the blood-nerve barrier and reconstitution of the perineurium following nerve grafting
TRANSCRIPT

Neuropathology and Applied Neurobiology 1979, 5, 469-483
THE BLOOD-NERVE BARRIER AND RECONSTITUTION
O F THE PERINEURIUM FOLLOWING NERVE GRAFTING
A . M. AHMED AND R. 0. WELLER" Department of Pathology (Neuropathology), Southampton University Hospital, Southampton SO9 4XY
Accepted for publication 22 February 1979
Ahmed A.M. & Weller R.O. (1979) Neuropathology and Applied Neurobiology 5, 469483
The blood-nerve barrier and reconstitution of the perineurium following nerve grafting
Sural nerve autografts were performed on intact rat sural nerves and on sural nerves excised proximal to the site of grafting. The effect of the presence or absence of regenerating axons upon reconstitution of the perineurium at the graft junctions and upon re-establishment of the blood-nerve barrier to horseradish peroxidase (HRP) were studied over the succeeding 3-24 weeks. Compartmentation of the nerve fascicle occurred at the graft junctions where the perineurium was damaged. Each compartment contained Schwann cells with or without axons and was surrounded by elongated fibroblast-like cells which resembled perineurial cells in the longer surviving animals. It was concluded that, (a) compartments form in a nerve at the site of perineurial damage even in the absence of axons; (b) although compart- mentation may be a mechanism for perineurial regeneration and reconstitution of the blood-nerve barrier, blood vessels and the cell layers forming compartments at graft junctions remain permeable to HRP for at least 6 months; and (c) the intact perineurium around the distal stump of a denervated nerve is permeable to HRP but the endoneurial blood vessels are not.
Introduction
The perineurium in normal peripheral nerves has been shown to act as a diffusion barrier between the epineurium and the endoneurium (Waggener, Bunn & Beggs, 1965; Olsson, 1966; Kristensson& Olsson, 1971 ; Soderfeldt& Olsson, 1973). Large nerve fascicles are surrounded by several layers of flattened perineurial cells separated by bands of collagen. Many of the outer layers appear to be incomplete but the inner layers near the endoneurium form an efficient barrier to the diffusion of tracers such as horseradish peroxidase both from and into the endoneurium (Klemm, 1970; Olsson & Reese, 1971). The cells in this layer form tight junctions (zonulae occlu- * Address correspondence to: Professor R. 0. Weller, Neuropathology, Level E, South Pathology & Laboratory Block, General Hospital, Southampton SO9 4XY, England.
0305-1846/79/1200-0469 $02.00 0 1979 Blackwell Scientific Publications 469

470 A . M. Ahmed and R. 0. Weller
dentes) with their neighbours (Thomas & Jones, 1967; Real, Luciano, & Spitznas, 1975). In addition to the perineurial barrier around the nerve, there is a blood-nerve barrier to the passage of peroxidase and other tracers from the blood into the endo- neurium (Olsson & Reese, 1971; Kristensson & Olsson, 1971).
When a nerve is injured, as by a crush, the perineurium is damaged and the blood- nerve barrier is altered so that fluorescent and electron dense tracers leak into the endoneurial compartment at the site of damage (Olsson & Kristensson, 1973; de la Motte & Allt, 1976). During the ensuing phase of nerve sprouting and regeneration, the blood-nerve barrier remains permeable to fluorescent protein-bound tracers at the site of the crush (Olsson & Kristensson, 1973). Histological and ultrastructural studies have shown that the nerve is divided into compartments at the crushed site where the perineurium is damaged. Each compartment contains several regenerating fibres and is surrounded by flattened cells (Thomas & Jones, 1967; Morris, Hudson & Weddell, 1972; Jurecka, Ammerer & Lassmann, 1975).
The present report is of an investigation of the blood-nerve barrier and reconsti- tution of the perineurium following nerve grafting. In order to evaluate the influence of regenerating axons upon perineurial repair and the blood-nerve barrier, grafts have been applied to both intact nerves and to nerves which have been excised proximally.
Material and methods
Forty-eight adult male Wistar rats (100-150 g) were used. The animals were divided into three groups, viz: two experimental groups (forty-five animals) and one control group (three animals). All surgical procedures were performed with aseptic techniques under a dissecting microscope while the animals were under Nembutal anaesthesia (40 mg/kg body weight).
Experimental animals
Group 1-grafts on intact nerves (30 animals): The left sural nerve was exposed in the mid-calf region and a 5 mm length of nerve was removed. A 1 cm length of nerve was taken from the contralateral sural nerve and used as a graft (Figure 1A). The ends of the nerve were held in position by gel foam soaked in thrombin solution containing 1-2 unitslml and then treated with concentrated fibrinogen.
Proximal sural nerve
A B
Junctions (proximal 6- distal)
Figure 1. Diagrammatic illustration of the operative procedure. (A) Group 1: a graft is inserted into the centrally intact sural nerve. (B) Group 2: a similar graft is inserted into the proximally excised sural nerve.
