interlamellar ca1 network in the hippocampus
TRANSCRIPT

Interlamellar CA1 network in the hippocampusSunggu Yanga,1, Sungchil Yangb,1, Thais Moreiraa, Gloria Hoffmanc, Greg C. Carlsond, Kevin J. Benderb,Bradley E. Algere, and Cha-Min Tanga,e,f,2
Departments of aNeurology and ePhysiology, University of Maryland School of Medicine, Baltimore, MD 21201; bCenter for Integrative Neuroscience,Department of Neurology, University of California, San Francisco, CA 94608; cDepartment of Biology, Morgan State University, Baltimore, MD 21251;fBaltimore Veterans Affairs Medical Center, Baltimore, MD 21201; and dCenter for Neurobiology and Behavior, University of Pennsylvania, Philadelpia,PA 19104-3403
Edited* by Susumu Tonegawa, Massachusetts Institute of Technology, Cambridge, MA, and approved July 25, 2014 (received for review March 28, 2014)
To understand the cellular basis of learning and memory, theneurophysiology of the hippocampus has been largely examinedin thin transverse slice preparations. However, the synaptic archi-tecture along the longitudinal septo-temporal axis perpendicular tothe transverse projections in CA1 is largely unknown, despite itspotential significance for understanding the information processingcarried out by the hippocampus. Here, using a battery of powerfultechniques, including 3D digital holography and focal glutamateuncaging, voltage-sensitive dye, two-photon imaging, electrophys-iology, and immunohistochemistry, we show that CA1 pyramidalneurons are connected to one another in an associational and well-organized fashion along the longitudinal axis of the hippocampus.Such CA1 longitudinal connections mediate reliable signal transferamong thepyramidal cells andexpress significant synaptic plasticity.These results illustrate a need to reconceptualize hippocampal CA1network function to include not only processing in the transverseplane, but also operations made possible by the longitudinal net-work. Our data will thus provide an essential basis for future com-putational modeling studies on information processing operationscarried out in the full 3D hippocampal network that underlies itscomplex cognitive functions.
longitudinal axons | CA1 association fibers | sequence memory | DHR
The hippocampus is widely used to study functional connectivityof the brain with the hope that principles that operate within
its relatively simple architecture may be extended to more com-plex cortical structures. Its manageable number of cell types alsoprovides an attractive opportunity to examine fundamental issuesin neuroscience such as the relationship between network circuitryand function. For example, considerable effort has been devotedtoward elucidating the circuitry supporting episodic memory—a property closely linked to the hippocampus. CA3 pyramidalneurons form extensive recurrent connections with each other (1).Such connections are able to learn to associate components of aninput pattern with each other (2), which, in turn, has greatlyinfluenced thinking on the mechanisms of memory formation andrecall (3). Under appropriate conditions, computer simulationsreveal that recurrent neural networks have the capacity to learntemporal sequences and to carry out pattern completion (4, 5).Interestingly, although they are quite near to the CA3 region, CA1pyramidal neurons reportedly form remarkably few associationalconnections (6, 7). This distinctive difference in network archi-tecture might suggest that, although area CA1 could serve todecode the output of CA3, it would not possess the intrinsic abilityfor autoassociational computations. This idea would imply that theability of CA1 to carry out independent information processingoperations may be more limited than that of CA3. However, evenafter removal of all input from area CA3, CA1 pyramidal neuronsstill have the capacity to transform location-modulated signalsfrom the entorhinal cortex into accurate spatial firing patterns (8).In addition, deficits in temporal sequence learning are more se-vere after selective lesions to CA1 than to CA3 (9). Finally, CA1 ismore closely linked to memory of temporal order of visual objectsand especially over long intervals (10). Thus, area CA1 appears tohave a greater ability for intrinsic information processing than
would be expected based on current understanding of its circuitry.Intrinsic processing could represent autoassociational computa-tions through direct excitatory synaptic contacts among the CA1pyramidal cells, but as noted, there is little evidence for suchconnectivity within CA1. This puzzle led us to reexamine the ap-parent sparseness of associational synaptic connections betweenCA1 pyramidal neurons using experimental techniques that werenot previously available for this investigation.The “trisynaptic circuits” (dentate gyrus: CA3–CA1) oriented
transversely to the hippocampal long axis, the basis of the “lamellarhypothesis” (11), has greatly influenced thinking about the struc-ture-function relationships of this structure. This hypothesis sug-gests that the hippocampus is organized as a stack of parallel,trisynaptic circuits. Although this view has been challenged by theobservation of fibers running across lamellae, especially in dentategyrus and CA3 area (12, 13), the hypothesis supported an explosionin the use of the transverse slice for electrophysiological studies ofthe hippocampus. However, axons oriented along the longitudinalaxis are unavoidably severed in the preparation of the transverseslice, meaning that these studies are heavily weighted in favor ofconclusions based on fibers traveling within the transverse plane.We used a whole hippocampus preparation, as well as longitudinaland transverse slice preparations, to obtain a more accurate pictureof synaptic connections among CA1 pyramidal neurons in threedimensions. Remarkably, we found prominent associational con-nectivity along the longitudinal axis. Furthermore, synapses of thelongitudinal network possess the capacity for synaptic plasticity that
Significance
It has generally been thought that CA1 cells form only negli-gible connections with each other along the longitudinal axisof the hippocampus. But if CA1 cells were interconnected in aneffective autoassociational network, this information wouldadd a critical new dimension to our understanding of cellularprocessing within this structure. Here, we report the existenceof a well-organized, longitudinally projecting synaptic networkamong CA1 pyramidal neurons. We further show that synapsesof this network are capable of supporting synaptic plasticity,including long-term potentiation, and a short-term memorymechanism called “dendritic hold and read.” These observa-tions will contribute to the construction of more realisticmodels of hippocampal information processing in behavior,memory, and other cognitive functions.
