assessment of epidermal nerve fibers
TRANSCRIPT

7Copyright @ 200 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.
REVIEW ARTICLE
Assessment of Epidermal Nerve Fibers: A New Diagnosticand Predictive Tool for Peripheral Neuropathies
Gigi J. Ebenezer, MBBS, MD, Peter Hauer, BS, Christopher Gibbons, MD,
Justin C. McArthur, MBBS, MPH, and Michael Polydefkis, MD, MHS
AbstractToday, skin biopsies can play an important role in the diagnosis
of peripheral nerve disorders and have yielded another diagnostic
tool for the neurologist. One of the commonly reported neuro-
pathologic abnormalities observed in skin biopsies is a reduction of
epidermal nerve density. Analyzing the changes in the morphology
and density of epidermal nerves is of immense diagnostic and
prognostic value in peripheral neuropathies. These changes also
provide an assessment of disease progression and of tissue
responses to regenerative treatments. Combined with immunohis-
tochemical studies, newly evolved skin biopsy and epidermal count
techniques have the potential to provide significant information
about the pathogenesis of many peripheral nervous system diseases.
They have great potential for impacts on both research and clinical
approaches to treatment. Evolution of a standardized and validated
counting methodology and significant advances in procuring skin
biopsies have opened up a wide spectrum of applications that make
the technology easy to apply in practice. The application of this
technology may lead to early detection of many common peripheral
nerve diseases and an enhanced understanding of disease onset and
progression. In this article we review the state of current research
and clinical practice in the use of skin biopsies and epidermal nerve
densities.
Key Words: Dermis, Epidermis, Peripheral neuropathies, Protein
gene product 9.5 (PGP 9.5), Schwann cell, Skin biopsy.
INTRODUCTIONThe skin is the largest organ in the body, covering the
entire external surface of the human body. It serves as aprotective barrier, facilitates in sensory perception, immuno-logic surveillance, and thermoregulation and controls insen-sible fluid loss. It constantly regenerates throughout life asits outer layers are shed and are replaced by inner layers.
The thickness of the skin varies according to anatomiclocation, sex, and age of the individual.
The skin is densely innervated (Fig. 1A) and sensoryimpulses from it are transmitted to the brain by fibers ofprimary sensory neurons located in trigeminal and dorsalroot ganglia. These neurons are a heterogeneous populationand include both mechanoreceptors and nociceptors. Noci-ceptive neurons detect noxious thermal, mechanical, andchemical stimuli that evoke a painful sensation and can warnof impending bodily injury. The peripherally directed axonsof dorsal root ganglia neurons run in peripheral nerves,terminate in skin, and innervate a variety of cutaneousstructures, including sweat glands (Fig. 1B), hair follicles,Merkel cells, Meissner’s corpuscles, blood vessels and theepidermis (1, 2).
In the skin, the nerve bundles typically course throughthe dermis vertically and form horizontal subepidermalneural plexuses in the papillary dermis. From the papillarydermal plexuses these nerves travel vertically, lose theirSchwann cell investment at the dermoepidermal junction,penetrate the epidermal basement membrane, ascendbetween the keratinocytes, and terminate in the keratin orsuperficial layers of the stratum spinosum as free nerveendings (Fig. 1C).
Cutaneous innervation consists mainly of unmyeli-nated fibers, which account for approximately 90% of alldermal nerve fibers (3). The unmyelinated axons areorganized into Remak bundles, which contain Schwann cellsenveloping 1 or more unmyelinated axons and the profile isenclosed by continuous basal lamina. The individual Remakbundles in the dermis typically contain 2 to 3 axons (Fig. 1D).Larger and deeper dermal nerves have a complete perineu-rium with a basal lamina and junctional complexes. Manysmaller and more superficial nerves lack perineurium and aresurrounded by collagen bundles oriented in the long axis ofthe nerve fibers, resembling endoneurial collagen.
Morphologically, the sensory axons within the skin areclassified into myelinated (A-fibers) and unmyelinated (C-fibers) fibers. Neurophysiologically, they are categorized asA-C and C-fibers. A-C fibers are small, thinly myelinatedaxons subserving mechanoreceptors and thermal receptorsthat transduce and transmit pain (nociception). C-fibers arevery thin unmyelinated axons that slowly transmit nocicep-tion sensation. Cool threshold detection is mediated by A-Cfibers and C-fibers and warm sensation by C-fibers (4).Slow-conducting unmyelinated (C) afferents also signal light
1059J Neuropathol Exp Neurol � Volume 66, Number 12, December 2007
J Neuropathol Exp NeurolCopyright � 2007 by the American Association of Neuropathologists, Inc.
Vol. 66, No. 12December 2007pp. 1059Y1073
From the Department of Neurology (GJE, PH, MP), Johns HopkinsUniversity, Baltimore, Maryland; Department of Neurology (CG), BethIsrael Deaconess Medical Center, Harvard Medical School, Boston, MA;and Departments of Neurology, Pathology and Epidemiology (JCM),Johns Hopkins University, Baltimore, Maryland.
Send correspondence and reprint requests to: Justin C. McArthur, MBBS,MPH, Interim Chair, Department of Neurology, Professor of Neurology,Pathology and Epidemiology, Johns Hopkins University, Meyer 6-113,600 N. Wolfe St., Baltimore MD 21287-7609; E-mail: [email protected]

7Copyright @ 200 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.
touch (5). Axonal conduction velocity depends on both themyelin thickness and nerve fiber diameter, and unmyelinatedC-fibers conduct impulses at about 1 m/s whereas myeli-nated A-fibers conduct at 40 to 1,000 m/s (2). Largemyelinated A> and AA fibers conduct vibration and lighttouch, and these fibers are not present in the epidermis (1).
Neurochemically, epidermal nerve endings can bebroadly categorized as either peptidergic or nonpeptidergic.Peptidergic axons densely express calcitonin gene-relatedpeptide (CGRP) along with other peptides including theneurotransmitter substance P. Nonpeptidergic afferents areidentified by their binding sites for the plant lectin IB4.Using genetically encoded axonal tracers expressed from theMrgrd locus, a subpopulation of small-diameter (presumedto be nociceptive, IB4+) nonpeptidergic epidermal axons thatexpress the molecular marker Mrgprd (Mrgprd+) have beenidentified in mice. These Mrgprd+ fibers course through the
stratum basalis and stratum spinosum and terminate in thestratum granulosum. In contrast, CGRP+ fibers terminate inthe stratum spinosum of the epidermis. Mrgprd+ fibers donot innervate any specific cutaneous sensory structure andthere is complete absence of Mrgprd+ fiber innervation in allother peripheral tissues. The central projections derived fromdistinct epidermal innervation zones terminate in adjacentlaminae in the dorsal spinal cord (6).
Sweat glands in the dermis are highly immunoreactiveto pan-axonal marker protein gene product 9.5)(Fig. 1B). The majority are vasoactive intestinal polypep-tide+ and some are CGRP+ but are sparsely innervated bysubstance P-immunoreactive fibers (7, 8). Characteristically,these fibers are cholinergic sympathetic axons with a fewnoradrenergic fibers. Cholinergic/noradrenergic coexpres-sion, which is a mode of autonomic regulation, is a uniquefeature of the primate peripheral nervous system and has not
FIGURE 1. Double-stainedconfocal images of protein gene product 9.5 (PGP 9.5) with p75 and electron microscopic findings ofcutaneous nerves in human controls. (A) Double-stained confocal images of PGP 9.5 (red) staining axons (arrows) combinedwith p75 (green) staining Remak Schwann cells and perineurium (broken arrows). Overlapping colocalization is yellow. Thedermal nerve bundles course through the dermis and form horizontal bundles at the level of the papillary dermis. Scale bar =20 Km. (Used, by permission. First published in Brain 2007;130(10):2703Y2714, modified). (B) PGP 9.5-stained axons (red)
the epidermis, but the p75 staining for Schwann cells (broken arrow) stops at the dermoepidermal junction. Scale bar = 20 Km.(D) A Remak bundle with Schwann cell (Sch) wrapping around 3 unmyelinated axons (arrows); the profile is enclosed bycontinuous basal lamina (broken arrow) and surrounded by collagen bundles (C). Scale bar = 1 Km. (Used, by permission. Firstpublished in Brain 2007;130(10):2703Y2714, modified).
Ebenezer et al J Neuropathol Exp Neurol � Volume 66, Number 12, December 2007
� 2007 American Association of Neuropathologists, Inc.1060
9.5 (PGP
densely innervating the sweat glands (S). Scale bar = 20 Km. (C)PGP 9.5-stained epidermal nerve fibers (arrows)(red) extend into

7Copyright @ 200 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.
yet been identified in other portions of the human sympa-thetic nervous system (9). Dermal blood vessels areinnervated by both sensory and autonomic fibers (sympa-thetic and parasympathetic), and most of these are unmyeli-nated (Fig. 2A). The autonomic fibers lie significantly closerto the endothelial cell layer and smooth muscle cellscompared with the sensory fibers (10).
Neurotrophic factors play an important role in thegrowth and maintenance of these terminal nerve endings.C-fiber nociceptors respond to trophic factors: nerve growthfactor (NGF), neurotrophin 3, glial cell line-derivedneurotrophic factor, and brain-derived neurotrophic factor(11, 12). NGF binds to 2 classes of transmembrane proteinreceptors: tyrosine kinase family (high-affinity receptor) andp75 (the low-affinity and pan-neurotrophin receptor). Thep75 receptor can be activated even in the absence of tyrosinekinase A. Expression of both NGF and p75 are important inthe regeneration of peripheral nerves after injury (13, 14).NGF and its receptor, tyrosine kinase A, are important forsurvival and target innervation of small diameter sensoryneurons and sympathetic neurons during development (15).Expression of NGF is maintained in the adult, especially intarget tissues such as skin keratinocytes, indicating apossible role in maintenance and plasticity of these neuronsin adulthood. Previous experiments in rats using an NGFantiserum infusion addressed this question (16) by demon-strating that the levels of NGF in sympathetic and sensoryneurons after axotomy partly regulate subsequent changes inneuropeptide expression. Yet, it remains unclear whetherNGF is important for maintaining survival or innervation ofsensory and sympathetic neurons in the adult nervoussystem.
ANALYSIS OF SKIN BIOPSIES INCUTANEOUS NERVE
Large sensory and motor nerve fibers in peripheralnerve disorders are typically assessed using electrodiagnos-
tic methods or sural nerve biopsy. However, electrodiag-nostic methods are not useful in assessing unmyelinatednerve fibers simply because this population is Binvisible[ tonerve conduction velocity studies. Within the epidermis, theindividual unmyelinated axons are separated from each otheras they pass between the keratinocytes, and at the lightmicroscopic level it is possible to identify and count axonsusing immunocytochemical stains. Before the advent ofsensitive pan-axonal markers such as PGP 9.5 (17), silverstains were used but were too insensitive for practical use.For almost 15 years skin biopsy has been used as a reliabletool to assess the epidermal and dermal nerve fibers. Theskin can be sampled at several anatomical sites simul-taneously, and biopsies can be repeated serially over time(18Y23).
