anatomical and electrotonic coupling in developing genioglossal motoneurons of the rat
TRANSCRIPT

Brain Research, 598 (1992) 127-137 127 © 1992 Elsevier Science Publishers B.V. All rights reserved 0006-8993/92/$05.00
BRES 18331
Anatomical and electrotonic coupling in developing genioglossal motoneurons of the rat
Emilio Mazza a Pedro A. Nfifiez-Abades a , . John M. Spielmann a and William E. Cameron a,b
Departments of a Behavioral Neuroscience and b Psychiatry University of Pittsburgh, Pittsburgh, PA (USA)
(Accepted 14 July 1992)
Key words: Motoneuron; Brainstem; Upper airway; Respiration; Electrical coupling; Dye-coupling; Postnatal development
Dye-, tracer- and electrotonic coupling were studied independently in genioglossal (GG) motoneurons using intracellular recordings in in vitro brainstem slices from rats postnatal ages 1-30 days. The subpopulation of GG motoneurons were retrogradely labeled after an injection of dextran-rhodamine into the posterior tongue. Dye-coupling was studied with Lucifer yellow injected into 55 motoneurons and tracer-coupling with neurobiotin injected into 89 presumptive GG motoneurons. Of the motoneurons injected with Lucifer yellow, only 6 of 41 cells (16.2%) exhibited dye-coupling; all occurred in animals less than 9 days old. In all but one instance, dye-coupling was restricted to only one other cell. No evidence of dye-coupling was found in the 14 cells injected in animals older than 8 days. Tracer-coupling (neurobiotin) was demonstrated in 12 of 30 cells (40%) from animals 1-2 days old and in 6 of 21 cells (28.6%) from animals 3-8 days old. Of the remaining 38 cells from animals 10 days of age and older, only one cell was found to be tracer-coupled. Cells injected with neurobiotin were coupled to an average of two other cells. Electrotonic coupling, as demonstrated with a short latency depolarization (SLD) in response to stimulation of hypoglossal axons, was found in developing GG motoneurons. These SLDs were revealed in 17 of 40 GG motoneurons (42.5%) examined in 1-8-day-old animals. There were no SLDs recorded in the 10 cells examined from animals of 10 days and older. The significance of coupling relative to patency of the newborn upper airways is discussed.
INTRODUCTION
In te rce l lu l a r communica t i ons via gap junc t ions have
an impor t an t funct ion dur ing the course of deve lop-
ment . A d i s rup t ion in this communica t i on dras t ica l ly
re ta rds d e v e l o p m e n t by compromis ing a pa thway for
nu t r i en t exchange and o the r small molecu les necessary
for growth t8. A p a r t f rom its b iochemica l role, gap
junc t ions have been shown to m e d i a t e e lec t ro ton ic
coupl ing be tween embryon ic cells 31, a p h e n o m e n o n
tha t pers is ts into pos tna ta l life. Wi th the except ion of
the h i p p o c a m p u s 25, the in fe r ior 01ivary nucleus 34, the
abducens nucleus t5 and the mesencepha l i c nucleus 5,
e lec t ro ton ic coupl ing is ra re ly seen in the adu l t mam-
mal; however , i t is p r eva l en t in the newborn . Pr io r to
es tabl i sh ing the full c o m p l e m e n t o f de scend ing pa th -
ways in the adul t , e l ec t ro ton ic coupl ing be tween mo-
t oneu rons may enhance synchroniza t ion of m o t o r activ- ity th rough shar ing of synapt ic input 4,13,42. This syn-
ch ron ized activity has been sugges ted to al low a par t ic-
u lar g roup of neu rons to funct ion as an ensemble z3.
The p re sence o f e l ec t ro ton ic coupl ing has been re-
p o r t e d for spinal m o t o n e u r o n s of the neona ta l rat 4't3'42,
the cat 15, and the frog 9'12'36'43. In the neona ta l rat and
the frog, it has been observed that e lec t ro tonica l ly
coup led m o t o n e u r o n s have a high degree of specifici ty
in that they pro jec t to the same muscle or funct ional ly
equiva lent (synergist ic) muscles 9'42'43. Ul t ras t ruc tu ra l
s tudies have sugges ted that the e lect r ica l in te rac t ion
be tw e e n m o t o n e u r o n s is m e d i a t e d via gap junc t iona l connec t ions 28'36'37. In t e rce l lu l a r communica t ion can
also be d e m o n s t r a t e d with dye-coupl ing be tween neu-
rons using the f luorescen t dye, Luci fer yel low 38 or with
t r ace r -coup l ing using neurob io t in 4°. Dye-coup l ing has
been shown to occur p r imar i ly be tween ceils known to be e lectr ical ly coup led 26,38. Brenowi tz et al. 7 have re-
p o r t e d concur ren t dye-coupl ing and e lec t ro ton ic cou-
p l ing in frog spinal mo toneu rons . Becke r and Navar-
Correspondence: W.E. Cameron, Department of Behavioral Neuroscience, 446 Crawford Hall, University of Pittsburgh, Pittsburgh, PA 15260, USA. Fax: (1) (412) 624 9198. * Permanent address: Laboratorio de Neurociencia, Dept. de Fisiologia y Biologia Animal, Universidad de Sevilla, Spain.

128
rete ~' have demonstrated that the presence of dye coupling is consistent with the transient expression of electrotonic coupling in developing rat lumbar moto- neurons ~.~.42.
Our interest in the coupling phenomenon stems from our desire to understand the factors that influ- ence the electrical properties of developing respiratory neurons. During respiration, the genioglossus muscle is activated just prior to the diaphragm and helps provide a patent upper airway 8. Dyscoordination between these two muscles may result in upper airway obstruction and a cessation of breathing 33. Another study showed that premature infants with mixed and obstructive ap- nea had a decrease in activation of the genioglossus muscle in response to an airway obstruction ~4. This inability to recruit the appropriate upper airway mus- cles during airway obstruction may play an important role in the genesis of apnea of prematurity.
In this study, we have asked the question does coupling exist between GG motoneurons of the new- born rat? We have provided evidence for dye-, tracer-, and electrotonic coupling in GG motoneurons during postnatal life.
MATERIALS AND METHODS
lntraceUular recording from motoneurons in the ventromedial hypoglossal nucleus were obtained using a brainstem slice prepara- tion from postnatal rats. Three separate protocols were used to determine the frequency of dye-, tracer- and electrotonic coupling in this motor nucleus.
Slice preparation Sprague-Dawley rats (wt. 6-115 g) were deeply anesthetized with
halothane, tracheotomized, and ventilated with 100% 0 2. The brain- stem slice preparation that we employed has been previously de- scribed by Aghajanian and Rasmussen 2. In brief, the animals were transcardially perfused with a cold (1-4°C) sucrose-artificial cerebral spinal fluid (ACSF) and then quickly decapitated. The brainstem was removed and transferred to a Vibraslice chamber filled with cold, oxygenated sucrose-ACSF and transversely sectioned at 300-400 p~m. The slice of brainstem studied was taken from approximately 1.0-1.5 mm caudal to the obex to the level of the inferior colliculi.
All slices were incubated in a holding chamber containing cold sucrose-ACSF for 35-45 rain and then transferred to a second holding chamber containing normal-ACSF at room temperature (21 _+ I°C). Individual slices were transferred to the recording chamber and superfused with normal ACSF. In order to counterbalance the increased CO 2 saturation at lower temperatures, the percentage of CO 2 in the gas bubbling the cold sucrose-ACSF was varied between 2 and 5% to maintain the pH between 7.3 and 7.4. At room temperatures, the normal ACSF was bubbled with 95% 0 2 / 5 % CO 2 (pH = 7.4).
The composition of sucrose-ACSF used during the perfusion, sectioning, and the initial incubation period was modified from Aghajanian and Rasmussen 2 and was composed of the following (in mM): 250 sucrose, 2.5 KCI, 1 NazHPO4, 25 NaHC O 3, 11 glucose, 2 MgSO 4, 2 CaC12. The normal-ACSF used for the subsequent por- tions of the experiment was composed of the following composition (in mM): 126 NaCI, 2.5 KCI, 1 Na2HPO 4, 25 NaHCO3, 11 glucose, 3 MgSO 4, 1 CaC12.
