ultrastructure of migrating spinal motoneurons in anuran larvae
TRANSCRIPT

Brain Research, 213 (1981) 30%318 307 © Elsevier/North-Holland Biomedical Press
U L T R A S T R U C T U R E OF M I G R A T I N G SPINAL M O T O N E U R O N S IN
A N U R A N LARVAE
I-WU CHU-WANG, RONALD W. OPPENHEIM and PAUL B. FAREL*
Neuroembryology Laboratory, North Carolina Department of Mental Health, Research Division, Raleigh, N.C. 2761 l and (P.B.F.) Department of Physiology, University of North Carolina, School of Medicine, Chapel Hill, N.C. 27514 (U.S.A.)
(Accepted October 16th, 1980)
Key words': spinal motoneuron - - migration - - ultrastructure - - anuran larva - - lateral motor column
SUMMARY
Motoneurons in the course of migration to the lateral motor column (LMC) were identified by horseradish peroxidase (HRP) applied to lumbar ventral roots in anuran larvae (Rana catesbeiana). The ultrastructural characteristics of these migrating moto- neurons were compared with motoneurons in the LMC that had completed their migration. Both migrating and LMC motoneurons tended to be bipolar with one process extending toward the ependyma and the other out towards the ventral root. Typically, centrally and peripherally directed processes in both migrating and LMC motoneurons contained many microtubules, mitochondria and rosette-like clusters of ribosomes, as did the soma. Four of the five types of synapses found on adult frog motoneurons were also found on migrating motoneurons in tadpoles. Radial glia, whose cell bodies are located in the ependyma, have processes extending to the spinal cord periphery. In less than 10 ~ of the preparations, radial glia were labeled with the H R P reaction product. These labeled glia were further distinguished from migrating motoneurons at the ultra- structural level by the presence of abundant filaments.
INTRODUCTION
Neuronal development is typically considered to be a sequential process in which cell birth and migration of the immature nerve cell are followed by differentiation and axon elongation. In contrast to this generalization, Levi-Montalcini 11 clearly demon- strated axonal outgrowth from sympathetic preganglionic neurons before they assumed
* To whom correspondence should be addressed.