Author contributions: Sunggu Yang and C.-M.T. designed research; Sunggu Yang,Sungchil Yang, T.M., and G.H. performed research; G.C.C. contributed new reagents/an-alytic tools; Sunggu Yang, Sungchil Yang, G.C.C., and K.J.B. analyzed data; and SungguYang, B.E.A., and C.-M.T. wrote the paper.
The authors declare no conflict of interest.
*This Direct Submission article had a prearranged editor.
Freely available online through the PNAS open access option.1Sunggu Yang and Sungchil Yang contributed equally to the work.2To whom correspondence should be addressed. Email: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1405468111/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1405468111 PNAS | September 2, 2014 | vol. 111 | no. 35 | 12919–12924
NEU
ROSC
IENCE

includes a novel memory mechanism we recently described calleddendritic hold and read (DHR) (14). These findings may help toexplain the intrinsic ability of area CA1 to process informationtransfer and provide novel data that will lead to more realisticmodels of hippocampal function in three dimensions.
ResultsLongitudinal Connections Between CA1 Pyramidal Neurons. The ax-onal arborization of individual CA1 pyramidal neurons was exam-ined by two-photon fluorescence imaging of cells filled with AlexaFluor 594 dye using themouse whole hippocampal preparation. Thispreparation maintains intact axonal processes and, thereby, permitsfollowing and reconstructing axonal processes in all dimensions. Wefound that the axonal processes were multiply branched and that, infact, all CA1 pyramidal neurons examined (n = 9/9) had a distinctprojection along the longitudinal axis. These processes are orientedperpendicularly to the transverse hippocampal lamellae (marked inred and with arrows in Fig. 1 A and B) and are readily distinguishedfrom previously described local associational processes that ramifywith a transverse orientation (6, 7, 15) (marked in green in Fig. 1).The longitudinal branch arises from the thick proximal axon thatgives rise to the transversely oriented axons (Fig. 1 and Fig. S1B). Allof the cells send a projection toward the subiculum; a branchheading to the fimbria, first described by Cajal (1911) (16), wasobserved in 8 of the 9 cells (Fig. 1 B and C). These subicular andfimbrial branches are consistent with the lamellar organizationalprinciple (11); however, the extensive and organized projection oflongitudinally projecting branches may be at variance with it (Fig.1C), depending on the actual physiological influence that thesebranches exert on their target cells (see ref. 13 for discussion).If these morphologically identified structures are part of an
operationally significant associational network of CA1 pyramidalcells, they should form excitatory synapses onto other CA1 pyra-midal cells. To examine the morphology and electrophysiology ofthe longitudinal network in more detail, a separate set of studieswas conducted by filling single cells with dye in the longitudinalhippocampal slice preparation (Fig. 2A). With two-photon imag-ing, we observed that the CA1 pyramidal neuron sends a thickprimary axon (PA; Fig. 2B, red arrow and Fig. S1) toward the
alveus, presumably heading for either the subiculum or the fimbria.A thin collateral arose from the primary axon and projected lon-gitudinally (Fig. 2B, red arrow and Fig. S1). The branch point waslocated on average 154 ± 30 μm from the cell body (n = 7), and thethin branches were followed in the longitudinal plane for up to 375± 22 μm (n = 7). Both PAs and LAs were studded with varicositiespresumed to represent presynaptic terminals. The longitudinalaxon (LA) either projected toward the ventral-temporal (referredto as “temporal”) end (n = 2/7) or toward the dorsal-septal(“septal”) end (n = 5/7). Similar results were obtained with re-construction of biocytin-labeled cells (Fig. 2C and Fig. S2). Thethick PAs were truncated in the longitudinal slice and appeared asshort stumps. They were best observed in slices made at an angle of45° with respect to the longitudinal axis (Fig. S1). LAs were signifi-cantly thinner than PAs, whereas the varicosity density per unitlength of LAs was higher than that of the PAs (Fig. S3). If the var-icosities actually represent sites of synaptic contact, then they shouldshow prominent voltage-gated calcium signals when the axons arestimulated. To test this prediction, we imaged the varicosities incalcium-sensitive, dye-filled cells and measured action potential-evoked calcium responses within varicosities on longitudinalbranches (white arrows in Fig. 2 B and D). The calcium responseswere reliablypropagated along the varicosities for up to 456 μmfromthe cell body without substantial decrement, demonstrating that theresponses were induced by propagating action potentials and not bypassive depolarizations from the neuronal somata (in which case theresponses would have declined monotonically because of the elec-trotonic decay of the depolarization) (Fig. S4).Hence, it is likely thatthe axonal varicosities are presynaptic transmitter release sites.It has been proposed that the previously described sparse as-
sociational projections to neighboring transverse lamellae inCA1 are primarily directed onto inhibitory interneurons (17, 18).To investigate whether the associational axonal processes de-scribed here represent excitatory-excitatory or excitatory-inhibitoryconnections, we looked for immunohistochemical evidence ofcontact with synaptic spines of pyramidal neurons. Spinophilin isexclusively localized at spine heads of hippocampal pyramidalneurons (19), and spines are rarely foundon inhibitory interneurons(some numerically small interneuronal subtypes are exceptions).
