chemoarchitecture of the anuran auditory midbrain
TRANSCRIPT

Brain Research Reviews 33 (2000) 179–198www.elsevier.com/ locate /bres
Full-length review
Chemoarchitecture of the anuran auditory midbraina , a ,1 b ,2*Heike Endepols , Wolfgang Walkowiak , Harald Luksch
a ¨ ¨ ¨Universitat zu Koln, Zoologisches Institut, Weyertal 119, 50923 Koln, Germanyb ¨RWTH Aachen, Institut f ur Biologie II, Kopernikusstr. 16, 52074 Aachen, Germany
Accepted 30 May 2000
Abstract
The anuran torus semicircularis consists of several subnuclei that are part of the ascending auditory pathway as well as audiomotorinterface structures. Additionally, recent anatomical studies suggest that the midbrain tegmentum is an integral part of the audiomotornetwork. To describe the chemoarchitecture of these nuclei, taking into account the toral subdivisions, we investigated the distribution ofserotonin, leucine–enkephalin, substance P, tyrosine–hydroxylase, dopamine D2-receptor, parvalbumin, aspartate, GABA, and estrogen-binding protein-immunoreactivity in the midbrain of Bombina orientalis, Discoglossus pictus and Xenopus laevis. In the torussemicircularis, the highest density of immunoreactive fibers and terminals for all transmitters was found in the laminar nucleus.Parvalbumin-like immunoreactivity was highest in the principal nucleus, and D2-receptor-like immunoreactivity was uniformly distributedthroughout the torus. In the tegmentum, axons and/or dendrites were stained with all antibodies except estrogen-binding protein.Additionally, heavily stained enkephalin and substance P-immunopositive fiber plexus were found in the lateral and dorsal tegmentum.The immunostainings revealed no qualitative differences between the three species. Immunopositive cell bodies were labeled in severalbrain areas, the connectivity of which with torus and tegmentum is discussed on the background of functional questions. The putativeneuromodulatory innervation of both the laminar nucleus of the torus semicircularis and the tegmentum may be the anatomical basis forthe influence of the animal’s endogenous state on the behavioral reaction to sensory stimuli. These data corroborate earlier anatomical andphysiological findings that the neurons of these nuclei are key elements in the audio-motor interface. 2000 Elsevier Science B.V. Allrights reserved.
Theme: Neurotransmitters, modulators, transporters, and receptors
Topic: Regional localization of receptors and transmitters
Keywords: Neuromodulator; Sensory-motor interface; Auditory pathway; Behavioral adaptation
Abbreviations: 5HT, serotonin; A, anterior thalamic nucleus; Acc, nucleus accumbens; ASP, aspartate; C, central thalamic nucleus; Cb, cerebellarnucleus; Cer, cerebellum; D2, dopamine D2-receptor; DA, dopamine; DB, nucleus of the diagonal band of Broca; df, dorsal field of spinal grey; DP, dorsalpallium; Ea, anterior entopeduncular nucleus; EBP, estrogen-binding protein; ENK, enkephalin; fr, fasciculus retroflexus; ft, fasciculi tegmentales; GABA,gamma-aminobutyric acid; Gc, griseum centrale rhombencephali; Hd, dorsal habenular nucleus; Hv, ventral habenular nucleus; Hyp, hypothalamus; Ipr,interpeduncular nucleus, rostral portion; LA, lateral amygdala; LC, locus coeruleus; LPd, lateral pallium, dorsal portion; LPv, lateral pallium, ventralportion; MA, medial amygdala; MP, medial pallium; MS, medial septum; NA, noradrenalin; Ndl, dorsolateral nucleus; NI, isthmal nucleus; Nis, secondaryisthmal nucleus; Npm, nucleus profundus mesencephali; Os, superior olive; P, posterior thalamic nucleus; PA, preoptic area; Prtr, pretrigeminal nucleus;PT, posterior tuberculum; Ptg, pretectal grey; PV, parvalbumin; Ra, raphe nucleus; Rcf, cuneiform reticular nucleus; SC, suprachiasmatic nucleus; Std,dorsal striatum; Stv, ventral striatum; SP, substance P; Tec, optic tectum; Teg, tegmentum; TH, tyrosine–hydroxylase; Tl, laminar nucleus of the torussemicircularis; Tm, magnocellular nucleus of the torus semicircularis; Tp, principal nucleus of the torus semicircularis; tv, tectal ventricle; vf, ventral fieldof spinal grey; VH, ventral hypothalamic nucleus; VM, ventromedial thalamic nucleus
*Corresponding author. Tel.: 149-221-4703-101; fax: 149-221-4704-889.E-mail addresses: [email protected] (H. Endepols), [email protected] (W. Walkowiak), [email protected] (H. Luksch).1Tel.: 149-221-470-3119; fax: 149-221-470-4889.2Tel.: 149-241-804-832, fax: 149-241-888-8133.
0165-0173/00/$ – see front matter 2000 Elsevier Science B.V. All rights reserved.PI I : S0165-0173( 00 )00029-1

180 H. Endepols et al. / Brain Research Reviews 33 (2000) 179 –198
Contents
1. Introduction ............................................................................................................................................................................................ 1802. Materials and methods ............................................................................................................................................................................. 1803. Results.................................................................................................................................................................................................... 181
3.1. Specificity ...................................................................................................................................................................................... 1813.2. Distribution of immunoreactive terminals and immunopositive somata within the torus semicircularis................................................... 1813.3. Synopsis of nuclei in the brain with somata immunoreactive for the neuromodulators studied............................................................... 187
4. Discussion .............................................................................................................................................................................................. 1904.1. Immunohistochemical findings ......................................................................................................................................................... 1904.2. Functional aspects of neurotransmitter /neuromodulator input to the torus ........................................................................................... 1934.3. Distribution of other neurotransmitters in the auditory midbrain ......................................................................................................... 1944.4. Functional anatomy of the torus semicircularis .................................................................................................................................. 194
Acknowledgements ...................................................................................................................................................................................... 195References................................................................................................................................................................................................... 195
1. Introduction midbrain. Furthermore, data from an earlier study on thedistribution of steroid-concentrating neurons in Xenopus
In the auditory pathway of anurans, the midbrain torus laevis [44,45] should be corroborated in other species. Wesemicircularis and tegmentum are of considerable interest therefore investigated the distribution of serotonin,as targets of neurotransmitters and neuromodulators. The leucine–enkephalin, substance P, tyrosine–hydroxylase,principal nucleus of the torus semicircularis is the main dopamine D2-receptor, parvalbumin, aspartate, GABA,projection area for ascending auditory afferents, whereas and estrogen-binding protein (estrogen-bp) immuno-the magnocellular nucleus and especially the laminar reactivity.nucleus constitute ascending and descending projections Many investigations have been carried out on neobat-with sensory, premotor and motor areas in all parts of the rachian species (e.g., Rana, Hyla). In order to broaden ourbrain [24,58,95]. Neurons of the midbrain tegmentum also knowledge about the Archaeobatrachia, we chose therespond to auditory stimulation and, similar to laminar species Bombina orientalis, Discoglossus pictus (Discog-nucleus neurons, project to sensory, premotor, and motor lossidae), and Xenopus laevis (Pipidae).regions [58]. All three subnuclei of the torus semicircularisas well as the tegmentum receive descending direct orindirect input from the striatum and the dorsal thalamus. 2. Materials and methodsEspecially the response of laminar nucleus neurons toauditory nerve stimulation could be altered by activation of A total of 61 adult animals was used in this studyforebrain efferents [21]. Since behavioral studies have comprising 30 Discoglossus pictus, 19 Xenopus laevis, andshown that torus semicircularis and tegmentum are in- 12 Bombina orientalis. The animals were taken from thevolved in audiomotor integration [86,87,95], shaping of laboratory’s breeding stock. For the detection of estrogen-neuronal responses in these nuclei may contribute essen- bp reproductively active specimen (females with well-tially to selection and modification of motor programs developed ovaries, males with thumb pads) were chosen.according to the internal state of the animal. In order to All antibodies were applied on animals of both sexes. Theunderstand the anatomical basis of these interfacing mecha- antibodies were obtained from different sources: polyclon-nisms, it is essential to work out the functional subar- al serotonin-antibody (rabbit) from Dianova (Hamburg,chitecture of the auditory midbrain in greater detail. Germany); polyclonal antibodies against substance P and
The central nervous system of anuran amphibians has leucine–enkephalin (both rabbit) from IBL (Hamburg,been the object of numerous immunohistochemical studies. Germany); polyclonal antibodies against aspartate (rabbit)However, most of these have concentrated on the visual and dopamine D2-receptor (rabbit), and the monoclonalmidbrain or have given overall distribution patterns with- antibody against tyrosine–hydroxylase (mouse) fromout taking the toral subdivisions into account. Therefore, Chemicon International (Temecula, CA). Two antibodiesthe aim of this study was to reveal the termination sites of against GABA, polyclonal and monoclonal, were used: thevarious neurotransmitter inputs to the torus semicircularis polyclonal antibody against GABA had been produced byand the midbrain tegmentum in detail. The nuclei giving Dr. T.G. Kingan (Columbia University) in rabbits and wasrise to these inputs should be identified by comparing the made available by Dr. J.J. Milde (University of Cologne,pattern of immunolabeled nuclei in the entire brain with Germany). The monoclonal GABA-antibody (Mab 3A12)the results of tracing studies on toral connectivity [24,57]. had been produced by Dr. C. Matute (University ofTo gain additional insight into the functional organization, Zaragoza, Spain) in mice and was a gift from Dr. K. Braunwe wanted to describe the localization of neurons immuno- (Leibnitz-Institute for Neurobiology, Magdeburg, Ger-positive for the different transmitters within the auditory many). It is now commercially available from Swant