308
their final dorsomedial position. Similarly, Hughes 9 showed that axon elongation of chick spinal motoneurons begins before migration is completed.
The recent use of retrogradely transported horseradish peroxidase (HRP) has confirmed conclusions drawn from these early studies. Heaton et al. 7 found that HRP injected into the first visceral arch of chick embryos retrogradely labeled trigeminal motoneurons before completion of their secondary migration from the common medial motor column into the trigeminal lateral motor nucleus. On the basis of this evidence, Heaton et al. 7 concluded that the axons of these migrating motoneurons had innervated their peripheral target before the cell bodies assumed their final position. In fetal rat, Grobstein 5 labeled migrating motoneurons by applying HRP to the ventral root. Farel and Bemelmans 4 retrogradely labeled both migrating and lateral motor column (LMC) spinal motoneurons by applying HRP to the ninth lumbar ventral root of bullfrog tadpoles; however, when HRP was applied to the limb bud, only motoneurons within the LMC contained the reaction product. In tadpole, then, limb motoneurons in the course of migration to the LMC have axons in the ventral root which have not yet reach- ed the limb bud.
Because migrating motoneurons can be reliably demonstrated in tadpoles of appropriate stages, the tadpole spinal cord can be studied to determine the extent of ultrastructural differentiation of migrating motoneurons and whether these migrating motoneurons receive synapses. The present study utilizes electron microscopic examina- tion of spinal motoneurons retrogradely labeled with HRP to compare the extent of differentiation of migrating motoneurons to that of motoneurons within the LMC. The primary conclusion drawn from this examination is that the two groups ofmotoneurons are quite similar in terms of their general morphology, organelles and the types of synapses they receive.
METHODS
Rana catesbeiana tadpoles between Stages VI and VIII 21 were anesthetized by immersion in 0.02 % Finquel (Ayerst). The ninth and tenth ventral roots, which provide innervation to the hind limb, were exposed by dorsal laminectomy. The roots were crushed 1 4 mm from the spinal cord with no. 5 watchmakers' forceps onto which HRP (Sigma, Type VI) had been recrystalized. Care was taken to minimize contact with the dorsal roots or spinal cord. The forceps were held in place around each root until the HRP had dissolved. Three to four days later, the tadpoles were perfused through the heart (3 % glutaraldehyde) or the lumbar enlargement was fixed directly by immersion (6 % glutaraldehyde). Phosphate buffer (0.1 M, pH 7.3) was used in both procedures.
The tissue remained in buffered glutaraldehyde for approximately 12 h at 4 °C and was then rinsed with phosphate buffer. The lumbar enlargements were supported with agar and 50/zm thick sections were cut with a Sorvall TC-2 tissue sectioner. Sec- tions were reacted according to the HRP procedure of Hanker et al. 6, postfixed in 1% OSO4, dehydrated through a graded ethanol series and propylene oxide, and finally embedded in Epon 812. Thick Epon sections (1-2/~m) were cut with glass knives and lightly stained with methylene blue-azure B for tissue orientation (see Results). Thin
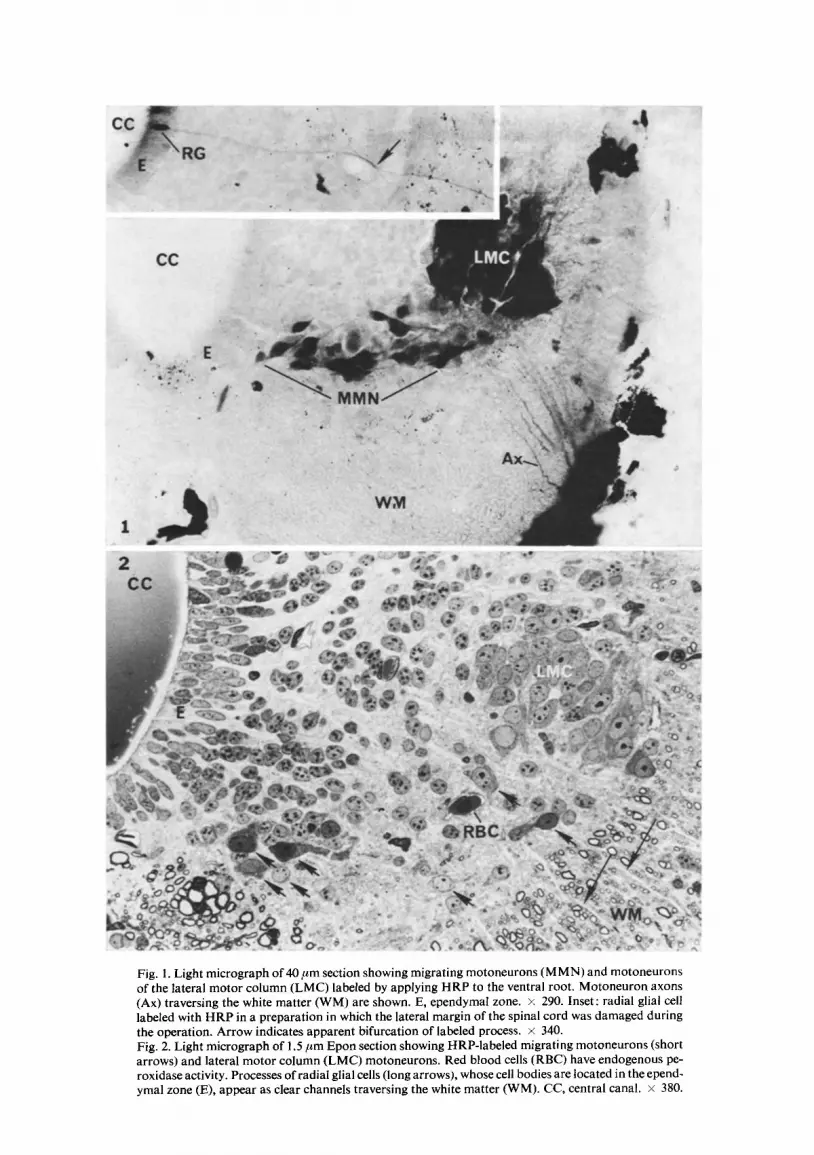
Fig. 1. Light micrograph of 40/~m section showing migrating motoneurons (MMN) and motoneurons of the lateral motor column (LMC) labeled by applying HRP to the ventral root. Motoneuron axons (Ax) traversing the white matter (WM) are shown. E, ependymal zone. x 290. Inset: radial glial cell labeled with HRP in a preparation in which the lateral margin of the spinal cord was damaged during the operation. Arrow indicates apparent bifurcation of labeled process, x 340. Fig. 2. Light micrograph of 1.5/~m Epon section showing HRP-labeled migrating motoneurons (short arrows) and lateral motor column (LMC) motoneurons. Red blood cells (RBC) have endogenous pe- roxidase activity. Processes of radial glial cells (long arrows), whose cell bodies are located in the epend- ymal zone (E), appear as clear channels traversing the white matter (WM). CC, central canal. × 380.

