ultrastructure of the circumoral nerve ring and the radial nerve cords in holothurians...
TRANSCRIPT

ORIGINAL ARTICLE
Vladimir S. Mashanov Æ Olga R. Zueva
Thomas Heinzeller Æ Igor Yu. Dolmatov
Ultrastructure of the circumoral nerve ring and the radial nerve cordsin holothurians (Echinodermata)
Received: 2 May 2005 / Accepted: 12 November 2005 / Published online: 1 February 2006� Springer-Verlag 2006
Abstract The circumoral nerve ring and the radial nervecords (RNCs) of Eupentacta fraudatrix and Pseudocnuslubricus (Holothuroidea) were examined as an exampleof holothurian nervous tissue. The RNC is composed ofouter ectoneural and inner hyponeural layers, which areinterconnected with one another via short neuralbridges. The circumoral nerve ring is purely ectoneural.Both ectoneural and hyponeural components are epi-thelial tubes with a thick neuroepithelium at one side. Athin ciliated non-neuronal epithelium complements theneuroepithelium to form a tube, thereby enclosing theepineural and hyponeural canals. The whole of the ec-toneural and hyponeural subsystems is separated fromthe surrounding tissue by a continuous basal lamina.The nerve ring and the ectoneural and hyponeural partsof the radial nerves are all neuroepithelia composed ofsupporting cells and neurons. Supporting cells areinterpreted as being glial cells. Based on ultrastructuralcharacters, three types of neurons can be distinguished:(1) putative primary sensory neurons, whose ciliumprotrudes into the epineural or hyponeural canal; (2)non-ciliated neurons with swollen rough endoplasmicreticulum cisternae; (3) monociliated neurons that areembedded in the trunk of nerve fibers. Different types ofsynapses occur in the neuropile area. They meet allmorphological criteria of classical chemical synapses.Vacuolated cells occur in the neuroepithelium of E.fraudatrix, but are absent in P. lubricus; their function isunknown. The cells of the non-neuronal epithelia thatoverlie the ectoneural and hyponeural canals are
hypothesized to belong to the same cell type as thesupporting cells of the neuroepithelium.
Keywords Nervous system Æ Holothuroidea ÆEctoneural–hyponeural connection Æ Synapses ÆGlial cells
Introduction
Echinodermata, together with Vertebrata, Cephalo-chordata, Tunicata, and Hemichordata, constitute theDeuterostomia. Since monophyly of this group is wellsupported (Adoutte et al. 2000), studies of the echino-derm nervous system can improve our understanding ofthe general principles of organization and general trendsin the evolution of the nervous system in deuterostomesin general.
It is generally believed that the echinoderm nervoussystem exhibits a number of unusual or unique featuresthat have not been reported for other metazoans. Theseare as follows:
(1) The echinoderm nervous system is composed ofseparate subsystems. It is thought that, in most in-stances, there is no direct anatomical connectionbetween the ectoneural and hyponeural subsystems.The ectoneural subsystem is believed to originatefrom the ectoderm during the ontogenesis, while thehyponeural subsystem seems to develop as a com-ponent of the coelomic mesoderm. Besides these twosubsystems, crinoids have an aboral subsystem,which is also reported to be mesodermally derived(Beklemishev 1964; Cobb 1987, 1990, 1995).
(2) The echinoderm nervous system has traditionallybeen regarded as being devoid of glial cells (Stubbsand Cobb 1981; Cobb 1987, 1990, 1995). However,some recent papers have challenged this view (Byrne1994; Viehweg et al. 1998; Heinzeller et al. 2004).
(3) Classic chemical synapses are extremely scarce inthe echinoderm nervous system (Cobb 1987, 1990;
V. S. Mashanov (&) Æ O. R. Zueva Æ I. Yu. DolmatovInstitute of Marine Biology FEB RAS,Palchevsky Str. 17, 690041 Vladivostok, RussiaE-mail: [email protected].: +7-4232-311178Fax: +7-4232-310900
T. HeinzellerLudwig-Maximilians-Universitat Munchen,Pettenkoferstrasse 11, 80336 Munchen, Germany
Zoomorphology (2006) 125: 27–38DOI 10.1007/s00435-005-0010-9

Peters and Campbell 1987; Byrne 1994; Heinzellerand Welsch 1994).
(4) There are no specialized central ganglia that coor-dinate echinoderm behavior. Any part of the radialnerve cord (RNC) can dominate and control wholeanimal behavior, and that control of behavior shiftsto other parts of the nervous system according tocircumstances (Cobb 1987, 1988, 1990, 1995;Heinzeller and Welsch 2001).However, in our opin-ion, the present state of knowledge regarding thehistology and ultrastructure of the echinoderm ner-vous system is not sufficient for drawing definiteconclusions. We regard the above statements asworking hypotheses, which should be tested as moredata become available.
The nervous system has been studied to a certainextent in four of five extant echinoderm groups (Cri-noidea, Asteroidea, Ophiuroidea, and Echinoidea). Onthe other hand, our knowledge of the holothurian ner-vous system is limited primarily to the nerves associatedwith viscera and body appendages (Bouland et al. 1982;McKenzie 1987, 1988; VandenSpiegel and Jangoux1987, 1993; Flammang and Jangoux 1992; Smiley 1994;VandenSpiegel et al. 1995; Mashanov and Dolmatov2001; Mashanov et al. 2004). There have been no papersthat focus specifically on the fine structure of the pe-ripharyngeal nerve ring and RNCs, which constitute themost prominent part of the holothuroidean nervoussystem. To fill this gap in our knowledge, we studied theultrastructure of these regions of the nervous system inthe dendrochirotid holothuroidean Eupentacta frauda-trix and Pseudocnus lubricus. For a detailed descriptionof macroanatomical features of the holothuroideannervous system the reader is referred to Hyman’s review(Hyman 1955), as well as to reports of a number of otherauthors (Cuenot 1948; Beklemishev 1964; Cobb 1987;Heinzeller and Welsch 2001). The nervous systems of P.lubricus and E. fraudatrix follow the general scheme.Therefore, we will give only a brief anatomical accountin this paper.
