anatomical features ofleiopelma embryos and larvae: implications for anuran evolution
TRANSCRIPT

Anatomical Features of Leiopelma Embryos and Larvae:Implications for Anuran EvolutionBen D. Bell,1 and Richard J. Wassersug2*
1School of Biological Sciences, Victoria University of Wellington, Wellington, New Zealand2Department of Anatomy and Neurobiology, Sir Charles Tupper Medical Building, Dalhousie University, Halifax,Nova Scotia, B3H 1X5 Canada
ABSTRACT A controversial issue in anuran systematicsis the relationship of Leiopelma to other anurans becauserecent phylogenetic constructions imply different relation-ships among the basal frogs. Of particular evolutionaryinterest is whether early development of Leiopelma re-sembles an ancestral salamander-like larva, an anurantadpole, or neither. In the 1950s, Neville G. Stephensonhypothesized that direct development is the primary modeof development in amphibians, based on the fact thatLeiopelma spp. lack a free-living (�feeding) larval stage.Although this hypothesis has not been generally accepted,it has not been formally refuted. We review Stephenson’swork on Leiopelma and examine the anatomy of embryos/“larvae” of the four extant Leiopelma species for evidenceof vestigial larval features that might refute the “direct-developing ancestor” hypothesis. We describe internal oralfeatures in early developmental stages of Leiopelma andcompare Leiopelma with a closely related basal anuran,Ascaphus, to assess whether their early developmentalstages share any derived features. In Leiopelma hochstet-teri, embryos/larvae have open gill slits and some faintrugosities around one gill slit that may be vestiges of gillrakers or filters. They also have more intestinal loops,indicative of an elongated alimentary tract, at earlierrather than late embryonic/larval stages. Collectively,these features support the view that the ancestor of Leio-pelma had a free-swimming, free-feeding, aquatic larva.The palatoquadrate of Leiopelma archeyi reorients ap-proximately 40° from a more horizontal to a more verticalposition through embryonic/“larval” development. Thisamount of cranial remodeling is intermediate betweenthat seen in salamanders (17–27°) and that reported forAscaphus (64°) and other basal frogs (71–78°) at metamor-phosis. We found no internal oral features that Leiopelmashares specifically with Ascaphus. However, Leiopelmaembryos have a ventrally positioned mouth and a down-turned rostrum, characteristic of Ascaphus and otherstream-adapted tadpoles. J. Morphol. 256:160–170, 2003.© 2003 Wiley-Liss, Inc.
KEY WORDS: Leiopelma; metamorphosis; tadpoles; evo-lution; development
A controversial issue in anuran systematics is therelationship of Leiopelma to other basal anurans(see Reiss, 1997, for a recent discussion). At theheart of this controversy is the fact that Leiopelmaspecies have either direct-developing or semiaquatic,
nonfeeding larvae. This means that they lack exter-nal larval features that might otherwise aid in re-solving their phylogenetic relationships. Althoughinternal oral features have not been examined pre-viously in Leiopelma embryos/“larvae,” they havebeen described for tadpoles of the comparably ar-chaic anuran genus, Ascaphus (Wassersug, 1980).The present study of internal oral morphology ofLeiopelma embryos/“larvae” provides an opportu-nity to reexamine these two genera and to assesswhether their early developmental stages share anyderived features. We reason that the presence inLeiopelma of vestiges of larval structures used foraquatic feeding by tadpoles in other genera wouldsupport the hypothesis that the ancestor of Leio-pelma had a free-living aquatic stage. “Direct devel-opment” in Leiopelma then would be a derivedrather than a plesiomorphic state. Anatomical fea-tures associated with suspension feeding in tadpolesinclude the gill filters, internal oral papillation, oralvalves (i.e., ventral and dorsal vela), and branchialfood traps. These are all larval-specific structuresused to sort and capture particulate matter drawninto the mouth by water currents (Sanderson andWassersug, 1993). Vestiges of the same anatomicalstructures have been found in many anuran generathat have endotrophic development and otherwisehave abandoned the freely feeding larval stage(Wassersug and Duellman, 1984; Wassersug andHeyer, 1988). Only one “direct-developing” genus,Eleutherodactylus, is known to lack all vestiges ofany tadpole-specific, suspension-feeding structures(Wassersug and Heyer, 1988), yet even in this genus
Contract grant sponsors: the Natural Sciences and EngineeringResearch Council of Canada and Victoria University of Wellington.
*Correspondence to: Richard J. Wassersug, Department of Anat-omy and Neurobiology, Sir Charles Tupper Medical Building, Dalhou-sie University, Halifax, Nova Scotia, B3H 1X5 Canada.E-mail: [email protected]
DOI: 10.1002/jmor.10082
JOURNAL OF MORPHOLOGY 256:160–170 (2003)
© 2003 WILEY-LISS, INC.

there are vestiges of metamorphosis (Callery andElinson, 2000; Callery et al., 2001).
Phylogeny of the Anura
Traditionally, anurans were divided into the “ar-chaic” Archaeobatrachia and the “advanced” Neoba-trachia (sensu Duellman, 1975). Laurent (1979)erected the suborder Mesobatrachia for those ar-chaeobatrachians whose morphology seemed transi-tional between the two suborders, whereas Ford andCannatella (1993), using morphological characters,made the Mesobatrachia a sister group of the Neo-batrachia (to form the Pipanura) and abolished theArchaeobatrachia. Their two most basal lineageswere Ascaphus truei and Leiopelma, together form-ing the Leiopelmatanura. Cannatella (1985) de-scribed synapomorphies of the Leiopelmatanura:elongate arms on the sternum; loss of the ascendingprocess of the palatoquadrate; sphenethmoid ossi-fies in the anterior position; the root of the facialnerve exits the braincase through the facial fora-men, anterior to the auditory capsule, rather thanvia the anterior acoustic foramen into the auditorycapsule (Slabbert and Maree, 1945; Stephenson EM,1951); and a palatoquadrate articulation with thebraincase via a pseudobasal process rather than abasal process (Pusey, 1943). Leiopelma is uniqueamong living frogs in having cartilaginous inscrip-tional ribs in the muscles of the abdomen.
Green et al. (1989) concluded on the basis of anallozyme study that Leiopelma is the sister-group toAscaphus and all other frogs (represented by Bom-bina and Discoglossus). Hay et al. (1995) examinedphylogenetic relationships of amphibian families in-ferred from DNA sequences of mitochondrial 12Sand 16S ribosomal RNA genes and found that theirdata support the monophyly of the three amphibianorders (Anura, Caudata, and Gymnophiona).Zardoya and Meyer (2001) reached a similar conclu-sion. Within the Anura, Hay et al. (1995) foundsupport for the monophyly of the two suborders Neo-batrachia and Archaeobatrachia (sensu Duellman,1975). Within the Archaeobatrachia, they foundclustering between the following pairs of taxa: Pelo-batidae and Pelodytidae; Pipidae and Rhinophryni-dae; Ascaphus and Leiopelmatidae; and Bombinaand Discoglossidae. The latter three pairs clusteredto the exclusion of the first group, disputing thevalidity of the taxa Mesobatrachia (Laurent, 1979)and Pipanura (Ford and Cannatella, 1993) and un-expectedly placed the Pelobatoidea as the most basalarchaeobatrachian lineage. This would place Orton’s(1953, 1957) Type 4 tadpole (jaw sheaths, labialteeth, sinistral spiracle), seen in the Pelobatoidea,as plesiomorphic for the Anura.