Fig. 3. Electron micrograph of a motoneuron in the lateral motor column. The nucleus (N) and cyto- plasm appear dense due to moderate amounts of the HRP reaction product. The motoneuron receives a type S (TS) synapse. Compare with the migrating motoneuron shown in Fig. 5. GA, Golgi apparatus. × 16,100.
Fig. 4. Electron micrograph of a motoneuron in the LMC showing peripheral process (Ax) extending into the lateral funiculus. Note numerous free rosette-like clusters of ribosomes in the axon as well as in the soma. Axon contains numerous microtubules and receives a type F (TF) synapse. N, nucleus. × 19,800.

3l l
sections (70--90 nm) were then cut from the selected area with a diamond knife, mounted on Formvar-coated slot grids, and stained with uranyl acetate and lead citrate for exa- mination with a Siemens Elmiskop 1A. With the exception of red blood cells and oc- casionally primary motoneurons 4, both of which are easily distinguished from LMC motoneurons, endogenous peroxidase activity was not seen.
RESULTS
As previously found 4, ventral root application of HRP retrogradely labeled motoneurons within the LMC and in the mantle zone. In the caudal regions of the labeled segment, motoneurons containing the reaction product formed an arc between the ventral ependyma and the LMC, giving the impression of migration along a defined path (Fig. 1). In the rostral regions of the labeled segment, migrating motoneurons are found more dorsally between the middle region of the ependyma and the L M C t Between these rostral and caudal areas of the labeled segment which contain migrating motoneurons is a region in which all labeled motoneurons are within the LMC. Most labeled motoneurons, both migrating and in the LMC, were bipolar.
In five of the more than 50 animals studied following HRP application to the ventral root, damage to the lateral marginal zone of the spinal cord resulted in labeling of radial glia (inset, Fig. 1). These cells were distinguished under the light microscope by the location of the cell body in the ependymal zone and the spoke-like projection from the ependymnl zone to the spinal cord surface (Fig. 2). The ultrastructure of radial glia is compared with that of migrating motoneurons below.
Fig. 2 illustrates a 1.5 #m Epon section lightly counterstained. Sections such as this were used to locate regions of HRP-labeled migrating motoneurons. In some in- stances, the HRP reaction product was clearly visible under light microscopy but could not be made out with certainty under the electron microscope. In such sections, the position of specific labeled motoneurons relative to other structures was determined under light microscopy and this map used to locate the same labeled motoneurons under the electron microscope.
General cytological features All ultrastructural observations of HRP-labeled migrating motoneurons were
taken from the migrating motoneuron (MMN) region in Fig. 1. Figs. 3 and 4 illustrate H RP-labeled motoneurons in the LMC. The cytoplasm and nucleus of the motoneuron shown in Fig. 3 appear somewhat electron dense due to the presence of moderate amounts of the diffuse HRP reaction product. The presence of label in the nucleus was an unexpected but consistent finding. Figs. 5 and 6 show HRP-labeled motoneurons in the course of migration. The ultrastructure of migrating motoneurons was similar to that of the motoneurons in the LMC. Typically, both groups have large, round nuclei, the matrix of which appears pale compared to surrounding neurons. Only a single nucleolus was present, as confirmed by serial sections. The cytoplasm of both the somata and processes contains abundant ribosomes, both in free rosette-like clusters and mem- brane-bound. The presence of numerous ribosomes in both processes of these typically

312
Fig. 5. Electron micrograph of migrating motoneuron receiving type F (TF) and type S (TS) synapses. Note similarity ot LMC motoneuron in Fig. 3. N, nucleus, x 13,500. Fig. 6. Electron micrograph of degenerating migrating motoneuron. Many such degenerating moto- neurons were seen, both in the lateral motor column and in the course of migration. The motoneuron receives type C (TC), type F (TF) and type S (TS) synapses. N, nucleus, x 13,550.