L;rperimental protocol Two to seven days prior to the intracellular experiments, animals
were anesthetized with ether and 3-5 ~1 of a 1(/% solution ol dextran-rhodamine (mol. wt. 10,000; Molecular Probes Inc.) was injected into the posterior aspects of their tongues. The purpose ol these injections made into the genioglossus muscle was not to label every GG motoneuron in the nucleus but to provide a fluorescent marker for positioning the intracellular electrode in a region of intense retrograde labeling. An epifluorescence at tachment to an upright microscope permitted us to visualize these neurons in the slice within the recording chamber. Larger injections ( > l0/~1) were found to label the dorsally located motoneurons innervating the intrinsic muscles of the tongue. Because there is a strict somatotopic organization within the hypoglossal nucleus 21, GG motoneurons could be identified by either the presence of retrograde label or their position within the labeled ventromedial compar tment of the hy- poglossal nucleus. The criteria for a successful impalement included: (i) a stable resting membrane potential more negative than - 5 4 mV; (ii) a spike height greater than 60 mV; and (iii) an action potential with positive overshoot. Dye- and tracer-coupling. Dye- and tracer-coupling were examined in GG motoneurons using intracellular injection of Lucifer yellow 3s or neurobiotin 2°, respectively. For the dye-coupling experiments, the tip of a glass electrode was filled with 10% Lucifer yellow CH (Sigma) in distilled H 2 0 and then back-filled with 1 M LiCI TM (resistances: 90-120 Mg2). Lucifer yellow was iontophoresed into the motoneuron by passing a constant hyperpolarizing current (0.3-0.6 nA) on which hyperpolarizing current pulses (0.1-0.4 nA) for 50 ms at 10 Hz was superimposed. For the tracer coupling experiments, the tip of a glass microelectrode was filled with 2% neurobiotin (Vector Laboratories) in 1 M methyl potassium sulfate 2° and backfilled with 1 M methyl potassium sulfate (resistances: 140-180 M,O). Neurobiotin was ion- tophoresed into the motoneuron by passing a depolarizing current pulse (0.2-1.0 hA) for a duration of 150 ms at 3.3 Hz. Care was taken to impale only a single cell per slice. The duration of ion- tophoretic injection ranged from 10-60 rain.
After intracellular injection of Lucifer yellow, the slice was re- turned to the holding chamber containing normal-ACSF at room temperature for 1-2 h. The slice was then fixed in 4% paraformalde- hyde and stored at 4°C for a period longer than 24 h. Following the fixation period, these slices were placed on a glass slide and were allowed to air dry. Finally, the slices were cleared with xylene and mounted with DPX (Fluka). Sections were viewed with epifluores- cence using a green filter (rhodamine) or a blue filter (Lucifer yellow). The criterion for dye-coupling with Lucifer yellow was confluence of this fluorescent label among motoneurons.
After intracellular injection of neurobiotin, the slices were like- wise incubated in normal-ACSF for several hours and fixed for 1-2 days in 4% paraformaldehyde at 4°C. Slices were transferred to 30% sucrose in phosphate buffer (pH = 7.4) at 4°C and stored overnight. Then, slices were pretreated with 1% Triton X-100 for 2 -3 h, and incubated with a standard ABC solution (Vector Laboratories) di- luted (1:500) in phosphate buffer for 3 h at room temperature. After rinsing, the horseradish peroxidase was reacted with nickel and diaminobenzidine (DAB) 1. Finally, slices were mounted on gela- tinized slides, dehydrated with serial alcohols, cleared with xylene and coverslipped with DPX (Fluka). Due to the density of the DAB reaction product, we did not at tempt to co-localize rhodamine in the cells labeled by neurobiotin injections. Electrotonic coupling. Glass microelectrodes were filled with 3 M potassium acetate (resistances: 70-90 M J2). Three signals were recorded for all cells: current, low- and high-gain DC-coupled signal of the membrane potential. A brief current pulse (13-19 hA; 100-200 ~s at 1 Hz) was employed to elicit an orthodromic action potential. Motoneurons were antidromically activated by a twisted pair nichrome electrode placed over the axon tract adjacent to the rootlet. This stimulating electrode was selected for those motoneu- rons with axons that remained within the plane of the slice and were relatively close to the surface. On average, 5-10 cells were impaled in order to find one that could be antidromically activated. An- tidromic action potentials were elicited by 1-10 mA current pulses (100 /zs at 10 Hz). In order to visualize small changes in membrane
129
potential, a digital sample-and-hold circuit was used to subtract the resting potential before amplification. This amplified signal consti- tuted the high-gain DC trace used in most of the analyses presented.
A variety of methods have been used to demonstrate electrotonic coupling. Two methods were employed in the present study. First, a subthreshold stimulation of the distal axons was used to activate
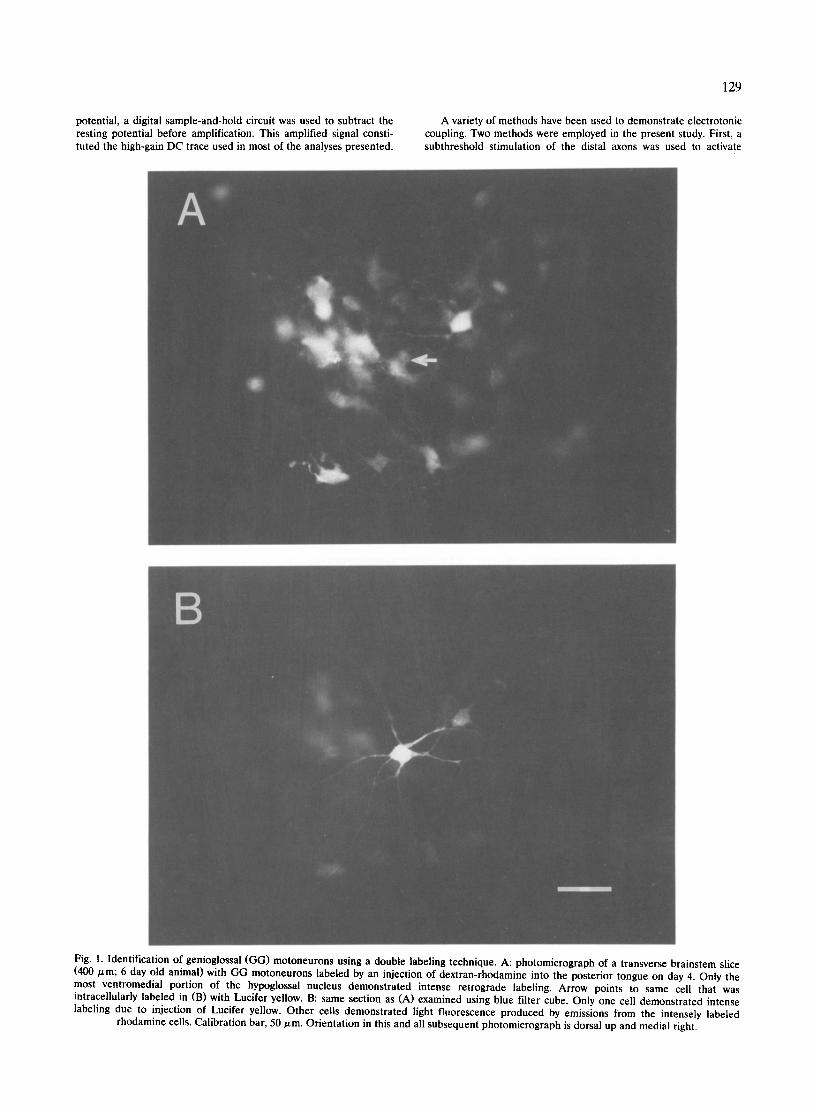
Fig. 1. Identification of genioglossal (GG) motoneurons using a double labeling technique. A: photomicrograph of a transverse brainstem slice (400 p,m; 6 day old animal) with GG motoneurons labeled by an injection of dextran-rhodamine into the posterior tongue on day 4. Only the most ventromedial portion of the hypoglossal nucleus demonstrated intense retrograde labeling. Arrow points to same cell that was intracellularly labeled in (B) with Lucifer yellow. B: same section as (A) examined using blue filter cube. Only one cell demonstrated intense labeling due to injection of Lucifer yellow. Other cells demonstrated light fluorescence produced by emissions from the intensely labeled
rhodamine cells. Calibration bar, 50 p~m. Orientation in this and all subsequent photomicrograph is dorsal up and medial right.
