interleukin1β mediates gdnf up-regulation upon dopaminergic injury in ventral midbrain cell...
TRANSCRIPT

www.elsevier.com/locate/ynbdi
Neurobiology of Disease 25 (2007) 92–104Interleukin-1β mediates GDNF up-regulation upon dopaminergicinjury in ventral midbrain cell cultures
Ana Saavedra,a Graça Baltazar,b and Emília P. Duartea,c,⁎
aCenter for Neuroscience and Cell Biology, University of Coimbra, Coimbra, PortugalbHealth Sciences Research Center, University of Beira Interior, Covilhã, PortugalcDepartment of Zoology, University of Coimbra, Coimbra, Portugal
Received 27 May 2006; revised 24 August 2006; accepted 25 August 2006Available online 5 October 2006
We recently proposed the involvement of diffusible modulators insignalling astrocytes to increase glial cell line-derived neurotrophicfactor (GDNF) expression after selective dopaminergic injury by H2O2
or L-DOPA. Here we report that interleukin-1β (IL-1β) is involved inthis crosstalk between injured neurons and astrocytes. IL-1β wasdetected only in the media from challenged neuron–glia cultures.Exogenous IL-1β did not change GDNF protein levels in astrocytecultures, and diminished GDNF levels in neuron–glia cultures. Thisdecrease was not due to cell loss, as assessed by the MTT assay andimmunocytochemistry. Neither H2O2 nor L-DOPA induced microgliaproliferation or appeared to change its activation state. The IL-1receptor antagonist (IL-1ra) prevented GDNF up-regulation inchallenged cultures, showing that IL-1β is involved in the signallingbetween injured neurons and astrocytes. Since IL-1ra decreased thenumber of dopaminergic neurons in H2O2-treated cultures, we proposethat IL-1 has a neuroprotective role in this system involving GDNFup-regulation.© 2006 Elsevier Inc. All rights reserved.
Keywords: Astrocytes; Crosstalk; Dopaminergic neurons; GDNF; IL-1β;Injury; Microglia; Neuroprotection
Introduction
Parkinson’s disease (PD) patients experience motor impairmentdue to the selective loss of dopaminergic neurons from thesubstantia nigra pars compacta and consequent dopaminedepletion in the caudate-putamen. In addition to dopaminergiccell loss, there is also a significant glial reaction composed mainlyof activated microglial cells and, to a lesser extend, of reactiveastrocytes (Teismann et al., 2003). A marked increase in the level
⁎ Corresponding author. Center for Neuroscience and Cell Biology,Department of Zoology-University of Coimbra, 3004-517 Coimbra,Portugal. Fax: +351 239 855789.
E-mail address: [email protected] (E.P. Duarte).Available online on ScienceDirect (www.sciencedirect.com).
0969-9961/$ - see front matter © 2006 Elsevier Inc. All rights reserved.doi:10.1016/j.nbd.2006.08.019
of pro-inflammatory cytokines in the brain and cerebrospinal fluidof PD patients is also well documented (reviewed by Nagatsu et al.,2000).
Cytokines are important intercellular messengers involved inneuron–glia interactions and in the crosstalk between injury-activated astrocytes and microglial cells, modulating the glialresponse to injury. Interleukin-1 (IL-1) is often considered theprototypic pro-inflammatory cytokine (Rothwell and Luheshi,2000). Whereas barely detectable in healthy brain (Vitkovic et al.,2000), IL-1β levels are increased in the brain of PD patients (Mogiet al., 1994), and it has been proposed that the increased levels ofIL-1 may contribute to the pathophysiology of PD (Nagatsu andSawada, 2005). In the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyr-idine (MPTP) animal model of PD, IL-1 levels were also found tobe increased (Ho and Blum, 1998; Mogi et al., 1998; Grunblatt etal., 2001; Mandel et al., 2002; Ciesielska et al., 2003). Althoughhistorically IL-1 has been associated with a pro-inflammatory role,several studies have highlighted the role of IL-1 in the regenerationof the central nervous system. IL-1 was implicated in the inductionof different neurotrophic factors after injury, including nervegrowth factor (DeKosky et al., 1996), ciliary neurotrophic factor(Herx et al., 2000), and insulin-like growth factor-1 (Mason et al.,2001). Taken together, these data support a role for IL-1 inencouraging repair. In fact, pro-inflammatory cytokines could playa neuroprotective role in PD. For instance, striatal application ofIL-1-releasing pellets after 6-hydroxydopamine (6-OHDA)-in-duced lesion in rats increased the spontaneous recovery of striataldopaminergic innervation (Wang et al., 1994). Other studiesshowed a positive correlation between striatal IL-1α levels anddopaminergic reinnervation after MPTP (Ho and Blum, 1998;Hebert et al., 2003). Using knockout mice, IL-1 and IL-6 have alsobeen shown to be involved in dopaminergic sprouting afterneurotoxin-induced denervation (Bolin et al., 2002; Parish et al.,2002). Furthermore, intranigral infusion of IL-1β was shown toactivate astrocytes and significantly protected nigral dopaminergiccell bodies from a subsequent 6-OHDA lesion (Saura et al., 2003).
Glial cell line-derived neurotrophic factor (GDNF) is a potentfactor for the protection of nigral dopaminergic neurons against

93A. Saavedra et al. / Neurobiology of Disease 25 (2007) 92–104
toxin-induced degeneration in animal models of PD and promotesrecovery from motor deficits (Gash et al., 1996; Kordower et al.,2000; Grondin et al., 2002; Dowd et al., 2005). Therefore, GDNFrepresents a potential therapeutic tool for PD, and some clinicaltrials have confirmed the efficacy of GDNF in patients with PD(Gill et al., 2003; Patel et al., 2005). In previous studies wedemonstrated that H2O2 and L-DOPA increased GDNF proteinlevels in a concentration- and time-dependent mode in neuron–gliacultures but not in astrocyte cultures. Since conditioned mediafrom H2O2- or L-DOPA-treated neuron–glia cultures increasedGDNF mRNA and protein levels in astrocyte cultures, weproposed that diffusible modulators, likely released by damagedneurons, signal astrocytes to increase GDNF expression (Saavedraet al., 2006). To understand the mechanisms involved inneurotrophic factor up-regulation after injury, it is important toidentify the molecular mediators involved in their production. Inthis work, we show that IL-1β is involved in GDNF up-regulationafter dopaminergic injury.
Material and methods
Cell culture
Animals were handled in accordance with the national ethicalrequirements for animal research and with the EuropeanConvention for the Protection of Vertebrate Animals Used forExperimental and Other Scientific Purposes. Cells cultures wereprepared as previously described (Saavedra et al., 2005, 2006).Briefly, postnatal day 1–3 Wistar rat pups were used to prepareventral midbrain astrocyte cultures, and 75,000 cells were platedonto poly-D-lysine and laminin-coated coverslips placed under0.8 cm2 holes in the bottom of 50 mm snap-top polystyrene Petridishes (BD Falcon). Once astrocytes were confluent, 25 μM 5-fluorodeoxyuridine with 70 μM uridine (FDU) was added to theculture medium to suppress cell growth. Three days before theneuronal cell preparation, the medium from astrocyte cultureswas changed to neuronal culture medium to allow theconditioning by astrocytes. The neuronal cultures were estab-lished by plating 80,000 substantia nigra cells onto a confluentmonolayer of ventral midbrain astrocytes. The cultures were keptat 37°C in a 5% CO2, 95% air atmosphere. Neuron–glia cultureswere used after 1 week in culture, and astrocyte cultures wereused after confluence was reached. Dopaminergic neuronsaccounted for 6.9±1.2% of the total neuronal population. Thispercentage is similar to the value reported by Burke et al. (1998)for cultures without added GDNF. The day prior to celltreatment, the culture medium of either neuron–glia cultures orastrocyte cultures was replaced by serum-free neuronal culturemedium.
ELISA assay for IL-1β
IL-1β levels in the conditioned media were determined using acommercially available ELISA kit (R&D Systems), as instructedby the manufacturer. The samples and standards were added to theplates coated with a polyclonal antibody specific for rat IL-1β, andincubated for 2 h at room temperature. The wells were then washedand further incubated for 2 h at room temperature with a polyclonalantibody against rat IL-1β conjugated to horseradish peroxidase.After washing, the plates were incubated for 30 min at roomtemperature with the substrate solution, containing H2O2 and
chromogen (tetramethylbenzidine). The enzyme reaction wasstopped with a hydrochloric acid solution, and the O.D. at450 nm was recorded, with wavelength correction set to 570 nm.