Fig. 1. Two-photon imaging of longitudinal axons in the whole hippocampal preparation. (A) The CA1 pyramidal cell region is viewed from above the hip-pocampus in the axis of the main apical trunk. A Neurolucida drawing of the axonal arborization of a CA1 pyramidal neuron is placed adjacent to the rawinverted fluorescence image. Higher magnification of parts of the fluorescence image is provided in Lower Inset. The longitudinal axons in the fluorescenceimages are identified by arrows. The relative position of the cell within the hippocampus is shown in Upper Inset. The four distinct parts of the axonal arbor arecolor coded. The longitudinal axon (red) can be distinguished from the local associational branches (green) by its perpendicular orientation to the main externalprojecting axons of the cell. (B) Example of another CA1 pyramidal neuron with a prominent longitudinal axon. The portion of its axonal process that projects tothe fimbria in this neuron was not found. (C) Summary of two-photon imaging demonstrating the organized pattern of longitudinal and projecting axons.
12920 | www.pnas.org/cgi/doi/10.1073/pnas.1405468111 Yang et al.
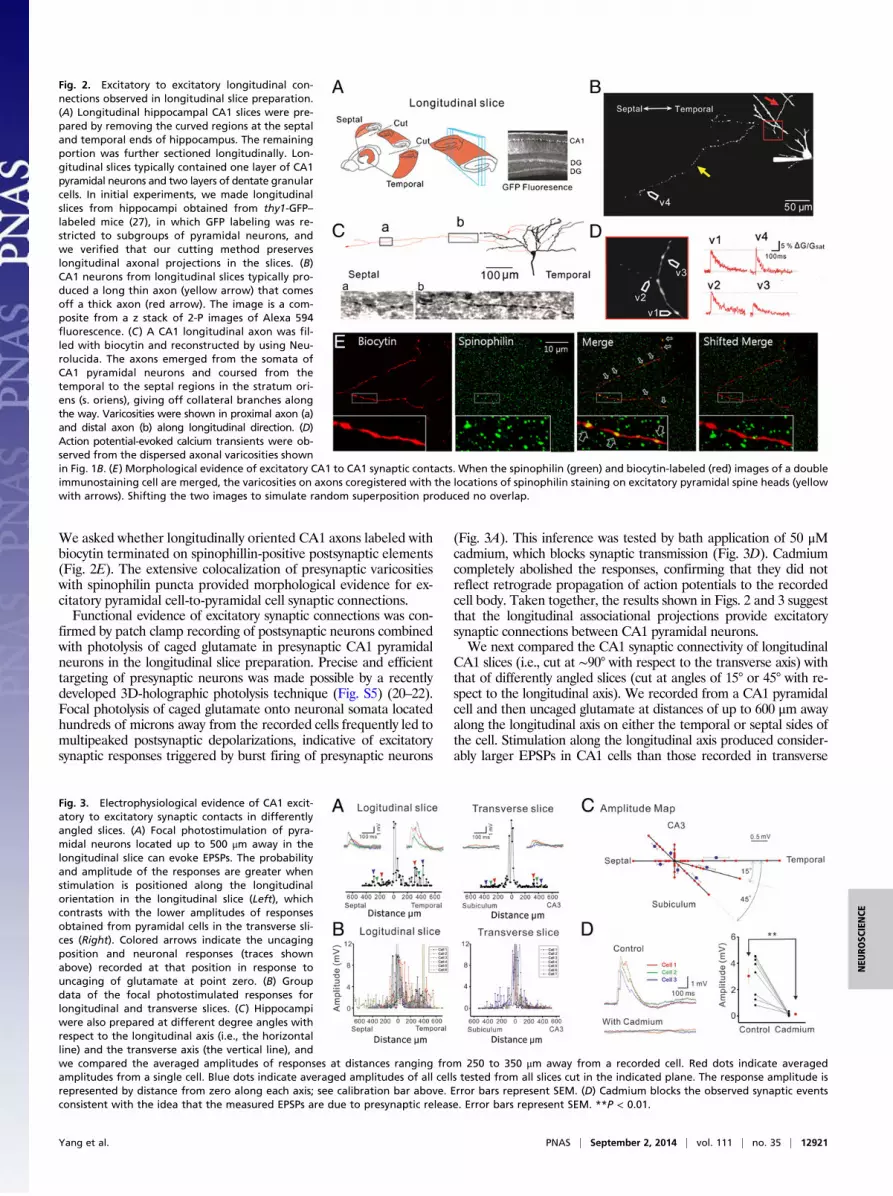
We asked whether longitudinally oriented CA1 axons labeled withbiocytin terminated on spinophillin-positive postsynaptic elements(Fig. 2E). The extensive colocalization of presynaptic varicositieswith spinophilin puncta provided morphological evidence for ex-citatory pyramidal cell-to-pyramidal cell synaptic connections.Functional evidence of excitatory synaptic connections was con-
firmed by patch clamp recording of postsynaptic neurons combinedwith photolysis of caged glutamate in presynaptic CA1 pyramidalneurons in the longitudinal slice preparation. Precise and efficienttargeting of presynaptic neurons was made possible by a recentlydeveloped 3D-holographic photolysis technique (Fig. S5) (20–22).Focal photolysis of caged glutamate onto neuronal somata locatedhundreds of microns away from the recorded cells frequently led tomultipeaked postsynaptic depolarizations, indicative of excitatorysynaptic responses triggered by burst firing of presynaptic neurons
(Fig. 3A). This inference was tested by bath application of 50 μMcadmium, which blocks synaptic transmission (Fig. 3D). Cadmiumcompletely abolished the responses, confirming that they did notreflect retrograde propagation of action potentials to the recordedcell body. Taken together, the results shown in Figs. 2 and 3 suggestthat the longitudinal associational projections provide excitatorysynaptic connections between CA1 pyramidal neurons.We next compared the CA1 synaptic connectivity of longitudinal
CA1 slices (i.e., cut at ∼90° with respect to the transverse axis) withthat of differently angled slices (cut at angles of 15° or 45° with re-spect to the longitudinal axis). We recorded from a CA1 pyramidalcell and then uncaged glutamate at distances of up to 600 μm awayalong the longitudinal axis on either the temporal or septal sides ofthe cell. Stimulation along the longitudinal axis produced consider-ably larger EPSPs in CA1 cells than those recorded in transverse
Fig. 2. Excitatory to excitatory longitudinal con-nections observed in longitudinal slice preparation.(A) Longitudinal hippocampal CA1 slices were pre-pared by removing the curved regions at the septaland temporal ends of hippocampus. The remainingportion was further sectioned longitudinally. Lon-gitudinal slices typically contained one layer of CA1pyramidal neurons and two layers of dentate granularcells. In initial experiments, we made longitudinalslices from hippocampi obtained from thy1-GFP–labeled mice (27), in which GFP labeling was re-stricted to subgroups of pyramidal neurons, andwe verified that our cutting method preserveslongitudinal axonal projections in the slices. (B)CA1 neurons from longitudinal slices typically pro-duced a long thin axon (yellow arrow) that comesoff a thick axon (red arrow). The image is a com-posite from a z stack of 2-P images of Alexa 594fluorescence. (C) A CA1 longitudinal axon was fil-led with biocytin and reconstructed by using Neu-rolucida. The axons emerged from the somata ofCA1 pyramidal neurons and coursed from thetemporal to the septal regions in the stratum ori-ens (s. oriens), giving off collateral branches alongthe way. Varicosities were shown in proximal axon (a)and distal axon (b) along longitudinal direction. (D)Action potential-evoked calcium transients were ob-served from the dispersed axonal varicosities shownin Fig. 1B. (E) Morphological evidence of excitatory CA1 to CA1 synaptic contacts. When the spinophilin (green) and biocytin-labeled (red) images of a doubleimmunostaining cell are merged, the varicosities on axons coregistered with the locations of spinophilin staining on excitatory pyramidal spine heads (yellowwith arrows). Shifting the two images to simulate random superposition produced no overlap.
Fig. 3. Electrophysiological evidence of CA1 excit-atory to excitatory synaptic contacts in differentlyangled slices. (A) Focal photostimulation of pyra-midal neurons located up to 500 μm away in thelongitudinal slice can evoke EPSPs. The probabilityand amplitude of the responses are greater whenstimulation is positioned along the longitudinalorientation in the longitudinal slice (Left), whichcontrasts with the lower amplitudes of responsesobtained from pyramidal cells in the transverse sli-ces (Right). Colored arrows indicate the uncagingposition and neuronal responses (traces shownabove) recorded at that position in response touncaging of glutamate at point zero. (B) Groupdata of the focal photostimulated responses forlongitudinal and transverse slices. (C) Hippocampiwere also prepared at different degree angles withrespect to the longitudinal axis (i.e., the horizontalline) and the transverse axis (the vertical line), andwe compared the averaged amplitudes of responses at distances ranging from 250 to 350 μm away from a recorded cell. Red dots indicate averagedamplitudes from a single cell. Blue dots indicate averaged amplitudes of all cells tested from all slices cut in the indicated plane. The response amplitude isrepresented by distance from zero along each axis; see calibration bar above. Error bars represent SEM. (D) Cadmium blocks the observed synaptic eventsconsistent with the idea that the measured EPSPs are due to presynaptic release. Error bars represent SEM. **P < 0.01.
Yang et al. PNAS | September 2, 2014 | vol. 111 | no. 35 | 12921
NEU
ROSC
IENCE

slices with glutamate uncaging given on either the transverse side(Fig. 3 B and C; longitudinal, 0.94 ± 0.23 mV, n = 19 slices vs.transverse, 0.27 ± 0.05 mV, n = 10 slices, χ2 =6.78, P = 0.009,Kruskal–Wallis ANOVA). The mean amplitudes of the responsesevoked by photolysis at different distances are plotted as a functionof the slice orientation in a group of cells (Fig. 3C). In Fig. 3C, re-sponse amplitudes are represented by distances along the given axis(either 0° = longitudinal, 15°, 45°, or 90° = transverse). A red dotrepresents the mean amplitude of a number of responses froma single cell located from 250 to 350 μmaway from the uncaging spotin a slice of the indicated orientation. A blue dot indicates the meanamplitude of the responses from all of the cells (all 250–350 μmawayfrom the uncaging spot) recorded in slices of the given orientation.Comparison of the blue dots shows that the largest responses areelicited in the longitudinal slices (horizontal line in Fig. 3C).To examine signaling along the longitudinal axis at the network
level we used voltage-sensitive dye imaging in the whole hippo-campus preparation (Fig. 4A). A stimulating electrode was posi-tioned in the superficial alvear field of the CA1 region. A singlestimulation evoked the expected early voltage signal (within 10ms),propagating within the transverse lamina toward the fimbria (Fig.4B) and the subiculum (Fig. 4C). A slower longitudinal signalemerged that peaked at ∼25 ms after stimulus and was gone by∼35 ms. Importantly, the longitudinal signal in CA1 was not sec-ondary to longitudinal activation in CA3. A stimulus train (fivestimuli at 100 Hz) evoked a similar early depolarization within thetransverse lamina but evoked a stronger and longer lasting de-polarization in the longitudinal axis between laminas. The groupdata for the spatial activation profile is shown in Fig. 4C (n = 5).These optical recordings further demonstrate the existence of arobust and functional associative excitatory network in region CA1.
