancesaxonal regeneration across 10- to 15-mm nerve gaps. Wehave examined the regenerative potential of novel resorb-able synthetic microfilaments that can be fabricated fordrug release. Release of neurotrophins, growth factor, orcyclic nucleotides could enhance further regeneration andsegregate axonal growth to more appropriate target loca-tions to increase functional reconnections, which is aconsistent problem with this type of nerve repair.
To increase the guidance environment in the tube,we have fabricated bioresorbable PLLA microfilaments.The fabrication process produces a continuous filament orthread, 40–100 �m in diameter that can be preloadedwith up to 60% aqueous phase by volume. Proteins in theaqueous phase can be loaded into filaments for slow releaseas the filaments degrade. Previous studies in our laboratoryhave shown that these microfilaments can be coated withlaminin and provide an improved substratum to organizeSchwann cell migration and direct axonal growth in vitro(Rangappa et al., 2000). We examined whether the mi-crofilament bundles alone could enhance nerve cable for-mation and provide improved axonal growth across vari-ous gap lengths. The results demonstrate that bundledfilaments enhance regeneration across 1.0-, 1.4-, and1.8-cm gaps in the rat sciatic nerve model. Nerve cableformation and axonal growth were more consistent acrossthese gaps with bundled filaments when compared tocontrols.
MATERIALS AND METHODS
PLLA Filament Preparation
Filament fabrication. Poly(L-lactic acid) (200 kD;Polysciences Inc., Warrington, PA) was dissolved in chloroform(Aldrich Chemical, St. Louis, MO) at a concentration of0.1 g/ml of chloroform and the mixed solution loaded into a
glass syringe (gas-tight syringes; Hamilton Co., Reno, NV) andplaced in a syringe pump (model KDS200; KD Scientific, NewHope, PA). A Viton tube (Cole-Parmer, Vernon Hills, IL)connected the syringe to a 25-gauge needle dispenser by stainlesssteel Luer fittings. We used only blunt-tipped needles (SmallParts Inc., Miami Lakes, FL), as the bevel cut on sharp needlespotentially could cause problems during extrusion.
The extrusion setup consisted of a glass tube, 25 mm indiameter by approximately 200 mm in length, immersed in asmall plastic container full of the coagulating bath fluid. A rubberseptum was placed on top of the glass tube, and the tube wasfilled completely with the coagulating bath by withdrawing airwith a separate syringe. Once the glass tube was completely full(the bottom still immersed in the fluid in the container), theblunt-tipped needle was then pierced through the rubber sep-tum and the syringe pump turned on to deliver 0.03 ml/min.The coagulation bath was a poor solvent for the polymer, yethighly miscible with the solvent used to dissolve the polymer.For the fibers used in this study, we used isopropyl alcohol as thecoagulation bath fluid. When the fiber exits the coagulating bathit was wound on a 8.25-cm diameter bobbin attached to thejaws of a modified 5-inch Garden Lathe (Sears-Craftsman model549-289000; Sears and Roebuck, Chicago, IL). A 24-V powersupply (model 5005R; Power Designs Inc., NY) drove a 0.03-horsepower DC motor (Pittman Motors, Harleysville, Pennsyl-vania) that replaced the original motor on the lathe. We firstwrapped the bobbin with paper so that at the completion of theextrusion run, we pulled the paper from the bobbin to removethe fiber intact. The angular velocity of the lathe was measuredusing an optical tachometer. The draw ratio was calculated as theratio of the linear velocity of the fiber measured at the take-upbobbin to the calculated mean linear flow rate of the emulsionwithin the dispensing needle, because die-swell is not observedtypically during these extrusions. At the end of the extrusionrun, fibers were removed from the lathe, left on the paper-roll,and placed under vacuum at roughly 4°C for at least 4 hr atroom temperature to help remove remaining solvents. The fiberwas stored in a desiccator or in a �20°C freezer until needed.
PLLA filament bundling and sterilization. PLLAfilament bundles were created by inserting PLLA fibers throughsterilized silicone tubes (A-M Systems, Carlsborg, WA; 1.4-,1.8-, and 2.2-cm lengths with an outside diameter of 2.5 mmand inside diameter of 1.5 mm). They were washed three timesin 1� phosphate buffered saline (PBS). Filaments were trimmedto 1.0, 1.4, and 1.8 cm and sterilized by 10� penicillin/streptomycin (Gibco, Carlsbad, CA) in PBS for 2 hr at 25°Cwith constant shaking. Filament bundles were rinsed and washedthree times in PBS, and then stored in PBS at 4°C until use.
Experimental Groups
In the first set of experiments, PLLA filament bundles ofvarious filament-packing densities were tested to bridge a 1.0-cmgap lesion. Four groups (n � 50) of animals were studied. Thecontrol group (n � 17) was implanted with silicone tubes withoutfilaments. The experimental groups had implanted silicone tubesthat were bundled as follows: 16–24 filaments (3.75% packingdensity, n � 7), 32–40 (7.5%, n � 12), 64–80 (15%, n � 7), and128–160 (30%, n � 7). The filament-packing density was deter-mined by comparing the cumulative cross-sectional area of thefilaments to the available area of the silicone tube.
228 Ngo et al.

A second set of experiments was designed to test whetherPLLA filament bundles could enhance regeneration across largergap lesions. Experimental groups were implanted with bundledPLLA filaments (7.5% filament-packing density) loaded intosilicone tubes to bridge 1.4-cm (n � 8) and 1.8-cm (n � 8) gaps.Similarly, control groups were implanted with silicone tubeswithout filaments to bridge 1.4-cm (n � 8) and 1.8-cm (n � 8)gaps.
Surgical Procedure
Adult female Sprague-Dawley rats (250–300 g) wereanesthetized by intraperitoneal injection of ketamine (66 mg/ml)/xylazine (6.66 mg/ml) at 0.1 ml/100 g. A 3-cm incision wasmade parallel to the right femur. After blunt dissection to exposethe sciatic nerve, a 2- to 5-mm section of the nerve was resected.PLLA filament bundles were then implanted to bridge either a1.0-, 1.4-, or 1.8-cm gap between the proximal and distal nervestumps. Superglue (Quicktite Loctite) was applied carefully toboth ends of the tube to secure the implant to both nervestumps. All tubes were filled with PBS upon deployment. Mus-cles were sutured with 4-0 coated VICRYL absorbable suture(Ethicon Inc., Piscataway, NJ), and stainless steel wound clips(Autoclip) were used to staple the skin together. Animals weremonitored and maintained for 10 weeks with full access to foodand water according to the animal care policy at the Universityof Kentucky Medical Center Division of Laboratory AnimalResources. Animals were monitored for any signs of infectionand paw biting of the right limb. All animals recovered success-fully from surgery. During the 10-week survival period, a fewanimals in each set of experiments developed mild autotomy.This included biting of the toenails or phalanges of the righthind paw. These animals were not excluded from the studybecause the injuries were not severe and they healed shortly afterElizabethan collars were applied and hind paws were treated.
Implant Retrieval and Histological Preparations
Animals were anesthetized using sodium pentobarbital(Nembutal, 0.1 ml/100 g), perfused transcardially with 0.9%sodium chloride and 0.5% sodium nitrite, and fixed with 2%paraformaldehyde and 0.5% glutaraldehyde in 0.1 M phosphatebuffer (PB), pH 7.5. Implants were extracted and incubated inthe same fixative for approximately 48 hr. Silicone tubes werethen removed by longitudinal dissection to expose the regener-ated nerves and filament bundles. Implants were washed withPB and prepared for microscopy studies.
Upon extraction of the implants after a 10-week implan-tation period, fibrous tissues ensheathed the outside of thesilicone tubes. The sheaths were observed on all silicone tubes inevery experiment. These tissues were removed along with thesilicone tubes before histological preparations of the regenerat-ing nerves that were encased in the tubes. Upon removal of thetissue sheath, fluid (clear to yellow in color) was seen insideempty and filament-filled silicone tubes. This fluid was drainedas the silicone tubes were removed.
Specimen segments for light microscopy were incubated3–4 hr in 1% osmium tetroxide (Electron Microscopy Sciences,Fort Washington, PA). After PB wash and sequential alcoholdehydration, specimens were immersed 2 hr in propylene oxide(Electron Microscopy Sciences), then incubated overnight in a1:1 mixture of propylene oxide and Spur resin (Polysciences).
Implants were embedded finally in 100% Spur resin and incu-bated overnight at 60°C.