MethodologySampling of skin to analyze epidermal nerves is done
essentially by using 2 techniques: skin blister and skin punchtechniques.
Skin Blister TechniqueA suction capsule is placed over the skin and the
epidermis is separated from the dermis at the level of theepidermal basement membrane (24). The technique is leastinvasive, painless, and bloodless, but the procedure is time-consuming and the relationship of dermal nerves to epi-dermal nerves cannot be studied.
Skin Punch TechniqueCutaneous nerve evaluation using 3-mm skin punches
(8, 22, 25) is easy to perform, no sutures are required, andthe test leaves behind only a very small scar. The majoradvantages of skin biopsies are intraepidermal nerves fibers(IENFs) can be quantified, dermal innervation and morpho-logic features can be evaluated, and sections can be used forultrastructural studies, polymerase chain reaction analysis,and laser pressure catapulting studies. In addition, from skin
FIGURE 2. Confocal images of AIII-tubulin staining dermal nerve bundles. (A) A neurovascular bundle with a nerve bundle(arrow) stained by AIII-tubulin (green) and small branching fibers innervating a blood vessel (red). Scale bar = 20 Km. (B) Adermal nerve bundle (arrows) stained by AIII-tubulin (green). Schwann cell nuclei are blue. Scale bar = 10 Km.
J Neuropathol Exp Neurol � Volume 66, Number 12, December 2007 Assessment of Epidermal Nerve Fibers
� 2007 American Association of Neuropathologists, Inc. 1061
7Copyright @ 200 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.
punches, epidermal sheets can be separated and subjected toprotein and mRNA analysis.
Processing and ImmunohistochemistryMost laboratories use either a PLP (paraformaldehyde,
lysine, and periodate) or Zamboni (2% paraformaldehydeand picric acid) fixative, as tissues fixed in formalin tend toproduce a fragmented appearance of nerve fibers (22, 26).PGP 9.5 is a cytoplasmic neuronal marker commonly used toinvestigate small caliber sensory fibers in skin biopsies, andanti-PGP 9.5 targets ubiquitin carboxyl-terminal esteraseL1, an enzyme found exclusively and ubiquitously in neu-rons (27). Other markers are antibodies against specific cyto-skeletal filaments (Fig. 2B), axonal membrane, and epitopes(28, 29).
Fifty-micrometer-thick sections are cut perpendicularto the epidermis to visualize both dermis and epidermisusing a freezing sliding microtome. At least 3 sections of 50Km from each biopsy are evaluated using bright field lightmicroscopy. Thick sections allow the visualization of
cutaneous nerves, as continuous wavy bundles and as singlenerves.
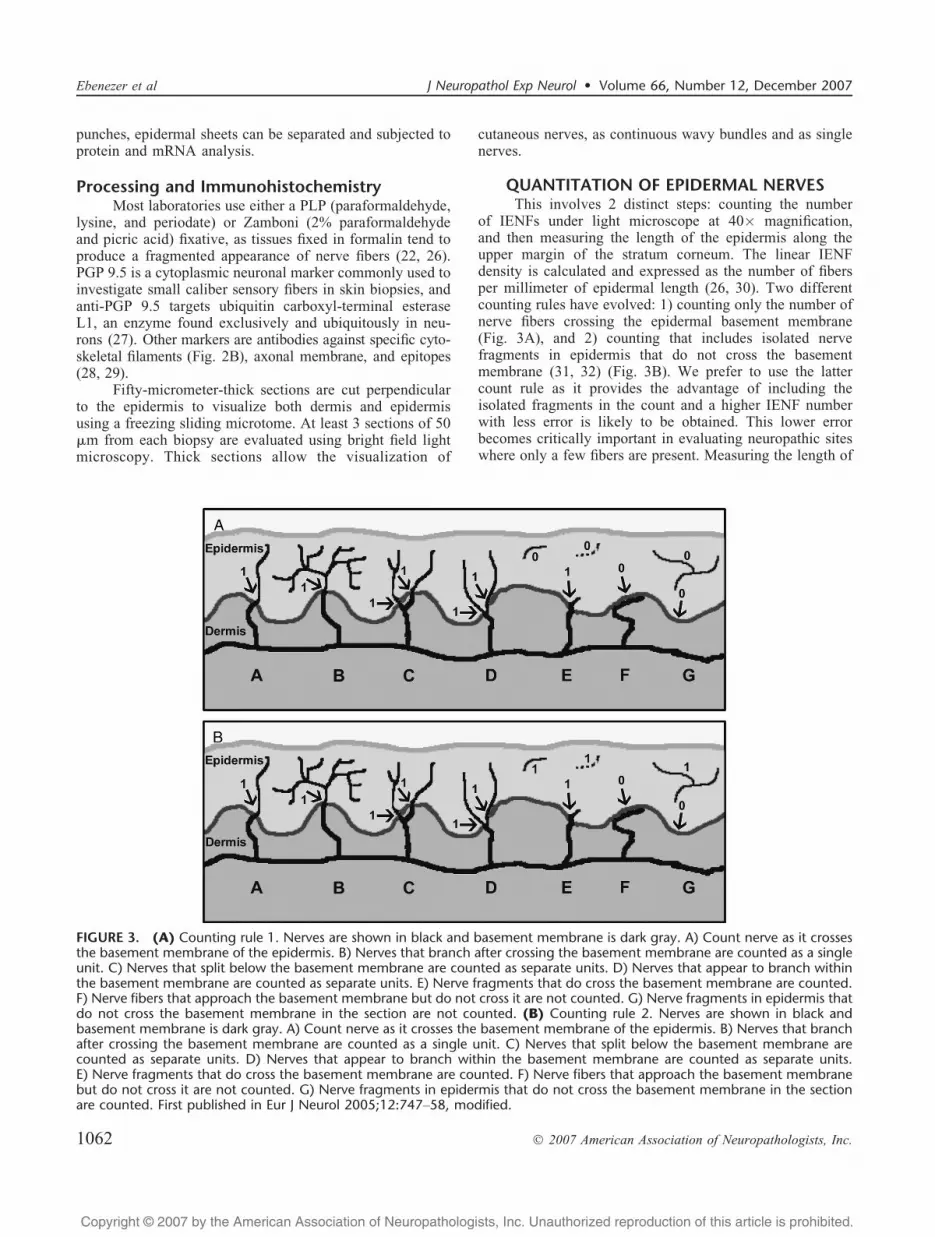
QUANTITATION OF EPIDERMAL NERVESThis involves 2 distinct steps: counting the number
of IENFs under light microscope at 40� magnification,and then measuring the length of the epidermis along theupper margin of the stratum corneum. The linear IENFdensity is calculated and expressed as the number of fibersper millimeter of epidermal length (26, 30). Two differentcounting rules have evolved: 1) counting only the number ofnerve fibers crossing the epidermal basement membrane(Fig. 3A), and 2) counting that includes isolated nervefragments in epidermis that do not cross the basementmembrane (31, 32) (Fig. 3B). We prefer to use the lattercount rule as it provides the advantage of including theisolated fragments in the count and a higher IENF numberwith less error is likely to be obtained. This lower errorbecomes critically important in evaluating neuropathic siteswhere only a few fibers are present. Measuring the length of
FIGURE 3. (A) Counting rule 1. Nerves are shown in black and basement membrane is dark gray. A) Count nerve as it crossesthe basement membrane of the epidermis. B) Nerves that branch after crossing the basement membrane are counted as a singleunit. C) Nerves that split below the basement membrane are counted as separate units. D) Nerves that appear to branch withinthe basement membrane are counted as separate units. E) Nerve fragments that do cross the basement membrane are counted.F) Nerve fibers that approach the basement membrane but do not cross it are not counted. G) Nerve fragments in epidermis thatdo not cross the basement membrane in the section are not counted. (B) Counting rule 2. Nerves are shown in black andbasement membrane is dark gray. A) Count nerve as it crosses the basement membrane of the epidermis. B) Nerves that branchafter crossing the basement membrane are counted as a single unit. C) Nerves that split below the basement membrane arecounted as separate units. D) Nerves that appear to branch within the basement membrane are counted as separate units.E) Nerve fragments that do cross the basement membrane are counted. F) Nerve fibers that approach the basement membranebut do not cross it are not counted. G) Nerve fragments in epidermis that do not cross the basement membrane in the sectionare counted. First published in Eur J Neurol 2005;12:747Y58, modified.
Ebenezer et al J Neuropathol Exp Neurol � Volume 66, Number 12, December 2007
� 2007 American Association of Neuropathologists, Inc.1062

7Copyright @ 200 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.
epidermis using both computerized image software and amicroscope intraocular lens ruler has produced similarresults (33). IENF density can also be derived simply bydividing the number of IENFs by 3 mm, and this result hascorrelated significantly with IENF density measured byimage analysis (34). Very good correlation exists betweenthe various numerical methods (35). The European Feder-ation of Neurological Studies task force has establishedguidelines and rules for counting and reporting IENFs(31, 32).
With strict counting rules and intensive training, a highdegree of inter- and intrarater reliability has been achievedin IENF density estimation. Density variations amongadjacent sections within the same site are minimal, and ahigh degree of correlation of IENF densities between 2punches at the same anatomical site has been demonstrated(26, 36). When the lowest 5th percentile of the normativerange in the distal leg is used as a cut-off value to identifynormal, the technique had a diagnostic efficiency of 88%, apositive predictive value of 75%, and a negative predictivevalue of 90% for the detection of small fiber sensoryneuropathy (26). Although this study used formalin fixation,IENF density differences between formalin and PLP fixationhave not been appreciative (37). In contrast, estimation ofIENF using confocal microscopy is expensive and laborintensive. Fifty- to 100-Km-thick sections are used forconfocal studies (23, 38, 39). As the IENF density dependson the optical sections created by the user, wide variations inresults occur, and normative data have not yet beenestablished using this technique.
IENF Density As a Diagnostic ToolIENF density normative data are available for only a
few anatomical sites: proximal thigh, distal thigh, distal leg,distal forearm, trunk and heel. Three major studies haveshown that in healthy subjects, IENF density is typicallyhigher in the proximal thigh than the distal leg with a normalproximal to distal length-dependent gradient in innervation(26, 37, 40) (Table, Fig. 4A, B).
