perinatal Δ 9-tetrahydrocannabinol exposure in rats modifies the responsiveness of midbrain...
TRANSCRIPT

Drug and Alcohol Dependence 42 (1996) 155.-166
Perinatal A9-tetrahydrocannabinol exposure in rats modifies the responsiveness of midbrain dopaminergk neurons in adulthoud to a
variety of challenges with dopaminergic drugs’
Lucia Garcia, Rosario de Miguel, Jo& A. Ramos, J. Javier Fernkndez-Ruiz*
Institutn Comphplzrtense de Drogodependencias, Department of Biochemistry, Facult~~ of Medicine. Complutense ~~niw~sit~~, ?KO4(l-Mndrid, Spain
Received IO April 1996; revised 10 July 1996; accepted IO July 1996
Abstract
The present study has been designed to explore further the existence of a persistent, but ‘silent’ alteration in the adult functionality of midbrain dopaminergic neurons following perinatal cannabinoid exposure. To this end, we evaluated the responsiveness of these neurons, measured at the neurochemical or behavioral levels, to pharmacological challenges with a variety of dopaminergic drugs administered to adult male and female rats that had been exposed to A9-tetrahydrocarmabinol (THC) or vehicle during the perinatal period. Results were as follows: In the first experiment, we tested the magnitude of motor inhibition caused by administration of dopaminergic receptor antagonists. The most interesting observation was that the administration of SCH 23390, a D, antagonist, produced a more marked motor inhibition, reflected by a greater decrease in the ambulation measured in an open-field test, in adult animals of both sexes when they had been exposed perinatally to THC. This did not occur with the motor inhibition caused by sulpiride, a D, antagonist. In the second experiment, we evaluated the sensitivity of midbrain dopaminergic neurons to amphetamine (AMPH), which causes, through different mechanisms, a decrease in dopamine (DA) metabolism. The most interesting observation was that adult females, when exposed perinatally to THC, exhibited a trend to lesser response to AMPH, in terms of decreasing DA metabolism, than oil-exposed females. This was observed in dopaminergic terminals reaching the limbic forebrain area, but not in those terminals reaching the striatum, and was a specific effect for THC-exposed adult females because it was not observed in THC-exposed adult males. In the third experiment, we evaluated the in vivo synthesis of DA in midbrain dopaminergic neurons by analyzing the magnitude of L-3,4-dihydroxyphenylalanine (L-DOPA) accumulation caused by the blockade of L-DOPA decarboxylase with NSD 1015. The most worthy finding was that. as occurred in the above experiment, adult females, when exposed perinatally to THC, tended to exhibit a higher ability to synthesize DA in vivo in the limbic forebrain but not in the striatum, as reflected by the increased L-DOPA accumulation observed after NSD 1015 administration. As in the above experiment, this was not seen in males. In summary, our results are consistent with the possible existence of subtle and sexually dimorphic changes in the sensitivity of midbrain dopdminergic neurons in adulthood caused by the exposure to THC during perinatal development. These silent changes could be revealed after the administration of drugs which specifically act on key processes of dopaminergic neurotransmission, such as the synthesis, reuptake and catabolism of DA and its binding to receptors.
Keywords: Cannabinoids; AY-tetrahydrocannabinol; Midbrain dopaminergic neurons: Dopamine: Tyrosine hydroxylase; D, and D, receptors: Motor behavior: SCH 23390; Sulpiride; Amphetamine: NSD 1015
1. Introduction * Corresponding author. Tel.: + 34 1 3941450; fax: + 34 I
3941691; e-mail: [email protected] ! Parts of this work have been previously presented in abstract
form and published in the Proceedings of 1995 Meeting of Interna- tional Cannabis Research Society (Scottsdale, AZ, USA).
We have demonstrated extensively that p&natal ex- posure to cannabinoids markedly afrects the matura- tion of several dopaminergic systems (Rodriguez de
0376-8716’96/$35.00 c 1996 Elsevier Science Ireland Ltd. All rights reserved PII S037h-G7 16(96)0 1276.8

156 L. Garcia et al. 1 Drug and Alcol~ol Dependence 42 (1996) 155-166
Fonseca et al., 1991; Fernandez-Ruiz et al., 1992, 1994; Bonnin et al., 1994). These effects appear early, even before the complete differentiation and maturation of dopaminergic projections into their target areas (Ro- driguez de Fonseca et al., 1992), in particular during the last third of gestation and first days of postnatal life, when cannabinoids seem to alter the expression of key genes for development of dopaminergic neurotrans- mission (Bonnin et al., 1995; Fernandez-Ruiz et al., 1996). Among the dopaminergic pathways affected most by the perinatal exposure to cannabinoids are the midbrain dopaminergic neurons (Rodriguez de Fonseca et al., 1991), in particular those neurons whose fibers reach the striatum and constitute the nigrostriatal do- paminergic system (Bonnin et al., 1994; Navarro et al., 1994), although we have also found changes in the other major component of midbrain dopaminergic fibers that reach the limbic forebrain nuclei and consti- tute the mesolimbic system (Rodriguez de Fonseca et al., 1991). The most characteristic effect, by its magni- tude and persistence, was a decrease in the activity of tyrosine hydroxylase (TH) in the striatum of males perinatally exposed to cannabinoids (Rodriguez de Fonseca et al., 1991; Fernandez-Ruiz et al., 1994). This effect was observed permanently along the peripubertal period (15-40 days after birth), in parallel with a slight decrease in motor activity (Navarro et al., 1994). Wal- ters and Carr (1986) also found decreases in the activity of this enzyme at immature ages after perinatal expo- sure to marihuana extracts. Accompanying this modifi- cation in TH activity, we have also found changes in gene expression of this enzyme in the midbrain (Bonnin et al., 1994), the area where cell bodies of nigrostriatal and mesolimbic TH-containing terminals are clustered, and increases in the density of both D, and D, recep- tors in the striatum (Rodriguez de Fonseca et al., 1991; Fernandez-Ruiz et al., 1994). As occurred with TH, this last effect was observed exclusively in males and tended to disappear after the withdrawal of the drug at wean- ing and, particularly, in adulthood (Navarro et al., 1994), presumably due to the appearance of compensa- tory mechanisms during the course of further develop- ment.
Despite the disappearance in adulthood of can- nabinoid-induced presynaptic and postsynaptic changes observed over the course of postnatal development, we suspect the persistence at this age of some irreversible alterations in the functionality of these neurons, result- ing in behavioral disturbances. These alterations might not be evident in basal conditions, but may be mani- fested in particular circumstances and in a sexually dimorphic manner. We had previously formulated this hypothesis (Navarro et al., 1994) based on a set of facts involving different key processes for dopaminergic neurotransmission. Thus: (i) the levels of TH-mRNA in the midbrain of adult males perinatally exposed to
cannabinoids were lesser than in controls (Bonnin et al., 1994), although TH activity was unaffected in the re- gions receiving terminals from midbrain dopaminergic cell bodies (Navarro et al., 1994; Fernandez-Ruiz et al., 1994); (ii) these animals exhibited significant changes in the motor response executed when exposed to novel stimuli in an open-field test (Navarro et al., 1994), in particular, in the frequency of stereotypy whose rela- tion to specific dopaminergic receptors seems to be clearly defined (Arnt, 1987); these changes did not exist in females (Navarro et al., 1994); (iii) the activity of midbrain dopaminergic neurons in adult male rats, that had been perinatally exposed to cannabinoids, was affected by a challenge with A9-tetrahydrocannabinol (THC), the prototypical psychoactive cannabinoid, to a lesser extent than naive rats (Fernandez-Ruiz et al., 1994) whereas adult females, when exposed perinatally to cannabinoids, responded to an adult challenge with THC as did naive rats (Fernandez-Ruiz et al., 1994). Recently, we have provided the first evidence to sup- port the hypothesis of the existence of an irreversible, although ‘silent’, alteration in the adult functionality of midbrain dopaminergic neurons originated as a conse- quence of perinatal THC exposure. We have observed that perinatal exposure to THC alters the ability of these neurons in adulthood to respond, particularly in males, to drugs which deplete dopamine (DA) stores and induce TH gene expression, such as reserpine and a-methyl-p-tyrosine (Garcia et al., 1996).