Microscopy Studies
Semi-thin sections for light microscopy studies were cutwith an ultramicrotome (Reichert-Jung) at a thickness of 0.6�m and stained in 1% toluene blue in 1% sodium borate. Forquantitation of axons, myelinated axons within the entire crosssection were counted. This was done because axon density wasnot uniform throughout the section. Composite images for eachsection were taken at 100–400� magnification using the NikonE800 light microscope Metamorph Imaging System 4.0 (Uni-versal Imaging, Downington, PA) and transferred to AdobePhotoshop 5.5 software to generate a montage of the entire crosssection. In Photoshop, a transparent layer with a grid pattern wassuperimposed on top of the image and the myelinated axonswere noted by placing a dot on this layer. This process wasrepeated until all axons were confirmed on the image. Myelin-ated axons were seen as black ovoid rings. To differentiatemyelinated axons from other organelles they were confirmedunder 400� magnification. The layer with the “dotted axons”was transferred to Metamorph to count the number of dots(axons) automatically. Cable areas were measured for each sam-ple using Metamorph Imaging System in pixels2 and convertedto mm2 units. Cable areas were measured at the distal end for the1.0-cm gap, at midsection and distal ends for the 1.4-cm gap,and at 2-mm intervals along the 1.8-cm gap. Note that animalshaving no nerve cables or having cables with no axons at thedistal end were considered in the analysis as having zero numberof axons.
Ultra-thin sections for transmission electron microscopy(TEM) analysis were cut at a thickness of 80–90 nm and stainedin uranyl acetate and lead citrate. TEM studies were carried outusing the Philips Tecnai transmission electron microscope at80 kV. Images were captured at 500–30,000� magnification.
Statistical Analysis
Data were analyzed using the SPSS 10.1 statistical software(SPSS, Inc.). Fisher’s exact tests were carried out to determinedifferences in success rates of nerve cable formation and axonalgrowth between control and test groups. One-way ANOVAwas used to evaluate differences in nerve cable areas and my-elinated axon counts for various filament-packing densities mea-sured at the distal end. Dunnett’s and Tukey HSD post-hocanalyses were used to determine differences within treatmentgroups. One-way ANOVA with repeated measures was used todetermine differences in nerve cable areas and myelinated axoncounts for various gap distances along the nerve cables of the1.4- and 1.8-cm gap groups. The Bonferroni-Holmes methodwas used to correct for the possibilities of errors inherent inmultiple test comparisons. P � 0.05 was the statistical signifi-cance criterion.
RESULTS
Filament-Packing Densities Across 1.0-cm GapThe ability of microfilaments to organize cellular
cable formation and increase axonal regeneration was ex-amined initially across a 1.0-cm sciatic nerve gap. Ourpreliminary studies showed that PLLA filaments could
Bioresorbable Filaments Promote Nerve Repair 229
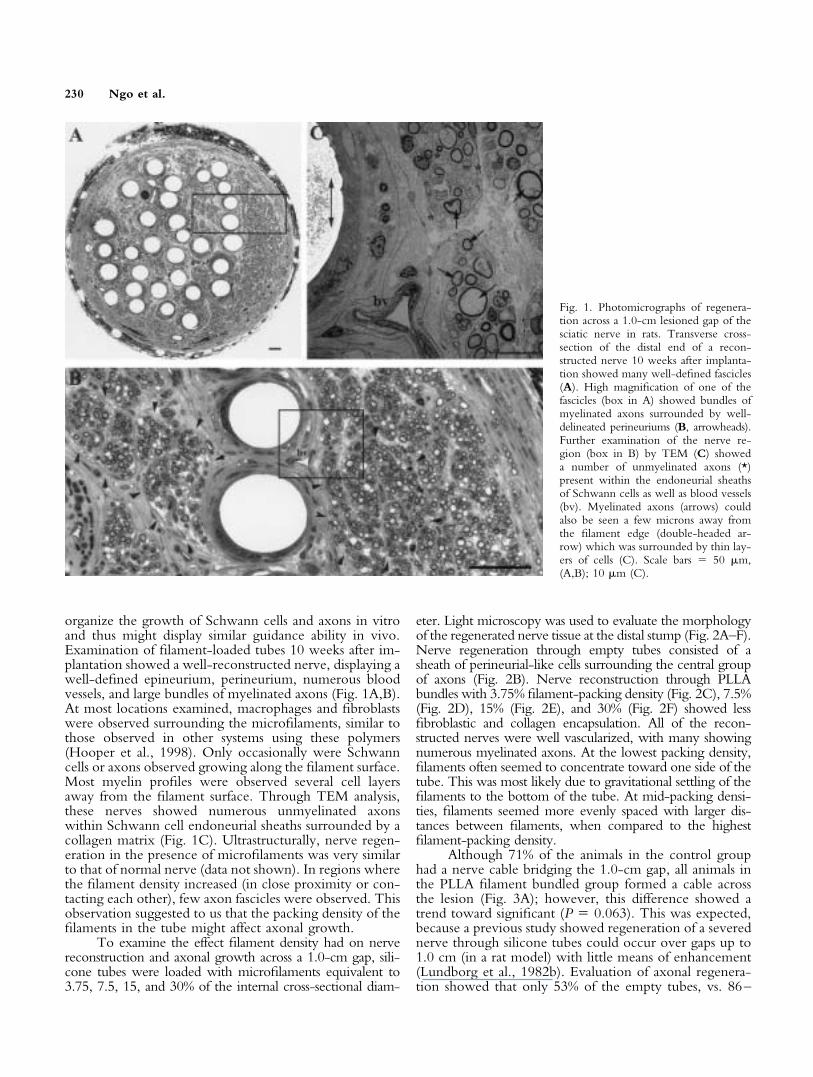
organize the growth of Schwann cells and axons in vitroand thus might display similar guidance ability in vivo.Examination of filament-loaded tubes 10 weeks after im-plantation showed a well-reconstructed nerve, displaying awell-defined epineurium, perineurium, numerous bloodvessels, and large bundles of myelinated axons (Fig. 1A,B).At most locations examined, macrophages and fibroblastswere observed surrounding the microfilaments, similar tothose observed in other systems using these polymers(Hooper et al., 1998). Only occasionally were Schwanncells or axons observed growing along the filament surface.Most myelin profiles were observed several cell layersaway from the filament surface. Through TEM analysis,these nerves showed numerous unmyelinated axonswithin Schwann cell endoneurial sheaths surrounded by acollagen matrix (Fig. 1C). Ultrastructurally, nerve regen-eration in the presence of microfilaments was very similarto that of normal nerve (data not shown). In regions wherethe filament density increased (in close proximity or con-tacting each other), few axon fascicles were observed. Thisobservation suggested to us that the packing density of thefilaments in the tube might affect axonal growth.
To examine the effect filament density had on nervereconstruction and axonal growth across a 1.0-cm gap, sili-cone tubes were loaded with microfilaments equivalent to3.75, 7.5, 15, and 30% of the internal cross-sectional diam-
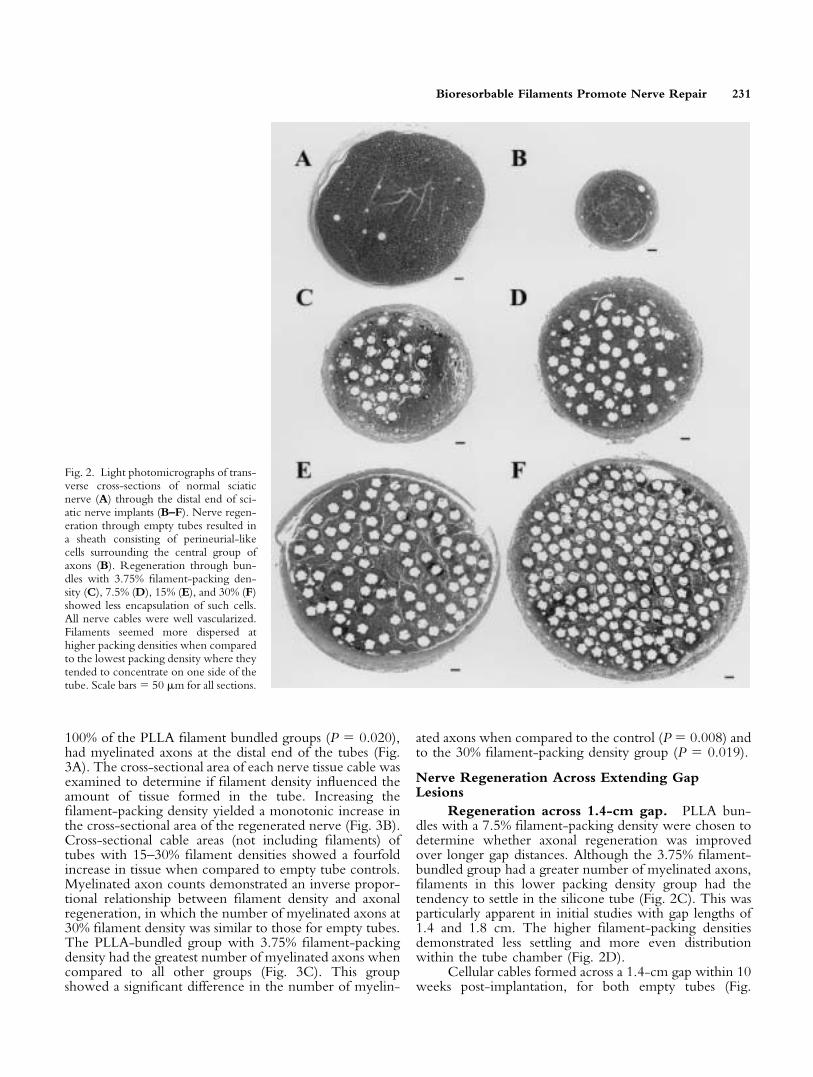
eter. Light microscopy was used to evaluate the morphologyof the regenerated nerve tissue at the distal stump (Fig. 2A–F).Nerve regeneration through empty tubes consisted of asheath of perineurial-like cells surrounding the central groupof axons (Fig. 2B). Nerve reconstruction through PLLAbundles with 3.75% filament-packing density (Fig. 2C), 7.5%(Fig. 2D), 15% (Fig. 2E), and 30% (Fig. 2F) showed lessfibroblastic and collagen encapsulation. All of the recon-structed nerves were well vascularized, with many showingnumerous myelinated axons. At the lowest packing density,filaments often seemed to concentrate toward one side of thetube. This was most likely due to gravitational settling of thefilaments to the bottom of the tube. At mid-packing densi-ties, filaments seemed more evenly spaced with larger dis-tances between filaments, when compared to the highestfilament-packing density.
