the group i mglu receptor agonist dhpg induces a novel form of ltd in the ca1 region of the...
TRANSCRIPT

Pergamon
PII: SOO28-3!Xr8(97)00181-0
Neuropharmmology, Vol. 36, No. 11/12, pp. 1517-1532, 1997 0 1998 Elsevier Science Ltd. All rights reserved
printed in Great Britain 0028-3908/98 $19.00 + 0.00
The Group I mGlu Receptor Agonist DHPG Induces a Novel Form of LTD in the CA1 Region of the
Hippocampus
M. J. PALMER,‘* A. J. IRVING,2 G. R. SEABROOK, D. E. JANE4 and G. L. COLLINGRIDGE’
‘Department of Anatomy, School of Medical Sciences, University of Bristol, Bristol BS8 ITD, U.K.; 2Department of Biomedical Sciences, institute of Medical Sciences, University of Aberdeen, Aberdeen AB2.5 220, U.K.; ‘Department of Pharmacology, Merck Sharp and Dohme, Neuroscience Research Centre, Terlings
Park, Harlow, Essex CM20 2QR, U.K. and 4Department of Pharmacology, School of Medical Sciences, University of Bristol, Bristol BS8 ITD, U.K.
(Accepted 20 September 1997)
Summary-The group I specific metabotropic glutamate (mGlu) receptor agonist (RS)-3,5_dihydroxyphe- nylglycine (DHIPG) (100 PM, 10 min) induced long-term depression (LTD) of synaptic transmission in the CA1 region of adult rat hippocampal slices, measured using a grease-gap recording technique. In “normal” (1 mM Mg2+-containing) medium, LTD (measured 30 min after washout of DHPG) was small (13 f 3%), but LTD was enhanced if DHPG was applied when the tissue was made hyperexcitable, either by omitting Mg2+ from the perfusate (35 + 3%) or by adding the GABAA receptor antagonist picrotoxin (29 + 2%). The N- methyl-D-aspartate (NMDA) receptor antagonist AP5 (100 PM) substantially reduced the generation of DHPG- induced LTD in. Mg 2+-free medium, but had little effect on LTD induced in the presence of picrotoxin. In Mg2+-free medium, the threshold concentration of DHPG required to induce LTD was between 1 and 3 PM. Neither agonists specific for group II (100 nM DCG-IV or 1 PM LY354740) or group III (10 PM L-AP4) mGlu receptors or a combined group I and II agonist (30-100 PM (lS,3R)-ACPD) induced LTD. However, an agonist (1 mM CHPG) which activates mGlus but not mGlui receptors did induce LTD. Surprisingly, DHPG-induced LTD was reversed by mGlu receptor antagonists, applied hours after washout of DHPG. DHPG-induced LTD did not occlude with LTD induced by synaptic activation (1200 stimuli delivered at 2 Hz), in Mg2+-free medium. These data show that activation of group I mGlu receptors (probably mGlu5) can induce LTD and that this mGlu receptor-mediated LTD may, or may not, require activation of NMDA receptors, depending on the experimental conditions. 0 1998 Elsevier Science Ltd. All rights reserved.
Keywords-Glutamate, metabotropic glutamate (mGlu) receptor, N-methyl-D-aspartate (NMDA) receptor, (RS)-3,5-dihydroxyphenylglycine (DHPG), (RS)-2-chloro-5-hydroxyphenylglycine (CHPG), (2S,l’RJ’R,3’R)- 2-(2’,3’-dicarboxycyclopropyl)glycine (DCG-IV), (S)-2-amino-4-phosphonobutanoic acid (L-AP4), hippo- campal, long-term depression (LTD), synaptic plasticity.
Long-term depression (LTD), like its counterpart long-
term potentiation (LTP), is the subject of intense
investigation, because: of the belief that long-term changes in synaptic efficiency are fundamental processes
involved in learning and memory (Bliss and Collingridge,
1993; Bear and Abraham, 1996). In the hippocampus, repetitive “low frequency” stimulation can reverse LTP by a process known as gdepotentiation (Barrionuevo et al., 1980; Staubli and Lynch, 1990; Fujii et al., 1991; Bashir and Collingridge, 1994.). A similar type of homosynaptic
*To whom correspondence should be addressed. Tel: (0117) 928 8392; Fax: (0117) 929 1687; E-mail: m.j.palmer@ bristol.ac.uk.