Materials and methods
Adult individuals of E. fraudatrix Djakonov et Bara-nova, 1958 (Holothuroidea, Dendrochirota) were col-lected from Vostok Bay, Sea of Japan, in May 2002.Juvenile individuals of P. lubricus H.L. Clark, 1901(Holothuroidea, Dendrochirota), a brooding species,collected at Swordfish Island, Juan de Fuca Strait,northeastern Pacific, in June 1998. Aquapharyngealbulbs of E. fraudatrix and small pieces of the body wall,together with the organ complex belonging to the radii,were fixed for 6 days at 4�C in 2.5% glutaraldehyde in0.05M cacodylate buffer at pH 7.6. Whole specimens ofP. lubricus were fixed in 2.5% glutaraldehyde in 0.1Mphosphate buffer (pH 7.2) for several days and then bi-sected sagittally into two halves. After fixation, the
specimens were rinsed in the respective buffer andpostfixed in OsO4 buffered in the same manner as therespective fixative, dehydrated in a graded ethanol seriesfollowed by acetone, and embedded in Araldite (Fluka).Sections were cut with glass knives using an Ultracut E(Reichert) ultratome. Semithin (0.8 lm) sections weretransferred onto gelatin-coated slides, stained with 1%toluidine blue in 1% aqueous sodium borate, andmounted in DPX (Fluka). The slides were examined andphotographed with a Jenamed 2 (Zeiss) light microscopeequipped with a Leica DC 150 digital camera. Ultrathin(50–70 nm) sections were stained with aqueous uranylacetate followed by lead citrate using an Ultrastainer(Reichert), and examined in a Zeiss EM 10 transmissionelectron microscope.
Results
General remarks
The RNCs run in the radii and span the body lengthfrom the anterior toward the posterior end (Fig. 1).They lie in the inner connective tissue layer of the bodywall and accompany the water-vascular canals, beingseparated from the latter by radial hemal lacunae(Fig. 2). Each RNC is composed of a thicker ectoneuraland a thinner hyponeural band (Figs. 1, 2a, b, 3), whichare separated by a thin partition of connective tissue.The two parts, however, come in direct connection viashort neural bridges that traverse the connective tissue(Figs. 1, 2b, 3, 4a). Both ectoneural and hyponeuralparts of the RNC are epithelial tubes, whose luminaconstitute an epineural and an hyponeural canal,respectively. Those epithelial walls that rest upon the
Fig. 1 Simplified schematic representation of the holothuroideannervous system. en ectoneural band of the radial nerve cord (RNC),hn hyponeural band of the RNC, nr circumoral nerve ring, rncRNC. Arrowheads point at short neural bridges that connect theectoneural and hyponeural subsystems. Tentacular and podialnerves are not shown. Not to scale
28

connective tissue partition are formed by a thick neu-roepithelium. A flat non-neuronal epithelium closes theepineural canal toward the body wall and the hypo-neural canal toward the hemal lacuna. Close to theanterior body end, at the bases of tentacles, the ecto-neural parts of the RNCs join each other to form aperipharyngeal nerve ring (Figs. 1, 2a). The hyponeuralsystem is restricted to the RNCs (Figs. 1, 2). At theposterior body end, the RNCs terminate and make nocontacts with one another.
Ultrastructure of the radial nerve cord and nerve ring
The two species share similar ultrastructural featuresand therefore will be described together unless uniquecharacters are worth mentioning. Both the RNCs andthe nerve ring are separated from the adjacent connec-tive tissue by a continuous basal lamina that follows theoutline of the nervous system and also covers the shortneural bridges, which connect the ectoneural and hyp-oneural subsystems (Figs. 3, 4a). The nerve ring andectoneural and hyponeural bands of the RNCs havebasically the same organization and are composed of aneuroepithelium, whose major components are sup-porting cells and neurons. The third cell type, vacuolatedcells, has been observed only in the apical part of theneuroepithelium of E. fraudatrix and is most abundantin the RNCs.
Supporting cells
Supporting cells constitute a framework that supportsother cell types of the neuroepithelium. They are trueepithelial cells that are composed of a cup-shaped apical
part that faces the lumen of the epineural or thehyponeural canal and a slender basal process that spansthe depth of the neuroepithelium and is connected to thebasal lamina through hemidesmosomes (Figs. 3, 4b, c).The apical surface of the cells bears a cilium anchored to abasal body and rare short microvilli that occasionallyanastomose with one another. Adluminal sheets spreadout of the perinuclear areas and are bound to neighboringcells (supporting cells or type I neurons, see below) byzonulae adhaerens and septate junctions. A spherical toelliptical nucleus with large patches of heterochromatinoccupies most of the cup-shaped apical part of the cell.The perinuclear cytoplasm contains few cisternae ofrough endoplasmic reticulum (RER), mitochondria,phagosomes, and electron-lucent vesicles. The supportingcells of the nerve ring and hyponeural part of the RNCshow clear signs of secretory activity. Their cytoplasmcontains numerous large secretory vacuoles (Fig. 4d)with fibrillar content, and a well-developed Golgi appa-ratus. Electron micrographs suggest that these cells candischarge their secretions into the lumen of the hypo-neural and ring epineural canals. A characteristic featureof the supporting cell is the bundle of intermediate fila-ments, which extends from the cup-shaped apical part tothe distal end of the basal process (Figs. 3, 4b, c). In somecells, the bundle exhibits a distinct transverse striationwith a period of about 310 nm. A few mitochondria andphagosomes are also occasionally encountered in thecytoplasm of the basal processes.