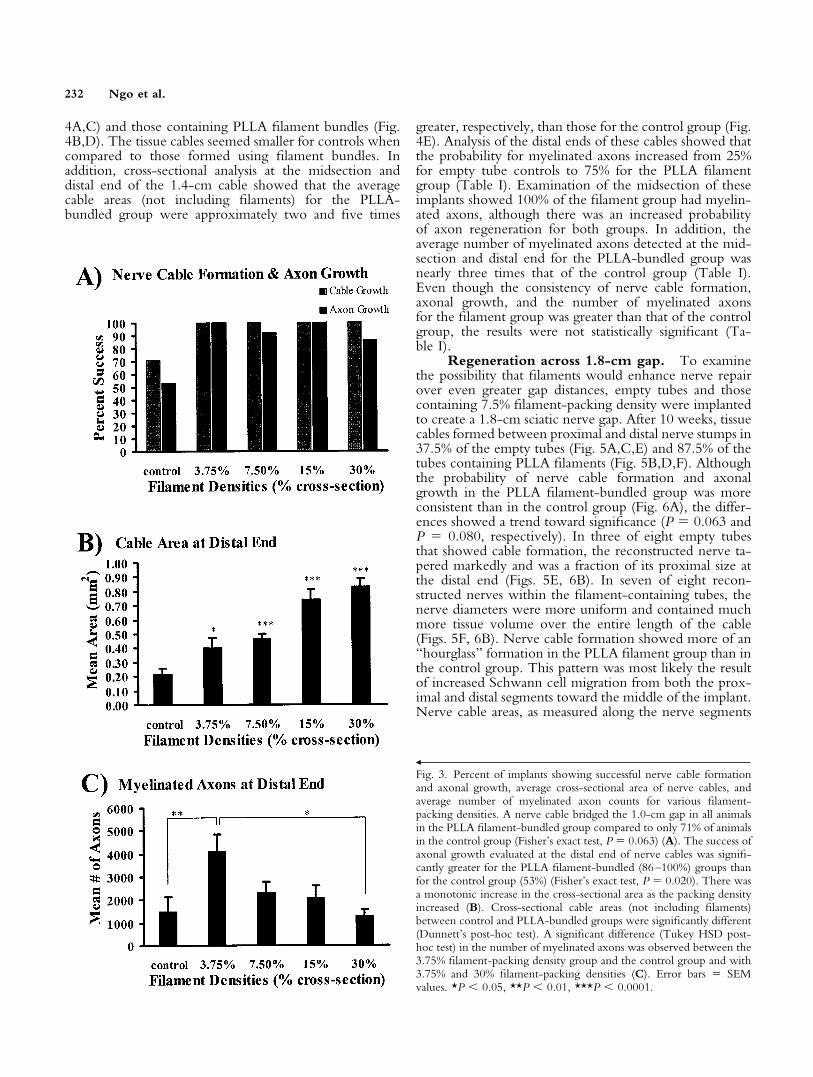
Although 71% of the animals in the control grouphad a nerve cable bridging the 1.0-cm gap, all animals inthe PLLA filament bundled group formed a cable acrossthe lesion (Fig. 3A); however, this difference showed atrend toward significant (P � 0.063). This was expected,because a previous study showed regeneration of a severednerve through silicone tubes could occur over gaps up to1.0 cm (in a rat model) with little means of enhancement(Lundborg et al., 1982b). Evaluation of axonal regenera-tion showed that only 53% of the empty tubes, vs. 86–
Fig. 1. Photomicrographs of regenera-tion across a 1.0-cm lesioned gap of thesciatic nerve in rats. Transverse cross-section of the distal end of a recon-structed nerve 10 weeks after implanta-tion showed many well-defined fascicles(A). High magnification of one of thefascicles (box in A) showed bundles ofmyelinated axons surrounded by well-delineated perineuriums (B, arrowheads).Further examination of the nerve re-gion (box in B) by TEM (C) showeda number of unmyelinated axons (*)present within the endoneurial sheathsof Schwann cells as well as blood vessels(bv). Myelinated axons (arrows) couldalso be seen a few microns away fromthe filament edge (double-headed ar-row) which was surrounded by thin lay-ers of cells (C). Scale bars � 50 �m,(A,B); 10 �m (C).
230 Ngo et al.
100% of the PLLA filament bundled groups (P � 0.020),had myelinated axons at the distal end of the tubes (Fig.3A). The cross-sectional area of each nerve tissue cable wasexamined to determine if filament density influenced theamount of tissue formed in the tube. Increasing thefilament-packing density yielded a monotonic increase inthe cross-sectional area of the regenerated nerve (Fig. 3B).Cross-sectional cable areas (not including filaments) oftubes with 15–30% filament densities showed a fourfoldincrease in tissue when compared to empty tube controls.Myelinated axon counts demonstrated an inverse propor-tional relationship between filament density and axonalregeneration, in which the number of myelinated axons at30% filament density was similar to those for empty tubes.The PLLA-bundled group with 3.75% filament-packingdensity had the greatest number of myelinated axons whencompared to all other groups (Fig. 3C). This groupshowed a significant difference in the number of myelin-
ated axons when compared to the control (P � 0.008) andto the 30% filament-packing density group (P � 0.019).
Nerve Regeneration Across Extending GapLesions
Regeneration across 1.4-cm gap. PLLA bun-dles with a 7.5% filament-packing density were chosen todetermine whether axonal regeneration was improvedover longer gap distances. Although the 3.75% filament-bundled group had a greater number of myelinated axons,filaments in this lower packing density group had thetendency to settle in the silicone tube (Fig. 2C). This wasparticularly apparent in initial studies with gap lengths of1.4 and 1.8 cm. The higher filament-packing densitiesdemonstrated less settling and more even distributionwithin the tube chamber (Fig. 2D).
Cellular cables formed across a 1.4-cm gap within 10weeks post-implantation, for both empty tubes (Fig.
Fig. 2. Light photomicrographs of trans-verse cross-sections of normal sciaticnerve (A) through the distal end of sci-atic nerve implants (B–F). Nerve regen-eration through empty tubes resulted ina sheath consisting of perineurial-likecells surrounding the central group ofaxons (B). Regeneration through bun-dles with 3.75% filament-packing den-sity (C), 7.5% (D), 15% (E), and 30% (F)showed less encapsulation of such cells.All nerve cables were well vascularized.Filaments seemed more dispersed athigher packing densities when comparedto the lowest packing density where theytended to concentrate on one side of thetube. Scale bars � 50 �m for all sections.