13(t
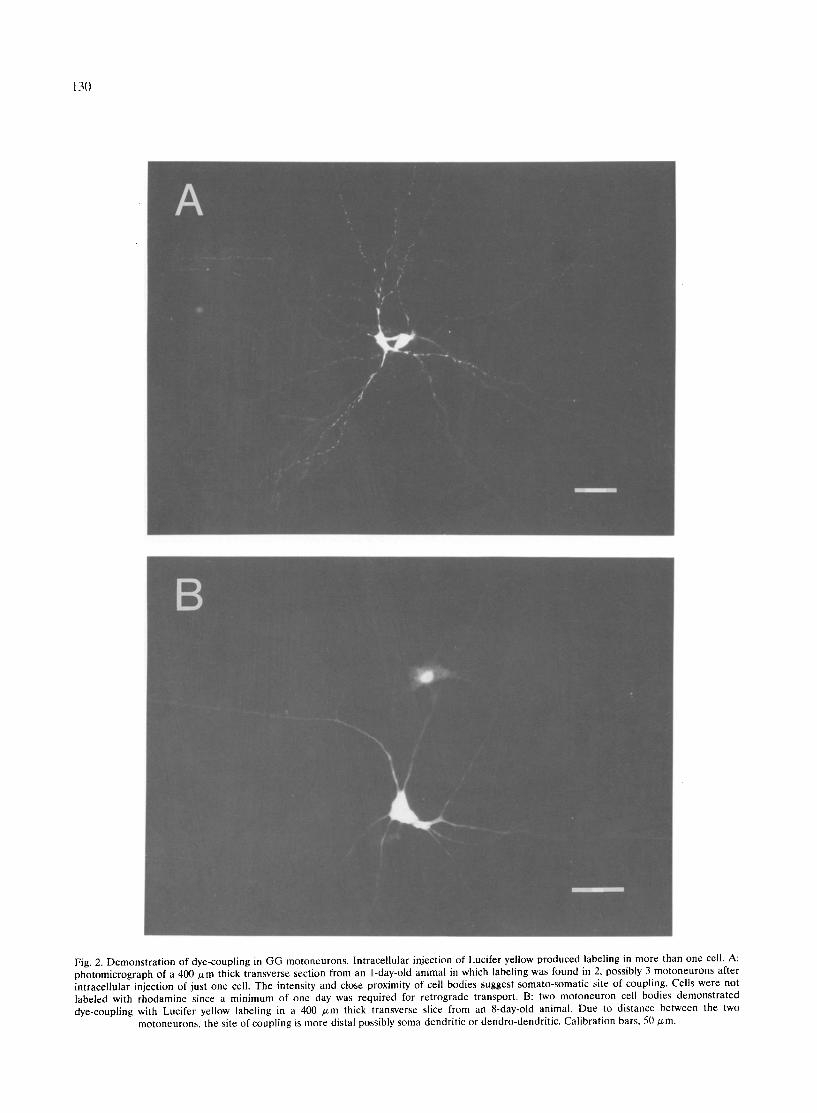
Fig. 2. Demonstrat ion of dye-coupling in GG motoneurons. Intracellular injection of Lucifer yellow produced labeling in more than one cell. A: photomicrograph of a 400 /zm thick transverse section from an 1-day-old animal in which labeling was found in 2, possibly 3 motoneurons after intracellular injection of just one cell. The intensity and close proximity of cell bodies suggest somato-somatic site of coupling. Cells were not labeled with rhodamine since a minimum of one day was required for retrograde transport. B: two motoneuron cell bodies demonstrated dye-coupling with Lucifer yellow labeling in a 400 /xm thick transverse slice from an 8-day-old animal. Due to distance between the two
motoneurons, the site of coupling is more distal possibly soma-dendritic or dendro-dendritic. Calibration bars, 50/zm.
lower threshold axons. The high-gain signal of the membrane poten- tial was averaged, triggered by the stimulus to the axons. In some instances, this stimulation revealed a depolarization that occurred within 5 ms following the antidromic stimulation. To exclude the possibility that this SLD was synaptic in nature, the SLD was measured at a series of membrane potentials, both hyperpolarized and depolarized relative to the resting membrane potential.
If no SLD was revealed by subthreshold antidromic stimulation, then the response to increasing strengths of suprathreshold stimula- tion (for action potential generation) was examined. In some cases, the SLD appeared as a small excursion on the repolarizing phase of the antidromic action potential. To eliminate the contribution of the antidromic potential, an orthodromic action potential was elicited prior to the antidromic stimulation in order to collide and cancel the antidromic invasion of the cell body 5. In this way, many motoneu- rons could be activated by suprathreshold stimulation and their contribution to the depolarization of the cell under study could be visualized independent of the antidromic action potential.
Because this SLD occurred during the falling phase of the orthodromic action potential, the SLD profile was extracted by subtracting the waveforms captured with and without the antidromic stimulation. The SLD amplitude and time course were measured after being acquired by an analog-to-digital board (RC Electronics) in a personal computer. The SLD amplitude was measured from resting membrane potential to peak voltage and the rate of rise determined as the slope of the line between 10% and 90% of maximum amplitude. The SLD latency was measured from both the antidromic stimulus artifact and the onset on the antidromic action potential.
Depolarizing potentials resulting from motor axon stimulation were identified as electrotonic in nature if they fulfilled the following criteria: (i) had a short latency ( < 5 ms with respect to stimulus artifact); (ii) were resistant to collision with an orthodromic action potential; and (iii) were insensitive to changes in resting membrane potential 5,13,43. Statistical analysis. The differences between the frequency of occur- rence of dye-, tracer- and electrotonic coupling were tested using a 2 x 2 table and calculating the Chi-square statistic. Differences were considered significant if P < 0.5.
R E S U L T S
Al l m o t o n e u r o n s r e c o r d e d w e r e l o c a t e d in t h e v e n -
t r o m e d i a l p o r t i o n o f t h e h y p o g l o s s a l n u c l e u s a n d m a n y
e x h i b i t e d r e t r o g r a d e r h o d a m i n e l a b e l (F ig . 1A) . Al l
i m p a l e m e n t s t a r g e t e d t h i s a r e a o f i n t e n s e l a b e l i n g .
D e f i n i t i v e i d e n t i f i c a t i o n as a G G m o t o n e u r o n r e q u i r e d
t h a t t h e ce l l b e l a b e l e d w i t h b o t h d e x t r a n - r h o d a m i n e
a n d L u c i f e r y e l l o w (Fig . 1B). F i f ty - f ive m o t o n e u r o n s
w e r e i o n t o p h o r e s e d w i t h L u c i f e r y e l l o w of w h i c h 15
( 2 7 % ) c o u l d b e d e f i n i t i v e l y i d e n t i f i e d as G G by t h e
p r e s e n c e o f d o u b l e l abe l . T h e b a l a n c e o f ce l l s w e r e
i d e n t i f i e d as m o t o n e u r o n s by t h e p r e s e n c e o f a n a x o n
t h a t c o u l d b e t r a c e d to exi t a t t h e v e n t r a l m e d u l l a r y
s u r f a c e . M o s t m o t o n e u r o n s l a b e l e d w i t h L u c i f e r ye l low
w e r e w i t h i n t h e a r e a o f r e t r o g r a d e l y l a b e l e d n e u r o n s .
