impaired hippocampus-dependent supplementary information
TRANSCRIPT

Impaired hippocampus-dependent and facilitated striatum-dependent behaviors in mice lacking the delta opioid receptor
Julie Le Merrer, Xavier Rezai, Grégory Scherrer, Jérôme A.J. Becker and Brigitte L. Kieffer
Supplementary information
Supplementary Material and Methods
Behavioral experiments
Experiments designed to compare Oprm1+/+ and Oprm1-/- mice were performed on
independent cohorts of naïve animals, except for the runway test that was performed in the
same cohort as rotarod experiment (easy condition). Rotarod testing was performed when the
animals were aged 7 weeks. After completion of this test (1 week), the animals were given a
1-week recovery period before food restriction started in view of runway training.
Elevated plus-maze (EPM): preliminary experiment. The EPM was a plus-shaped maze
elevated 52cm from base, with black Plexiglas floor, consisting of two open and two closed
arms (37x6 cm each) connected by a central platform (6x6 cm). The walls of the closed arms
were made of 18 cm-high clear acrylic. Light intensity in open arms was set at 15 lx. The
apparatus was placed over an infrared-lit platform. The movement and location of the mice
were analyzed by an automated tracking system equipped with an infrared-sensitive camera
(Videotrack; View Point, Lyon, France). All sessions were videotaped for further analyses.
The test started when the mouse was placed on the central platform facing a closed arm and
lasted 5 minutes. The distance travelled, time spent and entries in different parts of the
apparatus were counted automatically (videotracking). Head dips were scored manually on
video recordings to assess risk taking behavior. Three ratios were calculated as measures of
anxiety: an entry ratio (entries in open arms/total entries in arms), a distance ratio (distance
1

travelled in open arms/total distance travelled in arms) and a time ratio (time spent in open
arms/total time spent in arms). Total distance travelled in the apparatus (including central
platform) was used as an index of forward activity.
Novel object recognition: experiments 1 and 2. The experiments were conducted in 4 equal
square arenas (50x50 cm) separated by 35 cm-high opaque grey Plexiglas walls. Light
intensity of the room was set at 15 lx to facilitate exploration and minimize anxiety levels,
previously shown to be high in Oprd1-/- mice (Filliol et al. 2000) and naltrindole-treated
animals (Perrine et al. 2006; Saitoh et al. 2005). These conditions were chosen based on the
results of a preliminary experiment showing that Oprd1-/- mice do not display signs of
increased anxiety in the EPM under such light conditions (see above). The floor was a white
Plexiglas platform (View Point, Lyon, France), spread with sawdust. The room was equipped
with an overhead video camera connected to a computerized interface, allowing visualization
and recording of behavioral sessions on a computer screen in the adjacent room.
The experimental paradigm was adapted from (Carey et al. 2009), and lasted for 2 days. On
day 1, the animals were placed in an arena for a 15 min-habituation session with two copies of
an unfamiliar object (T-shaped plastic tubing, 1.5x3.5 cm). These objects were not used later
for recognition test. On day 2, the recognition test was performed. In experiment 1, this test
consisted of 3 trials of 10 minutes separated by 2 intertrial intervals of 5 minutes, during
which the animals were returned to their home cage (see Figure 1). In experiment 2, the third
trial (object phase) was omitted. Indeed, pilot studies revealed that, due to anxiogenic effects
of naltrindole (Perrine et al. 2006), exploration of the objects during this third phase was not
sufficient to allow a reliable measure of object recognition. On the first trial, or familiarization
phase, the mice were presented with two copies of an unfamiliar object. On the second trial,
or place phase, one of the two copies was displaced to a novel location in the arena. Finally,
2

on the third trial, or object phase, the copy that had not been moved on previous trial was
replaced by a novel object. Stimuli objects used in all previous experiments were Lego bricks,
plastic rings, dices or marbles (size 1.5-3x2-3 cm). The identity of the objects as well as the
spatial location in which these objects were positioned was balanced between subjects. The
number of visits and the time spent to explore each object were scored manually on video
recordings. A visit was acknowledged when the nose of the mouse came in direct contact with
an object. A percentage of discrimination was calculated for number of visits and time
exploring the objects as following: exploration of displaced or novel object / total exploration
* 100. The percentage of discrimination during familiarization phase was arbitrary calculated
for the object located in the right up corner of the arena. Animals that failed to explore the
objects more than 2 sec during familiarization phase were excluded from further analysis, as
well as mice that failed to explore one of the objects during place and object phases
(Experiment 1: 5 WT and 3 Oprd1-/- mice were excluded; Experiment 2: 4 WT and 4 Oprd1-/-
mice were excluded).
Place and response learning: experiments 3 and 4. Experiments were run in a cross-shaped
maze, adapted from (Passino et al. 2002). Elevated 40 cm above the floor, the maze consisted
of four arms (35 cm x 8 cm) with black Plexiglas floors enclosed in transparent Plexiglas
walls (15 cm), except for the terminal half of west and east arms. Removable sliding doors
made of black opaque Plexiglas delimited two starting boxes (10 x 8 cm) at the end of south
and north arms. Four more identical sliding doors separated each arm from the central
platform (8 x 8 cm). A food well (2 cm diameter) was inserted into the floor at 1 cm from the
distal end of east and west arms. The maze was located in a testing room that contained
several extra-maze visual cues, and maintained in a constant orientation during the
3