In normal individuals, higher IENF densities are foundin very young individuals (G20 years). The effect of gender,age, and height on distal leg innervation has been inves-tigated by several groups. A study that focused on subjectsaged 16 to 82 years (26) did not find an effect of increasingage other than subjects in the youngest decile (16Y20 years)having higher densities. Other studies of healthy controlsubjects have reported a mild inverse, gender-adjustedrelationship between IENF density and age with a decreaseof 0.6 to 1.8 fibers/mm per decade (37, 40). It appears thatexcluding the extremes of age, the effect of age onepidermal innervation is modest, if any. Similarly, therehas been variation in the role that gender plays with somestudies reporting no effect (26) and others finding mildincreases in epidermal innervation among females (37, 40).No significant relationship of race, height, or weight onIENF density has been reported (26, 37). Interestingly,height, presumably as a surrogate for axonal length, is a riskfactor for the development of diabetic neuropathy and hasbeen linked to reduced rates of IENF regeneration rates (48).
The epidermal nerve fiber density measure has shown highspecificity and reliability and published data in the normalpopulation could serve as a good baseline for detecting smallfiber sensory neuropathy (26, 40, 49, 50). Studies haveestimated IENF in the forearm, trunk, and heel, but thesedata have not been consistent (22, 41, 51).
In patients with sensory neuropathy patients the IENFdensity is consistently and significantly lower than that incontrols (Fig. 4C, D) (20, 23, 39, 50, 52Y54). The IENFdensity measurements were found to be more sensitive thansensory nerve conduction studies for diagnosing small fibersensory neuropathy (19, 23, 42, 43, 50, 55).
Morphologic ChangesMorphologic changes are seen in both epidermal and
dermal nerves in peripheral neuropathy and sometimes inotherwise healthy individuals from local effects of trauma.Although some small-sized swellings, beading, and varicos-ities may be present in the leg skin biopsy of normalindividuals, attenuation of fibers, large globular and fusiform-shaped swellings (Fig. 5A), dystrophic changes, and tortuousand increasingly complex branching are some of the morpho-logic changes that have been described in the skin biopsies ofpatients with neuropathy (3, 18, 22, 42, 44, 56, 57). Swellingswere more numerous in the calf of patients with small fiberneuropathy and also tended to be more numerous in diabeticthighs (57).
The higher density of swellings in the distal leg ofneuropathic patients correlates well with impaired heat-painthreshold, development of symptoms, and progression ofdisease (42, 44). These axonal swellings are thought topresage the subsequent loss of axons; that is, large swellingsserve as predictive markers of nerve fiber degeneration. In astudy of 28 patients with sensory complaints of unknownetiology, large axonal swellings on the initial skin biopsysubsequently showed a decline in IENF density on repeatedbiopsies. This decline was not observed in patients withoutnerve fiber swellings or in those with smaller nerve fiberswellings (18).
TABLE. Intraepidermal Nerve Fiber Density: Normative Rangefrom Different Laboratories
Author andReference Number No. of Subjects
PTENF/mm
Distal LegENF/mm
McArthur et al (26) 98 21.1 T 10.4 13.8 T 6.7
Omdal et al (36) 15 Not done 12.2 T 3.8
Umapathi et al (37) 84 20.4 T 6.9 11.7 T 4.1*
GLransson et al (40) 106 Not done 12.4 T 4.6Pan et al (41) 55 Not done 12.970 T .284
Lauria et al (42) 15 22.6 T 4.8 14.4 T 3.0
Shun et al (43) 38 Not done 9.359 T 3.466
Herrmann et al (44) 19 21.6 T 5.4 16.6 T 5.3
22.8 T 6.9 15.0 T 5.0†
Lauria et al (46) 15 24.37 T 5.80 14.46 T 4.45
*, Male.†, Female.ENF, epidermal nerve fiber; PT, proximal thigh.
J Neuropathol Exp Neurol � Volume 66, Number 12, December 2007 Assessment of Epidermal Nerve Fibers
� 2007 American Association of Neuropathologists, Inc. 1063

7Copyright @ 200 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.
Ultrastructural examination of skin biopsies frompatients with neuropathy have shown that dilated cytoskele-tal organelles, disintegration of axoplasmic organelles,watery axoplasm, and accumulation of abnormal mitochon-dria within axons and Schwann cells contribute to theformation of axonal swellings in terminal nerve endings(Fig. 5B). Pathologic changes were similar in all types ofsensory neuropathic cases examined without any specificrecognizable differentiating features. Thus, nerve fiberswellings are now considered to represent a predegenerativechange, and the presence of larger swellings at the lowerlimb can be used as a predictive marker to assess theprogression of neuropathy. Further studies are needed todescribe a methodology to quantify the swellings.
INTRAEPIDERMAL NERVE FIBERS DENSITY INPERIPHERAL NERVE DISEASE
Diabetic NeuropathyIt is estimated that the global prevalence of diabetes
mellitus in people of all ages will increase from 2.8% in theyear 2000 to 4.4% in 2030. The total number of people withdiabetes is projected to rise from 171 million in 2000 to 366million in 2030 (58). Epidemiologic studies suggest that
45% to 60% of all diabetic patients develop neuropathy (59,60). Abnormalities on nerve conduction studies (NCS) orelectromyographic changes were found in 18% to 25% ofpatients at the time of diabetes diagnosis (61).
Direct metabolic effects due to hyperglycemia, oxida-tive injury, mitochondrial dysfunction, and disruption ofaxonal transport by advanced glycation end products havebeen implicated in the pathogenesis of neuropathy (62, 63).
A number of epidermal nerve quantification studieshave been completed in diabetic patients and in experimentaldiabetic animals. IENF densities are reduced in type 2diabetic patients compared with those in age-matchedhealthy controls and correlate with changes in warm sensorydetection thresholds and the amplitude of the sural sensoryaction potential. The extent of denervation increases with theduration of type 2 diabetes (43). There is an inversecorrelation between IENF density and severity of neuropathy(20, 64), with progressively decreasing densities from prox-imal to distal sites (39). IENF density is also lower in thosepresenting with neuropathic pain compared with thosepresenting without such pain (64). Differences betweenpeople with type I versus type II diabetes have not beenidentified although severity of IENF loss has been linked tothe severity of glucose dysmetabolism (50).
FIGURE 4. Skin biopsies immunostained with PGP 9.5 showing length-dependent innervation of skin in the lower limb of ahealthy adult and in a patient with neuropathy. (A) PGP 9.5-stained section showing epidermal nerve fibers (arrow) in the distalleg of a healthy adult. Scale bar = 50 Km. (B) PGP 9.5-stained section showing epidermal nerve fibers (arrows) in the proximalthigh of a healthy adult. Scale bar = 50 Km. (C) PGP 9.5- and p75-stained section showing complete absence of epidermal fibersin the distal leg in a patient with chronic neuropathy. Axons = red; Schwann cells = green. Scale bar = 20 Km. (D) PGP 9.5-stained section showing an occasional epidermal nerve fiber (arrows) in the proximal thigh of a patient with chronic neuropathy.Axons = red; Schwann cells = green. Scale bar = 20 Km.
Ebenezer et al J Neuropathol Exp Neurol � Volume 66, Number 12, December 2007
� 2007 American Association of Neuropathologists, Inc.1064

7Copyright @ 200 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.
Increased incidence of impaired glucose tolerance isseen in neuropathy patients, particularly in those with painand sensory loss and is more common in patients withapparently idiopathic polyneuropathy than in the age-matched general population (61, 65). IENF density isreduced even in patients with neuropathy who have impairedglucose tolerance neuropathy with normal NCS (66). Theneuropathy associated with impaired glucose tolerance isgenerally milder than the neuropathy associated withdiabetes mellitus. Small nerve fibers are prominentlyaffected and may be the earliest detectable sign of neuro-pathy in glucose dysmetabolism (45, 50, 67, 68).
By using a human model of IENF regeneration, it hasbeen shown that reduced rates of nerve regeneration occur in
diabetic subjects without any evidence of neuropathy (29),providing a rationale to include subjects without overtneuropathy in trials of regenerative agents. Pancreas trans-plantation has not produced the expected major beneficialeffects on small nerve fibers, probably because of thereduced regeneration capacity of patients with advanceddiabetes. Severe depletion of epidermal and dermal fiberswas noted in both lower limbs during the post-transplantperiod (69). To register nerve fiber regeneration, prolongedobservation periods will probably be necessary, and IENFquantification can be used to assess the progress of poly-neuropathy during post-transplant follow up.
Although it is generally believed that diabetic neuro-pathy is due to chronic hyperglycemia, experience frompatients with insulinoma and experimental studies show thatperiods of hypoglycemia from overcontrol of glucose levelsmay also cause neuropathy in diabetic patients. The plantarnerves of diabetic eu-/hypoglycemic BB/Wor rats treatedwith insulin implants exhibited a distinct neuropathy, whichis accompanied by mild alterations in the epidermalinnervation of plantar skin and a more obviously abnormalnerve terminal pattern in plantar muscle (70).
Human Immunodeficiency Virus-AssociatedSensory Neuropathies
Various types of peripheral neuropathies have knownto be associated with human immunodeficiency virus (HIV)infection, and 10% to 35% of adults with AIDS are known todevelop HIV-associated sensory neuropathies (HIV-SN) (71,72). Two subtypes are manifest: one predominantly asso-ciated with HIV infection (HIV-associated distal sensoryneuropathy), and the other associated with specific neuro-toxic antiretroviral drugs (antiretroviral toxic neuropathy).IENF density is typically reduced in a length-dependentmanner in adult patients with HIV infection who have neuro-pathic signs independent of neuropathic symptoms (44).
Follow-up of HIV-infected medically asymptomatic,non-neuropathic individuals over a period of time has shownthat a lower leg IENF density, a higher cooling threshold,and a higher heat pain threshold for minimal pain werefound to be associated with a greater risk of transition tosymptomatic HIV-associated distal sensory neuropathy.There is a longitudinal association between leg IENF andquantitative sensory testing, and serial small fiber evaluationin chronic neuropathies can be useful in predicting patientsat risk for transition to HIV-associated distal sensoryneuropathy (73, 74). Decreased IENF density was associatedwith higher levels of neuropathic pain, higher plasma HIVRNA levels, and lower CD4 counts (54).