The finding of Hay et al. (1995) that Ascaphus andLeiopelma are sister-groups is in contrast to an ear-lier study of 12S rRNA sequence data by Hedges andMaxson (1993) and other previous work (Ford and
Cannatella, 1993; Green and Cannatella, 1993) thatfound no sister-relationship between Ascaphus andLeiopelma. Cannatella (1999) reported that a com-bined analysis of morphological characters and se-quence data yielded a tree that supports Ford andCannatella (1993). Haas (2001) analyzed the struc-ture of the mandibular arch musculature in larval,metamorphic, and postmetamorphic anurans, mak-ing comparisons with larvae of three caudate andone gymnophione species. He found that with majortransformations in early evolution of anuran larvaeAscaphus shares the plesiomorphic conditions withthe Caudata, but he did not examine Leiopelma.
Maglia et al. (2001) examined larval skeletons andosteological development in 22 anuran taxa (but notLeiopelma) and compared two hypotheses ofrelationships—that of Cannatella and one they pro-pose based on 41 morphological characters from lar-vae and 62 from adults. They found that severallarval features are highly conserved among someanurans, suggesting that there is some level of ca-nalization of morphology early in ontogeny. In con-trast, a number of morphologies varied amonggroups, supporting the fact that there have beenmajor evolutionary modifications to anuran larvalmorphologies early in ontogeny and in the earlyevolutionary history of anurans.
All of the literature reviewed above points to onefact—viz., the historical relationships of Leiopelmato Ascaphus and to the remaining Anura remainproblematic.
Stephenson’s Hypothesis
The development of Leiopelma is of particular evo-lutionary interest because it is unclear if the earlysize–shape trajectory of the embryo resembles a tad-pole, an ancestral salamander-like larva, or neither(Harris, 1999). Stephenson (1951a) suggested thatdirect development in Leiopelma was a primarymode of development in amphibians based on theabsence of a free-living (�feeding) larva and onefrom which the indirect development of anurans,with a “tadpole” stage and striking metamorphosis,might have been derived. Thirty years later, Bogart(1981) revisited this idea when he wrote that “Undercertain circumstances aquatic development [for an-urans] may also be derived from a terrestriallybreeding mode.” Stephenson’s hypothesis has notbeen generally accepted by herpetologists (e.g.,Green and Cannatella, 1993), but neither has it beenformally rejected (see discussion in Harris, 1999).
Green and Cannatella (1993) argued that the pat-tern of development in Leiopelma is toward abbre-viation of the tadpole stage and failure to developspecialized tadpole features. For direct developmentof Leiopelma to be primitive for anurans, Leiopelmawould have to be the sister-group of all other extantanurans. Cannatella (1999) examined tadpole cra-nial and musculoskeletal architecture and drew in-
161LEIOPELMA EMBRYOS AND LARVAE

ferences in relation to anuran phylogeny. His fourmost basal lineages of living Anura (Ascaphus truei,Leiopelma, Bombinatoridae, and Discoglossidae)share the Type-3 tadpole of Orton (1953, 1957)—jawsheaths, labial teeth, and medial spiracle. He notedthat this in itself is not very informative, as the taxapossessing the plesiomorphic Type-3 larvae are di-verse, including Leiopelma larvae with “direct” de-velopment and Ascaphus with a stream tadpole, al-though more typical pond tadpoles are present inBombina and Discoglossus. He further noted thatwhile it is generally assumed that the specializedmorphologies of Ascaphus tadpoles and Leiopelmalarvae are derived from some pond tadpole ancestor(Griffiths, 1963), phylogenetic analysis of larvalcharacters does not demand such an interpretation;some features of the Ascaphus tadpole might beeither plesiomorphic for Anura or apomorphic forAscaphus.
Carroll et al. (1999) noted again that, althoughsome species of Leiopelma lack aquatic larval stages,the presence of a tadpole generally is accepted asprimitive for the modern anuran families and fossilsof pipid larvae are known from as early as the LowerCretaceous (Estes et al., 1978). Callery et al. (2001)reviewed anuran direct development, discussing theecological factors contributing to its evolution, andthe developmental modifications that facilitate sucha simplified mode of embryogenesis for an amphib-ian. Their studies on Eleutherodactylus coqui iden-tified such modifications as developmental adapta-tions for dealing with increased egg size and loss oftadpole structures and they suggest how the ances-tral biphasic developmental pattern might havebeen rearranged during the evolution of direct de-velopment in this leptodactylid genus.
In summary, we not only do not know how closeLeiopelma is to the ancestral anuran, but currentinformation on its life history provides only equivo-cal insights into the life history of that ancestor.
Features of New Zealand Leiopelma
The endemic New Zealand frog genus Leiopelmacomprises seven species, three of which are extinct(Worthy, 1987; Bell et al., 1998). Following the ter-minology of Altig and Johnston (1989), all four ex-tant species of Leiopelma are endotrophic, L. hoch-stetteri being nidicolous, while L. archeyi, L.hamiltoni, and L. pakeka are all exoviviparous,hatchlings completing development on the male’sdorsum (Archey, 1922; Stephenson, 1955; Bell, 1985;Bell et al., 1998; Thibaudeau and Altig, 1999).* Thethree terrestrial species (L. archeyi, L. hamiltoni, L.pakeka) inhabit forests and open ridge tops and the
semiaquatic species (L. hochstetteri) occurs in wettersites alongside creeks and damp water courses (Bell,1978, 1982a,b; Bell et al., 1998). The Maud Islandfrog was formerly regarded as Hamilton’s frog, L.hamiltoni, but has recently been described as a cryp-tic species, L. pakeka, based on allozyme and mor-phometric data (Bell et al., 1998), although Hol-yoake et al. (1999), using partial 12s RNA and Cyt bsequences, found little variation between the twotaxa (�1% for Cyt b), favoring keeping them as onespecies.
The terrestrial species lay 1–19 eggs in moist de-pressions on land under rocks, logs, or vegetationand adult males undertake parental care of eggs andhatchlings (dorsal brooding). The semiaquatic Leio-pelma hochstetteri lays 10–22 eggs in wet seepsunder rocks, logs, and vegetation. Adults (parents?)usually remain nearby, but close parental care isabsent (Turbott, 1949; Bell, 1978, 1982a,b, 1985).Early embryos of L. hochstetteri are pale gray onwhitish-yellow yolks, while those of terrestrial taxaare whitish on deeper yellow yolks (Bell, 1985). Eggcapsules of terrestrial species expand to 10–15 mmdiameter prior to hatching, which occurs at a rela-tively late stage of development (ca. Gosner [1960]Stage 35–37), although this varies depending onhydrous conditions. At hatching, the embryo’s eyesare well-developed and the body has started to showadult pigmentation. After hatching, young moveonto the flanks and dorsum of the male, where theyremain relatively inactive.
In contrast, egg capsules of Leiopelma hochstetterimay reach 20 mm in diameter prior to hatching,which occurs at an earlier stage of development(Gosner Stage 27–29), when only eye pigment isdistinct. At this stage the hatchlings are mobile andadept swimmers, although they remain near theoviposition site during development (Bell, 1985).
Larvae of terrestrial species have less-developedtail fins, no webbing between the toes, only the basesof the forelimbs covered by the opercular (gular)fold, and a relatively large spherical yolk mass. Incontrast, Leiopelma hochstetteri larvae have asmaller yolk mass and a longer, more muscular tail,and thus appear more streamlined and adapted forlife in an (albeit shallow) aquatic environment (Ste-phenson, 1955; Bell, 1978, 1982a,b, 1985). They alsohave more pronounced tail fins and the forelimbs aremore fully protected by an opercular fold. There istoe webbing on both fore- and hindfeet of the larva;in the semiaquatic adult, webbing is retained onlyon the hindfeet.