H. Endepols et al. / Brain Research Reviews 33 (2000) 179 –198 181
(Bellinzona, Switzerland). The monoclonal antibody 3. Resultsagainst the estrogen-bp (rat) and its detection system waskindly provided by Abbott diagnostica (Wiesbaden, Ger- 3.1. Specificitymany). All other antisera were obtained from Dakodiagnostica (Hamburg, Germany). The procedure described above labeled neuronal cell
The animals were lethally anesthetized in a 1:50 (w/v) bodies, dendrites, and terminal structures with a strongsolution of tricaine methanesulfonate (MS 222) [76] and contrast to the background. In the case of the dopaminesubsequently perfused transcardially with cold 0.1 M D2-receptor, only somata and basal dendrites were stained;sodium-phosphate buffer (PB) (pH 7.4) followed by a no labeling of terminal structures could be achieved. Thesolution of 4% paraformaldehyde in PB. For the detection antibody against estrogen-bp labeled only the nuclei ofof GABA, animals were perfused with 0.05 M Tris buffer neurons. The control sections never showed any immuno-(TB) and fixated with 1% paraformaldehyde/2.5% glutar- reactivity. If not explicitly mentioned, the distribution ofdialdehyde in TB; the buffer system for the detection of the immunoreactive structures did not differ betweenestrogen-bp was a potassium-phosphate saline (0,01 M; pH species or sex.7.4). Brains were removed, post-fixed for 4–7 h and rinsedin buffer overnight. After an immersion in 15% sucrose in 3.2. Distribution of immunoreactive terminals andbuffer for 1 h and subsequent immersion in embedding immunopositive somata within the torus semicircularismedium (Reichert-Jung) for 30 min, brains were frozenrapidly. Frontal or sagittal sections (15–20 mm thick) were Serotonin (Fig. 1). In the torus semicircularis, axonalcut on a cryostat (Reichert-Jung, Frigocut 2800) and and terminal structures showing serotonin-like immuno-mounted on chrom–alum-gelatinated slides; every fifth reactivity were found almost exclusively in the laminarsection was collected on a separate slide for specificity test. nucleus (Fig. 10A); however, the total amount ofThe sections were air-dried at room-temperature for one h serotoninergic structures was not high. Within the marginand rinsed three times (10 min each) in PB, followed by an of the laminar nucleus, terminal labeling was strongest; inimmersion in a solution of 0.5% Triton-X in PB for 30 min the remaining toral nuclei, only few fibers could be(this step was omitted for the estrogen-bp and the GABA- detected. As an exception, the medial plane of the torusprocedure). After several rinses in PB, sections were was lightly labeled; nevertheless, this region (subependym-incubated with normal serum for 30 min on a shaker and al nucleus and central nucleus) is not ascribed to thesubsequently incubated with the primary antibody at 48C auditory torus and its function is still unclear. In the wholeovernight or for 2 h at room temperature; the control tegmentum immunoreactive fibers were found at lowsections were incubated with buffer (dilution buffer for all density, and thick fiber bundles and terminal plexus wereantibodies was PB containing 1% BSA). After three rinses labeled in the lateral parts of the tegmentum. Immuno-in PB (10 min each), the binding sites of the primary reactive somata were located in the ventral tegmentum andantibody were visualized with the PAP-technique. The in the nucleus profundus mesencephali.chromogen for the reaction was diaminobenzidine (DAB) Substance P (Fig. 2). The distribution of substancewith a heavy metal intensification following the protocol of P-like immunoreactivity was much like that described forAdams [1], the H O was provided by a glucose–oxidase serotonin concerning immunoreactive terminal structures2 2
reaction [88]. After the DAB procedure, sections were in the torus. In general, the label was strong in the laminardehydrated in ethanol and coverslipped with Corbit. The nucleus (Fig. 10B), much weaker in the magnocellularoptimal dilutions of the antibodies are listed below (Table nucleus and very weak in the principal nucleus, so that the1). For the demonstration of estrogen-bp, we followed the nuclei could easily be delineated on the basis of theinstructions of the detection kit but added a heavy metal labeling pattern. Immunoreactive somata were located inintensified DAB-procedure as described above. the principal and the magnocellular nucleus of the torus
Table 1Optimal dilutions of antibodies and detection complex
Antibody against Normal serum 18 antibody 28 antibody PAP–complex
serotonin 1:20 1:40 1:50 1:100substance P 1:20 1:2000 1:100 1:100leucine–enkephalin 1:20 1:2000 1:100 1:100parvalbumin 1:20 1:1000 1:100 1:100GABA (polyclonal) 1:20 1:1000 1:50 1:100GABA (monoclonal) 1:33 1:3000 1:100 1:100tyrosine–hydroxylase 1:33 1:400 1:100 1:100D2-receptor 1:33 1:500 1:100 1:100aspartate 1:33 1:500 1:100 1:100
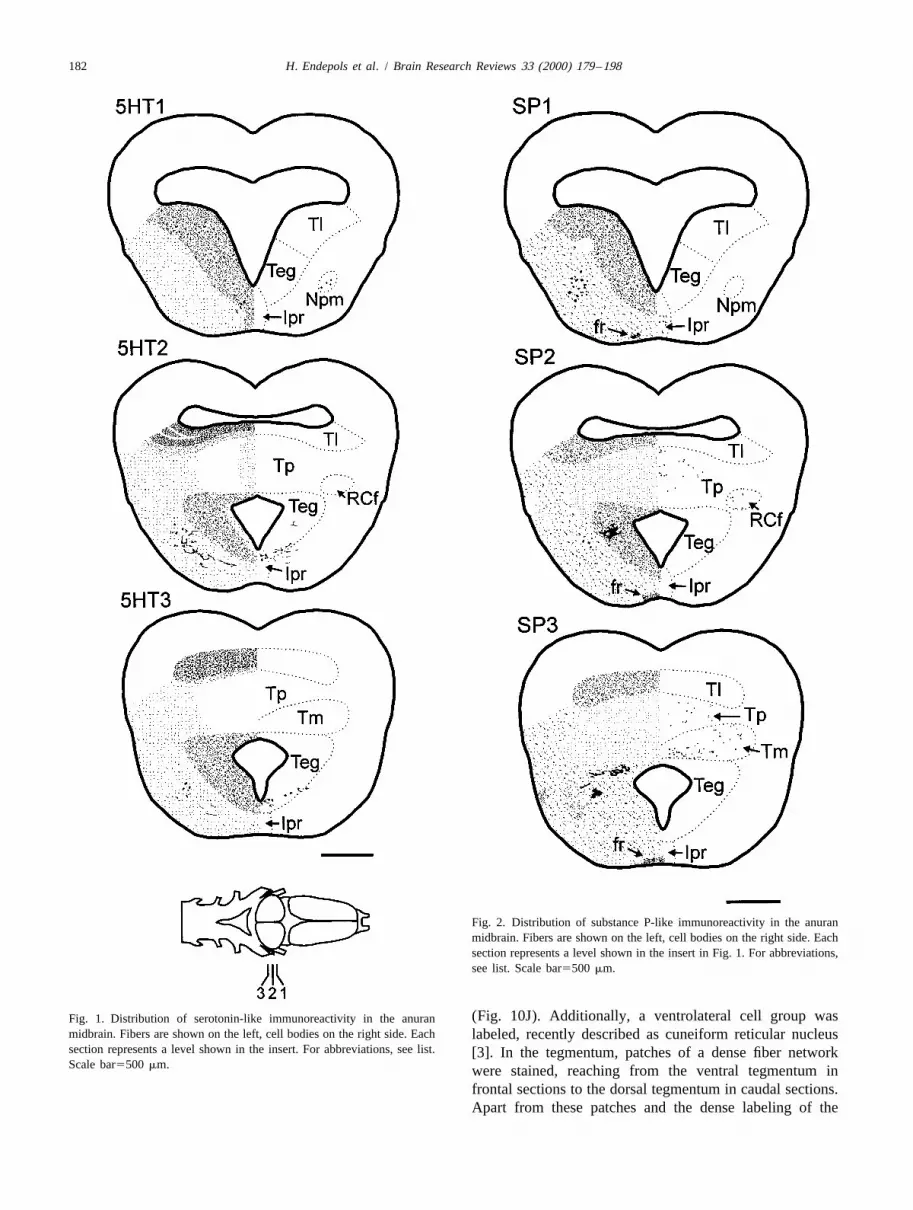
182 H. Endepols et al. / Brain Research Reviews 33 (2000) 179 –198
Fig. 2. Distribution of substance P-like immunoreactivity in the anuranmidbrain. Fibers are shown on the left, cell bodies on the right side. Eachsection represents a level shown in the insert in Fig. 1. For abbreviations,see list. Scale bar5500 mm.
(Fig. 10J). Additionally, a ventrolateral cell group wasFig. 1. Distribution of serotonin-like immunoreactivity in the anuranmidbrain. Fibers are shown on the left, cell bodies on the right side. Each labeled, recently described as cuneiform reticular nucleussection represents a level shown in the insert. For abbreviations, see list. [3]. In the tegmentum, patches of a dense fiber networkScale bar5500 mm. were stained, reaching from the ventral tegmentum in
frontal sections to the dorsal tegmentum in caudal sections.Apart from these patches and the dense labeling of the