experiment. Light intensity in the room was set at 15 lx. The floor and walls were cleaned
daily to limit intra-maze olfactory cues.
Place learning was evaluated in experiment 3 using a dual-solution cross-maze task, whereas
response learning was measured in experiment 4 using a single-solution cross-maze task (see
Packard 2009). In both experiments, mice were reduced to 85% of their ad lib feeding weights
over 7 days before maze training and maintained at this weight throughout the experiment.
The animals received sucrose reward tablets (5-10 per mouse; Formula 5TUT-formerly
PJFSC-20 mg, TestDiet, Richmond, USA) in their home cage for three consecutive days
before maze habituation. Habituation lasted three days. In experiment 3, access to the north
arm of the cross maze was blocked with a sliding door during this phase. In experiment 4, all
arms were accessible. The mice were placed in the south start box (experiment 3) or in the
south or north start box (counterbalanced, experiment4) and allowed to explore the maze for 5
min. On day 1, sucrose tablets were available throughout the apparatus. On day 2, a trail of
five tablets leading to the food cup was placed along the length of west and east arms. On day
3, tablets were present only in the food well at the end of baited arms.
In experiment 3, we adapted a classical dual-solution cross-maze protocol to assess spatial
strategy in rodents (Deipolyi et al. 2008; Packard 1999; Passino et al. 2002). Training (4 trials
per day) started immediately after habituation. The north arm was closed and mice were
released from the south arm, after a 15 s-confinement in the start box (see Figure 2). For half
of the animals, single food-pellet bait was located in the east arm, while the other half of the
animals received food in the west arm. After entering an arm, the door was closed and mice
were confined for at least 20 s or until food was consumed. If a mouse failed to eat the food
within 5 min, the trial was terminated. A correction procedure was used during the first two
training sessions only: mice making an incorrect response were allowed to trace back to the
baited maze arm and consume the food pellet. On probe trials (days 5, 12 and 19), mice were
4

released from the north arm and had access to the previously baited arm (place learning) or to
the opposed arm (response learning). Food was available in both arms (see Figure 2). Two
parameters were recorded: choice accuracy was expressed as the percentage of entries in the
baited arm during each session, and choice latency was recorded as the latency to enter an arm
of the maze (baited or not).
In experiment 4, a single-solution response learning task was designed based on (Packard
2009). For two days after habituation, mice received pre-training sessions of two trials per day
during which they were released (after a 15 s-confinement in start box) alternatively from
north and south arm (see Figure 2). During these sessions, mice were allowed to correct a
wrong choice and retrieve food in the baited arm. Then training sessions of 5 trials per day
started. Mice were released (after a 15 s-confinement in the start box) from north or south
arm, following a random sequence (for example: north-north-south-north-south). Each mutant
animal was matched to a WT mouse for the release sequence. North arm was closed when the
animals were released from south arm. Conversely, south arm was not available when mice
were released from north arm. Half of the animals received food when turning left, and the
other half received food when turning right. After entering an arm, the door was closed and
mice were confined for at least 20 s or until food was consumed. If a mouse failed to eat the
food within 5 min, the trial was terminated. A correction procedure was used during the two
pre-training sessions and the first training session: mice making an incorrect response were
allowed to trace back to the baited maze arm and consume the food pellet. Two parameters
were recorded: choice accuracy was expressed as the percentage of entries in the baited arm
during each session, and choice latency was recorded as the latency to enter an arm of the
maze (baited or not). In this experiment, one mutant male mouse failed to eat the sucrose
reward pellets and was hence excluded from testing.
5