In subjects with advanced HIV, IENF density corre-lates with severity of neuropathy severity and with the levelof neuropathic pain quantified by both the Gracely PainScale and the Visual Analogue Scale. IENF density at thedistal leg also correlates with sural amplitude, but IENF atboth distal leg and proximal thigh correlated negatively withtoe cooling detection threshold. In contrast to the study byPolydefkis et al (54), IENF density from patients receivinghighly active antiretroviral therapy was not associated withexposure to neurotoxic antiretroviral drugs, CD4 cell count,
FIGURE 5. Light and electron microscopic findings of cuta-neous nerves in chronic neuropathy patients. (A) PGP 9.5-stained skin section showing dermal axons with globularaxonal swellings bar = 50 Km. (B) Remak
both Schwann cell and axoplasm (Ax) containing abnormalmitochondria (broken arrows). Scale bar = 1 Km.
J Neuropathol Exp Neurol � Volume 66, Number 12, December 2007 Assessment of Epidermal Nerve Fibers
� 2007 American Association of Neuropathologists, Inc. 1065
bundle containing denervated Schwann cell (Sch, arrow) andScale(arrows).

7Copyright @ 200 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.
or plasma log HIV-RNA level. These findings suggest that inthis group of patients with advanced HIV, the abovepreviously established risk factors for HIV and HIV-SN areno longer determinants of the severity of HIV-SN in thecurrent highly active antiretroviral therapy era (74).
In a prospective cohort study HIV-infected patientswere examined at 2 different geographic locations toemphasize demographic differences. Reduced vibrationthresholds and epidermal nerve fiber densities had thehighest diagnostic efficiency of the laboratory indicators ofneuropathy examined, but were found to be relativelyinsensitive in isolation (75). IENF density in longitudinalstudies can be used to identify risk factors for neuropathyprogression and to monitor epidermal nerve fiber regener-ation in clinical trials of regenerative agents.
Sensory neuropathy also occurs in a significantproportion of HIV-infected pediatric patients (76, 77).Twenty-eight HIV-seropositive patients aged 5.6 to 18.2years were compared with 48 HIV-seronegative subjectsaged 6.5 to 17.8 years. NCS and quantitative sensory testingdid not significantly differ between patients and controlsubjects, but HIV-seropositive patients as a group showed ahigher IENF density than control subjects. This paradoxicalincrease in innervation indicates age-specific differences inthe expression of HIV-SN and the difficulty in establishingthe presence of peripheral neuropathy in the pediatric andadolescent group (C. Luciano, unpublished observation). Re-generation studies using human models of collateral (excisionmodel) and regenerative (capsaicin model) sprouting showedreduced rates of epidermal nerve fiber regeneration. Thisabnormality was more pronounced in those with evidence ofbaseline neuropathy but was also observed in those withnormal peripheral nerve function and no neuropathy symp-toms. The rate of regeneration was highly dependent upon thebaseline distal thigh IENF density but was independent ofCD4 cell count, HIV viral load, HIV duration, gender, race, orapolipoprotein E4 status. Regeneration was not furtherreduced in those subjects exposed to dideoxynucleosidemedications. Swellings were not a prominent feature amongsubjects in that study, suggesting that reductions in regener-ation may be the earliest change that occurs. These findingsalso imply that there is a progression from the introduction ofan insult such as infection with HIV to development of areduced regeneration rate followed by morphologic changesand finally the establishment of peripheral neuropathy. In thiscontext, an improvement in regeneration would be the earlieststep toward the improvement of nerve function in the settingof a regenerative neuropathy study (78).
To date, no agents have been shown to be effective inspeeding neuronal regeneration in humans although atherapeutic agent, acetyl-L-carnitine, which is vital formitochondrial dysfunction, has been reported to enhanceepidermal innervation in HIV-SN (79). A trial of recombi-nant human erythropoietin is now underway because of itspresumed neuroprotective properties (80, 81).
Idiopathic Small Fiber Sensory NeuropathyPatients with a clinical diagnosis of possible idiopathic
small fiber neuropathy often present with painful burning
feet, but on clinical examination show normal strength,proprioception, tendon reflexes, and normal electrophysio-logic responses. These patients often have a reduced IENFdensity at the calf with normal proximal nerve fiberdensities, indicating that idiopathic small fiber neuropathyis typically a length-dependent process (19, 82). Skin biop-sies can be useful to distinguish neuropathies from radicu-lopathies, which typically do not produce epidermal nervechanges because the damage occurs proximal to the dorsalroot ganglion. Follow-up skin biopsies of patients withidiopathic small fiber sensory neuropathy with durationsranging from 12 to 28 months show a decrease of IENFdensity in the leg, indicating the progressive destruction ofthese fibers in the disease (42).
Small and large fibers are not evenly distributed withincutaneous nerves, and this inconsistency increases thecomplexity of morphometric quantification of nerve fibersin biopsies such as those of the sural nerve. One studycomparing sural nerve biopsy to skin biopsy demonstratedthat a reduction in IENF density was the only evidence forsmall fiber neuropathy in 23% of patients (55). In 1 largestudy skin biopsy was found to be more sensitive than aquantitative sudomotor axon reflex test or quantitativesensory testing in diagnosing small fiber neuropathy (23).In another study with painful feet of unknown cause, anexcellent correlation was observed between results of thequantitative sudomotor axon reflex test, cooling abnormal-ities, and loss of IENFs (83). Although sural sensory nerveaction potentials have served as the traditional electro-physiologic marker of distal sensory polyneuropathy, inpatients with suspected distal sensory polyneuropathy whohave normal NCS measuring both medial plantar sural nerveaction potentials and small sensory fibers will serve ascomplementary tools in evaluation of distal SN (23, 84, 85).
LeprosyLeprosy is a chronic granulomatous disorder affecting
multiple peripheral nerves and skin. Cutaneous nerve fibersare affected in all types of leprosy (86, 87), resulting inanesthetic or hypoesthetic skin lesions. In a large multi-centric cohort study in multibacillary patients, unmyelinatedC-fibers were found to be more frequently affected thansmall myelinated AC fibers and NCS, and warm and coldthreshold measurements have emerged as the most promis-ing tools for early detection of leprosy (88). One of themajor challenges in leprosy control programs is to diagnoseindeterminate skin lesions, relapses, and dermatologic con-ditions that may mimic leprosy. Although no attempt hasbeen made to quantify epidermal nerve fibers in leprosy skinlesions, a large study evaluated cutaneous innervation andneuropeptides using immunohistochemical methods. Neuro-peptide immunoreactivity was seen in only 14% of theindeterminate leprosy specimens and was completely absentin other types of leprosy, highlighting the diagnosticsignificance of early disappearance of neuropeptide immu-noreactivity in leprosy lesions (89). Because the 3-mmpunch technique is simple to perform, nontechnical staff canbe trained in the quantification technique, and IENF densityof skin lesions could serve as an additional tool along with
Ebenezer et al J Neuropathol Exp Neurol � Volume 66, Number 12, December 2007
� 2007 American Association of Neuropathologists, Inc.1066

7Copyright @ 200 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.
skin smear examination and NCS to increase the diagnosticefficiency of detecting neuropathy in leprosy controlprograms.
Chemotherapy-Induced Peripheral NeuropathyChemotherapeutic agents produce both acute neuro-
toxicity and chronic neuropathy (90, 91). A number ofeffective chemotherapeutic agents, including platinumcompounds, taxanes, and vinca alkaloids produce length-dependent axonal injury and even worsen preexisting neu-ropathy (92). In an experimental rat model of paclitaxel- andvincristine-evoked painful peripheral neuropathies, a partialdegeneration of the sensory innervation of the epidermisand upregulation of PGP 9.5 in epidermal Langerhans cellshave been demonstrated in glabrous skin of the plantarhind paw (93). IENF density with total neuropathy scorealong the course of chemotherapy could help to predict thecourse and severity of chemotherapy-induced peripheralneuropathy.
Pyridoxine-Induced ToxicityExcess pyridoxine (vitamin B6) ingestion causes a
dose-dependent peripheral neuropathy characterized bynecrosis of dorsal root ganglion and degeneration of longmyelinated fibers in the sciatic nerve. In experimental rats aswell as in human subjects lower doses of pyridoxine alsoresult in development of small fiber involvement as theearliest and predominant abnormality, and higher doses leadto involvement of both large and small fibers (94Y96). Wehave observed 1 patient who developed a sensory neuro-pathy after consuming excessive amounts of pyridoxine.Serial skin biopsies demonstrated epidermal denervation thatrecovered over a period of several months (V. Choudhry,personal communication, 2007).
SKIN BIOPSY IN NON-LENGTH-DEPENDENTNEUROPATHIES
Sensory GanglionopathiesIn patients with sensory ganglion degeneration,
symptoms of ataxia and proprioceptive sensory losspredominate and the distribution of sensory impairment iswidespread and asymmetric (97Y99). A study comparedbiopsies from patients with ganglionopathy and axonalneuropathy with those from normal control subjects. Skinbiopsies at the proximal thigh and the distal leg in patientswith axonal neuropathies showed significantly lower valuesat the distal site of the leg, confirming the length-dependentloss of cutaneous innervation. In the patients with ganglion-opathy, in contrast, the degeneration of small-diametersensory fibers was seen equally in all locations (46). Itshould be noted that a severe length-dependent neuropathywill also show denervation at all sites and cannot be clearlydifferentiated from ganglionopathy on a neuropathologicbasis alone.
Postherpetic NeuralgiaHerpes zoster is caused by reactivation of latent
varicella zoster virus within the sensory ganglia, resulting
in destruction and degeneration of the respective spinal andperipheral axons (100). Several pathologic processes for thecause of postherpetic neuralgic pain have been suggested.IENF was examined in chronic postherpetic neuralgia-affected skin, and the loss of cutaneous innervation inverselycorrelated with allodynia. It is thought that survivingcutaneous primary afferent nociceptors spontaneouslybecame active and sensitized and contributed to postherpeticneuralgic pain and allodynia (101). Some have suggestedthat degeneration of primary afferent neurons causes centralhyperactivity (102). These studies expressed epidermaldensity as an average number of epidermal neurites/mm2
of skin surface area (101), a method of counting that isgenerally not used for other neuropathies.
SKIN BIOPSY IN INFLAMMATORYDEMYELINATING NEUROPATHIES
Guillain-Barre SyndromeGuillain-Barre Syndrome (GBS) or acute inflamma-
tory demyelinating polyneuropathy is an acute inflammatoryneuropathy, and, according to various clinical, neurophysio-logic, and pathologic studies large-diameter myelinatednerves are affected. Axonal degeneration of motor andsensory nerves has been described in the demyelinatingform of GBS (103). Skin biopsy from the distal leg ofpatients with the demyelinating form of GBS was inves-tigated for the involvement of small sensory fibers. IENFdensity was reduced in 55% of the cases in comparison withage- and gender-matched controls, and the values correlatedwith functional disabilities. These findings suggest that GBSis not a purely large-fiber neuropathy but that small-fibersensory and autonomic neuropathies exist in a significantproportion of patients with GBS (104).