MATERIALS AND METHODS
Leiopelma is protected in New Zealand and its embryos arerare in museum collections. Our study is based on a series of 42Leiopelma specimens (Table 1) obtained from reproductive stud-ies under permits from the New Zealand government. Eggs orlarvae of L. archeyi Turbott, 1942, and L. hochstetteri Fitzinger,1861, were collected from 1974–1987 either in the field (the
*A statement in Pough et al. (1997) that Leiopelma larvae feed inwater is not correct.
162 B.D. BELL AND R.J. WASSERSUG
Coromandel ranges) or from captive breeding colonies, while allspecimens of L. hamiltoni McCulloch, 1919, and L. pakeka Bell etal., 1998, were captive-bred progeny collected in New Zealandfrom 1977–1983. After collection, eggs were placed on moist filterpaper in Petri dishes and reared in incubators in the dark at10–16°C (Bell, 1978, 1982a,b, 1985). Because of the rarity of thespecies, specimens were collected sparingly, only a few beingkilled specifically for study (Table 1). The small sample of pre-served L. hamiltoni larvae (n � 2) allowed only limited compar-ison with the other species; however, additional observations
were made (by BDB) on live specimens of this and the otherspecies.
Specimens were fixed in neutral buffered formalin or in Tyler’ssolution (Tyler, 1962). They were dissected and described in asemiblinded fashion with the person describing the anatomy notinformed in advance of the species assignment for the specimens.Most basic measurements were taken on preserved specimens(Table 1), although total lengths of 33 larvae were taken prior tofixation. The specimens are held in the personal collection of RJWin the Department of Anatomy and Neurobiology, Dalhousie Uni-
TABLE 1. Leiopelma specimens examined, arranged according to approximate Gosner stagea
Collectionspecimenno.
Total lengthbefore fixation
(mm)
Totallength(mm)
Bodylength(mm)
Snout–ventlengthb
(mm)
Headwidth(mm)
Bodywidth(mm)
Max tailheightc
(mm)
Gosner stagea
Hindlimbsonly
Headonly
Wholespecimen
L. pakekap12 16.5 14.3 6.1 7.9 1.6 5 30p13 18.5 18.5 7.5 8.8 2.8 6.2 31p1 19.4 9.3 10.7 3.4 5.7 1.5 38p6 21.8 21.5 8.9 10 3.7 0.7 33p14 21.5 21 8.5 10.2 2.8 6.2 34p2 21.7 20.7 8.1 11.4 3.8 6 3.9 34p5 22.5 22.3 8.8 10.9 3.4 5.7 1.7 32 43p3 19 18.1 8.7 10.5 2.5 5.8 1.8 36 44p7 18.5 17.3 9.7 9.7 4.2 6 39 44p10 c.13 10.7 9.1 9.1 3.7 5.5 38 45p8 21 20.9 9.5 11.6 4.1 6.6 2.4 39 45p9 21.2 18.4 8.6 10.2 3.7 5.5 39 45p4 20.6 19.1 9.3 9.3 3.8 5.6 45p11 22 19 8.2 9.8 2.9 5.8 44–45p15 18 9.8 10.9 3 6.2 44–45
L. hamiltonis2 22 18.3 7.5 8.8 3.5 5.8 2.1 34s1 23.5 22 10.4 12.2 3.8 7.4 1.5 37 44
L. archeyia5 15.9 15.5 8 8 2.8 4.7 36 44a6 8.5 6.7 6.7 3.1 4.6 36 44a4 20.2 18.3 6.9 8.2 3.2 4.1 1.7 37 44a8 10 9.7 7.3 9.7 2.7 4.6 37 44a7 9.5 8.4 9.5 3.3 5.8 39 45a1 7.4 7.4 7.4 3.5 3.7 1.4 40 45a10 18.7 7.1 8.3 2.9 4.6 2.1 32a2 20.7 18 7.3 8.4 3.4 4.1 1.2 34a9 20.4 8.1 9.1 3.2 3 1.2 35a3 20.1 17.3 7 7.8 2.9 4.9 1.2 39
L. hochstetterih2 16.8 15.7 5.3 5.9 1.2 3.5 1.1 32h6 17.9 13.1 7.2 8.2 2.5 5.3 1.4 33h1 17.4 15.1 6 6.7 2.7 3.8 1.3 34h8 21.8 19.3 8 9 2.6 5.4 1.4 35h9 21.5 19.2 7.4 9 3.8 4.6 2.2 35h15 25 23.8 8.1 9.9 3.3 5 35h11 18.1 17.3 7.4 8.9 3.4 5 37h12 15.2 7.2 8.1 3 4.9 37h7 18 16 7.7 8.4 3.7 4.7 37 44h5 25.1 17.5 7.9 8.8 3.8 4.6 1.7 39 44h14 18.8 8.2 9 3.8 3.8 45h10 12.2 11.9 7.8 11.9 3.9 4.6 37 45h3 9.5 7.8 7.8 7.8 3.9 3.3 46h4 9.6 9.5 9.5 9.5 4 3.5 46h13 9.6 9.5 9 9.2 3.8 3.8 post 46
aBecause of accelerated development of the head relative to the limbs in Leiopelma, compared to more typical neobatrachian larvae(for which the Gosner [1960] staging table was designed), in many Leiopelma specimens the head and limbs were at different Gosnerstages. For those specimens, the stage according to the hind limb and the head are given separately. When there was no discrepancy,the single Gosner stage for that specimen as a whole is given in the last column.bThe anal tube extends beyond the body and along the tail, which accounts for these values being slightly higher than those recordedfor “body length.”cBecause of regression of the tail, tail height cannot be reliably measured in most specimens past stage 43.
163LEIOPELMA EMBRYOS AND LARVAE

versity, Canada, for ultimate deposition in the National Museumof Natural History, Smithsonian Institution, Washington, DC,USA.
Anatomy was examined with binocular light microscopy. Dis-sected specimens were lightly stained with crystal violet to high-light surface texture and secretory tissues. Drawings of internaloral surfaces were made with a camera lucida. The terminologyused follows that of Wassersug and Duellman (1984) and Was-sersug and Heyer (1988). Specimens were staged according toGosner (1960). This staging table makes use of both the differen-tiation of the limbs for early stages and the remodeling of the jawsto stage developmentally more advanced tadpoles. In direct-developing anurans the timing of limb and cranial developmentmay be accelerated relative to typical pond larvae, as well asshifted relative to each other. As a result, staging a direct-developing embryo according to limb criteria may yield a differentstage value than if the same specimen were staged according tocranial features. Rather than devising a Leiopelma-specific stag-ing table (as has been done for Eleutherodactylus; see Townsendand Stewart, 1985) we simply assigned stages based on both limband cranial criteria for problematic Leiopelma specimens (Table1).