Nerve cells
Cell bodies and processes of nerve cells fill the intersticesbetween the supporting cells (Fig. 3). The perikaryaare evenly distributed throughout the depth of the
Fig. 2 Semithin sections through the nerve ring and the radialnerve cord (RNC). a Pseudocnus lubricus. Longitudinal sectionthrough the nerve ring and adjacent RNC; arrow marks the blindanterior end of the hyponeural canal. b Eupentacta fraudatrix.Transverse section through the RNC; arrowheads show the shortnerves that connect the ectoneural and the hyponeural subsystems;
asterisk indicates the longitudinal connective tissue partitionseparating the two parts of the RNC. cec circular epineural canal,ec epineural canal, en ectoneural band of the RNC, h radial hemallacuna, hc hyponeural canal, hn hyponeural band of the RNC, lmlongitudinal muscle band, nr nerve ring, rc radial water-vascularcanal
29

neuroepithelium forming no distinct aggregations. Thefollowing cell types were distinguished based on theirultrastructure and position within the neuroepithelium.
Type I neurons occur in the ectoneural and hypo-neural subsystems of both species. Perikarya of theseneurons lie in the apical region of the neuroepitheliumand face the lumen of the epineural or the hyponeuralcanal (Figs. 3, 4e). They either are separated from oneanother by supporting cells or form clusters of two tothree cells (Figs. 3, 4e). The perikarya are connected toother cells (supporting cells or other type I neurons)through apicolateral zonulae adhaerens and septatejunctions. The apical plasma membrane of the perikar-yon forms a shallow pit, from which a cilium projectsinto the canal (Fig. 4e, f). The cilium is surrounded bycytoplasmic protrusions and microvilli. The axonalprocess arises from the opposite pole of the cell andintermingles with processes of other neurons. In thehyponeural subsystem of P. lubricus, we found a single
neuron whose perikaryon faces the lumen of the hypo-neural canal, but whose cilium protrudes between theunderlying nerve processes instead of projecting into thelumen.
The cytoplasm of the perikarya contains a highlydeveloped RER and mitochondria. Microtubules runfrom the basal body of the cilium, pass near the nucleus,and enter the axonal process. The Golgi complex liesbetween the nucleus and the basal bodies. Rosette-likeaggregations of glycogen, a few residual bodies, andsmall vesicles are evenly distributed throughout thecytoplasm. We distinguished light, dense, and dense-corevesicles (40–130 nm in diameter). Neurons differ in themorphology and number of vesicles; different types ofvesicles often coexist within single cells.
The cell bodies of other cell types are evenly distrib-uted throughout the depth of the neuroepithelium andnever reach the lumen of the ectoneural or hyponeuralcanals.
Fig. 3 Schematical reconstruction of a transverse section throughthe radial nerve cord (RNC), arrowheads show the short nervesthat connect the ectoneural and the hyponeural subsystems. blbasal lamina, ctp connective tissue partition, ec ectoneural canal,
en ectoneural band of the RNC, hc hyponeural canal, hnhyponeural band of the RNC, nne non-neuronal epithelium, npnerve processes, n1 type I neuron, n2 type II neuron, n3 type IIIneuron, sc supporting cell, vc vacuolated cell. Not to scale
30

Fig. 4 Eupentacta fraudatrix (a–e, g) and Pseudocnus lubricus (f).Neuroepithelia of the nerve ring and the radial nerve cord (RNC). aShort neural bridges between the ectoneural (en) and hyponeuralbands (hn) of the RNC, arrows indicate the continuous basallamina of the RNC. b Low-magnification micrograph showingsupporting cells in the ectoneural layer of the RNC. c Attachmentof the basal process (bp) of a supporting cell to the basal lamina(bl), arrowheads mark hemidesmosomes. d Large secretory vacuolein the supporting cell in the hyponeural subsystem. e Type I neuron
in the apical part of the ectoneural neuroepithelium of the RNC. fDetailed view of the apical part of type I neuron in the nerve ring. gType II (putative) neuron in the nerve ring. ap axonal process, cicilium, ctp connective tissue partition, dv dense/dense-cored vesicle,ec ectoneural canal, f bundles of intracellular filaments, ga Golgistack, hc hyponeural canal, nne non-neuronal epithelium, np nerveprocesses, n1 type I neuron, rer rough endoplasmic reticulum, scsupporting cell, v vacuole
31

Type II cells occur in the ectoneural and hyponeuralsubsystems in both species. These cells possess anirregularly shaped perikaryon and one or two processes.A distinguishing feature of this cell type is that thecytoplasm contains a well-developed Golgi apparatusand large swollen RER cisternae filled with a material ofmoderate electron density (Fig. 4g). Mitochondria andrare light and dense-core (50–100 nm in diameter) vesi-cles occur both in the processes and in the perinuclearcytoplasm. This cell type seems to have no cilia.