Chronic Inflammatory DemyelinatingPolyneuropathy
When IENF densities and thermal thresholds inpatients with chronic inflammatory demyelinating polyneu-ropathy were tested, patients with chronic inflammatorydemyelinating polyneuropathy showed lower IENF densitythan that of control subjects. The low IENF density andelevated thermal threshold were associated with autonomicsymptoms. Patients with chronic inflammatory demyelinat-ing polyneuropathy have small-fiber sensory and autonomicneuropathies in addition to the immunologic inflammation inlarge diameter myelinated fibers (105).
SarcoidosisSarcoidosis is a chronic progressive granulomatous
multisystem disorder that may affect virtually any part of thenervous system, and patients can present with peripheralnerve pain and fatigue (106, 107). Patients with sarcoidosiswith normal nerve conduction studies had shown reducedtemperature sensitivity and low IENF in the leg comparedwith control subjects. Small fiber sensory neuropathy occursin sarcoidosis, and reduced IENF may be the only detectableabnormality in patients with sarcoidosis presenting withperipheral nerve pain (53).
J Neuropathol Exp Neurol � Volume 66, Number 12, December 2007 Assessment of Epidermal Nerve Fibers
� 2007 American Association of Neuropathologists, Inc. 1067

7Copyright @ 200 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.
Hepatitis C InfectionThe most frequent form of hepatitis C infection
(HCV)-associated neuropathy is distal sensory or sensori-motor polyneuropathy (108). Most cases occur in associationwith cryoglobulinemia (109) and vasculitis (110). A multi-centric study in an unselected, untreated referral populationwith HCV infection has shown that electrophysiologicperipheral neuropathy occurs in 15.3% of patients (111).Coinfection with HIV and HCV has become increasinglyprevalent and distal symmetric polyneuropathy is the mostcommon form of peripheral neuropathy in HIV and HCVinfections. However, in a cohort study of 147 HIV-infectedadults, HCV was not found to be significantly associatedwith HIV-related sensory neuropathies (75).
Vasculitic NeuropathyVasculitis is one of the rare causes of peripheral
neuropathy, and clinical manifestations include motor weak-ness, reduced sensitivity, and neuropathic pain. Smallmyelinated nerves were shown to be more susceptible toischemia than larger myelinated fibers in experimentallyinduced ischemia in Wistar rats (112). Patients with motorand sensory impairments have shown significant IENFreduction, demonstrating that the depletion and degenerationof small-diameter sensory nerves in vasculitis occur asfrequently as those of large-diameter nerves (113, 114).
SKIN BIOPSY IN AUTOIMMUNE DISEASESSkin biopsies have played an important role in
identifying small fiber neuropathy in chronic inflammatoryautoimmune diseases.
Sjogren SyndromeSensory neuropathy with prominent ataxia and gan-
glionitis are well-recognized forms of neuropathy associatedwith Sjogren syndrome. Sural biopsies show a wide range inloss of both large and small myelinated and unmyelinatedaxons (98, 115). A study of 20 consecutive patients withSjogren neuropathy showed that 60% of the subjects hadnonpatchy symptoms and a non-length-dependent pattern ofepidermal nerve fiber loss, suggesting that patients with thisdisorder commonly have a small-fiber sensory neuronopathyrather than a Bdying-back[ axonopathy (116). This hypoth-esis has been further confirmed in a population-based studyin a small group of patients (117).
Systemic Lupus ErythematosusThe prevalence of peripheral neuropathy in systemic
lupus erythematosus (SLE) varies from 5% to 27%, and it ischaracterized by a mild sensory or sensorimotor neuropathy(47, 118, 119). Pure small-diameter nerve fiber neuropathyoccurs in SLE with a significant reduction in IENF density.Disease activity, disease duration, or routine hematologic,biochemical, or immunologic variables did not influence thenumber of IENFs in patients with SLE (36, 120).
The degree of loss of small-diameter nerve fibersamong patients with these chronic inflammatory autoim-mune diseases varies. Sixty patients with SLE, 61 patientswith primary Sjogren syndrome, and 52 patients with
rheumatoid arthritis were compared with 106 healthysubjects. Densities were significantly less in patients withSLE compared with patients with rheumatoid arthritis orhealthy subjects. Nerve fiber densities were also reduced inpatients with primary Sjogren syndrome compared withhealthy subjects. Only 8 patients (13%) with SLE, 2 patients(3%) with primary Sjogren syndrome, and 2 patients (4%)with rheumatoid arthritis have shown densities below thelower reference limit of 3.4 fibers/mm (121). Despite thelower average mean values, very few patients actually meeta diagnosis of small fiber neuropathy.
Celiac DiseaseCeliac disease is an autoimmune disease of the small
intestine and the gastrointestinal symptoms result fromingestion of gluten in genetically susceptible people. Neuro-logic complications are estimated to occur in 10% ofaffected patients (122), with sensory neuropathy being oneof the most frequently reported neurologic abnormalities(123). Despite the association between celiac disease andsmall fiber neuropathy, a definitive causal relationship hasnot been established. Interestingly, in patients presentingwith asymmetric numbness and paresthesias, low IENFdensities occur at the thigh, forearm, and distal leg,suggesting a widespread involvement of small fibers (124).Detection of anti-gliadin antibodies can be useful indiagnosis (125).
Friedreich AtaxiaThis is an autosomal recessive cardioneurodegener-
ative disorder resulting from an inability to produce theprotein frataxin. It is due to extensive trinucleotide repeatexpansions in the first intron of the gene encoding frataxin,resulting in defective transcriptions and protein deficiency(126). Patients with Friedreich ataxia have been evaluated byskin biopsy and have shown significant loss of epidermalfibers, reduced autonomic innervation to sweat glands,arrector pilorum muscles, and arterioles in parallel withtheir clinical findings of impaired thermal sensitivity, tactilethresholds, and decreased mechanical pain detection (127).
Fabry DiseaseFabry disease is an X-linked recessive disorder caused
by deficiency of >-galactosidase that results in accumulationof neutral glycosphingolipids (principally ceramide trihexo-side) primarily in endothelial cells (128) of blood-brain/nerve barriers within the central or peripheral nervoussystems (129). In the peripheral nervous system it producesa painful small fiber neuropathy (130Y132). Patients withFabry disease may have reduced IENF density at distalbiopsy sites with relatively preserved proximal nerve fiberdensity (133). Twenty-five heterozygous male subjects withFabry disease did not show an increase in epidermalinnervation density after >-galactosidase A (agalsidase->)enzyme replacement at 6 months. After an additional yearof therapy, there was a significant reduction in IENFdensity in the patient group as a whole, perhaps attributableto the declining glomerular filtration rate and the effects ofuremia on peripheral nerves. Thermal thresholds remained
Ebenezer et al J Neuropathol Exp Neurol � Volume 66, Number 12, December 2007
� 2007 American Association of Neuropathologists, Inc.1068

7Copyright @ 200 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.
unchanged. This study suggests that longer treatmentperiods may be needed for nerve fiber regeneration orthat enzyme replacement therapy alone is not sufficient toreverse the pathology of this disease (134).
Congenital Insensitivity to Pain With AnhydrosisCongenital insensitivity to pain with anhydrosis or
hereditary sensory neuropathy type IV (135) is a very raredisorder and clinical manifestations include mental retarda-tion, congenital analgesia that results in self-mutilation,multiple scars, fractures, and anhidrosis (136). Ultrastruc-tural and morphometric studies of sural nerves show loss ofboth unmyelinated and small myelinated fibers (137, 138).Skin biopsy examination in a 10-year-old girl has shownextensive loss of epidermal nerve fibers and denervation ofdermal adnexal structures including sweat glands andarrector pili (138).
Abnormalities in Dermal InnervationGlabrous skin is richly innervated by myelinated fibers
in the dermis and morphologically these fibers appearsimilar to the myelinated fibers in sciatic and sural nerves.Quantification of myelinated fibers and mechanoreceptors inglabrous skin in normal subjects has shown that thecutaneous segments of AA fibers undergo repeated branch-ing and shortening of internodal length during their coursefrom the nerve trunk (139).
Charcot-Marie-Tooth disease is caused by mutationsin myelin proteins and evaluation of myelinated dermalfibers in skin has revealed pathologic abnormalities inaxolemmal molecular architecture (140, 141). Patients withanti-myelin-associated glycoprotein neuropathy have shownIgM deposits on dermal myelinated fibers with a greaterprevalence at the distal site of extremities. These findingshighlight the use of glabrous skin in identifying abnormal-ities in inherited demyelinating and chronic demyelinatingneuropathies (142).
Many studies have shown reduced sweat gland inner-vation in patients with peripheral neuropathy (20, 22, 105,128, 139, 143). A few researchers have tried differentmethods to quantify and measure sweat gland innervation.Decreased nerve fiber length around sweat glands in diabeticpatients (52), reduced innervation in patients with Rosssyndrome (144, 145), and familial dysautonomia (146) havebeen demonstrated. Larger studies with validation of thequantification methods in both healthy control subjects andin diseased conditions are needed for quantification ofdermal innervation in sweat glands.
USE OF SKIN BIOPSIES IN HUMANREGENERATION STUDIES OF SENSORY NERVES
As the epidermis is highly innervated by small sensorynerves, skin biopsies provide easy accessibility to study theeffect of chemical, thermal, and mechanical trauma on theregeneration and degeneration of sensory nerves. Two typesof nerve injury models have evolved: an Bincision model[ inwhich, following transection of subepidermal plexes, thesprouting epidermal axons outside the incision line completethe reinnervation by 30 to 75 days. In the Bexcision model[
removal of the incised cylinder of skin resulted in adenervated area, and no complete reinnervation of epidermiswas seen even after 23 months (147). The effect of atherapeutic compound, neuroimmunophilin ligand timcodardimesylate, on collateral nerve sprouting was measuredusing the excision model. Though the compound did notimprove collateral sprouting, the model was consistentacross treatment groups and had a low coefficient ofvariation (148).
Application of capsaicin, an ingredient of hot chillipepper, to the skin produces a reproducible chemicalaxotomy of the epidermal nerves. Capsaicin can be admin-istered intradermally or topically. Serial biopsies haveshown that nerve regeneration after topical capsaicin isfaster than after intradermal delivery (149, 150). ThisBcapsaicin model[ has been used to study the degenerative(150) and regenerative properties of epidermal fibers (29)after chemical axotomy. The rate of regeneration of IENFwas lower in diabetic subjects irrespective of the presence orabsence of neuropathy, indicating that abnormalities inperipheral nerve function are present early in diabetes, evenbefore signs or symptoms develop (29). In a randomized,double-blind, placebo-controlled trial, the effect of timcodardimesylate, a putative regenerative neurophilin compound,on the regeneration of epidermal fibers after capsaicin nerveinjury was investigated. There was no difference in theregeneration rate between the treatment and placebo arms,but the baseline IENF density and the height and race of theparticipants influenced the rate of regeneration (48). Thesemodels hold potential for use in clinical trials to study theeffect of regenerative drugs and growth factors on sensorynerve fibers.