Cellular Memory Mechanisms of the Interlamella CA1 Network. Thepotential functional roles carried out by the longitudinal path-way will depend in part on the physiological properties of the
synapses formed on the target cells. We first examined theirability to support plastic mechanisms such as long-term poten-tiation (LTP). Application of high frequency stimulation (HFS)in longitudinal slices resulted in a robust LTP to 210.7 ± 3.6% ofthe control value (n = 5 slices; Fig. 5B) along the course of thelongitudinally oriented fibers. This data suggests that LTP canstrengthen interlamellar CA1-to-CA1 connections as well as thewell-established CA3-to-CA1 connections in the transverse plane.We next examined whether the synapses of the interlamellar
network can express DHR (14). DHR was previously demon-strated to occur on apical oblique dendrites (14). Because theinterlamellar network forms synapses mainly on basal dendrites,we tested for DHR on basal dendrites in the longitudinal slicepreparation (Fig. 5A). Photolysis of caged glutamate was directedonto select CA1 basal dendrites to create the bound-but-blockedstate of the NMDAR (blue arrow and blue circles in Fig. 5 A, i).This uncaging simulates the effects of temporally and spatiallydispersed EPSPs arriving on the dendrite, which are unable toproduce a depolarization large enough to remove the Mg2+ blockof the NMDARs. Next, a stronger second stimulus (“gatingstimulus”, purple arrows and circles) that, by itself, would produceonly a modest depolarization, was delivered to the same dendrite.However, if the gating stimulus is given within a several hundred-millisecond delay after the priming stimulus, its response ismarkedly potentiated, because the NMDARs put into the bound-but-blocked state by the priming stimulus become unblocked bythe gating stimulus and initiate a regenerative local dendritic spike(Fig. 5 A, i). In essence a molecular memory of previous synapticactivity is stored in the bound-but-blocked state and is “read out”by the gating stimulus in the form of a larger gating response.Previously, DHR was only demonstrated with photolysis of caged
glutamate to prime the dendrite. We next sought to determinewhether synaptically induced EPSPs produced by the longitudinalaxons could also act in the priming role. To test this, we placeda stimulating electrode in s.oriens temporal to a CA1 neuronrecorded under whole-cell current clamp conditions in a longitudi-nal slice (Fig. 5 A, ii). A moderately strong photolysis-mediatedgating stimulus (red circles) was directed onto a target basal den-drite alone or 200 ms after priming by synaptic stimulation (Fig. 5A, ii Top). The photolytic gating response was significantly poten-tiated if it occurred after the priming EPSP (control: 4.27 ± 0.54mV vs. DHR: 7.17 ± 0.73 mV; paired t test; n = 5) (Fig. 5 A, iiMiddle). The potentiated local dendritic spike could subsequentlydrive a somatic action potential even when the gating depo-larization alone without priming could not (Fig. 5 A, ii Bottom).This data are consistent with the idea that the CA1 interlamellarnetwork can support DHR mediated memory mechanisms.
DiscussionThe experiments described here provide anatomical, electrophysi-ological, and electro-optical evidence of the existence of well-or-ganized associational connections among CA1 pyramidal neurons.Two-photon imaging of intracellularly labeled CA1 pyramidalneurons shows that every CA1 pyramidal neuron that was examinedsends out a longitudinally oriented axon that branches off, and isperpendicular to, the thick, well-established projection that goestoward the subiculum and fimbria. The subicular and fimbrial ax-onal projections agree with established data (17), but the longitu-dinally projecting collateral has not been described. Note thatalthough we use the term “longitudinal” as others have, that thisterm was done partly for convenience: The most favorable orien-tation for excitatory effects was in the longitudinal plane; however,stimulation at angles oblique to the longitudinal plane was alsoquite effective in activating CA1 pyramidal cells. The delineationbetween transverse and longitudinal is not sharp, but rather a mat-ter of degree in CA1, as it is in CA3. The synaptic contacts made bythe longitudinal projections are excitatory, as evidenced by immu-nostaining and electrophysiology. It is likely that the thinner axons
Fig. 4. The longitudinal propagation of electrical signals in area CA1 in thewhole hippocampus preparation. (A) The stimulating electrode was placedin the alveus over area CA1. The image of voltage-sensitive dye (VSD) signalsis shown in Right and represents the average of 500 individual frames. (B) VSDresponses to a single stimulation (Upper) and multiple stimulation (five stimuliat 100 Hz) (Lower) are shown as snapshots of activation at 3, 10, 25, and 35ms.(C) Group data of the spatial imaging profile of VSD to CA1 activation illus-trate signal propagation along transverse and longitudinal direction.