Western blot
GDNF protein levels were determined by Western blot analysisof samples obtained by cell homogenization in a lysis buffercontaining 25 mM Tris, 2.5 mM EDTA, 2.5 mM EGTA, 1% TritonX-100, 1 mM DTT, 1 mM PMSF and 25 μg/ml leupeptin. Proteinconcentration was determined using the Bradford method withbovine serum albumin as standard (Bio-Rad). Usually 15–25 μgprotein were separated by SDS-PAGE using a 12% resolving gelunder reducing conditions, and electrotransferred onto polyviny-lidene fluoride membranes (Millipore). After blocking with 5%skimmed milk powder in TBS-T (0.5% Tween 20 in a 20 mM Trisand 137 mM NaCl solution), for 60 min at room temperature, themembranes were incubated overnight at 4°C with a rabbit anti-GDNF primary antibody (Santa Cruz Biotechnology; 1:1000)diluted in TBS-T containing 1% skimmed milk powder. To controlfor protein loading, the membranes were probed with a mouse anti-β-actin antibody (Sigma; 1:10,000). After rinsing, blots wereincubated for 60 min, at room temperature, with an anti-rabbitantibody conjugated to alkaline phosphatase (Amersham LifeSciences), diluted 1:20,000 in TBS-T containing 1% skimmed milkpowder. Protein bands were detected using the EnhancedChemiFluorescence (ECF) system (Amersham Life Sciences) andquantified by densitometric analysis using the Quantity Onesoftware (Bio-Rad).
Total RNA extraction and reverse transcription
Total RNA was extracted from cell cultures using the TRIzol®
reagent according to the manufacturer's protocol (Invitrogen). Theisolated RNA was dissolved in 20 μl diethylpyrocarbonate-treatedwater and stored at −80°C. To remove any residual genomic DNA,RNA samples were treated for 15 min at room temperature withDNase I (Invitrogen), which was then inactivated by incubation at65°C for 10 min. To assess RNA integrity, randomly chosensamples were analyzed in an Agilent 2100 Bioanalyzer using aRNA NanoLabChip® (RNA 6000 Nano Assay, Agilent Techno-logies). The RNA integrity number was greater than 7 in allanalyzed samples.
Single-stranded cDNAs were synthesized using TaqMan®
Reverse Transcription Reagents (Roche Molecular Systems) byincubating total RNA (0.5 μg of DNase-treated RNA) for 10 min at25°C, and 30 min at 48°C, in a final volume of 25 μl. The reactionwas stopped by incubating 5 min at 95°C. All samples were storedat −20°C until analysis. For control purposes, non-templatesamples were subjected to reverse transcription.
Real-time PCR
Real-time PCR was performed to monitor the expression ofGDNF and of a housekeeping gene, the 18S ribosomal RNA(TaqMan ribosomal RNA Control Reagents) using the TaqMantechnology, and the results analyzed with a 7900 HT SequenceDetector System (Applied Biosystems). Primers and TaqManprobe were selected from Genebank (accession number L15305)and designed using the Primer Express software (AppliedBiosystems): forward primer-5′ GACTTGGGTTTGGGCTAC-

94 A. Saavedra et al. / Neurobiology of Disease 25 (2007) 92–104
GAA 3′; reverse primer-5′ ATTGTCTCGGCCGCTTCAC 3′;TaqMan® probe-5′ 6-FAM AAGGAGGAACTGATCTTTCGA-TATTGTAGCGGTTC 3′ TAMRA. Specificity of the primers wasconfirmed by a BLAST search. The amplification reaction mixture(25 μl) contained the TaqMan® Universal PCR Master Mixcomponents and 5 μl of the cDNA sample, 300 nM of each primer,and 250 nM TaqMan® probe (Applied Biosystems). The thermalcycling conditions consisted in 10 min at 95°C, proceeding with 40cycles of 95°C for 15 s, and 60°C for 1 min. The size of the PCRproduct was determined in an Agilent 2100 Bioanalyzer using aDNA 1000 LabChip® Kit (Agilent Technologies). The GDNFmRNA levels were normalized using the 18S ribosomal RNA andexpressed relative to control using the ΔΔCt method.
MTT assessment of cell viability
After treatment, cultures were incubated with 3-(4,5-di-methylthiazal-2-yl)-2,5-diphenyl-tetrazolium bromide (MTT)(0.5 mg/ml) in Krebs medium for 60–90 min at 37°C in theincubator chamber. MTT is converted by viable cells to a water-insoluble precipitate that was dissolved in 0.04 M HCl inisopropanol, and colorimetrically quantified using a microplatereader (O.D. 570 nm; wavelength correction set to 620 nm).
Immunocytochemistry
The cells were prefixed in Minimal Essential Medium withfew drops of 4% paraformaldehyde, and fixed in 4% parafor-maldehyde for 10 min. The cells were then permeabilized with0.2% Triton X-100 in phosphate buffered saline (PBS) for10 min. Blocking was performed by incubating with 0.2% gelatinin PBS with 0.5% Tween-20 for 90 min at room temperature. Thecells were then incubated for 90 min at room temperature withthe following primary antibodies, according to the aim of theexperiment: goat anti-IL-1β (R&D Systems; 1:50), rabbit anti-IL-1RI (Santa Cruz Biotechnology; 1:50), mouse anti-glial fibrillaryassociated protein (GFAP) (Sigma; 1:800), rabbit anti-GFAP(Dako; 1:400), mouse anti-tyrosine hydroxylase (TH) (Calbio-chem; 1:10,000), rabbit anti-microtubule associated protein 2(MAP2) (Chemicon; 1:1000), mouse anti-OX-42 (Serotec; 1:50).After washing, the cells were incubated for 90 min with theappropriate secondary antibodies conjugated to Alexa Fluor® 488(Molecular Probes; 1 μg/ml) or to Alexa Fluor® 594 (MolecularProbes; 2 μg/ml). To assess the specificity of the immunostaining,the primary antibody was omitted in some coverslips. In theseconditions, no specific immunofluorescence was detected. Forquantification purposes, in experiments designed for the assess-ment of dopaminergic cell viability, 15 randomly assigned fieldswere analyzed for each condition on a Zeiss inverted microscopeunder a 200× magnification, and the cells immunoreactive to THand MAP2 counted. In each independent experiment, at least 100MAP2-positive cells were counted in each condition. Thepopulation of dopaminergic neurons was reported as thepercentage of the total neuronal population labelled by MAP2antibody. For microglial quantification, after the immunostainingprocess, the coverslips were incubated for 5 min with 1 μg/mlHoechst 33342 (Molecular Probes) in PBS. In these experiments,quantification was performed by counting, under a 400×magnification, in randomly assigned fields, the number of OX-42-positive cells and the total number of nuclei, assessed withHoechst 33342.
GDNF neutralization studies
Ventral midbrain neuron–glia cultures were incubated for 24 hwith 200 or 400 μM L-DOPA in the presence or in the absence ofthe anti-GDNF antibody (Santa Cruz Biotechnology, 0.4 μg/ml).About 10 h later, the application of the antibody was repeated toensure that only residual GDNF would be available to bind to itsreceptor. The cultures were then processed for TH and MAP2immunocytochemistry as described above.
Data analysis and statistics
Data are expressed as percentages of values obtained in controlconditions, and are presented as mean±SEM of at least threeexperiments, performed in independent cell cultures. Statisticalanalysis was performed using t-test or one-way ANOVA followedby Dunnett's or Bonferroni's test, as indicated in the figure legends.Values of P<0.05 were considered significant.
Results
H2O2 and L-DOPA increased extracellular levels of IL-1β inneuron–glia but not in astrocyte cultures
We have previously shown that H2O2 and L-DOPA inducedopaminergic cell loss in neuron–glia cultures (Saavedra et al.,2006). Since up-regulation of IL-1β expression has been shownto occur following several types of injury, we investigated theeffect of H2O2 and L-DOPA on IL-1β levels in the culture mediafrom ventral midbrain astrocyte or neuron–glia cultures. IL-1βlevels were under the detection limit in samples from astrocytecultures, both in control and challenged cultures. In neuron–gliacultures, IL-1β was also undetectable in control cultures, but itincreased in a time-dependent manner in the media from culturesincubated with H2O2 or L-DOPA (Fig. 1). In cultures challengedwith 100 μM H2O2, IL-1β peaked at 1 h (190.7±17.8 pg/ml).After 2 h, IL-1β returned to levels similar to those registered at30 min and remained stable thereafter. In L-DOPA-treatedcultures, the maximum increase was observed later andcorresponded to a higher IL-1β level than in H2O2-treatedcultures (276.0±22.4 pg/ml at 3 h).
Effects of exogenous IL-1β on GDNF levels in astrocyte and inneuron–glia cultures
In previous studies, we have shown that H2O2 and L-DOPAconcentration-dependently increase GDNF levels in neuron–gliacultures, but not in astrocyte cultures (Saavedra et al., 2006).Since we observed that IL-1β levels increased in the culturemedia from neuron–glia cultures exposed to H2O2 and L-DOPA,but not in astrocyte cultures under the same conditions, weinvestigated whether IL-1β was involved in GDNF up-regulation.We determined the effect of exogenous IL-1β on GDNF levels inthese two cell cultures. Ventral midbrain astrocyte or neuron–gliacultures were incubated for 24 h with IL-1β (0.1–50 ng/ml) andGDNF levels assessed by Western blot analysis. We found thatIL-1β up to 50 ng/ml had no effect on GDNF protein levels inastrocyte cultures (Fig. 2B), whereas 1, 10 and 50 ng/ml IL-1βsignificantly decreased GDNF cell content in neuron–gliacultures (Figs. 2A and B). The effects of 1 and 10 ng/ml IL-1β were similar, inducing a 20% reduction in GDNF levels,

Fig. 1. Effects of H2O2 and L-DOPA on IL-1β levels in the culture media of ventral midbrain neuron–glia cultures. The cells were incubated with 100 μMH2O2
or 200 μM L-DOPA for up to 6 h and the conditioned media collected and stored at −20°C until analysis. IL-1β levels in the culture media were measured usingan ELISA kit. IL-1β levels were under detection limit (n.d.—not detected) in samples from control neuron–glia cultures. Data are shown as the mean±SEM ofup to five independent experiments. Statistical analysis was performed using one-way ANOVA followed by Dunnett's test. ⁎⁎P<0.01 as compared to control.