CONCLUSIONSSkin biopsy analysis of nerve fibers is a technique that
has opened a new window to help visualize the previouslyinaccessible small unmyelinated fibers. Over the last 2decades there has been a dramatic increase in our under-standing of cutaneous innervation, leading to improveddiagnostic and therapeutic interventions. The EuropeanFederation of Neurological Societies Task Force has for-mulated guidelines for IENF quantification (32) and sum-marized the utility of the technique. Normative data fromhealthy subjects matched for age and gender and fromappropriate anatomical sites should be used to interpretneuropathy. Quality control measures must be followed at alllevels to ensure the optimal examination of skin biopsies.The laboratory techniques are not automated, and artifacts orerrors can potentially be introduced at the stage of section-ing, staining, or interpreting. Intra- and interobserver ratingsand quality control between laboratories should be periodi-cally assessed. Skin biopsies could become a critical tool forstudies of axon-Schwann cell interactions that are vital forthe survival and maintenance of axons. Our data suggest thatSchwann cells are lost during prolonged denervation andalso in chronic peripheral neuropathies (3). In neuroregener-ative trials it is vital to identify group of patients who retainSchwann cells and who are most likely to respond toregenerative drugs. Identifying sites with low or absent
J Neuropathol Exp Neurol � Volume 66, Number 12, December 2007 Assessment of Epidermal Nerve Fibers
� 2007 American Association of Neuropathologists, Inc. 1069

7Copyright @ 200 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.
IENF or with more normal IENF would help in identifyingpatients who would progress and/or benefit from therapeuticintervention.
ACKNOWLEDGMENTSThe authors thank Barbara Freeman and Christopher
Wright for expert technical assistance.
REFERENCES1. Julius D, Basbaum AI. Molecular mechanisms of nociception. Nature
2001;413:203Y102. Oaklander AL, Siegel SM. Cutaneous innervation: Form and function.
J Am Acad Dermatol 2005;53:1027Y373. Ebenezer GJ, McArthur JC, Thomas D. Denervation of skin in
neuropathies: The sequence of axonal and Schwann cell changes inskin biopsies. Brain 2007;130:2703Y14
4. Simone DA, Kajander KC. Responses of cutaneous A-fiber nocicep-tors to noxious cold. J Neurophysiol 1997;77:2049Y60
5. Olausson H, Lamarre Y, Backlund H, et al. Unmyelinated tactileafferents signal touch and project to insular cortex. Nat Neurosci 2002;5:900Y904
6. Zylka MJ, Rice FL, Anderson DJ. Topographically distinct epidermalnociceptive circuits revealed by axonal tracers targeted to Mrgprd.Neuron 2005;45:17Y25
7. Donadio V, Nolano M, Provitera V, et al. Skin sympathetic adrenergicinnervation: An immunofluorescence confocal study. Ann Neurol2006;59:376Y81
8. Kennedy WR, Wendelschafer-Crabb G, Brelje TC. Innervation andvasculature of human sweat glands: An immunohistochemistry-laserscanning confocal fluorescence microscopy study. J Neurosci 1994;14:6825Y33
9. Weihe E, Schutz B, Hartschuh W, Anlauf M, Schafer MK, Eiden LE.Coexpression of cholinergic and noradrenergic phenotypes in humanand nonhuman autonomic nervous system. J Comp Neurol 2005;492:370Y79
10. Ruocco I, Cuello AC, Parent A, Ribeiro-da-Silva A. Skin bloodvessels are simultaneously innervated by sensory, sympathetic, andparasympathetic fibers. J Comp Neurol 2002;448:323Y36
11. Bennett DL. Neurotrophic factors: Important regulators of nociceptivefunction. Neuroscientist 2001;7:13Y17
12. Mendell LM, Albers KM, Davis BM. Neurotrophins, nociceptors, andpain. Microsc Res Tech 1999;45:252Y61
13. Song XY, Zhou FH, Zhong JH, Wu LL, Zhou XF. Knockout ofp75NTR impairs re-myelination of injured sciatic nerve in mice. JNeurochem 2006;96:833Y42
14. Terenghi G. Peripheral nerve regeneration and neurotrophic factors. JAnat 1999;194(Pt 1):1Y14
15. Smeyne RJ, Klein R, Schnapp A, et al. Severe sensory andsympathetic neuropathies in mice carrying a disrupted Trk/NGFreceptor gene. Nature 1994;368:246Y49
16. Shadiack AM, Sun Y, Zigmond RE. Nerve growth factor antiseruminduces axotomy-like changes in neuropeptide expression in intactsympathetic and sensory neurons. J Neurosci 2001;21:363Y71
17. Dalsgaard CJ, Rydh M, Haegerstrand A. Cutaneous innervation in manvisualized with protein gene product 9.5 (PGP 9.5) antibodies.Histochemistry 1989;92:385Y90
18. Gibbons CH, Griffin JW, Polydefkis M, et al. The utility of skinbiopsy for prediction of progression in suspected small fiber neuro-pathy. Neurology 2006;66:256Y58
19. Holland NR, Stocks A, Hauer P, et al. Intraepidermal nerve fiberdensity in patients with painful sensory neuropathy. Neurology 1997;48:708Y11
20. Kennedy WR, Wendelschafer-Crabb G, Johnson T. Quantitation ofepidermal nerves in diabetic neuropathy. Neurology 1996;47:1042Y48
21. McArthur JC, Griffin JW. Another tool for the neurologist’s toolbox.Ann Neurol 2005;57:163Y67
22. McCarthy BG, Hsieh ST, Stocks A, et al. Cutaneous innervation insensory neuropathies: Evaluation by skin biopsy. Neurology 1995;45:1848Y55
23. Periquet MI, Novak V, Collins MP, et al. Painful sensory neuropathy:Prospective evaluation using skin biopsy. Neurology 1999;53:1641Y47
24. Kennedy WR, Nolano M, Wendelschafer-Crabb G, Johnson TL,Tamura E. A skin blister method to study epidermal nerves inperipheral nerve disease. Muscle Nerve 1999;22:360Y71
25. Wang L, Hilliges M, Jernberg T, Wiegleb-Edstrom D, Johansson O.Protein gene product 9.5-immunoreactive nerve fibres and cells inhuman skin. Cell Tissue Res 1990;261:25Y33
26. McArthur JC, Stocks EA, Hauer P, Cornblath DR, Griffin JW.Epidermal nerve fiber density: Normative reference range anddiagnostic efficiency. Arch Neurol 1998;55:1513Y20
27. Wilkinson KD, Lee KM, Deshpande S, Duerksen-Hughes P, Boss JM,Pohl J. The neuron-specific protein PGP 9.5 is a ubiquitin carboxyl-terminal hydrolase. Science 1989;246:670Y73
28. Lauria G, Borgna M, Morbin M, et al. Tubule and neurofilamentimmunoreactivity in human hairy skin: Markers for intraepidermalnerve fibers. Muscle Nerve 2004;30:310Y16
29. Polydefkis M, Hauer P, Sheth S, Sirdofsky M, Griffin JW, McArthurJC. The time course of epidermal nerve fibre regeneration: Studies innormal controls and in people with diabetes, with and withoutneuropathy. Brain 2004;127:1606Y15
30. Stocks EA, McArthur JC, Griffen JW, Mouton PR. An unbiasedmethod for estimation of total epidermal nerve fibre length. JNeurocytol 1996;25:637Y44
31. Kennedy WR, Wendelschafer-Crabb G, Polydefkis M, McArthur JC.Pathology and quantitation of cutaneous innervation. In: Dyck PJ,Thomas PK, eds. Peripheral Neuropathy. Philadelphia: Saunders,2005:869Y96
32. Lauria G, Cornblath DR, Johansson O, et al. EFNS guidelines on theuse of skin biopsy in the diagnosis of peripheral neuropathy. Eur JNeurol 2005;12:747Y58
33. Wilder-Smith EP, Chow A. Comparison of a simple methodfor quantitation of intraepidermal nerve fibres with a standardimage analysis method using hypothenar skin. J Neurol 2006;253:1011Y15
34. Chien HF, Tseng TJ, Lin WM, et al. Quantitative pathology ofcutaneous nerve terminal degeneration in the human skin. ActaNeuropathol (Berl) 2001;102:455Y61
35. Hilliges M, Johansson O. Comparative analysis of numerical estima-tion methods of epithelial nerve fibers using tissue sections. J PeripherNerv Syst 1999;4:53Y57
36. Omdal R, Mellgren SI, Goransson L, et al. Small nerve fiberinvolvement in systemic lupus erythematosus: A controlled study.Arthritis Rheum 2002;46:1228Y32
37. Umapathi T, Tan WL, Tan NC, Chan YH. Determinants of epidermalnerve fiber density in normal individuals. Muscle Nerve 2006;3:742Y46
38. Besne I, Descombes C, Breton L. Effect of age and anatomical site ondensity of sensory innervation in human epidermis. Arch Dermatol2002;138:1445Y50
39. Pittenger GL, Ray M, Burcus NI, McNulty P, Basta B, Vinik AI.Intraepidermal nerve fibers are indicators of small-fiber neuropathy inboth diabetic and nondiabetic patients. Diabetes Care 2004;27:1974Y79
40. Goransson LG, Mellgren SI, Lindal S, Omdal R. The effect of ageand gender on epidermal nerve fiber density. Neurology 2004;62:774Y77
41. Pan CL, Lin YH, Lin WM, Tai TY, Hsieh ST. Degeneration ofnociceptive nerve terminals in human peripheral neuropathy. Neuro-report 2001;12:787Y92
42. Lauria G, Morbin M, Lombardi R, et al. Axonal swellings predict thedegeneration of epidermal nerve fibers in painful neuropathies.Neurology 2003;61:631Y36
43. Shun CT, Chang YC, Wu HP, et al. Skin denervation in type 2diabetes: Correlations with diabetic duration and functional impair-ments. Brain 2004;127:1593Y1605
44. Herrmann DN, McDermott MP, Henderson D, Chen L, Akowuah K,Schifitto G. Epidermal nerve fiber density, axonal swellings and QSTas predictors of HIV distal sensory neuropathy. Muscle Nerve 2004;29:420Y27
45. Umapathi T, Tan WL, Loke SC, et al. Intraepidermal nerve fiber
Ebenezer et al J Neuropathol Exp Neurol � Volume 66, Number 12, December 2007
� 2007 American Association of Neuropathologists, Inc.1070

7Copyright @ 200 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.