The present work has been designed to complete these first results. Thus, we have examined the respon- siveness, tested at neurochemical or behavioral levels, of these neurons when challenged pharmacologically with a variety of drugs, injected at well-defined active doses, that affect key processes for dopaminergic neu- rotransmission, such as DA metabolism and reuptake or D, and D, DA receptors. To this end, we have performed three different experiments. In the first ex- periment, we evaluated the degree of motor inhibition (measured in the open-field test) caused by the adminis- tration of specific antagonists for either D, or D, DA receptors. As mentioned previously, both receptors were affected by perinatal THC exposure during the course of postnatal development (Rodriguez de Fon- seca et al., 1991). DA antagonists have been largely reported to produce a set of motor deficits (catalepsy, inactivity), through the blockade of these receptors in the areas receiving terminals from midbrain dopaminer- gic neurons, when injected to adult rats (Campbell and Baldessarini, 1981; McDougall et al., 1990; Meyer, 1993; Kostrzewa and Kastin, 1993; Garrett and Holtz- man, 1994) and also when administered during the perinatal period (Scalzo et al., 1989).
In the second experiment, we evaluated the respon- siveness of midbrain dopaminergic neurons to the ad- ministration of amphetamine (AMPH). This drug has

been reported to produce dopaminergic hyperactivity when injected to adult rats: (i) by reducing the DA reuptake process; (ii) by inhibiting the activity of monoamine oxidase; or (iii) via its stimulant effects on the release of an extravesicular pool of newly synthe- sized DA. In the present study, we have paid special attention to a common consequence, from a neuro- chemical point of view, of this triad of effects of AMPH: the decrease in DA intraneuronal metabolism reflected in the loss of L-3,4-dihydroxyphenylacetic acid (DOPAC) production (Elverfors and Nissbrandt, 1992). This process has been reported to be affected signih- cantly by THC during development (Rodriguez de Fon- seca et al., 1991). In the third experiment, we tested the ability of midbrain dopaminergic neurons to synthesize DA in vivo, in view of the important effects of perinatal THC exposure on TH gene expression and activity. Thus, we analyzed L-3,4-dihydroxyphenylalanine (L- DOPA) accumulation following NSD 1015induced blockade of L-DOPA decarboxylase, a classical test for analyzing in vivo synthesis of DA (Steger et al., 1989; Rowlett et al., 1993).
2. Methods
2.1. Animals
Female virgin Wistar rats were housed in a room with a controlled photoperiod (08.00-20.00 h light) and temperature (23 + 1’C). They had free access to stan- dard food (Panlab, Barcelona, Spain) and water. At adult age ( > 8 weeks of life; 150-200 g), daily vaginal smears were taken between lO:OO-12:00 h, and only those animals exhibiting three or more consistent 4-day cycles were used in this study. Females in the proestrous phase were allowed to stay with a male for mating, and a new vaginal smear was taken on the next day. Those animals showing the presence of sperm cells were accepted as probably pregnant and used for the cannabinoid exposure studies. The day on which sperm plugs were found was designated as the first day of gestation.
2.2. Perinu fat THC e.pmwe
THC, kindly supplied by the National Institute on Drug Abuse (USA), was prepared in a sesame oil solution for administration. Pregnant females received a daily oral dose of THC (5 mg/kg) from the day 5 of gestation, as in our previous studies (Bonnin et al., 1995; Fernandez-Ruiz et al., 1996; Garcia et al., 1996). This dose is an extrapolation from current estimates of moderate exposure to this compound in humans, cor- recting for differences in route of administration and body surface area (Nahas, 1984). Control rats were fed
with vehicle alone. This treatment was maintained until day 24 after birth, the day on which rats were weaned. Then, males and females were housed separately (4 per cage) and left to reach adult age ( > 10 weeks). During the whole treatment period, we recorded a set of gesta- tional and lactational parameters, such as maternal and neonatal weight gain, maternal food and water intake, gestational length, placental and fetal weights, litter size and maternal plasma THC concentrations, in order to show that potential adult changes were not due to a possible cannabinoid-induced nutritional deficit during the perinatal life. These results have been previously published (Bonnin et al.. 1995; Fernandez-Ruiz et al., 1996) and are not included in the present report.
Males and females that had been exposed perinatally to THC or vehicle were used when adult ( > 10 weeks: females were in diestrus) for three different pharmaco- logical challenges. In the three cases, animals were assigned randomly to adult treatment conditions, being always the four different perinatal groups (females+ oil: females + THC: males -+ oil; males + THC) ob- tained from at least three different oil- or THC-exposed litters. Experiments with these animals were always performed between lO:OO~- 13:00 h. In experiment I, one third of the animals of each of these four groups was i.p. injected with a specific D, antagonist, SCH 23390 (purchased from Research Riochemicals Inc.. USA), used at a dose of 0.1 “g/kg, another third was injected with a specitic Dz antagonist, sulpiride (purchased from Sigma, USA), used at a dose of 30 mg!kg, and the last third was treated with vehicle. 30 min later, animals were tested behaviorally (open-field test). All doses were chosen because they were effective in markedly dimin- ishing motor behavior in adult rats (they produeed more than 70% of reduction in ambulatory activity in both sexes) in dose-response studies conducted previ- ously in our laboratory (data not shown) and also in previous reports (McDougall et al., 1990: Garrett and Holtzman, 1994). In experiment II, half of each perina- tal group was S.C. injected with AMPH (purchased from Research Biochemicals, USA), used at a dose of 10 mg/kg and the other half of each group was treated with vehicle. 30 min later, animals were sacrificed. The dose of AMPH was chosen because it was effective in markedly diminishing intraneuronal DA metabolism in adult rats in a previous report (Elverfors and Niss- brandt. 1992). and in dose-response studies conducted previously in our laboratory (data not shown). In ex- periment III. half of each perinatal group was i.p. injected with NSD 1015 (purchased from Research Biochemicals, USA), used at a dose of 100 mg/kg, and the other half of each group was injected with vehicle. 30 min after treatment, animals were sacrificed. The

158 L. Garcia et al. 1 Drug and Alcohol Dependence 42 (1996) 155-166
dose of NSD 1015 was chosen because it was effec- tive in inhibiting L-DOPA decarboxylase in adult rats, as previously reported (Steger et al., 1989; Rowlett et al., 1993) and validated in our laboratory (data not shown). Drugs were dissolved in distilled water (ex- periment I), although sulpiride was dissolved initially in a drop of glacial acetic acid, or saline (experiments II and III), and injected in a volume of 2-5 ml/kg (the differences in the volumes of injection depended on the experiment). In order to control a potential effect of acetic acid used for solubilizing sulpiride in experiment I, some control animals were injected with acidified distilled water in the same proportion as for preparing sulpiride solution; as these animals did not exhibit differences as compared to animals injected with distilled water alone, both were combined for the statistical analysis and presentation.
2.4. Sampling
In experiments II and III, animals were sacrificed and their brains removed and dissected according to Glowinski and Iversen (1966). The striatum and lim- bit forebrain (this contains olfatory tubercles, rostra1 limbic nuclei, the nucleus accumbens, septal nuclei and parts of the anterior amygdaloid nuclei) were dis- sected, weighed and immediately frozen and stored at - 70°C until assayed for DA indices.
2.5. Behavioral studies
In experiment I, animals were subjected to an open-field test 30 minutes after treatment. This con- sisted of a circular arena composed of hardboard (di- ameter: 90 cm) with a surround (height: 30 cm), both made of transparent polyvinylchloride. The floor was divided into five inner and eight outer parts using circles and radial segments. On the day of the test, the experimental animal was placed in the centre of the arena and its spontaneous activity was recorded on a TV-video system. In the pilot dose-response ex- periments, the duration of the test was 10 min, di- vided into two periods of 5 min. It was observed that the motor inhibition caused by increasing doses of the DA antagonists injected in both sexes was, in terms of percentages, similarly distributed in both periods, suggesting that the influence of the stress derived from the novel environment was not a determinant. Thus, we decided to perform the behavioral testing of the main experiment only during the first 5 min. The advantage of this option is the increase in the number of animals per each daily testing session, reducing the influence of interassay variation without markedly in- creasing the total time of each session and, hence, avoiding the effects of natural fluctuations during the
photoperiod. The apparatus was washed out with an odoriferous solution after each rat had been tested. The following parameters were analyzed: (i) sponta- neous locomotor activity (ambulation)-number of sector crossings (a single line-crossing was defined as the rat placing the four paws into an adjacent quad- rant); (ii) frequency (score) of spontaneous stereotypic behaviors (grooming, rearing and shaking); and (iii) time spent in inactivity. The scoring of the different behaviors was carried out by investigators who had no knowledge of the treatment of each rat.