H. Endepols et al. / Brain Research Reviews 33 (2000) 179 –198 183
fasciculus retroflexus (a ventromedially descending fiber ventricle and the ventral surface of the brain in the mosttract from the habenular nuclei to the interpeduncular rostral midbrain sections. This nucleus has been describednucleus), immunopositive fibers were homogeneously dis- as rostral portion of the interpeduncular nucleus [46] ortributed throughout the tegmentum in a density comparable nucleus of the medial longitudinal fasciculus [73].to the magnocellular nucleus. Labeled cells formed a Leucine–enkephalin (Fig. 3). The density of leucine–narrow group, located between the ventromedial tip of the enkephalin-immunoreactive fibers was high in the laminar
nucleus (Fig. 10C), much lower in the magnocellularnucleus and lowest in the principal nucleus. The arealabeled in the laminar nucleus was smaller than thesubstance P-immunoreactive zone but still broad comparedto the labeling pattern obtained with the serotonin-anti-body. Immunoreactive somata were found in the laminarnucleus and the magnocellular nucleus. In the tegmentum,the density of labeled fibers was high near the Sylvianaqueduct, and only a little lower in all other parts of thetegmentum. Thick patches of immunoreactive structureswere found — similar to the substance P-labeling — in theventral and the dorsal tegmentum. Cell bodies were labeledin the ventral and the dorsal tegmentum.
Tyrosine–hydroxylase (Fig. 4). Tyrosine–hydroxylase-immunostaining produced strong labeling of somata, den-drites and axons with a very low background. In allsubnuclei of the torus semicircularis labeled fibers werefound; their density was highest in the laminar nucleus(Fig. 10D). The axons passed between the cell layers,forming horizontally oriented ‘stripes’. Relatively highdensity of tyrosine–hydroxylase-immunoreactive terminalsoccurred also in the medial part of the torus, where thestained puncta formed vertically oriented ‘stripes’. Cellbodies were not labeled in the torus. In the wholetegmentum fibers were stained, with the highest densitymedial (close to the Sylvian aqueduct) and ventrolateral.Immunopositive cell bodies formed a narrow unpairedgroup in the ventromedial part of the rostral tegmentumdirectly below the ventral tip of the Sylvian aqueduct(rostral portion of the interpeduncular nucleus). In the mostrostral sections their location overlapped with the sub-stance P-immunoreactive neurons. The thick dendritesand/or axons of the tyrosine–hydroxylase-labeled cellscould be found throughout the ventrolateral tegmentum upto the lateral torus.
Dopamine D2-receptor (Fig. 5). The D2-receptor-anti-body stained cell bodies with high intensity, basal den-drites with moderate, and distal dendrites with lowerintensity. It is not clear whether some axons were alsolabeled — immunonegative axons often appeared white infront of the gray neuropil. In the torus semicircularis,numerous cell bodies were labeled in the laminar, princi-pal, and magnocellular nucleus. The neuropil was homoge-neously stained with only few unlabeled fibers. A highdensity of immunopositive neurons was found in the dorsaland ventral tegmentum (Fig. 10I), and in the interpeduncu-lar nucleus. Lateral from the periventricular cell masses,
Fig. 3. Distribution of LEU–enkephalin-like immunoreactivity in thethe unlabeled fasciculi tegmentales were found, cuttinganuran midbrain. Fibers are shown on the left, cell bodies on the rightthrough radial oriented lightly stained dendrites (andside. Each section represents a level shown in the insert in Fig. 1. For
abbreviations, see list. Scale bar5500 mm. maybe axons). More lateral, immunonegative fibers were

184 H. Endepols et al. / Brain Research Reviews 33 (2000) 179 –198
Fig. 4. Distribution of tyrosine–hydroxylase-like immunoreactivity in the Fig. 5. Distribution of dopamine D2-receptor-like immunoreactivity inanuran midbrain. Fibers are shown on the left, cell bodies on the right the anuran midbrain. Neuropil staining is shown on the left, cell bodies onside. Each section represents a level shown in the insert in Fig. 1. For the right side. Each section represents a level shown in the insert in Fig.abbreviations, see list. Scale bar5500 mm. 1. For abbreviations, see list. Scale bar5500 mm.
localized parallel to the lateral surface of the brain. labeling of additional structures could be achieved. Im-Ventromedially in the caudal midbrain, the interpeduncular munostaining with the monoclonal antibody produced anucleus was stained with a dark background on which the dense network of labeled somata, fibers and terminalcrossing of unlabeled fibers was clearly visible. structures. All neurons were surrounded by immuno-
Gamma-aminobutyric acid (GABA) (Fig. 6). With the positive puncta, presumably terminal boutons, so thatpolyclonal GABA-antibody, only somata were stained; no unlabeled cells were clearly outlined. A high density of

H. Endepols et al. / Brain Research Reviews 33 (2000) 179 –198 185
semicircularis, and in the cuneiform reticular nucleus. Inthe tegmentum, immunopositive fibers were found at highdensity at the ventrolateral surface of the midbrain andbetween the periventricular cell bodies. From the latter,small bundles of stained radial fibers emerged, which cutthrough the unlabeled longitudinal fasciculi tegmentalesand fanned out more lateral. Within this lateral network ofradial fibers, thick and heavily stained structures withlongitudinal orientation could be detected. It seemed thatthese structures were axons and/or dendrites from a groupof GABAergic neurons that had migrated out of theperiventricular mass of tegmental cells, projecting rostradinto the diencephalon. This small nucleus was locatedrostral from the nucleus of the lateral lemniscus and maybe identical with the caudal portion of the nucleus profun-dus mesencephali [17]. Some GABA-immunoreactivesomata were also located in dorsal and ventral parts of therostral tegmentum.
Parvalbumin (Fig. 7). The antibody against parvalbuminstained a dense web of fibers in the principal nucleus of thetorus semicircularis (Fig. 10F). Label was strongest at therostral margin of the principal nucleus, thus constituting asharp border to the laminar nucleus that was completelydevoid of immunoreactive structures. Cell bodies withparvalbumin-like immunoreactivity were situated in themagnocellular nucleus (reaching with their dendrites intothe principal nucleus), in the transition zone betweencaudal laminar nucleus and optic tectum, and in theventrolateral torus (cuneiform reticular nucleus [3]).Among the periventricular tegmental cells only few fiberswere labeled, whereas more medially, close to the Sylvianaqueduct, the density of stained fibers was relatively high.Immunopositive somata were found in the ventral tegmen-tum and in the same laterally migrated tegmental nucleuswhere GABAergic neurons had been labeled.
Aspartate (Fig. 8). In the torus semicircularis, aspartateimmunostaining produced a strong fiber labeling in thelaminar, magnocellular, and principal nucleus; immuno-positive cell bodies were found in the same nuclei. In thetegmentum, the density of labeled fibers was high withinthe periventricular cell mass and at the ventrolateral brainsurface. A few labeled radial fibers cut through thelongitudinal fasciculi tegmentales, but did not reach as farlateral as the GABAergic radial axons. Numerous somatawere stained in the dorsal and ventral tegmentum (Fig.10G), and some aspartate-immunoreactive cells have mi-Fig. 6. Distribution of GABA-like immunoreactivity in the anurangrated laterally.midbrain. Fibers are shown on the left, cell bodies on the right side. Each
section represents a level shown in the insert in Fig. 1. For abbreviations, Estrogen-binding protein (Fig. 9). The distribution ofsee list. Scale bar5500 mm. estrogen-bp’s in anuran brains has not yet been investi-
gated with immunocytochemical methods; therefore, adescription of immunoreactive structures throughout the
immunoreactive fibers were found in the laminar nucleus entire brain will be given. The antibody labeled the nucleiof the torus semicircularis (Fig. 10E); in the principal and of neurons in several brain areas of all species studied. Inmagnocellular nucleus the fiber density was lower. Cell the torus semicircularis, immunoreactive cells were locatedbodies were labeled with both antibodies in the magnocel- exclusively in the laminar nucleus; no immunoreactivelular and laminar nuclei, in the medial zone of the torus cells were found in the tegmentum. Additional cells were

186 H. Endepols et al. / Brain Research Reviews 33 (2000) 179 –198
Fig. 8. Distribution of aspartate-like immunoreactivity in the anuranFig. 7. Distribution of parvalbumin-like immunoreactivity in the anuranmidbrain. Fibers are shown on the left, cell bodies on the right side. Eachmidbrain. Fibers are shown on the left, cell bodies on the right side. Eachsection represents a level shown in the insert in Fig. 1. For abbreviations,section represents a level shown in the insert in Fig. 1. For abbreviations,see list. Scale bar5500 mm.see list. Scale bar5500 mm.
labeled in the lateral septum, the ventral striatum, the geminal nucleus, the secondary isthmal nucleus and in thepreoptic area, the caudal portion of the paraventricular spinal cord were missing in Xenopus laevis. A detailedorgan of the hypothalamic infundibulum, the pretrigeminal comparison between the sexes of the species studied wasnucleus, the caudal portion of the secondary isthmal not useful due to the small sample that does not suffice fornucleus, and in a cell column in the dorsal spinal cord a statistical analysis; however, sex-specific differences incaudal of the obex. Immunopositive neurons in the pretri- the total amount of immunopositive cells probably exist in

H. Endepols et al. / Brain Research Reviews 33 (2000) 179 –198 187
Fig. 9. Distribution of estrogen-binding protein-like immunoreactivity in the anuran midbrain. For better visibility, labeled cells are presented twice aslarge. Each section represents a level shown in the insert. For abbreviations, see list. Scale bar5500 mm.
the preoptic area. In the laminar nucleus of Discoglossus 3.3. Synopsis of nuclei in the brain with somatapictus and Bombina orientalis, labeled cells were mostly immunoreactive for the neuromodulators studieddispersed into the dorsal area of the nucleus. In Xenopuslaevis, the total amount of immunopositive cells in the The distribution of somata in the anuran brain that arelaminar nucleus was considerable larger, and the cells immunoreactive for the neuromodulators studied has beenformed a band that reached from the ventral torus far into described in various studies and will therefore not bethe region of the tectum opticum (Fig. 10H). However, mentioned in detail. Table 2 gives an overview of oursince the torus in Xenopus extends relatively far caudally findings to provide a basis for the following discussion ofto the tectum, these cells probably belong to the laminar the afferent sources of modulatory input to the laminarnucleus as well. nucleus of the torus semicircularis.

188 H. Endepols et al. / Brain Research Reviews 33 (2000) 179 –198
Fig. 10. Immunoreactive fibers and cell bodies in the midbrain of Discoglossus pictus (D.p.), Bombina orientalis (B.o.) and Xenopus laevis (X.l.). A:serotonin (D.p.), B: substance P (B.o.), C: LEU–enkephalin (D.p.), D: tyrosine–hydroxylase (D.p.), E: GABA (D.p.), F: parvalbumin (D.p.), G: aspartate(D.p.), H: estrogen-binding protein (X.l.) I: dopamine D2-receptor (B.o.), J: substance P (X.l.). K: schematic frontal midbrain section (D.p.); italic lettersshow the location of photographs A–F (laminar and principal nucleus), and G1I (tegmentum). L: schematic parasagittal section (X.l.); italic letters showthe location of photographs H (laminar and principal nucleus) and J (principal nucleus). For abbreviations, see list. Scale bar in J (for A–J)5100 mm; scalebar in K and L5500 mm.