Type III neurons are the most abundant nerve cells inboth species. All these cells bear a cilium, whose basalbodies are located between the nucleus and the axonalhillock region (Fig. 5a). Electron-lucent (40–100 nm indiameter), electron dense, and dense-core (80–130 nm indiameter) vesicles are usually colocalized in the cyto-plasm of single cells (Fig. 5b); any of them being themost abundant in individual neurons. The number ofvesicles per cell varies widely. The cytoplasm oftencontains glycogen rosettes and residual bodies. The de-gree of development of Golgi complex, RER, andmitochondria also differs widely among the neurons ofthis type. Some cells have a very narrow rim of theperinuclear cytoplasm with very scarce vesicles andmembrane organelles. Other cells possess a highlydeveloped Golgi apparatus composed of several dict-yosomes, numerous vesicles, mitochondria, and RERcisternae (Fig. 5b). We also encountered nerve cellswhose perinuclear cytoplasm had very scarce membraneorganelles but contained large deposits of glycogenclusters (Fig. 5c).
Nerve processes
Numerous densely packed nerve processes fill theinterstices between the supporting cells, neuronal peri-karya, and the basal lamina of the neuroepithelium. Inthe ectoneural parts of the radial cords, the processesare oriented mostly in the longitudinal direction, i.e.,parallel to the long axis of the RNC, although someaxons run parallel to the basal processes of the sup-porting cells, and some others are transverse or oblique.In the hyponeural subsystem, most of the nerve pro-cesses also run in the longitudinal direction, althoughthere are transverse bundles in the basal region of theneuroepithelium. In the nerve ring, most nerve fibersare arranged in circular tracts that join the radialnerves; a small number of processes run in otherdirections. The processes usually measure 0.1–0.3 lm indiameter, although they often form varicosities up to1.1–1.7 lm thick (Fig. 5d). At least some of the nervefibers give off collaterals. The cytoplasm of the nerveprocesses contains microtubules, mitochondria, RERcisternae, different types of vesicles, as well as rareglycogen inclusions and residual bodies. Some of thenerve fibers look empty in electron micrographs. It isworth noting that different types of vesicles often maycoexist in a single fiber.
Synapses
In both species, synapses occur throughout the neuro-epithelia of the nerve ring and RNCs. They closelyresemble classical chemical synapses (Fig. 6). Pre- andpostsynaptic membranes are thickened and lie at a dis-tance of about 15 nm from each other. The diameter ofthe active zone ranges from 0.3 to 0.9 lm. All synapsesare polarized, i.e., the vesicles are crowded close to thepresynaptic membrane, but absent at the postsynapticside. Vesicles of different types can coexist in individualpresynaptic endings (Fig. 6a). It is worth noting that anyregion of the plasma membrane, including the mem-brane of the perikaryon, can form synapses on otherneurons (Fig. 6b). The following morphological types ofsynapses can be distinguished.
In simple synapses, the presynaptic membrane con-tacts the postsynaptic membrane through one activezone associated with one cluster of vesicles (Fig. 6a, b).One of the varieties of the simple synapse in the speciesstudied is the ensheathing synapse, which consists of apresynaptic ending that forms a continuous cover overthe postsynaptic terminal. In this synapse, vesicles areclustered over the entire inner surface of the presynapticmembrane (Fig. 6c). Adjacent nerve processes occa-sionally form en passant synapses that do not involve anerve terminal and may occur between any parts of twoparallel fibers (Fig. 6d).
The remaining synapses were complex, being com-posed of either a presynaptic ending forming two (ormore) distinct simple synapses onto different postsyn-aptic terminals (Fig. 6e), or, the opposite situation, two(or more) distinct presynaptic endings synapsing withthe same postsynaptic neuron (Fig. 6f).
Non-neuronal epithelium
The floor of the epineural canal and the roof of thehyponeural canal consist of a thin flattened unilayerednon-neuronal epithelium, which is totally devoid ofnerve cell bodies and processes (Figs. 3, 7a). The epi-thelial cells are held together with adhaerens junctionsand rest on a thickened basal lamina, being attached tothe latter through hemidesmosomes. An elongated nu-cleus is usually aligned along the long axis of the cell.The cells bear a cilium and no or very few microvilli(Fig. 7b). The cytoplasm contains a Golgi complex,relatively well-developed RER, mitochondria, and vac-uoles filled with heterogeneous material. These vacuolescan reach the apical plasma membrane and seem to re-lease their contents into the lumen of the canal (Fig. 7b).The apical plasma membrane can form pinocytotic pits.The epithelial cells of the non-neuronal epitheliumcontain bundles of intermediate filaments that are sim-ilar to those in the supporting cells of the neuroepithe-lium. In some cells, they show a transverse striation witha period of about 300–310 nm (Fig. 7a). However, incontrast to supporting cells, these bundles are aligned
32

either parallel to the apical cell surface or at an acuteangle to it.
Vacuolated cells
Vacuolated cells (Fig. 7c) occur in the nerve ring andRNC of E. fraudatrix, but are absent in the juvenile P.lubricus material. These large (up to 12.5 lm in diame-ter) spherical cells lie in the apical region of the neu-roepithelium, but never reach the lumen of ectoneuraland hyponeural canals and are not joined to adjacentcells through intracellular junctions. The cytoplasm ofthese cells is filled with small to large light vacuoles of
varying shape, which sometimes fuse with one another.A subspherical nucleus is located either in the centralpart of the cell or pushed to the periphery. The inter-vacuolar cytoplasmic regions contain scarce mitochon-dria and RER cisternae.