density as a marker of early diabetic neuropathy. Muscle Nerve 2007;35:591Y98
46. Lauria G, Sghirlanzoni A, Lombardi R, Pareyson D. Epidermal nervefiber density in sensory ganglionopathies: Clinical and neurophysio-logic correlations. Muscle Nerve 2001;24:1034Y39
47. Omdal R, Henriksen OA, Mellgren SI, Husby G. Peripheralneuropathy in systemic lupus erythematosus. Neurology 1991;41:808Y11
48. Polydefkis M, Sirdofsky M, Hauer P, Petty BG, Murinson B,McArthur JC. Factors influencing nerve regeneration in a trial oftimcodar dimesylate. Neurology 2006;66:259Y61
49. Smith AG, Howard JR, Kroll R, et al. The reliability of skin biopsywith measurement of intraepidermal nerve fiber density. J Neurol Sci2005;228:65Y69
50. Sumner CJ, Sheth S, Griffin JW, Cornblath DR, Polydefkis M. Thespectrum of neuropathy in diabetes and impaired glucose tolerance.Neurology 2003;60:108Y11
51. Lauria G, Holland N, Hauer P, Cornblath DR, Griffin JW, McArthurJC. Epidermal innervation: Changes with aging, topographic location,and in sensory neuropathy. J Neurol Sci 1999;164:172Y78
52. Hirai A, Yasuda H, Joko M, Maeda T, Kikkawa R. Evaluation ofdiabetic neuropathy through the quantitation of cutaneous nerves. JNeurol Sci 2000;172:55Y62
53. Hoitsma E, Marziniak M, Faber CG, et al. Small fibre neuropathy insarcoidosis. Lancet 2002;359:2085Y86
54. Polydefkis M, Yiannoutsos CT, Cohen BA, et al. Reduced intra-epidermal nerve fiber density in HIV-associated sensory neuropathy.Neurology 2002;58:115Y19
55. Herrmann DN, Griffin JW, Hauer P, Cornblath DR, McArthur JC.Epidermal nerve fiber density and sural nerve morphometry inperipheral neuropathies. Neurology 1999;53:1634Y40
56. Kennedy WR, Wendelschafer-Crabb G. The innervation of humanepidermis. J Neurol Sci 1993;115:184Y90
57. Wendelschafer-Crabb G, Kennedy WR, Walk D. Morphologicalfeatures of nerves in skin biopsies. J Neurol Sci 2006;242:15Y21
58. Wild S, Roglic G, Green A, Sicree R, King H. Global prevalence ofdiabetes: Estimates for the year 2000 and projections for 2030.Diabetes Care 2004;27:1047Y53
59. Dyck PJ, Kratz KM, Karnes JL, et al. The prevalence by stagedseverity of various types of diabetic neuropathy, retinopathy, andnephropathy in a population-based cohort: The Rochester DiabeticNeuropathy Study. Neurology 1993;43:817Y24
60. Ziegler D, Gries FA, Spuler M, Lessmann F. The epidemiologyof diabetic neuropathy: Diabetic Cardiovascular Autonomic Neuro-pathy Multicenter Study Group. J Diabetes Complications 1992;6:49Y57
61. Lehtinen JM, Uusitupa M, Siitonen O, Pyorala K. Prevalence ofneuropathy in newly diagnosed NIDDM and nondiabetic controlsubjects. Diabetes 1989;38:1307Y13
62. Brownlee M. Biochemistry and molecular cell biology of diabeticcomplications. Nature 2001;414:813Y20
63. King RH; The role of glycation in the pathogenesis of diabeticpolyneuropathy; Mol Pathol 2001;54:400Y8
64. Sorensen L, Molyneaux L, Yue DK. The relationship among pain,sensory loss, and small nerve fibers in diabetes. Diabetes Care 2006;29:883Y87
65. Singleton JR, Feldman EL. Insulin-like growth factor-I in musclemetabolism and myotherapies. Neurobiol Dis 2001;8:541Y54
66. Smith AG, Ramachandran P, Tripp S, Singleton JR. Epidermal nerveinnervation in impaired glucose tolerance and diabetes-associatedneuropathy. Neurology 2001;57:1701Y4
67. Polydefkis M, Griffin JW, McArthur J. New insights into diabeticpolyneuropathy. JAMA 2003;290:1371Y76
68. Singleton JR, Smith AG. Therapy insight: Neurological complicationsof prediabetes. Nat Clin Pract Neurol 2006;2:276Y82
69. Boucek P, Havrdova T, Voska L, et al. Severe depletion ofintraepidermal nerve fibers in skin biopsies of pancreas transplantrecipients. Transplant Proc 2005;37:3574Y75
70. Mohseni S, Lillesaar C, Theodorsson E, Hildebrand C. Hypoglycaemicneuropathy: Occurrence of axon terminals in plantar skin and plantarmuscle of diabetic BB/Wor rats treated with insulin implants. ActaNeuropathol (Berl) 2000;99:257Y62
71. Cornblath DR, McArthur JC. Predominantly sensory neuropathy inpatients with AIDS and AIDS-related complex. Neurology 1988;38:794Y96
72. So YT, Holtzman DM, Abrams DI, Olney RK. Peripheral neuropathyassociated with acquired immunodeficiency syndrome: Prevalence andclinical features from a population-based survey. Arch Neurol 1988;45:945Y48
73. Herrmann DN, McDermott MP, Sowden JE, et al. Is skin biopsy apredictor of transition to symptomatic HIV neuropathy? A longitudinalstudy. Neurology 2006;66:857Y61
74. Zhou L, Kitch DW, Evans SR, et al. Correlates of epidermal nervefiber densities in HIV-associated distal sensory polyneuropathy.Neurology 2007;68:2113Y19
75. Cherry CL, Skolasky RL, Lal L, et al. Antiretroviral use and otherrisks for HIV-associated neuropathies in an international cohort.Neurology 2006;66:867Y73
76. Araujo AP, Nascimento OJ, Garcia OS. Distal sensory polyneuropathyin a cohort of HIV-infected children over five years of age. Pediatrics2000;106:e35
77. Floeter MK, Civitello LA, Everett CR, Dambrosia J, Luciano CA.Peripheral neuropathy in children with HIV infection. Neurology1997;49:207Y12
78. Hahn K, Triolo A, Hauer P, McArthur JC, Polydefkis M. Impairedreinnervation in HIV infection following experimental denervation.Neurology 2007;68:1251Y56
79. Hart AM, Wilson AD, Montovani C, et al. Acetyl-L-carnitine: Apathogenesis based treatment for HIV-associated antiretroviral toxicneuropathy. AIDS 2004;18:1549Y60
80. Hoke A, Keswani SC. Neuroprotection in the PNS: Erythropoietin andimmunophilin ligands. Ann NY Acad Sci 2005;1053:491Y501
81. Keswani SC, Leitz GJ, Hoke A. Erythropoietin is neuroprotective inmodels of HIV sensory neuropathy. Neurosci Lett 2004;371:102Y5
82. Holland NR, Crawford TO, Hauer P, Cornblath DR, Griffin JW,McArthur JC. Small-fiber sensory neuropathies: Clinical course andneuropathology of idiopathic cases. Ann Neurol 1998;44:47Y59
83. Novak V, Freimer ML, Kissel JT, et al. Autonomic impairment inpainful neuropathy. Neurology 2001;56:861Y68
84. Herrmann DN, Ferguson ML, Pannoni V, Barbano RL, Stanton M,Logigian EL. Plantar nerve AP and skin biopsy in sensory neuro-pathies with normal routine conduction studies. Neurology 2004;63:879Y85
85. Nodera H, Logigian EL, Herrmann DN. Class of nerve fiber involve-ment in sensory neuropathies: Clinical characterization and utilityof the plantar nerve action potential. Muscle Nerve 2002;26:212Y17
86. Chandi SM, Chacko CJ. An ultrastructural study of dermal nervesin early human leprosy. Int J Lepr Other Mycobact Dis 1987;55:515Y20
87. Job CK. Mechanism of nerve destruction in tuberculoid-borderlineleprosy: An electron-microscopic study. J Neurol Sci 1973;20:25Y38
88. Van Brakel WH, Nicholls PG, Das L, et al. The INFIR Cohort Study:Assessment of sensory and motor neuropathy in leprosy at baseline.Lepr Rev 2005;76:277Y95
89. Karanth SS, Springall DR, Lucas S, et al. Changes in nerves andneuropeptides in skin from 100 leprosy patients investigated byimmunocytochemistry. J Pathol 1989;157:15Y26
90. Lehky TJ, Leonard GD, Wilson RH, Grem JL, Floeter MK.Oxaliplatin-induced neurotoxicity: Acute hyperexcitability andchronic neuropathy. Muscle Nerve 2004;29:387Y92
91. Wilson RH, Lehky T, Thomas RR, Quinn MG, Floeter MK, Grem JL.Acute oxaliplatin-induced peripheral nerve hyperexcitability. J ClinOncol 2002;20:1767Y74
92. Chaudhry V, Chaudhry M, Crawford TO, Simmons-O’Brien E, GriffinJW. Toxic neuropathy in patients with pre-existing neuropathy.Neurology 2003;60:337Y40
93. Siau C, Xiao W, Bennett GJ. Paclitaxel- and vincristine-evokedpainful peripheral neuropathies: Loss of epidermal innervation andactivation of Langerhans cells. Exp Neurol 2006;201:507Y14
94. Berger AR, Schaumburg HH, Schroeder C, Apfel S, Reynolds R. Doseresponse, coasting, and differential fiber vulnerability in human toxicneuropathy: A prospective study of pyridoxine neurotoxicity. Neurol-ogy 1992;42:1367Y70
J Neuropathol Exp Neurol � Volume 66, Number 12, December 2007 Assessment of Epidermal Nerve Fibers
� 2007 American Association of Neuropathologists, Inc. 1071

7Copyright @ 200 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.