Bioresorbable Filaments Promote Nerve Repair 231
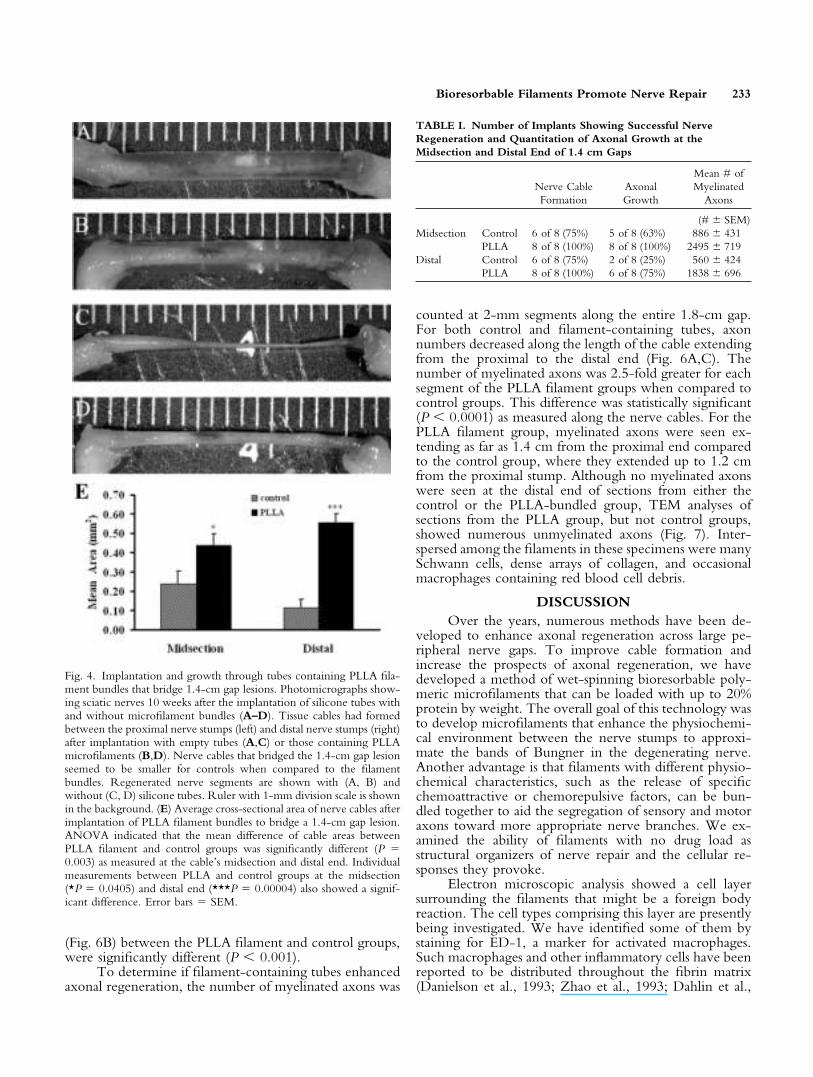
4A,C) and those containing PLLA filament bundles (Fig.4B,D). The tissue cables seemed smaller for controls whencompared to those formed using filament bundles. Inaddition, cross-sectional analysis at the midsection anddistal end of the 1.4-cm cable showed that the averagecable areas (not including filaments) for the PLLA-bundled group were approximately two and five times
greater, respectively, than those for the control group (Fig.4E). Analysis of the distal ends of these cables showed thatthe probability for myelinated axons increased from 25%for empty tube controls to 75% for the PLLA filamentgroup (Table I). Examination of the midsection of theseimplants showed 100% of the filament group had myelin-ated axons, although there was an increased probabilityof axon regeneration for both groups. In addition, theaverage number of myelinated axons detected at the mid-section and distal end for the PLLA-bundled group wasnearly three times that of the control group (Table I).Even though the consistency of nerve cable formation,axonal growth, and the number of myelinated axonsfor the filament group was greater than that of the controlgroup, the results were not statistically significant (Ta-ble I).
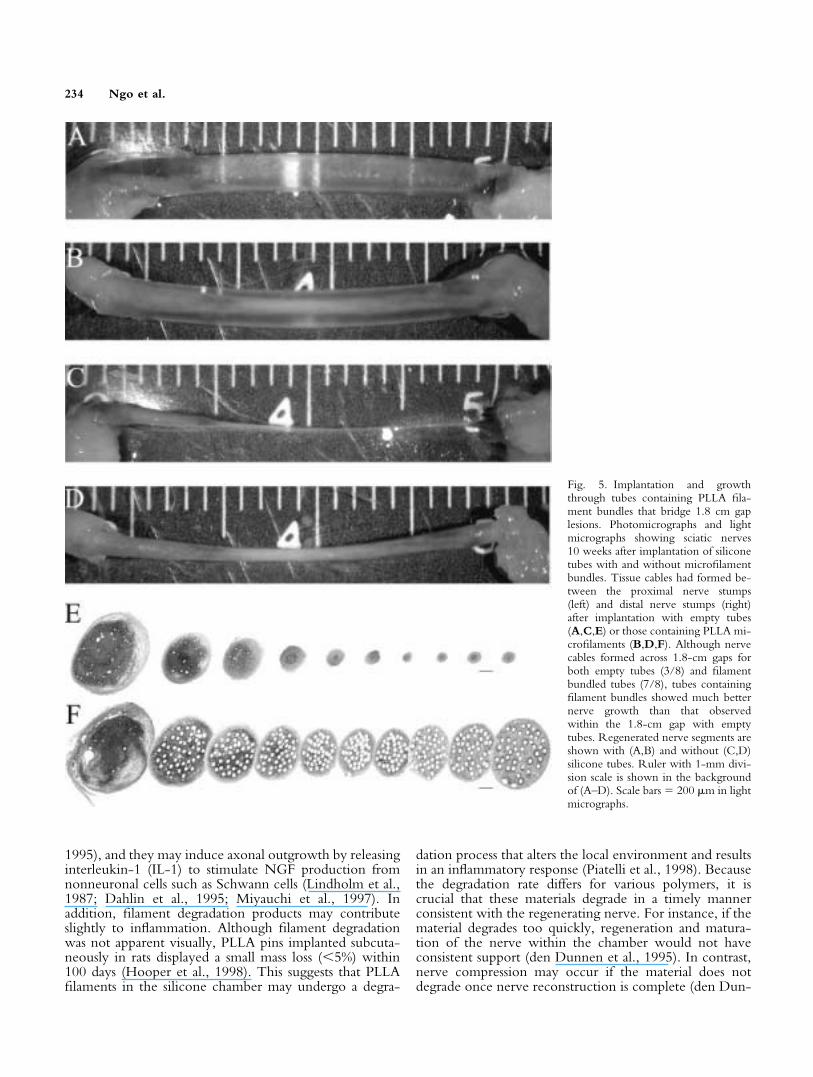
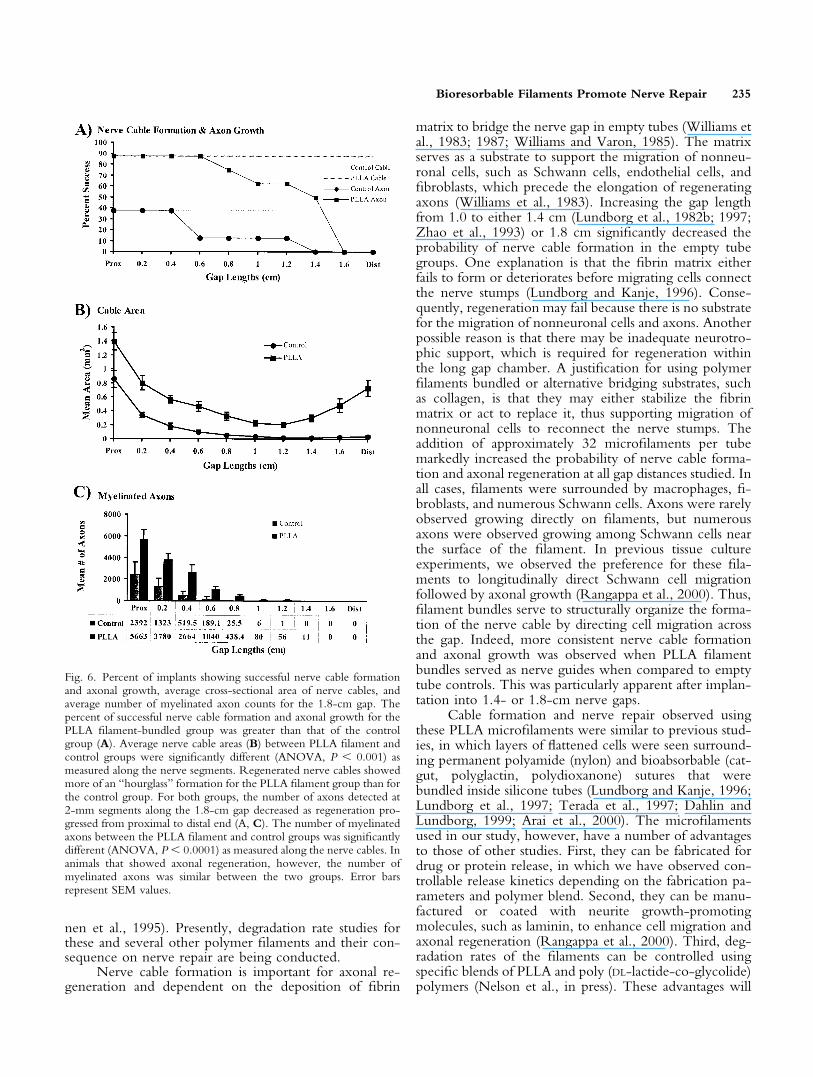
Regeneration across 1.8-cm gap. To examinethe possibility that filaments would enhance nerve repairover even greater gap distances, empty tubes and thosecontaining 7.5% filament-packing density were implantedto create a 1.8-cm sciatic nerve gap. After 10 weeks, tissuecables formed between proximal and distal nerve stumps in37.5% of the empty tubes (Fig. 5A,C,E) and 87.5% of thetubes containing PLLA filaments (Fig. 5B,D,F). Althoughthe probability of nerve cable formation and axonalgrowth in the PLLA filament-bundled group was moreconsistent than in the control group (Fig. 6A), the differ-ences showed a trend toward significance (P � 0.063 andP � 0.080, respectively). In three of eight empty tubesthat showed cable formation, the reconstructed nerve ta-pered markedly and was a fraction of its proximal size atthe distal end (Figs. 5E, 6B). In seven of eight recon-structed nerves within the filament-containing tubes, thenerve diameters were more uniform and contained muchmore tissue volume over the entire length of the cable(Figs. 5F, 6B). Nerve cable formation showed more of an“hourglass” formation in the PLLA filament group than inthe control group. This pattern was most likely the resultof increased Schwann cell migration from both the prox-imal and distal segments toward the middle of the implant.Nerve cable areas, as measured along the nerve segments
Š
