anuran skin and basking behavior: the case of the treefrog bokermannohyla alvarengai (bokermann,...
TRANSCRIPT

Anuran Skin and Basking Behavior: the Case of theTreefrog Bokermannohyla alvarengai (Bokermann, 1956)
Fernanda C. Centeno,1 Marta M. Antoniazzi,2* Denis V. Andrade,1 Roberto T. Kodama,2
Juliana M. Sciani,3 Daniel C. Pimenta,3 and Carlos Jared2
1Departamento de Zoologia, Instituto de Biociencias, Universidade Estadual Paulista (UNESP), Av. 24A, 1515, CEP13506-900, Rio Claro, Brazil2Laborat�orio de Biologia Celular, Instituto Butantan, Av. Vital Brasil, 1500, CEP 05503-000, S~ao Paulo, Brazil3Laborat�orio de Bioqu�ımica e Biof�ısica, Instituto Butantan, Av. Vital Brasil, 1500, CEP 05503-000, S~ao Paulo,Brazil
ABSTRACT We investigated the morphology of theskin and the biochemistry of the lipids in the skinsecretion of Bokermannohyla alvarengai, a montanetreefrog that is known to bask regularly, motionless infull sunlight for extended periods of time. Our primarygoal was to identify structural and biochemical modifi-cations that might assist this frog species to accommo-date the conflicting demands for heat exchange andwater balance while basking. The modulation of heatexchange in basking B. alvarengai involves changes inskin coloration. We found that this response was sup-ported by a prominent monolayer of large iridophores,whose light reflectance property is adjusted by theresponse of intervening melanophores. Mucosubstancesand lipid compounds, mainly consisted of saturatedfatty acids and presumably secreted from granularglands, were detected on the skin of B. alvarengai.These compounds formed an extra-epidermal layer overthe animal’s dorsal surface that might assist in the pre-vention of excessive water loss through evaporation.Additionally, we found well-developed skin folds at theventral region of the frogs that lead to an increment ofsurface area. This feature combined with the extensivehypervascularization, also noticed for the skin of B.alvarengai, may play an important role in water reab-sorption. The suite of structural and biochemical modi-fications identified for the integument of B. alvarengaiseems to conjugate aspects relevant to both, heatexchange and water balance, allowing for this speciesto explore basking as an efficient thermoregulatorystrategy. J. Morphol. 000:000–000, 2015. VC 2015 Wiley
Periodicals, Inc.
KEY WORDS: Amphibia; Anura; thermoregulation;integument morphology; skin color change
INTRODUCTION
Temperature and water balance are the impor-tant features shaping the life history of amphib-ians. Indeed, body temperature is perhaps themost important ecophysiological variable affectingthe performance of anurans (Angilletta et al.,2002). Also, it can be particularly challenging forterrestrial species maintaining an adequate waterbalance given the characteristically highly water-
permeable skin of the group (Shoemaker et al.,1992). Temperature and water balance regulationinterfere with each other and the maintenance ofan adequate body temperature and hydration levelcan represent a trade-off between the mutual con-straints and benefits of each of these physiologicalfunctions. Thus, the ecological and evolutionarysuccess of anuran species that inhabit environ-ments with challenging thermal and water condi-tions will be accompanied by a diversity ofbehavioral, physiological, and morphological spe-cializations (Jared et al., 2005).
As ectotherms, anurans depend primarily on exter-nal heat sources to regulate their body temperature(Cowles, 1940; Bogert, 1949), mainly through behav-ioral adjustments (Cowles and Bogert, 1944; Bratt-strom, 1963; Lillywhite, 1971; Lillywhite et al., 1973;Hutchinson and Maness, 1979; Hillman et al., 2009).Basking is one of the main mechanisms by whichmost ectothermic organisms increase body tempera-ture, exposing whole or part of the body to direct sun-light (Muths and Corn, 1997). However, in
Contract grant sponsor: FAPESP; Grant number: 2009/15712-0(F.C.C); Contract grant sponsor: FAPESP; Grant number: 2010/05473-6 (D.V.A.); Contract grant sponsor: CNPq (C.J., D.C.P.,D.V.A. and M.M.A.), and Secretaria de Estado da Sa�ude do Estadode S~ao Paulo (R.T.K.).
Fernanda C. Centeno is currently at Department of Zoology, Uni-versity of Otago, P.O. Box 56, Dunedin, 9054, New Zealand.
Conflict of Interest: The authors state no conflict of interest.
*Correspondence to: Marta Maria Antoniazzi; Laborat�orio deBiologia Celular, Instituto Butantan, Av. Vital Brasil, 1500, CEP05503-000, S~ao Paulo, Brazil. E-mail: [email protected]
Received 3 March 2015; Revised 30 April 2015;Accepted 5 May 2015.
Published online 00 Month 2015 inWiley Online Library (wileyonlinelibrary.com).DOI 10.1002/jmor.20407
VC 2015 WILEY PERIODICALS, INC.
JOURNAL OF MORPHOLOGY 00:00–00 (2015)

amphibians, basking potentially increases the risk ofexcessive water loss through evaporation; thus, onlyfew anurans species are known to bask and, for thosebasking is usually accompanied by an improvementin skin resistance to evaporation (Bentley, 1966; Lilly-white, 2006). Also, basking anurans often exhibitphysiological color changes that modulate the amountof absorbed solar energy (Hoppe, 1979; Stegen et al.,2004) and consequently, affect body temperature andevaporative water loss (EWL).
The anuran skin, therefore, is the undisputedinterface for exchanging heat and water betweenthe animal and the environment, and it is herethat one would expect to encounter more obviousfunctional compromises in basking species. In thiscontext, the skin of anurans that bask is likely tohave structural specializations that can be corre-lated with their life history (Toledo and Jared,1993). We study specializations of the skin of Boker-mannohyla alvarengai, a large South Americantreefrog endemic to the campos rupestres (rockymeadows) above 1,000 m (Sazima and Bokermann,1977; Eterovick and Sazima, 2004) in southeasternBrazil. Despite being nocturnal, this species isknown to bask regularly on rock outcrops, wherethey can remain motionless for extended periods oftime completely exposed to full sunlight and wind(Sazima and Bokermann, 1977). While basking, B.
alvarengai changes skin color markedly, turningfrom a mottled cryptic pattern to a chalky white(Fig. 1A, B), a color change that is involved in heatexchange modulation (Tattersall et al., 2006).