Characteristics of dye-coupling Fi f ty - f ive p r e s u m p t i v e G G m o t o n e u r o n s w e r e la-
b e l e d w i t h L u c i f e r y e l l o w in 30 a n i m a l s f r o m 1 - 2 4 days
p o s t n a t a l . O f t h e m o t o n e u r o n s i n j e c t e d , six s h o w e d
e v i d e n c e o f d y e - c o u p l i n g ( 1 1 % ) . T h e s e d y e - c o u p l e d
m o t o n e u r o n s w e r e f o u n d in 41 m o t o n e u r o n s t a k e n
131
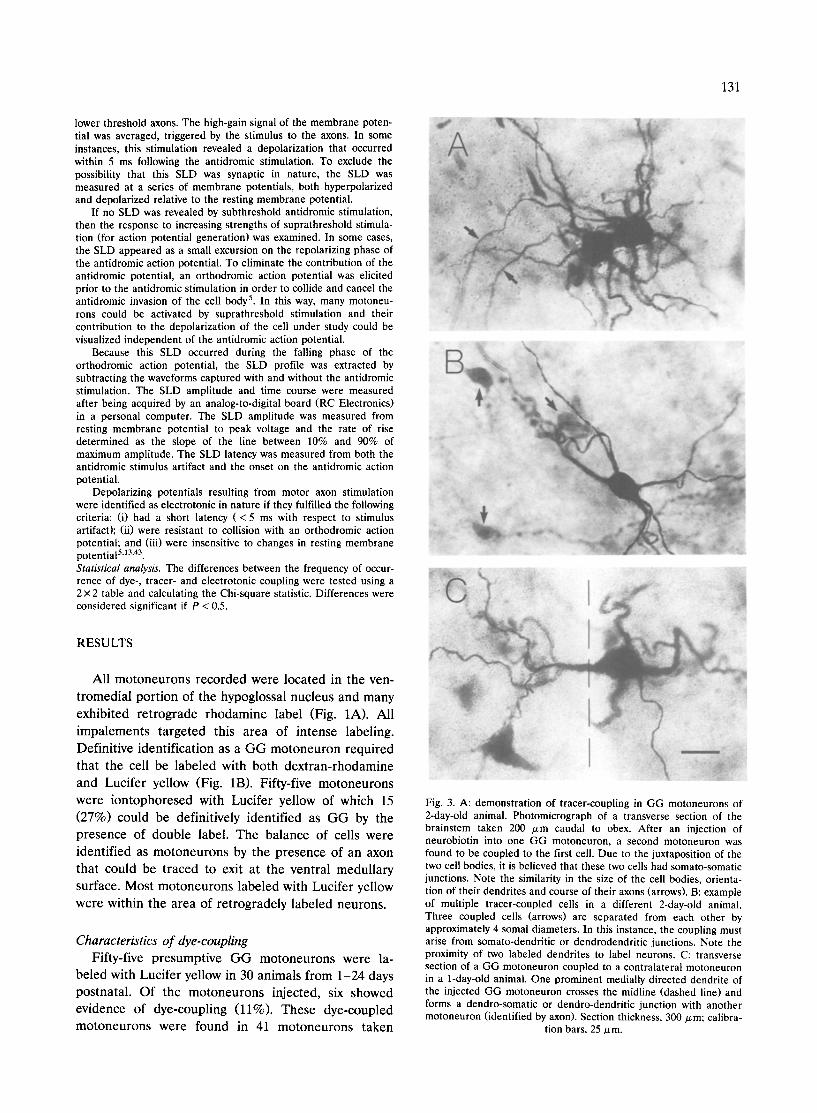
Fig. 3. A: demonstration of tracer-coupling in GG motoneurons of 2-day-old animal. Photomicrograph of a transverse section of the brainstem taken 200 /xm caudal to obex. After an injection of neurobiotin into one GG motoneuron, a second motoneuron was found to be coupled to the first cell. Due to the juxtaposition of the two cell bodies, it is believed that these two cells had somato-somatic junctions. Note the similarity in the size of the cell bodies, orienta- tion of their dendrites and course of their axons (arrows). B: example of multiple tracer-coupled ceils in a different 2-day-old animal. Three coupled cells (arrows) are separated from each other by approximately 4 somal diameters. In this instance, the coupling must arise from somato-dendritic or dendrodendritic junctions. Note the proximity of two labeled dendrites to label neurons. C: transverse section of a GG motoneuron coupled to a contralateral motoneuron in a 1-day-old animal. One prominent medially directed dendrite of the injected GG motoneuron crosses the midline (dashed line) and forms a dendro-somatic or dendro-dendritic junction with another motoneuron (identified by axon). Section thickness, 300 ~m; calibra-
tion bars, 25/xm.
132
from the 1-8-day-old animals (15%). No dye-coupling was seen in the 14 cells studied in animals 10 days or older. Fig. 2A demonstrates dye-coupling in a 1-day-old animal. In this photomicrograph, two and possibly three adjacent dye-coupled motoneurons of the ventromedial hypoglossal nucleus are shown. Two of the motoneu- rons are intensely labeled with a third motoneuron having a less intense labeling. Cells were not labeled with dextran-rhodamine in this 1 day-old animal be- cause retrograde labeling required at least 1-2 days post-injection survival period. In Fig. 2B, dye-coupling is demonstrated between two motoneurons separated by a larger intersomal distance (c.f. Fig 2A). It is believed that the brightly labeled (injected) cell is cou- pled to the less intensely labeled (non-injected) cell. Although these motoneurons were not retrogradely labeled, they were presumed to be GG motoneurons based on their ventromedial location within the hy- poglossal nucleus.
With one exception, the spread of Lucifer yellow was restricted to only one other cell. The apparent site of coupling differed among the cells in which dye-cou- pling was demonstrated. In Fig. 2A, the site of dye- coupling appears to be somato-somatic based on the close apposition of the cell bodies. In Fig. 2B, the coupled cell bodies are at a distance from one another suggesting that the site of coupling could be dendro- dendritic a n d / o r dendro-somatic.
Characteristics of tracer-coupling Eighty-nine presumptive GG motoneurons were la-
beled with neurobiotin in 41 animals ages 1-30 days old. Of the injected motoneurons, nineteen showed evidence of tracer-coupling (21%). Twelve of 30 cells exhibiting tracer-coupling were from 1-2-day-old ani- mals (40%). Six of 21 coupled cells were from animals 3-8 days old (29%) and only 1 of 38 cells (3%) was found to be coupled in animals 10 days and older.
In contrast to dye-coupling, cells injected with neu- robiotin exhibited coupling to an average of 2.2 + 0.7 cells (_+S.D.; range: 1-6) at 1-2 days old and an average of 1.8 _+ 0.2 cells (range: 1-4) in animals 3 -8 days old. One coupled cell, in the oldest age group, was coupled with two other cells. The tracer-coupled neu- rons exhibited preferentially somato-somatic or so- mato-proximal dendritic coupling (9 of 12 coupled cells at 1-2 days; 4 of 6 coupled cells at 3 -8 days). Fig. 3A depicts the somato-somatic coupling between two cells in a two-day old animal. However, it was also possible to find coupling between cells that were separated by greater than 4-5 somal diameters. Fig. 3B shows tracer-coupling between the injected cell and three other motoneurons that are spatially separated. One
I
i
2
1 mV
1
5 ms
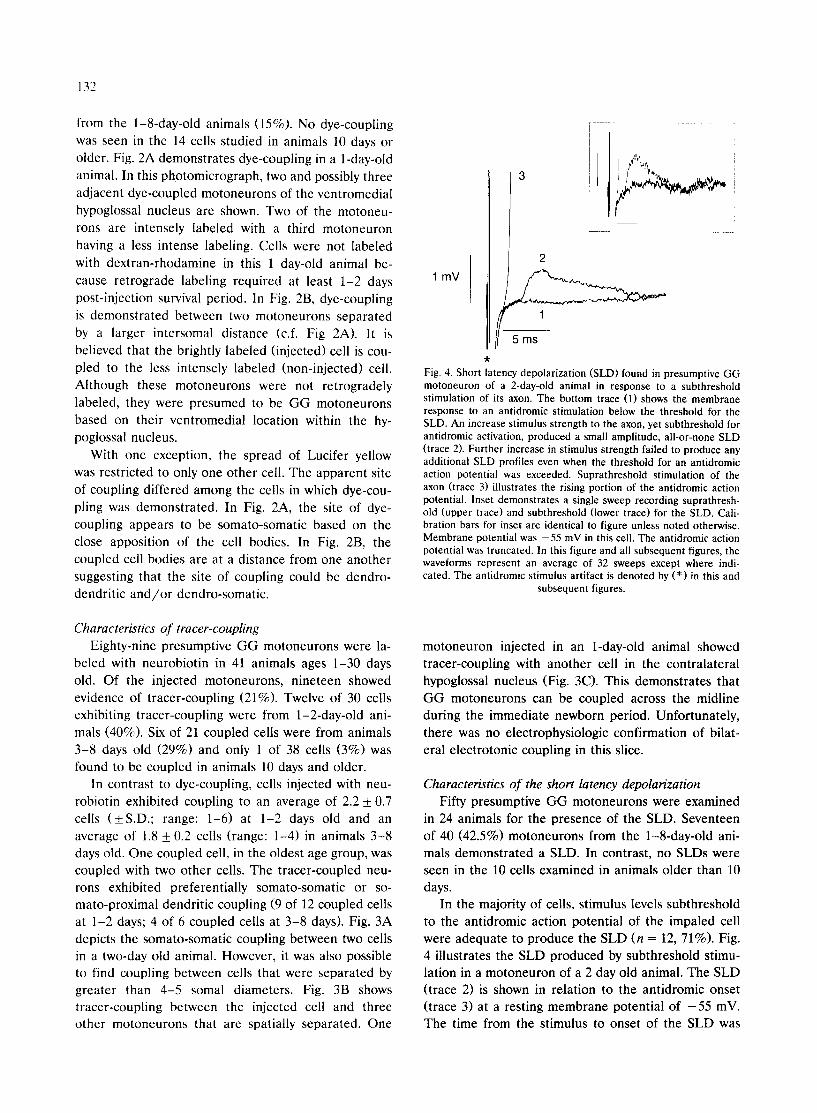
Fig. 4. Short latency depolarization (SLD) found in presumptive GG motoneuron of a 2-day-old animal in response to a subthreshold stimulation of its axon. The bottom trace (1) shows the membrane response to an antidromic stimulation below the threshold for the SLD. An increase st imulus strength to the axon, yet subthreshold for antidromic activation, produced a small amplitude, all-or-none SLD (trace 2). Further increase in st imulus strength failed to produce any additional SLD profiles even when the threshold for an antidromic action potential was exceeded. Suprathreshotd stimulation of the axon (trace 3) illustrates the rising portion of the antidromic action potential. Inset demonstra tes a single sweep recording suprathresh- old (upper trace) and subthreshold (lower trace) for the SLD. Cali- bration bars for inset are identical to figure unless noted otherwise. Membrane potential was - 55 mV in this cell. The antidromic action potential was truncated. In this figure and all subsequent figures, the waveforms represent an average of 32 sweeps except where indi- cated. The antidromic stimulus artifact is denoted by (*) in this and
subsequent figures.