Fig. 3. Percent of implants showing successful nerve cable formationand axonal growth, average cross-sectional area of nerve cables, andaverage number of myelinated axon counts for various filament-packing densities. A nerve cable bridged the 1.0-cm gap in all animalsin the PLLA filament-bundled group compared to only 71% of animalsin the control group (Fisher’s exact test, P � 0.063) (A). The success ofaxonal growth evaluated at the distal end of nerve cables was signifi-cantly greater for the PLLA filament-bundled (86–100%) groups thanfor the control group (53%) (Fisher’s exact test, P � 0.020). There wasa monotonic increase in the cross-sectional area as the packing densityincreased (B). Cross-sectional cable areas (not including filaments)between control and PLLA-bundled groups were significantly different(Dunnett’s post-hoc test). A significant difference (Tukey HSD post-hoc test) in the number of myelinated axons was observed between the3.75% filament-packing density group and the control group and with3.75% and 30% filament-packing densities (C). Error bars � SEMvalues. *P � 0.05, **P � 0.01, ***P � 0.0001.
232 Ngo et al.
(Fig. 6B) between the PLLA filament and control groups,were significantly different (P � 0.001).
To determine if filament-containing tubes enhancedaxonal regeneration, the number of myelinated axons was
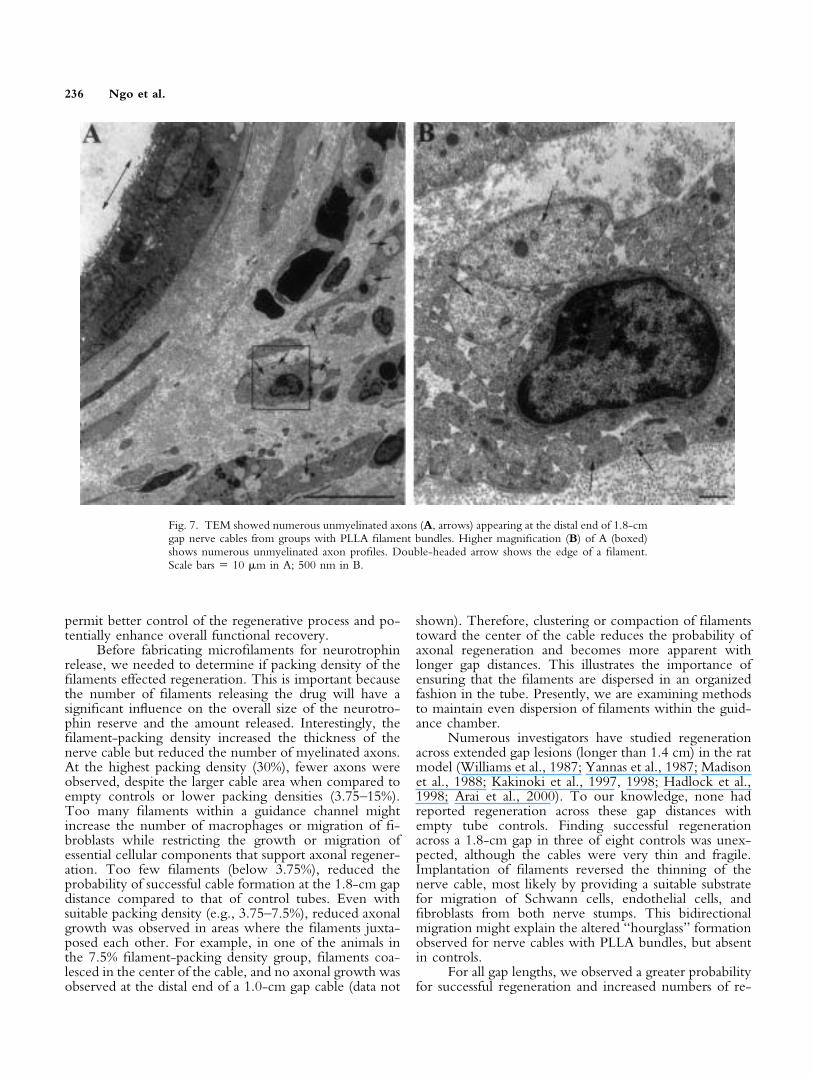
counted at 2-mm segments along the entire 1.8-cm gap.For both control and filament-containing tubes, axonnumbers decreased along the length of the cable extendingfrom the proximal to the distal end (Fig. 6A,C). Thenumber of myelinated axons was 2.5-fold greater for eachsegment of the PLLA filament groups when compared tocontrol groups. This difference was statistically significant(P � 0.0001) as measured along the nerve cables. For thePLLA filament group, myelinated axons were seen ex-tending as far as 1.4 cm from the proximal end comparedto the control group, where they extended up to 1.2 cmfrom the proximal stump. Although no myelinated axonswere seen at the distal end of sections from either thecontrol or the PLLA-bundled group, TEM analyses ofsections from the PLLA group, but not control groups,showed numerous unmyelinated axons (Fig. 7). Inter-spersed among the filaments in these specimens were manySchwann cells, dense arrays of collagen, and occasionalmacrophages containing red blood cell debris.
DISCUSSIONOver the years, numerous methods have been de-
veloped to enhance axonal regeneration across large pe-ripheral nerve gaps. To improve cable formation andincrease the prospects of axonal regeneration, we havedeveloped a method of wet-spinning bioresorbable poly-meric microfilaments that can be loaded with up to 20%protein by weight. The overall goal of this technology wasto develop microfilaments that enhance the physiochemi-cal environment between the nerve stumps to approxi-mate the bands of Bungner in the degenerating nerve.Another advantage is that filaments with different physio-chemical characteristics, such as the release of specificchemoattractive or chemorepulsive factors, can be bun-dled together to aid the segregation of sensory and motoraxons toward more appropriate nerve branches. We ex-amined the ability of filaments with no drug load asstructural organizers of nerve repair and the cellular re-sponses they provoke.