Blood-nerve barrier and perineurium following nerve grafting 471
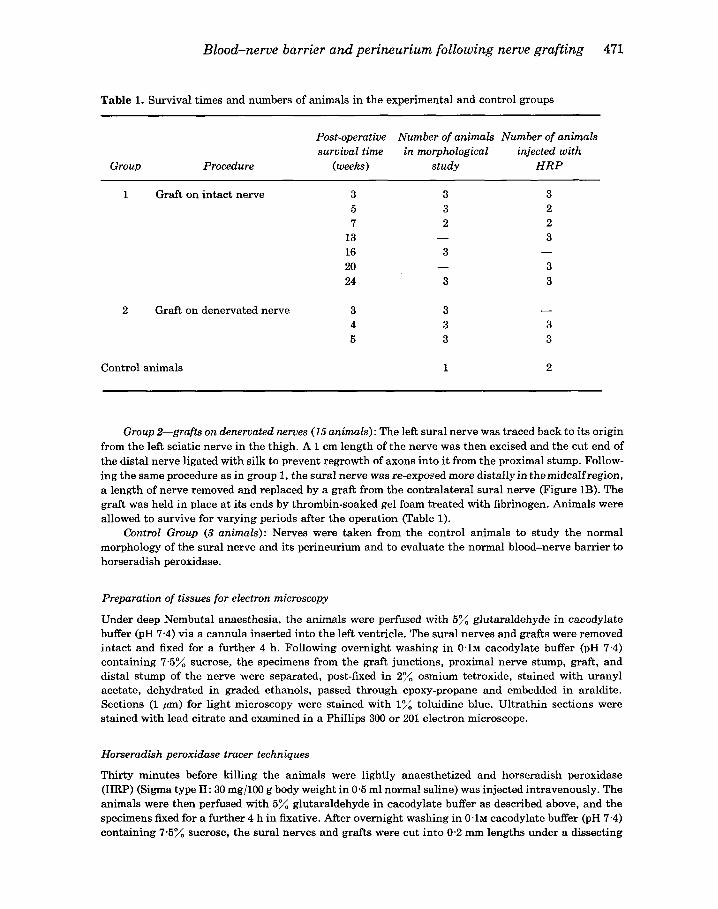
Table 1. Survival times and numbers of animals in the experimental and control groups
Post-operative Number of animals Number of animals survival time in morphological injected with
Group Procedure (weeks) study HRP
1 Graft on intact nerve 3 3 5 3 7 2
13 16 3 20 24 3
-
-
2 Graft on denervated nerve 3 3 4 3 5 3
Control animals 1
3 2 2 3
3 3
-
- 3 3
2
Group 2-grafts on denervated nerves (15 animals): The left sural nerve was traced back to its origin from the left sciatic nerve in the thigh. A 1 cm length of the nerve was then excised and the cut end of the distal nerve ligated with silk to prevent regrowth of axons into it from the proximal stump. Follow- ing the same procedure as in group 1, the sural nerve was re-exposed more distally in themidcalfregion, a length of nerve removed and replaced by a graft from the contralateral sural nerve (Figure 1B). The graft was held in place at its ends by thrombin-soaked gel foam treated with fibrinogen. Animals were allowed to survive for varying periods after the operation (Table 1).
Control Group (3 animals): Nerves were taken from the control animals to study the normal morphology of the sural nerve and its perineurium and to evaluate the normal blood-nerve barrier to horseradish peroxidase.
Preparation of tissues for electron microscopy
Under deep Nembutal anaesthesia, the animals were perfused with 5% glutaraldehyde in cacodylate buffer (pH 7.4) via a cannula inserted into the left ventricle. The sural nerves and grafts were removed intact and fixed for a further 4 h. Following overnight washing in 0 . 1 ~ cacodylate buffer (pH 7.4) containing 7.5% sucrose, the specimens from the graft junctions, proximal nerve stump, graft, and distal stump of the nerve were separated, post-fixed in 2% osmium tetroxide, stained with uranyl acetate, dehydrated in graded ethanols, passed through epoxy-propane and embedded in araldite. Sections (1 pm) for light microscopy were stained with 1% toluidine blue. Ultrathin sections were stained with lead citrate and examined in a Phillips 300 or 201 electron microscope.