depression can also be induced without first inducing LTP, and is particularly pronounced in slices obtained from young animals (Dudek and Bear, 1992; Mulkey and Malenka, 1992). This will be referred to as homosynaptic LTD to distinguish it from depotentiation where necessary. Several studies have addressed the role of different glutamate receptor subtypes in the induction of these two forms of synaptic depression, but no clear consensus has emerged. In particular, there is confusion over the relative importance of N-methyl-D-aspartate (NMDA) receptors and metabotropic glutamate (mGlu) receptors. The induction of both depotentiation (Fujii et al., 1991) and homosynaptic LTD (Dudek and Bear, 1992; Mulkey and Malenka, 1992; Kemp and Bashir, 1997) can be prevented by the NMDA receptor
1517

1518 M. J. Palmer et al.
antagonist (R)-2-amino-5-phosphonopentanoate (AP5), but both depotentiation (Bashir and Collingridge, 1994) and homosynaptic LTD (Bolshakov and Siegelbaum, 1994; O’Mara et al., 1995; Kemp and Bashir, 1997) can also be induced in the presence of this antagonist. Similarly the mGlu receptor antagonist (S)-a-methyl4- carboxyphenylglycine (MCPG) can block the induction of depotentiation (Bashir et al., 1993; Bashir and Collingridge, 1994) and homosynaptic LTD (Bolshakov and Siegelbaum, 1994; O’Mara et al., 1995; Oliet et al., 1997) but does not invariably do so (Selig et al., 1995; Oliet et al., 1997).
A complementary strategy for studying the possible role of a glutamate receptor in the induction of synaptic plasticity is to apply a specific agonist. For example, the broad-spectrum mGlu receptor agonist (l&3@-l-amino- cyclopentane- 1,3-dicarboxylate (( lS,3R)-ACPD) can be used to induce LTP under certain circumstances (Bortolotto and Collingridge, 1993, 1995; Manahan- Vaughan and Reymann, 1995). (lS,3R)-ACPD can also induce LTD under certain conditions. It has been shown to induce LTD in area CA1 of slices obtained from very young animals (3-7-day-old rats) when paired with depolarization (Bolshakov and Siegelbaum, 1994), and from slices obtained from 8-12-day-old rats (Overstreet et al., 1997). It has also been shown to induce LTD in the medial perforant path input to the dentate gyrus in slices obtained from adult (25-35-day-old) rats (O’Mara et al., 1995).
During the course of an investigation into the role of mGlu receptor subtypes in regulating synaptic transmis- sion in area CA1 of slices obtained from adult (4-10- week-old) rats, we noted that (RS)-3,5-dihydroxyphenyl- glycine (DHPG), a specific agonist of group I mGlu receptors (Ito et al., 1992; Schoepp et al., 1994) could induce a long-lasting depression of synaptic transmis- sion, in the absence of added Mg2+ in the perfusate (Harvey et al., 1996). We have termed this effect DHPG- induced LTD, following from the terminology adopted for (l&3@-ACPD-induced long-lasting depression of synaptic transmission (O’Mara et al., 1995; Overstreet et al., 1997), and present here a characterization of this form of plasticity.
A preliminary account of some of this work has appeared in abstract form (Irving et al., 1996).
MATERIALS AND METHODS
Experiments were performed on 400-pm thick trans- verse hippocampal slices obtained from female rats of 140-200 g in weight (approximately 4-10 weeks of age). Area CA3 was surgically removed and slices perfused with medium (28-3O”C), which composed (mM): NaCl 124; KC1 3; NaHCOs 26; NaH2POd 1.25; MgS04 1; CaC12 2; D-ghCOSe 10 (bubbled with 95% 02/5% CO2; pH 7.4); at a rate of approximately 2 ml/mm. Drugs were applied in the perfusate. The Schaffer collateral- commissural pathway was stimulated continuously using
single shocks delivered once every 30 sec. Electrical activity was recorded using a grease-gap recording technique, as described previously (Blake et al., 1988; Harvey and Collingridge, 1993). In brief, the alveus was partially dissected from the slice and placed under the wick of a Ag/AgCl electrode. The wick and alveus assembly was covered by an insulating grease mixture and d.c. potentials were recorded between this electrode and a similar one placed in contact with the perfusate. The maximum slope of field excitatory postsynaptic potentials (fEPSPs) was plotted versus time on-line and data were subsequently analysed off-line using programs written in-house (Anderson and Collingridge, 1997).
(RS)-DHPG, (RS)-CHPG, DCG-IV, L-AP4, (l&3@- ACPD, (S)-MCPG, (S)-4CPG, MTPG, MPPG and (R)- AP5 were obtained from Tocris Cookson (Bristol, U.K.) and picrotoxin was purchased from Sigma (Poole, Dorset, U.K.). LY354740 was a gift from David Lodge (Eli Lilly).