Electron microscopic analysis showed a cell layersurrounding the filaments that might be a foreign bodyreaction. The cell types comprising this layer are presentlybeing investigated. We have identified some of them bystaining for ED-1, a marker for activated macrophages.Such macrophages and other inflammatory cells have beenreported to be distributed throughout the fibrin matrix(Danielson et al., 1993; Zhao et al., 1993; Dahlin et al.,
Fig. 4. Implantation and growth through tubes containing PLLA fila-ment bundles that bridge 1.4-cm gap lesions. Photomicrographs show-ing sciatic nerves 10 weeks after the implantation of silicone tubes withand without microfilament bundles (A–D). Tissue cables had formedbetween the proximal nerve stumps (left) and distal nerve stumps (right)after implantation with empty tubes (A,C) or those containing PLLAmicrofilaments (B,D). Nerve cables that bridged the 1.4-cm gap lesionseemed to be smaller for controls when compared to the filamentbundles. Regenerated nerve segments are shown with (A, B) andwithout (C, D) silicone tubes. Ruler with 1-mm division scale is shownin the background. (E) Average cross-sectional area of nerve cables afterimplantation of PLLA filament bundles to bridge a 1.4-cm gap lesion.ANOVA indicated that the mean difference of cable areas betweenPLLA filament and control groups was significantly different (P �0.003) as measured at the cable’s midsection and distal end. Individualmeasurements between PLLA and control groups at the midsection(*P � 0.0405) and distal end (***P � 0.00004) also showed a signif-icant difference. Error bars � SEM.
TABLE I. Number of Implants Showing Successful NerveRegeneration and Quantitation of Axonal Growth at theMidsection and Distal End of 1.4 cm Gaps
Nerve CableFormation
AxonalGrowth
Mean # ofMyelinated
Axons
(# � SEM)Midsection Control 6 of 8 (75%) 5 of 8 (63%) 886 � 431
PLLA 8 of 8 (100%) 8 of 8 (100%) 2495 � 719Distal Control 6 of 8 (75%) 2 of 8 (25%) 560 � 424
PLLA 8 of 8 (100%) 6 of 8 (75%) 1838 � 696
Bioresorbable Filaments Promote Nerve Repair 233
1995), and they may induce axonal outgrowth by releasinginterleukin-1 (IL-1) to stimulate NGF production fromnonneuronal cells such as Schwann cells (Lindholm et al.,1987; Dahlin et al., 1995; Miyauchi et al., 1997). Inaddition, filament degradation products may contributeslightly to inflammation. Although filament degradationwas not apparent visually, PLLA pins implanted subcuta-neously in rats displayed a small mass loss (�5%) within100 days (Hooper et al., 1998). This suggests that PLLAfilaments in the silicone chamber may undergo a degra-
dation process that alters the local environment and resultsin an inflammatory response (Piatelli et al., 1998). Becausethe degradation rate differs for various polymers, it iscrucial that these materials degrade in a timely mannerconsistent with the regenerating nerve. For instance, if thematerial degrades too quickly, regeneration and matura-tion of the nerve within the chamber would not haveconsistent support (den Dunnen et al., 1995). In contrast,nerve compression may occur if the material does notdegrade once nerve reconstruction is complete (den Dun-
Fig. 5. Implantation and growththrough tubes containing PLLA fila-ment bundles that bridge 1.8 cm gaplesions. Photomicrographs and lightmicrographs showing sciatic nerves10 weeks after implantation of siliconetubes with and without microfilamentbundles. Tissue cables had formed be-tween the proximal nerve stumps(left) and distal nerve stumps (right)after implantation with empty tubes(A,C,E) or those containing PLLA mi-crofilaments (B,D,F). Although nervecables formed across 1.8-cm gaps forboth empty tubes (3/8) and filamentbundled tubes (7/8), tubes containingfilament bundles showed much betternerve growth than that observedwithin the 1.8-cm gap with emptytubes. Regenerated nerve segments areshown with (A,B) and without (C,D)silicone tubes. Ruler with 1-mm divi-sion scale is shown in the backgroundof (A–D). Scale bars � 200 �m in lightmicrographs.
234 Ngo et al.
nen et al., 1995). Presently, degradation rate studies forthese and several other polymer filaments and their con-sequence on nerve repair are being conducted.
Nerve cable formation is important for axonal re-generation and dependent on the deposition of fibrin
matrix to bridge the nerve gap in empty tubes (Williams etal., 1983; 1987; Williams and Varon, 1985). The matrixserves as a substrate to support the migration of nonneu-ronal cells, such as Schwann cells, endothelial cells, andfibroblasts, which precede the elongation of regeneratingaxons (Williams et al., 1983). Increasing the gap lengthfrom 1.0 to either 1.4 cm (Lundborg et al., 1982b; 1997;Zhao et al., 1993) or 1.8 cm significantly decreased theprobability of nerve cable formation in the empty tubegroups. One explanation is that the fibrin matrix eitherfails to form or deteriorates before migrating cells connectthe nerve stumps (Lundborg and Kanje, 1996). Conse-quently, regeneration may fail because there is no substratefor the migration of nonneuronal cells and axons. Anotherpossible reason is that there may be inadequate neurotro-phic support, which is required for regeneration withinthe long gap chamber. A justification for using polymerfilaments bundled or alternative bridging substrates, suchas collagen, is that they may either stabilize the fibrinmatrix or act to replace it, thus supporting migration ofnonneuronal cells to reconnect the nerve stumps. Theaddition of approximately 32 microfilaments per tubemarkedly increased the probability of nerve cable forma-tion and axonal regeneration at all gap distances studied. Inall cases, filaments were surrounded by macrophages, fi-broblasts, and numerous Schwann cells. Axons were rarelyobserved growing directly on filaments, but numerousaxons were observed growing among Schwann cells nearthe surface of the filament. In previous tissue cultureexperiments, we observed the preference for these fila-ments to longitudinally direct Schwann cell migrationfollowed by axonal growth (Rangappa et al., 2000). Thus,filament bundles serve to structurally organize the forma-tion of the nerve cable by directing cell migration acrossthe gap. Indeed, more consistent nerve cable formationand axonal growth was observed when PLLA filamentbundles served as nerve guides when compared to emptytube controls. This was particularly apparent after implan-tation into 1.4- or 1.8-cm nerve gaps.
Cable formation and nerve repair observed usingthese PLLA microfilaments were similar to previous stud-ies, in which layers of flattened cells were seen surround-ing permanent polyamide (nylon) and bioabsorbable (cat-gut, polyglactin, polydioxanone) sutures that werebundled inside silicone tubes (Lundborg and Kanje, 1996;Lundborg et al., 1997; Terada et al., 1997; Dahlin andLundborg, 1999; Arai et al., 2000). The microfilamentsused in our study, however, have a number of advantagesto those of other studies. First, they can be fabricated fordrug or protein release, in which we have observed con-trollable release kinetics depending on the fabrication pa-rameters and polymer blend. Second, they can be manu-factured or coated with neurite growth-promotingmolecules, such as laminin, to enhance cell migration andaxonal regeneration (Rangappa et al., 2000). Third, deg-radation rates of the filaments can be controlled usingspecific blends of PLLA and poly (DL-lactide-co-glycolide)polymers (Nelson et al., in press). These advantages will
Fig. 6. Percent of implants showing successful nerve cable formationand axonal growth, average cross-sectional area of nerve cables, andaverage number of myelinated axon counts for the 1.8-cm gap. Thepercent of successful nerve cable formation and axonal growth for thePLLA filament-bundled group was greater than that of the controlgroup (A). Average nerve cable areas (B) between PLLA filament andcontrol groups were significantly different (ANOVA, P � 0.001) asmeasured along the nerve segments. Regenerated nerve cables showedmore of an “hourglass” formation for the PLLA filament group than forthe control group. For both groups, the number of axons detected at2-mm segments along the 1.8-cm gap decreased as regeneration pro-gressed from proximal to distal end (A, C). The number of myelinatedaxons between the PLLA filament and control groups was significantlydifferent (ANOVA, P � 0.0001) as measured along the nerve cables. Inanimals that showed axonal regeneration, however, the number ofmyelinated axons was similar between the two groups. Error barsrepresent SEM values.
Bioresorbable Filaments Promote Nerve Repair 235
permit better control of the regenerative process and po-tentially enhance overall functional recovery.
Before fabricating microfilaments for neurotrophinrelease, we needed to determine if packing density of thefilaments effected regeneration. This is important becausethe number of filaments releasing the drug will have asignificant influence on the overall size of the neurotro-phin reserve and the amount released. Interestingly, thefilament-packing density increased the thickness of thenerve cable but reduced the number of myelinated axons.At the highest packing density (30%), fewer axons wereobserved, despite the larger cable area when compared toempty controls or lower packing densities (3.75–15%).Too many filaments within a guidance channel mightincrease the number of macrophages or migration of fi-broblasts while restricting the growth or migration ofessential cellular components that support axonal regener-ation. Too few filaments (below 3.75%), reduced theprobability of successful cable formation at the 1.8-cm gapdistance compared to that of control tubes. Even withsuitable packing density (e.g., 3.75–7.5%), reduced axonalgrowth was observed in areas where the filaments juxta-posed each other. For example, in one of the animals inthe 7.5% filament-packing density group, filaments coa-lesced in the center of the cable, and no axonal growth wasobserved at the distal end of a 1.0-cm gap cable (data not
shown). Therefore, clustering or compaction of filamentstoward the center of the cable reduces the probability ofaxonal regeneration and becomes more apparent withlonger gap distances. This illustrates the importance ofensuring that the filaments are dispersed in an organizedfashion in the tube. Presently, we are examining methodsto maintain even dispersion of filaments within the guid-ance chamber.