95A. Saavedra et al. / Neurobiology of Disease 25 (2007) 92–104
whereas 50 ng/ml IL-1β reduced GDNF cell content by 40%(Fig. 2B).
The decrease in GDNF cell content could be due to GDNFrelease induced by IL-1β since IL-1 was shown to increaseneurotrophic factor release from cultured astrocytes (Gadient et al.,1990; Appel et al., 1997; Juric and Carman-Krzan, 2001; Albrechtet al., 2002). However, using an ELISA kit (Promega), we wereunable to detect GDNF in the culture media, either from control orfrom cultures incubated with 50 ng/ml IL-1β.
Given that increased IL-1β levels in the culture media wereobserved as early as 30 min after incubation with H2O2 or L-DOPA(Fig. 1), and within few minutes following binding to cells IL-1induces several biochemical events (Andre et al., 2005;
Fig. 2. Effects of exogenous IL-1β on GDNF protein (A, B) and mRNA (C) levelswith the indicated concentrations of IL-1β and protein extracts were prepared andGDNF antibody (A, B). (A) Representative Western blot for GDNF cell content in ndata by densitometric analysis. The results are expressed as percentage of controls,six independent experiments performed in triplicate. (C) Astrocyte cultures were incsample, 0.5 μg of RNAwas reverse transcribed and analyzed by real-time PCR for Gresults. The expression levels from three to four independent experiments are presStatistical analysis was performed using one-way ANOVA followed by Dunnett's
Samuelsson et al., 2005), we examined the effect of IL-1β onGDNF mRNA levels in astrocyte cultures by real-time PCR.Astrocyte cultures were incubated up to 3 h with 0.3 ng/ml IL-1β,but no significant changes were observed on GDNF mRNA levels(Fig. 2C). The IL-1β concentration 0.3 ng/ml was selected for thisstudy because it was the highest concentration tested and notinducing decrease in GDNF cell content in neuron–glia cultures(Fig. 2B).
IL-1β, even at high concentration, did not decrease cell viability
Since IL-1β is mostly considered a pro-inflammatory cytokine,we examined whether the observed reduction of GDNF cell
. Ventral midbrain astrocyte or neuron–glia cultures were incubated for 24 hseparated by SDS-PAGE followed by immunoblot analysis using an anti-euron–glia cultures exposed to IL-1β. (B) Quantification of the Western blotincubated in the absence of IL-1β. Data shown are the means±SEM of up toubated with 0.3 ng/ml IL-1β for up to 3 h, and total RNA extracted. For eachDNF and for a housekeeping gene, the 18S ribosomal RNA, to normalize theented as mean±SEM relative to controls incubated in the absence of IL-1β.test. ⁎P<0.05 and ⁎⁎P<0.01 as compared to control.
96 A. Saavedra et al. / Neurobiology of Disease 25 (2007) 92–104
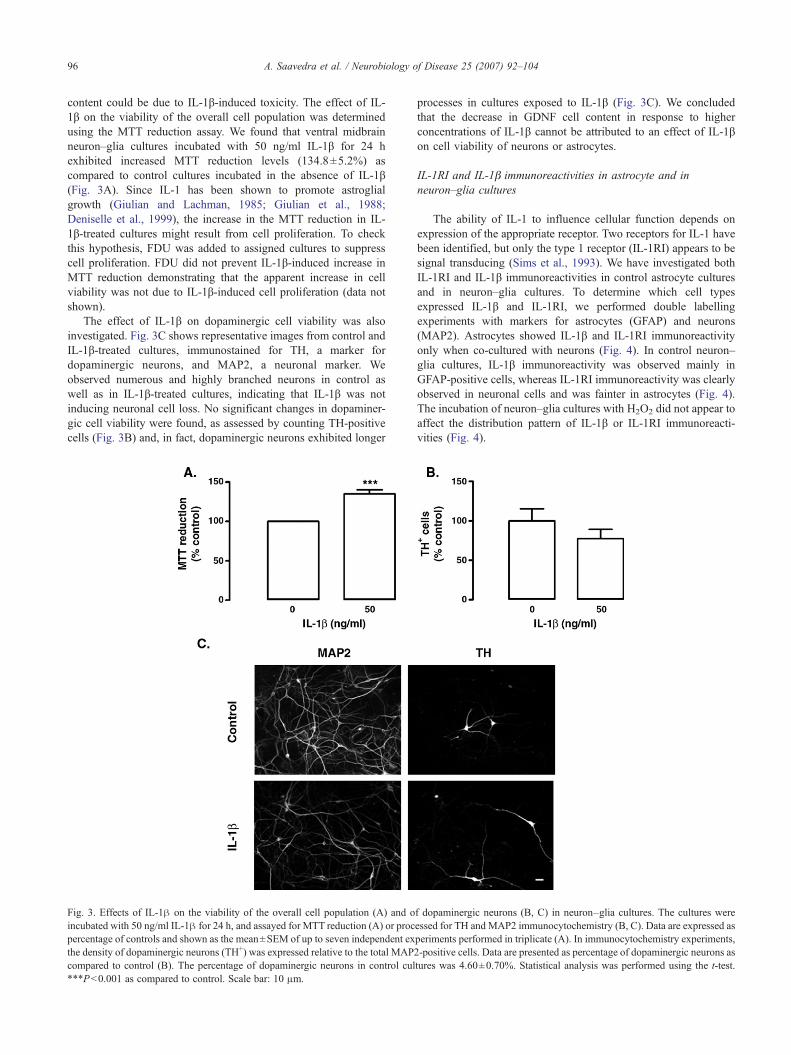
content could be due to IL-1β-induced toxicity. The effect of IL-1β on the viability of the overall cell population was determinedusing the MTT reduction assay. We found that ventral midbrainneuron–glia cultures incubated with 50 ng/ml IL-1β for 24 hexhibited increased MTT reduction levels (134.8±5.2%) ascompared to control cultures incubated in the absence of IL-1β(Fig. 3A). Since IL-1 has been shown to promote astroglialgrowth (Giulian and Lachman, 1985; Giulian et al., 1988;Deniselle et al., 1999), the increase in the MTT reduction in IL-1β-treated cultures might result from cell proliferation. To checkthis hypothesis, FDU was added to assigned cultures to suppresscell proliferation. FDU did not prevent IL-1β-induced increase inMTT reduction demonstrating that the apparent increase in cellviability was not due to IL-1β-induced cell proliferation (data notshown).
The effect of IL-1β on dopaminergic cell viability was alsoinvestigated. Fig. 3C shows representative images from control andIL-1β-treated cultures, immunostained for TH, a marker fordopaminergic neurons, and MAP2, a neuronal marker. Weobserved numerous and highly branched neurons in control aswell as in IL-1β-treated cultures, indicating that IL-1β was notinducing neuronal cell loss. No significant changes in dopaminer-gic cell viability were found, as assessed by counting TH-positivecells (Fig. 3B) and, in fact, dopaminergic neurons exhibited longer
Fig. 3. Effects of IL-1β on the viability of the overall cell population (A) and oincubated with 50 ng/ml IL-1β for 24 h, and assayed for MTT reduction (A) or procpercentage of controls and shown as the mean±SEM of up to seven independent exthe density of dopaminergic neurons (TH+) was expressed relative to the total MAPcompared to control (B). The percentage of dopaminergic neurons in control cul⁎⁎⁎P<0.001 as compared to control. Scale bar: 10 μm.
processes in cultures exposed to IL-1β (Fig. 3C). We concludedthat the decrease in GDNF cell content in response to higherconcentrations of IL-1β cannot be attributed to an effect of IL-1βon cell viability of neurons or astrocytes.
IL-1RI and IL-1β immunoreactivities in astrocyte and inneuron–glia cultures
The ability of IL-1 to influence cellular function depends onexpression of the appropriate receptor. Two receptors for IL-1 havebeen identified, but only the type 1 receptor (IL-1RI) appears to besignal transducing (Sims et al., 1993). We have investigated bothIL-1RI and IL-1β immunoreactivities in control astrocyte culturesand in neuron–glia cultures. To determine which cell typesexpressed IL-1β and IL-1RI, we performed double labellingexperiments with markers for astrocytes (GFAP) and neurons(MAP2). Astrocytes showed IL-1β and IL-1RI immunoreactivityonly when co-cultured with neurons (Fig. 4). In control neuron–glia cultures, IL-1β immunoreactivity was observed mainly inGFAP-positive cells, whereas IL-1RI immunoreactivity was clearlyobserved in neuronal cells and was fainter in astrocytes (Fig. 4).The incubation of neuron–glia cultures with H2O2 did not appear toaffect the distribution pattern of IL-1β or IL-1RI immunoreacti-vities (Fig. 4).
f dopaminergic neurons (B, C) in neuron–glia cultures. The cultures wereessed for TH and MAP2 immunocytochemistry (B, C). Data are expressed asperiments performed in triplicate (A). In immunocytochemistry experiments,2-positive cells. Data are presented as percentage of dopaminergic neurons astures was 4.60±0.70%. Statistical analysis was performed using the t-test.