Results were analysed with paired and unpaired Student’s t tests as appropriate. n signifies the number of times a result was obtained which, unless otherwise stated, is the same as the number of slices tested. Each slice was obtained from a separate rat.
RESULTS
DHPG can induce LTD
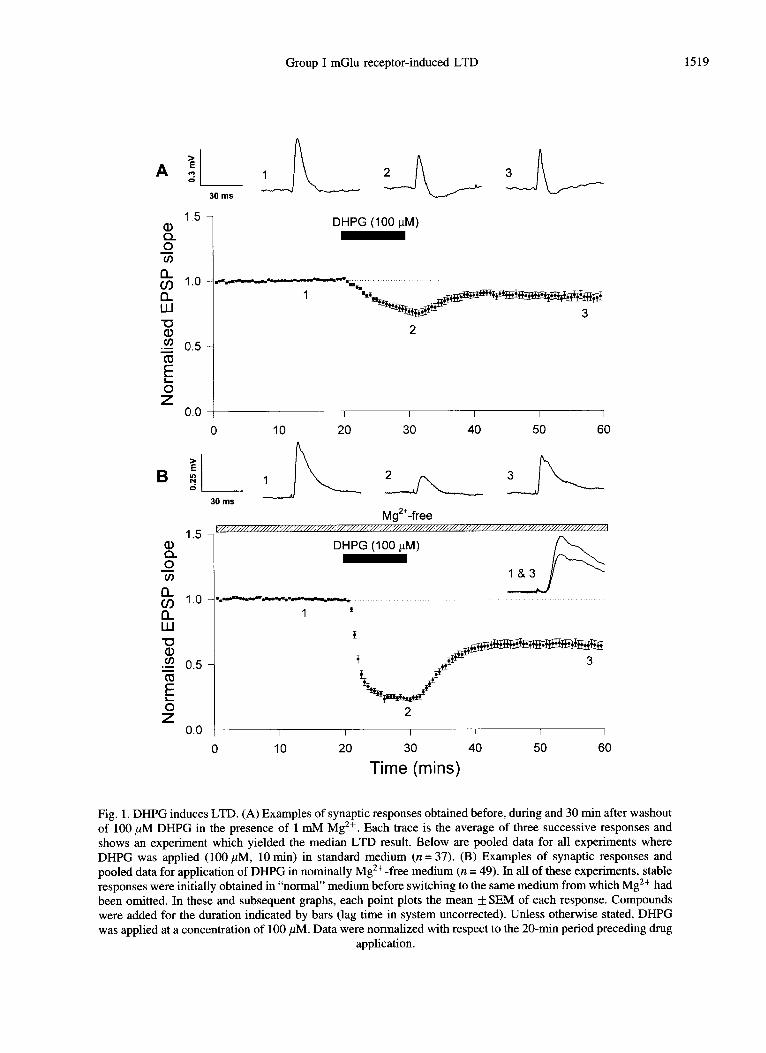
In the presence of 1 mM Mg2+ and absence of a GABA* receptor antagonist (hereafter termed “normal” medium), DHPG (100 ,uM, 10 min) caused a small depression of synaptic transmission (n = 37) which showed either partial or complete reversibility upon washout. In both cases, a stable response was obtained approximately 10 min following the start of washout of DHPG. When the data for all 37 slices were pooled, DHPG was found to lead, on average, to a sustained depression of 13 f 3% (p ~0.001; quantified 30 min following the start of washout). This sustained depression is termed DHPG-induced LTD. If LTD is defined as a stable depression of 10% or greater (at 30 min), then approximately 40% (15/37) of slices exhibited LTD. A single example and pooled data showing DHPG-induced LTD in normal medium is illustrated in Fig. l(A).