MATERIAL AND METHODSThe Habitat
Campos rupestres is a phytophysiognomic type of the Brazil-ian Cerrado; typically, it occurs above 1,000 m, and is predomi-nantly characterized by low-growing vegetation with a fewshrubs and small trees amid rocky outcrops (Fig. 1C). The localclimate is the altitudinal tropical with rainy summers of thetype Cwb (following K€oppen’s classification; K€oppen, 1900), andwell-established dry season. The high seasonality is character-ized by rainy periods between October and March, with meantemperature from 12.5 2 29.6�C and 288 mm of mean rainfall,and drier periods between April and September, with meantemperature from 8.6 2 26.8�C and 33 mm of mean rainfall.
Data Collection
Six females (snout-vent length5 74.4 2 86.0 mm; weight 5
31.7 2 57.4 g) Bokermannohyla alvarengai (Bokermann, 1956)were collected during three expeditions to the Serra do Cip�o(19812’–19820’S; 43830’–43840’W) in the southern portion of theEspinhaco Mountain Range, in the state of Minas Gerais betweenNovember 2011 and July 2012; sampling occurred during bothrainy and dry periods (SISBIO, 22170-1/2010). Right after beingcollected and before being euthanized, three specimens were leftcompletely exposed to the sunlight for a 10 min period (hereafter,called “bright”), and three were held in the dark for 30 min
Fig. 1. Bokermannohyla alvarengai, a basking individual with a bright skin color (A). The sameindividual with a dark skin pattern a few minutes after having been removed from the sun (B). Gen-eral view of campos rupestres at Serra do Cip�o (MG, Brazil) where B. alvarengai is found (C).
2 F.C. CENTENO ET AL.
Journal of Morphology

(hereafter, called “dark”); these time periods are sufficient for thephysiological color change to occur (Tattersall et al., 2006). Thefrogs then were injected with a lethal dose of thiopental (0.03 ml/g) and immediately photographed for color documentation. Piecesof skin (0.5 3 0.5 cm) were excised from the dorsal, mental, pecto-ral, ventral, and inguinal regions, and immediately fixed in 4%paraformaldehyde in phosphate-buffered saline, pH 7.2 (Jun-queira, 1995), for light microscopy (LM), and in Karnovsky fixa-tive solution (5% glutaraldehyde 1 4% paraformaldehyde insodium cacodilate buffer, 0.1 mol l21, pH 7.2; Karnovsky, 1965),for scanning and transmission electron microscopy (SEM andTEM). The procedures with the bright specimens were conductedin the sunlight, whereas those with the dark specimens were con-ducted in a minimal light to preserve the skin coloration. Aftertissue collection, the anurans were fixed in 10% formalin, andmaintained in 70% ethyl alcohol. The voucher specimens aredeposited in the C�elio F. B. Haddad amphibian collection, Depar-tamento de Zoologia, Instituto de Biociencias, UniversidadeEstadual Paulista, Campus Rio Claro, S~ao Paulo, Brazil.
The animals were handled according to the procedures indi-cated in the Guidelines for Animal Experimentation establishedby the Brazilian College for Animal Experimentation.
Morphological Analysis
The skin samples were examined in the Laborat�orio de BiologiaCelular of Instituto Butantan. All photomicrographs wereobtained with an Olympus BX51 microscope equipped with a CCDcamera Olympus (Q Color 5), coupled with the software Image-Pro Express (Media Cybernetics, Inc., Rockville, MD, USA).
Light Microscopy
Skin fragments fixed in paraformaldehyde were dehydratedin crescent ethanol series (70, 95, and 100%), and embedded inglycol methacrylate (Leica, Deerfield, IL, USA), sectioned 2 mmthick, and stained with toluidine blue-fuchsin for general obser-vations. Skin samples also were embedded in paraffin, sec-tioned 5 mm thick, and stained with hematoxylin-eosin forgeneral morphological observation, von Kossa for detection ofcalcium, and Sudan Black B for identification of lipids.
Transmission Electron Microscopy
Skin fragments fixed in Karnovsky solution were postfixed in1% osmium tetroxide and precontrasted in 0.5% uranylacetate 1 13.3% sucrose. After dehydration in crescent ethanolseries, the samples were embedded in epoxy resin (ElectronMicroscopy Sciences, Hatfield, PA, USA) in flat molds. Ultrathinsections (60 2 70 nm) were obtained in a Sorvall MT 6000 ultrami-crotome, collected on copper grids, contrasted in aqueous 2% ura-nyl acetate and lead citrate, and examined using a LEO 906ETEM (Carl Zeiss, Germany). Images were captured by a CCD cam-era MegaView III, using the software Item- universal TEM Imag-ing Platform (Olympus Soft Imaging Solutions GmbH, Germany).
Scanning Electron Microscopy
The sections fixed in Karnovsky fixative were postfixed in 1%osmium tetroxide and cleaned ultrasonically at a frequency of 40Hz in a solution containing sodium alkyl benzene sulfonate(Reckitt Benckiser, Brazil). After have been washed in distilledwater, the samples were dehydrated in crescent ethanol series,dried in critical point drier equipment CPD 030 (Leica Microsys-tems, Heerbrugg, Switzerland), mounted in aluminum stubs,and covered with gold in sputtering equipment SCD 050 (LeicaMicrosystems, Wetzlar, Germany). The samples were then exam-ined in a Quanta 250 SEM (FEI Company, Hillsboro, OR, USA).
Biochemical Analysis of Lipids in the SkinSecretion
Skin secretion of Bokermannohyla alvarengai was extractedfrom male and female specimens (n 5 4), by gentle manual
stimulation of the skin of each individual immersed in a beakercontaining 50 ml of ultrapure water. The pooled secretion wasthen lyophilized and 2 mg (dry weight) was resuspended in1 ml ultrapure water. A liquid–liquid extraction was performedto obtain molecules soluble in lipids. A mixture of chloroformand methanol (1:2) was added (3.7 ml) in the secretion andhomogenized for 10 min. Then, 1.25 ml chloroform was addedand mixed for 1 min. Water ultrapure (1.25 ml) was added andthe solution was centrifuged at 8,000 g for 5 min. The upperphase was separated from the bottom phase, which was thenconcentrated and analyzed by gas chromatography–mass spec-trometry (GC–MS).