Fig. 4. IL-1β and IL-1RI immunoreactivities in ventral midbrain astrocyte or neuron–glia cultures. Astrocyte (control cultures) and neuron–glia cultures (controland incubated for 24 h with 100 μM H2O2) were processed for IL-1β, IL-1RI, GFAP and MAP2 immunocytochemistry. Astrocytes showed IL-1β and IL-1RIimmunoreactivity in neuron–glia cultures but not in astrocyte cultures. Scale bar: 10 μm.
97A. Saavedra et al. / Neurobiology of Disease 25 (2007) 92–104
IL-1ra prevented GDNF up-regulation in challenged neuron–gliacultures
To further address the role of IL-1β on GDNF up-regulationafter injury, we studied the effects of the IL-1 receptor antagonist(IL-1ra) on GDNF levels in challenged ventral midbrain neuron–glia cultures. Cell cultures were incubated for 24 h with 100 μMH2O2 or 200 μM L-DOPA in the presence or in the absence of the
recombinant rat IL-1ra (10 or 100 ng/ml; R&D Systems), withoutpre-incubation, and processed for GDNF immunoblotting. In bothH2O2- and L-DOPA-treated cultures, the presence of 10 ng/ml IL-1ra prevented GDNF up-regulation: GDNF expression decreasedfrom 134.7±6.1 to 86.5±16.3% in H2O2-treated cultures and from137.9±2.9 to 103.6±4.0% in L-DOPA-treated cultures (Fig. 5). Wefound no significant differences in the effects of 10 and 100 ng/mlIL-1ra on GDNF levels in H2O2 or L-DOPA-challenged cultures,
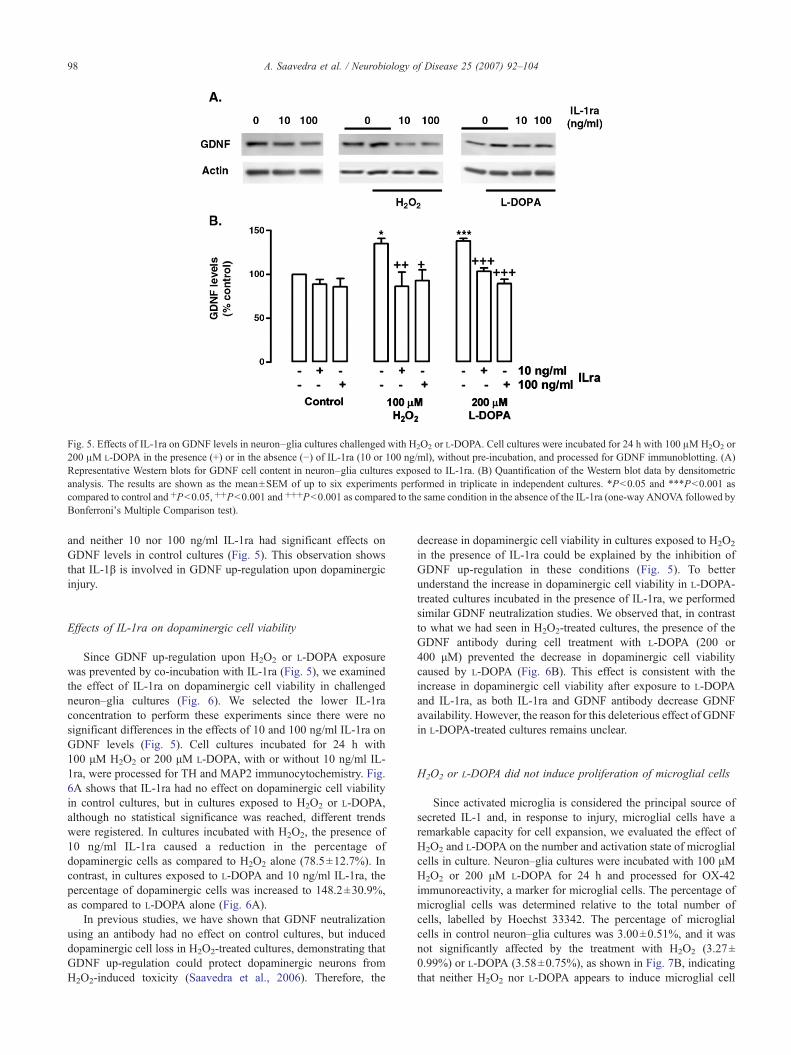
Fig. 5. Effects of IL-1ra on GDNF levels in neuron–glia cultures challenged with H2O2 or L-DOPA. Cell cultures were incubated for 24 h with 100 μMH2O2 or200 μM L-DOPA in the presence (+) or in the absence (−) of IL-1ra (10 or 100 ng/ml), without pre-incubation, and processed for GDNF immunoblotting. (A)Representative Western blots for GDNF cell content in neuron–glia cultures exposed to IL-1ra. (B) Quantification of the Western blot data by densitometricanalysis. The results are shown as the mean±SEM of up to six experiments performed in triplicate in independent cultures. ⁎P<0.05 and ⁎⁎⁎P<0.001 ascompared to control and +P<0.05, ++P<0.001 and +++P<0.001 as compared to the same condition in the absence of the IL-1ra (one-way ANOVA followed byBonferroni's Multiple Comparison test).
98 A. Saavedra et al. / Neurobiology of Disease 25 (2007) 92–104
and neither 10 nor 100 ng/ml IL-1ra had significant effects onGDNF levels in control cultures (Fig. 5). This observation showsthat IL-1β is involved in GDNF up-regulation upon dopaminergicinjury.
Effects of IL-1ra on dopaminergic cell viability
Since GDNF up-regulation upon H2O2 or L-DOPA exposurewas prevented by co-incubation with IL-1ra (Fig. 5), we examinedthe effect of IL-1ra on dopaminergic cell viability in challengedneuron–glia cultures (Fig. 6). We selected the lower IL-1raconcentration to perform these experiments since there were nosignificant differences in the effects of 10 and 100 ng/ml IL-1ra onGDNF levels (Fig. 5). Cell cultures incubated for 24 h with100 μM H2O2 or 200 μM L-DOPA, with or without 10 ng/ml IL-1ra, were processed for TH and MAP2 immunocytochemistry. Fig.6A shows that IL-1ra had no effect on dopaminergic cell viabilityin control cultures, but in cultures exposed to H2O2 or L-DOPA,although no statistical significance was reached, different trendswere registered. In cultures incubated with H2O2, the presence of10 ng/ml IL-1ra caused a reduction in the percentage ofdopaminergic cells as compared to H2O2 alone (78.5±12.7%). Incontrast, in cultures exposed to L-DOPA and 10 ng/ml IL-1ra, thepercentage of dopaminergic cells was increased to 148.2±30.9%,as compared to L-DOPA alone (Fig. 6A).
In previous studies, we have shown that GDNF neutralizationusing an antibody had no effect on control cultures, but induceddopaminergic cell loss in H2O2-treated cultures, demonstrating thatGDNF up-regulation could protect dopaminergic neurons fromH2O2-induced toxicity (Saavedra et al., 2006). Therefore, the
decrease in dopaminergic cell viability in cultures exposed to H2O2
in the presence of IL-1ra could be explained by the inhibition ofGDNF up-regulation in these conditions (Fig. 5). To betterunderstand the increase in dopaminergic cell viability in L-DOPA-treated cultures incubated in the presence of IL-1ra, we performedsimilar GDNF neutralization studies. We observed that, in contrastto what we had seen in H2O2-treated cultures, the presence of theGDNF antibody during cell treatment with L-DOPA (200 or400 μM) prevented the decrease in dopaminergic cell viabilitycaused by L-DOPA (Fig. 6B). This effect is consistent with theincrease in dopaminergic cell viability after exposure to L-DOPAand IL-1ra, as both IL-1ra and GDNF antibody decrease GDNFavailability. However, the reason for this deleterious effect of GDNFin L-DOPA-treated cultures remains unclear.
H2O2 or L-DOPA did not induce proliferation of microglial cells
Since activated microglia is considered the principal source ofsecreted IL-1 and, in response to injury, microglial cells have aremarkable capacity for cell expansion, we evaluated the effect ofH2O2 and L-DOPA on the number and activation state of microglialcells in culture. Neuron–glia cultures were incubated with 100 μMH2O2 or 200 μM L-DOPA for 24 h and processed for OX-42immunoreactivity, a marker for microglial cells. The percentage ofmicroglial cells was determined relative to the total number ofcells, labelled by Hoechst 33342. The percentage of microglialcells in control neuron–glia cultures was 3.00±0.51%, and it wasnot significantly affected by the treatment with H2O2 (3.27±0.99%) or L-DOPA (3.58±0.75%), as shown in Fig. 7B, indicatingthat neither H2O2 nor L-DOPA appears to induce microglial cell

Fig. 6. Effects of IL-1ra (A) and GDNF neutralization (B) on dopaminergiccell viability in neuron–glia cultures challenged with H2O2 or L-DOPA. Cellcultures were incubated for 24 h with 100 μM H2O2 or 200 μM L-DOPA inthe presence or in the absence of IL-1ra (10 ng/ml), without pre-incubation(A), or in the presence (+) or in the absence (−) of anti-GDNF antibody (B),and processed for TH and MAP2 immunocytochemistry. The density ofdopaminergic neurons (TH+) was evaluated as percentage of MAP2immunoreactive cells. Data are presented as percentage of dopaminergicneurons as compared to control (B) or to the same condition in the absenceof IL-1ra (A), and shown as the mean±SEM of at least three experimentsperformed in independent cultures. ⁎P<0.05, ⁎⁎⁎P<0.01 as compared tocontrol, and ++P<0.01 and +++P<0.001 as compared to the same conditionin the absence of the neutralizing antibody (one-way ANOVA followed byBonferroni's Multiple Comparison test).