H. Endepols et al. / Brain Research Reviews 33 (2000) 179 –198 189
Fig. 10. (continued)

190 H. Endepols et al. / Brain Research Reviews 33 (2000) 179 –198
Table 2aNuclei with immunopositive somata in the anuran brain
Serotonin Substance P Leucine–enkephalin Tyrosine–hydroxylase
Paraventricular organ of the Striatum Olfactory bulb (D.p., X.l.) Olfactory bulb
hypothalamic infundibulum Lateral amygdala (D.p.) Lateral pallium Preoptic area
Nucleus profundus Anterior preoptic nucleus Striatum Suprachiasmatic nucleus
mesencephali Anterior thalamic nucleus Medial septum (D.p., X.l.) Ventromedial thalamic nucleus
Raphe nucleus Ventromedial thalamic nucleus (D.p., X.l.) Lateral amygdala (D.p., X.l.) Posterior thalamic nucleus
Posterior tuberculum (D.p., X.l.) Anterior preoptic nucleus Posterior tubercle
Ventral hypothalamic nucleus Magnocellular preoptic nucleus Dorsal hypothalamic nucleus
Principal nucleus Suprachiasmatic nucleus Rostral portion of ventromedial tegmentum
Optic tectum Anterior thalamic nucleus (D.p.) Locus coeruleus
Rostroventral tegmentum Ventromedial thalamic nucleus (D.p.) Nucleus of the solitary tract
Dorsocaudal tegmentum Ventral hypothalamic nucleus
Optic tectum (D.p., B.o.)Laminar nucleus
Magnocellular nucleus
Tegmentum
Nucl. cerebelli (D.p., X.l.)Spinal cord
a D.p.: Discoglossus pictus; B.o.: Bombina orientalis; X.l.: Xenopus laevis.
4. Discussion the ventral tegmentum and in the nucleus profundusmesencephali is similar to the results in ranids [2,16,106].
4.1. Immunohistochemical findings However, there is a discrepancy in two structures in whichimmunopositive neurons have been found by other authors,
Eight of the antisera (i.e., serotonin, leucine–enkephalin, namely the ventrolateral torus semicircularis (cuneiformsubstance P, GABA, parvalbumin, tyrosine–hydroxylase, and subcuneiform nucleus; [3]) and the optic tectum [56].dopamine D2-receptor, and aspartate) yielded a specific In these nuclei, we never found immunopositive somata inlabeling of somata, dendrites, axonal and terminal struc- any of the three species studied here.tures. Our results did not reveal principal species-specific Data concerning the distribution of serotonin-immuno-differences in the distribution of immunoreactive structures reactive terminal structures in the amphibian brain arein the torus semicircularis. The monoclonal antibody sparse because most authors concentrate on immuno-against the estrogen-bp had not been applied in amphibian reactive somata or terminals in the spinal cord and thematerial before, but yielded a clearly detectable label of optic tectum. Using an autoradiographic approach,cell nuclei in the areas already described by Kelley et al. Diederen et al. [16] found a high concentration of immuno-[45]. reactive structures in the tectum opticum but not in the
To delineate putative sources for the neurotransmitter / torus; however, this finding might be ascribed to the poorneuromodulatory afferents to the torus semicircularis we resolution of the autoradiographic method. In salamanders,will combine our immunohistochemical data with data on serotonin-positive somata and fibers were reported in thethe connectivity of the anuran torus semicircularis pub- ventral tegmentum [15].lished by various authors (e.g., [23,24,57,72,99]). It has to The probable source for the serotoninergic innervationbe emphasized that most of these studies have not taken of the laminar nucleus can be delineated as the nucleusthe toral subdivision into account, and many studies have raphe of the medulla oblongata since it contains immuno-reported contradictory findings. We will therefore discuss positive somata and projects to the torus semicircularisall connections that have been reported to include every [57] and to the tegmentum (unpublished observations). Apossible afferent source. contribution of neurons in the paraventricular nucleus of
The connections of the anuran tegmentum have not yet the hypothalamic infundibulum is questionable; the ventralbeen investigated in detail. However, there is a wealth of hypothalamus has been reported to project to the laminardata on efferent projections from other brain areas that nucleus [74,98], but this finding could not be confirmed byinclude the tegmentum [39,49,50,61,62,65,92,100]. Still, Feng and Lin [24].our knowledge about the sources of tegmental afferents is Substance P. Several studies have investigated thepresumably incomplete. distribution of substance P-like immunoreactivity in anuran
A summary of the putative neuromodulatory input into brains [2,3,14,41,47,64]. However, these authors studiedthe laminar nucleus and the tegmentum is shown in Fig. ranid or hylid brains and concentrated on the spinal cord11. and the tectum opticum, respectively. Similar to our
Serotonin. The localization of immunoreactive somata in findings, immunoreactive terminal structures concentrated
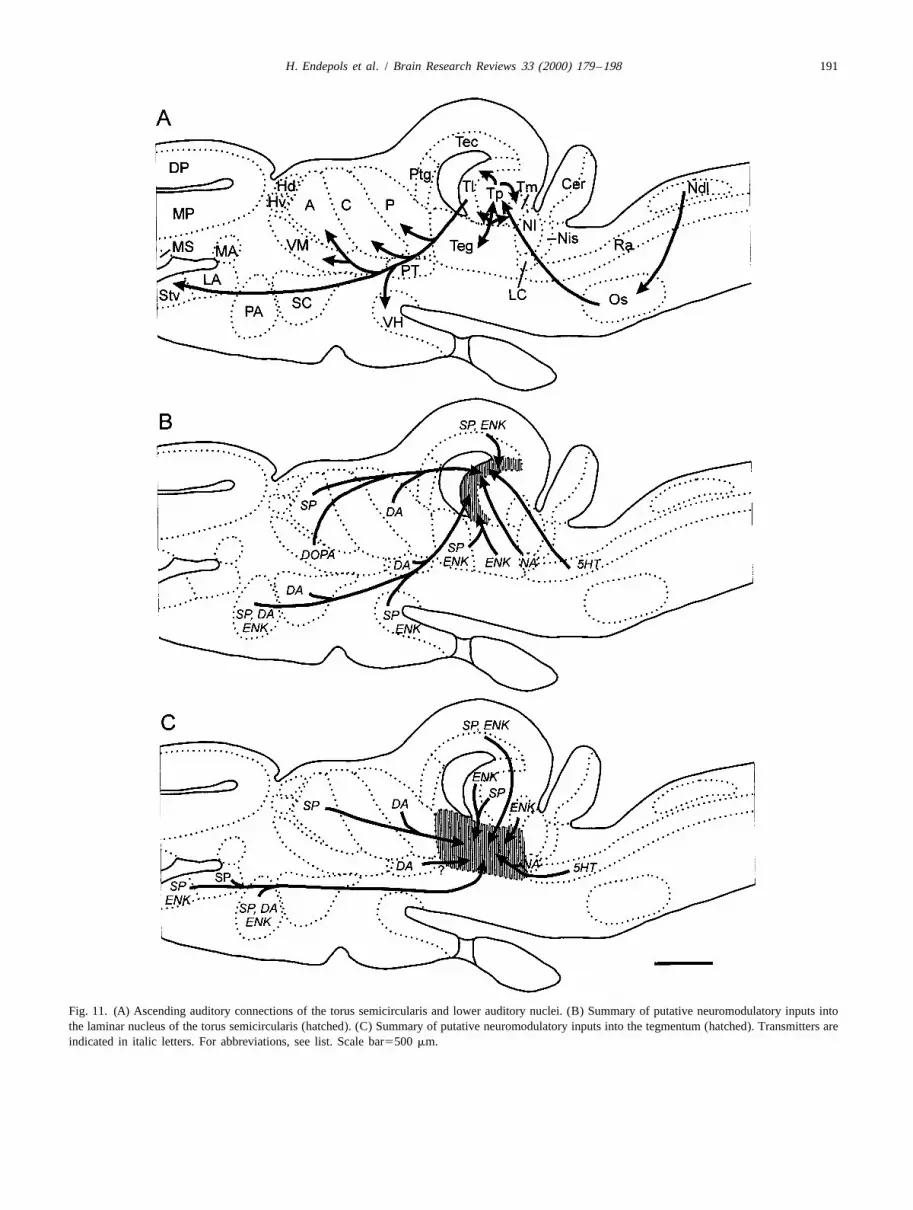
H. Endepols et al. / Brain Research Reviews 33 (2000) 179 –198 191
Fig. 11. (A) Ascending auditory connections of the torus semicircularis and lower auditory nuclei. (B) Summary of putative neuromodulatory inputs intothe laminar nucleus of the torus semicircularis (hatched). (C) Summary of putative neuromodulatory inputs into the tegmentum (hatched). Transmitters areindicated in italic letters. For abbreviations, see list. Scale bar5500 mm.