Gas chromatography–mass spectrometry analysis was per-formed on a gas chromatography system (7890A, Agilent Tech-nologies) equipped with a flame ionization detector (FID),coupled to a 5975C mass detector (Agilent Technologies). Sam-ples were injected in the splitless mode (1 ml) at an injectortemperature of 1208C. Molecules were separated by a HP-5MScolumn (30 m 3 0.25 mm, 0.25 mm), with oven temperatureprogrammed from 708C (isothermal for 2 min) to 3208C, at 58C/min (total run 60 min). Helium was used as carrier gas, at aflow rate of 1.7 ml/min. Mass spectra were obtained throughelectron impact (70 eV) and results were compared to NationalInstitute of Standards and Technology database (http://www.nist.gov/pml/data/asd.cfm), to obtain the molecule identity, andmanually checked.
RESULTS
The skin of Bokermannohyla alvarengai is typi-cal of that of other anurans. The skin is 245 6 11mm thick in the middle dorsal region and 354 6 35mm in the ventral (pectoral) region. Both dorsallyand ventrally, the skin is composed of an epider-mis, a dermis with a stratum spongiosum and astratum compactum, and a subcutaneous tissue(Fig. 2A). Interspersed muscle fibers transversethe skin to connect the base of the epidermis withsubcutaneous tissue. The dorsal epidermis com-prises of 6 or 7 cell layers (Fig. 2B,C), whereas theventral epidermis has 5 or 6 layers.
A pigment layer underlies the dorsal epidermis.It is composed of a thin layer of xantophores ontop of a monolayer of large iridophores (27 6 2 mm3 10 6 1 mm), and an inferior layer of melano-phores, with prominent cytoplasmic processes thatembrace the iridophores (Fig. 2B,C). In animalskilled in the darkness, pigments were well dis-persed within the cytoplasmic processes of themelanophores (Fig. 2C), whereas in animals killedin the light, pigments were concentrated in thenuclear region (Fig. 2B). The prominent pigmentlayer is typical of the dorsum, which also hassome melanophores around the glands, near thecalcified dermal layer (Figs. 2B and 3D) and in thesubcutaneous tissue (Figs. 2D and 3E). The ven-tral skin has melanophores that are sparsely dis-tributed in equivalent positions to those of thedorsal skin. A few pigment cells also occur in theinguinal skin.
Granular and two types of mucous glands arelocated below the pigment cells, as in other anu-rans (Fig. 2A,B). The granular glands usually areround or bottle-shaped, and are much larger than
3BASKING BEHAVIOR IN TREEFROG
Journal of Morphology

the mucous glands; they are syncytial and full ofspherical homogeneous-sized eosinophilic granules(Fig. 2B,C), which are reactive to bromophenolblue (Fig. 2G) and have a moderate affinity toSudan Black B (Fig. 2E). In transmission electronmicrographs, the granules are quite electron-dense(Fig. 3A,B). The mucous glands are acinar and
composed of a simple secretory epithelium and alumen. One type (m1) is usually larger; it is char-acterized by a wide lumen and a short secretoryepithelium constituted by cells of only one type,which are very reactive to PAS and alcian blue(Figs. 2B,F and 3B). The other type (m2) is usuallysmaller; it has a narrow lumen and is composed of
Fig. 2. Bokermannohyla alvarengai. (A) General view of the dorsal skin. Note the large number of granular glands and the adipo-cytes in the subcutaneous layer. The arrows point to mucous glands, and iridophores are represented by asterisk (*). Paraffin section,HE staining. (B) Detail of the dorsal skin of an individual euthanized while exposed to the light. The iridophores (*) are totallyexposed to the light owing to the concentration of the pigments in the nuclear region of the melanophore (arrowheads). The arrowpoints to the calcified dermal layer. Historesin section, toluidine-fuchsin staining. (C) Detail of the dorsal skin of an individual eutha-nized while in the dark. The iridophores (*) are hidden by the dispersal of the pigments in the melanophore cytoplasmic processes(arrowheads). The arrow points to the nuclear region of a melanophore. Historesin section, toluidine-fuchsin staining. (D) Detail ofthe dorsal skin showing the subcutaneous tissue full of adipocytes, and the presence of melanophores (arrowheads). Paraffin section,HE staining. (E) General view of the dorsal skin in a section equivalent of Figure 2A, showing a moderate affinity of the granularglands to Sudan Black staining. The arrow points to mucous gland. Paraffin section, Sudan Black histochemistry method. (F) Pecto-ral skin, focusing the two types of mucous glands after being submitted to alcian blue, pH 2.5. Paraffin section, alcian blue, pH 2.5histochemistry method. (G) Pectoral skin, focusing the two types of mucous glands and a granular gland after being submitted tobromophenol blue reaction. Paraffin section, bromophenol blue histochemistry method. E, epidermis; g, granular gland; li, adipocytes;m1, mucous gland type 1; m2, mucous gland type 2; sc, stratum compactum; ss, stratum spongiosum.