2.6. Neurochemical analyses
DA, DOPAC and L-DOPA contents were analyzed using HPLC with electrochemical detection. Striata and limbic forebrains were homogenized in ice-cold 0.2 N perchloric acid containing 0.5 mM sodium bisulfite and 0.45 mM EDTA (30 vol for the striatum and 10 vol for the limbic forebrain). Dihydroxybenzy- lamine was added as an internal standard. The ho- mogenates were then centrifuged and the supernatants injected into the HPLC system. The HPLC system consisted of a Spectra-Physics 8810 isocratic pump. The column was a RP-18 (Spherisorb ODS-2; 125 mm, 4.6 mm, 5 pm particle size). The mobile phase consisted of 100 mM citric acid, 100 mM sodium acetate, 1.2 mM octane sulphonate, 1 mM EDTA and 5% methanol (pH 3.9) and the flow rate was 0.8 ml/min. The effluent was monitored with a coulo- chemical detector (Coulochem II, ESA) using a proce- dure of oxidation/reduction (conditioning cell: + 360 mV; analytical cell # 1: + 50 mV; analytical cell # 2: - 340 mV). The signal was recorded from the analytical cell # 2, with a sensitivity of 50 nA (10 pg per sample), on a Spectra-Physics 4290 integrator and the results were given as area under the peaks. Values were expressed as ng/mg of tissue.
2.7. Statistics
Data from the three experiments were mainly as- sessed by 3-way analysis of variance (variable 1: sex; variable 2: perinatal treatment; variable 3: adult chal- lenge), using the computer package BMDP, followed by a post-hoc test (Student-Newman-Keuls test). Ad- ditionally, data were transformed to percentage of change over the mean value of the corresponding ve- hicle-injected group and, then, subjected to 2-way analysis of variance (variable 1: sex; variable 2: peri- natal treatment) also followed by the Student-New- man-Keuls test. This was done to evaluate better the differences or to enhance some trends, as a function of sex and perinatal treatment, in the magnitude of changes caused by the different adult challenges.
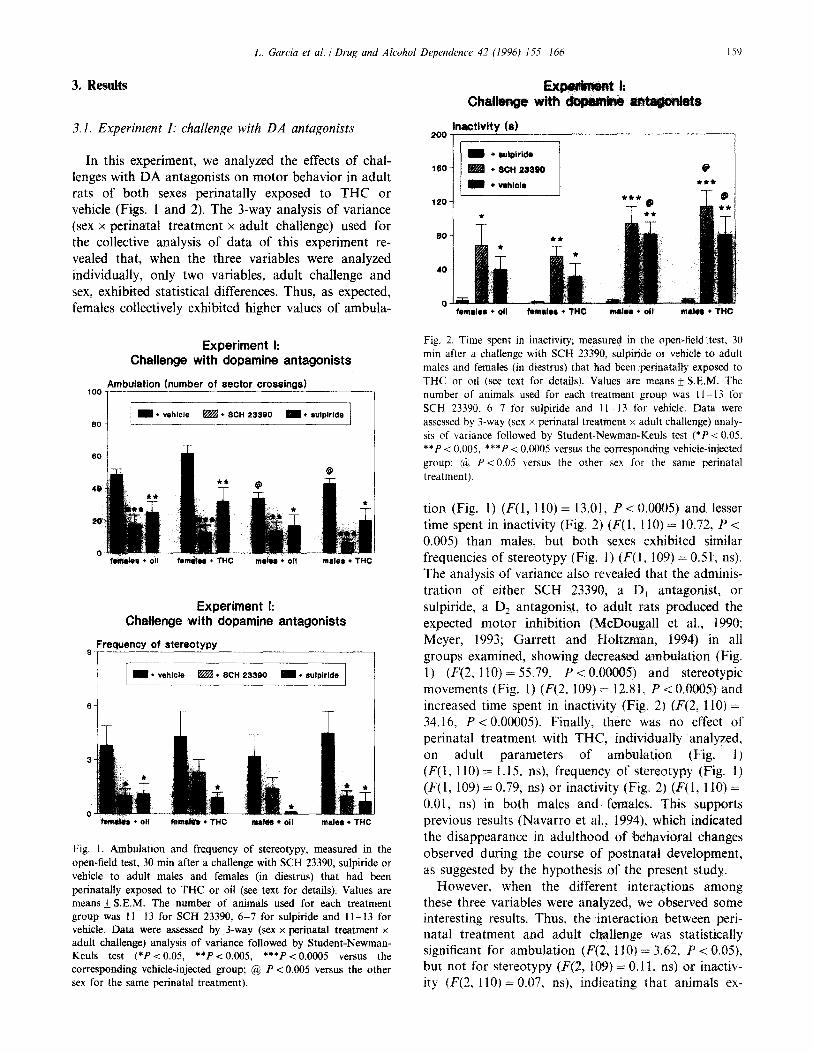
L. Garcia et al. 1 Drug and Alcohol Dependence 42 (1996) 155 166
3. Results
3.1. Experiment I: challenge with DA antagonists
In this experiment, we analyzed the effects of chal- lenges with DA antagonists on motor behavior in adult rats of both sexes perinatally exposed to THC or vehicle (Figs. 1 and 2). The 3-way analysis of variance (sex x perinatal treatment x adult challenge) used for the collective analysis of data of this experiment re- vealed that, when the three variables were analyzed individually, only two variables, adult challenge and sex, exhibited statistical differences. Thus, as expected, females collectively exhibited higher values of ambula-
Experiment I: Challenge with dopamine antagonists
60
0 fmwates + Oil femfw l THC m&4 l oil mater l THC
Experiment I: Challenge with dopamine antagonists
Freauencv of stereotvDY - I
- l vehicle &i%! l SCH 23390 = + rulplride
6
0 fsfn&s l Oil lam* * THC mat64 l oil molar + THC
Fig. 1. Ambulation and frequency of stereotypy, measured in the open-field test. 30 min after a challenge with SCH 23390, sulpiride or vehicle to adult males and females (in diestrus) that had been perinatally exposed to THC or oil (see text for details). Values are means k S.E.M. The number of animals used for each treatment group was 11 - 13 for SCH 23390, 6-7 for sulpiride and 1 l- 13 for vehicle. Data were assessed by 3-way (sex x perinatal treatment x adult challenge) analysis of variance followed by Student-Newman- Keuls test (*Pi 0.05, **P ~0.005, ***PC 0.0005 versus the corresponding vehicle-injected group; @ P < 0.005 versus the other sex for the same perinatal treatment).
Ex Challgnse wBh
inactivltv Is) 200
l- 160
120
60
40
0
m *w$irid4 + SCH 29890
m + which
li n
Fig. 2. Time spent in inactivity. measured in the open-field test. 30 min after a challenge with SCH 23390, sulpiride or vehicle to adult males and females (in diestrus) that had been perinatally exposed to THC or oil (see text for details). Values are means f S.E.M. The number of animals used for each treatment group was II-13 for SCH 23390. 6 7 for sulpiride and 1 l- 13 for vehicle. Data were assessed by 3-way (sex x perinatal treatment x adult challenge) analy- sis of variance followed by Student-Newman-Keuls test (*P < 0.05. **P < 0.005, ***P i 0.0005 versus the corresponding vehicle-injected group: ($ P < 0.05 versus the other sex for the same perinatal treatment).
tion (Fig. 1) (F(l, 110) = 13.01, P<O.O005) and lesser time spent in inactivity (Fig. 2) (F(1, I 10) = 10.72, P < 0.005) than males, but both sexes exhibited similar frequencies of stereotypy (Fig. 1) (F(1~ 109) = 0.5 1, ns). The analysis of variance also revealed that the adminis- tration of either SCH 23390, a D, antagonist, or sulpiride, a D, antagonist, to adult rats produced the expected motor inhibition (McDougall et al., 1990: Meyer, 1993; Garrett and Holtzman, 1994) in all groups examined, showing decreased ambulation (Fig. 1) (F(2, 110) = 55.79, P < 0.00005) and stereotypic movements (Fig. 1) (F(2, 109) = 12.81, P < 0.0005) and increased time spent in inactivity (Fig. 2) (F(2, 110) = 34.16. P < 0.00005). Finally, there was no effect of perinatal treatment with THC, individually analyzed,
adult parameters of ambulation (Fig. 1) Fl(l, 110) = 1.15, ns), frequency of stereotypy (Fig. I) (F(1, 109) = 0.79, ns) or inactivity (Fig. 2) (F( 1, 110) = 0.01, ns) in both males and females. This supports previous results (Navarro et al., 1994), which indicated the disappearance in adulthood of behavioral changes observed during the course of postnatal development, as suggested by the hypothesis of the present study.