192 H. Endepols et al. / Brain Research Reviews 33 (2000) 179 –198
in the laminar nucleus of the torus semicircularis, and tional tegmental afferents originate in the posterior tuber-labeled somata were found in the principal nucleus of culum is likely, but so far unknown.Rana pipiens [3,47]. Tyrosine–hydroxylase-immunoreactive terminals in both
The substance P-ergic input to the torus may have torus and tegmentum comprise dopaminergic as well asseveral sources as many immunopositive nuclei have been noradrenergic structures. Purely noradrenergic arereported to project to the laminar nucleus, i.e., the preoptic tyrosine–hydroxylase-positive cells in the locus coeruleus,area, the anterior thalamic nucleus, the ventral hypo- whereas the nucleus of the solitary tract consists ofthalamus, the optic tectum and the anteroventral tegmen- noradrenergic as well as dopaminergic cells. The TH-irtum [11,24,98]. A contribution of principal neurons to the neurons in the ventromedial thalamic nucleus are thoughtsubstance P-like terminal zone is rather unlikely as the to be DOPAergic and all other TH-ir nuclei are purelyprincipal nucleus projects not or only very weakly into the dopaminergic [31,32].laminar zone [58]. Dopamine D2-receptor. So far, dopamine D2-receptors
Substance P-positive nuclei that project to the tegmen- have been demonstrated only in the olfactory bulb [18] andtum are the striatum, the preoptic area [64], the anterior the retina [70,93] of anurans. The occurrence of D2-thalamic nucleus, and the principal nucleus of the torus receptors in the anuran midbrain in this study correspondssemicircularis. The dense fiber plexus in the tegmentum with the distribution of tyrosine–hydroxylase-immuno-may arise from substance P-ergic neurons in the striatum reactive terminals which were abundant in torus semicir-[91]. cularis and tegmentum. The high density of cells with
Leucine–enkephalin. Studies with antibodies directed dopamine D2-receptor-like immunoreactivity underlinesagainst leucine–enkephalin in ranids have yielded results the importance of dopaminergic modulation in the anurancomparable to our data [2,3,47,52]. In the latter three, brain.frontal midbrain sections of Rana pipiens and Rana GABA. Several studies have investigated the distributionesculenta show a labeled terminal pattern corresponding to of GABAergic structures in amphibian brainsour findings, but immunoreactive somata in the torus and [5,28,71,82,85], but immunoreactive neurons in the mag-the tegmentum were only reported in one study [3]. nocellular nucleus have not been mentioned yet as most
Many of the leucine–enkephalin-immunopositive nuclei authors concentrated on other brain areas, e.g., the opticproject to the torus: preoptic area, ventral hypothalamus, tectum. However, if the distribution of immunoreactiveoptic tectum, and the anterodorsal and posteroventral perikarya in these brain areas in our study is compared totegmentum. Additionally, intrinsic projections from the the findings of the authors listed above, the pattern ofmedially located immunopositive neurons of the laminar labeling is well in agreement, indicating that the antibodiesand magnocellular nucleus could be involved as well. used binds to the appropriate epitope.However, as the amount of immunopositive cells in the Parvalbumin. Studies investigating the distribution oftorus is rather low, the massive innervation probably arises parvalbumin-like immunoreactivity in amphibian brainsmainly from the other sources. Taking the strength of the have not been published yet. Parvalbumin has been shownafferent projections into consideration, our data point to be colocalized with GABA in various systems [6,9], andtowards a dominance of tegmental origin of leucine– it can be speculated that the parvalbumin-like immuno-enkephalinergic terminals in the laminar nucleus. reactive cells in the magnocellular nucleus and the tegmen-
The dense fiber networks in the tegmentum represent tum are identical to the neurons stained with the antibodyterminal structures of fibers descending via the lateral against GABA. As further evidence (double labelingforebrain bundle [50], the somata of which are located in studies) is lacking, this interpretation can not be confirmedthe striatum and the lateral amygdala. Further en- yet. The cells of the magnocellular nucleus that showkephalinergic tegmental afferents may originate in the parvalbumin-like immunoreactivity are typical neurons ofpreoptic area (unpublished observations), the optic tectum this nucleus with large somata and a multipolar dendriticand in the laminar and magnocellular nucleus of the torus arborization. In an intracellular study [58], a similarsemicircularis. magnocellular cell type has been found that has a large
Tyrosine–hydroxylase. Our immunostainings with the dendritic field and an extensive axonal arborization withintyrosine–hydroxylase antibody corroborate the results from the entire principal nucleus. However, the terminal struc-ranids [31,64,106], and Xenopus laevis [32]. Catechol- tures found in the principal nucleus may arise fromaminergic input to the torus could possibly derive from the additional sources throughout the brain as well (e.g.,preoptic area, suprachiasmatic nucleus, ventromedial or dorsolateral nucleus and superior olive).posterior thalamus, posterior tuberculum, and the locus Aspartate. Though other authors have already suggestedcoeruleus. The ventromedial tegmentum does not project that aspartate acts as a transmitter in amphibians [13,30,89]to the torus [24]. Catecholaminergic nuclei that project to this is the first immunohistochemical demonstration ofthe tegmentum are the preoptic area (unpublished observa- aspartatergic neurons in the anuran brain. Several studiestions) and the posterior thalamic nucleus. Whether addi- in mammals support the hypothesis that glutamate and

H. Endepols et al. / Brain Research Reviews 33 (2000) 179 –198 193
aspartate are co-localized transmitters detectable in nerve tion or feedback-modulation of sensory pathways, butcell bodies [33,104,108] and synaptic terminals [36,109]. further evidence is still missing.However, some of the labeled aspartate may be part of the Though enkephalins seem to be involved in modulation‘metabolic pool’ rather than part of the ‘transmitter pool’ of auditory brainstem responses [35,96], their role has notof the cell [77]. been clarified yet. Neurons immunoreactive to enkephalins
Estrogen-binding protein. Immunohistochemical studies have often been found to be intrinsic implying a functionwith antibodies against sex-steroids in anurans have not in the local circuitry of sensory systems; this could explainbeen published yet. Autoradiographic methods have been the abundance of immunoreactive cell bodies withinapplied to localize steroid hormone-concentrating cells in various nuclei throughout the brain. However, the totalthe brain of Xenopus laevis and Rana pipiens amount of immunopositive somata within the torus[44,45,69,84]. After injection of tritiated estradiol in semicircularis is rather low and probably not sufficient toXenopus, Morrell et al. [69] found labeled cells in the explain the rich terminal web in the laminar nucleus,telencephalon, preoptic area, ventral thalamus, hypo- especially as the principal nucleus does not seem to projectthalamic infundibulum, and laminar nucleus of the torus into the laminated zone [57]. It is most likely that there is asemicircularis. With the exception of the thalamic cell leucine–enkephalinergic projection into the torus, probablygroup, this finding is in accordance with the situation we originating in the tegmental nuclei. As the anterodorsalfound in Xenopus laevis. However, in Discoglossus pictus tegmentum receives an enkephalinergic innervation fromand Bombina orientalis we found additional neurons in the telencephalic sources [52] and projects to the deep layerspretrigeminal nucleus (a premotor vocal nucleus) and the of the optic tectum and the laminar nucleus, these elementsspinal cord. We cannot exclude methodological reasons for might be part of an enkephalinergic brain subsystem.these discrepancies since a comparison between au- Besides their direct action on postsynaptic receptors, thetoradiography and immunohistochemistry has not been catecholamines dopamine, noradrenalin, and adrenalinpossible. (tyrosine–hydroxylase-like immunoreactivity) are known
to interact with the classical transmitters released at thesame or at neighboring synapses. Pre- and post-synaptic
4.2. Functional aspects of neurotransmitter / interactions of catecholamines with glutamate [10,25,81]neuromodulator input to the torus as well as with GABA [67,81] have been demonstrated in
mammals. In the auditory system, noradrenalin can modu-The delineation of possible neurotransmitter /neuro- late the excitability of neurons in the superior olivary
modulatory sources allows some considerations on the complex [96]. In the frog spinal cord, adrenalin andfunctional implications of the laminar nucleus and the noradrenalin modulate the glutamatergic and aspartatergictegmentum. Both candidates for afferent serotoninergic reflex transmissions [102], and it seems likely that cat-input (paraventricular organ of the infundibulum and echolamines also modify the glutamatergic /aspartatergicnucleus raphe) are part of general arousal brain systems. In and GABAergic transmissions in torus and tegmentum.mammals, serotonin has been ascribed a variety of func- GABAergic inhibition in the anuran torus semicircularistions depending on the receptor subtypes [101], among mediates inhibitory frequency tuning which leads to shap-others the modulation of auditory computation in brain- ing of the frequency selectivity of torus neurons [29,37].stem centers [12,19,27] and auditory cortex [43]. In the IC Furthermore, GABA is involved in binaural inhibition onthe modulatory effects even depend on the auditory signal the level of the torus semicircularis and is important forpresented [42]. Since detailed studies in amphibians have direction dependence in frequency tuning [107]. In mam-shown complex effects on motoneurons [40] and on tectal mals and birds, inhibitory mechanisms have also beenneurons [60], the systemical role of serotonin seems to be shown to be essential in the shaping of neuronal responsethe regulation of arousal state, modulating motor as well as characteristics in several nuclei of the auditory pathwaysensory systems. [8,20,68]. The finding that in anurans at least a part of the
Substance P, first thought to be correlated to pain magnocellular neurons are GABAergic allows furtherperception, is nowadays believed to be involved in various speculations on the functional role of this nucleus. A recentprocesses including presynaptic modification. While a intracellular study of toral neurons has shown a subtype ofspecialized receptor is still unknown, there is good evi- magnocellular neurons with an axonal arborization thatdence that at the cellular level substance P is usually covers large areas of the principal area of the torus [58].colocalized with at least one ‘classical’ transmitter which These neurons, if identical with the GABAergic subpopu-often seems to be serotonin [54] (but see [79]) or GABA lation, could modulate the representation of auditory[34]. In mammals, substance P depolarizes olivocochlear stimuli in the torus. The calcium-binding protein parval-neurons, possibly via descending projections from the bumin has been reported to be colocalized with GABA ininferior colliculus or the auditory cortex [96,97]. This various brain areas. In the torus semicircularis, we have nofinding might indicate a role of substance P in automodula- clear evidence for a colocalization yet. However, the dense

194 H. Endepols et al. / Brain Research Reviews 33 (2000) 179 –198
parvalbumin-positive terminal web in the principal nucleus studied; however, as we did not find any species-dependentmight be attributed to the intracellularly characterized differences in the localization of immunoreactive terminalsmagnocellular subtype mentioned above. in our study, this data will probably also account for
As has been described above, behavioral reactions of anurans in general.anuran amphibians strongly depend on the hormonal As a general conclusion, neuromodulatory afferents tosituation that typically changes seasonally. Kelley [44] the torus semicircularis focus on the laminar nucleus.already pointed out that the laminar nucleus of the torus Another interesting generalization that can be made is thatsemicircularis might be a key element for the hormonal the density of immunoreactive structures in the tegmentuminfluence on acoustic communication. Steroid hormones is almost always comparable with the density found in themay alter the expression of genes and could thus induce laminar nucleus. This holds true for each of the transmit-morphological as well as physiological changes, for exam- ters we investigated as well as for the substances studied inple, dendritic outgrowth, expression of dendritic spines and the papers mentioned above. This finding is of specialchanges in the susceptibility towards neuromodulators by interest since the tegmentum has got massive reciprocalchanging the transcription of specific receptor proteins connections with the laminar nucleus of the torus, similar[66,90,103]. It would be very interesting to investigate connectivity with nuclei throughout the entire brain, andwhether estrogen-receptor positive neurons have mor- comparable axonal projection patterns of individual neu-phological features varying seasonally. Our finding that the rons [95].antibody (that is available commercially) works well inamphibians offers the possibility for double-labeling 4.4. Functional anatomy of the torus semicircularisstudies to investigate these questions.
The differentiation of the toral subnuclei has been4.3. Distribution of other neurotransmitters in the difficult since the first denomination [80]. Many inconsis-auditory midbrain tent findings in the literature, especially data on the
connectivity of the auditory pathway, can be ascribed toVarious studies have focused on the distribution of the rather inaccurate differentiation between a ‘laminated’
neurotransmitters and neuromodulators in the amphibian area and a ‘clustered but occasionally laminated’ zonebrain (Table 3). A generalization of these data has to be defining the laminar nucleus and the principal nucleus,done with caution since different species have been respectively. However, as the degree of lamination in the
Table 3aDistribution of other neurotransmitters in the auditory midbrain
Neurotransmitter Torus Tegmentum Species References
fib cb fib cb
acetylcholine Tl1 Tl1 – Teg(1) Rana pipiens [38](via acetylcholine Tp11 Tp1
esterase) Tm1 Tm1
acetylcholine – Teg1 Teg11 Rana perezi, Xenopus laevis [63](via choline acetyl Tp(1) Ipr1transferase) Tm(1)atrial Tl1 – Teg11 Teg1 Rana ridibunda, [26,75]natriuretic factor Ipr11 Ipr1 Hyla japonicabombesin Tl(1) – Teg1 – Rana pipiens, Rana catesbeiana [47,55]dopamine Tl11 – Teg1 – Rana [31]
Ipr(1) ridibundagalanin Tl11 Teg1 Rana esculenta, [51]
Tp(1) Tp(1) Ipr1 Ipr1 Xenopus laevisTm(1)
histamine Tl(1) – Teg1 – Xenopus laevis, Rana catesbeiana [4,55]Ipr1
neuromedin U Tl1 Tl11 Teg1 – Rana esculenta [59]Tm1
pituitary adenylate- Tl1 Teg1 Rana [105]cyclase Tp(1) Tp(1) Ipr1 ridibundaactivating polypeptide Tm(1)proneuropeptide Y Tl11 Teg11 Teg1 Rana esculenta, [53]
Tp(1) Xenopus laevissomatostatin Tl1 Tl(1) Teg11 Teg11 Rana ridibunda, [3,48]
Tp(1) Ipr1 Ipr11 Rana pipiensa cb: cell bodies; fib: fibers; (1) sparse; 1 moderately dense; 11 highly dense; 2 no immunoreactive fibers or cell bodies.