4 F.C. CENTENO ET AL.
Journal of Morphology
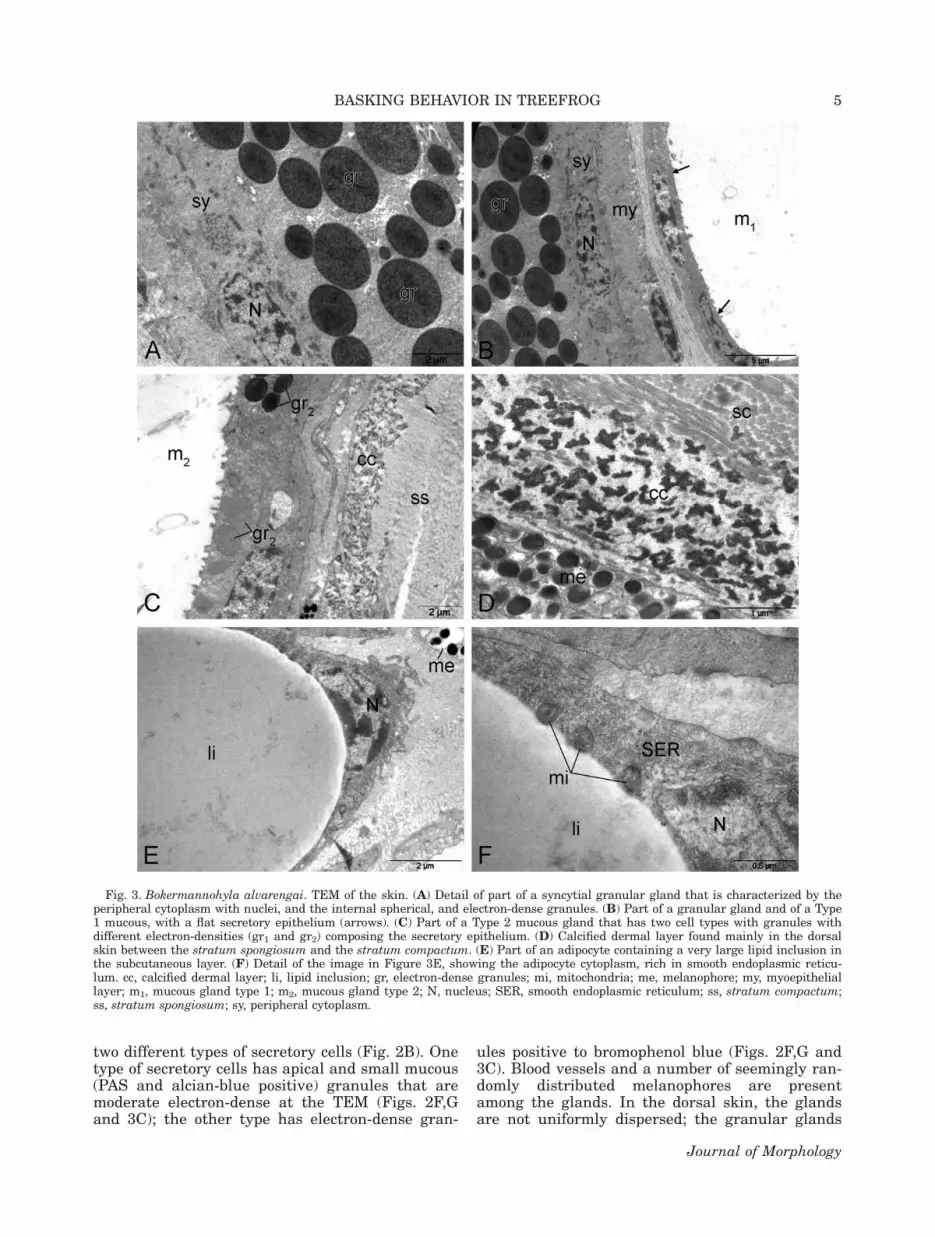
two different types of secretory cells (Fig. 2B). Onetype of secretory cells has apical and small mucous(PAS and alcian-blue positive) granules that aremoderate electron-dense at the TEM (Figs. 2F,Gand 3C); the other type has electron-dense gran-
ules positive to bromophenol blue (Figs. 2F,G and3C). Blood vessels and a number of seemingly ran-domly distributed melanophores are presentamong the glands. In the dorsal skin, the glandsare not uniformly dispersed; the granular glands
Fig. 3. Bokermannohyla alvarengai. TEM of the skin. (A) Detail of part of a syncytial granular gland that is characterized by theperipheral cytoplasm with nuclei, and the internal spherical, and electron-dense granules. (B) Part of a granular gland and of a Type1 mucous, with a flat secretory epithelium (arrows). (C) Part of a Type 2 mucous gland that has two cell types with granules withdifferent electron-densities (gr1 and gr2) composing the secretory epithelium. (D) Calcified dermal layer found mainly in the dorsalskin between the stratum spongiosum and the stratum compactum. (E) Part of an adipocyte containing a very large lipid inclusion inthe subcutaneous layer. (F) Detail of the image in Figure 3E, showing the adipocyte cytoplasm, rich in smooth endoplasmic reticu-lum. cc, calcified dermal layer; li, lipid inclusion; gr, electron-dense granules; mi, mitochondria; me, melanophore; my, myoepitheliallayer; m1, mucous gland type 1; m2, mucous gland type 2; N, nucleus; SER, smooth endoplasmic reticulum; ss, stratum compactum;ss, stratum spongiosum; sy, peripheral cytoplasm.
5BASKING BEHAVIOR IN TREEFROG
Journal of Morphology

are concentrated in some regions where they arelarge and bottle-shaped (Fig. 2A).
A continuous calcified dermal layer about3.8 6 0.8 mm thick lies just below the glands in thedorsal skin (Fig. 2B). In TEM, it is characterizedas a heterogeneous aggregation of irregularelectron-dense vesicles (Fig. 3C,D). Commonly thelayer is overlain by melanophores (Fig. 3D).Although more obvious in the dorsal skin, a dis-crete noncontinuous calcified layer also occurs inthe ventral skin. It is most evident in the cranialregion and less so toward the caudal region, andnever been observed in the inguinal region.
The stratum compactum, which is composed ofthick collagen fibers that are arranged in abasket-like interlacing, lies below the stratumspongiosum (Fig. 2A). The subcutaneous tissue iscomposed of loose connective tissue containingblood vessels, nerves, and many large lipid drop-
lets; the latter are grouped in some regions butabsent in others (Fig. 2A,D,E). These lipid dropletsare present in great numbers in the dorsal andpectoral skin, but absent in the ventral and ingui-nal regions. When observed with TEM, the drop-lets look like as typical adipocytes with aperipheral nucleus and a narrow region of cyto-plasm that contains smooth endoplasmic reticulumsurrounding a large spherical lipid inclusion (Fig.3E,F).
The surface layers of the skin have a uniformarrangement across the body. The outermost epi-dermal cells are polygonal and strongly connected.The skin microdermatoglyphics reveals a charac-teristic labyrinthic design. Glandular pores occurin the intercellular spaces in round or crow’s feetarrangements (Fig. 4A). The skin of the pectoraland particularly the inguinal region is character-ized by many folds forming a pattern of elevations
Fig. 4. Bokermannohyla alvarengai. SEM of the skin. (A) Surface of the dorsal skin showing the typical labyrinth design of thepolygonal epidermal cells. (B) Verrucae hydrophilica in the pectoral region. (C) Verrucae in the inguinal region. (D) Detail of a ver-ruca in the pectoral region showing the surface of the epidermal cells full of small tips (*). (E) Detail of the tips (*) in a verruca ofthe inguinal region. The arrows point to glandular pores. ce, cellular limits.