However, when the different interactions among these three variables were analyzed, we observed some interesting results. Thus, the interaction between peri- natal treatment and adult challenge was statistically significant for ambulation (F(2, 110) =r 3.62, P < 0.05), but not for stereotypy (F(2, 109) = 0.11, ns) or inactiv- ity (F(2, 110) = 0.07, ns), indicating that animals ex-
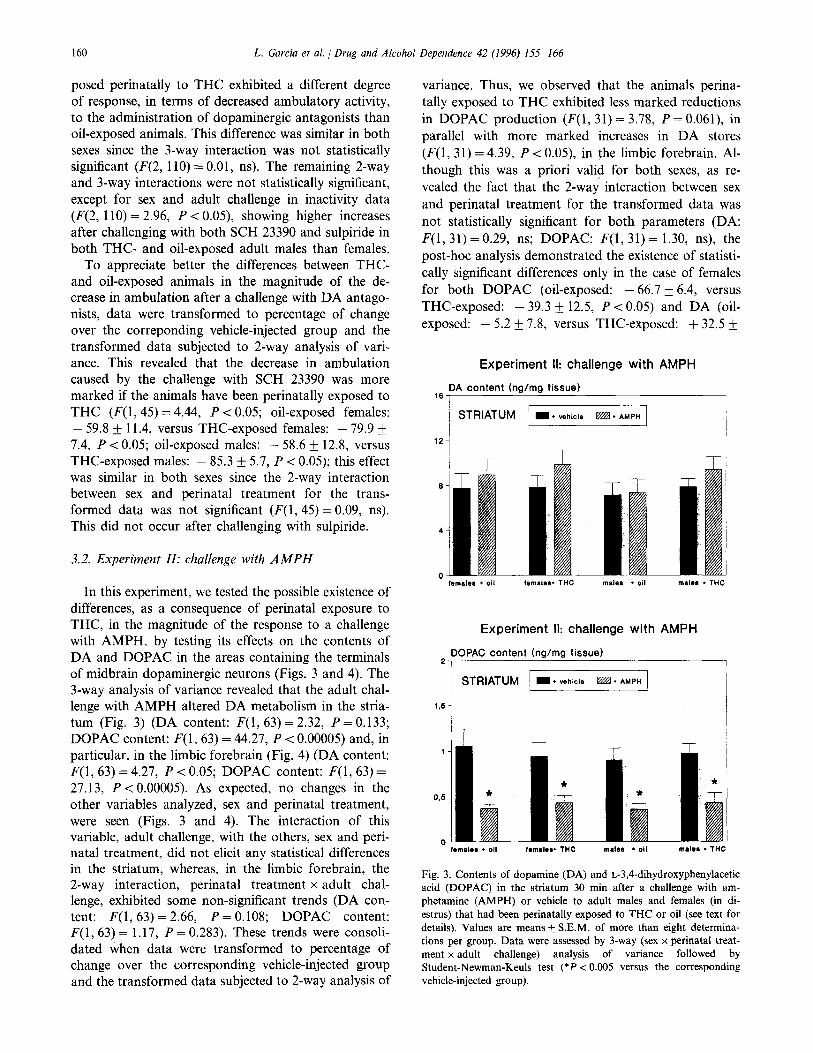
160 L. Garcia et al. / Drug and Alcohol Dependence 42 (1996) 155-166
posed perinatally to THC exhibited a different degree of response, in terms of decreased ambulatory activity, to the administration of dopaminergic antagonists than oil-exposed animals. This difference was similar in both sexes since the 3-way interaction was not statistically significant (F(2, 110) = 0.01, ns). The remaining 2-way and 3-way interactions were not statistically significant, except for sex and adult challenge in inactivity data (F(2, 110) = 2.96, P < 0.05), showing higher increases after challenging with both SCH 23390 and sulpiride in both THC- and oil-exposed adult males than females.
To appreciate better the differences between THC- and oil-exposed animals in the magnitude of the de- crease in ambulation after a challenge with DA antago- nists, data were transformed to percentage of change over the correponding vehicle-injected group and the transformed data subjected to 2-way analysis of vari- ance. This revealed that the decrease in ambulation caused by the challenge with SCH 23390 was more marked if the animals have been perinatally exposed to THC (F( 1,45) = 4.44, P < 0.05; oil-exposed females: - 59.8 + 11.4, versus THC-exposed females: - 79.9 + 7.4, P < 0.05; oil-exposed males: - 58.6 f 12.8, versus THC-exposed males: - 85.3 + 5.7, P < 0.05); this effect was similar in both sexes since the 2-way interaction between sex and perinatal treatment for the trans- formed data was not significant (F(1,45) = 0.09, ns). This did not occur after challenging with sulpiride.
3.2. Experiment II: challenge with AMPH
In this experiment, we tested the possible existence of differences, as a consequence of perinatal exposure to THC, in the magnitude of the response to a challenge with AMPH, by testing its effects on the contents of DA and DOPAC in the areas containing the terminals of midbrain dopaminergic neurons (Figs. 3 and 4). The 3-way analysis of variance revealed that the adult chal- lenge with AMPH altered DA metabolism in the stria- turn (Fig. 3) (DA content: F(1,63) = 2.32, P = 0.133; DOPAC content: F(1, 63) = 44.27, P < 0.00005) and, in particular, in the limbic forebrain (Fig. 4) (DA content: F(1,63) = 4.27, P < 0.05; DOPAC content: F(1,63) = 27.13, P < 0.00005). As expected, no changes in the other variables analyzed, sex and perinatal treatment, were seen (Figs. 3 and 4). The interaction of this variable, adult challenge, with the others, sex and peri- natal treatment, did not elicit any statistical differences in the striatum, whereas, in the limbic forebrain, the 2-way interaction, perinatal treatment x adult chal- lenge, exhibited some non-significant trends (DA con- tent: F(1, 63) = 2.66, P = 0.108; DOPAC content: F(1, 63) = 1.17, P = 0.283). These trends were consoli- dated when data were transformed to percentage of change over the corresponding vehicle-injected group and the transformed data subjected to 2-way analysis of
variance. Thus, we observed that the animals perina- tally exposed to THC exhibited less marked reductions in DOPAC production (P(1, 31) = 3.78, P = 0.061), in parallel with more marked increases in DA stores (F(1, 31) = 4.39, P < 0.05), in the limbic forebrain. Al- though this was a priori valid for both sexes, as re- vealed the fact that the 2-way interaction between sex and perinatal treatment for the transformed data was not statistically significant for both parameters (DA: F(1,31) = 0.29, ns; DOPAC: F(1, 31) = 1.30, ns), the post-hoc analysis demonstrated the existence of statisti- cally significant differences only in the case of females for both DOPAC (oil-exposed: - 66.7 _+ 6.4, versus THC-exposed: - 39.3 IfI 12.5, P < 0.05) and DA (oil-
t exposed: - 5.2 f 7.8, versus THC-exposed: + 32.2
Experiment II: challenge with AMPH
,6 DA content hg/mg tissue)
I / STRIATUM m * vehicle m l AMPH
12 -
Experiment II: challenge with AMPH
2 DOPAC content (ng/mg tissue) I
~ STRIATUM / I+ vehicle RW l AMPH
105 1
male0 + THC
Fig. 3. Contents of dopamine (DA) and L-3,4-dihydroxyphenylacetic acid (DOPAC) in the striatum 30 min after a challenge with am- phetamine (AMPH) or vehicle to adult males and females (in di- es&us) that had been perinatally exposed to THC or oil (see text for details). Values are means + S.E.M. of more than eight determina- tions per group. Data were assessed by 3-way (sex x perinatal treat- ment x adult challenge) analysis of variance followed by Student-Newman-Keuls test (*P < 0.005 versus the corresponding vehicle-injected group).