Horseradish peroxidase tracer techniques
Thirty minutes before killing the animals were lightly anaesthetized and horseradish peroxidase (HRP) (Sigma type 11: 30 mg/l00 g body weight in 0.5 ml normal saline) was injected intravenously. The animals were then perfused with 5% glutaraldehyde in cacodylate buffer as described above, and the specimens fixed for a further 4 h in fixative. After overnight washing in 0 . 1 ~ cacodylate buffer (pH 7.4) containing 7.5% sucrose, the sural nerves and grafts were cut into 0.2 mm lengths under a dissecting
472 A . M . Ahmed and R. 0. Weller
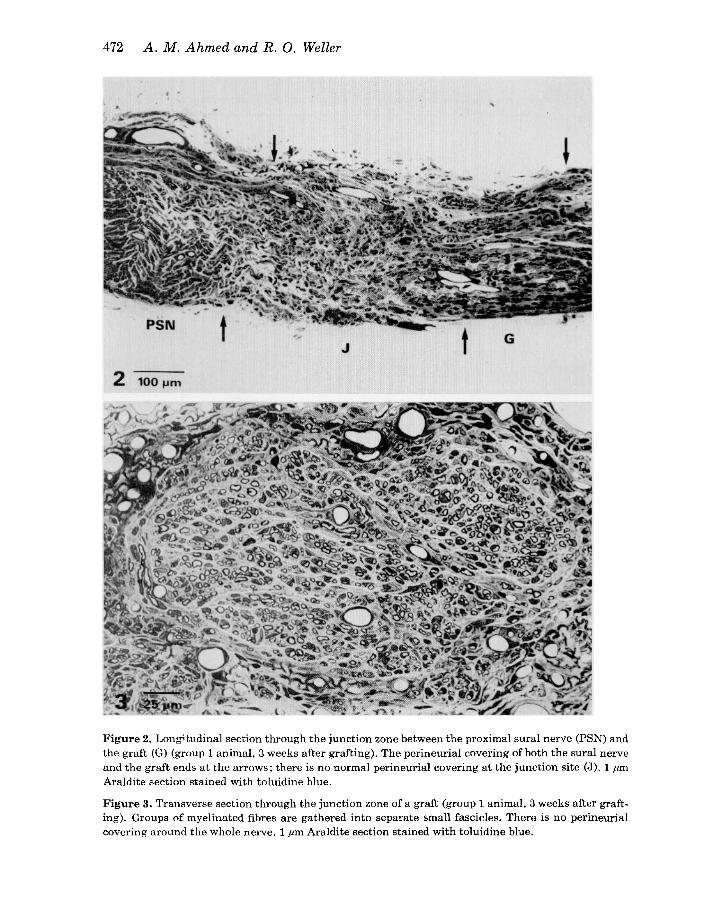
Figure 2. Longitudinal section through the junction zone between the proximal sural nerve (PSN) and the graft (G) (group 1 animal, 3 weeks after grafting). The perineurial covering of both the sural nerve and the graft ends at the arrows; there is no normal perineurial covering at the junction site (J)$ 1 pm Araldite section stained with toluidine blue.
Figure 3. Transverse section through the junction zone of a graft (group 1 animal, 3 weeks after graft- ing). Groups of myelinated fibres are gathered into separate small fascicles. There is no perineurial covering around the whole nerve. 1 pm Araldite section stained with toluidine blue.

Blood-nerve barrier and perineurium following nerve grafting 473
microscope and the pieces were incubated in diaminobenzidine (DAB) and hydrogen peroxide to localize the peroxidase activity (Graham & Karnovsky, 1966). The specimens of nerve were then processed and embedded in Araldite as described above, but the uranyl acetate staining was omitted. Sections (1 pm) were examined unstained or after staining with 1% toluidine blue. Ultrathin sections were examined unstained in Phillips 300 or 201 electron microscopes.
Control animals were injected with 0.5 ml of saline instead of horseradish peroxidase and lengths of nerve were incubated in DAB and hydrogen peroxide to localize endogenous peroxidase activity.
Results
The grafted nerves were examined after periods of 3-24 weeks (see Table 1).
G R O U P G GRAFTS ON I N T A C T NERVES
By 3 weeks, nerve sprouts had grown across the junction sites into nerve grafts and into the distal stumps. Examination of longitudinal sections of the proximal and distal nerve graft junction sites at this time and at 24 weeks revealed a deficiency in the perineurium (Figure 2). Between the severed ends of the perineurium, the nerve was divided into small fascicles or compartments each containing a few myelinated or non-myelinated fibres and their Schwann cells (Figure 3). The arrangements of the compartments is shown diagrammatically in Figure 4. Where the perineurium was intact in the proximal stump (Figure 4A) there were no compartments. An asymmetrical deficiency of the perineurium (Figure 4B) resulted in the formation of compartments on this side only. In the centre of the graft junction, where the peri- neurium was totally deficient around the periphery of the nerve, compartmentation extended throughout the whole nerve cross-section (Figure 4C). On approaching the intact perineurium of the graft, the central area of the nerve became devoid of com-
Junction
PSN G
A B C D E
Figure 4. Diagrammatic representation of the distribution of compartments in relation to perineurial deficiencies in group 1 animals. Proximal sural nerve (PSN), graft (G). For explanation see text.