RESULTSDevelopment in Leiopelma hochstetteri
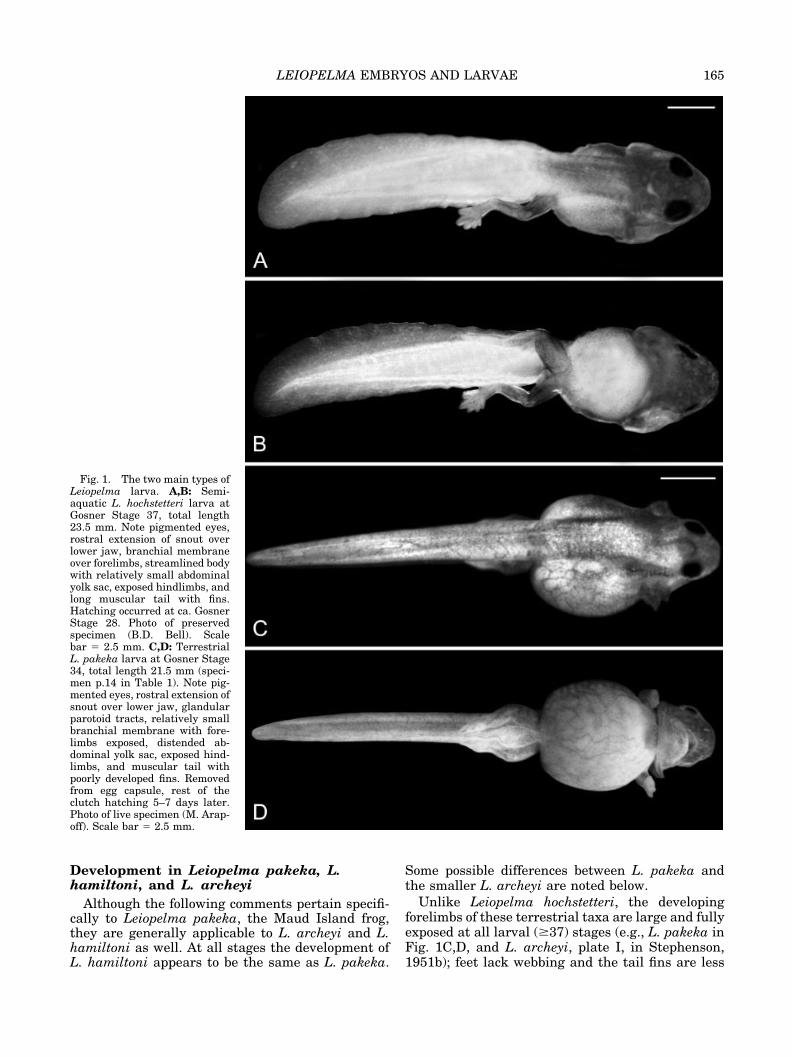
General description. Our Leiopelma hochstet-teri tadpoles (Fig. 1A,B) closely fit the external de-scription provided by Stephenson (1955) and onlymajor external features, plus new internal details,are recounted here. The pale gray L. hochstetteriembryos develop into larvae in which the first prom-inent pigment is around the eyes (Bell, 1985), al-though body pigment becomes increasingly evidentat later stages of development (by Stages 35–37).The developing forelimbs are protected by the largeopercular (gular) fold and are not exposed untillater, although by leg Stage 37 they may protrudethrough the ventral opening of the fold. The devel-oping appendicular skeleton can be observedthrough the translucent tissue of the limbs. Themouth lacks any oral disc, denticles, or other labialstructures. Instead it is a broad, simple arched slit.The oral orifice is ventrally positioned, with thesnout extending far rostral to it, particularly in theyounger stages—i.e., 1.5 mm beyond the jaw sym-physis at Stage 32/33. As a result, the snout curvesventrally, forming a lip ventral to the internal nares.The tail is muscular. The small dorsal and ventralfins are subequal in size. At Stage 37, when the finsare most prominent, each is approximately as tall asthe muscular portion of the tail along most of thelength of the tail. The fins are proportionally smallerin both younger and older specimens.
Because of the brittle nature of the formalin-fixedyolk and the extremely thin gut wall in these spec-imens, we could not measure the length of the ali-mentary tract. However some qualitative observa-tions on the viscera were made in three of thedissected specimens. At Stage 32, the gut tube hasapproximately 2.5 horseshoe-shaped loops. This in-creases to 3 or 4 such loops by Stage 34/35. Thealimentary tract appears to be absolutely longer atthis stage. By Stage 37, only two horseshoe loops are
discernible. At all of the stages examined, the liver issmall compared to that of exotrophic tadpoles. Nomanicotto glandulare—the precursor of the gastricantrum in exotrophic tadpoles—is discernable at thelevel of magnification in our dissections.
Lungs, gills, and gill slits. The lung buds aresmall and uninflated in tadpoles of Stage 37 oryounger. The lung buds are thin, pointed sacks, withaverage dimensions of 1.5 mm long and 0.3 mm widein the Stage-32 specimen. They increase slightly, to2.3 mm long and 0.6 mm wide, by Stage 37. No gillfilaments were detected. A crystal violet solutioninjected under pressure into the mouth of a Stage-32larva did not exit the pharynx, suggesting that thegill slits are not patent at this stage. At Stage 35,however, injected dye is expelled as a weak, asym-metric (stronger on the right) stream, and at Stage37 the dye comes out both sides approximatelyequally. By Stage 44/45 the opercular/gular fold isreduced to a faint gular ridge and there is no evi-dence of gill slits deep to it. Dye injection confirmsthe absence of patent gill slits at these metamorphicstages. In none of the tadpoles does dye exit thenares.
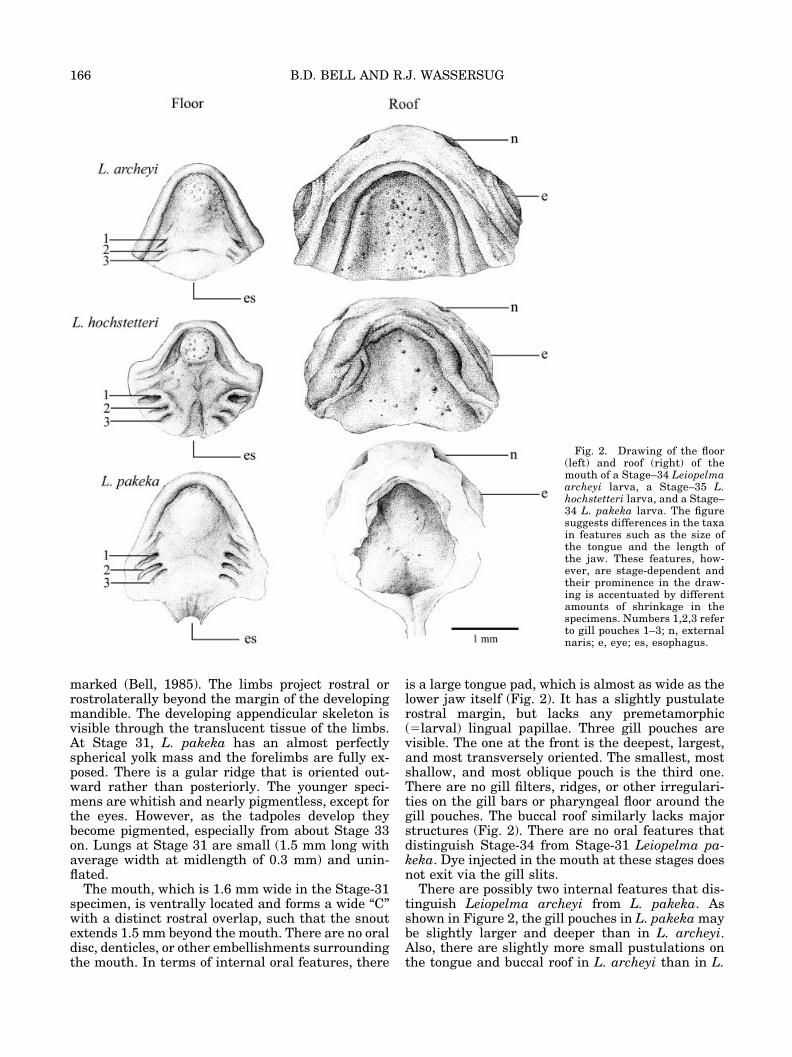
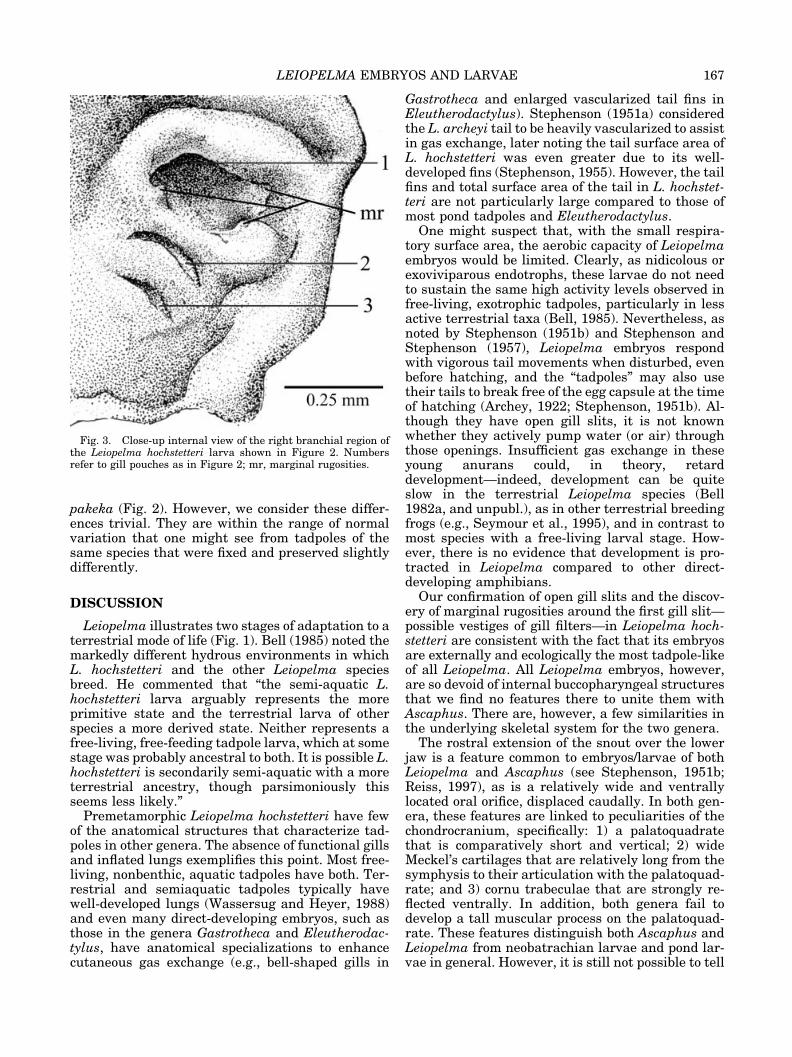
Internal oral features. Figure 2 shows the buc-cal floor and roof of a Stage-35 tadpole. The full archof the lower jaw is filled with the developing tonguepad. The tongue is a large, approximately roundmass with a faintly pustulate rostral margin. Thetongue lacks any projections comparable to thepremetamorphic lingual papillae of exotrophic tad-poles. The central portion of the buccal floor isslightly elevated. The only other features on thebuccal floor are three gill slits/pouches on each sideand a few (ca. 12–20) faint pustulations randomlylocated about the buccal floor. The first (most ros-tral) gill pouch is the deepest, largest, and mosttransversely oriented. The last is the shallowest,smallest, and most obliquely oriented. Under highmagnification, a few (2 or 3) slight and asymmetricrugosities line the posterior margin of the first gillslit (Fig. 3).