H. Endepols et al. / Brain Research Reviews 33 (2000) 179 –198 195
torus semicircularis seems to be species-specific and necessarily restricted to a single key nucleus in thecoupled to factors like genome size [83], the general midbrain, but rather a common principle in the entirebenefit of such a classification was questionable. Our study audio-motor transmission. The connectivity of the auditoryshows that a delineation of toral subdivisions, especially pathway [24,58] implies that auditory evoked excitation isbetween the laminar and the principal nucleus, is possible processed in both parallel and distributive fashion, leadingwith immunocytochemical methods. In the species we to a complex network with numerous nuclei involved. Byhave examined so far, considerable differences in the modulating these neurons that have widespread connect-relative size of these nuclei were not found; however, the ions with sensory and non-sensory targets, the responseexact outline of the subnuclei in a given species may differ characteristics of the entire network will be altered,and should therefore be investigated in detail to differen- possibly allowing audio-motor shortcuts that are inacces-tiate correctly. sible during a different state of the system.
Within the auditory pathway, the torus semicircularishas an outstanding position due to its size, its connectivity,and the physiology of its neurons. The connectivity of the
Acknowledgementsmagnocellular nucleus and especially the laminar nucleus[24,57,58] imply a predominant role in the process of
We thank Dr. K. Braun (Magdeburg), Dr. J.J. Mildeaudio-motor interfacing. Of these two nuclei, the laminar(Cologne), and Dr. A. Schmidt (Bremen) for providingnucleus seems to be the more promising candidate becauseseveral antibodies, as well as Abbott Diagnostica (Wies-of the cytoarchitecture of its neurons and the comparablybaden, Germany) for the generous gift of the estrogen-bpstrong descending efferents. The functional role of thedetection kit. Dr. J.J. Milde, Dr. R. Wegerhoff (Kiel), andmagnocellular nucleus is still vague; however, the findingM. Wallstein gave many helpful comments. This work wasof neurons with GABA-like immunoreactivity in thissupported in part by the Deutsche Forschungsgemein-structure and the cytoarchitecture of the neurons imply anschaft.intrinsic function additional to ascending and descending
projections.The massive input of transmitters that are known to act
as neuromodulators in other systems, and the presence of Referenceshormone-sensitive neurons in the laminar nucleus stronglyconfirm the importance of this structure. It has already [1] J.C. Adams, Heavy metal intensification of DAB-based HRPbeen shown that acoustical stimulation has an influence on reaction product, J. Histochem. Cytochem. 29 (1981) 775.
[2] D.S.H. Adli, B.M. Rosenthal, G.L. Yuen, R.H. Ho, W.L.R. Cruce,hormone level [7] and that the hormonal level is cruciallyImmunohistochemical localization of substance P, somatostatin,controlling the vocal reproductive behavior [78,94]. Ster-enkephalin, and serotonin in the spinal cord of the northern leopardoid hormones may influence the dendritic arborization offrog, Rana pipiens, J. Comp. Neurol. 275 (1988) 106–117.
target cells, thus possibly altering the connectivity as well [3] D.S.H. Adli, S.L. Stuesse, W.L.R. Cruce, Immunohistochemistry andas the input strength of afferent connections. Additionally, spinal projections of the reticular formation in the northern leopard
frog, Rana pipiens, J. Comp. Neurol. 404 (1999) 387–407.the short-term variations of the animal’s endogenous state[4] M.S. Airaksinen, P. Panula, Comparative neuroanatomy of the(arousal, distress, rest) have a strong influence on acousti-
histaminergic system in the brain of the frog Xenopus laevis, J.cally guided behavior. The neuromodulatory afferents toComp. Neurol. 292 (1990) 412–423.
the laminar nucleus might be the neuronal substrate for all [5] M. Antal, Distribution of GABA-immunoreactivity in the opticthese influences. It is interesting to note that the modulat- tectum of the frog: a light and electron microscopic study, Neuro-
science 42 (1991) 879–891.ory input to the laminar nucleus terminates in the lami-[6] L. Arckens, A. Rosier, C.W. Heizmann, G.A. Orban, F. Vandesande,nated zone, i.e., on the somata and the tangential dendrites
Partial colocalization of the GABA-A receptor with parvalbuminof the laminar neurons and not onto the radial dendritesand calbindin D-28k in neurons of the visual cortex and the dorsal
that deeply reach into the principal nucleus [22,57,95]. The lateral geniculate nucleus of the cat, J. Chem. Neuroanat. 8 (1994)functional significance of this finding is unclear but might 1–10.
[7] J. Brzoska, H.-J. Obert, Acoustic signals influencing the hormoneimply a segregation of classical transmitter input andproduction of the testes in the grass frog, J. Comp. Phys. 140 (1980)modulatory input onto the laminar cells.25–29.It has to be emphasized that the putative modulatory
[8] J.H. Casseday, D. Ehrlich, E. Covey, Neural tuning for soundafferents to the laminar nucleus can not account for all the duration: Role of inhibitory mechanisms in the inferior colliculus,plasticity seen in the behavioral reactions of anurans to Science 264 (1994) 847–850.
[9] M.R. Celio, Parvalbumin in most GABA-containing neurons of theacoustic stimuli. The tegmentum probably representsrat cerebral cortex, Science 231 (1986) 995–997.another level of sensory and (pre-) motor integration and
[10] C. Cepeda, Z. Radisavljevic, W. Peacock, M.S. Levine, N.A.receives comparable input (with the exception of steroidBuchwald, Differential modulation by dopamine of responses
hormones), and even in the auditory brainstem nuclei evoked by excitatory amino acids in human cortex, Synapse 11neuromodulatory afferents can be found. Thus, modulation (1992) 330–341.of neuronal activity in the audio-motor pathway is not [11] B.H. Chahoud, M.J. Cordier-Picouet, P. Clairambault, Larval de-