12922 | www.pnas.org/cgi/doi/10.1073/pnas.1405468111 Yang et al.

and their orientation with respect to the canonical transverse or-ganization of hippocampal architecture led to this pathway beingoverlooked in earlier studies. Despite its more limited arborization,its ability to affect potent excitatory signaling is clearly evident fromvoltage sensitive dye imaging. The imaging results show that lon-gitudinal spread of excitation in CA1 does not require precedingCA3 excitation. Furthermore, the CA1 associational network sup-ports forms of synaptic plasticity, such as LTP, as well as DHR.These properties suggest that this system may be an integral com-ponent of the larger 3D information processing network ofthe hippocampus.
Comparison of CA1 and CA3 Associational Connection. Although weshow here that CA1, as well as CA3 and the dentate gyrus, pos-sesses associational connections, substantial differences amongthem suggest that they probably serve different functions. The CA3and dentate mossy cell projections are extensive, and their signals,which propagate relatively rapidly (e.g., Fig. S6), could recruit largepopulations of target cells (either inhibitory or excitatory) andaffect wide areas of the hippocampus in concert. The extensiveCA3 recurrent connections may be suitable for dealing with largeinformation transfer and mediate feedback memory mechanisms.The relatively restricted projection of the CA1 autoassocia-
tional network suggests a more focused targeting, perhaps in-volving coordinated activity within smaller modules of cells. TheCA1 associational axons are shorter in length than the CA3axons and their signals propagate with significant time delays(Fig. 4). These characteristics would allow the CA1 system toparticipate in information processing functions that the largerCA3 and mossy cell longitudinal association systems are not ca-pable of carrying out. For example, during exploratory behavior orrapid eye movement sleep, traveling theta waves propagate lon-gitudinally throughout the entire extent of the CA1 region of theintact hippocampus, but with a 180° phase shift from the septal tothe temporal poles (23). Similar phase precession is a hallmark ofhippocampal place cell behavioral learning in rats learning to runamaze (24). Although the anatomical substrate for traveling thetawave propagation is unknown, Patel et al. (23) speculate thata longitudinal chain of “weakly coupled oscillators” could accountfor their observations. This idea would be an example of the typeof processing that the slower and more spatially restricted axonalconnections in the CA1 longitudinal system seem well suited toserve; whether this network is actually involved in propagation oftraveling theta waves is of course unknown at this point.Alternatively, the limited CA1 associational connections could
theoretically form a delay-line network comprising a linear chainof serially connected, nearly identical CA1 pyramidal neurons(14). A unique feature of such linear chains is that they cantransform a time sequence into a spatial sequence as informationis iteratively passed from one cell to its downstream neighbors.
The resulting “time-to-space” transform is effectively an efficientsequence memory mechanism, like the shift register that exists atthe input stage of processors of all modern computers to serve asinput memory buffers (25) Computers need memory buffersbecause information typically arrives as a time sequence overa finite interval and its meaning can only be revealed if eachsequence is viewed as a whole and in proper order. Whether thebrain uses a delay-line type circuit for sequence memory is notknown. However, behavioral studies have suggested that areaCA1 possesses the capacity for sequence memory (9, 10). Theresults here suggest that the CA1 longitudinal associationalnetwork possess two necessary requisites for a delay-line typememory. First, it contains linear chains of similar CA1 neurons.Second, the basal dendrites that participate in this associationalnetwork possess a robust newly described short-term memoryand conditional retrieval mechanism, DHR (Fig. 5A). DHRoperating on basal dendrites can function in a manner similarlyto digital flip-flops in the shift registers of a computer. Directdemonstration of delay-line memory in the brain will have towait for video imaging studies involving large populations offluorescently labeled neurons in behaving animals.Which of these or other phenomena are mediated by the CA1
longitudinal system will have to be addressed in future work.Perhaps most importantly, it should be kept in mind that theassociational systems in all of the hippocampal subregions arepart of a “three-dimensional series of connections,” in whichdivergence and convergence are major organizational principles(18). Amaral and Lavenex (18) suggest further that the prom-inent associational connections within hippocampal subregionsconstitutes a substrate that allows each subregion to act “semi-independently from, as well as in concert with” other regions.The deeper significance of our observations may be that the CA1region, previously thought to be incapable of taking part insepto-temporal processing operations, can now, by virtue of itsown, distinctive longitudinal associational network, be recog-nized as a potential key partner in them.
Relevance for the Lamellar Hypothesis of Hippocampal Architecture.As noted, longitudinal interconnections are well known to existamong CA3 pyramidal cells and dentate gyrus mossy cells, butthe likelihood of significant interconnections among CA1 cells hasbeen considered “negligible” (13). Does their existence necessarilyconflict with the lamellar hypothesis of hippocampal organizationput forward by Andersen et al. (11)? Amaral and Witter (12) arguethat, indeed, the longitudinal connectivity directly contradicts thelamellar hypothesis. In contrast, Sloviter and Lømo (13) argue thatit is what the longitudinal connections actually do, their functionalimpact, that is the most important factor, and that a lamella shouldnot be defined in strictly anatomical terms. For example, if thelongitudinal projections were mainly targeted to inhibitory
Fig. 5. Short-term and long-term memory mecha-nisms of the CA1 longitudinal network. (A, i) Basaldendrites of CA1 pyramidal neurons can support DHR.A low level of glutamate is first diffusely released byphotolysis on a single basal dendrite to create a pop-ulation of NMDA receptors in the “bound-but-blocked” state. The timing and location of this“priming” stimulus are marked by light blue arrowsand circles, respectively. A second independent stimu-lus is provided to produce a moderate local de-polarization. The timing and location of this “gating”stimulus aremarked by the purple arrows and circles. Ifthe gating stimulus was coupled to the priming stim-ulus within an interval of hundreds of milliseconds,a local dendritic spike was triggered. (A, ii) Electricalstimulation of the longitudinally oriented axons can also prime basal dendrites for DHR. The local dendritic spike produced by DHR of a single dendrite can thentrigger a somatic action potential (lower trace). (B) LTP can be induced in synapses of the longitudinal network in response to a brief high frequency stimulus (100 Hz).