motoneuron injected in an 1-day-old animal showed tracer-coupling with another cell in the contralateral hypoglossal nucleus (Fig. 3C). This demonstrates that GG motoneurons can be coupled across the midline during the immediate newborn period. Unfortunately, there was no electrophysiologic confirmation of bilat- eral electrotonic coupling in this slice.
Characteristics of the short latency depolarization Fifty presumptive G G motoneurons were examined
in 24 animals for the presence of the SLD. Seventeen of 40 (42.5%) motoneurons from the 1-8-day-old ani- mals demonstrated a SLD. In contrast, no SLDs were seen in the 10 ceils examined in animals older than 10 days.
In the majority of cells, stimulus levels subthreshold to the antidromic action potential of the impaled cell were adequate to produce the SLD (n = 12, 71%). Fig. 4 illustrates the SLD produced by subthreshold stimu- lation in a motoneuron of a 2 day old animal. The SLD (trace 2) is shown in relation to the antidromic onset (trace 3) at a resting membrane potential of - 5 5 mV. The time from the stimulus to onset of the SLD was
133
3.4 ms and from the antidromic onset to SLD was
approximately 2 ms. In some cells, the threshold for producing the SLD
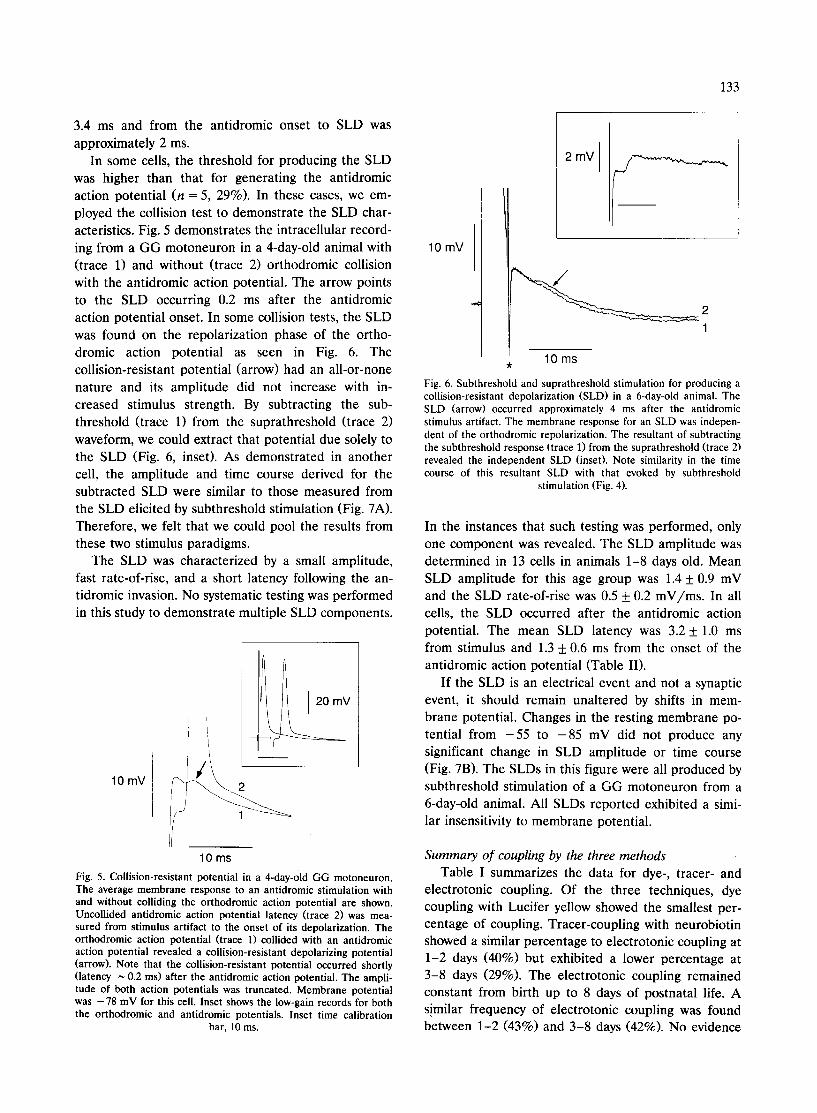
was higher than that for generating the antidromic action potential (n = 5, 29%). In these cases, we em- ployed the collision test to demonstrate the SLD char- acteristics. Fig. 5 demonstrates the intracellular record- ing from a GG motoneuron in a 4-day-old animal with (trace 1) and without (trace 2) orthodromic collision with the antidromic action potential. The arrow points to the SLD occurring 0.2 ms after the antidromic action potential onset. In some collision tests, the SLD was found on the repolarization phase of the ortho- dromic action potential as seen in Fig. 6. The collision-resistant potential (arrow) had an all-or-none nature and its amplitude did not increase with in- creased stimulus strength. By subtracting the sub- threshold (trace 1) from the suprathreshold (trace 2) waveform, we could extract that potential due solely to the SLD (Fig. 6, inset). As demonstrated in another cell, the amplitude and time course derived for the subtracted SLD were similar to those measured from the SLD elicited by subthreshold stimulation (Fig. 7A). Therefore, we felt that we could pool the results from these two stimulus paradigms.
The SLD was characterized by a small amplitude, fast rate-of-rise, and a short latency following the an- tidromic invasion. No systematic testing was performed in this study to demonstrate multiple SLD components.
10 mV 2
10 m s
Fig. 5. Collision-resistant potential in a 4-day-old GG motoneuron. The average membrane response to an antidromic stimulation with and without colliding the orthodromic action potential are shown. Uncollided antidromic action potential latency (trace 2) was mea- sured from stimulus artifact to the onset of its depolarization. The orthodromic action potential (trace 1) collided with an antidromic action potential revealed a collision-resistant depolarizing potential (arrow). Note that the collision-resistant potential occurred shortly (latency ~ 0.2 ms) after the antidromic action potential. The ampli- tude of both action potentials was truncated. Membrane potential was - 7 8 mV for this cell. Inset shows the low-gain records for both the orthodromic and antidromic potentials. Inset time calibration
bar, 10 ms.
2 mV
10 mV
10 ms y¢
Fig. 6. Subthreshold and suprathreshold stimulation for producing a collision-resistant depolarization (SLD) in a 6-day-old animal. The SLD (arrow) occurred approximately 4 ms after the antidromic stimulus artifact. The membrane response for an SLD was indepen- dent of the orthodromic repolarization. The resultant of subtracting the subthreshold response (trace 1) from the suprathreshold (trace 2) revealed the independent SLD (inset). Note similarity in the time course of this resultant SLD with that evoked by subthreshold
stimulation (Fig. 4).