95. Callizot N, Warter JM, Poindron P. Pyridoxine-induced neuropathyin rats: A sensory neuropathy that responds to 4-methylcatechol.Neurobiol Dis 2001;8:626Y35
96. Perry TA, Weerasuriya A, Mouton PR, Holloway HW, Greig NH.Pyridoxine-induced toxicity in rats: A stereological quantification ofthe sensory neuropathy. Exp Neurol 2004;190:133Y44
97. Dalakas MC. Chronic idiopathic ataxic neuropathy. Ann Neurol 1986;19:545Y54
98. Griffin JW, Cornblath DR, Alexander E, et al. Ataxic sensoryneuropathy and dorsal root ganglionitis associated with Sjogren_ssyndrome. Ann Neurol 1990;27:304Y15
99. Lauria G, Pareyson D, Grisoli M, Sghirlanzoni A. Clinical andmagnetic resonance imaging findings in chronic sensory gangliono-pathies. Ann Neurol 2000;47:104Y9
100. Head H, Campbell AW, Kennedy PG. The pathology of herpes zosterand its bearing on sensory localization. Rev Med Virol 1997;7:131Y43
101. Rowbotham MC, Yosipovitch G, Connolly MK, Finlay D, Forde G,Fields HL. Cutaneous innervation density in the allodynic form ofpostherpetic neuralgia. Neurobiol Dis 1996;3:205Y14
102. Oaklander AL. The density of remaining nerve endings in human skinwith and without postherpetic neuralgia after shingles. Pain 2001;92:139Y45
103. Asbury AK, Arnason BG, Adams RD. The inflammatory lesion inidiopathic polyneuritis: Its role in pathogenesis. Medicine (Baltimore)1969;48:173Y215
104. Pan CL, Tseng TJ, Lin YH, Chiang MC, Lin WM, Hsieh ST.Cutaneous innervation in Guillain-Barre syndrome: Pathology andclinical correlations. Brain 2003;126:386Y97
105. Chiang MC, Lin YH, Pan CL, Tseng TJ, Lin WM, Hsieh ST.Cutaneous innervation in chronic inflammatory demyelinating poly-neuropathy. Neurology 2002;59:1094Y98
106. Newman LS, Rose CS, Maier LA. Sarcoidosis. N Engl J Med 1997;336:1224Y34
107. Sharma OP. Fatigue and sarcoidosis. Eur Respir J 1999;13:713Y14108. Tembl JI, Ferrer JM, Sevilla MT, Lago A, Mayordomo F, Vilchez JJ.
Neurologic complications associated with hepatitis C virus infection.Neurology 1999;53:861Y64
109. Nemni R, Sanvito L, Quattrini A, Santuccio G, Camerlingo M, CanalN. Peripheral neuropathy in hepatitis C virus infection with andwithout cryoglobulinaemia. J Neurol Neurosurg Psychiatry 2003;74:1267Y71
110. Cacoub P, Saadoun D, Limal N, Leger JM, Maisonobe T. Hepatitis Cvirus infection and mixed cryoglobulinaemia vasculitis: A review ofneurological complications. AIDS 2005;19( Suppl 3): S128Y34
111. Santoro L, Manganelli F, Briani C, et al. Prevalence and character-istics of peripheral neuropathy in hepatitis C virus population. J NeurolNeurosurg Psychiatry 2006;77:626Y29
112. Parry GJ, Brown MJ. Selective fiber vulnerability in acute ischemicneuropathy. Ann Neurol 1982;11:147Y54
113. Chao CC, Hsieh ST, Shun CT, Hsieh SC. Skin denervation andcutaneous vasculitis in eosinophilia-associated neuropathy. ArchNeurol 2007;64:959Y65
114. Lee JE, Shun CT, Hsieh SC, Hsieh ST. Skin denervation in vasculiticneuropathy. Arch Neurol 2005;62:1570Y73
115. Mori K, Iijima M, Sugiura M, et al. Sjogren_s syndrome associatedpainful sensory neuropathy without sensory ataxia. J Neurol NeurosurgPsychiatry 2003;74:1320Y22
116. Chai J, Herrmann DN, Stanton M, Barbano RL, Logigian EL. Painfulsmall-fiber neuropathy in Sjogren syndrome. Neurology 2005;65:925Y27
117. Goransson LG, Herigstad A, Tjensvoll AB, Harboe E, Mellgren SI,Omdal R. Peripheral neuropathy in primary Sjogren syndrome: Apopulation-based study. Arch Neurol 2006;63:1612Y15
118. Huynh C, Ho SL, Fong KY, Cheung RT, Mok CC, Lau CS. Peripheralneuropathy in systemic lupus erythematosus. J Clin Neurophysiol1999;16:164Y68
119. Omdal R, Loseth S, Torbergsen T, Koldingsnes W, Husby G, MellgrenSI. Peripheral neuropathy in systemic lupus erythematosusVAlongitudinal study. Acta Neurol Scand 2001;103:386Y91
120. Goransson LG, Tjensvoll AB, Herigstad A, Mellgren SI, Omdal R.Small-diameter nerve fiber neuropathy in systemic lupus erythemato-sus. Arch Neurol 2006;63:401Y4
121. Goransson LG, Brun JG, Harboe E, Mellgren SI, Omdal R. Intra-epidermal nerve fiber densities in chronic inflammatory autoimmunediseases. Arch Neurol 2006;63:1410Y13
122. Wills A, Hovell CJ. Neurological complications of enteric disease. Gut1996;39:501Y4
123. Chin RL, Sander HW, Brannagan TH, et al. Celiac neuropathy.Neurology 2003;60:1581Y85
124. Brannagan TH 3rd, Hays AP, Chin SS, et al. Small-fiber neuro-pathy/neuronopathy associated with celiac disease: Skin biopsyfindings. Arch Neurol 2005;62:1574Y78
125. Alaedini A, Okamoto H, Briani C, et al. Immune cross-reactivity inceliac disease: Anti-gliadin antibodies bind to neuronal synapsin I. JImmunol 2007;178:6590Y95
126. Bencze KZ, Kondapalli KC, Cook JD, et al. The structure and functionof frataxin. Crit Rev Biochem Mol Biol 2006;41:269Y91
127. Nolano M, Provitera V, Crisci C, et al. Small fibers involvement inFriedreich’s ataxia. Ann Neurol 2001;50:17Y25
128. Brady RO, Gal AE, Bradley RM, et al. Enzymatic defect in Fabry’sdisease: Ceramidetrihexosidase deficiency. N Engl J Med 1967;276:1163Y67
129. Kaye EM, Kolodny EH, Logigian EL, Ullman MD. Nervous systeminvolvement in Fabry’s disease: Clinicopathological and biochemicalcorrelation. Ann Neurol 1988;23:505Y9
130. Grunnet ML, Spilsbury PR. The central nervous system in Fabry’sdisease: An ultrastructural study. Arch Neurol 1973;28:231Y34
131. Kocen RS, Thomas PK. Peripheral nerve involvement in Fabry’sdisease. Arch Neurol 1970;22:81Y88
132. Onishi A, Dyck PJ. Loss of small peripheral sensory neurons inFabry disease: Histologic and morphometric evaluation of cutaneousnerves, spinal ganglia, and posterior columns. Arch Neurol 1974;31:120Y27
133. Scott LJ, Griffin JW, Luciano C, et al. Quantitative analysis ofepidermal innervation in Fabry disease. Neurology 1999;52:1249Y54
134. Schiffmann R, Hauer P, Freeman B, et al. Enzyme replacementtherapy and intraepidermal innervation density in Fabry disease.Muscle Nerve 2006;34:53Y56
135. Dyck PJ, Mellinger JF, Reagan TJ, et al. Not ’indifference to pain’ butvarieties of hereditary sensory and autonomic neuropathy. Brain 1983;106(Pt 2):373Y90
136. Rosemberg S, Marie SK, Kliemann S. Congenital insensitivity to painwith anhidrosis (hereditary sensory and autonomic neuropathy typeIV). Pediatr Neurol 1994;11:50Y56
137. Goebel HH, Veit S, Dyck PJ. Confirmation of virtual unmyelinatedfiber absence in hereditary sensory neuropathy type IV. J NeuropatholExp Neurol 1980;39:670Y75
138. Nolano M, Crisci C, Santoro L, et al. Absent innervation of skin andsweat glands in congenital insensitivity to pain with anhidrosis. ClinNeurophysiol 2000;111:1596Y1601
139. Nolano M, Provitera V, Crisci C, et al. Quantification of myelinatedendings and mechanoreceptors in human digital skin. Ann Neurol2003;54:197Y205
140. Li J, Bai Y, Ghandour K, et al. Skin biopsies in myelin-relatedneuropathies: Bringing molecular pathology to the bedside. Brain2005;128:1168Y77
141. Sabet A, Li J, Ghandour K, et al. Skin biopsies demonstrate MPZsplicing abnormalities in Charcot-Marie-Tooth neuropathy 1B. Neu-rology 2006;67:1141Y46
142. Lombardi R, Erne B, Lauria G, et al. IgM deposits on skin nerves inanti-myelin-associated glycoprotein neuropathy. Ann Neurol 2005;57:180Y87
143. Facer P, Mathur R, Pandya SS, Ladiwala U, Singhal BS, Anand P.Correlation of quantitative tests of nerve and target organ dysfunctionwith skin immunohistology in leprosy. Brain 1998;121( Pt 12):2239Y47
144. Nolano M, Provitera V, Perretti A, et al. Ross syndrome: A rare or amisknown disorder of thermoregulation? A skin innervation study on12 subjects. Brain 2006;129:2119Y31
145. Sommer C, Lindenlaub T, Zillikens D, Toyka KV, Naumann M.Selective loss of cholinergic sudomotor fibers causes anhidrosis inRoss syndrome. Ann Neurol 2002;52:247Y50
146. Hilz MJ, Axelrod FB, Bickel A, et al. Assessing function andpathology in familial dysautonomia: Assessment of temperature
Ebenezer et al J Neuropathol Exp Neurol � Volume 66, Number 12, December 2007
� 2007 American Association of Neuropathologists, Inc.1072

7Copyright @ 200 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.
perception, sweating and cutaneous innervation. Brain 2004;127:2090Y98
147. Rajan B, Polydefkis M, Hauer P, Griffin JW, McArthur JC. Epidermalreinnervation after intracutaneous axotomy in man. J Comp Neurol2003;457:24Y36
148. Hahn K, Sirdofsky M, Brown A, et al. Collateral sprouting of humanepidermal nerve fibers following intracutaneous axotomy. J PeripherNerv Syst 2006;11:142Y47
149. Nolano M, Simone DA, Wendelschafer-Crabb G, Johnson T, Hazen E,Kennedy WR. Topical capsaicin in humans: Parallel loss of epidermalnerve fibers and pain sensation. Pain 1999;81:135Y45
150. Simone DA, Nolano M, Johnson T, Wendelschafer-Crabb G,Kennedy WR. Intradermal injection of capsaicin in humansproduces degeneration and subsequent reinnervation of epidermalnerve fibers: Correlation with sensory function. J Neurosci 1998;18:8947Y59
J Neuropathol Exp Neurol � Volume 66, Number 12, December 2007 Assessment of Epidermal Nerve Fibers
� 2007 American Association of Neuropathologists, Inc. 1073