6 F.C. CENTENO ET AL.
Journal of Morphology

and grooves and giving skin the appearance of“mountains” and “valleys” (Figs. 4B,C and 5A).Scanning electron micrographs reveal that the
surfaces of the folds have many small irregularlydispersed pits with diameters around 3.5 mm (Fig.4D,E).
In addition to having folds, the inguinal skinhas an external epidermal layer that is less corni-fied than that of the rest of the body, and withcells possessing a clear cytoplasm and a morerounded (less flat) shape (Fig. 5B). In TEM, thesecells have moderate electron-density and pycnoticnuclei (Fig. 5C). Many other clear cells areobserved in the three more superficial epidermallayers. The vascular system in the inguinal regionis prominent. Thick vessels in the subcutaneoustissue branch toward the surface, giving rise to alarge number of capillaries that course into theepidermis above; in some cases, these capillariesare only two or three cell layers from the surfaceof the body (Fig. 5A–C). The combination of ingui-nal skin peculiarities including the presence offolds, an extensive capillary network, and lack of acornified layer characterizes the structure termedverrucae hydrophilica.
Analysis by GC–MS of the organic extractobtained from Bokermannohyla alvarengai skinsecretion showed a complex profile of molecules, asshown in Figure 6A,B. Most of lipids are saturatedfatty acids, as shown in Table 1 with the most
Fig. 5. Bokermannohyla alvarengai. (A) General aspect of theverrucae in the inguinal region. Note the large number of bloodvessels coming from the subcutaneous tissue and giving rise tocapillaries (*) that penetrate the epidermis. LM. Historesin sec-tion, toluidine-fuchsin staining. (B) Detail of the capillaries (*)penetrating the epidermis. Note the clear cells (arrow) coveringthe surface of the skin. LM, Historesin section, toluidine-fuchsinstaining. (C) A detailed view of a capillary (*) near the epidermissurface of the inguinal skin. Note the clear cell forming theexternal layer, with a pycnotic nucleus (arrow). TEM. ce, cellularlimits; E, epidermis; m, mucous glands; sc, stratum compactum.v, blood vessels.
Fig. 6. Organic fraction of the skin secretion of Bokermanno-hyla alvarengai analyzed by GC–MS (FID detector). (A) Thelipid peaks identified are numbered from 1 to 11. (B) Anenlarged region of Figure 6A showing Peaks 3 2 10.
7BASKING BEHAVIOR IN TREEFROG
Journal of Morphology

abundant being the fatty acids phenol, 2,4-bis(1,1-dimethyl)- (Peak 2) and 1,2-benzenedicarboxylicacid, bis(2-methylpropyl)ester (Peak 8). Severalother intense peaks in the profile could be identi-fied, such as hexadecanoic acid, methyl ester (pal-mitic acid) (Peak 10), and octadecanoic acid,methyl ester (estearic acid) (Peak 11) and phthalicacid, decyl methyl ester (Peak 3).
DISCUSSION
The highly permeable skin of anurans generallylimits their ability to use basking as a means ofincreasing body temperature because exposure tothe sun usually results in increased rates of EWL(Jameson, 1966; Shoemaker et al., 1992). Somespecies, however, seem to circumvent this problembehaviorally by basking near or in shallow bodiesof water, which enables an easy transition betweenthe sunlight and more amenable conditions (Bratt-strom, 1963; Lillywhite, 1970; Tracy et al., 1993).However, a few anuran species can remain station-ary and exposed to direct solar radiation for pro-longed periods away from water sources withoutcompromising their water balance (Lillywhite,1970). This is certainly the case of Bokermanno-hyla alvarengai. Here, we focus on constraints andbenefits associated with body temperature andwater balance regulation and how they are relatedto particular structural modifications found in theskin of this basking treefrog.
Basking anurans have a suite of behavioral,physiological, and morphological specializationsthat allow them to employ a thermoregulatorystrategy otherwise unavailable to most amphib-ians. For example, physiological ability to changecolor in Bokermannohyla alvarengai modulatesheat absorption (see also Tattersall et al., 2006).In this species, as well as in other basking anu-rans, the ability to change color is linked to theorganization of chromatophores in the dermis ofthe dorsal skin associated with the dispersal/
aggregation response of the pigmentary organelles(e.g., Kobelt and Linsenmair, 1986; King et al.,1994; Withers, 1995). Ordinarily, a dermal chroma-tophore unit consists of a central iridophorecapped by a xanthophore and underlain by a largemelanophore with dendrites extending betweenthe iridophores and xanthophores (Bagnara et al.,1968). The iridophores are responsible for palebrightening coloration through the light-reflectingorganelles composed of stacks of purine-containingreflecting platelets, whereas the melanophoresplay a role in darkening the animal by rearrange-ment of melanin in the cytoplasm (Bagnara et al.,1968). To produce a dark-colored skin, melano-somes migrate from a basal position into the cellu-lar processes involving the iridophores, therebyobscuring the reflecting capability of the brightlycolored pigments within them. During basking incontrast, melanosomes are concentrated aroundthe cell body underneath the iridophores allowingfor light to be reflected by the crystals within theiridophore causing the whitening of the skin.
These responses are obvious in the differencesin the structure of light and dark skin in Boker-mannohyla alvarengai (Fig. 2B,C), for ability ofthis species to alter its skin coloration dramati-cally. We found that the iridophores in the skin ofB. alvarengai are prominent and numerous, form-ing a dense layer of purine with a palisade-likeappearance unique to this species. We speculatethat this arrangement increases cutaneous reflec-tance, which in turn would increase the efficiencyof physiological color change in modulating heatabsorption/reflection. The number of pigment cellsper chromatophore unit varies among anuran spe-cies (Bagnara et al., 1968), but independent oftheir configuration, the ability of iridophores toreflect light provides an excellent basis for temper-ature regulation (Withers et al. 1982; Kobelt andLinsenmair 1986; Schmuck et al., 1988).
Despite the fact that Bokermannohyla alvaren-gai is not considered a “waterproof” species (Tat-tersall et al., 2006), it has a lower rate of EWLthan other nonbasking hylid frogs (e.g., Wygoda,1984); this suggests that mechanisms exist toreduce water loss through evaporation. In thisregard, the presence of both mucosubstances andthe lipoid secretion in the skin of B. alvarengai isthought to assist in the prevention of excessiveEWL by forming an extra-epidermal layer. Last,our biochemical analysis of the skin secretionclearly shows the existence of different lipid com-pounds that can influence thermoregulation andwater balance.