Discussion
Gross anatomy
The general trends in the evolution of the metazoannervous system include the centralization of the nervoustissue, submergence of the latter below the epidermis,
Fig. 5 Eupentacta fraudatrix (a, c, d) and Pseudocnus lubricus (b).a Type III neuron (n3) in the ectoneural part of the radial nervecord (RNC). b High magnification view of a type III neuron in thenerve ring showing co-localization of different type of vesicles in thecytoplasm. c Perikaryon with glycogen deposits in the nerve ring.
d Varicosity (var) of a nerve fiber in the ectoneural part of theRNC. ci cilium, bp basal process, dv dense/dense-cored vesicle, gaGolgi stack, gl glycogen, lv light vesicles, mt mitochondrion, mtbmicrotubules, rer rough endoplasmic reticulum, v vacuole
33
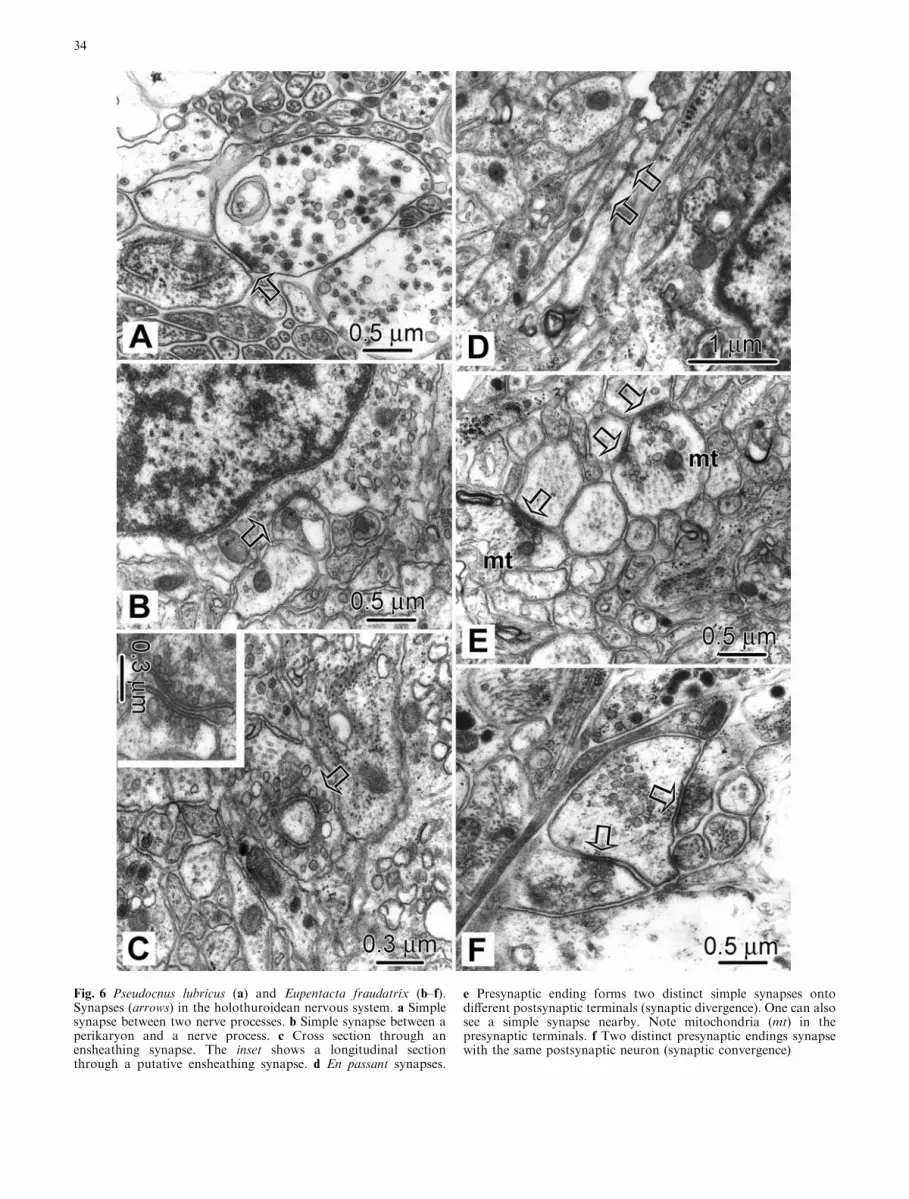
Fig. 6 Pseudocnus lubricus (a) and Eupentacta fraudatrix (b–f).Synapses (arrows) in the holothuroidean nervous system. a Simplesynapse between two nerve processes. b Simple synapse between aperikaryon and a nerve process. c Cross section through anensheathing synapse. The inset shows a longitudinal sectionthrough a putative ensheathing synapse. d En passant synapses.
e Presynaptic ending forms two distinct simple synapses ontodifferent postsynaptic terminals (synaptic divergence). One can alsosee a simple synapse nearby. Note mitochondria (mt) in thepresynaptic terminals. f Two distinct presynaptic endings synapsewith the same postsynaptic neuron (synaptic convergence)
34

and development of advanced integration centers asso-ciated with complex sense organs (Beklemishev 1964).The first two trends can be easily seen in echinoderms.For instance, the basiepithelial ectoneural nerve plexusis integrated into the epidermis of asteroids all over thebody surface and reaches the highest concentration inthe epidermis of the ambulacral grooves thereby formingthe radial cords, which are connected with one anotherby a peristomial nerve ring (Hyman 1955; Kawaguti1965; Chia and Koss 1994). In the remaining Eleuther-ozoa—echinoids, ophiuroids, and holothuroideans—theectoneural system detaches from the epidermis and liesbeneath the body surface (Hyman 1955; Smith 1984;Markel and Roser 1991; Byrne 1994). As a result, asubcutaneous nerve tube has evolved, whose internalwall is composed of a neuroepithelium, which is overlainby a non-neuronal epineural epithelium.