474 A . M . Ahmed and R. 0. Weller
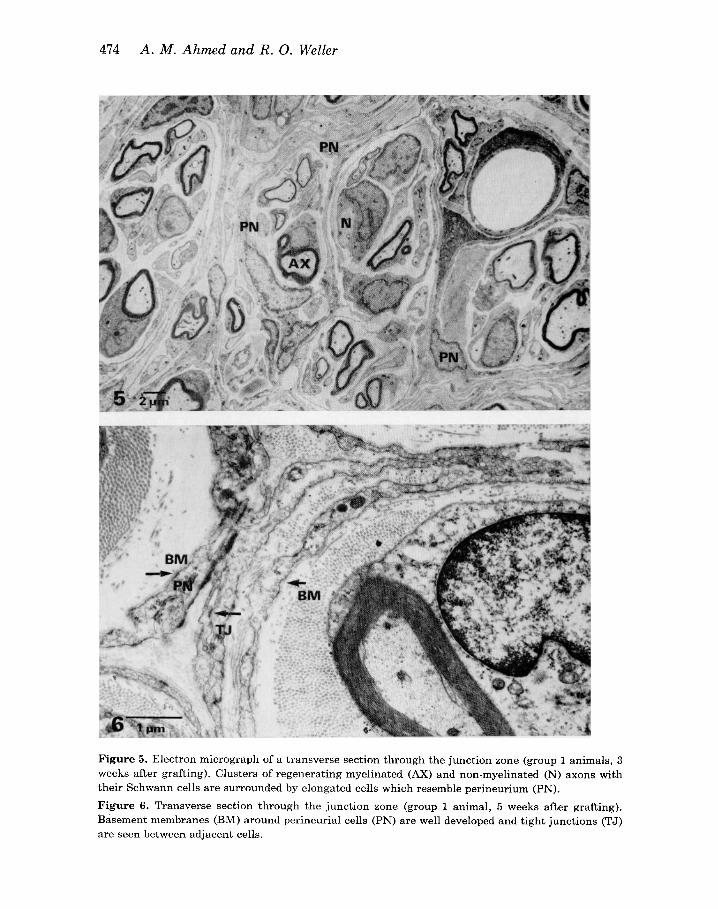
Figure 5 . Electron micrograph of a transverse section through the junction zone (group 1 animals, 3 weeks after grafting). Clusters of regenerating myelinated (AX) and non-myelinated (N) axons with their Schwann cells are surrounded by elongated cells which resemble perineurium (PN). Figure 6. Transverse section through the junction zone (group 1 animal, 5 weeks after grafting). Basement membranes (BM) around perineurial cells (PN) are well developed and tight junctions (TJ) are seen between adjacent cells.
Blood-nerve barrier and perineurium following nerve grafting 475
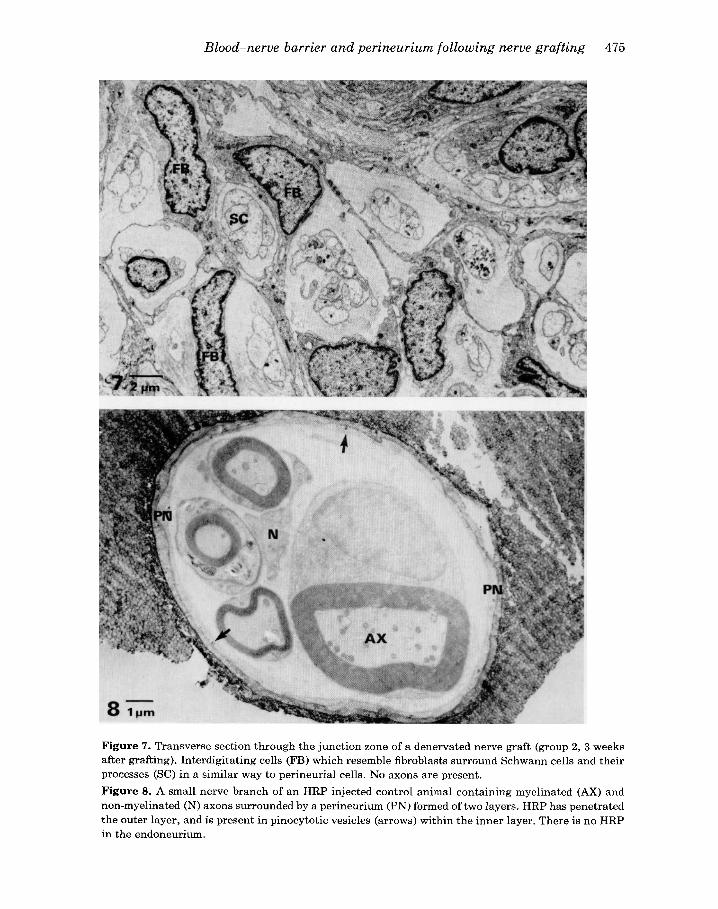
Figure 7. Transverse section through the junction zone of a denervated nerve graft (group 2, 3 weeks after grafting). Interdigitating cells (FB) which resemble fibroblasts surround Schwann cells and their processes (SC) in a similar way to perineurial cells. No axons are present. Figure 8. A small nerve branch of an HRP injected control animal containing myelinated (AX) and non-myelinated (N) axons surrounded by a perineurium (PN) formed of two layers. HRP has penetrated the outer layer, and is present in pinocytotic vesicles (arrows) within the inner layer. There is no HRP in the endoneurium.

476 A . M . Ahmed and R. 0. Weller
partments and the latter remained only around the outer edge of the nerve (Figure 4D). Compartments were not present where the perineurium of the nerve or the graft was intact (Figure 4E).