Gas chromatography–mass spectrometry analy-sis demonstrated that the main lipid componentsin the skin secretion of Bokermannohyla alvaren-gai are saturated fatty acids. Nine of the elevenmain peaks revealed in the analysis seem to becomponents of the skin secretion; the remaining
TABLE 1. List of compounds found in Bokermannohylaalvarengai skin secretion, identified by GC–MS
Molecule n8 Compound name RT* (min)
1 1,3-Benzenediol,o-(4-methylbenzoyl)-o’-(2—methoxybenzoyl)-
13.8
2 Phenol, 2,4-bis(1,1-dimethylethyl)- 21.593 Phthalic acid, decyl methyl ester 25.224 Tetradecane,2,6,10-trimethyl- 25.75 Methoxyacetic acid,4-tetradecyl ester 26.26 Trichloroacetic acid, hexadecyl ester 26.67 1-nonene,4,6,8-trimethyl 27.28 1,2-Benzenedicarboxylic acid,
bis(2-methylpropyl) ester29.32
9 Decane, 2,3,5,8-tetramethyl- 29.7510 Hexadecanoic acid, methyl ester 30.3911 Octadecanoic acid, methyl ester 34.09
*RT, Retention time.
8 F.C. CENTENO ET AL.
Journal of Morphology

two, i.e., Peak 3 (phthalic acid, decyl methyl ester)and Peak 8 [1,2-benzenedicarboxylic acid, bis(2-methylpropyl) ester], are common components ofplants (Taskin, 2013; Vimalavady and Kadavul,2013) and are probably contaminants resultingfrom the preparation of the samples. One of themost abundant lipid molecules found in the skinsecretion of B. alvarengai is sphenol, 2,4-bis(1,1-dimethyl)- (Peak 2), which is a chemical interme-diate for the synthesis of UV stabilizers or antioxi-dants. This finding suggests that the skinsecretion of this treefrog may afford some protec-tion against the deleterious effects associated tothe prolonged exposition to solar radiation (Yoonet al., 2006; Gao et al., 2011).
The fatty substances identified in the skin secre-tion of Bokermannohyla alvarengai probably arisein the granular glands that histochemical analysisdemonstrated to be positive to lipids; these maypartially “waterproof” the skin surface. Manyother frogs change the permeability of the skin bysecreting a lipid coating on the body surface. How-ever, the nature and the origin of the lipid secre-tion differ considerably among species (Lillywhite,2006). Lipid secretion in some nonwaterproof spe-cies of the genus Hyla occurs only via granularglands as it does B. alvarengai (Barbeau and Lilly-white, 2005); in contrast, in Polypedates maculatusonly the mucous glands secrete lipid (Lillywhiteet al., 1997). Among waterproof frogs, a specializedtype of gland spread throughout the skin is dedi-cated to lipid production in the genus Phyllome-dusa (Blaylock et al., 1976; McClanahan et al.,1978), whereas in Litoria caerulea, the lipid is pro-duced both in specialized glands and within themucous glands (Christian et al., 1988; Christianand Parry, 1997). Thus, it seems that lipid secre-tion is not limited to a one type of dermal gland;instead, a few different glands may be involved inthe process (Warburg et al., 2000). Independent ofthe secretion pathway, the presence of extra-epidermal lipids seems to be a convergent adaptiveresponse found among several distantly relatedgenera of arboreal frogs inhabiting arid or season-ally arid environments (Barbeau and Lillywhite,2005), as the case in B. alvarengai.
In addition to extra-epidermal lipid substance,Bokermannohyla alvarengai has many large adipo-cytes grouped in some parts of the subcutaneouslayer of the dorsal and the pectoral skin. It hasbeen proposed that these cutaneous droplets oflipid are important for protecting frogs from desic-cation because the permeability of the skin towater is influenced by lipid content (Schmid andBarden, 1965; Toledo and Jared, 1993; Lillywhite,2006). In fact, some waterproof species seem touse an unbroken layer of cutaneous lipids as awaterproofing mechanism (e.g., Amey and Grigg,1995). However, the role of the discontinuous layerof adipocytes observed in B. alvarengai and other
nonwaterproof species is less clear; it has beensuggested to be linked to energy storage (Wygodaet al., 1987). We cannot resolve the relative contri-bution of this adipocyte layer to the maintenanceof water balance versus energy storage at presentbecause the functions are not mutually exclusive.Perhaps both are at play in B. alvarengai,although the discontinuous adipocyte layer wouldcontribute less to skin resistance in this speciesthan in those frogs with a continuous layer.
As observed in other anuran species (Dreweset al., 1977), Bokermannohyla alvarengai has well-developed verrucae hydrophilica in the pelvicregion and a large number of folds in the mentaland pectoral regions; as a result, the entire venteris “roughened” increasing substantially the surfacearea. Consequently, the pronounced developmentof channels throughout the venter combined withthe thinning of the epidermal barrier typical ofthe verrucae undoubtedly facilitate rapid wateruptake by osmotic water absorption from moistsubstrates, moving water in all directions of theskin surface (McClanahan and Baldwin, 1969;Roth, 1973; Baldwin, 1974; Lillywhite and Licht,1974; Toledo and Jared, 1993; Hillyard et al.,1998; Wells, 2007). At present, we have no expla-nation for the function of the small pits dispersedon the verrucae surface of B. alvarengai, but theirlocation on the verrucae suggest that they may beassociated with water absorption. The pits havenot been described before; their anatomical detailsand function clearly deserve further investigation.
The skin of Bokermannohyla alvarengai ishypervascularized, especially in the pelvic region.This condition may enable the distribution ofwater being absorbed in this region to the otherparts of the body (McClanahan and Baldwin,1969; Roth, 1973; Drewes et al., 1977; Navaset al., 2004). Vascularization characterizes the der-mis and capillaries rarely penetrate the epidermis(Lillywhite and Maderson, 1988). However, vascu-larization extends into outermost layer of the skinin B. alvarengai; the rich capillary network ispresent in the epidermis and nearly reaches theskin surface. Given the elaboration of this vascu-lar network in the pelvic region in close associa-tion with the verrucae, it seems logical that thenetwork plays an important role in waterreabsorption.