In echinoderms, the ectoneural subsystem is usuallyaccompanied by the hyponeural subsystem. There is apositive correlation between the degree of developmentof the hyponeural subsystem and that of muscles, whichis why this subsystem is thought to be purely motor(Hyman 1955; Beklemishev 1964; Cobb 1987; Heinzellerand Welsch 2001). Holothuroidean species possesspowerful body wall muscles and a rigid peripharyngealskeleton. Accordingly, only the RNCs are combinedwith the hyponeural layer, and no hyponeural ring canbe observed. The hyponeural system of the Eleutherozoais often referred to as a basiepithelial plexus associatedwith the lining epithelium of the hyponeural (perihemal)coelomic canals (Hyman 1955; Byrne 1994; Chia andKoss 1994; Heinzeller and Welsch 2001). In Asteroidaand Ophiuroida, myoepithelial cells were shown to beinterspersed among supporting and nerve cells of thehyponeural radial nerve (van Hehn 1970; Byrne 1994).On the other hand, in both holothuroidean speciesstudied, the hyponeural cord shows no signs of meso-dermal origin and is very similar to the ectoneural cordin terms of histological structure and cell composition.
It has been stressed repeatedly by Cobb (1987, 1990)that there is no direct anatomical connection betweenthe ectoneural and hyponeural subsystems. However,the possibility that there is a physiological interactionvia chemical synapses across the basal lamina (and athin layer of connective tissue) has not been ruled out.Rare instances of direct connection between the twosubsystems (Cobb and Stubbs 1981, 1982; Stubbs andCobb 1981; Peters 1985; Heinzeller and Welsch 2001)have been considered to be exceptions that do not refutethe general rule (Cobb 1987). Contrarily, in both
holothuroidean species studied, the hyponeural and ec-toneural bands of the RNC are clearly connected viashort neural bridges that are composed of nerve cell
Fig. 7 Eupentacta fraudatrix. Non-neuronal epithelium coveringthe hyponeural and ectoneural canals. a Cell of the non-neuronalepithelium in the ectoneural part of the RNC. bHigh-magnificationview of the apical cytoplasm of a cell in the hyponeural non-neuronal epithelium. c Vacuolated cell. bl basal lamina, bp basalprocesses of supporting cells, ci cilium, ec epineural canal, f bundlesof intracellular filaments, ga Golgi apparatus, hc hyponeural canal,np nerve processes, rb residual body, v vacuole, vc vacuolated cell
c
35

perikarya and processes. Moreover, the ectoneural andhyponeural parts of the RNC as well as the neuralbridges that connect the two subsystems are covered bya continuous basal lamina that separates them from theadjacent tissues. Thus, our data on histology andstructural interrelationships between the ectoneural andhyponeural subsystems suggest that, contrary to thegeneral belief, it is quite possible that the two parts ofthe holothuroidean nervous system originate from thesame source. The only way to resolve this contradictionis to carry out a scrupulous study of the developingnervous system. However, whatever the case, the ec-tonerural and hyponeural tissues of E. fraudatrix and P.lubricus should be referred to as two interconnectedparts of the whole, anatomically continuous nervoussystem rather than two separate subsystems.
Neuroepithelium
In both holothuroidean species studied, as in all theother echinoderms, irrespective of whether they possessopen or closed ambulacra, the ectoneural subsystemexhibits the organization of a neuroepithelium (Bek-lemishev 1964; Kawaguti 1965; Markel and Roser 1991;Chia and Koss 1994; Viehweg et al. 1998; Heinzeller andWelsch 2001). In holothuroidean, the hyponeural bandis similar to the ectoneural tissue both in histologicalstructure and cell composition. In both subsystems, theframework of the neuroepithelium is composed of sup-porting cells, whose major feature is the presence ofbundles of intermediate filaments, which run the heightof the cells from the apical to the basal surfaces. Similarcells were found in the ectoneural subsystem of manyother echinoderms (Markel and Roser 1991; Byrne 1994;Chia and Koss 1994; VandenSpiegel et al. 1995; Viehweget al. 1998), but often were not called glial cells by theauthors. Therefore, it has long been stressed that theechinoderm nervous tissue has no glia (Stubbs and Cobb1981; Cobb 1990, 1995). However, Viehweg et al. (1998)showed that the supporting cells of the ectoneural cordof the asteroid Asterias rubens were labeled specificallywith a polyclonal antiserum directed against Reissner’ssubstance, which is commonly produced and secreted bythe radial glia in chordates. In ophiuroid species, thesecells seem to exhibit positive CRF- and S-100-likeimmunoreactivity (Heinzeller et al. 2004). These obser-vations suggest that the supporting cells are true glialelements. They provide support to neurons and seem tobe capable of phagocytosis and secretory activity. Sup-porting cells of echinoderms are therefore similar toependymal cells of vertebrates and are probablyhomologous to the latter. In metazoans, nerve cellsmight have initially evolved as a component of primitiveepithelia. There were initially no specialized glial cells,although, epithelial cells of that primitive tissues couldaccomplish some additional functions to support andnourish the intraepithelial neurons. Later, the cell bodiesand processes of the nerve cells presumably formed a
highly developed basiepithelial plexus, while the epithe-lial cells probably gave rise to the ependymal glia ofvertebrates (Borisov et al. 1986) and ependyma-like glialcells of echinoderms.
The cells of the non-neuronal epithelia that line theepineural and hyponeural canals in E. fraudatrix and P.lubricus are morphologically similar to the supportingcells of the neuroepithelium. Both these cell types con-tain bundles of intracellular filaments in their cytoplasm.Although the non-neuronal epithelium is flattened andcontains no neurons, its constituent cells may be a var-iant of the ependyma-like supporting cells.