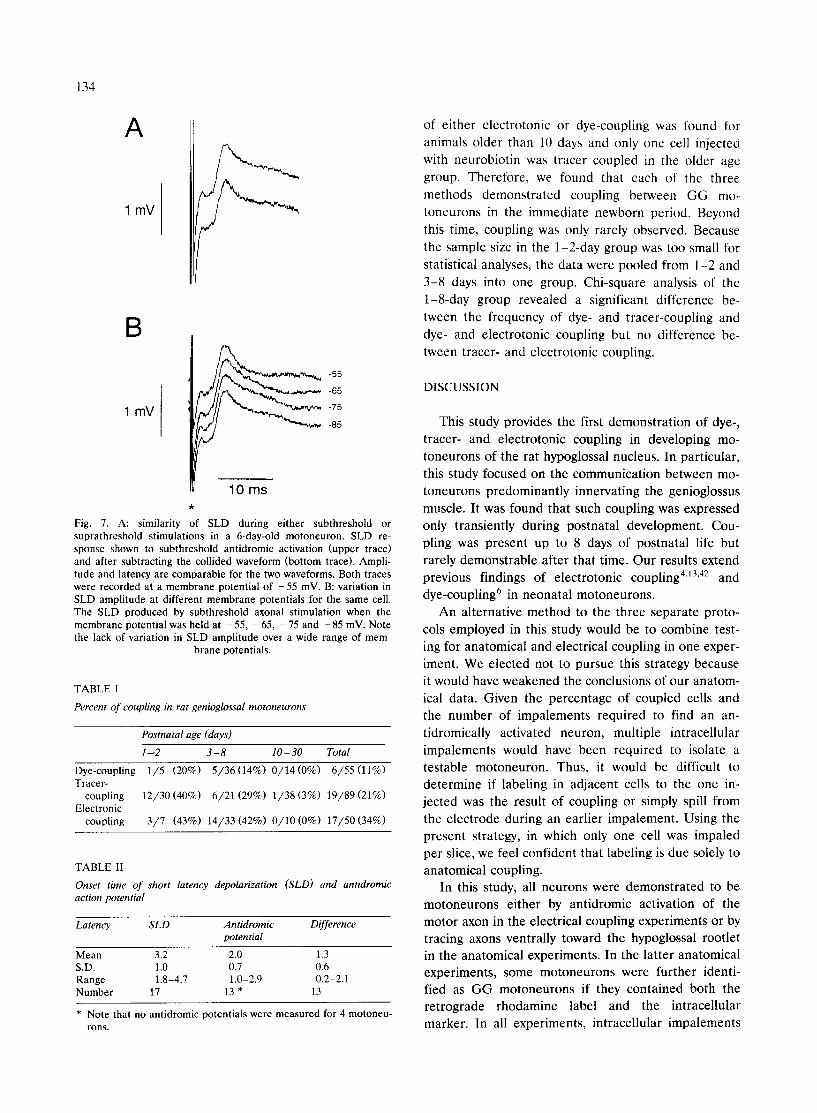
In the instances that such testing was performed, only one component was revealed. The SLD amplitude was determined in 13 cells in animals 1-8 days old. Mean SLD amplitude for this age group was 1.4 + 0.9 mV and the SLD rate-of-rise was 0.5 + 0.2 mV/ms. In all cells, the SLD occurred after the antidromic action potential. The mean SLD latency was 3.2 + 1.0 ms from stimulus and 1.3 + 0.6 ms from the onset of the antidromic action potential (Table II).
If the SLD is an electrical event and not a synaptic event, it should remain unaltered by shifts in mem- brane potential. Changes in the resting membrane po- tential from - 5 5 to - 8 5 mV did not produce any significant change in SLD amplitude or time course (Fig. 7B). The SLDs in this figure were all produced by subthreshold stimulation of a GG motoneuron from a 6-day-old animal. All SLDs reported exhibited a simi- lar insensitivity to membrane potential.
Summary of coupling by the three methods Table I summarizes the data for dye-, tracer- and
electrotonic coupling. Of the three techniques, dye coupling with Lucifer yellow showed the smallest per- centage of coupling. Tracer-coupling with neurobiotin showed a similar percentage to electrotonic coupling at 1-2 days (40%) but exhibited a lower percentage at 3-8 days (29%). The electrotonic coupling remained constant from birth up to 8 days of postnatal life. A similar frequency of electrotonic coupling was found between 1-2 (43%) and 3-8 days (42%). No evidence
134
A
1 mV
B
1 mV
-55
-65
-75
-85
10 ms
Fig. 7. A: similarity of SLD during either subthreshold or suprathreshold stimulations in a 6-day-old motoneuron. SLD re- sponse shown to subthreshold antidromic activation (upper trace) and after subtracting the collided waveform (bottom trace). Ampli- tude and latency are comparable for the two waveforms. Both traces were recorded at a membrane potential of - 5 5 mV. B: variation in SLD amplitude at different membrane potentials for the same cell. The SLD produced by subthreshold axonal stimulation when the membrane potential was held at - 55, - 65, - 75 and - 85 mV. Note the lack of variation in SLD amplitude over a wide range of mem-
brane potentials.
TABLE I
Percent o f coupling in rat genioglossal motoneurons
Postnatal age (days)
1 -.2 3 - 8 10 - 30 Total
Dye-coupling 1 /5 (20%) 5 / 36 (14%) 0 / 1 4 (0%) 6 /55 (11%) Tracer-
coupling 12/30 (40%) 6 /21 (29%) 1/38 (3%) 19/89 (21%) Electronic
coupling 3 / 7 (43%) 14/33 (42%) 0 / 1 0 (0%) 17/50 (34%)
TABLE II
Onset time of short latency depolarization (SLD) and antidromic action potential
Latency S L D Antidromic Difference potential
Mean 3.2 2.0 1.3 S.D. 1.0 0.7 0.6 Range 1.8-4.7 1.0-2.9 0.2-2.1 Number 17 13 * 13
• Note that no antidromic potentials were measured for 4 motoneu- rons.
of either electrotonic or dye-coupling was found for animals older than 10 days and only one cell injecte0 with neurobiotin was tracer coupled in the older age group. Therefore, we found that each of the three methods demonstrated coupling between GG mo- toneurons in the immediate newborn period. Beyond this time, coupling was only rarely observed. Because the sample size in the 1-2-day group was too small for statistical analyses, the data were pooled from 1-2 and 3-8 days into one group. Chi-square analysis of the 1-8-day group revealed a significant difference be- tween the frequency of dye- and tracer-coupling and dye- and electrotonic coupling but no difference be- tween tracer- and electrotonic coupling.
DISCUSSION
This study provides the first demonstration of dye-, tracer- and electrotonic coupling in developing mo- toneurons of the rat hypoglossal nucleus. In particular, this study focused on the communication between mo- toneurons predominantly innervating the genioglossus muscle. It was found that such coupling was expressed only transiently during postnatal development. Cou- pling was present up to 8 days of postnatal life but rarely demonstrable after that time. Our results extend previous findings of electrotonic coupling 4J3"42 and
dye-coupling 6 in neonatal motoneurons. An alternative method to the three separate proto-
cols employed in this study would be to combine test- ing for anatomical and electrical coupling in one exper- iment. We elected not to pursue this strategy because it would have weakened the conclusions of our anatom- ical data. Given the percentage of coupled cells and the number of impalements required to find an an- tidromically activated neuron, multiple intracellular impalements would have been required to isolate a testable motoneuron. Thus, it would be difficult to determine if labeling in adjacent cells to the one in- jected was the result of coupling or simply spill from the electrode during an earlier impalement. Using the present strategy, in which only one cell was impaled per slice, we feel confident that labeling is due solely to anatomical coupling.
In this study, all neurons were demonstrated to be motoneurons either by antidromic activation of the motor axon in the electrical coupling experiments or by tracing axons ventrally toward the hypoglossal rootlet in the anatomical experiments. In the latter anatomical experiments, some motoneurons were further identi- fied as GG motoneurons if they contained both the retrograde rhodamine label and the intracellular marker. In all experiments, intraceltular impalements

135
were made in the ventromedial hypoglossal nucleus, the region of intense retrograde labeling. This region of the nucleus has been shown to contain almost exclu- sively GG motoneurons in previous studies in the rat 21'39. It is possible that this region of the hypoglossal
nucleus might also contain a few motoneurons inner- vating another extrinsic tongue muscle, the geniohy- oideus. Because the geniohyoid motoneurons are coac- tivated with GG motoneurons to protrude the tongue and dilate the upper airways (functional synergist), it was felt that their inclusion in the sampling would not detract from the goal of understanding control of the
upper airways. In our results, Lucifer yellow showed a 15% dye-
coupling in animals 1-8 days old while electrotonic coupling was approximately 43%. The disparity be- tween the frequency of electrotonic and dye-coupling has also been demonstrated in motoneurons of the rat lumbar spinal cord 6'42 and in motoneurons innervating the frog triceps muscles 32'43. In the rat, 77% of the
lumbar motoneurons demonstrated a SLD from birth to 3 days old while only 18% of motoneurons showed the presence of dye-coupling. Between the ages of 4 and 7 days, the SLD was present in 68% of the motoneurons whereas dye-coupling was only present in 3% of the motoneurons. In the frog, 71% of the triceps motoneurons demonstrated SLD while only 5% showed dye-coupling.