Enhanced DHPG-induced LTD under conditions of increased excitability
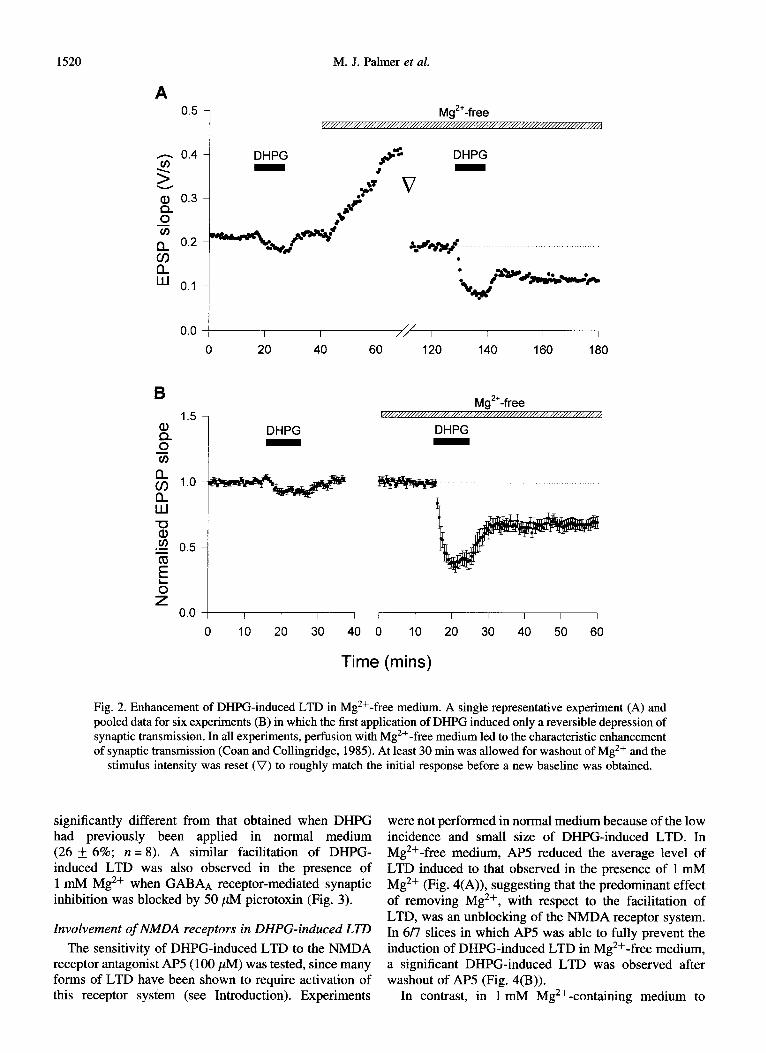
In the absence of Mg2+, DHPG induced LTD in over 80% (41/49) of slices. When the data for all 49 slices were pooled, DHPG was found to induce LTD of 35 f 3% (p <O.OOOl; Fig. l(B)). In a subset of these experiments, DHPG was applied in Mg2+-free medium to slices which had already been exposed to DHPG prior to washout of Mg2+ (and where no LTD had been induced). In 6/8 of these slices, substantial LTD (32 ) 4%; n = 6) was observed in Mg 2+-free medium (Fig. 2). The level of LTD obtained when DHPG was applied for the first time in Mg 2+-free medium (35 f 4%, n = 25) was not
Group I mGlu receptor-induced LTD 1519
DHPG (100 FM)
B;I Aa A 30 Ills
Ma*+-free
DHPG (100 pM)
I
0 I
10 I I I / I
20 30 40 50 60
Time (mins)
Fig. 1. DHPG induces LTD. (A) Examples of synaptic responses obtained before, during and 30 min after washout of 100 PM DHPGi in the presence of 1 mM Mg *+ Each trace is the average of three successive responses and . shows an experiment which yielded the median LTD result. Below are pooled data for all experiments where DHPG was applied (100 PM, 10 min) in standard medium (n = 37). (B) Examples of synaptic responses and pooled data for application of DHPG in nominally Mg 2+-free medium (n = 49). In all of these experiments, stable responses were initially obtained in “normal” medium before switching to the same medium from which Mg2+ had been omitted. In these and subsequent graphs, each point plots the mean f SEM of each response. Compounds were added for the duration indicated by bars (lag time in system uncorrected). Unless otherwise stated, DHPG was applied at a concentration of 100 PM. Data were normalized with respect to the 20-min period preceding drug
application.
1520 M. J. Palmer et al.
A 0.5 Mg*‘-free
0.0 /// 0 20 40 60 120 140 160 180
B Mg*‘-free
1.5
iti DHPG DHPG
0 z
I I I I I
0 10 20 30 40
I I I I I I I
0 10 20 30 40 50 60
Time (mins)
Fig. 2. Enhancement of DHPG-induced LTD in Mg ‘+-free medium. A single representative experiment (A) and pooled data for six experiments (B) in which the first application of DHPG induced only a reversible depression of synaptic transmission. In all experiments, perfusion with Mg’+-free medium led to the characteristic enhancement of synaptic transmission (Coan and Collingridge, 1985). At least 30 min was allowed for washout of Mg2+ and the
stimulus intensity was reset (V) to roughly match the initial response before a new baseline was obtained.
significantly different from that obtained when DHPG had previously been applied in normal medium (26 + 6%; 12 = 8). A similar facilitation of DHPG- induced LTD was also observed in the presence of 1 mM Mg2+ when GABAA receptor-mediated synaptic inhibition was blocked by 50 PM picrotoxin (Fig. 3).