Numerous investigators have studied regenerationacross extended gap lesions (longer than 1.4 cm) in the ratmodel (Williams et al., 1987; Yannas et al., 1987; Madisonet al., 1988; Kakinoki et al., 1997, 1998; Hadlock et al.,1998; Arai et al., 2000). To our knowledge, none hadreported regeneration across these gap distances withempty tube controls. Finding successful regenerationacross a 1.8-cm gap in three of eight controls was unex-pected, although the cables were very thin and fragile.Implantation of filaments reversed the thinning of thenerve cable, most likely by providing a suitable substratefor migration of Schwann cells, endothelial cells, andfibroblasts from both nerve stumps. This bidirectionalmigration might explain the altered “hourglass” formationobserved for nerve cables with PLLA bundles, but absentin controls.
For all gap lengths, we observed a greater probabilityfor successful regeneration and increased numbers of re-
Fig. 7. TEM showed numerous unmyelinated axons (A, arrows) appearing at the distal end of 1.8-cmgap nerve cables from groups with PLLA filament bundles. Higher magnification (B) of A (boxed)shows numerous unmyelinated axon profiles. Double-headed arrow shows the edge of a filament.Scale bars � 10 �m in A; 500 nm in B.
236 Ngo et al.

generating axons with filament implants compared to con-trols. For gap lengths of 1.0 and 1.4 cm, the number ofmyelinated axons observed in the distal stump were 2.6-and 3.2-fold greater than those observed in empty tubecontrols. At a gap length of 1.8 cm, few myelinated axons,but multiple unmyelinated axons, were observed at thedistal end. As sections of nerve cables were examinedtoward the proximal stump, myelinated axons were de-tected, with the number increasing toward the proximalstump. To determine the farthest length at which myelin-ated axons had regenerated, sections were examined at2-mm increments from the distal to the proximal stump.The number of myelinated axons tapered off with increas-ing distance from the proximal stump, indicating thataxonal regeneration and myelination were still takingplace.
Although myelinated axons were not detected at thedistal end for PLLA filament bundles with nerve cablesspanning the entire 1.8-cm gap, unmyelinated axons wereobserved at this location. These structures were similar tothose seen previously in similar studies (Morris et al., 1972;Williams et al., 1983), which referred to them as “regen-erating units”, representing groups of axons still in a re-generating phase. With time, these axons should matureand become myelinated, and axonal regeneration across1.8-cm gap lengths would be successful. Future studieswould include long-term implantation periods and func-tional studies with the PLLA filament bundles across1.8-cm gaps to test the efficacy of these filament bundles asnerve guidance devices. For those studies, we have begunfabricating microporous bioresorbable tubes to replace thesilicone tubes. Preliminary studies demonstrate a greaterrate of axonal growth along filaments in these tubes com-pared to growth in silicone tubes. We also have fabricatedPLLA/PGA filaments that dissolve more rapidly to releaseneurotrophin to enhance further regeneration across1.8-cm long gaps.
ACKNOWLEDGMENTSWe thank Dr. H. Truszczynska for statistical consul-
tation and Drs. N. Rangappa and J. Cai for helpful dis-cussions.
REFERENCESArai T, Lundborg G, Dahlin LB. 2000. Bioartificial nerve graft for bridging
extended nerve defects in rat sciatic nerve based on resorbable guidingfilaments. Scand J Plast Reconstr Surg Hand Surg 34:101–108.
Aldini NN, Perego G, Cella GD, Maltarello MC, Fini M. Rocca M,Giardino R. 1996. Effectiveness of a bioabsorbable conduit in the repairof peripheral nerve. Biomaterials 17:959–962.
Archibald SJ, Krarup C, Shefner J, Li ST, Madison RD. 1991. A collagen-based nerve guide conduit for peripheral nerve repair: an electrophysio-logical study of nerve regeneration in rodents and nonhuman primates.J Comp Neurol 306:685–696.
Bailey SB, Eichler ME, Villadiego A, Rich KM. 1993. The influence offibronectin and laminin during Schwann cell migration and peripheralnerve regeneration through silicone chambers. J Neurocytology 22:176–184.
Bain JR, Mackinnon SE, Hudson AR, Falk RE, Falk JA, Hunter DA.1988. The peripheral nerve allograft: a dose-response curve in the ratimmunosuppressed with cyclosporin A. Plast Reconstr Surg 82:447–457.
Chamberlain LJ, Yannas IV, Arrizabalaga A, Hsu HP, Norregaard TV,Spector M. 1998. Early peripheral nerve healing in collagen and siliconetube implants: myofibroblasts and the cellular response. Biomaterials19:1393–1403.
Chiu DT, Janecka I, Krizek TJ, Wolff M, Lovelace RC. 1982. Autogenousvein grafts as a conduit for nerve regeneration. Surgery 91:226–233.
Dahlin LB, Lundborg G. 1999. Bridging defects in nerve continuity:influence of variations in synthetic fiber composition. J Mater Sci MaterMed 10:549–553.
Dahlin LB, Zhao Q, Bjursten LM. 1995. Nerve regeneration in siliconetubes: distribution of macrophages and interleukin-1a in the formed fibrinmatrix. Restor Neurol Neurosci 8:199–203.
Danielsen N, Dahlin LB, Thomsen P. 1993. Inflammatory cells and me-diators in the silicone chamber model for regeneration. Biomaterials14:1180–1185.
da Silva CF, Madison R, Dikkes P, Chiu TH, Sidman RL. 1985. An invivo model to quantify motor and sensory peripheral nerve regenerationusing bioresorbable nerve guides tubes. Brain Res 342:307–315.
DeFranzo AJ, Morykwas MJ, LaRosse JR, Jennings DA, Challa V, ArgentaLC. 1994. Autologous denatured muscle as a nerve graft. J ReconstrMicrosurg 10:145–151.
Dellon AL, Mackinnon SE. 1988. An alternative to the classical nerve graftfor the management of the short nerve gap. Plast Reconstr Surg 82:849–856.
den Dunnen WF, Stokroos I, Blaauw EH, Holwerda A, Pennings AJ,Robinson PH, Schakenraad JM. 1996. Light-microscopic and electron-microscopic evaluation of short-term nerve regeneration using a biode-gradable poly(DL-lactide-epsilon-caprolactone) nerve guide. J BiomedMater Res 31:105–115.
den Dunnen WF, van der Lei B, Robinson PH, Holwerda A, Pennings AJ,Schakenraad JM. 1995. Biological performance of a degradable poly(lacticacid-epsilon-caprolactone) nerve guide: influence of tube dimensions.J Biomed Mater Res 29:757–766.
den Dunnen WF, van der Lei B, Schakenraad JM, Stokroos I, Blaauw E.Bartels H, Pennings AJ, Robinson PH. 1997. Poly(DL-lactide-epsilon-caprolactone) nerve guides perform better than autologous nerve grafts.Microsurgery 17:348–357.
Dubuisson AS, Foidart-Dessalle M, Reznik M, Grosdent JC, Stevenaert A.1997. Predegenerated nerve allografts versus fresh nerve allografts in nerverepair. Exp Neurol 148:378–387.
Evans PJ, Mackinnon SE, Best TJ, Wade JA, Awerbuck DC, Makino AP,Hunter DA, Midha R. 1995. Regeneration across preserved peripheralnerve grafts. Muscle Nerve 18:1128–1138.
Fiodart-Dessalle M, Dubuisson A, Lejeune A, Severyns A, Manassis Y,Deiree P, Crielaard JM, Bassleer R, Lejeune G. 1997. Sciatic nerveregeneration through venous or nervous grafts in the rat. Exp Neurol148:236–246.
Gattuso JM, Glasby MA, Gschmeissner SE, Norris RW. 1989. A compar-ison of immediate and delayed repair of peripheral nerve using freeze-thawed autologous skeletal muscle grafts—in the rat. Br J Plast Surg42:306–313.
Gibson KL, Daniloff JK. 1989. Comparison of sciatic nerve regenerationthrough silicone tubes and nerve allografts. Microsurgery 10:126–129.