99A. Saavedra et al. / Neurobiology of Disease 25 (2007) 92–104
proliferation. Microglial cells appeared in an intermediate state ofactivation, exhibiting large cell bodies with multiple, thicklamellipodia. Moreover, no obvious morphological changes wereobserved in challenged cultures as compared to control cultures(Fig. 7A). We did not observe ameboid-shaped microglial cells incultures treated with H2O2 or L-DOPA suggesting that there wasnot a strong microglial reaction. Since astrocytes were the vastmajority of cells expressing IL-1β in our cultures (Fig. 4), theincrease in IL-1β levels observed in conditioned media fromchallenged neuron–glia cultures was likely a contribution fromastrocytes.
Discussion
In previous studies, we showed that the selective injury todopaminergic neurons with H2O2 or L-DOPA increased GDNFexpression in neuron–glia cultures, but not in astrocyte cultures.Since the conditioned media from challenged neuron–glia cultures
increased GDNF in astrocyte cultures, we proposed the involve-ment of diffusible modulators in signalling astrocytes to increaseGDNF expression (Saavedra et al., 2006). Here we show that IL-1βis involved in GDNF up-regulation in response to H2O2 or L-DOPA in ventral midbrain cell cultures, suggesting a neuropro-tective role for this cytokine, in addition to its more oftenrecognized deleterious action.
Neuronal injury induced IL-1β release
In this work, IL-1β was not detectable in the media from controlcultures, but marked increases were observed in the media fromneuron–glia cultures exposed to H2O2 or L-DOPA (Fig. 1). Theseresults are consistent with other works showing an increase in IL-1expression following injury to the nigrostriatal pathway (Ho andBlum, 1998; Mogi et al., 1998; Grunblatt et al., 2001; Mandel etal., 2002; Ciesielska et al., 2003; Hebert et al., 2003). Theseevidences implicate IL-1 in the process of neuronal death and/orrepair and recovery.
In contrast, H2O2 and L-DOPA did not increase IL-1β in mediafrom astrocyte cultures, in agreement with the lack of IL-1βimmunoreactivity in these cultures (Fig. 4). We found thatastrocytes do not express IL-1β or the IL-1RI unless they arecultured in the presence of neurons (Fig. 4). In neuron–gliacultures, IL-1β immunoreactivity was observed mainly in astro-cytes, both in control and in challenged cultures (Fig. 4), but IL-1βwas only detectable in the media from challenged cultures (Fig. 1).This suggests that IL-1β release from astrocytes is induced byinjured neurons. The observation that IL-1β immunoreactivity wasmainly found in astrocytes is consistent with previous reportssupporting the ability of astrocytes to produce IL-1 (Giulian et al.,1986; Sairanen et al., 1997; Mason et al., 2001).
The expression of pro-inflammatory cytokines has always beenassociated with microglial activation, but there are evidences thatastrocytes, as well as neurons, can synthesize IL-1 (Fontana et al.,1982; Lieberman et al., 1989; Tchelingerian et al., 1993; Nieto-Sampedro and Berman, 1987; Lemke et al., 1999; Acarin et al.,2000). In our model, IL-1β immunoreactivity was found mainly inastrocytes present in neuron–glia cultures (Fig. 4). In addition, wefound increased IL-1β levels in the culture media from challengedneuron–glia cultures in conditions under which neither H2O2 norL-DOPA induced an increase in the number of microglial cells(Fig. 7B), nor ameboid-shaped microglial cells were observed.These observations suggest that there is not a strong microglialreaction in our model. Since microglial cells accounted for onlyabout 3% of the total number of cells, both in control and inchallenged cultures, they are a minor population as compared toastrocytes, suggesting that their contribution to IL-1β production isnot substantial. The presence of microglia in an intermediate stateof activation in our postnatal cell culture, even in controlconditions (Fig. 7A), is likely related to the fact that in theperinatal period microglia is not yet at a resting state (Hanisch,2002).
IL-1β is involved in GDNF up-regulation after dopaminergic injury
Several lines of evidence in this work support the idea that IL-1β is involved in GDNF up-regulation: (i) the time-course of IL-1βaccumulation in the media paralleled the increases in GDNFmRNA upon H2O2 or L-DOPA challenge observed previously(Saavedra et al., 2006); (ii) IL-1ra prevented GDNF up-regulation

Fig. 7. Effects of H2O2 and L-DOPA on microglial cells in culture. Neuron–glia cultures were incubated with 100 μM H2O2 or 200 μM L-DOPA for 24 h andprocessed for OX-42 immunocytochemistry. (A) Effects on microglial activation. Scale bar: 10 μm. (B) Effects on the density of microglial cells in culture. Afterexposure to H2O2 or L-DOPA, cells were processed for OX-42 immunocytochemistry (OX-42+) and the percentage of microglial cells determined relative to thetotal number of cells assessed with Hoechst 33342. The data shown are the percentages of microglial cells relative to control and expressed as mean±SEM of atleast three experiments performed in independent cultures. The percentage of microglial cells in control neuron–glia cultures was 3.00±0.51%, and it was notsignificantly affected by the treatment with H2O2 (3.27±0.99%) or L-DOPA (3.58±0.75%).
100 A. Saavedra et al. / Neurobiology of Disease 25 (2007) 92–104
in response to H2O2 or L-DOPA (Fig. 5); (iii) the effect of IL-1raon dopaminergic cell viability in challenged cultures (Fig. 6A) wassimilar to the effect of GDNF neutralization (Fig. 6B and Saavedraet al., 2006).
However, we did not observe any increase in GDNF mRNA orprotein levels in response to exogenous IL-1β, either in astrocyte orneuron–glia cultures (Fig. 2). In fact, GDNF protein levelsdecreased in neuron–glia cultures exposed to IL-1β concentrationshigher than 0.3 ng/ml (Fig. 2). This might be due to GDNF releaseinduced by IL-1β since IL-1 induction of neurotrophic factorsynthesis was also shown to be accompanied by an increase inprotein secretion (Lindholm et al., 1987; Spranger et al., 1990;Araujo and Cotman, 1992) and IL-1β was shown to induce GDNFsynthesis/release in mouse cultured astrocytes (Appel et al., 1997),rat C6 glioblastoma cells (Appel et al., 1997; Verity et al., 1998)and in a human glioblastoma cell line (Verity et al., 1999).Nevertheless, we were unable to detect GDNF in the culture mediafrom IL-1β-treated cultures using an ELISA assay.
The lack of IL-1RI immunoreactivity in control astrocytecultures (Fig. 4) is in agreement with reports showing that few IL-1RI-positive glial cells were detected in control astrocyte cultures orin the normal adult brain (Ericsson et al., 1995; Sairanen et al., 1997;Friedman, 2001). Yet astrocytes proliferate and produce cytokinesand growth factors in response to IL-1, indicating that the expressionof the receptor may be induced. IL-1 binding using radiolabelledligands has been detected in mouse astrocytes in culture (Ban et al.,1993; Rubio, 1994), and in cultured rat astrocytes IL-1RI mRNAand protein expression was induced by IL-1β itself (Friedman,2001). Therefore, we can conclude that the lack of effect of IL-1β on
GDNF levels in astrocyte cultures (Fig. 2B) is not related to theabsence of IL-1RI immunoreactivity in astrocytes (Fig. 4).
The lack of effect of exogenous IL-1β on GDNF expression inastrocyte or neuron–glia cultures (Fig. 2) is not incompatible withour proposal of the involvement of IL-1β on GDNF up-regulation,which is supported by the effect of IL-1ra on GDNF levels inchallenged cultures (Fig. 5). Other factors released into the mediumin response to dopaminergic injury might be required to the effectof IL-1β on GDNF expression. If other factors, in addition to IL-1β, are required, IL-1ra is expected to prevent GDNF up-regulation, but the addition of exogenous IL-1β to control cultureswould not increase GDNF expression. In some systems, theinduction of growth factors by IL-1β has been shown to require thesynergistic effect of other cytokines (Gadient et al., 1990; Takamiet al., 2005).
The signalling pathways of IL-1 are compatible with theinduction of GDNF transcription. Activation of two signallingpathways, the mitogen-activated protein kinase signalling cascadesand the nuclear factor kappa-B (NF-κB) pathway, have emerged asmajor routes through which IL-1RI stimulation results in genetranscription events (O’Neill and Greene, 1998). NF-κB activationin glia mediates the regulation of cytokines and growth factors byIL-1 (Sparacio et al., 1992; Friedman et al., 1996; Heese et al.,1998; Andre et al., 2005). Moreover, GDNF transcripts were foundto increase after IL-1β treatment and the NF-κB binding site wassuggested to be involved in the response of the mouse GDNF geneto cytokines (Tanaka et al., 2000). In addition, a consensus NF-κBbinding site was also identified in the human GDNF promoter(Woodbury et al., 1998; Baecker et al., 1999).