All buccal floor papillae are absent. No ventralvelum, branchial food traps, or gill filters were ob-served. The buccal roof at Stages 31–34 is similarlysimple (Fig. 2). Some 20 very faint pustulations arescattered about the central portion of the buccalroof. There is a faint transverse ridge that separatesthe caudal border of the buccal floor from the esoph-ageal funnel proper. Otherwise, the buccal roof, likethe buccal floor, is featureless; i.e., there is no sign ofpre- or postnarial papillae, narial valves, a medianridge, dorsal velum, secretory pits, or ciliary groove.
At Stage 37 the gill clefts are shallower andslightly reduced in size compared to the youngerstages. The only other buccopharyngeal feature thatdistinguishes Stage 37 Leiopelma hochstetteri fromyounger specimens is the presence of a single, tinypapilla on each side, on the recurved portion of thesnout, just ventral to the internal nares.
164 B.D. BELL AND R.J. WASSERSUG
Development in Leiopelma pakeka, L.hamiltoni, and L. archeyi
Although the following comments pertain specifi-cally to Leiopelma pakeka, the Maud Island frog,they are generally applicable to L. archeyi and L.hamiltoni as well. At all stages the development ofL. hamiltoni appears to be the same as L. pakeka.
Some possible differences between L. pakeka andthe smaller L. archeyi are noted below.
Unlike Leiopelma hochstetteri, the developingforelimbs of these terrestrial taxa are large and fullyexposed at all larval (�37) stages (e.g., L. pakeka inFig. 1C,D, and L. archeyi, plate I, in Stephenson,1951b); feet lack webbing and the tail fins are less
Fig. 1. The two main types ofLeiopelma larva. A,B: Semi-aquatic L. hochstetteri larva atGosner Stage 37, total length23.5 mm. Note pigmented eyes,rostral extension of snout overlower jaw, branchial membraneover forelimbs, streamlined bodywith relatively small abdominalyolk sac, exposed hindlimbs, andlong muscular tail with fins.Hatching occurred at ca. GosnerStage 28. Photo of preservedspecimen (B.D. Bell). Scalebar � 2.5 mm. C,D: TerrestrialL. pakeka larva at Gosner Stage34, total length 21.5 mm (speci-men p.14 in Table 1). Note pig-mented eyes, rostral extension ofsnout over lower jaw, glandularparotoid tracts, relatively smallbranchial membrane with fore-limbs exposed, distended ab-dominal yolk sac, exposed hind-limbs, and muscular tail withpoorly developed fins. Removedfrom egg capsule, rest of theclutch hatching 5–7 days later.Photo of live specimen (M. Arap-off). Scale bar � 2.5 mm.
165LEIOPELMA EMBRYOS AND LARVAE
marked (Bell, 1985). The limbs project rostral orrostrolaterally beyond the margin of the developingmandible. The developing appendicular skeleton isvisible through the translucent tissue of the limbs.At Stage 31, L. pakeka has an almost perfectlyspherical yolk mass and the forelimbs are fully ex-posed. There is a gular ridge that is oriented out-ward rather than posteriorly. The younger speci-mens are whitish and nearly pigmentless, except forthe eyes. However, as the tadpoles develop theybecome pigmented, especially from about Stage 33on. Lungs at Stage 31 are small (1.5 mm long withaverage width at midlength of 0.3 mm) and unin-flated.
The mouth, which is 1.6 mm wide in the Stage-31specimen, is ventrally located and forms a wide “C”with a distinct rostral overlap, such that the snoutextends 1.5 mm beyond the mouth. There are no oraldisc, denticles, or other embellishments surroundingthe mouth. In terms of internal oral features, there
is a large tongue pad, which is almost as wide as thelower jaw itself (Fig. 2). It has a slightly pustulaterostral margin, but lacks any premetamorphic(�larval) lingual papillae. Three gill pouches arevisible. The one at the front is the deepest, largest,and most transversely oriented. The smallest, mostshallow, and most oblique pouch is the third one.There are no gill filters, ridges, or other irregulari-ties on the gill bars or pharyngeal floor around thegill pouches. The buccal roof similarly lacks majorstructures (Fig. 2). There are no oral features thatdistinguish Stage-34 from Stage-31 Leiopelma pa-keka. Dye injected in the mouth at these stages doesnot exit via the gill slits.