Guenard V, Kleiman N, Morrissey TK, Bunge RP, Aebischer P. 1992.Syngenic Schwann cells derived from adult nerves seeded in semiperme-able guidance channels enhance peripheral nerve regeneration. J Neurosci12:3310–3320.
Hadlock T, Elisseeff J, Langer R, Vacanti J, Cheney M. 1998. A tissue-engineered conduit for peripheral nerve repair. Arch Otolaryngol HeadNeck Surg 124:1081–1086.
Hafiz M, Lee S, Wolf P, Haghighi P. 1985. A comparative study ofautologous peripheral nerve grafts. Microsurgery 6:135–140.
Hooper KA, Macon ND, Kohn J. 1998. Comparative histological evalua-tion of new tyrosine-derived polymers and poly (L-lactic) as a function ofpolymer degradation. J Biomed Mater Res 41:443–454.
Bioresorbable Filaments Promote Nerve Repair 237

Kakinoki R, Nishijima N, Ueba Y, Oka M, Yamamuro T, Nakamura T.1997. Nerve regeneration over a 25 mm gap in the rat sciatic nerves usingtubes containing blood vessels: the possibility of clinical application. IntOrthop 21:332–336.
Kakinoki R, Nishijima N, Ueba Y, Oka M, Yamamuro T, Nakamura T.1998. Nerve regeneration over a 20-mm gap through a nerve conduitcontaining blood vessels in rats: the influence of interstump distance onnerve regeneration. J Neurosurg Sci 42:11–21.
Keeley RD, Nguyen KD, Stephanides MJ, Podilla J, Rosen JM. 1991. Theartificial nerve graft: a comparison of blended elastomer-hydrogel withpolyglycolic acid conduits. J Reconstr Microsurg 7:93–100.
Labrador RO, Buti M, Navarro X. 1998. Influence of collagen and laminingels concentration on nerve regeneration after resection and tube repair.Exp Neurol 149:234–252.
Leenslag AJ, Pennings JW, Bos RRM, Rozema FR, Boering G. 1987.Resorbable materials of poly(L-lactide): in vivo and in vitro degradation.Biomaterials 8:311–314.
Levi ADO, Sonntag VKH, Dickman C, Mather J, Li RH, Cordoba SC,Bichard B, Berens M. 1997. The role of cultured Schwann cells grafts inthe repair of gaps within the peripheral nervous systems of primates. ExpNeurol 143:25–36.
Li ST, Archibald SJ, Krarup C, Madison RD. 1992. Peripheral nerve repairwith collagen conduits. Clin Mater 9:195–200.
Lindholm D, Heumann R, Meyer M, Thoenen H. 1987. Interleukin-1regulates synthesis of nerve growth factor in nonneuronal cells of ratsciatic nerve. Nature 330:658–659.
Lundborg G, Dahlin LB, Danielson N. 1982a. Nerve regeneration across anextended gap: a neurobiological view of nerve repair and the possibleinvolvement of neurotrophic factors. J Hand Surg 7:580–587.
Lundborg G, Dahlin LB, Danielson N, Gelberman RH, Longo FM, PowellHC, Varon S. 1982b. Nerve regeneration in silicone chambers: influenceof gap length and of distal stump components. Exp Neurol 76:361–375.
Lundborg G, Dahlin LB, Dohi D, Kanje M, Terada N. 1997. A new typeof “bioartificial” nerve graft for bridging extended defects in nerves.J Hand Surg (Br) 22:299–303.
Lundborg G, Kanje M. 1996. Bioartificial nerve grafts. A prototype. ScandJ Plast Reconstr Surg Hand Surg 30:105–110.
Mackinnon SE, Dellon AL. 1990. Clinical nerve reconstruction with abioabsorbable polyglycolic acid tube. Plast Reconstr Surg 85:419–424.
Madison R, da Silva CF, Dikkes P, Chiu TH, Sidman RL. 1985. Increasedrate of peripheral nerve regeneration using bioresorbable nerve guides anda laminin-containing gel. Exp Neurol 88:767–772.
Madison RD, da Silva CF, Dikkes P. 1988. Entubulation repair withprotein additives increases the maximum nerve gap distance successfullybridged with tubular prostheses. Brain Res 447:325–334.
Miyauchi A, Kanje M, Danielsen N, Dahlin LB. 1997. Role of macro-phages in the stimulation and regeneration of sensory nerves by transposedgranulation tissue and temporal aspects of the response. Scand J PlastReconstr Surg Hand Surg 31:17–23.
Molander H, Engkvist O, Hagglund J, Olsson Y, Torebjork E. 1983. Nerverepair using a polyglactin tube and nerve graft: an experimental study inthe rabbit. Biomaterials 4:276–280.
Morris JH, Hudson AR, Weddell G. 1972. A study of degeneration andregeneration in the divided rat sciatic nerve based on electron microscopy.II. The development of the “regenerating unit”. Z Zellforsch MikroskAnat 124:103–130.
Nelson KD, Romero A, Waggoner P, Crow B, Borneman A, Smith GM.A technique paper for wet-spinning poly(l-lactic acid) and poly(dl-lactide-co-glycolide) monofilament fibers. Tissue Eng (in press).
Norris RW, Glasby MA, Gattuso JM, Bowden RE. 1988. Peripheral nerverepair in humans using muscle autografts. A new technique. J Bone JointSurg Br 70:530–533.
Piattelli A, Scarano X, Coraggio F, Matarasso S. 1998. Early tissue reactionsto polylactic acid resorbable membranes: a histological and histochemicalstudy in rabbits. Biomaterials 19:889–896.
Pistner H, Bendix DR, Muhling J, Reuther JF. 1993. Poly(L-lactide): along-term degradation study in vivo. Part III. Analytical characterization.Biomaterials. 14:291–298.
Rangappa, N, Romero A, Nelson KD, Eberhart RC, Smith GM. 2000.Laminin-coated poly(L-lactide) filaments induce robust neurite out-growth while providing directional orientation. J Biomed Mater Res51:625–634.
Rich KM, Alexander TD, Pryor JC, Hollowell JP. 1989. Nerve growthfactor enhances regeneration through silicone chambers. Exp Neurol105:162–170.
Rodriguez FJ, Gomez N, Perego G, Navarro X. 1999. Highly permeablepolylactide-caprolactone nerve guides enhance peripheral nerve regener-ation through long gaps. Biomaterials 20:1489–1500.
Seckel BR, Chiu TH, Nyilas E, Sidman RL. 1984. Nerve regenerationthrough synthetic biodegradable nerve guides: regulation by the targetorgan. Plast Reconstr Surg 74:173–181.
Terada N, Bjursten LM, Dohi D, Lundborg G. 1997. Bioartificial nervegrafts based on absorbable guiding filament structures—early observa-tions. Scand J Plast Reconstr Surg Hand Surg 31:1–6.
Tong XJ, Hirai KI, Shimada H, Mizutani Y, Izumi T, Toda N, Yu P. 1994.Sciatic nerve regeneration navigated by laminin-fibronectin doublecoated biodegradable collagen grafts in rats. Brain Res 663:155–162.
Wells MR, Kraus K, Batter DK, Blunt DG, Weremowitz J, Lynch SE,Antoniades HN, Hansson HA. 1997. Gel matrix vehicles for growthfactor application in nerve gap injuries repaired with tubes: a comparisonof biomatrix, collagen, and methylcellulose. Exp Neurol 146:395–402.
Williams LR. 1987. Exogenous fibrin matrix precursors stimulates thetemporal progress of nerve regeneration within a silicone chamber. Neu-rochem Res 12:851–860.
Williams LR, Danielsen N, Muller H, Varon S. 1987. Exogenous matrixprecursors promote functional nerve regeneration across a 15-mm gapwithin a silicone chamber in the rat. J Comp Neurol 264:284–290.
Williams LR, Longo FM, Powell HC, Lundborg G, Varon S. 1983.Spatial-temporal progress of peripheral nerve regeneration within a sili-cone chamber: parameters for a bioassay. J Comp Neurol 218:460–470.
Williams LR, Varon S. 1985. Modification of the fibrin matrix formationin situ enhances nerve regeneration in silicone chambers. J Comp Neurol231:209–220.
Yannas, IV, Orgill DP, Silver J, Norregaard TV, Zervas NT, Schoene WC.1987. Regeneration of sciatic nerve across 15 mm gap by use of apolymeric template. In: Gebelein CG, editor. Biomedical polymers. NewYork: Plenum. p 1–9.
Zhao Q, Dahlin LB, Kanje M, Lundborg G. 1993. Repair of the transectedrat sciatic nerve: matrix formation within implanted silicone tubes. RestorNeurol Neurosci 5:197–204.
238 Ngo et al.