Electron microscope studies of junction sites at 3-24 weeks showed how the regenerating myelinated and non-myelinated axons were gathered into clusters enclosed in separate compartments (Figure 5). At 3 weeks after grafting, the cells forming the compartments resembled fibroblasts in their nuclear appearance, in their complement of cytoplasmic organelles, density of cytoplasmic staining and in their long tapering cell processes, but they differed from the fibroblasts of normal endo- neurium in their orientation where they surrounded clusters of axons and Schwann cells. Furthermore, even at this time, basement membrane covered much of the cell surfaces; by 5 weeks there were many pinocytotic vesicles in their cytoplasm and zonulae occludentes had formed between some of the cell processes (Figure 6). In these three features they resembled perineurial cells. The mixture of cell charac- teristics suggests that these perineurial cells may have developed from fibroblasts. Compartments differed in size and complexity and almost without exception they contained no blood vessels. Capillaries were present within the nerve, but they were situated between the compartments.
G R O U P 8 - G R A F T S O N D E N E R V A T E D N E R V E S
No regeneration of nerve fibres was observed in the distal stumps or grafts of any of the denervated sural nerves examined at 3-5 weeks. The perineurium around the distal stumps and the grafts remained morphologically intact, but it was disrupted at the graft junctions where the nerve trunk in all animals was divided into compart- ments each containing one or two Schwann cells or their processes (Figure 7). As in the regenerating intact sural nerve, the cells forming the compartments had many of the features of fibroblasts. The compartment walls were usually only one or two layers thick but junctional complexes between the cells were present. Many of the elongated cell processes lacked basement membrane and pinocytotic vesicles at 3 weeks after grafting. In these features the cells more closely resembled fibroblasts than perineurial cells. By 5 weeks, however, the elongated cells had acquired base- ment membranes and contained many pinocytotic vesicles and thereby resembled perineurial cells. Compartments in group 2 animals remained less well-formed when compared with compartments in group 1 animals.
B L O O D - N E R V E B A R R I E R T O H O R S E R A D I S H P E R O X I D A S E ( H R P ) .
N O R M A L N E R V E
In the normal sural nerve HRP was observed in epineurial connective tissue and in the outer layers of the perineurium, but it did not penetrate the inner layers of the perineurium and did not leak from the endoneurial vessels. Even in the very small branch in Figure 8 which contained only four myelinated fibres and a few non- myelinated axons there was no penetration of HRP into the endoneurium. There were
Blood-nerve barrier and perineurium following nerve grafting 477
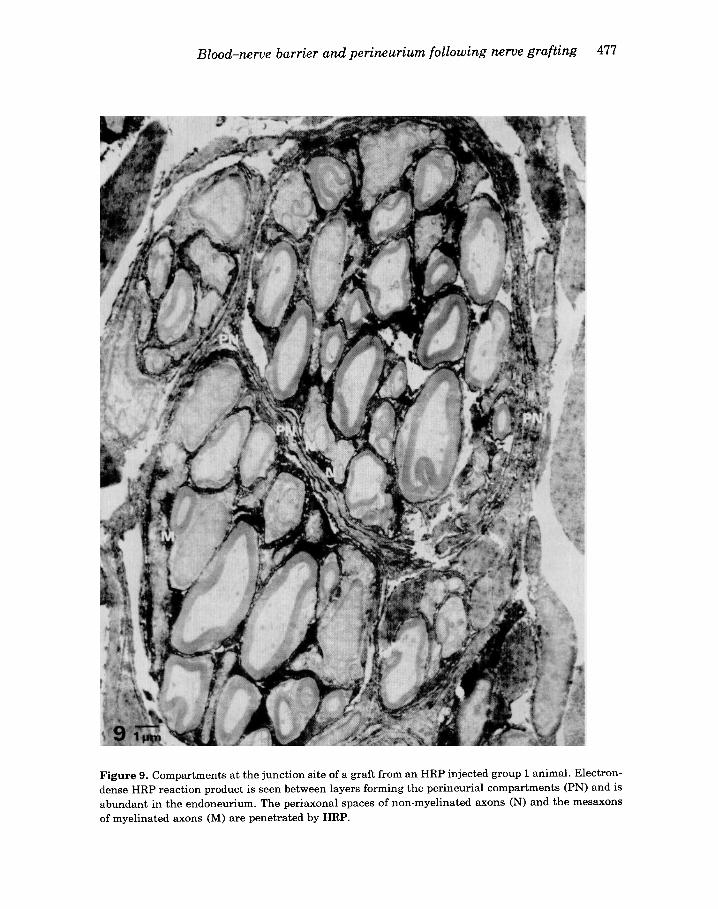
Figure 9. Compartments at the junction site of a graft from a n HRP injected group 1 animal. Electron- dense HRP reaction product is seen between layers forming the perineurial compartments (PN) and is abundant in the endoneurium. The periaxonal spaces of non-myelinated axons (N) and the mesaxons of myelinated axons (M) are penetrated by HRP.

478 A . M . Ahmed and R . 0. Weller
no blood vessels in this small nerve branch and it was surrounded by only two layers of perineurium. HRP had passed through the outer layer of the perineurium, but none was seen in the endoneurium. A few pinocytotic vesicles within the innermost layer of the perineurium did, however, contain HRP.