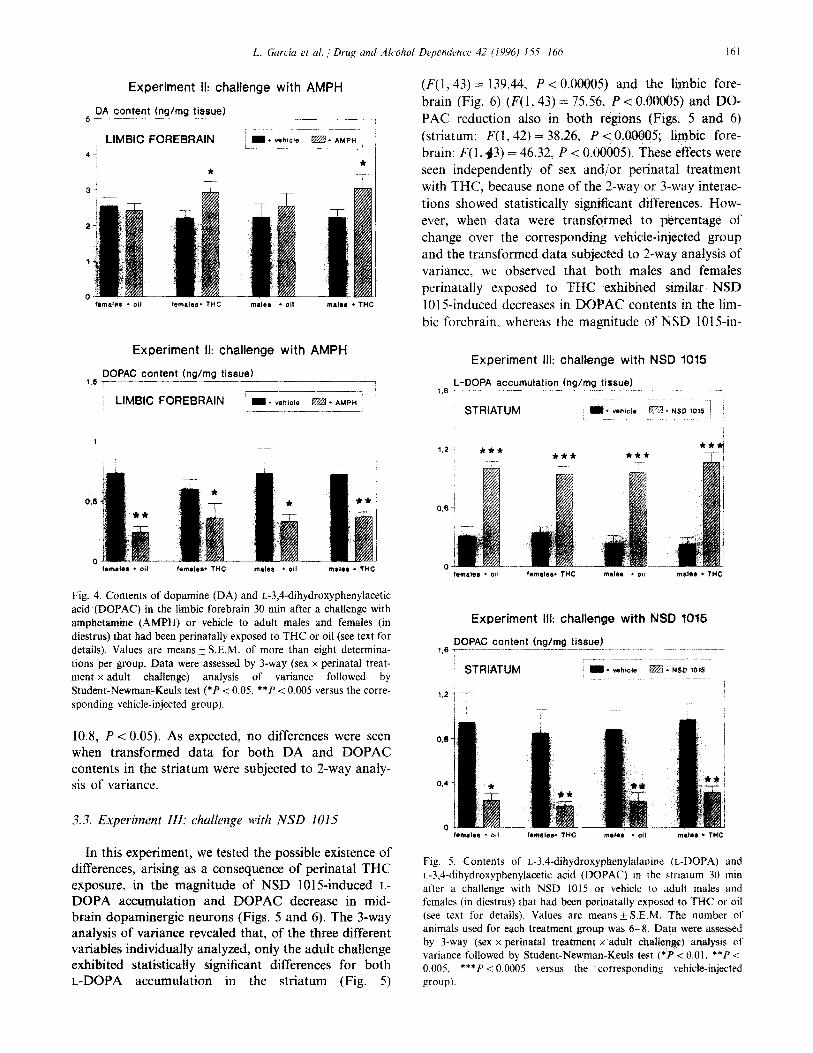
L. Garcia et al. / Drug and Alcohol Dependence 42 (1996) 1.5.5 166 IhI
Experiment II: challenge with AMPH
5 PA content hg/mg tissue) --__--_ ---7
ri l vahicte I
LlMBiC FOREBRAIN Em.AMPH /
47 L----L /
* *
Experiment II: challenge with AMPH
DOPAC content (ng/mg tissue) 1.5 r __--____ ---
‘~ LIMBIC FOREBRAIN -___
I I* vehicle m + AMPH _____
Fig. 4. Contents of dopamine (DA) and L-3,4-dihydroxyphenylacetic acid (DOPAC) in the limbic forebrain 30 min after a challenge with amphetamine (AMPH) or vehicle to adult males and females (in diestrus) that had been perinatally exposed to THC or oil (see text for details). Values are means + S.E.M. of more than eight determina- tions per group. Data were assessed by 3-way (sex x perinatal treat- ment x adult chahenge) analysis of variance followed by Student-Newman-Keuls test (*P < 0.05, **P < 0.005 versus the corre- sponding vehicle-injected group).
10.8, P < 0.05). As expected, no differences were seen when transformed data for both DA and DOPAC contents in the striatum were subjected to 2-way analy- sis of variance.
3.3. Experiment III: challenge with NSD 1015
In this experiment, we tested the possible existence of differences, arising as a consequence of perinatal THC exposure, in the magnitude of NSD 1015-induced L- DOPA accumulation and DOPAC decrease in mid- brain dopaminergic neurons (Figs. 5 and 6). The 3-way analysis of variance revealed that, of the three different variables individuahy analyzed, only the adult challenge exhibited statistically significant differences for both L-DOPA accumulation in the striatum (Fig. 5)
(F(l,43) = 139.44, P < 0.00005) and the limbic fore- brain (Fig. 6) (F(1,43) = 75.56, P < 0.00005) and DO- PAC reduction also in both regions (Figs. 5 and 6) (striatum: F(1,42) = 38.24, P < 0.00005; limbic fore- brain: F(1,43) = 46.32, P < 0.00005). These effects were seen independently of sex and/or perinatal treatment with THC, because none of the 2-way or 3-way interac- tions showed statistically sigmficant differences. How- ever, when data were transformed to percentage of change over the corresponding vehicle-injected group and the transformed data subjected to 2-way analysis of variance, we observed that both males and females perinatally exposed to THC exhibited similar NSD 1015-induced decreases in DOPAC contents in the lim- bit forebrain, whereas the magnitude of NSD 1015-in-
Experiment III: challenge with NSD 1015
L-DOPA accumulation (ng/mg tissue) ,*s --- -.--.---- ~--~-__; ._... ~~.- ..~
Experiment Ill: challenge with NSD 1015
DOPAC content (ng/mg tissue) ,,s -.----~~-...- -~-.---~----. __-.---__ ------. - _--_
STRIATUM I * v&id* m * NSD 30% ~
1.27 I
Fig. 5. Contents of r.-3,4-dihydroxyphenylalanine (L-DOPA) and L-3,4-dihydroxyphenylacetic acid (DOPAC) in the striarum 30 min after a challenge with NSD 1015 or vehicle to adult males and females (in diestrus) that had been perinatally exposed to THC or oil (see text for details). Values are means + S.E.M. The number of animals used for each treatment group was 6--8~ Data were assessed by 3-way (sex x perinatal treatment x adult challenge) analysis of variance followed by Student-Newman-Ret& test (*P < 0.01, **P < 0.005. ***P < 0.0005 versus the corresponding vehicle-injected group).