There are possibly two internal features that dis-tinguish Leiopelma archeyi from L. pakeka. Asshown in Figure 2, the gill pouches in L. pakeka maybe slightly larger and deeper than in L. archeyi.Also, there are slightly more small pustulations onthe tongue and buccal roof in L. archeyi than in L.
Fig. 2. Drawing of the floor(left) and roof (right) of themouth of a Stage–34 Leiopelmaarcheyi larva, a Stage–35 L.hochstetteri larva, and a Stage–34 L. pakeka larva. The figuresuggests differences in the taxain features such as the size ofthe tongue and the length ofthe jaw. These features, how-ever, are stage-dependent andtheir prominence in the draw-ing is accentuated by differentamounts of shrinkage in thespecimens. Numbers 1,2,3 referto gill pouches 1–3; n, externalnaris; e, eye; es, esophagus.
166 B.D. BELL AND R.J. WASSERSUG
pakeka (Fig. 2). However, we consider these differ-ences trivial. They are within the range of normalvariation that one might see from tadpoles of thesame species that were fixed and preserved slightlydifferently.
DISCUSSION
Leiopelma illustrates two stages of adaptation to aterrestrial mode of life (Fig. 1). Bell (1985) noted themarkedly different hydrous environments in whichL. hochstetteri and the other Leiopelma speciesbreed. He commented that “the semi-aquatic L.hochstetteri larva arguably represents the moreprimitive state and the terrestrial larva of otherspecies a more derived state. Neither represents afree-living, free-feeding tadpole larva, which at somestage was probably ancestral to both. It is possible L.hochstetteri is secondarily semi-aquatic with a moreterrestrial ancestry, though parsimoniously thisseems less likely.”
Premetamorphic Leiopelma hochstetteri have fewof the anatomical structures that characterize tad-poles in other genera. The absence of functional gillsand inflated lungs exemplifies this point. Most free-living, nonbenthic, aquatic tadpoles have both. Ter-restrial and semiaquatic tadpoles typically havewell-developed lungs (Wassersug and Heyer, 1988)and even many direct-developing embryos, such asthose in the genera Gastrotheca and Eleutherodac-tylus, have anatomical specializations to enhancecutaneous gas exchange (e.g., bell-shaped gills in
Gastrotheca and enlarged vascularized tail fins inEleutherodactylus). Stephenson (1951a) consideredthe L. archeyi tail to be heavily vascularized to assistin gas exchange, later noting the tail surface area ofL. hochstetteri was even greater due to its well-developed fins (Stephenson, 1955). However, the tailfins and total surface area of the tail in L. hochstet-teri are not particularly large compared to those ofmost pond tadpoles and Eleutherodactylus.
One might suspect that, with the small respira-tory surface area, the aerobic capacity of Leiopelmaembryos would be limited. Clearly, as nidicolous orexoviviparous endotrophs, these larvae do not needto sustain the same high activity levels observed infree-living, exotrophic tadpoles, particularly in lessactive terrestrial taxa (Bell, 1985). Nevertheless, asnoted by Stephenson (1951b) and Stephenson andStephenson (1957), Leiopelma embryos respondwith vigorous tail movements when disturbed, evenbefore hatching, and the “tadpoles” may also usetheir tails to break free of the egg capsule at the timeof hatching (Archey, 1922; Stephenson, 1951b). Al-though they have open gill slits, it is not knownwhether they actively pump water (or air) throughthose openings. Insufficient gas exchange in theseyoung anurans could, in theory, retarddevelopment—indeed, development can be quiteslow in the terrestrial Leiopelma species (Bell1982a, and unpubl.), as in other terrestrial breedingfrogs (e.g., Seymour et al., 1995), and in contrast tomost species with a free-living larval stage. How-ever, there is no evidence that development is pro-tracted in Leiopelma compared to other direct-developing amphibians.
Our confirmation of open gill slits and the discov-ery of marginal rugosities around the first gill slit—possible vestiges of gill filters—in Leiopelma hoch-stetteri are consistent with the fact that its embryosare externally and ecologically the most tadpole-likeof all Leiopelma. All Leiopelma embryos, however,are so devoid of internal buccopharyngeal structuresthat we find no features there to unite them withAscaphus. There are, however, a few similarities inthe underlying skeletal system for the two genera.
The rostral extension of the snout over the lowerjaw is a feature common to embryos/larvae of bothLeiopelma and Ascaphus (see Stephenson, 1951b;Reiss, 1997), as is a relatively wide and ventrallylocated oral orifice, displaced caudally. In both gen-era, these features are linked to peculiarities of thechondrocranium, specifically: 1) a palatoquadratethat is comparatively short and vertical; 2) wideMeckel’s cartilages that are relatively long from thesymphysis to their articulation with the palatoquad-rate; and 3) cornu trabeculae that are strongly re-flected ventrally. In addition, both genera fail todevelop a tall muscular process on the palatoquad-rate. These features distinguish both Ascaphus andLeiopelma from neobatrachian larvae and pond lar-vae in general. However, it is still not possible to tell
Fig. 3. Close-up internal view of the right branchial region ofthe Leiopelma hochstetteri larva shown in Figure 2. Numbersrefer to gill pouches as in Figure 2; mr, marginal rugosities.
167LEIOPELMA EMBRYOS AND LARVAE

whether they are shared derived features of Asca-phus and Leiopelma or primitive features of Anura(cf. Wassersug and Hoff, 1982).
The absence in Leiopelma of structures related toaquatic feeding is surprising on two accounts. First,most other species of anurans with nonfeeding, en-dotrophic larvae still retain vestiges of suspension-feeding organs (Wassersug and Duellman, 1984;Wassersug and Heyer, 1988). The anurans with thefewest vestiges of buccopharyngeal structures usedin aquatic feeding are those of the direct-developingleptodactylid genus, Eleutherodactylus. Yet even inthat genus there is an egg tooth, which presumablyis derived from the keratinized mouthparts of itsancestor. Similarly, the large tail fin ofEleutherodactylus—a respiratory organ—is under-stood to have been derived from the tail fin of anaquatic ancestor. Callery and Elinson (2000) foundseveral other features in Eleutherodactylus thatsuggest vestiges of a larval stage and metamorpho-sis in that genus.
The simple buccopharyngeal structure of Leio-pelma supports the idea that this genus is quitedistant—morphologically, if not taxonomically—from the remaining extant Anura. It does not, how-ever, support Stephenson’s hypothesis that the tad-pole stage emerged at an evolutionary step aboveLeiopelma.
Stephenson’s hypothesis may be reformulated andrefined as a question: Did the ancestor of Leiopelmahave an aquatic larva and, if so, to what extent didthat larva resemble a modern tadpole? The first partof this question, we believe, was answered by Ste-phenson himself (1951a,b, 1955), when he observedthat Leiopelma embryos retained: 1) gill slits (whichwe have now confirmed are open ca. Stages 31–37);2) a muscular tail with tadpole-like tail-fin propor-tions in L. hochstetteri; and 3) an opercular (gular)fold, particularly developed in L. hochstetteri. All ofthese features are readily understood as features ofaquatic amphibian larvae. Their presence suggeststhat the ancestor of Leiopelma had an aquatic larva;however, they provide little insight as to how muchthat larva resembled a modern exotrophic tadpole.That issue, in turn, hinges on what one considers theessence of anuran larvae and the anuran life cycle.