Fig. 7. Electron micrograph of migrating motoneuron showing peripherally directed process containing numerous ribosomes and microtubules and receiving a type S (TS) synapse. N, nucleus. × 12,100. Fig. 8. Electron micrograph of centrally directed process of migrating motoneuron receiving type F (TF), type NfS (TNfS) and type S (TS) synapses. Note that a large amount of reaction product is present in this process. × 11,900. Fig. 9. Electron micrograph of a portion of cell body and centrally directed process of migrating moto- neuron receiving a type C (TC) synapse on the soma. Note the similarity of this process to that directed peripherally (Fig. 7). × 12,700.

314
bipolar neurons represents a departure from the usual finding that this organelle is restricted to the cell body and proximal dendrites of neurons. Both the central and peripheral processes of motoneurons in the course of migration and in the LMC con- tained many microtubules (Figs. 4 and 7-9).
Many of the motoneurons examined, both migrating and in the LMC, appeared to be degenerating, as evidenced by dilation of membrane systems (such as the rough endoplasmic reticulum, nuclear envelope and the Golgi apparatus), darkening of the cytoplasm (Fig. 6), and chromatin condensation in the nucleus (type II degeneration of Chu-Wang and Oppenheiml). Whether these motoneurons are dying as a result of the deliberate damage to their axons during HRP application or as part of naturally occurring histogenetic cell death is not known.
Synapses on L M C and migrating motoneurons
Synapses were readily found on both migrating motoneurons and those on the periphery of the LMC and included 4 of the 5 synaptic morphologies distinguished in adult frog spinal cord by Sotelo and Grofova20: type S, in which the presynaptic ter- minal contains spherical vesicles; type F, in which the presynaptic terminal contains tubular vesicles in aldehyde-fixed material (Figs. 5-8); type C, long boutons in which a subsynaptic cistern runs along the postsynaptic membrane (Figs. 6 and 9); and type NfS, in which the presynaptic terminal contains spherical vesicles and a central core of neurofilaments and mitochondria (Fig. 8). Type NfF, similar to type NfS but containing tubular vesicles, was not observed, but this synaptic type is rarely seen even in adult tissue 2°. Synapses were observed on peripheral (Fig. 7) and central (Fig. 8) processes as well as on the somata of migrating cells (Figs. 5, 6 and 9). Type C, S and F synapses were found on both migrating and LMC motoneurons. Only one example of Type NfS was seen, and this synapse was on a migrating motoneuron.
Although a quantitative analysis was not performed, clear differences in the numbers of synapses on different cells were observed. Migrating motoneurons showing signs of degeneration had up to 9 synapses visible in a single section. Healthy appearing migrating motoneurons had fewer synapses, the maximum observed being four. Synap- ses were rarely found on LMC motoneurons except for those motoneurons located on the periphery of the LMC bordering the lateral marginal zone. One such peripherally located LMC motoneuron received 4 synapses, but usually these cells had only one or two synapses. Similarly, during early stages of synapse formation in chick 13 and mouse z2 spinal motoneurons, synapses were virtually always observed only on cells located at the lateral border of the LMC adjacent to the prospective white matter.
Limb motoneurons in the course of migration thus have a substantial synaptic complement, and these synapses match closely the morphological types seen in the adult.
Radial glia
Radial glia have processes which extend from the cell bodies in the ependymal zone to the periphery of the spinal cord and thus could be mistaken for migrating moto- neurons, particularly if they contained the HRP reaction product (see ref. 10 in Fig. 7). Several features distinguish radial glia from migrating motoneurons, both at the light

315
Fig. I0. Electron micrograph of radial glial cell somata in ependymal zone. These cell bodies and their processes are of ten joined by desmosomes (D). Arrow indicates radial glial process running toward cord periphery. Central canal (not shown) is to the left of the figure. N, nucleus, x 9050. Fig. 11. Electron micrograph of radial glial processes in white matter (see long arrows, Fig. 2). Glial filaments (Mf) in bundles are characteristic of these processes. Arrows indicate direction of periphery. x 16,100.