N E R V E GRAFTS
Group 1-grafts on intact nerves
At 3 weeks, no leakage of HRP was observed in regions of the proximal stump sur- rounded by normal perineurium, nor in the graft where the regenerating nerve fibres had penetrated and the perineurium was intact. Where the perineurium had been replaced by compartments a t the junctions, there was considerable leakage of HRP into the endoneurium in all animals from 3 to 24 weeks. HRP was present in the epineurium and in the endoneurium within the compartments. In animals with longer survival times, central areas of the graft junction site were free of compartments and showed no permeation by HRP. Two or three layers of flattened cells resembling perineurium surrounded each compartment and HRP was seen in the gaps between the cell layers and within pinocytotic vesicles in the cells (Figure 9). The endoneurium was heavily stained for HRP which had also penetrated the mesaxons and periaxonal spaces around non-myelinated axons. Pinocytotic vesicles containing HRP were seen not only in the perineurial cells but also in Schwann cells (Figure 10). Blood vessels between the compartments and in the surrounding epineurium appeared to be permeable to HRP. Small blood vessels adjacent to the compartments often had HRP within pinocytotic vesicles of endothelial cells and in the basement membrane (Figure 10). Intercellular junctions between the endothelial cells contained HRP which suggests that protein was leaking between the cells. Endogenous peroxidase activity within macrophages was also seen in graft material and in nerves without HRP.
Group 2-grafts on denervated nerves
HRP was found in the endoneurium within the compartments at the junction sites 3-5 weeks after grafting, as in the nerves in group 1. A patchy distribution of HRP was also observed in the endoneurium in the distal end of the denervated nerve and
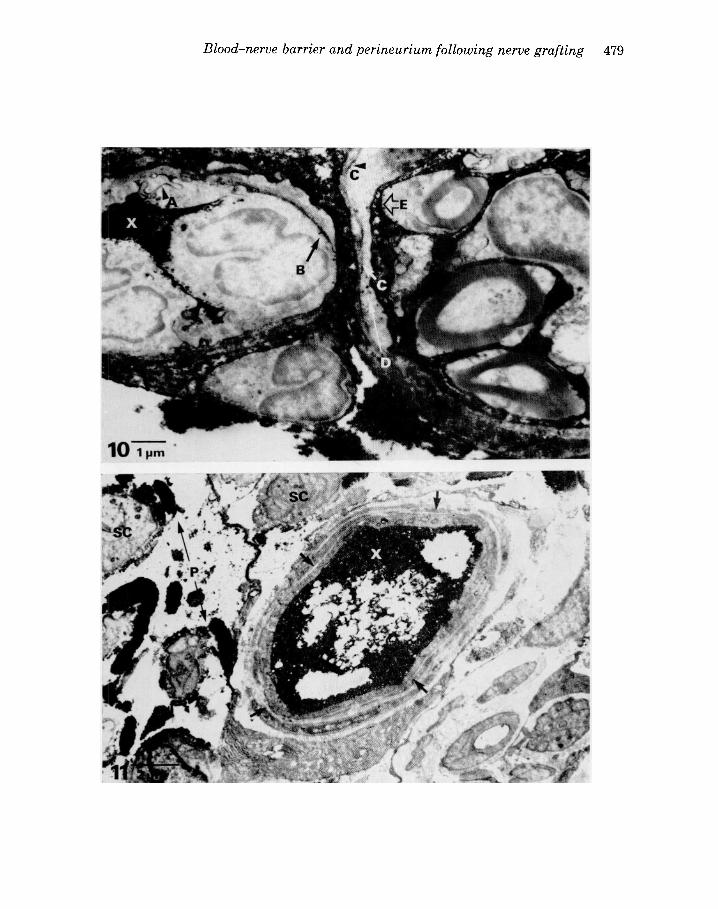
Figure 10. A group 1 animal injected with HRP showing a capillary adjacent to a compartment with reaction product in the lumen (X), between the endothelial cells (A) and in the basement membrane (B). HRP is also seen between layers of the compartment boundary cells (C) in pinocytotic vesicles within these cells (D) and within Schwann cells (E).
Figure 11. Transverse section of the distal stump of denervated nerve in an HRP injected group 2 animal. Reaction product is seen within the lumen of an endoneurial capillary (X). The basement mem- brane (short arrows) lacks HRP staining which suggests that the vessel is not leaking peroxidase. Patches of HRP reaction product are seen free within the endoneurium (P). Schwann cell processes (SC) in the endoneurium contain no axons.
Blood-nerve barrier and perineurium following nerve grafting 479

480 A . M . Ahmed and R . 0. Weller
in the graft at 4 weeks, even when the perineurium appeared structurally intact and at sites remote from direct trauma. HRP penetrated all layers of the perineurium but as no HRP was observed in capillary basement membrane (Figure ll), it probably did not leak from endoneurial blood vessels except at the junction zones.
Discussion
When nerve autografts are performed, the continuity of three major elements of the nerve is necessarily interrupted. Axons are divided, the perineurium is disrupted at the graft junctions and, inevitably, the endoneurial blood vessels are severed. In the present study, regeneration of axons was found to occur readily from the proximal stump of the intact sural nerve in the group 1 animals and by 3 weeks they had grown through the length of the grafts. Also by 3 weeks, the grafts had become revascular- ized and the blood-nerve barrier re-established; neither the intact perineurium nor the endoneurial blood vessels were permeable to peroxidase at this stage.