Skill motor learning: experiment 5. Mice were aged 7 weeks at the beginning of this
experiment, to limit weight bias. The animals were placed on a rotarod apparatus (Bioseb,
Valbonne, France) accelerating from 4 to 40 rpm in 5 min. In the “easy” condition, the rod
was covered with insulation tubing, which external perimeter was 5 cm. In the “hard”
condition, tubing was removed and external perimeter of the rod was 3 cm. Light intensity in
the room was 40 lx.
Behavioral testing lasted five days. On day 1, mice were habituated to rotation on the rod
under a constant speed of 4 rpm, until they were able to stay on the rod more than 180 s under
the “easy” condition or 90 s under the “hard” condition. From day 2 to day 5, mice were
tested for three trials a day (60-s intertrial interval) on consecutive days. Each trial started by
placing the mice on the rod and beginning rotation at constant 4 rpm-speed for 60 s. Then the
accelerating program was launched, and trial ended for a particular mouse when falling off the
rod. Time stayed on the rod was automatically recorded.
Locomotor activity under pharmacological challenge: experiment 6. Locomotor activity
was assessed in clear Plexiglas boxes (21 × 11× 17 cm) placed over a white Plexiglas
infrared-lit platform. Light intensity of the room was set at 15 lx. The trajectories of the mice
were analyzed and recorded via an automated tracking system equipped with an infrared-
sensitive camera (Videotrack; View Point, Lyon, France). To focus on forward activity, only
movements which speed was over 6 cm/s were taken into account for the measure of
locomotor activity.
Behavioral testing started when the animals were placed in the activity boxes for a 60 min-
habituation period. Habituation allowed the animals to reach a low and stable level of basal
activity and ensured reliable subsequent measure of drug-induced locomotor effects. Mice
were injected with either vehicle, the D1/D3 dopamine receptor agonist SKF-81297 (0.5, 1 or
6

2.5 mg/kg, s.c.) or the D2/D5 dopamine receptor agonist quinpirole (0.05, 0.1 or 0.5 mg/kg,
s.c.), and locomotor activity was monitored for further 60 min. Doses were chosen based on a
review of literature (Guzman et al. 2011; Napolitano et al. 2010; Smith et al. 2005).
Food seeking: experiments 7 and 8. In experiment 7, food seeking was assessed in a runway
(70 x 8 cm) with black Plexiglas floor enclosed in transparent Plexiglas walls (15 cm) and
elevated 40 cm above the floor. Removable sliding doors made of black opaque Plexiglas
delimited two boxes (10 x 8 cm), a start box and a goal box, at each end of the alley. A food
well (2 cm diameter) was inserted into the floor at 1 cm from the end of the goal box. Light
intensity in the room was set at 15 lx.
We adapted a runway task (Barbano et al. 2009; Pecina et al. 2003) to match as closely as
possible the conditions of the cross-maze tasks that we used in experiments 3 and 4. The
animals (WT – n=5 females, 5 males; Oprd1-/- - n=5 females, 5 males) were reduced to 85%
of their ad lib feeding weights over 7 days before runway training and maintained at this
weight throughout the experiment. They received sucrose reward tablets (5-10 per mouse;
Formula 5TUT-formerly PJFSC-20 mg, TestDiet, Richmond, USA) in their home cage for
three consecutive days before habituation. Habituation lasted three days. The mice were
placed in the start box and allowed to explore the alley for 5 min. On day 1, sucrose pellets
were available throughout the apparatus. On day 2, a trail of five pellets leading to the food
cup was placed along the alley. On day 3, pellets were present only in the food well. Training
(4 trials per day) started immediately after habituation. Mice were released after a 15 s-
confinement in the start box. A single food-pellet bait was located in the food well of the goal
box. After entering this goal box, the door was closed and mice were confined for at least 20 s
or until food was consumed. If a mouse failed to eat the food within 5 min, the trial was
terminated. An observer recorded the amount of time needed for each mouse to reach the goal
7

box (running latency). Observation of retreat behavior in Oprd1-/- mice (not scored) evoked an
approach-avoidance conflict in these animals (Geist and Ettenberg 1997), consistent with their
high levels of anxiety (Filliol et al. 2000). To reduce anxiety levels in this task, we thus
abolished confinement in end box during two additional “challenge” sessions. Under these
novel conditions, the animals performed as many trials as they could (starting with 15 s-
confinement in the start box) in a fixed delay of 4 min.
In experiment 8, food seeking was assessed in 4 equal square arenas (50x50 cm) separated by
35 cm-high opaque grey Plexiglas walls. The floor was a white Plexiglas platform, covered
with 5 cm of fresh sawdust. Three pellets of ordinary lab chow were placed on a white square
tissue in the center of the arena. Light intensity in the center of the arenas was set at 60 lx.
A novelty-suppressed feeding protocol was adapted from (Zhou et al. 2010). Mice (Exposed
to arena: WT – n=5 females, 4 males; Oprd1-/- - n=4 females, 4 males; control: WT – n=6
females, 8 males; Oprd1-/- - n=6 females, 8 males) were food-deprived (no food - water only)
for 24 h before the test, and isolated in a standard housing cage for 30 min before testing.
Animals were tested one at a time. The test started when a mouse was placed in a corner of
one of the arenas and allowed to explore for a maximum of 15 min. A food approach was
counted each time the nose of the mouse came in contact with the food pellets. Latency to
feed was measured as the time necessary to bite a food pellet. Immediately after an eating
event, the mouse was transferred back to home cage (free from other cage-mates) and allowed
to feed on lab chow for 5 min. Food consumption in the home cage was measured. Non-
exposed control mice undergone 24-h fasting and 30 min isolation in a standard housing cage
but were not introduced into the testing arenas. They were instead immediately placed back in
their home cage (free from other cage-mates) and allowed to feed on lab chow for 5 min.
Gene expression
8