Yang et al. PNAS | September 2, 2014 | vol. 111 | no. 35 | 12923
NEU
ROSC
IENCE

interneurons (26), then excitation of principal cells would createzones of lateral inhibition that would, in effect, sharpen the func-tional borders of the lamellae and support, rather than contradict,the lamellar hypothesis (13). Alternatively, they suggest that longi-tudinal connections could simply serve to coordinate activity acrosslamellae. Although not part of the original lamellar hypothesis, suchcoordination would be in keeping with it, necessitating only that thislocal mechanism be added to the overall hippocampal model.Clearly, data on the actual physiological influences of axonal
projections, and their anatomical arrangements, are crucial forresolving such issues. For instance, our results essentially discon-firm the postulate that the longitudinal associational CA1 networkprimarily contacts interneurons. Stimulation of these fibers causedsignificant excitation, not inhibition, in their downstream targetcells. In addition, the CA1 longitudinal system demonstratesconsiderable capacity for the kinds of synaptic plasticity thoughtto be important for information storage. If this pathway simplyacted as a relay to coordinate the actions of lamellae, simple ex-citation of the downstream target cells would seem to be sufficient;there is no obvious role for synaptic plasticity if that is all they do.Hence, although we cannot rule out that coordination across la-mellae is a function of theCA1 longitudinal pathway, its capacity toundergo significant and subtle synaptic plasticity permits it to playmore sophisticated roles in hippocampal information processing.Regardless of how the physiological details are resolved, our
findings have important implications for the understanding ofthe hippocampus as a region capable of processing informationthroughout its full 3D extent. Indeed, as suggested (12), it mayno longer be productive to view the evidence as being either“for” or “against” the lamellar hypothesis. This hypothesis, al-though highly important and influential, leading as it did to themassive and productive output of work done on the transversehippocampal slice, is simply too limited to encompass the rich-ness of the anatomical and physiological data.
MethodsBrain Slice and Whole Hippocampus Preparation for Physiology. Sprague–Dawley rats (postnatal age: 4–6 wk) for brain slices and C57BL/6 mice (post-natal day 10–12) for whole hippocampus imaging were used. Additionaldetails are provided in SI Methods.
Whole-Cell Patch Recording. Whole-cell patch recordings were obtained byusing anAxon instruments Axoclamp 700BAmplifier (Molecular Devices), andpClamp Version 10.2 software (Molecular Devices) or Igor Pro (WaveMetrics)was used for data acquisition. Additional details are provided in SI Methods.
Three-Dimensional Digital Holography Uncaging. The procedures for digitalholographic photolysis have been described in detail in an earlier methodspaper (20). Digital holographic photolysis is a method that readily permitsflexible and accurate uncaging of glutamate and, therefore, stimulation ofcells at multiple discrete dendritic sites. Unlike conventional uncaging tech-niques, the holographic method permits glutamate photolysis to be directedprecisely at multiple sites and depths simultaneously. Additional details areprovided in SI Methods.
Two-Photon Imaging. The laser was tuned to 810 nm for Ca2+ imaging. Epi-fluorescence and transfluorescence signals were captured through a 60×,1.0 N.A. objective and a 1.4 N.A. oil immersion condenser (Olympus). Fluo-rescence was split into red and green channels by using dichroic mirrors andband-pass filters. Green fluorescence (Fluo-5F) and Red fluorescence (Alexa594) were captured with H10440PA and R9110 PMTs, respectively . Data werecollected in linescan mode (2.5 ms per line, including mirror flyback). For Ca2+
imaging, datawere presented as averages of 10 events per site, and expressedas Δ(G/R)/(G/R)max × 100, where (G/R)max is the maximal fluorescence insaturating (2mM) Ca2+. Ca2+ transient peakswere calculated from the peak ofthe fluorescence rise after stimulus onset.
VSD Imaging. Whole hippocampi from rats (p7–p12) were stained with0.125 mg/mL JPW3031 in artificial CSF for 20 min and imaged in an oxy-genated interface chamber (34 °C) by using an 80 × 80 CCD camera (Neu-roCCD; RedShirtImaging). Epi-illumination was provided by a custom LEDilluminator. Compared with the more commonly used photodiode array, theCCD chip well size (215,000 e−) requires use of relatively low light intensities,which minimized photodynamic damage. A 4× objective lens (0.28 N.A.;Olympus) imaged a 2.5 × 2.5-mm region in the hippocampal area CA1 (32 ×32-μm region imaged per pixel).