Damage to the perineurium resulted in the formation of compartments within the nerve. This phenomenon has been observed at the end of the proximal stump of severed nerves (Morris et al., 1972), at graft junction sites by Pollard & Fitzpatrick (1973) and in the present study. Thomas & Bhagat (1978) also described compartments in nerves where the perineurium had been disrupted by removal of the endoneurial contents from the nerve. Following grafting in our rats, compartments were formed by elongated fibroblast-like cells and contained four or five regenerating myelinated fibres together with groups of non-myelinated axons. Blood vessels were situated between compartments rather than within them, although some capillaries were partly surrounded by elongated cells. The formation of compartments may partly compensate for the absence of perineurium or may be a mechanism by which the perineurium regenerates. Although compartments may fuse eventually to form an intact perineurium, they were still present at the graft junctions 24 weeks after grafting, so it is possible that the perineurium in these circumstances never com- pletely regains its normal structure. No compartments formed in the middle of the nerve fascicles, except in the centre of the junction site; usually compartments were concentrated at the periphery of the fascicle throughout the length of the graft junction.
There has been much discussion upon the origin of perineurial cells. In the present study, the compartments appear to be formed by cells which are indistinguishable initially from fibroblasts; these cells later acquire the morphological characteristics of perineurial cells with the appearance of basement membrane and junctional com- plexes between the cells. By comparison the perineurium of the developing nerve forms as a layer around the whole fascicle and does not pass through a phase of compartment formation (Gamble & Breathnach, 1965). Some authors have suggested from the structure and function of the normal perineurium that its cells are derived from neural crest or are an extension of the leptomeninges (Masson, 1942; Benke & Rohlich, 1964; Andres, 1967; Shanthaveeruppa & Bourne, 1969). The observations in

Blood-nerve barrier and perineurium following nerve grafting 481
the present study agree with those of Gamble & Breathnach (1965) on developing perineurium and of Thomas & Bhagat (1978), and these authors suggest that peri- neurial cells can develop from fibroblasts. It is possible, however, that regenerating perineurial cells merely pass through a stage of differentiation where they resemble fibroblasts.
The stimulus for the formation of compartments was thought to be the regrowth of axons into the damaged area of nerve (Thomas & Bhagat, 1978), but we have found that compartments form around Schwann cells and their processes in the absence of regenerating axons. Migration of Schwann cells from the ends of severed nerves has been observed by Young, Holmes & Saunders (1940), by Rexed (1942), and by Thomas (1966). We found that Schwann cells migrate into the gaps at the graft junctions even when there is no axonal regeneration. Thus, in the grafts on to denervated nerves, Schwann cells filled the gap at the junctions and the compartments formed around them. They were not as well formed as in the intact nerve, but they did occur and this suggests that regeneration of axons is not the only stimulus for the formation of perineurial compartments.
Morris et al. (1972) suggested that the formation of compartments was a mechan- ism for rapidly reconstituting perineurium and blood-nerve barrier in damaged nerves. However, several studies using tracers have shown that re-establishment of the blood-nerve barrier is very slow at sites of perineurial damage (Olsson & Kristensson, 1973; de la Motte & Allt, 1976). Here it has been found that the vessels were quite separate from the compartments and were permeable to horseradish peroxidase which passed readily through junctions between the endothelial cells. In this characteristic they resembled epineurial vessels from which they were probably derived. HRP also passed through the layers of perineurial cells forming the compartments; in this characteristic they resembled the perineurium in developing mice where similarly there is no barrier to the passage of HRP (Kristensson & Olsson, 1971).
Although the perineurium around the regenerated nerve and its graft was im- permeable to horseradish peroxidase, the perineurium around the distal stump of the denervated nerves were permeable to it. The presence of axons is thus necessary for the re-establishment of the perineurial barrier.
The exact biological role of perineurial compartment formation is unclear. Although the compartments are permeable to HRP, they may play a role in regulating ion transport in a similar way to the control of calcium transport by normal peri- neurium (Ochs & Worth, 1978). Compartmentation, on the other hand, serves to prevent scattering of axons and Schwann cells when a break in the perineurium occurs. Following the construction of this perineurial cellular network, progress towards the reconstruction of an intact peripheral perineurium may coincide with the change in the accompanying blood vessels as the blood-nerve barrier is re-established.
Acknowledgements
We wish to thank Dr J. Mitchell, Professor G. Kerkut and the staff of the Electron

402 A. M. Ahmed and R. 0. Weller
Microscope Unit, Southampton General Hospital for their advice and criticism. Mrs Olive Huber kindly typed the manuscript.