Fig. 8. Hypothetical model for the involvement of IL-1β on GDNF up-regulation upon dopaminergic injury in ventral midbrain cultures. Damageddopaminergic (DA) neurons induce IL-1β release from astrocytes. Howcompromised dopaminergic neurons signal astrocytes is not clear butdiffusible signals are involved since we have shown that conditioned mediafrom challenged neuron–glia cultures have the ability to up-regulate GDNFexpression in astrocyte cultures (Saavedra et al., 2006). IL-1β, acting in anautocrine or a paracrine way, binds to IL-1 receptors present in astrocytesand triggers GDNF up-regulation. The role of microglia needs furtherclarification.
101A. Saavedra et al. / Neurobiology of Disease 25 (2007) 92–104
IL-1β and dopaminergic cell viability
IL-1β is considered a pro-inflammatory cytokine proposed tocontribute to neuronal injury. However, we did not observe anyevidence for IL-1β-induced toxicity in our cultures. On thecontrary, exposure to IL-1β increased cell viability as assessed byMTT reduction (Fig. 3A). This effect was not due to cellproliferation since it was not prevented by the mitotic inhibitorFDU. Furthermore, immunocytochemistry studies showed noevidence for damage to MAP2- or TH-positive neurons. Actually,no significant changes in dopaminergic cell density were observed(Fig. 3B), and dopaminergic neurons exhibited longer processes incultures exposed to IL-1β (Fig. 3C). A neurotrophic effect of IL-1β was also observed by Akaneya et al. (1995) who demonstratedthat this cytokine has a survival promoting effect on embryonicdopaminergic neurons. In the intact nigrostriatal system, IL-1β isnot toxic since the number of TH-positive cell bodies and fibersdid not change in the substantia nigra after injection of IL-1β(Castano et al., 2002; Depino et al., 2005). Furthermore,intranigral infusion of IL-1β protected dopaminergic cell bodiesfrom a subsequent 6-OHDA lesion (Saura et al., 2003). Moreover,IL-1 is involved in dopaminergic sprouting after neurotoxin-induced denervation (Parish et al., 2002). The data shown in thepresent report suggest that these protective effects might involveGDNF up-regulation.
The lack of IL-1β-induced toxicity was also observed in othersystems where IL-1 up to 100 ng/ml was not toxic (Carlson et al.,1999), and IL-1 was in fact shown to protect neurons fromexcitotoxicity (Strijbos and Rothwell, 1995; Carlson et al., 1999).Using in vivo models, other studies have shown that administrationof IL-1 does not by itself cause neuronal damage (Yamasaki et al.,1995; Loddick and Rothwell, 1996; Lawrence et al., 1998; Daviset al., 2005).
The effect of IL-1ra on dopaminergic cell viability in H2O2-treated cultures is compatible with a neuroprotective role for IL-1β,but in cultures incubated with L-DOPA the presence of IL-1raimproved the viability of dopaminergic neurons (Fig. 6A). Thisdifferential effect of IL-1ra on dopaminergic cell viability ispeculiar as the effect on GDNF levels was similar in H2O2- and inL-DOPA-treated cultures (Fig. 5). However, this unexpected resultwas similar to the effect of GDNF neutralization on the viability ofdopaminergic cells in L-DOPA-treated cultures (Fig. 6B). Thereason for this observation is not clear, but it might be related to anoversupply of dopamine in L-DOPA-treated cultures. In the intactnigrostriatal system, GDNF has been shown to increase dopaminelevels (Hudson et al., 1995; Beck et al., 1996; Martin et al., 1996;Xu and Dluzen, 2000). GDNF was also shown to up-regulate THexpression at both mRNA and protein level, to delay thedegradation of TH mRNA and to enhance TH enzymatic activity(Xiao et al., 2002). In addition, GDNF significantly increases THphosphorylation, which is known to increase TH activity(Salvatore et al., 2001; Lindgren et al., 2002; Kobori et al.,2004). Therefore, we propose that GDNF up-regulation in responseto L-DOPA can increase TH levels and/or activity resulting inenhanced production of dopamine that, together with the dopamineresulting from the decarboxylation of exogenous L-DOPA, inducedtoxicity to dopaminergic neurons. Therefore, neutralizing GDNF orpreventing its up-regulation with the IL-1ra protected dopaminer-gic neurons from L-DOPA-induced toxicity (Fig. 6).
In summary, although evidences exist from other reports thatIL-1β induces GDNF synthesis/release (Appel et al., 1997; Verity
et al., 1998, 1999), exogenous IL-1β was used and some of theseworks were performed in cell lines. In contrast, in the present work,we show the involvement of endogenously produced IL-1β inGDNF up-regulation after mild toxic damage to dopaminergicneurons in vitro. Since IL-1β was only detected in the media fromchallenged neuron–glia cultures, and astrocytes are the vastmajority of cells expressing IL-1β in the cultures, we propose thatdamaged neurons triggered IL-1β release from astrocytes (Fig. 8).Once released, IL-1β can bind to its receptors present both inneurons and in astrocytes and increase GDNF expression as aneuroprotective strategy. Therefore, IL-1β might have neuropro-tective effects mediated by GDNF up-regulation. The knowledgeof the mechanisms involved in GDNF expression and release andthe identification of the intercellular mediators involved in theneuron–glia crosstalk upon dopaminergic injury might revealtargets for the development of therapeutic tools to use in thetreatment of PD patients.
Acknowledgments
We are grateful to Dr. Célia Aveleira (Center of Ophtalmologyof Coimbra, IBILI, Faculty of Medicine, University of Coimbra,Portugal) for providing the IL-1β and the IL-1RI antibodies and toDr. Paulo Santos (Center for Histocompatibility, Coimbra,Portugal) for his help with the real-time PCR experiments. Thiswork was supported by the Bissaya Barreto Foundation and theFoundation for Science and Technology, Portugal (Grant SFRH/BD/5337/2001 to Ana Saavedra).
References
Acarin, L., Gonzalez, B., Castellano, B., 2000. Neuronal, astroglial andmicroglial cytokine expression after an excitotoxic lesion in theimmature rat brain. Eur. J. Neurosci. 12, 3505–3520.

102 A. Saavedra et al. / Neurobiology of Disease 25 (2007) 92–104
Akaneya, Y., Takahashi,M., Hatanaka, H., 1995. Interleukin-1 beta enhancessurvival and interleukin-6 protects against MPP+ neurotoxicity incultures of fetal rat dopaminergic neurons. Exp. Neurol. 136, 44–52.
Albrecht, P.J., Dahl, J.P., Stoltzfus, O.K., Levenson, R., Levison, S.W.,2002. Ciliary neurotrophic factor activates spinal cord astrocytes,stimulating their production and release of fibroblast growth factor-2,to increase motor neuron survival. Exp. Neurol. 173, 46–62.
Andre, R., Pinteaux, E., Kimber, I., Rothwell, N.J., 2005. Differentialactions of IL-1 alpha and IL-1 beta in glial cells share common IL-1signalling pathways. NeuroReport 16, 153–157.
Appel, E., Kolman, O., Kazimirsky, G., Blumberg, P.M., Brodie, C., 1997.Regulation of GDNF expression in cultured astrocytes by inflammatorystimuli. NeuroReport 8, 3309–3312.
Araujo, D.M., Cotman, C.W., 1992. Basic FGF in astroglial, microglial, andneuronal cultures: characterization of binding sites and modulation ofrelease by lymphokines and trophic factors. J. Neurosci. 12, 1668–1678.
Baecker, P.A., Lee, W.H., Verity, A.N., Eglen, R.M., Johnson, R.M., 1999.Characterization of a promoter for the human glial cell line-derivedneurotrophic factor gene. Brain Res. Mol. Brain Res. 69, 209–222.
Ban, E.M., Sarlieve, L.L., Haour, F.G., 1993. Interleukin-1 binding sites onastrocytes. Neuroscience 52, 725–733.
Beck, K.D., Irwin, I., Valverde, J., Brennan, T.J., Langston, J.W., Hefti, F.,1996. GDNF induces a dystonia-like state in neonatal rats and stimulatesdopamine and serotonin synthesis. Neuron 16, 665–673.
Bolin, L.M., Strycharska-Orczyk, I., Murray, R., Langston, J.W., Di, MD.,2002. Increased vulnerability of dopaminergic neurons in MPTP-lesioned interleukin-6 deficient mice. J. Neurochem. 83, 167–175.
Burke, R.E., Antonelli, M., Sulzer, D., 1998. Glial cell line-derivedneurotrophic growth factor inhibits apoptotic death of postnatalsubstantia nigra dopamine neurons in primary culture. J. Neurochem.71, 517–525.
Carlson, N.G., Wieggel, W.A., Chen, J., Bacchi, A., Rogers, S.W., Gahring,L.C., 1999. Inflammatory cytokines IL-1 alpha, IL-1 beta, IL-6, andTNF-alpha impart neuroprotection to an excitotoxin through distinctpathways. J. Immunol. 163, 3963–3968.