In the newborn rat, dye-coupling was found at a significantly lower frequency in the first 8 days of life than tracer-coupling. Even more interestingly, the per- centage o f tracer-coupled motoneurons in animals 1-8 days old (35%) more accurately reflects the percentage of electrotonic coupling (43%) found during that same period than dye-coupling. Neurobiotin has been shown to provide a better estimate of the electrical coupling between some retinal neurons than Lucifer yellow. Vaney 4° proposed that the difference in passage of the larger Lucifer yellow molecule as compared to neurobi- otin in type B horizontal cells could be due to channel size or permeability. Alternatively, a reduction in the number of channels could restrict the amount of Lu- cifer yellow that would diffuse into an adjacent cell making visualization of the dye difficult. The present study provides the first comparison between the fre- quency of tracer- and electrotonic coupling in a de- fined population of neurons.
Using Lucifer yellow, there was only one instance in which coupling was found with more than one other cell. In contrast, neurobiotin injected motoneurons were tracer-coupled to an average of two other neu- rons in the hypoglossal nucleus. During the newborn period, the SLDs of GG motoneurons were comprised
of single, all-or-none components while lumbar mo-
toneurons exhibit multiple SLD components during this same period 42. The multiple component SLDs in these lumbar motoneurons indicate that more than two cells were electrically coupled. Like the percent of cells with SLDs, the number of multiple component SLD in lumbar motoneurons decreases with age. Although we did not test systematically for multiple components of the SLD using graded stimuli, in those cases in which we did, we found no evidence for electrotonic coupling
between more than one cell. There are several possible explanations for these
observations. First, neurobiotin may show tracer path- ways that are not available to ionic exchange. The agreement between frequency of tracer- and electro- tonic coupling would tend to argue against that possi- bility. Second, the cells that were electrically coupled to the injected cell may have had axons with similar thresholds to stimulation. In this case, we would be unable to discriminate based on graded stimulus, if the SLD was the product of one or more cells. Whereas the hemisected cord used by Walton and Navarrete 42
had axons that were all contained in the section, the slice preparation used in the present study may have potentially reduced the number of electrically coupled cells detected because some of the axons may not have been located within the plane of the section. Finally, there may have been more examples of complex SLDs than we were able to uncover using our stimulus proto- cols. It is difficult to discern between the last three possibilities and further study is required.
There was one case of tracer coupling between GG motoneurons observed in a 21-day-old animal. Both dye- and electrotonic coupling data suggest that there is no coupling beyond 8 days of postnatal life. How- ever, it should be noted that the total sample in these two experimental series (n = 24) is less than the sample size for the tracer-coupling experiments. Based on a 3% occurrence of tracer coupling, it feasible that our limited electrophysiological sample of 10 adult mo- toneurons could have missed the few electrotonically coupled adult cells. Therefore, it is possible that cou- pling is a less frequent occurrence in older animals but still a persistent feature of the adult hypoglossal nu- cleus. It is doubtful that coupling between so few motoneurons has a functional role in the controlling the adult tongue and may simply represent a vestige of the developmental process.
The decrease in SLD frequency might be used as an indicator of the stage of maturation. In lumbar mo- toneurons, Walton and Navarrete 42 described a contin- ual decrease in the percent of cells expressing a SLD. Thirty-one percent of lumbar motoneurons exhibited

136
SLDs between 8-13 days while no G G motoneurons were found with an SLD during that period. This
difference may reflect the less mature stage of the
lumbar motoneurons compared to a brainstem popula- tion. These differences in the SLD frequency support
the hypothesis of a rostrocaudal sequence of develop- ment originally described by Kingsbury 19 and later
corroborated by the cell birth data of Nornes and Das 3° and Altman and Bayer 3.
The SLDs of G G motoneurons were always found to follow the onset of the antidromic action potential
with a mean latency of 1.3 ms. This temporal relation- ship indicates that the ceils coupled to the injected
cells have axons that conduct more slowly. If axonal conduction velocity is correlated with cell body size as described for developing spinal motoneurons I J, then it
might be proposed that these slowly conducting cells
are also small cells and that they are preferentially coupled to larger cells. Since there are few if any
muscle afferents, and therefore, no gamma-motor ax- ons, in the hypoglossal nerve 17'24, we can conclude that
this is coupling between adjacent a-motoneurons. However, our figures do not demonstrate a large dif-
ference in cell size between the dye-or tracer-coupled neurons. Alternatively, the ceils with slower conducting axons may simply be less mature with respect to the myelination of their axons 42, and therefore, the less
mature motoneurons may be coupled to the faster
conducting, more mature motoneurons. The mean SLD amplitude of G G motoneurons was
1.38 mV. This value was more comparable to the 1.31 mV value found immediately after birth for lumbar motoneurons 42 than the 1.10 mV and 0.56 mV ampli-
tudes found for 4 -7 and 8-13 days, respectively. The mean SLD rate-of-rise for G G motoneurons was 0.49
m V / m s and this value was intermediate to the values reported for lumbar motoneurons at 4 -7 days (0.64 mV) and 8-13 days (0.36 mV). Based on the quantita- tive parameters of amplitude and rate-of-rise for SLD
waveforms, electrotonic coupling in these brainstem motoneurons appear to be comparable in nature to those reported in lumbar motoneurons. However, there is a temporal difference in the expression of coupling in the two motor pools. Coupling of these brainstem motoneurons was found to disappear earlier in postna- tal development than it did in the lumbar motoneu- rons. This implies that the first week of postnatal life represents the final stage of uncoupling process in brainstem motoneurons and may explain, in part, for the lack of multiple components of the SLD.
A decrease in electrotonic coupling during postnatal development is a common feature of the CNS having been reported in lumbar spinal motoneurons 42 and the
neocortex m. Loss of coupling between motoneurons after the first week of development is paralleled by the
disappearance of gap junctions between developing muscle fibers 35. This correlation suggests that muscle
fibers match the functional requirements of the mo-
toneuron that supplies them, and that with further development, differentiation and the refinement of
motoneuron activity goes hand in hand with the devel- opment of muscle fiber properties 29'4~.
A major role of electrotonic coupling is in the syn- chronization of neuronal firing. This function has been
demonstrated in the adult hippocampus where there is a low threshold to an epileptogenic discharge initiated in the cortex 25. Llinfis and Sasaki 22 demonstrated syn-
chronous neuronal firing in the inferior olive via elec- trotonic coupling. In the frog lumbar motoneurons, it is
suggested that electrotonic coupling provides a means for bilateral synchronization of muscle groups involved in activities such as swimming and jumping 12. Based on
our example of tracer-coupling that bridged the mid- line, the tongue may be controlled in a similar manner
to ensure bilateral activation. Electrotonic coupling among G G motoneurons
would serve to synchronize their collective discharge
and produce a more robust, uniform protrusion of the tongue. This contraction would, in turn, dilate the oropharynx and lessen the resistance to airflow. In human hand muscles, the subjects have been shown to synchronize the discharge of their motoneurons in or- der to sustain a maximal voluntary contraction 27. Syn-
chronization, in this case, permits the subject to gener- ate a larger force in a more efficient fashion. During the period when new synaptic inputs are being formed, it would be an advantage to the newborn to synchro- nize the discharge of its respiratory motoneurons in
order to optimize muscle performance. Loss of electrotonic coupling with age would release
motoneuron control of muscle fibers to individual mo- tor units. This is an important developmental step for the emergence of the graded and independent recruit- ment of motor units required for fine motor control 42.
We would suggest that the transition from shared excitation early in postnatal life to individualized in- puts from premotor centers may present a time period when neither mechanism is fully capable of maintain- ing adequate activity to prevent the collapse of the upper airways. This transition may be a factor in cases
of obstructive apnea reported in the premature and
newborn infants.
Acknowledgements. The authors wish to thank Mr. Fang He for his technical assistance. This work was supported by grants from the NIH (HD 22703) and the BRSG Fund of Magee-Womens Hospital. E.M. was supported by a Chancellor's Undergraduate Research

137
Fellowship from the University of Pittsburgh and P.N.-A. by a fellowship from the Comision Interministerial de Ciencia y Tecnolo- gia, Ministerio de Educacion y Ciencia, Spain.
R E F E R E N C E S
1 Adams, J.C., Technical considerations on the use of HRP as a neuronal marker, Neuroscience, 2 (1977) 141-145.
2 Aghajanian, G.K. and Rasmussen, K., Intracellular studies in the facial nucleus illustrating a simple new method for obtaining viable motoneurons in adult rat brain slices, Synapse, 3 (1989) 331-338.