316
and electron microscopic levels. Radial glial processes appear to run from the ependyma to the cord periphery in a fashion resembling spokes on a wheel (inset, Fig. 1, Fig. 2, long arrows) while the peripheral processes of migrating motoneurons are usually directed toward the LMC. The cell bodies of radial glia are readily found within the ependyma and frequently have desmosomal specializations (Fig. 10). Both the cell body and peri- pheral process typically possess numerous filaments. In the white matter (region in- dicated by long arrows, Fig. 2), radial glia processes appear as well-organized, homo- genous structures densely packed with filaments (Fig. 11). In contrast, the central and peripheral processes of migrating motoneurons contain abundant ribosomes and microtubules.
DISCUSSION
This study confirms earlier work that migrating motoneurons can be labeled by ventral root HRP application 4 and shows that these motoneurons are as differentiated as those that have completed their migration. The types of organelles found in migrating and LMC motoneurons appear identical, and little indication of a qualitative difference in the synapses they receive was observed. The HRP-reaction product was generally light and diffusely distributed throughout both the cytoplasm and nucleus, in contrast to results obtained in embryonic chick 2 where the label is localized to the cytoplasm and, while also diffuse, is usually more dense.
The finding that migrating motoneurons can be found from middle limb-bud to late premetamorphic stages 3, combined with preliminary evidence showing [3H]- thymidine incorporation into motoneurons located within the LMC following a two- day post-injection survival time (Farel, in preparation), implies a continuous stream of motoneurons running into the LMC from the ependymal zone over a considerable period of development. These results also indicate that ectopic HRP-labeled moto- neurons are truly migrating rather than undergoing a postmigratory translocationta, 19.
Radial glia were easily distinguished from migrating motoneurons at the ultra- structural level by the absence of microtubules and ribosomes in their processes and by the presence of numerous filaments. Radial glia have been found to provide a substrate for neuronal migration in studies of the developing central nervous system (e.g. refs. 15-17). In tadpole, the course of migrating motoneurons cuts tangentially across the spoke-like processes of the radial glia, making it unlikely that these processes help direct or guide the motoneurons to the LMC (see Figs. 1 and 2). The possibility remains, how- ever, that initial axonal outgrowth through the LMC to the ventral root may follow the course of radial glial fibers.
The finding that, with the exception of synaptic types found only rarely in the adult, the same morphological classes of synapses are found on migrating and mature moto- neurons was unexpected considering the immaturity of the spinal cord as a whole. At the stages examined, segmental primary afferent projections are just beginning to penetrate the dorsal gray and the mean latency of the LMC motoneuron response to electrical stimulation of primary afferent fibers is greater than 10 msec while the response of primary motoneurons to the same stimulus is about 5 msec. Electrophysiological