Tissue collection. As previously described (Goeldner et al. 2011; Le Merrer et al. 2012),
brains were removed and placed into a matrix with 1 mm division (ASI Instruments, Warren,
MI, USA) cooled on ice. Slices were rapidly collected and structures were dissected based on
the stereotaxic atlas of mouse brain (Paxinos and Franklin 2001). Bilateral punches were
taken from two consecutive slices of the caudate putamen (CPu) and nucleus accumbens
(NAc). Dorsal hippocampus was dissected bilaterally from two consecutive brain slices
(HPC). These tissues were immediately frozen on dry ice and kept at -80°C until use.
Real-time quantitative PCR analysis. For each structure of interest and each genotype
(n=10 mice per genotype), tissue collected bilaterally from one male and one female mouse
was pooled in the same sample (n=5 samples per genotype). Therefore, real-time quantitative
PCR (qRT-PCR) was performed on 5 independent samples per genotype. RNA was extracted
and purified using the MIRNeasy mini-kit (Qiagen, Courtaboeuf, France) according to the
manufacturer's instructions. Total RNA (2.5 μg) was treated for 30 min at 37 °C by DNase I
RNase-free (5 U; Invitrogen, Carlsbad, USA) in first-strand Superscript buffer (Invitrogen)
and the reaction was stopped by incubating the mix for 5 min at 75 °C. RNA was pre-
incubated with anchored-oligo-dT primer (8 µM), random hexamer (16 µM) and dNTPs
(500 μM each) in a volume of 30 μL for 5 min at 65 °C. First-strand Superscript buffer,
dithiothreitol (0.01 M) and Superscript II (400 U; Invitrogen) were added in a final volume of
50 μL for 50 min at 42 °C. The reaction was stopped by 15 min incubation at 70 °C. qRT-
PCR was performed in quadruplets on a LightCycler 480 Real-Time PCR (Roche, Manheim,
Germany) using iQ SYBR Green supermix (Bio-Rad, Marnes-la-Coquette, France) kit with
0.25 µl cDNA in a 12.5 µl final volume. Gene-specific primers were designed using primer3
(http://frodo.wi.mit.edu/cgi-bin/primer3/primer3_www.cgi) to obtain a 100- to 150-bp
9

product. Each primer pair was validated for specificity and checked for unique melting curve
of each amplicon (see Table S1 for primer sequences). Thermal cycling parameters were
5 min at 95 °C followed by 40 cycles of 15 s at 95 °C, 15 s at 60 °C and 30 s at 72 °C.
Relative expression ratios were normalized to the level of actin as the reference gene and the
2−ΔΔCt method was applied to evaluate differential expression level (Livak and Schmittgen
2001).
Drugs
The nonpeptide antagonist of delta opioid receptors naltrindole hydrochloride was obtained
from Sigma (Sigma-Aldrich, Lyon, France). The D1/D3 dopamine receptor agonist SKF-
81297 (2,3,4,5-tetrahydro-6-chloro-7,8-dihydroxy-phenyl-1H-3-benzazepine) and the D2/D5
dopamine receptor agonist quinpirole were purchased from Tocris (Tocris Bioscience, Bristol,
UK). Compounds were dissolved in sterile isotonic saline solution (NaCl 0.9%) and injected
s.c. in a volume of 10 ml/kg. Doses refer to salt weight.
Statistical analyses
Behavioral experiments. Results from preliminary EPM experiment
Novel object recognition data (experiments 1-2) were expressed as percentage of time spent
exploring or number of visits to the displaced or novel object. During the first phase, this
percentage was arbitrary calculated for the object located in the right and/or top half of the
arena. These data were analyzed using three-way ANOVA, with genotype and gender as
between-subject factors and phase as within-subject factor. Data from cross-maze and runway
tasks (experiment 3, 4 and 7) were analyzed using three-way ANOVA with genotype and
10