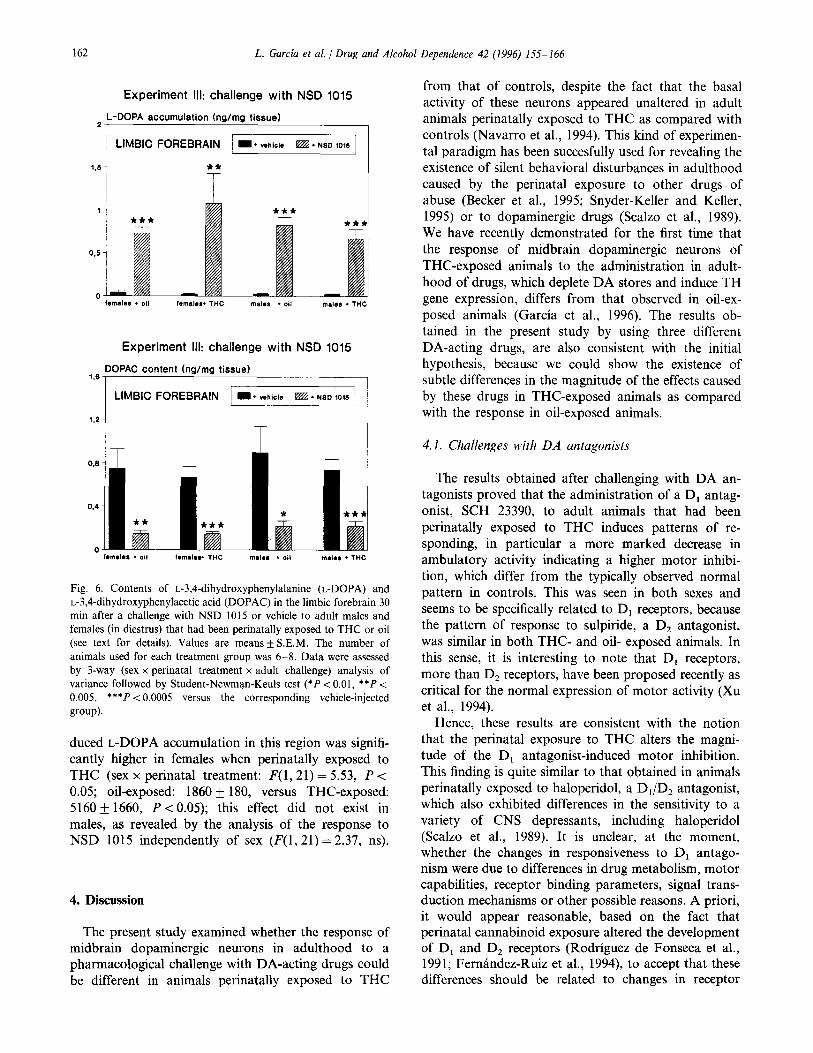
162 L. Garcia et al. /Drug and Alcohol Dependence 42 (1996) 1.55-166
Experiment III: challenge with NSD 1015
2 L-DOPA accumulation (ng/mg tissue)
Experiment Ill: challenge with NSD 1015
1.6- DOPAC content (ng/mg tissue)
LIMBIC FOREBRAIN I I. vehic~s W. NSD mm
Fig. 6. Contents of L-3,4-dihydroxyphenylalanine (L-DOPA) and L-3,4-dihydroxyphenylacetic acid (DOPAC) in the limbic forebrain 30 min after a challenge with NSD 1015 or vehicle to adult males and females (in diestrus) that had been perinatally exposed to THC or oil (see text for details). Values are means + S.E.M. The number of animals used for each treatment group was 6-8. Data were assessed by 3-way (sex x perinatal treatment x adult challenge) analysis of variance followed by Student-Newman-Keuls test (*P < 0.01, **P < 0.005, ***P < 0.0005 versus the corresponding vehicle-injected grow).
duced L-DOPA accumulation in this region was signifi- cantly higher in females when perinatally exposed to THC (sex x perinatal treatment: F(1,21) = 5.53, P < 0.05; oil-exposed: 1860 f 180, versus THC-exposed: 5160 + 1660, P < 0.05); this effect did not exist in males, as revealed by the analysis of the response to NSD 1015 independently of sex (P(1,21) = 2.37, ns).
4. Discussion
The present study examined whether the response of midbrain dopaminergic neurons in adulthood to a pharmacological challenge with DA-acting drugs could be different in animals perinatally exposed to THC
from that of controls, despite the fact that the basal activity of these neurons appeared unaltered in adult animals perinatally exposed to THC as compared with controls (Navarro et al., 1994). This kind of experimen- tal paradigm has been succesfully used for revealing the existence of silent behavioral disturbances in adulthood caused by the perinatal exposure to other drugs of abuse (Becker et al., 1995; Snyder-Keller and Keller, 1995) or to dopaminergic drugs (Scalzo et al., 1989). We have recently demonstrated for the first time that the response of midbrain dopaminergic neurons of THC-exposed animals to the administration in adult- hood of drugs, which deplete DA stores and induce TH gene expression, differs from that observed in oil-ex- posed animals (Garcia et al., 1996). The results ob- tained in the present study by using three different DA-acting drugs, are also consistent with the initial hypothesis, because we could show the existence of subtle differences in the magnitude of the effects caused by these drugs in THC-exposed animals as compared with the response in oil-exposed animals.
4.1. Challenges with DA antagonists
The results obtained after challenging with DA an- tagonists proved that the administration of a D, antag- onist, SCH 23390, to adult animals that had been perinatally exposed to THC induces patterns of re- sponding, in particular a more marked decrease in ambulatory activity indicating a higher motor inhibi- tion, which differ from the typically observed normal pattern in controls. This was seen in both sexes and seems to be specifically related to D, receptors, because the pattern of response to sulpiride, a D, antagonist, was similar in both THC- and oil- exposed animals. In this sense, it is interesting to note that D, receptors, more than D, receptors, have been proposed recently as critical for the normal expression of motor activity (Xu et al., 1994).
Hence, these results are consistent with the notion that the perinatal exposure to THC alters the magni- tude of the D, antagonist-induced motor inhibition. This finding is quite similar to that obtained in animals perinatally exposed to haloperidol, a D,/D, antagonist, which also exhibited differences in the sensitivity to a variety of CNS depressants, including haloperidol (Scalzo et al., 1989). It is unclear, at the moment, whether the changes in responsiveness to D, antago- nism were due to differences in drug metabolism, motor capabilities, receptor binding parameters, signal trans- duction mechanisms or other possible reasons. A priori, it would appear reasonable, based on the fact that perinatal cannabinoid exposure altered the development of D, and D, receptors (Rodriguez de Fonseca et al., 199 1; Fernandez-Ruiz et al., 1994), to accept that these differences should be related to changes in receptor

L. Garcia et al. I Drug and Alcohol Dependence 42 (1996) 155-I 66 163
binding parameters in areas recipient of midbrain do- paminergic outputs. However, we have recently re- ported that D, receptor binding parameters were unchanged in the striatum and the limbic forebrain of adult males perinatally exposed to THC (Navarro et al., 1994; Fernandez-Ruiz et al., 1994), despite the increase shown during the immature period (Rodriguez de Fonseca et al., 1991; Fernandez-Ruiz et al., 1992). In adult females exposed perinatally to THC, we observed a statistically significant, but very small, increase in the density of D, receptors in the striatum but not in the limbic forebrain (Navarro et al., 1994; Fernandez-Ruiz et al., 1994), with no changes in affinity. Taking into account these previous data, it appears unlikely that the SCH 23390~induced higher decrease in the ambulatory activity of adult males, when exposed perinatally to THC, might be related to changes in affinity or number of D, receptors, since there were no relevant changes in the binding characteristics of D, receptors in these animals (Navarro et al., 1994). This can be also applied to adult THC-exposed females, because, although we could observe changes in the receptor binding parame- ters. such as an increase in the density of D, in the striatum (Navarro et al., 1994) it is unlikely that this might explain a higher degree of motor inhibition. The higher density of these receptors in the striatum of THC-exposed females should lead to a higher propor- tion of unblocked D, receptors after SCH 23390 admin- istration than in control females, thereby allowing a higher receptor activation by DA and more ‘normal’ motor behavior, rather than a more marked motor inhibition.
Hence, the higher decrease in the ambulatory activity caused by SCH 23390 in THC-exposed adult males and females seems to be unrelated to changes in binding parameters for this receptor. This would be similar to that seen with the behavioral sensitization to drugs that act by directly or indirectly stimulating dopaminergic sensitivity, which require the stimulation of DA recep- tors. although unaccompanied by changes in DA recep- tor affinity or density (Rowlett et al., 1993). Another possibilities could be the existence of changes in the availability of D, receptors or in the molecular mecha- nisms coupled to these receptors. Regarding the first of these two possibilities, there could be differences in the proportion of receptors that might be incorporated into the postsynaptic membrane, and, thus, easily available for neurotransmitter binding, and the proportion of receptors that might be incorporated into the intracellu- lar membranes, presumably in recycling processes, and unable to bind the neurotransmitter in vivo. However, the in vitro binding analysis is not able to discriminate between both receptor locations (both are marked with the radioactive probe) because of the use of crude membrane fractions. The possible existence of differ- ences at the postreceptor level, i.e., coupling to their
intracellular signal mechanisms, leading to differences in the magnitude of their biological action, could be also considered. This might be supported by the recent studies from Herkenham et al. (1991), who have re- ported that DA receptors, mainly D, receptors, are present in the same striatal neurons where cannabinoid receptors seem to be located, and by the fact that both cannabinoid and DA receptors act at the intracellular level by modifying the CAMP contents (for review, see Fernandez-Ruiz et al., 1992; Schwartz et al., 1992).