The vestige of a possible gill ruffle or raker in theLeiopelma hochstetteri “larva” (Fig. 3) is the onlybuccopharyngeal surface feature related to suspen-sion feeding and grazing that we have identified inthe genus. This feature, however, is too diminutiveand equivocal to clarify whether Leiopelma’s ances-tor had suspension-feeding larvae—i.e., true gener-alized tadpoles. Nevertheless, we believe that thispossibly vestigial larval structure supports the viewthat the ancestor of Leiopelma had an anuran-likemetamorphosis. Ironically, the strongest evidencefor this conclusion comes again from Stephenson’sown work and not from this new observation.
Anurans are characterized, and distinguishedfrom other amphibians, by a metamorphic transitionthat is both abrupt and extreme. Wassersug andHoff (1982) argued that the more a tadpole differsmorphologically from its adult, the more abruptshould be its metamorphosis. They used the remod-eling of the anuran jaw suspension at metamorpho-sis to illustrate this thesis.
Tadpoles are unusual among aquatic vertebratesin having a long and relatively horizontal palato-quadrate bar. Linked to this unusual jaw suspensionare short, anteriorly displaced Meckel’s cartilages—hence, the characteristic, small mouth of tadpoles.Immediately caudal to Meckel’s cartilage in tad-poles, and underlying the floor of their buccal cavity,is a large hyoid plate. The vertical oscillations of thisplate—i.e., the tadpole’s buccal pump (reviewed inSanderson and Wassersug, 1993)—pulls water intothe mouth for feeding and respiration. This is essen-tial for their suspension-feeding way of life.
At metamorphosis, the palatoquadrate is short-ened and its distal end swings backward. This givesthe jaw suspension a more typically vertical orien-tation and allows Meckel’s cartilage to elongate—aprerequisite for converting a small-mouthed mi-crophagous tadpole into a large-mouthed macropha-gous frog. Stephenson’s (1951a) own illustrations ofthe chondrocranium in Leiopelma archeyi reveal anincrease in the angle between the base of the neu-rocranium and the palatoquadrate from approxi-mately 44° in a late intracapsular embryo to approx-imately 84° 1 week posthatching. This amounts to ashift of approximately 40° in a period comparable tothe time that many anurans go through metamor-phic climax. By comparison, the salamanders Am-bystoma and Hynobius, which do not have theabrupt metamorphic remodeling seen in Anura,have a maximum ontogenetic shift in the orientationof the palatoquadrate of only 17° and 27°, respec-tively (Wassersug and Hoff, 1982). Ascaphus, in con-trast, has a metamorphic realignment of its palato-quadrate of 64°, and other archaeobatrachian frogs(Alytes, Bombina, and Discoglossus) of 71–78°. Thismeans that the magnitude of metamorphic remod-eling of the suspensorium in Leiopelma is interme-diate between urodeles and other archaic Anura.
One other hint of an anuran-style metamorphosisin the ancestor of Leiopelma is the looping pattern ofthe premetamorphic alimentary tract in L. hochstet-teri. If we are correct that the gut tube in L. hoch-stetteri first elongates and then shortens betweenStages 37 and 44/45, this would be circumstantialevidence that Leiopelma’s ancestral alimentarytract had caenogenic specializations. Specifically,Leiopelma’s ancestor had an elongated alimentarytract that is characteristic of primarily herbivorousvertebrates and lost at metamorphosis, just likemodern tadpoles.
If we assume, for the sake of discussion, that theancestor of Leiopelma had a tadpole-like larva, then
168 B.D. BELL AND R.J. WASSERSUG

what type of tadpole was it? Although only yet-to-be-discovered fossils can answer that question, thecurrent ecology of L. hochstetteri suggests that itwould be a stream tadpole. Among the features ofthe L. hochstetteri semiaquatic “larva” that are alsofeatures of stream tadpoles are: a downturnedsnout, a ventral mouth, small and uninflated lungs,and low tail fins of relatively uniform height alongthe length of the tail. Coincidentally, along withrelatively long Meckel’s cartilages and relativelyshort and vertical palatoquadrate, these are all fea-tures that L. hochstetteri larvae share with Asca-phus tadpoles.
By examining anatomical features of the buccalcavity and pharynx, we have addressed the issue ofwhether the “direct” development seen in livingLeiopelma is a primitive feature of anurans or aderived feature of this particular genus. Althoughfew authors (other than Stephenson, 1951a) wouldsupport the idea that direct development is primi-tive for all anurans, especially given the widespreadoccurrence of this life history mode in diverse anu-ran lineages, this scenario is a formal possibility andas such should be entertained. Moreover, we knowrelatively little about the state of “tadpole” charac-ters in most direct-developing anurans, and it is ofinterest to see what remnants of these features existin those taxa.
An additional feature suggesting a tadpole ances-try for Leiopelma is the early contact between itsceratohyal and the palatoquadrate, later lost (Ste-phenson, 1951a, 1955). This articulation is a derivedfeature of tadpoles associated with the buccal-pumping mechanism and, apparently, it does notoccur in any other tetrapod.
In conclusion, we have found limited evidence ofvestigial buccopharyngeal features in premetamor-phic Leiopelma that supports the suggestion thatLeiopelma evolved from an amphibian with atadpole-like (e.g., microphagous, suspension-feeding) larva and an abrupt, anuran-like metamor-phosis. Additional evidence—taken largely fromStephenson’s own studies—suggests that Leiopel-ma’s ancestor minimally had an aquatic larva. Fur-thermore, the metamorphic transformation of thechondrocranium in that ancestral larva was likely tohave been intermediate between that seen in mod-ern urodeles and that of extant archaeobatrachiananurans. Finally, we have not found any ineluctable,derived features of the buccopharyngeal region thatare uniquely shared by Leiopelma and Ascaphus. Wehave, however, identified several features commonto the premetamorphic stages of both genera.
ACKNOWLEDGMENTS
We thank K. Oseen, N. Major, and J. Cookson forassistance in preparing the manuscript. R. Barkerprepared the illustrations. M. Fejtek assisted withcomposition of the photographic plate. N.J. Mitchell
and K. Oseen provided constructive comments ondrafts of the manuscript. S.N. Keall kindly arrangedfor the shipment of larval specimens. We thank theNew Zealand Department of Conservation and theformer New Zealand Wildlife Service for approvalfor research on Leiopelma frogs.
LITERATURE CITED
Altig R, Johnston, GF. 1989. Guilds of anuran larvae: relation-ships among developmental modes, morphologies, and habitats.Herpetol Monogr 3:81–109.
Archey G. 1922. The habit and life history of Leiopelma hochstet-teri. Rec Canterbury (NZ) Mus 2:59.
Bell BD. 1978. Observations on the ecology and reproduction ofthe New Zealand native frogs. Herpetologica 34:340–354.
Bell BD. 1982a. The amphibian fauna of New Zealand. In: New-man DG, editor. New Zealand herpetology. NZ Wildl Serv OccPubl 2:27–89.
Bell BD. 1982b. New Zealand frogs. Herpetofauna 14:1–21.Bell BD. 1985. Development and parental care in the endemic
New Zealand frogs. In: Grigg G, Shine R, Ehmann H, editors.The biology of Australasian frogs and reptiles. Chipping Nor-ton, NSW: Surrey Beatty & Sons. p 269–278.
Bell BD, Daugherty CH, Hay JM. 1998. Leiopelma pakeka, n. sp.(Anura: Leiopelmatidae), a cryptic species of frog from MaudIsland, New Zealand, and a reassessment of the conservationstatus of L. hamiltoni from Stephens Island. J R Soc NZ 28:39–54.
Bogart JP. 1981. How many times has terrestrial breedingevolved in anuran amphibians? Monit Zool Ital (NS) Suppl15:29–40.
Callery EM, Elinson RP. 2000. Thyroid hormone-dependentmetamorphosis in a direct developing frog. Proc Natl Acad SciUSA 97:2615–2620.