It has being suggested that the calcified dermallayer in anurans reduces EWL (Elkan, 1968; Tol-edo and Jared, 1993). Thus, the continuous calci-fied layer in the dorsum of Bokermannohylaalvarengai might prevents excessive EWL in thisregion, which usually is exposed, particularly dur-ing basking. In contrast, the discontinuous layerin the ventral skin and its absence in the inguinalregion might enhance water absorption on thesesurfaces. But the calcified layer lies deep to thevascular plexus of the stratum spongiosum; thus it
9BASKING BEHAVIOR IN TREEFROG
Journal of Morphology

seems that it is poorly located to assist in waterconservation (Drewes et al., 1977).
Basking in anurans is assumed to confer a num-ber of thermoregulatory benefits (Brattstrom,1979). This is especially true for anurans thatthrive at moderate to high altitudes wheredecreases in the average ambient temperatureoccur with high levels of solar radiation such as atcampos rupestres where Bokermannohyla alvaren-gai is found. Given these conditions, trade-offsmay occur between thermo- and osmoregulationthat favor the appearance of the basking behaviorin anurans, and in fact, basking is more commonin species that occur at relatively high altitudes(Vences et al., 2002). For one of such species, ourstudy revealed morphological and anatomical fea-tures that are thought to be convenient to accom-modate heat acquisition via basking withoutcompromising water balance.
ACKNOWLEGMENTS
ICMBio furnished the permit to collect the ani-mals (SISBIO, 22170-1/2010). The authors thankParque Nacional da Serra do Cip�o (PNSC) forresearch assistance.
LITERATURE CITED
Angilletta MJ, Niewiarowski PH, Navas CA. 2002. The evolu-tion of thermal physiology in ectotherms. J Thermal Biol 27:249–268.
Amey AP, Grigg GC. 1995. Lipid-reduced evaporative waterloss in two arboreal hylid frogs. Comp Biochem Physiol APhysiol 111:283–291.
Bagnara J, Taylor J, Hadley M. 1968. The dermal chromato-phore unit. J Cell Biol 38:67–79.
Baldwin RA. 1974. The water balance response of the pelvic“patch” of Bufo punctatus and Bufo boreas. Comp BiochemPhysiol A Physiol 47:1285–1295.
Barbeau TR, Lillywhite HB. 2005. Body wiping behaviors asso-ciated with cutaneous lipids in hylid tree frogs of Florida.J Exp Biol 208:2147–2156.
Bentley PJ. 1966. Adaptations of Amphibia to arid environ-ments. Science152:619–623.
Blaylock LA, Ruibal R, Plattaloia K. 1976. Skin structure andwiping behavior of phyllomedusine frogs. Copeia 1976:283–295.
Bogert CM. 1949. Thermoregulation in reptiles, a factor in evo-lution. Evolution 3:195–211.
Brattstrom BH. 1963. A preliminary review of the thermalrequirements of amphibians. Ecology 44:238–255.
Brattstrom BH. 1979. Amphibian temperature regulation stud-ies in the field and laboratory. Am Zool 19:345–356.
Christian K, Parry D. 1997. Reduced rates of water loss andchemical properties of skin secretions of the frogs Litoriacaerulea and Cyclorana australis. Aust J Zool 45:13–20.
Christian K, Parry D, Green B. 1988. Water loss and an extrae-pidermal lipid barrier in the Australian tree frog Litoria caer-ulea. Am Zool 28:17A.
Cowles RB. 1940. Additional implications of reptilian sensitivityto high temperatures. Am Nat 74:542–561.
Cowles RB, Bogert CM. 1944. A preliminary study of the ther-mal requirements of desert reptiles. Bull Am Mus Nat Hist83:261–296.
Drewes RC, Hillman SS, Putnam RW, Sokol OM. 1977. Water,nitrogen and ion balance in the African treefrog Chiromantis
petersi Boulenger (Anura: Rhacophoridae), with commentsonthe structure of the integument. J Comp Physiol B 116:257–267.
Elkan E. 1968. Mucopolysaccharides in the anuran defenceagainst desiccation. J Zool 155:19–53.
Eterovick PA, Sazima I. 2004. Anf�ıbios da Serra do Cip�o, MinasGerais—Brasil. Belo Horizonte, Brasil: PUC Minas. 151 p.
Gao Y, Gu Y, Wei Y. 2011. Determination of polymer additives-antioxidant and ultraviolet (UV) absorbers by high-performance liquid chromatography coupled with UV photo-diode array detection in food simulants. J Agric Food Chem59:12982–12989.
Hillman SS, Withers PC, Drewes RC, Hillyard SD. 2009. Eco-logical and environmental physiology of amphibians. NewYork: Oxford University Press. 464 p.
Hillyard SD, Hoff KS, Propper C. 1998. The water absorptionresponse: A behavioral assay for physiological processes interrestrial amphibians. Physiol Zool 71:127–138.
Hoppe DM. 1979. The influence of color on behavioral thermo-regulation and hydroregulation. In: Burtt EH, editor. TheBehavioral Significance of Color. New York: Garland STPMPress. pp. 37–62.
Hutchinson VH, Maness JD. 1979. The role of behavior in tem-perature acclimation and tolerance in ectotherms. Am Zool19:367–384.
Jameson DL. 1966. Rate of weight loss of tree frogs at varioustemperatures and humidities. Ecology 47:605–613.
Jared C, Antoniazzi MM, Navas CA, Katchburian E,Freym€uller E, Tambourgi DV, Rodrigues MT. 2005. Head co-ossification, phragmosis and defence in the casque-headedtree frog Corythomantis greeningi. J Zool 265:1–8.
Junqueira LCU. 1995. Histology revisited. Technical improve-ment promoted by the use of hydrophilic resin embedding.Cienc Cult (Sao Paulo) 47:92–95.
Karnovsky MJA. 1965. A formaldehyde glutaraldehyde fixativeof high osmolarity for use in electron microscopy. J Cell Biol27:137.
King RB, Hauff S, Phillips JB. 1994. Physiological color changein the green treefrog: responses to background brightnessand temperature. Copeia 1994:422–432.
Kobelt F, Linsenmair KE. 1986. Adaptations of the reed frogHyperolius viridiflavus (Amphibia, Anura, Hyperoliidae) toits arid environment. 1. The skin of Hyperolius viridiflavusnitidulus in wet and dry season conditions. Oecologia 68:533–541.