Nerve cells
In the RNCs and nerve ring of E. fraudatrix and P.lubricus, the perikarya of nerve cells seem to be distrib-uted evenly throughout the neuroepithelium and showno clustering into ganglionic units. It remains unclearwhether the neurons are randomly distributed or, to-gether with the glial supporting cells, form a specificpattern, as is the case in asteroid arms (Viehweg et al.1998). In the neuroepithelium of holothuroidean species,we distinguish the following nerve cell types:
(1) Type I cells whose cell bodies are in contact with thelumen of the epineural or hyponeural canal, whereasthe axonal process arises from the basal pole of thecell. Similar cells occur in the epidermis in all echi-noderm classes (Peters and Campbell 1987; Byrne1994; Chia and Koss 1994; Heinzeller and Welsch1994; VandenSpiegel et al. 1995) and are commonlyagreed to be primary sensory neurons (propriocep-tors and chemoreceptors) (Cobb 1987; Markel andRoser 1991), although there is little convincingphysiological evidence for this (Heinzeller and Wel-sch 2001). It is worth noting that in E. fraudatrix andP. lubricus these cells are found both in the ecto-neural subsystem and in the hyponeural tissue. Ifthey do serve as receptors, the general belief that theechinoderm hyponeural subsystem is purely motormay be inaccurate (Hyman 1955; Beklemishev 1964;Cobb 1987).
(2) Type II cells whose cytoplasm contains swollen RERcisternae are never in contact with the lumen of theepineural and hyponeural canals. To our knowledge,such cells have not been found in the echinodermnervous system before. We refer to them as putativenerve cells, since they possess processes and vesiclesin the cytoplasm; however, this cell type deservesfurther investigation.
(3) Type III neurons are the most abundant nerve cellsin the holothuroidean species studied. They are al-ways embedded in the trunk of nerve fibers and beara cilium, which originates close to the axon hillockregion. These neurons are morphologically similarto the nerve cells of the radial nerves of ophiuroidsand echinoids (Cobb and Stubbs 1982; Markel and
36

Roser 1991) and seem to be the predominant nervecell type in echinoderms.
Synapses
Classical chemical synapses have been found very rarelyin Echinodermata (Cobb 1987, 1990; Peters andCampbell 1987; Byrne 1994; Heinzeller and Welsch1994). Interneuronal transmission is believed to occurusually via simple varicose endings, which are filled withvesicles and do not show morphological specializationsof the pre- and postsynaptic membranes and cytoplasm(Cobb 1987, 1990). This paper is the first to report onthe presence of synapses in the nervous system of Ho-lothuroidea. These structures occur both in the nervering and RNCs of E. fraudatrix and P. lubricus and meetthe morphological criteria of a classical chemical syn-apse. Moreover, these synapses are not all the same.Besides simple and en passant synapses, which involvethe transmission from a presynaptic cell to a postsyn-aptic neuron, there are also combinations of simplesynapses that probably enable a cell to receive inputsfrom two (or more) other cells (synaptic convergence)or, alternatively, to supply two to many others (synapticdivergence).
Centralization of the nervous system
There are two different viewpoints concerning the cen-tralization of the echinoderm nervous system. Smith(1965, 1966) stated that the circumoral nerve ring andimmediately adjacent nerve cords act as control centersthat drive the behavior of the entire animal. Later, thisidea was challenged (Cobb and Stubbs 1982; Cobb 1987,1988, 1990; Heinzeller and Welsch 2001). According tothe current, widely accepted hypothesis, the echinodermnervous system is not centralized. The circumoral ringseems merely to connect the radial cords together and tomediate the interaction between them. In both holo-thuroidean species examined, the ectoneural ring isidentical to the ectoneural bands of the RNCs in termsof both histological organization and cell composition.Thus, the morphological data suggest that the holothu-roidean nerve ring and RNCs have equal functionalimportance, and there is no evidence for the nerve ringhaving a central coordinating function, although holo-thuroidean clearly possess a permanent anterior bodyend.
Acknowledgments We are grateful to Philip Lambert of the RoyalBritish Columbia Museum Victoria, Canada, for providing us withfixed specimens of juvenile P. lubricus and to Beate Aschauer(LMU, Munich) for her inestimable technical assistance. We areespecially indebted to Prof. I.C. Wilkie (Glasgow CaledonianUniversity, Scotland) for his critical reading of the manuscript. Thework was supported by the grant of the Russian Foundation forBasic Research (project no. 05-04-48715) to I.Yu. Dolmatov and bythe DAAD scholarship to V.S. Mashanov.