4.2. Challenges with AMPH
The administration of AMPH to adult rodents pro- duced a well-characterized reduction in DA metabolism in midbrain dopaminergic terminals, which originates in a variety of potential sites of action for this drug (see Section 1). It is possible to evaluate the degree of AMPH action by examining the attenuation induced by this drug in DOPAC production, since this metabolite is formed intraneuronally from the reuptaken or newly synthesized DA (Elverfors and Nissbrandt, 1992). In the present work, we have observed that AMPH ad- ministration produced a marked attenuation in the DOPAC production, ranging around 60%. similar to that observed by Elverfors and Nissbrandt (1992). It can be asserted that the magnitude of the effects of AMPH, in terms of attenuation of DOPAC production, was similar in oil- and THC-exposed animals of both sexes, in view of the absence of statistical significance in the interaction between perinatal treatment and adult challenge, as well as in the 3-way interaction (sex x perinatal treatment x adult challenge). However, when data were transformed to percentages of change, we could appreciate that the effect of AMPH was slightly less marked when this drug was administered to ani- mals that had been exposed perinatally to THC. The fact that this effect was seen only after transformation of the data. might indicate that the effect is subtle rather than pronounced. To determine its physiological relevance would require further experiments. Anyway, this subtle effect was seen only in the midbrain do- paminergic terminals reaching the Iimbic forebrain area, but not in those terminals reaching the caudate- putamen area, indicating that it is specific for mesolim- bit neurons. Moreover, it appears to occur only in adult females, but not in adult males. Our group has reported the existence of marked sexual differences in the response to cannabinoids when administered peri- natally in rats (for review, see Fernandez-Ruiz et al.. 1992, 1994, 1996), and others have obtained similar findings in animals exposed perinataliy to other drugs of abuse such as cocaine, even when challenged phar- macologically in adulthood as in the present study (Snyder-Keller and Keller, 1995; for review, see Fernan- dez-Ruiz et al.. 1992, 1994). However an interesting

164 L. Garcia et al. 1 Drug and Alcohol Dependence 42 (1996) 155-166
aspect derived from our results after the AMPH chal- lenge is that females were more responsive in adult- hood, whereas previous reports demonstrated that males were more affected during development and that the effects in developing females were slight and tran- sient (for review, see Fernandez-Ruiz et al., 1992, 1994, 1996). In this sense, it is important to note that the maturation of midbrain dopaminergic neurons, in terms of increasing DOPAC production, during the first days after birth was accelerated profoundly in THC-exposed females but not in THC-exposed males (Bonnin et al., unpublished results). We interpreted this effect as aris- ing from an early maturation of DA reuptake process induced by THC in females, which allowed a higher DOPAC production. It is possible that this accelerated maturation during the postnatal period might also re- sult in a higher number of DA reuptake sites in adult- hood, leading to an expected reduced effect of DA reuptake blockers as AMPH.
The above finding, a potential increase in the number of DA reuptake sites in THC-exposed females, might explain at the molecular level the differences in the responsiveness to AMPH, but other possibilities should be also considered. For example AMPH at the dose used in the present study presumably might inhibit monoamine oxidase activity leading to reduction of DOPAC production. To our knowledge, there is no evidence for possible changes in the activity of this enzyme as a function of perinatal THC exposure that might explain the subtle differences in the magnitude of AMPH action. Another possibillity could be changes at the level of DA synthesis; we have seen that animals perinatally exposed to THC exhibited AMPH-induced increases in the DA content in the limbic forebrain, but not in the striatum, an effect which is difficult a priori to explain. These changes did not exist in oil-exposed animals. In this respect, Elversfors and Nissbrandt (1992) have suggested the existence of an indirect AMPH stimulatory effect on DA synthesis. This could be produced as a consequence of the decrease in the concentrations of extravesicular intraneuronal DA caused by AMPH, which, in turn, may cause an in- crease of synthesis due to reduced end-product inhibi- tion. It would be expected that THC-exposed animals-in particular females, in which the DA metabolism was reduced to a lesser extent than in oil-exposed animals-might secondarily develop an in- creased DA synthesis leading to higher DA contents. This finding seems to be strengthened by the data obtained in experiment III of the present study, which suggested that mesolimbic dopaminergic terminals of THC-exposed females had a subtlely higher rate of DA synthesis than oil-exposed animals, as will be discussed below.
4.3. Challenges with NSD 1015
The administration of NSD 1015, an inhibitor of L-DOPA decarboxylase, to adult rats produced the well-known stimulatory effect on L-DOPA concentra- tions, that has been used as an index of in vivo DA synthesis (Rowlett et al., 1993). As ocurred in the above experiment, it can be asserted that the magnitude of L-DOPA accumulation was similar in oil- and THC-ex- posed animals of both sexes, in view of the absence of statistical significance in the 2-way and 3-way interac- tions. However, the transformation of data to percent- age of change allowed to appreciate that the magnitude of L-DOPA accumulation might be subtly higher in the midbrain dopaminergic terminals reaching the limbic forebrain area of THC-exposed females as compared with oil-exposed females. This did not exist in THC-ex- posed males and in the other constituent of midbrain dopaminergic terminals, the nigrostriatal system. Fur- ther experiments must determine the physiological rele- vance of this observation.
It is important to note that we have recently reported that the activity of TH in the limbic forebrain of THC-exposed females was similar to that measured in oil-exposed animals (Fernandez-Ruiz et al., 1994). However, it is important to consider that the analysis of TH activity was performed in vitro by measuring the amounts of L-DOPA formed when brain homogenates were incubated with saturating concentrations of L-ty- rosine. It would be possible that, although the in vitro analysis leads to no changes, the in vivo analysis might show the existence of an altered synthesis capability, presumably due to circumstances that do not exist in the in vitro assay: substrate availability, intracellular location and others. Certainly, this might be occurring in THC-exposed females, which subtly exhibited a higher capability to accumulate L-DOPA when the activity of L-DOPA-decarboxylase had been blocked with NSD 1015.
5. Conclusions
In summary, our results are consistent with the possi- ble existence of subtle changes in the sensitivity of midbrain dopaminergic neurons in adulthood caused by the exposure to THC during perinatal development. These silent changes can be revealed after the adminis- tration of drugs which specifically act on key processes of dopaminergic neurotransmission, such as the metabolism and reuptake of DA and its binding to receptors. A particular feature was the existence of a possible sexual dimorphism in these silent changes in the sensitivity of midbrain dopaminergic neurons. The neurochemical basis for these differences remains to be determined, but it is important to note that these might

L. Garcia et al.’ / Drug and Alcohol Dependence 42 (1996) 155166 165
presumably underly a variety of potential long-term disturbances, observed in adult animals perinatally ex- posed to cannabinoids, in the adult functionality of dopamine-related processes, such as stress, motor be- havior and reward (Dalterio, 1980; Mokler et al., 1987; Navarro et al., 1994; for review see Dalterio, 1986; Fernrindez-Ruiz et al., 1992). In this sense, we have recently observed that both adult male and fe- male rats became more intensely opiate-dependent in studies of self-administration, when exposed perina- tally to THC (Ambrosio, FernBndez-Ruiz and Ramos, unpublished results), an effect that might be tentatively related to the above mentioned changes in mesolimbic dopaminergic neurons, in view of the well-known involvement of these neurons in reward- ing effects of habit-forming drugs (Herz and Shippen- berg, 1989).
Acknowledgements
This work has been supported by grants from CI- CYT (SAF-94-0540). The authors are indebted to the National Institute on Drug Abuse (USA) for kindly supplying THC.
References
Amt. J. (1987) Behavioral studies of dopamine receptors: evidence for regional selectivity and receptor multiplicity. In: Receptor Biochemistry and Methodology, Vol. 8, Dopamine Receptors (Creese. I. and Fraser, C.M., eds.), pp. 199%220. Alan R. Liss, New York.
Becker, H.C., Weathersby, R.T. and Hale, R.L. (1995) Prenatal ethanol exposure alters sensitivity to the effects of apomorphine given alone and in combination with ethanol on locomotor ac- tivity in adult male mouse offspring. Neurotox. Teratol. 17, 57 64.