References
ANDRES K.H. (1967) o b e r die Feinstruktur der Arachnoidea und Dura Mater von Mammalia. Zeitschrift fiir Zellforschung und Mikroskopische Anatomie (Berlin) 97, 271-295
BENKE B. & ROHLICH P. (1964) Elektronenmikroskopische Untersuchungen an den Hiillen der Rucken- markswurzeln. I: Hintere Wurzel. Journal f u r Hirnforschung 7, 87-98
DE LA MOTTE D. & ALLT G. (1976) Crush injury to peripheral nerve: An electron microscope study employing horseradish peroxidase. Acta Neuropathologica (Berlin) 36, 9-19
GAMBLE H.J. & BREATHNACH A.S. (1965) An electron microscope study of human foetal peripheral nerves. Journal of Anatomy (London) 99, 573-584
GRAHAM R.C. & KARNOVSKY M.J. (1966) The early stages of absorption of injected horseradish peroxi- dase in the proximal tubules of mouse kidney : Ultrastructural cytochemistry by a new technique. Journal of Histochemistry and Cytochemistry 14, 291-302
JURECKA W., AMMERER H.P. & LASSMANN H. (1975) Regeneration of a transected peripheral nerve. An autoradiographic and electron microscopic study. Acta Neuropathologica (Berlin) 32, 299-312
KLEMM H. (1970) Das Perineurium als Diffusionsbarriere gegeniiber Peroxydase bei epi- und endo- neuraler Applikation. Zeitschrift fur Zellforschung und Mikroskopische Anatornie (Berlin) 108, 431-445
KRISTENSSON K. & OLSSON Y. (1971) The perineurium as a diffusion barrier to protein tracers. Differ- ences between mature and immature animals. Acta Neuropathologica (Berlin) 17, 127-138
MASSON P. (1942) I. Tumeurs encapsulees e t bknignes des nerfs. Canadian Biological Reviews 1,209-343 MORRIS J.H., HUDSON A.R. & WEDDELL, G. (1972) A study of degeneration and regeneration in divided
rat sciatic nerve based on electron microscopy. IV. Changes in fascicular microtopography, perineurium and endoneurial fibroblasts. Zeitschrift fur Zellforschung und Mikroskopische Anatornie (Berlin) 124, 165-203
OCHS S. & WORTH R.M. (1978) Axoplasmic transport in normal and pathological systems. In Physiology and Pathobiology of Axons, pp. 251-264, Ed. Waxman S.G. Raven Press, New York
OLSSON Y. (1966) Studies on vascular permeability in peripheral nerves. I: Distribution of circulating fluorescent serum albumin in normal, crushed and sectioned rat sciatic nerve. Acta Neuropatho- Zogica (Berlin) 7, 1-15
OLSSON Y. & KRISTENSSON K. (1973) The perineurium as a diffusion barrier to protein tracers following trauma to nerves. Acta Neuropathologica (Berlin) 23, 108-111
OLSON Y., KRISTENSSON K. & KLATZO J. (1971) Permeability of blood vessels and connective tissue sheaths in the peripheral nervous system to exogenous proteins. Acta Neuropathologica (Berlin), Supplement V, 61-69
OLSSON Y. & REESE T.S. (1971) Permeability of vasa nervorum and perineurium in mouse sciatic nerve studied by fluorescence and electron microscopy. Journal of Neuropathology and Experimental Neurology 30, 105-119
POLLARD J.D. & FITZPATRICK L. (1973) A comparison of the effects of irradiation and immunosuppressive agents on regeneration through peripheral nerve allografts. An ultrastructural study. Acta Neuropathologica (Berlin) 23, 166-180
REAL E., LUCIANO, L. & SPITZNAS M. (1975) Freeze fracture faces of the perineural sheath of the rabbit sciatic nerve. Journal of Neurocytology 4, 261-270
REXED B. (1942) Uber die Aktivitat der Schwannschen Zellen bei der Nervenregeneration. I: Die Uberbruckung neuritloser nervenlucken. Zeitschrift fur Zellforschung und Mikroskopische Ana- tomie (Berlin) 51, 177-205

Blood-nerve barrier and perineurium following nerve grafting 483
SHANTHAVEERUPPA T.R. & BOURNE G.H. (1966) Perineurial epithelium: a new concept of its role in the integrity of the peripheral nervous system. Science 154, 1464-1467
SODERFELDT B. & OLSSON Y. (1973) The perineurium as a diffusion barrier to protein tracers in human peripheral nerve. Acta Neuropathologica (Berlin) 25, 120-126
THOMAS P.K. (1966) The cellular response to nerve injury. I: The cellular outgrowth from the distal stump of transected nerve. Journal of Anatomy (London) 100, 287-303
THOMAS P.K. & BHAGAT S. (1978) The effect of extraction of the intrafascicular contents of peripheral nerve trunks on perineurial structure. Acta Neuropathologica (Berlin) 43, 135-141
THOMAS P.K. & JONES D.G. (1967) The cellular response to nerve injury. 2: Regeneration of the peri- neurium after nerve section. Journal of Anatomy (London) 101, 45-55
WAGGENER J.P., BUNN S.M. & BEGGS J. (1965) The diffusion of ferritin within the peripheral nerve sheath: an electron microscopy study. Journal of Neuropathology and Experimental Neurology 24, 430443
YOUNG J.Z., HOLMES W. & SAUNDERS F.K. (1940) Nerve regeneration. Importance of the peripheral stump and the value of nerve grafts. Lancet 2 , 128-130
E