References
Adoutte A, Balavoine G, Lartillot N, Lespinet O, Prud’homme B,de Rossa R (2000) The new animal phylogeny: reliability andimplications. Proc Natl Acad Sci USA 97:4453–4456
Beklemishev VN (1964) Principles of comparative anatomy ofinvertebrates (in Russian). 3rd edn. Nauka, Moscow
Borisov IN, Dunaev PV, Bazhanov AN (1986) Phylogenetic prin-ciples of histological organization of animals (in Russian).Nauka, Novosibirsk
Bouland C, Massin C, Jangoux M (1982) The fine structure of thebuccal tentacles of Holothuria forskali (Echinodermata, Holo-thuroidea). Zoomorphology 101:133–149
Byrne M (1994) Ophiuroidea. In: Harisson FW, Chia FS (eds)Microscopic anatomy of invertebrates, vol. 14. Echinodermata.Wiley-Liss, New York, pp 247–344
Chia FS, Koss R (1994) Asteroidea. In: Harisson FW, Chia FS(eds) Microscopic anatomy of invertebrates, vol. 14. Echino-dermata. Wiley-Liss, New York, pp 169–246
Cobb JLS (1987) Neurobiology of the Echinodermata. In: Ali MA(eds) Nervous systems in invertebrates. Plenum Press, NewYork, pp 438–525
Cobb JLS (1988) A preliminary hypothesis to account for theneural basis of behaviour in echinoderms. In: Burke RD (eds)Echinoderm biology. Rotterdam, Balkema, pp 565–573
Cobb JLS (1990) Enigmas of echinoderm nervous systems. In:Anderson PAV (eds) Evolution of the first nervous systems.Plenum Press, New York, pp 329–337
Cobb JLS (1995) The nervous systems of Echinodermata: recentresults and new approaches. In: Breidbach O, Kutsch W (eds)The nervous systems of invertebrates: an evolutionary andcomparative approach. Birkhauser, Basel, Boston, Berlin, pp407–424
Cobb JLS, Stubbs T (1981) The giant neurone system inOphiuroids. I. The general morphology of the radial nervecords and the circumoral nerve ring. Cell Tissue Res 219:197–207
Cobb JLS, Stubbs T (1982) The giant neurone system in Ophiu-roids. III. The detailed connections of the circumoral nervering. Cell Tissue Res 226:675–687
Cuenot L (1948) Anatomie, ethologie et systematique des Echino-derms. In: Grasse PP (ed) Traite de Zoologie. II. Masson, Paris,pp 3–363
Flammang P, Jangoux M (1992) Functional morphology of thelocomotory podia of Holothuria forskali. Zoomorphology111:167–178
von Hehn G (1970) Uber den Feinbau des hyponeuralen Nerven-systems des Seesternes (Asterias rubens L.). Z Zellforsch105:137–154
Heinzeller T, Welsch U (1994) Crinoidea. In: Harrison FW, ChiaFS (eds) Microscopic anatomy of invertebrates, vol. 14. Echi-nodermata. Wiley-Liss, New York, pp 9–148
Heinzeller T, Welsch U (2001) The echinoderm nervous system andits phylogenetic interpretation. In: Roth G, Wullimann MF(eds) Brain evolution and cognition. Wiley-Spektrum, NewYork, pp 41–75
Heinzeller T, Ashauer B, Mashanov V (2004) Immunolocalizationof nNOS, CRF and S-100 in ecto/hyponeural cells and fibers ofOphiothrix fragilis. In: Heinzeller T, Nebelsick JH (eds) Echi-noderms: Munchen. Taylor & Francis Group, London, pp 335–338
Hyman LH (1955) The invertebrates. IV. Echinodermata. Thecoelomate Bilateria. McGraw-Hill Book Co. Inc., New York
Kawaguti S (1965) Electron microscopy on the radial nerve of thestarfish. Biol J Okayama Univ 11:41–52
Markel K, Roser U (1991) Ultrastructure and organization of theepineural canal and nerve cord in sea urchins (Echinodermata,Echinoida). Zoomorphology 110:267–279
Mashanov VS, Dolmatov IYu (2001) Ultrastructure of the ali-mentary canal in five-month-old pentactulae of the holothurianEupentacta fraudatrix. Russ J Mar Biol 27:320–328
37

Mashanov VS, Dolmatov IYu, Frolova LT (2004) Structure of thedigestive tube in the holothurian Eupentacta fraudatrix (Holo-thuroidea: Dendrochirota). Russ J Mar Biol 30:366–374
McKenzie JD (1987) The ultrastructure of the tentacles of elevenspecies of dendrochirote holothurians studied with special ref-erence to the surface coats and papillae. Cell Tissue Res248:187–199
McKenzie JD (1988) Ultrastructure of the tentacles apodousholothurian Leptosynapta spp. With special reference to theepidermis and surface coats. Cell Tissue Res 251:387–397
Peters BH (1985) The innervations of spines in the sea-urchinEchinus esculentus L. An electron-microscopic study. Cell Tis-sue Res 239:219–228
Peters BH, Campbell AC (1987) Morphology of the nervous andmuscular system in the heads of pedicellariae from the seaurchin Echinus esculentus L. J Morphol 193:35–51
Smiley S (1994) Holothuroidea. In: Harisson FW, Chia FS (eds)Microscopic anatomy of invertebrates, vol. 14. Echinodermata.Wiley-Liss, New York, pp 401–472
Smith JE (1965) Echinodermata. In: Bullock TH, Horridge GA(eds) Structure and function in the nervous system of inverte-brates, vol. 11. Freeman & Co., San Francisco, pp 1520–1558
Smith JE (1966) The form and function of the nervous system. In:Boolootian RA (ed) Echinoderm physiology. Wiley, New York,pp 503–512
Smith AB (1984) Classification of the Echinodermata. Paleontol-ogy 27(3):431–459
Stubbs T, Cobb JLS (1981) The giant neuron system in Ophiuroids.II. The hyponeural motor tracts. Cell Tissue Res 220:373–385
VandenSpiegel D, Jangoux M (1987) Cuvierian tubules of the ho-lothuroid Holothuria forskali: a morphofunctional study. MarBiol 96:263–275
VandenSpiegel D, Jangoux M (1993) Fine structure and behaviorof the so-called Cuvierian organs in the holothuroid genusActinopyga (Echinodermata). Acta Zool 74:43–50
VandenSpiegel D, Flammang P, Fourmeau D, Jangoux M (1995)Fine structure of the dorsal papillae in the holothurioid Hol-othuria forskali (Echinodermata). Tissue Cell 27:457–465
Viehweg J, Naumann WW, Olsson R (1998) Secretory radial gliacells in the ectoneural system of the sea star Asterias rubens(Echinodermata). Acta Zool 79:119–131
38