196 H. Endepols et al. / Brain Research Reviews 33 (2000) 179 –198
´velopment of tectal efferents and afferents in Xenopus laevis [31] A. Gonzalez, W.J.A.J. Smeets, Comparative analysis of dopamine(Amphibia Anura), J. Hirnforsch. 37 (1996) 519–535. and tyrosine hydroxylase immunoreactivities in the brain of two
[12] H. Cransac, J.M. Cottet-Emard, S. Hellstrom, L. Peyrin, Specific amphibians, the anuran Rana ridibunda and the urodele Pleurodelessound-induced noradrenergic and serotonergic activation in central waltlii, J. Comp. Neurol. 303 (1991) 457–477.auditory structures, Hear. Res. 118 (1998) 151–156. ´[32] A. Gonzalez, W.J.A.J. Smeets, Catecholamine systems in the CNS of
[13] R.A. Davidoff, J.C. Hackman, A.M. Holohean, J.L. Vega, D.X. amphibians, in: W.J.A.J. Smeets, A. Reiner (Eds.), Phylogeny andZhang, Primary afferent acitivity, putative excitatory transmitters development of catecholamine systems in the CNS of vertebrates,and extracellular potassium levels in frog spinal cord, J. Physiol. Cambridge University Press, 1994, pp. 77–102.(Lond.) 397 (1988) 291–306. [33] A. Gonzalo-Ruiz, J.M. Sanz, A.R. Lieberman, Immunohistochemi-
[14] E.A. Debski, M. Constantine-Paton, The development of non-retinal cal studies of localization and co-localization of glutamate, aspartateafferent projections to the frog optic tectum and the substance P and GABA in the anterior thalamic nuclei, retrosplenial granularimmunoreactivity of tectal connections, Dev. Brain Res. 72 (1993) cortex, thalamic reticular nucleus and mammillary nuclei of the rat,21–39. J. Chem. Neuroanat. 12 (1996) 77–84.
[15] U. Dicke, M. Wallstein, G. Roth, 5-HT-like immunoreactivity in the [34] A.M. Graybiel, Neurotransmitters and neuromodulators in the basalbrains of plethodontid and salamandrid salamanders (Hydromantes ganglia, Trends Neurosci. 13 (1990) 244–254.italicus, Hydromantes genei, Plethodon jordani, Desmognathus [35] K. Gregorowicz, S. Kosinski, W.Z. Traczyk, Alteration of brain stemochrophaeus, Pleurodeles waltl): an immunohistochemical and
auditory evoked potentials after intracerebroventricular administra-biocytin double-labelling study, Cell Tissue Res. 287 (1997) 513–
tion of met–enkephalin in rabbits, Acta Physiol. Pol. 41 (1990)523.
63–70.[16] J.H.B. Diederen, H.G.B. Vullings, M. Terlou, Catecholaminergic and
[36] V. Gundersen, F.A. Chaudhry, J.G. Bjaalie, F. Fonnum, O.P.serotoninergic brain structures in european green frogs: an au-Ottersen, J. Storm-Mathisen, Synaptic vesicular localization andtoradiographical study, Bas. App. Histochem. 29 (1985) 297–308.exocytosis of L-aspartate in excitatory nerve terminals: a quantitative[17] H.J. ten Donkelaar, Anurans, in: R. Nieuwenhuys, H.J. ten Don-immunogold analysis in rat hippocampus, J. Neurosci. 18 (1998)kelaar, C. Nicholson (Eds.), The Central Nervous System of6059–6070.Vertebrates, Vol. 2, Springer, Berlin, Heidelberg, New York, 1998,
[37] J.C. Hall, GABAergic inhibition shapes frequency tuning andpp. 1151–1314.modifies response properties in the auditory midbrain of the leopard[18] P. Duchamp-Viret, V. Coronas, J.C. Delaleu, E. Moyse, A. Duchamp,frog, J. Comp. Physiol. A 185 (1999) 479–491.Dopaminergic modulation of mitral cell activity in the frog olfactory
[38] J.C. Hall, M.C. Bunker, Acetylcholinesterase staining in the auditorybulb: a combined radioligand binding-electrophysiological study,brainstem nuclei of the leopard frog, Rana pipiens, Neurosci. Lett.Neuroscience 79 (1997) 203–216.182 (1994) 222–226.[19] U. Ebert, J. Ostwald, Serotonin modulates auditory information
[39] J.C. Hall, A.S. Feng, Evidence for parallel processing in the frog’sprocessing in the cochlear nucleus of the rat, Neurosci. Lett. 145auditory thalamus, J. Comp. Neurol. 258 (1987) 407–419.(1992) 51–54.
[40] A.M. Holohean, J.C. Hackman, R.A. Davidoff, An in vitro study of[20] U. Ebert, J. Ostwald, GABA can improve acoustic contrast in the ratthe effects of serotonin on frog primary afferent terminals, Neurosci.ventral cochlear nucleus, Exp. Brain Res. (1995) 310–322.Lett. 113 (1990) 175–180.[21] H. Endepols, W. Walkowiak, Influence of descending forebrain
[41] M.F. Humphrey, G.M.C. Renshaw, P.D. Kitchener, L.D. Beazley,projections on processing of acoustic signals and audiomotorSubstance P, bombesin, and leucine–enkephalin immunoreactivitiesintegration in the anuran midbrain, Eur. J. Morphol. 37 (1999)are restored in the frog tectum after optic nerve regeneration, J.182–184.Comp. Neurol. 354 (1995) 295–305.[22] A.S. Feng, Morphology of neurons in the torus semicircularis of the
[42] L.M. Hurley, G.D. Pollak, Serotonin differentially modulates re-northern leopard frog, Rana p. pipiens, J. Morph. 175 (1983)sponses to tones and frequency-modulated sweeps in the inferior253–269.colliculus, J. Neurosci. 19 (1999) 8071–8082.[23] A.S. Feng, J.C. Hall, D.M. Gooler, Neural basis of sound pattern
[43] G. Juckel, U. Hegerl, M. Molnar, V. Csepe, G. Karmos, Auditoryrecognition in Anurans, Progr. Neurobiol. 34 (1990) 313–329.evoked potentials reflect serotonergic neuronal activity--a study in[24] A.S. Feng, W.Y. Lin, Differential innervation pattern of threebehaving cats administered drugs acting on 5-HT1A autoreceptors indivisions of frog auditory midbrain (torus semicircularis), J. Comp.the dorsal raphe nucleus, Neuropsychopharmacology 21 (1999)Neurol. 306 (1991) 613–630.710–716.[25] B. Ferry, P.J. Magistretti, E. Pralong, Noradrenaline modulates
[44] D.B. Kelley, Auditory and vocal nuclei in the frog brain concentrateglutamate-mediated neurotransmission in the rat basolateralsex hormones, Science 207 (1980) 553–555.amygdala in vitro, Eur. J. Neurosci. 9 (1997) 1356–1364.
[45] D.B. Kelley, I. Lieberburg, B.S. McEwen, D.W. Pfaff, Autoradio-[26] M. Feuilloley, L. Yon, K. Kawamura, S. Kikuyama, J. Gutkowska,graphic and biochemical studies of steroid hormone-concentratingH. Vaudry, Immunocytochemical localization of atrial natriureticcells in the brain of Rana pipiens, Brain Res. 140 (1978) 287–305.factor (ANF)-like peptides in the brain and heart of the treefrog
[46] M. Kemali,V. Braitenberg, Atlas of the frog’s brain. Springer-Verlag,Hyla japonica: effect of weightlessness on the distribution ofBerlin, Heidelberg, New York, 1969, 74 pp.immunoreactive neurons and cardiocytes, J. Comp. Neurol. 330
[47] R.O. Kuljis, H.J. Karten, Laminar organization of peptide-like(1993) 32–47.immunoreactivity in the anuran optic tectum, J. Comp. Neurol. 212[27] K.K. Fitzgerald, D.H. Sanes, Serotonergic modulation of synapses in(1982) 188–201.the developing gerbil lateral superior olive, J. Neurophysiol. 81
[48] A. Laquerriere, P. Leroux, B.J. Gonzalez, C. Bodeant, R. Benoit, H.(1999) 2743–2752.Vaudry, Distribution of somatostatin receptors in the brain of the[28] M.F. Franzoni, P. Morino, The distribution of GABA-like immuno-frog, Rana ridibunda: Correlation with the localization of somatos-reactive neurons in the brain of the newt Triturus cristatus and thetatin-containing neurons, J. Comp. Neurol. 280 (1989) 451–467.green frog, Rana esculenta, Cell Tissue Res. 255 (1989) 155–166.
´ ´[29] Z.M. Fuzessery, J.C. Hall, Role of GABA in shaping frequency [49] G. Lazar, Efferent pathways of the optic tectum in the frog, Actatuning and creating FM sweep selectivity in the inferior colliculus, J. Biol. Acad. Sci. Hung. 20 (1969) 171–183.
´ ´Neurophys. 76 (1996) 1059–1073. [50] G. Lazar, T. Kozicz, Morphology of neurons and axon terminals[30] M.G. Giovanni, G. Lacey, G. Pepeu, A. Nistri, Release of endoge- associated with descending and ascending pathways of the lateral
neous glutamate and aspartate from the frog spinal cord in vitro, forebrain bundle in Rana esculenta, Cell Tissue Res. 260 (1990)Eur. J. Pharmacol. 195 (1991) 47–53. 535–548.