317
evidence indicates some of the synapses onto L M C motoneurons could be derived from descending fibers 3.
Henrikson and Vaughn s found that at certain stages o f development in embryonic
mouse spinal cord synapses occur between cells not synaptically connected in the adult
(e.g. between neurons and glia). The same phenomenon has been found in chick embryo
spinal cord 14. On the basis o f such evidence, Henrikson and Vaughn s suggested that a
hierarchy of synaptic affinities exists in development such that early-appearing synapses formed between two elements not connected in the adult may later be broken when more
appropriate targets are available. Al though in these studies the transient connections
are between neuronal and non-neuronal elements, transient synapses between neurons have also been identified in a number o f developing systemslL Thus, following this
reasoning, the synapses onto migrating neurons may represent transient connections during a developmental period o f constantly changing patterns o f synaptic association.
This conclusion is supported by the present finding that synapses onto migrating moto-
neurons are much more plentiful than onto motoneurons in the L M C that have com-
pleted migration.
ACKNOWLEDGEMENTS
The authors wish to thank Ms. Sibyl Bemelmans for her help in surgical procedu-
res and tissue preparation. Supported by PHS Gran t NS16030 to P.B.F., N S F Grant BNS24088 and research funds f rom the N.C. Depar tment of Mental Health to R.W,O.
Ancillary support was provided by PHS program project Grant NS14899.
REFERENCES
1 Chu-Wang, I.-W. and Oppenheim, R. W., Cell death of motoneurons in the chick embryo spinal cord I. A light and electron microscopic study of naturally occurring and induced cell loss during development, J. comp. NeuroL, 177 (1978a) 33-58.
2 Chu-Wang, I.-W. and Oppenheim, R. W., Uptake, intraaxonal transport and fate of horseradish peroxidase in embryonic spinal neurons of the chick, J. comp. Neurol., 193 (1980) 753-776.
3 Farel, P. B., Reflex development in anuran larvae: electrophysiological and anatomical correlates, NeuroscL Abstr., 6 (1980) 847.
4 Farel, P. B. and Bemelmans, S. E., Retrograde labeling of migrating spinal motoneurons in bullfrog larvae, Neurosci. Lett., 18 (1980) 133-136.
5 Grobstein, C. S., The development of afferents and motoneurons in rat spinal cord, Neurosci. Abstr., 5 (1979) 161.
6 Hanker, J. S., Yates, P. E., Metz, C. B. and Rustioni, A., A new specific, sensitive and non-carcinog- enic reagent for the demonstration of horseradish peroxidase, Histochem. J., 9 (1977) 789-792.
7 Heaton, M. B., Moody, S. A. and Kosier, M. E., Peripheral innervation by migrating neuroblasts in the chick embryo, Neurosci. Lett., 10 (1978) 55-59.
8 Hendrikson, C. K. and Vaughn, J. E., Fine structural relationships between neurites and radial glial processes in developing mouse spinal cord, J. Neurocytol., 3 (1974) 659-675.
9 Hughes, A., The development of the neural tube of the chick embryo. A study with the ultraviolet microscope, J. EmbryoL exp. Morph., 3 (1955) 305-325.
10 Lamb, A. H., Retrograde axonal transport of horseradish peroxidase for determining motor pro- jection patterns to the developing limb in Xenopus, Brain Research, 134 (1977) 197-212.
11 Levi-Montalcini, R., The origin and development of the visceral system in the spinal cord of the chick embryo, d. MorphoL, 86 (1950) 253-277.
12 Oppenheim, R. W., Neuronal cell death and some related regressive phenomena during neurogene-

318
sis: a selective review and progress report. In W. M. Cowan (Ed.), Studies in Developmental Neuro- biology: Essays in Honor of Viktor Hamburger, Oxford University Press, New York, in press.
13 Oppenheim, R. W., Chu-Wang, I.-W. and Foelix, R. F., Some aspects of synaptogenesis in the spinal cord of the chick embryo: a quantitative electron microscopic study, J. comp. Neurol., 161 (1975) 383-418.
14 Oppenheim, R. W., Chu-Wang, I.-W. and Maderdrut, J. L., Cell death of motoneurons in the chick embryo spinal cord III. The differentiation of motoneurons prior to their induced degeneration following limb-bud removal, J. comp. NeuroL, 177 (1978) 87-112.
15 Rakic, P., Guidance of neurons migrating to the fetal monkey neocortex, Brain Research, 33 (1971) 471-476.
16 Rakic, P., Neuron-glia relationship during granule cell migration in developing cerebellar cortex: a Golgi and electron microscopic study in Macacus rhesus., J. comp. NeuroL, 141 (1971) 283-312.
17 Rakic, P., Mode of cell migration to the superficial layers of fetal monkey neocortex, J. comp. NeuroL, 145 (1972) 61-84.
18 Rakic, P., Prenatal development of the visual system in rhesus monkey, Phil. Trans. B, 278 (1977) 245-260.
19 Sidman, R. L. and Rakic, P., Neuronal migration, with special reference to the human brain: a review, Brain Research, 62 (1973) 1-35.
20 Sotelo, C. and Grofova, I., Ultrastructural features of the spinal cord. In R. Llin~is and W. Precht (Eds.), Frog Neurobiology, Springer-Verlag, New York, 1977.
21 Taylor, A. C. and Kollros, J. J., Stages in the normal development of Rana pipiens larvae, Anat. Rec., 94 (1946) 7-24.
22 Vaughn, J. E., Sims, T. and Nakashima, M., A comparison of the early development of axodendritic and axosomatic synapses upon embryonic mouse spinal motor neurons, J. comp. Neurol., 175 (1977) 79-100.