Castano, A., Herrera, A.J., Cano, J., Machado, A., 2002. The degenerativeeffect of a single intranigral injection of LPS on the dopaminergicsystem is prevented by dexamethasone, and not mimicked by rh-TNF-alpha, IL-1beta and IFN-gamma. J. Neurochem. 81, 150–157.
Ciesielska, A., Joniec, I., Przybylkowski, A., Gromadzka, G., Kurkowska-Jastrzebska, I., Czlonkowska, A., Czlonkowski, A., 2003. Dynamics ofexpression of the mRNA for cytokines and inducible nitric synthase in amurine model of the Parkinson’s disease. Acta Neurobiol. Exp. (Wars)63, 117–126.
Davis, A.E., Campbell, S.J., Wilainam, P., Anthony, D.C., 2005. Post-conditioning with lipopolysaccharide reduces the inflammatory infiltrateto the injured brain and spinal cord: a potential neuroprotectivetreatment. Eur. J. Neurosci. 22, 2441–2450.
DeKosky, S.T., Styren, S.D., O'Malley, M.E., Goss, J.R., Kochanek, P.,Marion, D., Evans, C.H., Robbins, P.D., 1996. Interleukin-1 receptorantagonist suppresses neurotrophin response in injured rat brain. Ann.Neurol. 39, 123–127.
Deniselle,M.C., Lavista-Llanos, S., Ferrini,M.G., Lima, A.E., Roldan, A.G.,De Nicola, A.F., 1999. In vitro differences between astrocytes of controland wobbler mice spinal cord. Neurochem. Res. 24, 1535–1541.
Depino, A., Ferrari, C., Pott Godoy, M.C., Tarelli, R., Pitossi, F.J., 2005.Differential effects of interleukin-1beta on neurotoxicity, cytokineinduction and glial reaction in specific brain regions. J. Neuroimmunol.168, 96–110.
Dowd, E., Monville, C., Torres, E.M., Wong, L.F., Azzouz, M., Mazarakis,N.D., Dunnett, S.B., 2005. Lentivector-mediated delivery of GDNFprotects complex motor functions relevant to human Parkinsonism in arat lesion model. Eur. J. Neurosci. 22, 2587–2595.
Ericsson, A., Liu, C., Hart, R.P., Sawchenko, P.E., 1995. Type 1 interleukin-1 receptor in the rat brain: distribution, regulation, and relationship tosites of IL-1-induced cellular activation. J. Comp. Neurol. 361,681–698.
Fontana, A., Kristensen, F., Dubs, R., Gemsa, D., Weber, E., 1982.Production of prostaglandin E and an interleukin-1 like factor bycultured astrocytes and C6 glioma cells. J. Immunol. 129, 2413–2419.
Friedman, W.J., 2001. Cytokines regulate expression of the type 1interleukin-1 receptor in rat hippocampal neurons and glia. Exp. Neurol.168, 23–31.
Friedman, W.J., Thakur, S., Seidman, L., Rabson, A.B., 1996. Regulation ofnerve growth factor mRNA by interleukin-1 in rat hippocampalastrocytes is mediated by NFkappaB. J. Biol. Chem. 271, 31115–31120.
Gadient, R.A., Cron, K.C., Otten, U., 1990. Interleukin-1 beta and tumornecrosis factor-alpha synergistically stimulate nerve growth factor(NGF) release from cultured rat astrocytes. Neurosci. Lett. 117,335–340.
Gash, D.M., Zhang, Z., Ovadia, A., Cass, W.A., Yi, A., Simmerman, L.,Russell, D., Martin, D., Lapchak, P.A., Collins, F., Hoffer, B.J.,Gerhardt, G.A., 1996. Functional recovery in parkinsonian monkeystreated with GDNF. Nature 380, 252–255.
Gill, S.S., Patel, N.K., Hotton, G.R., O'Sullivan, K., McCarter, R., Bunnage,M., Brooks, D.J., Svendsen, C.N., Heywood, P., 2003. Direct braininfusion of glial cell line-derived neurotrophic factor in Parkinsondisease. Nat. Med. 9, 589–595.
Giulian, D., Lachman, L.B., 1985. Interleukin-1 stimulation of astroglialproliferation after brain injury. Science 228, 497–499.
Giulian, D., Baker, T.J., Shih, L.C., Lachman, L.B., 1986. Interleukin 1 ofthe central nervous system is produced by ameboid microglia. J. Exp.Med. 164, 594–604.
Giulian, D., Young, D.G., Woodward, J., Brown, D.C., Lachman, L.B.,1988. Interleukin-1 is an astroglial growth factor in the developing brain.J. Neurosci. 8, 709–714.
Grondin, R., Zhang, Z., Yi, A., Cass, W.A., Maswood, N., Andersen, A.H.,Elsberry, D.D., Klein, M.C., Gerhardt, G.A., Gash, D.M., 2002.Chronic, controlled GDNF infusion promotes structural and functionalrecovery in advanced parkinsonian monkeys. Brain 125, 2191–2201.
Grunblatt, E., Mandel, S., Maor, G., Youdim, M.B., 2001. Gene expressionanalysis in N-methyl-4-phenyl-1,2,3,6-tetrahydropyridine mice modelof Parkinson’s disease using cDNA microarray: effect of R-apomor-phine. J. Neurochem. 78, 1–12.
Hanisch, U.K., 2002. Microglia as a source and target of cytokines. Glia 40,140–155.
Hebert, G., Arsaut, J., Dantzer, R., Demotes-Mainard, J., 2003. Time-courseof the expression of inflammatory cytokines and matrix metalloprotei-nases in the striatum and mesencephalon of mice injected with 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine, a dopaminergic neurotoxin. Neu-rosci. Lett. 349, 191–195.
Heese, K., Hock, C., Otten, U., 1998. Inflammatory signals induceneurotrophin expression in human microglial cells. J. Neurochem. 70,699–707.
Herx, L.M., Rivest, S., Yong, V.W., 2000. Central nervous system-initiated inflammation and neurotrophism in trauma: IL-1 beta isrequired for the production of ciliary neurotrophic factor. J. Immunol.165, 2232–2239.
Ho, A., Blum, M., 1998. Induction of interleukin-1 associated withcompensatory dopaminergic sprouting in the denervated striatum ofyoung mice: model of aging and neurodegenerative disease. J. Neurosci.18, 5614–5629.
Hudson, J., Granholm, A.C., Gerhardt, G.A., Henry, M.A., Hoffman, A.,Biddle, P., Leela, N.S., Mackerlova, L., Lile, J.D., Collins, F., 1995.Glial cell line-derived neurotrophic factor augments midbrain dopami-nergic circuits in vivo. Brain Res. Bull. 36, 425–432.
Juric, D.M., Carman-Krzan, M., 2001. Interleukin-1 beta, but not IL-1 alpha,mediates nerve growth factor secretion from rat astrocytes via type I IL-1receptor. Int. J. Dev. Neurosci. 19, 675–683.
Kobori, N., Waymire, J.C., Haycock, J.W., Clifton, G.L., Dash, P.K., 2004.Enhancement of tyrosine hydroxylase phosphorylation and activity byglial cell line-derived neurotrophic factor. J. Biol. Chem. 279, 2182–2191.
Kordower, J.H., Emborg, M.E., Bloch, J., Ma, S.Y., Chu, Y., Leventhal, L.,McBride, J., Chen, E.Y., Palfi, S., Roitberg, B.Z., Brown, W.D., Holden,

103A. Saavedra et al. / Neurobiology of Disease 25 (2007) 92–104
J.E., Pyzalski, R., Taylor, M.D., Carvey, P., Ling, Z., Trono, D.,Hantraye, P., Deglon, N., Aebischer, P., 2000. Neurodegenerationprevented by lentiviral vector delivery of GDNF in primate models ofParkinson’s disease. Science 290, 767–773.
Lawrence, C.B., Allan, S.M., Rothwell, N.J., 1998. Interleukin-1beta andthe interleukin-1 receptor antagonist act in the striatum to modifyexcitotoxic brain damage in the rat. Eur. J. Neurosci. 10, 1188–1195.
Lemke, R., Hartlage-Rubsamen, M., Schliebs, R., 1999. Differential injury-dependent glial expression of interleukins-1 alpha, beta, and interleukin-6 in rat brain. Glia 27, 75–87.
Lieberman, A.P., Pitha, P.M., Shin, H.S., Shin, M.L., 1989. Production oftumor necrosis factor and other cytokines by astrocytes stimulated withlipopolysaccharide or a neurotropic virus. Proc. Natl. Acad. Sci. U. S. A.86, 6348–6352.
Lindgren, N., Goiny, M., Herrera-Marschitz, M., Haycock, J.W., Hokfelt,T., Fisone, G., 2002. Activation of extracellular signal-regulatedkinases 1 and 2 by depolarization stimulates tyrosine hydroxylasephosphorylation and dopamine synthesis in rat brain. Eur. J. Neurosci.15, 769–773.
Lindholm, D., Heumann, R., Meyer, M., Thoenen, H., 1987. Interleukin-1regulates synthesis of nerve growth factor in non-neuronal cells of ratsciatic nerve. Nature 330, 658–659.