gender as between-subject factors and session as within-subject factor. Pearson’s 2 test was
used to compare the percentage of place learners between Oprd1-/- and WT mice in
experiment 3. Data from the skill motor learning task (experiment 5) were analyzed using
five-way ANOVA with genotype, gender and condition (“hard” versus “easy”) as between
subject factors and trials and sessions as within-subject factors. Locomotor activity data
(experiment 6) were analysed using three-way ANOVA, with genotype, gender and dose of
dopamine agonist as between-subject factors. Finally, data from novelty-suppressed feeding
experiment (experiment 8) were analyzed using two-way ANOVA with genotype and gender
as between factors (latency to first eat, approaches) or three-way ANOVA with genotype,
gender and condition (arena exposure or not) as between factors. Statistical significance was
set at p<0.05 for all tests (Statistica 9.0, StatSoft, Maisons Alfort).
Gene expression. qRT-PCR data were transformed prior to statistical analysis to obtain a
symmetrical distribution centered on 0 (corresponding to no change in gene expression) using
the following formula: if x<1, y=1-1/x; if x>1, y=x-1 (x: qPCR data; y: transformed data) (Le
Merrer et al. 2012). A Student’s t test was then performed to assess their statistical
significance. Calculated p values indicated probability for a regulation to differ from 0. Only
regulations over +1.20 or below -1.20 (fold change) were retained as significant.
Supplementary Results
Oprd1-/- mice do not display signs of elevated anxiety when tested under low light
conditions in the EPM
11

In a preliminary experiment, we evaluated signs of anxiety in mutant mice and their controls
using the EPM test under low light conditions (15 lx).
At 15 lx, WT and Oprd1-/- mice displayed similar numbers of entries (genotype: F1,12<1;
gender: F1,12<1; arm: F1,12<1; arm x gender: F1,12=12.91, p<0.01), similar distance travelled
(genotype: F1,12=3.84, NS; gender: F1,12=6.37, p<0.05; arm: F1,12=14.49, p<0.0001; arm x
gender: F1,12=10.18, p<0.001) and time spent (genotype: F1,12<1; gender: F1,12<1; arm:
F1,12=7.36, p<0.05; arm x gender: F1,12=8.59, p<0.05) in the open and closed arms of the EPM
(see Figure S1a,b and c), resulting in similar entry (genotype: F1,12<1; gender: F1,12=13.86,
p<0.001), distance (genotype: F1,12<1; gender: F1,12=8.17, p<0.05) and time (genotype: F1,12<1;
gender: F1,12=8.68, p<0.05) ratios (Figure S1d). Females appeared more anxious than males,
independently from the genotype. Control and mutant mice travelled the same distance in the
apparatus (genotype: F1,12=3.10, NS; gender: F1,12=9.16, p<0.05), demonstrating similar
forward activity in this test (females in both genotype being less active than males). Finally,
delta opioid receptor null mice performed significantly more head dips than WT animals
(genotype: F1,12=9.24, p<0.05; gender: F1,12<1), suggesting that risk taking behavior may be
increased in these animals.
Increased food seeking in mice lacking delta opioid receptors
In experiments 7 and 8, we evaluated whether motivation to obtain a food reward was
modified delta opioid receptor null mice.
In a runway task (experiment 7), running latencies were similar across training sessions in
Oprd1-/- and WT mice (Figure S3A), and were slower to decrease in females than in males
(Three-way ANOVA, genotype: F1,12<1; gender: F1,12=11.14, p<0.01; session: F16,192=6.71,
p<0.0001; session x gender: F16,192=3.03, p<0.001). We then performed two consecutive
challenge sessions, during which the animals were allowed to retrieve as many food pellets as
12

they could in 4 min, and were no longer confined in goal arm. Under these conditions, mutant
mice obtained significantly more pellets (genotype: F1,12=40.59, p<0.0001) and reached the
food quicker (genotype: F1,12=5.91, p<0.05) than their WT counterparts.
In a novelty-suppressed feeding task (experiment 8), Oprd1-/- and WT mice displayed
equivalent latencies to start feeding in the center of the arena (two-way ANOVA, genotype:
F1,13<1) (Figure S3B). Mutant mice, however, approached the food more often than WT
controls (genotype: F1,13=36.65, p<0.0001). Back in their home cage, mice exposed to the
bright arena ate less lab chow than non-exposed controls (three-way ANOVA; genotype:
F1,37<1; arena exposure: F1,37=4.39, p<0.05). Subsequent two-way ANOVA revealed that
mutant mice consumed less food than their WT counterparts when placed back in their home
cage immediately after the novelty-suppressed feeding test (genotype: F1,13=6.86, p<0.05).
Data from experiments 7 and 8 indicate that motivation for food is not reduced in Oprd1-/-
mice, but instead appears increased, as suggested by facilitated performance in challenge
sessions of experiment 7 and increased number of food approaches in experiment 8.
Transcriptional regulations in the ventral striatum of Oprd1-/- animals
In the nucleus accumbens (NAc), delta receptor deletion modified the expression of 10 genes,
coding for actors of GABA signaling (up-regulation: Slc6a1; down-regulation: Gabra4;
Gabbr2), a glutamate transporter and an element of the post-synaptic densities (up-regulation:
Slc1a6; down-regulation: Dlg2), enzymes involved in monoamine metabolism (up-regulation:
Maoa, Ache) and MSN markers (up-regulation: Pde10a; down-regulation: Adora2a, Cnr1).
The expression of only 3 genes was found regulated in both the CPu and NAc: Grm4, Slc6a11
(up-regulated) and Slc6a4 (down-regulated in the CPu, up-regulated in the NAc).
13