Extracellular Field Recording. Extracellular multiple unit recordings wereobtained by using glass pipettes filled with ACSF (3-7MΩ). Synaptic responseswere evoked with 15–60 μA, 0.2-ms current pulses delivered through a con-centric bipolar stimulating electrode (FHC, 100 μm o.d.). Baseline responseswere recorded by using half-maximal stimulation intensity at 0.033 Hz. HFSfor LTP was induced by 100 Hz stimulation (1-s duration).
Biocytin and Spinophillin Histochemistry. At the end of an electrophysiologyexperiment, the slices injected with biocytin were reacted in avidin–biotin–peroxidase complex. Labeled neurons were reconstructed in 3D by using amotorized microscopy interfaced with the Neurolucida software. For spi-nophilin histochemistry, tissue with biocytin was rinsed and placed intorabbit anti-spinophilin serum overnight at room temperature, and then in-cubated with a Cy2-conjugated goat anti-rabbit. Additional details are pro-vided in SI Methods.
1. Li XG, Somogyi P, Ylinen A, Buzsáki G (1994) The hippocampal CA3 network: An invivo intracellular labeling study. J Comp Neurol 339(2):181–208.
2. Marr D (1971) Simple memory: A theory for archicortex. Philos Trans R Soc Lond B BiolSci 262(841):23–81.
3. Gluck MA (1996) Computational models of hippocampal function in memory. Hip-pocampus 6(6):565–566.
4. Levy WB (1996) A sequence predicting CA3 is a flexible associator that learns and usescontext to solve hippocampal-like tasks. Hippocampus 6(6):579–590.
5. Yang S, et al. (2013) Integrity of mGluR-LTD in the associative/commissural inputs toCA3 correlates with successful aging in rats. J Neurosci 33(31):12670–12678.
6. Knowles WD, Schwartzkroin PA (1981) Local circuit synaptic interactions in hippo-campal brain slices. J Neurosci 1(3):318–322.
7. Deuchars J, Thomson AM (1996) CA1 pyramid-pyramid connections in rat hippocampusin vitro: Dual intracellular recordings with biocytin filling. Neuroscience 74(4):1009–1018.
8. Brun VH, et al. (2002) Place cells and place recognition maintained by directentorhinal-hippocampal circuitry. Science 296(5576):2243–2246.
9. Hunsaker MR, Lee B, Kesner RP (2008) Evaluating the temporal context of episodicmemory: The role of CA3 and CA1. Behav Brain Res 188(2):310–315.
10. Hoge J, Kesner RP (2007) Role of CA3 and CA1 subregions of the dorsal hippocampuson temporal processing of objects. Neurobiol Learn Mem 88(2):225–231.
11. Andersen P, Bliss TV, Skrede KK (1971) Lamellar organization of hippocampal path-ways. Exp Brain Res 13(2):222–238.
12. Amaral DG, Witter MP (1989) The three-dimensional organization of the hippocam-pal formation: A review of anatomical data. Neuroscience 31(3):571–591.
13. Sloviter RS, Lømo T (2012) Updating the lamellar hypothesis of hippocampal orga-nization. Front Neural Circuits 6:102.
14. Santos MD, et al. (2012) Dendritic hold and read: A gated mechanism for short terminformation storage and retrieval. PLoS ONE 7(5):e37542.
15. Alger BE, Teyler TJ (1977) A monosynaptic fiber track studied in vitro: Evidence ofa hippocampal CA1 associational system? Brain Res Bull 2(5):355–365.
16. y Cajal SR (1909-1911) Histologie du Systeme Nerveux de I’Homme et des Vertebres(Maloine, Paris), Vol 2, pp 986–993.
17. Knowles WD, Schwartzkroin PA (1981) Axonal ramifications of hippocampal Ca1pyramidal cells. J Neurosci 1(11):1236–1241.
18. Amaral D, Lavenex P (2007) The Hippocampus Book (Oxford Univ Press, Oxford), pp 27–110.19. Allen PB, Ouimet CC, Greengard P (1997) Spinophilin, a novel protein phosphatase 1
binding protein localized to dendritic spines. Proc Natl Acad Sci USA 94(18):9956–9961.20. Yang S, et al. (2011) Three-dimensional holographic photostimulation of the dendritic
arbor. J Neural Eng 8(4):046002.21. Lutz C, et al. (2008) Holographic photolysis of caged neurotransmitters. Nat Methods
5(9):821–827.22. Yang S, Emiliani V, Tang CM (2014) The kinetics of multibranch integration on the
dendritic arbor of CA1 pyramidal neurons. Front Cell Neurosci 8:127.23. Hasselmo ME (2008) Grid cell mechanisms and function: Contributions of entorhinal
persistent spiking and phase resetting. Hippocampus 18(12):1213–1229.24. Patel J, Fujisawa S, Berényi A, Royer S, Buzsáki G (2012) Traveling theta waves along
the entire septotemporal axis of the hippocampus. Neuron 75(3):410–417.25. Givone DD (2003) Digital Principles and Design (McGraw–Hill, New York).26. Acsády L, Kamondi A, Sík A, Freund T, Buzsáki G (1998) GABAergic cells are the major
postsynaptic targets of mossy fibers in the rat hippocampus. J Neurosci 18(9):3386–3403.27. Feng G, et al. (2000) Imaging neuronal subsets in transgenic mice expressing multiple
spectral variants of GFP. Neuron 28(1):41–51.
12924 | www.pnas.org/cgi/doi/10.1073/pnas.1405468111 Yang et al.