Bonnin. A., de Miguel, R., Rodriguez-Manzaneque, J.C., Fern&- dez-Ruiz, J.J.. Santos, A. and Ramos, J.A. (1994) Changes in tyrosine hydroxylase gene expression in mesencephalic cate- cholaminergic neurons of immature and adult male rats perina- tally exposed to cannabinoids. Dev. Brain Res. 81, 147-150.
Bonnin, A., de Miguel, R., HernBndez, M.L., Ramos, J.A. and FernBndez-Ruiz, J.J. (1995) The prenatal exposure to A9-te- trahydrocannabinol affects the gene expression and the activity of tyrosine hydroxylase during early brain development. Life Sci. 56, 2177.2184.
Campbell, 4. and Baldessarini, R.J. (1981) Effects of maturation and aging on behavioral responde to haloperidol in the rat, Psychopharmacology 73, 219-222.
Dalterio. S.L. (1980) Perinatal or adult exposure to cannabinoids alters male reproductive functions in mice. Pharmacol. Biochem. Behav. 12. 14% 153.
Daltcrio. S.L. (1986) Cannabinoid exposure: effects on develop- ment. Neurobehav. Toxicol. Teratol. 8, 345- 352.
Elverfors. A. and Nissbrandt, H. (1992) Effects of D-amphetamine on dopaminergic neurotransmission; a comparison between the substantia nigra and the striatum. Neuropharmacology 31, 661 - 670
FernBndez-Ruiz. J.J., Rodriguez de Fonseca, I-., Navarro. M. and Ramos. J.A. (1992) Maternal cannabinoid exposure and brain development: changes in the ontogeny of dopaminergic neurons. In: Neurobiology and Neurophysiology of Cannabinoids, Bio- chemistry and Physiology of Substance Abuse. Vol. IV (Bartke, A. and Murphy, L.L.. eds.). pp. 1 I9 164. CRC Press. Boca Raton, FL.
FernPndez-Ruiz, J.J.. Bonnin, A., Cebeira. M. and Ramos. J.A. ( 1994) Ontogenic and adult changes in the activity of’ hypotha- lamic and extrahypothalamic dopeminer& neurons after peri- natal cannabinoid exposure. In: Strategies for Studying Brain Disorders, Vol. I. (Palomo, T. and Archer. 7-., eds.), pp. 357 390. Farrand Press, England.
FernBndez-Ruiz. J.J.. Bonnin. A.. de Miguel, R., Castro, J.G. and Ramos. J.A. (1996) Perinatal exposure to marihuana or to its main psychoactive constituent. A’-tetrahydrocannabinol, affects the development of brain dopaminergic neurons. In: Procced- ings of the International Conference on Drugs of Abuse Re- cent Advances in Clinical and Experimental Research. (Tavareh. M..4. and Ali. SF.. eds.), Arquivos de Medicina tin press).
Garcia. L.. Castro. J.G. Bonnin, A.. de Miguel. R.. Ramos. J.A. and Fernindez-Ruiz. J.J. (1996) Exposure to psychoactive com- pounds of marihuana during perinatal development alters the responsiveness of midbrain dopaminergic ncurona in adulthood to drugs m#hich deplete dopamine stores ;md induce tyrosine hydrox) lase gent expression. In: f’roceedings of the Internw- tional Conference on Drugs of Abuse Recent Advances in Clinical and Experimental Research. (Tav:,rec M.A. dnd .Ali. S.F.. cds.). Arquivos de Medicina (in press)
Garrett. B.E. and Holtrman. S.G. (1994) r>, and I& dopamine receptor antagonist block caffeine-mduced stimulation of loco- motor acticily in rats. Pharmacol. Biochem Beha\. 47. 89 94.
Glowinskl, J. and Iverscn. L.L. (1966) C;rtzcholamine regional metabolism in rat brain. J. Neurochem. 1.3. 055 669.
Herkenham, M.. Lynn. ‘4.8.. de Costa. B.R. :tnd Richfield, L.K. (199 I) I\leuronal localization of cannabinold receptors in the basal ganglia of the rat. Brain Res. 547, 76”’ 274.
Herz. A. itnd Shippenberg. S.T. (lY8Yi Neurochemical aspects of addiction: opioids and other drugs of abus< In: Molecular and Cellular Aspects of the Drug Addictions. ~Goldslein. ,4.. cd.). pp. I I I 155. Springer-Verlag, New York.
Kostrzewa. R.M. and Kastin. A.J. I 1993) ‘P!r-MIF-I attenuates development of tolerance to spipernne-!ilduccd catalepsy in rats. Brain Res. Bull. 31. 707 712.
McDougall, S.A.. Arnold. T.F. and Nonnem,in. A.J. rl9911) On- togcn) of locomotor activity and grooming II) the young rat: role of dopamine DI and D2 receptors. Eui J Phannacol. 186, 123 1.30
Meyer, M.E. ( 1993) El’fccts of mtraaccumbeni dopammc agonist SKI: 38393 and antagonist SCH 133130 on locomotor activity in rats. Pharmacol. Biocbem. Behav. 4.5. 843 sJ5
Mokler. D.A.. Robinson. S.E., Johnson J.11.. Hong. J.S. and Rosecrans. .J.A. (19X:) Neonatal administration of <i’-tctrahy- drocannabinol alters the neurochemical re\ponsc to stress in the adult Fischer-344 rat. Neurotoxicol. Teratol 9. 331 226.
Nahas. G.G. (1984) Toxicology and Pharnracolog>. In: Marihuana in Science and Medicine (Nahas. G G.. i,d 1. pp 102 247. Raven Press. New York.
Navarro. M.. Rodriguez de Fonseca. t‘.. Hernandel. M.I.., Ramos. J.A. and FernBndez-Ruiz. J.J. (1994) Motor behavior and ni- grostriatal dopaminergic activity in adult rats perinatally ex- posed to cannabinoid>. Pharmacol. Biochem. Behav. 47. 47 58.
Rodriguez de Fonscca. F.. Cabeira. M.. Fzrnindez-Ruiz. J.J.. ?iacarro. M. and Ramos. J.A. ( I991 ) Effects of pre- and peri- natal exposure to hashish extracts on the ontogeny of bl-ain dopaminergic neurons Neuroscience 3.7. 7 I : ‘:I 3,

166 L. Garcia et al. /Drug and Alcohol Dependence 42 (1996) 155-166
Rodriguez de Fonseca, F., Hernandez, M.L., de Miguel, R., Fernan- dez-Ruiz, J.J. and Ramos, J.A. (1992) Early changes in the development of dopaminergic neurotransmission after maternal exposure to cannabinoids. Pharmacol. Biochem. Behav. 41, 469- 414.
Rowlett, J.K., Mattingly, B.A. and Bardo, M.T. (1993) Neurochemi- cal correlates of behavioral sensitization following repeated apo- morphine treatment: assessment of the role of D, dopamine receptor stimulation. Synapse 14, 160- 168.
Scalzo, F.M., Ali, SF. and Holson, R.R. (1989) Behavioral effects of prenatal haloperidol exposure. Pharmacol. Biochem. Behav. 34, 127-131.
Schwartz, J.C., Giros, B., Martres, M.P. and Sokoloff, P. (1992) The dopamine receptor family: molecular biology and pharmacology. Sem. Neurosci. 4. 99-108.
Snyder-Keller, A.M. and Keller Jr., R.W. (1995) Prenatal cocaine alters later sensitivity to cocaine-induced seizures. Neuro-Sci. Lett. 191, 149-152.
Steger, R.W., Esquifino, AL, Fernandez-Ruiz, J.J. and Bartke, A. (1989). Prolonged neuroendocrine effects of a brief exposure of adult male rats to diethylstilbestrol. Neuroendocrinology 49, 191- 196.
Walters, D.E. and Carr, L.A. (1986) Changes in brain catecholamine mechanisms following perinatal exposure to marihuana. Pharma- col. Biochem. Behav. 25, 763-768.
Xu, M., Moratalla, R., Gold, L.H., Hiroi, N., Koob, G.F., Gray- biel, A.M. and Tonegawa, S. (1994) Dopamine D, receptor mutant mice are deficient in striatal expression of dynorphin and in dopamine-mediated behavioral responses. Cell 79, 729- 142.