Discussion experiments 7 and 8
Increased food seeking in mice lacking delta opioid receptors
In the runway task (Figure S2a), latencies to reach the food were not modified in mutant
animals as compared to WT controls when experimental conditions matched these of the
cross-maze tasks (confinement in end box). However, under challenge conditions (no
confinement), Oprd1-/- mice obtained more food pellets than WT mice, suggesting increased
food seeking. This result may reflect competing high levels of anxiety (avoidance behavior)
and high motivation for food (approach behavior) in mutant animals (Aupperle and Paulus
2010; Montgomery 1955; Powell et al. 2004). By omitting confinement in the runway
experiment, we aimed at reducing anxiety levels, which are elevated in Oprd1-/- mice (Filliol
et al. 2000). This manipulation may have unmasked high motivation for food in mutants.
Consistent with this, in the novelty-suppressed feeding experiment (Figure S2b), these
animals took as long as WT controls to start eating in the arena, but approached food pellets
more often (and retreated), revealing conflicting avoidance and approach behaviors (Powell et
al. 2004). Decreased food intake of mutant animals in their home cage was likely due to high
anxiety levels after arena exposure, since not observed when this exposure was omitted. From
food-seeking experiments, therefore, we cannot exclude that motivation to obtain food is
increased in Oprd1-/- mice. Increased motivation for food may have contributed to ameliorate
response learning in a single-solution response cross-maze task (experiment 4), but failed to
ameliorate place learning in a dual-solution task (experiment 3).
ReferencesAupperle RL, Paulus MP (2010) Neural systems underlying approach and avoidance in
anxiety disorders. Dialogues Clin Neurosci 12: 517-31.Barbano MF, Le Saux M, Cador M (2009) Involvement of dopamine and opioids in the
motivation to eat: influence of palatability, homeostatic state, and behavioral paradigms. Psychopharmacology 203: 475-87.
14

Carey AN, Lyons AM, Shay CF, Dunton O, McLaughlin JP (2009) Endogenous kappa opioid activation mediates stress-induced deficits in learning and memory. The Journal of neuroscience : the official journal of the Society for Neuroscience 29: 4293-300.
Deipolyi AR, Fang S, Palop JJ, Yu GQ, Wang X, Mucke L (2008) Altered navigational strategy use and visuospatial deficits in hAPP transgenic mice. Neurobiol Aging 29: 253-66.
Filliol D, Ghozland S, Chluba J, Martin M, Matthes HW, Simonin F, Befort K, Gaveriaux-Ruff C, Dierich A, LeMeur M, Valverde O, Maldonado R, Kieffer BL (2000) Mice deficient for delta- and mu-opioid receptors exhibit opposing alterations of emotional responses. Nat Genet 25: 195-200.
Geist TD, Ettenberg A (1997) Concurrent positive and negative goalbox events produce runway behaviors comparable to those of cocaine-reinforced rats. Pharmacol Biochem Behav 57: 145-50.
Goeldner C, Lutz PE, Darcq E, Halter T, Clesse D, Ouagazzal AM, Kieffer BL (2011) Impaired emotional-like behavior and serotonergic function during protracted abstinence from chronic morphine. Biol Psychiatry 69: 236-44.
Guzman MS, De Jaeger X, Raulic S, Souza IA, Li AX, Schmid S, Menon RS, Gainetdinov RR, Caron MG, Bartha R, Prado VF, Prado MA (2011) Elimination of the vesicular acetylcholine transporter in the striatum reveals regulation of behaviour by cholinergic-glutamatergic co-transmission. PLoS Biol 9: e1001194.
Le Merrer J, Befort K, Gardon O, Filliol D, Darcq E, Dembele D, Becker JA, Kieffer BL (2012) Protracted abstinence from distinct drugs of abuse shows regulation of a common gene network. Addiction biology 17: 1-12.
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25: 402-8.
Montgomery KC (1955) The relation between fear induced by novel stimulation and exploratory behavior. J Comp Physiol Psychol 48: 254-60.
Napolitano F, Bonito-Oliva A, Federici M, Carta M, Errico F, Magara S, Martella G, Nistico R, Centonze D, Pisani A, Gu HH, Mercuri NB, Usiello A (2010) Role of aberrant striatal dopamine D1 receptor/cAMP/protein kinase A/DARPP32 signaling in the paradoxical calming effect of amphetamine. The Journal of neuroscience : the official journal of the Society for Neuroscience 30: 11043-56.
Packard MG (1999) Glutamate infused posttraining into the hippocampus or caudate-putamen differentially strengthens place and response learning. Proc Natl Acad Sci U S A 96: 12881-6.
Packard MG (2009) Exhumed from thought: basal ganglia and response learning in the plus-maze. Behavioural brain research 199: 24-31.
Passino E, Middei S, Restivo L, Bertaina-Anglade V, Ammassari-Teule M (2002) Genetic approach to variability of memory systems: analysis of place vs. response learning and fos-related expression in hippocampal and striatal areas of C57BL/6 and DBA/2 mice. Hippocampus 12: 63-75.
Paxinos G, Franklin K (2001) The Mouse Brain in Stereotaxic coordinates, 2d edition edn. Academic Press, San Diego
Pecina S, Cagniard B, Berridge KC, Aldridge JW, Zhuang X (2003) Hyperdopaminergic mutant mice have higher "wanting" but not "liking" for sweet rewards. The Journal of neuroscience : the official journal of the Society for Neuroscience 23: 9395-402.
Perrine SA, Hoshaw BA, Unterwald EM (2006) Delta opioid receptor ligands modulate anxiety-like behaviors in the rat. Br J Pharmacol 147: 864-72.
15