3 Altman, J. and Bayer, S.A., Development of the brain stem in the rat. I. thymidine-radiographic study of the time of origin of neurons of the lower medulla, J. Comp. Neurol., 194 (1980) 1-35.
4 Arasaki, K., Kudo, N. and Nakanishi T., Firing of spinal mo- toneurons due to electrical interaction in the rat: an in vitro study, Exp. Brain Res., 54 (1984) 437-445.
5 Baker, R. and Llin~s, R., Electrotonic coupling between neu- rones in the rat mesencephalic nucleus, J. Physiol., 212 (1971) 45 -63.
6 Becker, D.L. and Navarette, R., Dye between lumbar motoneu- rones in the embryonic and neonatal rat spinal cord: an in vitro study, J. Physiol., 423 (1990) 98P.
7 Brenowitz, G.L., Collins, W.F. and Erulkar, S.D., Dye-coupled and electrically coupled neurones in frog spinal cord, Brain Res., 274 (1983) 371-375.
8 Brouillette, R.T. and Thach, B.T., Control of genioglossal muscle inspiratory activity, J. Appl. Physiol., 49 (1980) 801-808.
9 Collins, W.F., Organization of electrical coupling between frog lumbar motoneurons, J. Neurophysiol., 49 (1983) 730-744.
10 Connors, B.W., Benardo, L.S. and Prince, D.A., Coupling be- tween neurons of the developing rat neocortex, J. Neurosci., 3 (1983) 773-782.
11 Cullheim, S. and Ulfhake, B., Relations between cell body size, axon diameter and axon conduction velocity of triceps surae alpha-motoneurons during the postnatal development in the cat, J. Comp. Neurol., 188 (1979) 679-686.
12 Erulkar, S.D. and Soller, R.W., Interactions among lumbar mo- toneurons on opposite sides of the frog spinal cord: morphologi- cal and electrophysiological studies, J. Comp. Neurol., 192 (1980) 473-488.
13 Fulton, B.P., Miledi, R. and Takahashi, T., Electrical synapses between motoneurons in the spinal cord of the newborn rat, Proc. Roy. Soc. Lond., 208 (1980) 115-120.
14 Gauda, E.B., Miller, M.J., Carlo, W.A., Difiore, J.M., Johnsen, D.C. and Martin, R.J., Genioglossus response to airway occlusion in apneic versus nonapneic infants, Pediatr. Res., 22 (1987) 683- 687.
15 Gogan, P., Gueritaud, J.P., Horcholle-Bossavit, G. and Tyc- Dumont, S., Electrotonic coupling between motoneurons in the abducens nucleus of the cat, Exp. Brain Res., 21 (1974) 139-154.
16 Grace, A.A. and Llin~is, R.R., Morphological artifacts induced in intracellularly stained neurons by dehydration: circumvention us- ing rapid dimethyl sulfoxide clearing, Neuroscience, 16 (1985) 461-475.
17 Green, J.D. and Negishi, K., Membrane potentials in hypoglossal motoneurons, J. Neurophysiol., 26 (1963) 835-856.
18 Guthrie, S.C. and Gilula, N.B., Gap junctional communication and development, Trends. Neurol. Sci., 12 (1989) 12-16.
19 Kingsbury, B.F., The 'law' of cephalocaudal differential growth in its application to the nervous system, J. Comp. Neurol., 56 (1932) 431-464.
20 Kita, H. and Armstrong, W., A biotin-containing compound N- (2-aminoethyl) biotinamide for intracellular labeling and neu- ronal tracing studies: comparison with biocytin, Z Neurosci. Meth- ods, 37 (1991) 141-150.
21 Krammer, E.B., Rath, T. and Lischka, M.F., Somatotopic organi- zation of the hypoglossal nucleus: a HRP study in the rat, Brain Res., 170 (1979) 533-537.
22 Llimls, R. and Sasaki, K., The functional organization of the olivo-cerebellar system as examined by multiple Purkinje cell recordings, Eur. J. Neurosci., 1 (1989) 587-602.
23 Llin~is, R.R., Electrotonic transmission in the mammalian central nervous system. In M.V.L. Bennett and D.C. Spray (Eds.), Gap Junctions, Cold Spring Harbor Laboratory, Cold Spring Harbor, 1985, pp. 337-353.
24 Lodge, D., Duggan, T.J. and Caddy, K.W.T., Concerning recur- rent collaterals and afferent fibers in the hypoglossal nerve of the rat, Exp. Neurol., 41 (1973) 63-75.
25 MacVicar, B.A. and Dudek, F.E., Dye-coupling between CA3 pyramidal cells in slices of rat hippocampus, Brain Res., 196 (1980) 494-497.
26 MacVicar, B.A. and Dudek, F.E., Electrotonic coupling between pyramidal cells: a direct demonstration in rat hippocampal slices, Science, 213 (1981) 782-784.
27 Milner-Brown, H.F., Stein, R.V. and Lee, R.G., Synchronization of human motor units: possible roles of exercise and supraspinal reflexes, EEG Clin. Neurophys., 38 (1975) 245-254.
28 Motorina, M.V., Electrotonic synapses in the spinal cord of mammals, Neurosci. Behac. Physiol., 19 (1989) 72-78.
29 Navarrete, R. and Vrbowl, G., Changes of activity patterns in slow and fast muscle during postnatal development, Dec. Brain Res., 8 (1983) 11-19.
30 Nornes, H.O. and Das, G.D., Temporal pattern of neurogenesis in spinal cord of rat. I. an autoradiographic study-time and sites of origin and migration and settling patterns of neuroblasts, Brain Res., 73 (1974) 121-138.
31 Potter, D.D., Furshpan, E.J. and Lennox, R.S., Connections between cells of the developing squid as revealed by electrophysi- ological methods, Proc. Natl. Acad. Sci. USA, 55 (1966) 328-335.
32 Powell, S.L. and Westerfield, M., The absence of specific dye- coupling among frog spinal neurons, Brain Res., 294 (1984) 9-14.
33 Remmers, J.E., Degroot, W.J., Sauerland, E.K. and Anch, A.M., Pathogenesis of upper airway occlusion during sleep, J. Appl. Physiol., 44 (1978) 931-942.
34 Sasaki, K. and Llin~s, R., Dynamic electrotonic coupling in mammalian inferior olive as determined by simultaneous multiple Purkinje cell recording, J. Biophys., 47 (1985) 53a.
35 Schmalbruch, H., Skeletal muscles muscle fibers of newborn rats are coupled by gap junctions, Dec. Biol., 91 (1982) 485-490.
36 Sonnhof, U., Richter, D.W. and Taugner, R., Electrotonic cou- pling between frog spinal motoneurons. An electrophysiological and morphological study, Brain Res., 138 (1977) 197-215.
37 Sotelo, C. and Taxi, J., Ultrastructural aspects of electrotonic junctions in the spinal cord of the frog, Brain Res., 17 (1970) 137-141.
38 Stewart, W.W., Functional connections between cells as revealed by dye-coupling with a highly fluorescent naphthalimide tracer, Cell, 14 (1978) 741-759.
39 Uemura-Sumi, M., Itoh, M. and Mizuno, N., The distribution of hypoglossal motoneurons in the dog, rabbit and rat, Anat. Em- bryol., 177 (1988) 389-394.
40 Vaney, D.I., Many diverse types of retinal neurons show tracer coupling when injected with biocytin or Neurobiotin, Neurosci. Lett., 125 (1991) 187-190.
41 Vrbov~i, G., Navarrete, R. and Lowrie, M., Matching of muscle properties and motoneurone firing patterns during early stages of development, J. Exp. Biol., 115 (1985) 113-123.
42 Walton, K.D. and Navarrette, R., Postnatal changes in motoneu- rone electrotonic coupling studied in the in vitro rat lumbar spinal cord, J. Physiol., 433 (1991) 283-305.
43 Westerfield, M. and Frank, E., Specificity of electrical coupling among neurons innervating forelimb muscles of the adult bull- frog, J. Neurophysiol., 48 (1982) 904-913.
Note added in p roo f . T h e a u t h o r s w i s h to a c k n o w l e d g e t h e c o n t r i b u t i o n o f a p a p e r [ V i a n a , F., G i b b s , L. a n d B e r g e r ,
A . J . , Neuroscience, 3 (1990) 8 2 9 - 8 4 1 ] to t h e d e v e l o p m e n t o f t h e d o u b l e - l a b e l i n g m e t h o d o l o g y u s e d in t h e p r e s e n t
s tudy .