K€oppen W. 1900. Versuch einer klassifikation der klimate, vor-zugsweise nach ihren beziehungen zur pflanzenwelt. Geogr Z6:593–611.
Lillywhite HB. 1970. Behavioural temperature regulation inthe bullfrog, Rana catesbeiana. Copeia 1970:158–168.
Lillywhite HB. 1971. Temperature selection by the bullfrog,Rana catesbeiana. Comp Biochem Physiol A Physiol 40:213–227.
Lillywhite HB. 2006. Water relations of tetrapod integument.J Exp Biol 209:202–226.
Lillywhite HB, Licht P. 1974. Movement of water over toadskin: Functional role of epidermal sculpturing. Copeia 1974:165–171.
Lillywhite HB, Maderson PFA. 1988. Structure and permeabil-ity of integument. Am Zool 28:945–962.
Lillywhite HB, Licht P, Chelgren P. 1973. The role of behavioralthermoregulation in the growth energetics of the toad, Bufoboreas. Ecology 54:375–383.
Lillywhite HB, Mittal AK, Garg TK, Agrawal N. 1997. Wipingbehavior and its ecophysiological significance in the IndianTree Frog Polypedates maculatus. Copeia 1997:88–100.
McClanahan LL, Baldwin R. 1969. Rate of water uptakethrough the integument of the desert toad, Bufo punctatus.Comp Biochem Physiol 28:381–389.
McClanahan LL, Stinner JW, Shoemaker VH. 1978. Skin lipids,water loss and energy metabolism in a South American treefrog (Phyllomedusa sauvagei). Physiol Zool 51:179–187.
10 F.C. CENTENO ET AL.
Journal of Morphology

Muths E, Corn PS. 1997. Basking by adult boreal toads (Bufoboreas boreas) during the breeding season. J Herpetol 31:426–428.
Navas CA, Antoniazzi MM, Jared C. 2004. A preliminaryassessment of anuran physiological and morphological adap-tation to the Caatinga, a Brazilian semiarid environment. In:Morris S, Vosloo A, editors. Animals and Environments. Pro-ceedings of the Third International Conference of Compara-tive Physiology and Biochemistry, Vol. 1275. Cambridge:Elsevier. pp 298–305.
Roth JJ. 1973. Vascular supply to the ventral pelvic region ofanurans as related to water balance. J Morphol 140:443–460.
Sazima I, Bokermann WCA. 1977. Anf�ıbios da Serra do Cip�o,Minas Gerais, Brasil. 3, Observac~oes sobre a biologia de Hylaalvarengai Bok. (Anura, Hylidae). Rev Bras Biol 37:413–417.
Schmid WD, Barden RE. 1965. Water permeability and lipid con-tent of amphibian skin. Comp Biochem Physiol 15:423–427.
Shoemaker VH, Hillman SS, Hillyard SD, Jackson DC,McClanahan LL, Withers PC, Wygoda ML. 1992. Exchange ofwater, ions, and respiratory gases in terrestrial amphibians.In: Feder ME, Burggren WW, editors. Environmental physiol-ogy of the amphibians. Chicago: The University of ChicagoPress. pp 125–150.
Schmuck R, Kobelt F, Linsenmair KE. 1988. Adaptations of thereed frog Hyperolius viridiflavus (Amphibia: Anura: Hypero-liidae) to its arid environment: V. Iridophores and nitrogenmetabolism. J Comp Physiol B 158:537–546.
Stegen JC, Gienger CM, Sun L. 2004. The control of colorchange in the pacific tree frog, Hyla regilla. Can J Zool 82:889–896.
Taskin H. 2013. Detection of volatile aroma compounds ofMorchella by headspace gas chromatography mass spectrome-try (HS-GC/MS). Not Bot Hort Agrobot 41:122–125.
Tattersall GJ, Eterovick PC, Andrade DV. 2006. Tribute to R.G. Boutilier: Skin color and body temperature changes in
basking Bokermannohyla alvarengai (Bokermann 1956).J Exp Biol 209:1185–1196.
Toledo RC, Jared C. 1993. Cutaneous adaptations to water bal-ance in amphibians. Comp Biochem Physiol A Physiol 105:593–608.
Tracy CR, Christian KA, O’Connor MP, Tracy CR. 1993. Behav-ioral thermoregulation by Bufo americanus: The importanceof the hydric environment. Herpetologica 49:375–382.
Vences M, Gal�an P, Vieites DR, Puente M, Oetter K, Wanke S.2002. Field body temperatures and heating rates in a mon-tane frog population: The importance of black dorsal patternfor thermoregulation. Ann Zool Fennici 39:209–220.
Vimalavady A, Kadavul K. 2013. Phytocomponents identifiedon the various extracts of stem of Hugonia mystax L. (Lina-ceae). Eur J Exp Biol 3:73–80.
Yoon M, Jeong T, Park D, Xu M, Oh H, Song K, Lee WS, ParkH. 2006. Antioxidant effects of quinolone alkaloids and 2,4-di-tert-butylphenol isolated from Scolopendra subspinipes. BiolPharm Bull 29:735–739.
Warburg MR, Rosenberg M, Roberts JR, Heatwole H. 2000.Cutaneous glands in the Australian hylid Litoria caerulea(Amphibia, Hylidae). Anat Embryol 201:341–348.
Wells KD. 2007. The ecology and behavior of amphibians. Chi-cago: University of Chicago Press. 1400 p.
Withers PC. 1995. Evaporative water loss and color change inthe Australian desert tree frog Litoria rubella (Amphibia:Hylidae). Rec West Austr Mus 17:277–281.
Withers PC, Hillman SS, Drewes RC, Sokol OM. 1982. Waterloss and nitrogen excretion in sharp nosed reed frogs (Hypero-lius nasutus – Anura, Hyperoliidae). J Exp Biol 97:335–343.
Wygoda ML. 1984. Low cutaneous evaporative water loss inarboreal frogs. Physiol Zool 57:329–337.
Wygoda ML, Garman RH, Howard CE. 1987. Cutaneous andsubcutaneous adipose tissue in anuran amphibians. Copeia1987:1031–1035.
11BASKING BEHAVIOR IN TREEFROG
Journal of Morphology