Powell SB, Geyer MA, Gallagher D, Paulus MP (2004) The balance between approach and avoidance behaviors in a novel object exploration paradigm in mice. Behavioural brain research 152: 341-9.
Saitoh A, Yoshikawa Y, Onodera K, Kamei J (2005) Role of delta-opioid receptor subtypes in anxiety-related behaviors in the elevated plus-maze in rats. Psychopharmacology 182: 327-34.
Smith DG, Tzavara ET, Shaw J, Luecke S, Wade M, Davis R, Salhoff C, Nomikos GG, Gehlert DR (2005) Mesolimbic dopamine super-sensitivity in melanin-concentrating hormone-1 receptor-deficient mice. The Journal of neuroscience : the official journal of the Society for Neuroscience 25: 914-22.
Zhou M, Rebholz H, Brocia C, Warner-Schmidt JL, Fienberg AA, Nairn AC, Greengard P, Flajolet M (2010) Forebrain overexpression of CK1delta leads to down-regulation of dopamine receptors and altered locomotor activity reminiscent of ADHD. Proc Natl Acad Sci U S A 107: 4401-6.
Supplementary figure legends
Figure S1. Oprd1-/- and WT mice show similar levels of anxiety when tested in the elevated
plus-maze under low light conditions (15 lx). Control and mutant animals display similar (a)
number of entries, (b) distance travelled and (c) time spent in open and closed arms of the
EPM. (d) Entry, distance and time ratios are equivalent in Oprd1-/- and WT mice. (e) WT and
mutant mice travel the same distance in the EPM, (f) but the later perform more head dips
than their WT counterparts. Data are expressed as mean ± esm. Genotype effect: one solid
star: p<0.05 (two-way ANOVA); Arm effect: one open star: p<0.05, two open stars:
p<0.01(three-way ANOVA).
Figure S2. Choice latency decreases over sessions in cross-maze tasks. (a) In the dual
solution cross-maze task, Oprd1-/- and WT mice display progressively shorter choice latencies
as they learned the task, females being slower than males in both mouse lines. (b) In the single
solution response task, choice latency is shorter to decrease over sessions in WT than in
mutant animals, females displaying longer latencies than males. Data are expressed as mean
(± esm) choice latency over 4 (a) or 5 (b) daily trials.
16