H. Endepols et al. / Brain Research Reviews 33 (2000) 179 –198 197
´ ´[51] G. Lazar, Z.S. Liposits, P. Toth, S.L. Trasti, J.L. Maderdrut, I. [72] T.J. Neary, Forebrain auditory pathways in ranid frogs, in: B.Merchenthaler, Distribution of galanin-like immunoreactivity in the Fritzsch, M.J. Ryan, W. Wilczynski, T.E. Hetherington, W. Wal-brain of Rana esculenta and Xenopus laevis, J. Comp. Neurol. 310 kowiak (Eds.), The Evolution of the Amphibian Auditory System,(1991) 45–67. Wiley & Sons, New York, 1988, pp. 233–252.
´ ´[52] G. Lazar, J.L. Maderdrut, I. Merchenthaler, Some enkephalinergic [73] T.J. Neary, R.G. Northcutt, Nuclear organization of the bullfrogpathways in the brain of Rana esculenta: an experimental analysis, diencephalon, J. Comp. Neurol. 213 (1983) 262–278.Brain Res. 521 (1990) 238–246. [74] T.J. Neary, W. Wilczynski, Autoradiographic demonstration of
´ ´[53] G. Lazar, J.L. Maderdrut, S.L. Trasti, Z. Liposits, P. Toth, T. hypothalamic efferents in the bullfrog, Rana catesbeiana, Anat. Rec.Kozicz, I. Merchenthaler, Distribution of proneuropeptide Y-derived 187 (1977) 665.peptides in the brain of Rana esculenta and Xenopus laevis, J. [75] P. Netchitailo, M. Feuilloley, G. Pelletier, F. Leboulenger, M.Comp. Neurol. 327 (1993) 551–571. Cantin, J. Gutkowska, H. Vaudry, Atrial natriuretic factor-like
[54] S.E. Leeman, Substance P, in: G. Adelman (Ed.), Encyclopedia of immunoreactivity in the central nervous system of the frog, Neuro-¨Neuroscience, Birkhauser, Basel, Boston, Stuttgart, 1987, pp. 1154– science 22 (1987) 341–359.
1156. [76] E.A. Ohr, Tricaine methanesulfonate-I. pH and its effect on anes-[55] J.P. Leger, W.B. Mathieson, Development of bombesin-like and thetic potency, Comp. Biochem. Physiol. 54 (1976) 13–17.
histamine-like innervation in the bullfrog (Rana catesbeiana) central [77] O.P. Ottersen, J. Storm-Mathisen, Different neuronal localization ofnervous system, Brain Behav. Evol. 49 (1997) 63–77.
aspartate-like and glutamate-like immunoreactivities in the hip-[56] Q. Liu, E.A. Debski, Origins of serotonin-like immunoreactivity in
pocampus of rat, guinea-pig and Senegalese baboon (Papio papio),the optic tectum of Rana pipiens, J. Comp. Neurol. 352 (1995)
with a note on the distribution of gamma-aminobutyrate, Neuro-280–296.
science 16 (1985) 589–606.[57] H. Luksch, S. Schmidt-Burr, W. Walkowiak, Cytoarchitecture and[78] M. Penna, R.R. Capranica, J. Somers, Hormone-induced vocalefferent connections of the torus semicircularis in the anuran species
behavior and midbrain auditory sensitivity in the green treefrog HylaDiscoglossus pictus, in: N. Elsner, H. Penzlin (Eds.), Proc. 19thcinerea, J. Comp. Physiol. 170 (1992) 73–82.¨Gottingen Neurobiol. Conf., Thieme, Stuttgart, New York, 1991, p.
¨[79] V.A. Pieribone, L. Brodin, T. Hokfelt, Innunohistochemical analysis127.of the relation between 5-Hydroxytryptamine and neuropeptide-[58] H. Luksch, W. Walkowiak, Morphology and axonal projectionimmunoreactive elements in the spinal cord of an amphibianpatterns of auditory neurons in the midbrain of the painted frog,(Xenopus laevis), J. Comp. Neurol. 341 (1994) 492–506.Discoglossus pictus, Hearing Res. 122 (1998) 1–17.
[80] H.D. Potter, Mesencephalic auditory region of the bullfrog, J.´ ´[59] J.L. Maderdrut, G. Lazar, T. Kozicz, I. Merchenthaler, DistributionNeurophysiol. 28 (1965) 1132–1154.of neuromedin U-like immunoreactivity in the central nervous
[81] E. Pralong, R.S.G. Jones, Interactions of dopamine with glutamate-system of Rana esculenta, J. Comp. Neurol. 369 (1996) 438–450.and GABA-mediated synaptic transmission in the rat entorhinal[60] A.A. Malayev, E.A. Debski, Serotonin modulates induced synapticcortex in vitro, Eur. J. Neurosci. 5 (1993) 760–767.activity in the optic tectum of the frog, Brain Res. 781 (1998)
[82] I. Reichenberger, P. Streit, O.P. Ottersen, N. Dieringer, GABA- and167–181.glycine-like immunoreactivities in the cerebellum of the frog,´ ´[61] O. Marın, A. Gonzalez, W.J.A.J. Smeets, Basal ganglia organizationNeurosci. Lett. 154 (1993) 89–92.in amphibians: efferent connections of the striatum and the nucleus
[83] G. Roth, J. Blanke, D.B. Wake, Cell size predicts morphologicalaccumbens, J. Comp. Neurol. 380 (1997) 23–50.complexity in the brains of frogs and salamanders, Proc. Natl. Acad.´ ´[62] O. Marın, A. Gonzalez, W.J.A.J. Smeets, Anatomical substrate ofSci. USA 91 (1994) 4796–4800.amphibian basal ganglia involvement in visuomotor behaviour, Eur.
[84] E.J. Roy, M.A. Wilson, D.B. Kelley, Estrogen-induced progestinJ. Neurosci. 9 (1997) 2100–2109.receptors in the brain and pituitary of the south african clawed frog´ ´[63] O. Marın, W.J.A.J. Smeets, A. Gonzalez, Distribution of cholineXenopus laevis, Neuroendocrinology 42 (1986) 51–56.acetyltransferase immunoreactivity in the brain of anuran (Rana
[85] K.K. Rybicka, S.B. Udin, Ultrastructure and GABA immuno-perezi, Xenopus laevis) and urodele (Pleurodeles waltl) amphibians,reactivity in layers 8 and 9 of the optic tectum of Xenopus laevis,J. Comp. Neurol. 382 (1997) 499–534.Eur. J. Neurosci. 6 (1994) 1567–1582.´ ´[64] O. Marın, A. Gonzalez, W.J.A.J. Smeets, Basal ganglia organization
[86] R.S. Schmidt, Mating call phonotaxis in female American toads:in amphibians: chemoarchitecture, J. Comp. Neurol. 392 (1998)Lesions of central auditory system, Brain Behav. Evol. 32 (1988)285–312.119–128.[65] T. Masino, P. Grobstein, Tectal connectivity in the frog Rana
[87] R.S. Schmidt, Releasing (unclasping) in male American toads: apipiens: tectotegmental projections and a general analysis of topog-neural substrate in the lateral subtoral tegmentum, Brain Behav.raphic organization, J. Comp. Neurol. 291 (1990) 103–127.Evol. 36 (1990) 307–314.[66] B.S. McEwen, S.E. Alves, Estrogen actions in the central nervous
[88] S. Shu, G. Ju, L. Fan, The glucose oxidase-DAB-nickel method insystem, Endocrin. Rev. 20 (1999) 279–307.peroxidase histochemistry of the nervous system, Neurosci. Lett. 85[67] T. Momiyama, J.A. Sim, Modulation of inhibitory transmission by(1988) 169–171.dopamine in rat basal forebrain nuclei: activation of presynaptic
[89] H. Straka, I. Reichenberger, N. Dieringer, Size-related properties ofD1-like dopaminergic receptors, J. Neurosci. 16 (1996) 7505–7512.vestibular afferent fibers in the frog: uptake of and immunoreactivity[68] P. Monsivais, L. Yang, E.W. Rubel, GABAergic inhibition in nucleusfor glycine and aspartate /glutamate, Neuroscience 70 (1996) 685–magnocellularis: Implications for phase locking in the avian auditory696.brainstem, J. Neurosci. 20 (2000) 2954–2963.
[90] C.D. Toran-Allerand, M. Singh, G. Setalo Jr., Novel mechanisms of[69] J.I. Morrell, D.B. Kelley, D.W. Pfaff, Autoradiographic localizationestrogen action in the brain: new players in an old story, Front.of hormone-concentrating cells in the brain of an amphibian,Neuroendocrinol. 20 (1999) 97–121.Xenopus laevis. II. Estradiol, J. Comp. Neurol. 164 (1975) 63–77.
[91] C.L. Veenman, D. Crzan, H. Kern, P. Wahle, P. van Mier, The[70] Z. Muresan, J.C. Besharse, D2-like dopamine receptors in am-anatomical substrate of telencephalic function, Adv. Anat. Embryol.phibian retina: localization with fluorescent ligands, J. Comp.Cell. Biol. 117 (1989) 1–110.Neurol. 331 (1993) 149–160.
¨[71] C. Naujoks-Manteuffel, W. Himstedt, G. Glasener-Cipollone, Dis- [92] N.P. Vesselkin, T.V. Ermakova, N.B. Kenigfest, M. Goikovic, Thetribution of GABA-immunoreactive neurons in the brain of adult striatal connections in frog Rana temporaria: an HRP study, J.and developing salamanders (Pleurodeles waltli, Triturus alpestris), Hirnforsch. 21 (1980) 381–392.Cell Tissue Res. 276 (1994) 485–501. [93] H.J. Wagner, B.G. Luo, M.A. Ariano, D.R. Sibley, W.K. Stell,

198 H. Endepols et al. / Brain Research Reviews 33 (2000) 179 –198
Localization of D2 dopamine receptors in vertebrate retinae with [103] C.S. Woolley, Effects of estrogen in the CNS, Curr. Opin.anti-peptide antibodies, J. Comp. Neurol. 331 (1993) 469–481. Neurobiol. 9 (1999) 349–354.
[94] W. Walkowiak, Neuroethology of anuran call recognition, in: B. [104] K. Yingcharoen, E. Rinvik, J. Storm-Mathisen, O.P. Ottersen,Fritzsch, M.J. Ryan, W. Wilczynski, T.E. Hetherington, W. Wal- GABA, glycine, glutamate, aspartate and taurine in the perihypo-kowiak (Eds.), The Evolution of the Amphibian Auditory System, glossal nuclei: an immunocytochemical investigation in the catWiley & Sons, New York, 1988, pp. 485–510. with particular reference to the issue of amino acid colocalization,
[95] W. Walkowiak, H. Luksch, Sensory motor interfacing in acoustic Exp. Brain. Res. 78 (1989) 345–357.behavior of anurans, Am. Zool. 34 (1994) 685–695. [105] L. Yon, M. Feuilloley, N. Chartrel, A. Arimura, J.M. Conlon, A.
[96] X. Wang, D. Robertson, Effects of bioamines and peptides on Fournier, H. Vaudry, Immunohistochemical distributuion and bio-neurones in the ventral nucleus of trapezoid body and rostral logical activity of pituitary adenylate cyclase-activating polypep-periolivary regions of the rat superior olivary complex: an in vitro tide (PACAP) in the central nervous system of the frog Ranainvestigation, Hear. Res. 106 (1997) 20–28. ridibunda, J. Comp. Neurol. 324 (1992) 485–499.
[97] X. Wang, D. Robertson, Substance P-induced inward current in [106] M. Yoshida, I. Nagatsu, Y. Kondo, N. Karasawa, T. Ohno, M.identified auditory efferent neurons in rat brain stem slices, J. Spatz, T. Nagatsu, Immunohistocytochemical localization of theNeurophysiol. 80 (1998) 218–229. neurons containing catecholamine-synthesizing enzymes and
[98] W. Wilczynski, Afferents to the midbrain auditory center in the serotonin in the brain of the bullfrog (Rana catesbeiana), Actabullfrog, Rana catesbeiana, J. Comp. Neurol. 198 (1981) 421–433. Histochem. Cytochem. 16 (1983) 245–258.
[99] W. Wilczynski, Brainstem auditory pathways in anuran amphibians, [107] H. Zhang, J. Xu, A.S. Feng, Effects of GABA-mediated inhibitionin: B. Fritzsch, M.J. Ryan, W. Wilczynski, T.E. Hetherington, W. on direction-dependent frequency tuning in the frog inferiorWalkowiak (Eds.), The Evolution of the Amphibian Auditory colliculus, J. Comp. Physiol. A 184 (1999) 85–98.System, Wiley & Sons, New York, 1988, pp. 209–232. [108] N. Zhang, F. Walberg, J.H. Laake, B.S. Meldrum, O.P. Ottersen,
[100] W. Wilczynski, R.G. Northcutt, Connections of the bullfrog Aspartate-like and glutamate-like immunoreactivities in the inferiorstriatum: efferent projections, J. Comp. Neurol. 214 (1983) 333– olive and climbing fibre system: a light microscopic and semiquan-343. titative electron microscopic study in rat and baboon (Papio
[101] L.O. Wilkinson, C.T. Dourish, Serotonin and animal behavior, in: anubis), Neuroscience 38 (1990) 61–80.S.J. Peroutka (Ed.), Serotonin Receptor Subtypes: Basic and [109] M. Zhou, C.L. Peterson, Y.B. Lu, J.V. Nadler, Release of glutamateClinical Aspects, Wiley-Liss, New York, 1991, pp. 147–210. and aspartate from CA1 synaptosomes: selective modulation of
[102] C.J. Wohlberg, J.C. Hackman, R.A. Davidoff, Epinephrine and aspartate release by ionotropic glutamate receptor ligands, J.norepinephrine modulate neuronal responses to excitatory amino Neurochem. 64 (1995) 1556–1566.acids and agonists in frog spinal cord, Synapse 1 (1987) 202–207.