Callery EM, Fang H, Elinson RP. 2001. Frogs without polliwogs:evolution of anuran direct development. BioEssays 23:233–241.
Cannatella DC. 1985. A phylogeny of primitive frogs (archaeoba-trachians). Dissertation, University of Kansas.
Cannatella DC. 1999. Architecture: cranial and axial musculo-skeleton. In: McDiarmid R, Altig R, editors. Tadpoles: the biol-ogy of anuran larvae. Chicago: University of Chicago Press. p52–91.
Carroll RL, Kuntz A, Albright K. 1999. Vertebral developmentand amphibian evolution. Evol Dev 1:36–48.
Duellman WE. 1975. On the classification of frogs. Occ Pap MusNat Hist Univ Kansas 42:1–14.
Estes R, Spinar Z, Nevo E. 1978. Early Cretaceous pipid tadpolesfrom Israel (Amphibia: Anura). Herpetologica 34:374–393.
Fitzinger LJ. 1861. Eine neue Batrachier-Gattung aus New-Seeland. Verhandlungen der Zoologisch-botanischen Gesell-schaft in Wien 2:217.
Ford LS, Cannatella DC. 1993. The major clades of frogs. Herpe-tol Monogr 7:94–117.
Gosner KL. 1960. A simplified table for staging anuran embryosand larvae with notes on identification. Herpetologica 16:183–190.
Green DM, Cannatella, DC. 1993. Phylogenetic significance of theamphicoelous frogs, Ascaphidae and Leiopelmatidae. Ethol EcolEvol 5:233–245.
Green DM, Sharbel TF, Hitchmough RA, Daugherty CH. 1989.Genetic variation in the genus Leiopelma and relationships toother primitive frogs. Z Zool Syst Evol Forsch 27:65–79.
Griffiths I. 1963. The phylogeny of the Salientia. Biol Rev 38:241–292.
Haas A. 2001. Mandibular arch musculature of anuran tadpoles,with comments on homologies of amphibian jaw muscles. JMorphol 247:1–33.
Harris RN. 1999. The anuran tadpole: evolution and mainte-nance. In: McDiarmid R, Altig R, editors. Tadpoles: the biology
169LEIOPELMA EMBRYOS AND LARVAE

of anuran larvae. Chicago: University of Chicago Press. p 279–294.
Hay JM, Ruvinsky I, Hedges SB, Maxson LR. 1995. Phylogeneticrelationships of amphibian families inferred from DNA se-quences of mitochondrial 12S and 16S ribosomal RNA genes.Mol Biol Evol 12:928–937.
Hedges SB, Maxson LR. 1993. A molecular perspective on liss-amphibian phylogeny. Herpetol Monogr 7:27–42.
Holyoake AB, Waldman B, Gemmell N. 1999. A re-examination ofthe species status of Leiopelma hamiltoni and L. pakeka. Pro-ceedings of the Society for Research on Amphibians and Rep-tiles in New Zealand, Abstracts. New Zealand J Zool 26:258.
Laurent RF. 1979. Esquisse d’une phylogenese des anoures. BullSoc Zool Fr 104:397–422.
Maglia AM, Pugener LA, Trueb L. 2001. Comparative develop-ment of anurans: using phylogeny to understand ontogeny. AmZool 4:538–551.
McCulloch AR. 1919. A new discoglossid frog from New Zealand.Trans NZ Inst 51:447–449.
Orton GI. 1953. The systematics of vertebrate larvae. Syst Zool2:63–75.
Orton GI. 1957. The bearing of larval evolution on some problemsin frog classification. Syst Zool 6:79–86.
Pough FH, Andrews RM, Cadle JE, Crump ML, Savitzky A, WellsK. 1997. Herpetology. Englewood Cliffs, NJ: Prentice Hall.
Pusey HK. 1943. On the head of the leiopelmid frog, Ascaphustruei. I. The chondrocranium, jaws, arches, and muscles of apartly-grown larva. Q J Microsc Sci 84:105–185.
Reiss JO. 1997. Early development of chondrocranium in thetailed frog Ascaphus truei (Amphibia: Anura): implications foranuran palatoquadrate homologies. J Morphol 231:63–100.
Sanderson SL, Wassersug RJ. 1993. Convergent and alternativedesigns for vertebrate suspension feeding. In: Hanken J, HallBK, editors. The vertebrate skull: functional and evolutionarymechanisms, vol. 3. Chicago: University of Chicago Press. p37–112.
Seymour RS, Mahony MJ, Knowles R. 1995. Respiration of em-bryos and larvae of the terrestrially breeding frog Kyarranusloveridgei. Herpetologica 51:369–376.
Slabbert GK, Maree WA. 1945. The cranial morphology of theDiscoglossidae and its bearing upon the phylogeny of the prim-itive Anura. Ann Univ Stellen 23a:91–97.
Stephenson EM. 1951. The anatomy of the head of the NewZealand frog, Leiopelma. Trans Zool Soc London 27:255–305.
Stephenson NG. 1951a. On the development of the chondrocra-nium and visceral arches of Leiopelma archeyi. Trans Zool SocLond 27:203–253.
Stephenson NG. 1951b. Observations on the development of theamphicoelous frogs, Leiopelma and Ascaphus. J Linn Soc (Zool)Lond 42:18–28.
Stephenson NG. 1955. On the development of the frog, Leiopelmahochstetteri Fitzinger. Proc Zool Soc Lond 124:785–795.
Stephenson EM, Stephenson NG. 1957. Field observations on theNew Zealand frog, Leiopelma Fitzinger. Trans R Soc NZ 84:867–882.
Thibaudeau G, Altig R. 1999. Endotrophic anurans: developmentand evolution. In: McDiarmid R, Altig R, editors. Tadpoles: thebiology of anuran larvae. Chicago: University of Chicago Press.p 170–188.
Townsend DS, Stewart MM. 1985. Direct development in Eleu-therodactylus coqui (Anura: Leptodactylidae): a staging table.Copeia 1985:423–436.
Turbott EG. 1942. The distribution of the genus Leiopelma inNew Zealand with a description of a new species. Trans R SocNZ 71:247–253.
Turbott EG. 1949. Discovery of the breeding habits of Leiopelmahochstetteri Fitzinger. Rec Auckland Inst Mus 3:373–376.
Tyler MJ. 1962. On the preservation of anuran tadpoles. Aust JSci 25:222.
Wassersug R. 1980. Internal oral features of larvae from eightanuran families: functional, systematic, evolutionary and eco-logical considerations. Misc Publ Mus Nat Hist Univ Kansas68:1–146.
Wassersug RJ, Duellman WE. 1984. Oral structures and theirdevelopment in egg-brooding hylid frog embyos and larvae:evolutionary and ecological implications. J Morphol 182:1–37.
Wassersug RJ, Heyer WR. 1988. A survey of internal oral fea-tures of leptodactyloid larvae (Amphibia: Anura). SmithsonianContrib Zool 457:1–99.
Wassersug RJ, Hoff K. 1982. Developmental changes in the ori-entation of the anuran jaw suspension: a preliminary explora-tion into the evolution of metamorphosis. Evol Biol 15:223–246.
Worthy TH. 1987. Osteology of Leiopelma (Amphibia, Leiopelma-tidae) and descriptions of three new subfossil Leiopelma spe-cies. J R Soc NZ 17:201–251.
Zardoya R, Meyer A. 2002. On the origin and phylogenetic rela-tionships among living amphibians. Proc Nat Acad Sci USA98:7380–7383.
170 B.D. BELL AND R.J. WASSERSUG