Figure S3. Food seeking appears increased in Oprd1-/- mice. (a) In the runway task, running
latencies decrease similarly over training sessions in mutant and WT animals, females being
slower than males in both lines. Under challenge conditions, however, Oprd1-/- mice gain
more food pellets and displayed shorter feeding latencies than WT mice. (b) In the novelty-
suppressed feeding task, latencies to feed are equivalent between WT and mutant mice. The
later, however, approach the food more often than their WT counterparts. Back in their home
cage immediately after the test, Oprd1-/- mice eat less than WT mice. This effect likely reflects
high anxiety levels in mutant animals, as it disappears when the animals are not exposed to
the test arena (control condition). Data are expressed as mean ± esm. Genotype effect: one
solid star: p<0.05, three solid stars: p<0.001 (two-way ANOVA); one open star: p<0.05, three
open stars: p<0.001(one-way ANOVA).
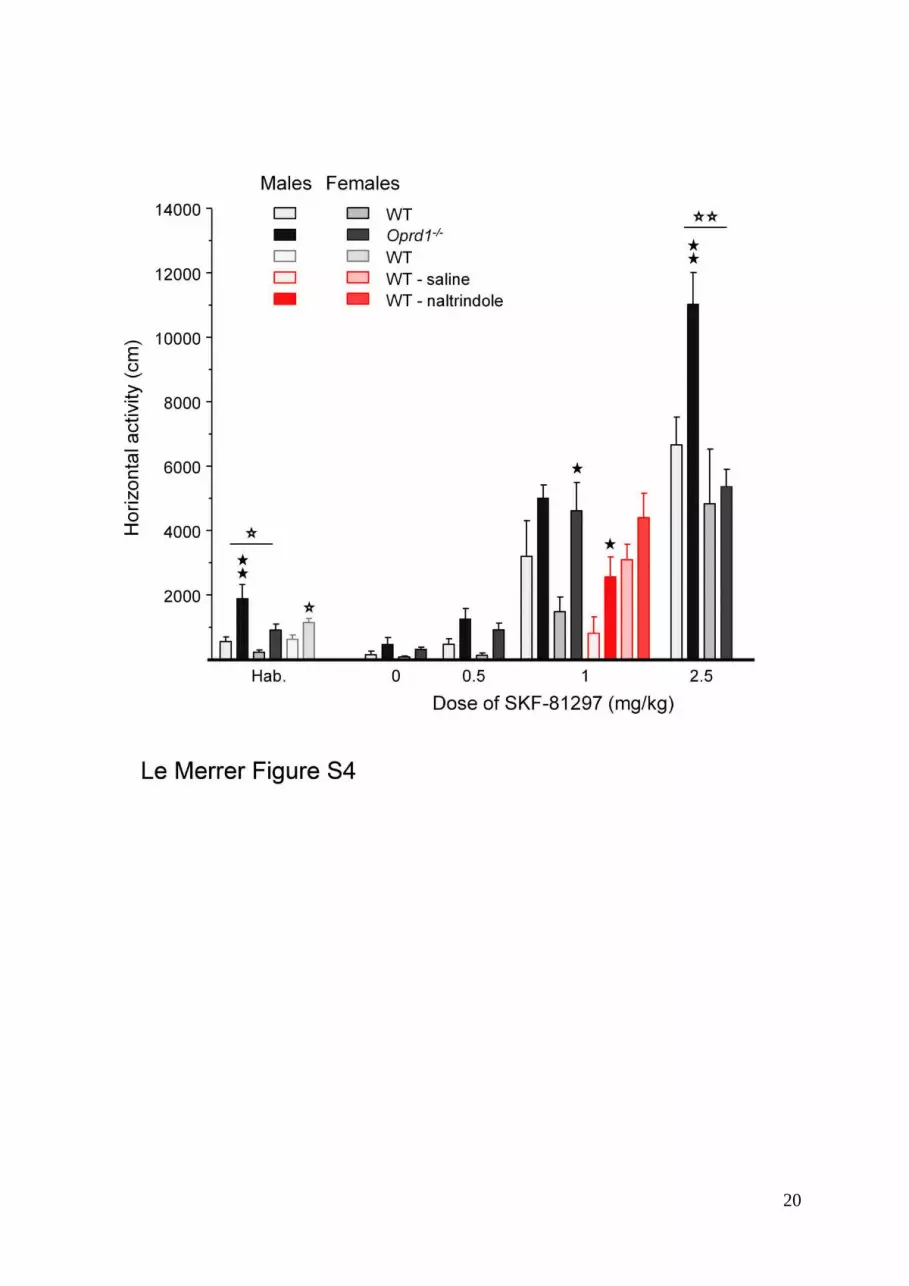
Figure S4. Genetic or pharmacological inactivation of delta opioid receptors results in
increased sensitivity to the locomotor effects of the D1/D5 agonist SKF-81297. This effect or
a trend towards this effect is seen both in male and female mice, except for the highest dose of
SKF-81297 in Oprd1-/- mice, at which only males displayed higher locomotor response as
compared to WT animals. Genotype/treatment effect: one solid star: p<0.05, two solid stars:
p<0.01; gender effect: one open star: p<0.05, two open stars: p<0.01(one- or two-way
ANOVA).
Supplementary table legends
Table S1. List of primers used for qRT-PCR.
17

Table S2. Transcription of genes coding for a set of 67 genes, including actors of GABA,
glutamate or monoamine signaling and neuronal markers, was evaluated in the hippocampus
(HPC), caudate putamen (CPu, dorsal striatum) and nucleus accumbens (NAc, ventral
striatum) of Oprd1-/- mice.
Data are presented as fold-change Oprd1-/- versus WT mice (median ± sem). Student’s t-tests
were performed on transformed data (see Material and Methods) to determine whether fold
changes differed from 0 (no regulation: corresponds to ±1 in table). Significant regulations are
highlighted in bold.
Table S3. Animal numbers used in experiment 6: effects of dopamine receptor agonists on
locomotor activity.
18

19
20