Loddick, S.A., Rothwell, N.J., 1996. Neuroprotective effects of humanrecombinant interleukin-1 receptor antagonist in focal cerebral ischae-mia in the rat. J. Cereb. Blood Flow Metab. 16, 932–940.
Mandel, S., Grunblatt, E., Maor, G., Youdim, M.B., 2002. Early and lategene changes in MPTP mice model of Parkinson’s disease employingcDNA microarray. Neurochem. Res. 27, 1231–1243.
Martin, D., Miller, G., Cullen, T., Fischer, N., Dix, D., Russell, D., 1996.Intranigral or intrastriatal injections of GDNF: effects on monoaminelevels and behavior in rats. Eur. J. Pharmacol. 317, 247–256.
Mason, J.L., Suzuki, K., Chaplin, D.D., Matsushima, G.K., 2001.Interleukin-1beta promotes repair of the CNS. J. Neurosci. 21,7046–7052.
Mogi, M., Harada, M., Kondo, T., Riederer, P., Inagaki, H., Minami,M., Nagatsu, T., 1994. Interleukin-1 beta, interleukin-6, epidermalgrowth factor and transforming growth factor-alpha are elevated inthe brain from parkinsonian patients. Neurosci. Lett. 180,147–150.
Mogi, M., Togari, A., Ogawa, M., Ikeguchi, K., Shizuma, N., Fan, D.,Nakano, I., Nagatsu, T., 1998. Effects of repeated systemic administra-tion of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) to miceon interleukin-1beta and nerve growth factor in the striatum. Neurosci.Lett. 250, 25–28.
Nagatsu, T., Sawada, M., 2005. Inflammatory process in Parkinson’sdisease: role for cytokines. Curr. Pharm. Des. 11, 999–1016.
Nagatsu, T., Mogi, M., Ichinose, H., Togari, A., 2000. Cytokines inParkinson’s disease. J. Neural Transm. 143–151.
Nieto-Sampedro, M., Berman, M.A., 1987. Interleukin-1-like activity in ratbrain: sources, targets, and effect of injury. J. Neurosci. Res. 17,214–219.
O'Neill, L.A., Greene, C., 1998. Signal transduction pathways activated bythe IL-1 receptor family: ancient signaling machinery in mammals,insects, and plants. J. Leukocyte Biol. 63, 650–657.
Parish, C.L., Finkelstein, D.I., Tripanichkul, W., Satoskar, A.R., Drago, J.,Horne, M.K., 2002. The role of interleukin-1, interleukin-6, and glia ininducing growth of neuronal terminal arbors in mice. J. Neurosci. 22,8034–8041.
Patel, N.K., Bunnage, M., Plaha, P., Svendsen, C.N., Heywood, P., Gill,S.S., 2005. Intraputamenal infusion of glial cell line-derived neuro-trophic factor in PD: a two-year outcome study. Ann. Neurol. 57,298–302.
Rothwell, N.J., Luheshi, G.N., 2000. Interleukin 1 in the brain: biology,pathology and therapeutic target. Trends Neurosci. 23, 618–625.
Rubio, N., 1994. Demonstration of the presence of an interleukin-1 receptoron the surface of murine astrocytes and its regulation by cytokines andTheiler's virus. Immunology 82, 178–183.
Saavedra, A., Baltazar, G., Carvalho, C.M., Duarte, E.P., 2005. GDNFmodulates HO-1 expression in substantia nigra postnatal cell cultures.Free Radical Biol. Med. 39, 1611–1619.
Saavedra, A., Baltazar, G., Santos, P., Carvalho, C.M., Duarte, E.P., 2006.Selective injury to dopaminergic neurons up-regulates GDNF insubstantia nigra postnatal cell cultures: role of neuron–glia crosstalk.Neurobiol. Dis. 23, 533–542.
Sairanen, T.R., Lindsberg, P.J., Brenner, M., Siren, A.L., 1997. Globalforebrain ischemia results in differential cellular expression ofinterleukin-1beta (IL-1beta) and its receptor at mRNA and proteinlevel. J. Cereb. Blood Flow Metab. 17, 1107–1120.
Salvatore, M.F., Waymire, J.C., Haycock, J.W., 2001. Depolarization-stimulated catecholamine biosynthesis: involvement of protein kinasesand tyrosine hydroxylase phosphorylation sites in situ. J. Neurochem.79, 349–360.
Samuelsson, M., Fisher, L., Iverfeldt, K., 2005. beta-Amyloid andinterleukin-1beta induce persistent NF-kappaB activation in rat primaryglial cells. Int. J. Mol. Med. 16, 449–453.
Saura, J., Pares, M., Bove, J., Pezzi, S., Alberch, J., Marin, C., Tolosa, E.,Marti, M.J., 2003. Intranigral infusion of interleukin-1beta activatesastrocytes and protects from subsequent 6-hydroxydopamine neurotox-icity. J. Neurochem. 85, 651–661.
Sims, J.E., Gayle, M.A., Slack, J.L., Alderson, M.R., Bird, T.A., Giri, J.G.,Colotta, F., Re, F., Mantovani, A., Shanebeck, K., 1993. Interleukin 1signaling occurs exclusively via the type I receptor. Proc. Natl. Acad.Sci. U. S. A. 90, 6155–6159.
Sparacio, S.M., Zhang, Y., Vilcek, J., Benveniste, E.N., 1992. Cytokineregulation of interleukin-6 gene expression in astrocytes involvesactivation of an NF-kappa B-like nuclear protein. J. Neuroimmunol. 39,231–242.
Spranger, M., Lindholm, D., Bandtlow, C., Heumann, R., Gnahn, H.,Naher-Noe, M., Thoenen, H., 1990. Regulation of nerve growthfactor (NGF) synthesis in the rat central nervous system:comparison between the effects of interleukin-1 and various growthfactors in astrocyte cultures and in vivo. Eur. J. Neurosci. 2,69–76.
Strijbos, P.J., Rothwell, N.J., 1995. Interleukin-1 beta attenuates excitatoryamino acid-induced neurodegeneration in vitro: involvement of nervegrowth factor. J. Neurosci. 15, 3468–3474.
Takami, Y., Motoki, T., Yamamoto, I., Gohda, E., 2005. Synergisticinduction of hepatocyte growth factor in human skin fibroblasts by theinflammatory cytokines interleukin-1 and interferon-gamma. Biochem.Biophys. Res. Commun. 327, 212–217.
Tanaka, M., Ito, S., Kiuchi, K., 2000. Novel alternative promoters of mouseglial cell line-derived neurotrophic factor gene. Biochim. Biophys. Acta1494, 63–74.
Tchelingerian, J.L., Quinonero, J., Booss, J., Jacque, C., 1993. Localizationof TNF alpha and IL-1 alpha immunoreactivities in striatal neurons aftersurgical injury to the hippocampus. Neuron 10, 213–224.
Teismann, P., Tieu, K., Cohen, O., Choi, D.K., Wu, dC., Marks, D., Vila, M.,Jackson-Lewis, V., Przedborski, S., 2003. Pathogenic role of glial cellsin Parkinson’s disease. Mov. Disord. 18, 121–129.
Verity, A.N., Wyatt, T.L., Hajos, B., Eglen, R.M., Baecker, P.A.,Johnson, R.M., 1998. Regulation of glial cell line-derived neuro-trophic factor release from rat C6 glioblastoma cells. J. Neurochem.70, 531–539.
Verity, A.N., Wyatt, T.L., Lee, W., Hajos, B., Baecker, P.A., Eglen, R.M.,Johnson, R.M., 1999. Differential regulation of glial cell line-derivedneurotrophic factor (GDNF) expression in human neuroblastoma andglioblastoma cell lines. J. Neurosci. Res. 55, 187–197.
Vitkovic, L., Bockaert, J., Jacque, C., 2000. Inflammatory cytokines:neuromodulators in normal brain? J. Neurochem. 74, 457–471.
Wang, J., Bankiewicz, K.S., Plunkett, R.J., Oldfield, E.H., 1994.Intrastriatal implantation of interleukin-1. Reduction of parkinsonismin rats by enhancing neuronal sprouting from residual dopaminergicneurons in the ventral tegmental area of the midbrain. J. Neurosurg. 80,484–490.

104 A. Saavedra et al. / Neurobiology of Disease 25 (2007) 92–104
Woodbury, D., Schaar, D.G., Ramakrishnan, L., Black, I.B., 1998. Novelstructure of the human GDNF gene. Brain Res. 803, 95–104.
Xiao, H., Hirata, Y., Isobe, K., Kiuchi, K., 2002. Glial cell line-derivedneurotrophic factor up-regulates the expression of tyrosine hydrox-ylase gene in human neuroblastoma cell lines. J. Neurochem. 82,801–808.
Xu, K., Dluzen, D.E., 2000. The effect of GDNF on nigrostriataldopaminergic function in response to a two-pulse K(+) stimulation.Exp. Neurol. 166, 450–457.
Yamasaki, Y., Matsuura, N., Shozuhara, H., Onodera, H., Itoyama, Y.,Kogure, K., 1995. Interleukin-1 as a pathogenetic mediator of ischemicbrain damage in rats. Stroke 26, 676–680.