Involvement of NMDA receptors in DHPG-induced LTD
The sensitivity of DHPG-induced LTD to the NMDA receptor antagonist AP5 (100 PM) was tested, since many forms of LTD have been shown to require activation of this receptor system (see Introduction). Experiments
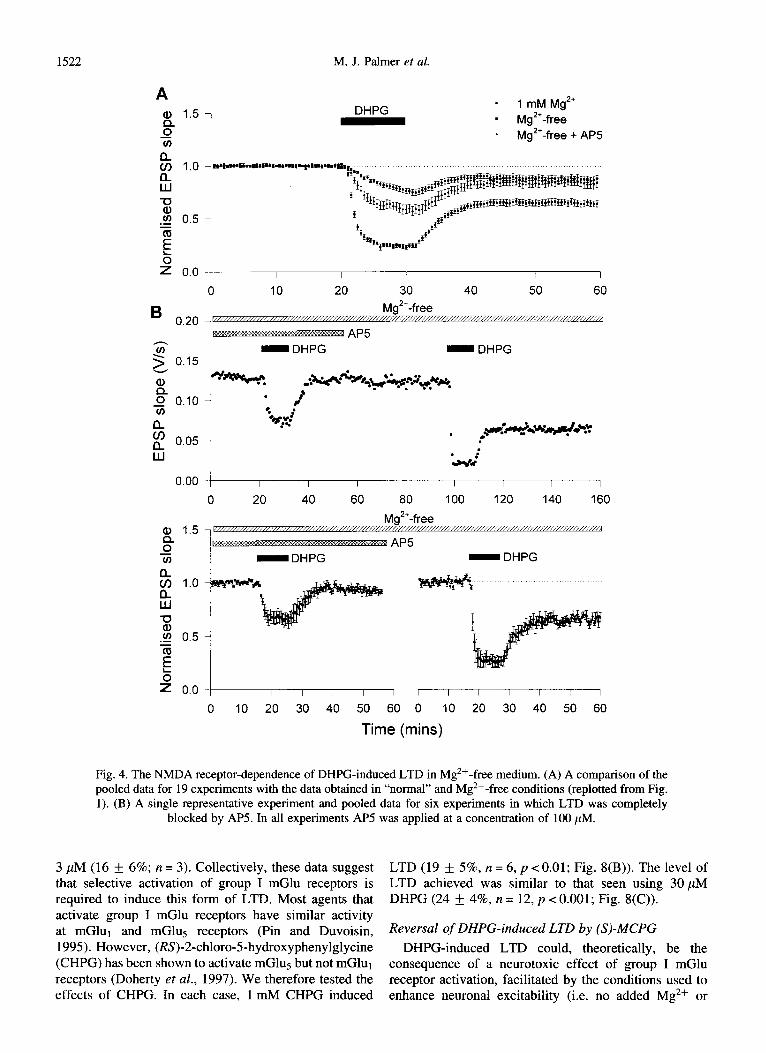
were not performed in normal medium because of the low incidence and small size of DHPG-induced LTD. In Mg2+-free medium, AP5 reduced the average level of
LTD induced to that observed in the presence of 1 mM Mg2+ (Fig. 4(A)), suggesting that the predominant effect of removing Mg2+, with respect to the facilitation of LTD, was an unblocking of the NMDA receptor system. In 6/7 slices in which AP5 was able to fully prevent the induction of DHPG-induced LTD in Mg2+-free medium, a significant DHPG-induced LTD was observed after washout of AP5 (Fig. 4(B)).
In contrast, in 1 mM Mg2+-containing medium to

Group I mGlu receptor-induced LTD 1521
A 0.25
Picrotoxin
*5;‘ 0.2’0 DHPG
I -
DHPG
2
v -
0 20 40 60 100 120 140 160
Picrotoxin
f 0.0 , I I I I I I
0 10 20 30 40 50 60
DHPG
I I I I I I I I
0 10 20 30 40 50 60 70
Time (mins)
Fig. 3. Enhancement of DHPG-induced LTD by the blockade of GABA* receptor-mediated synaptic inhibition. A single representative experiment (A) and pooled data for six experiments (B). In most experiments, perfusion with picrotoxin (50 PM) led to the characteristic enhancement of synaptic transmission (Herron et al., 1985). In these experiments, the stimulus intensity was reset (V) to roughly match the initial response before a new baseline was
obtained.
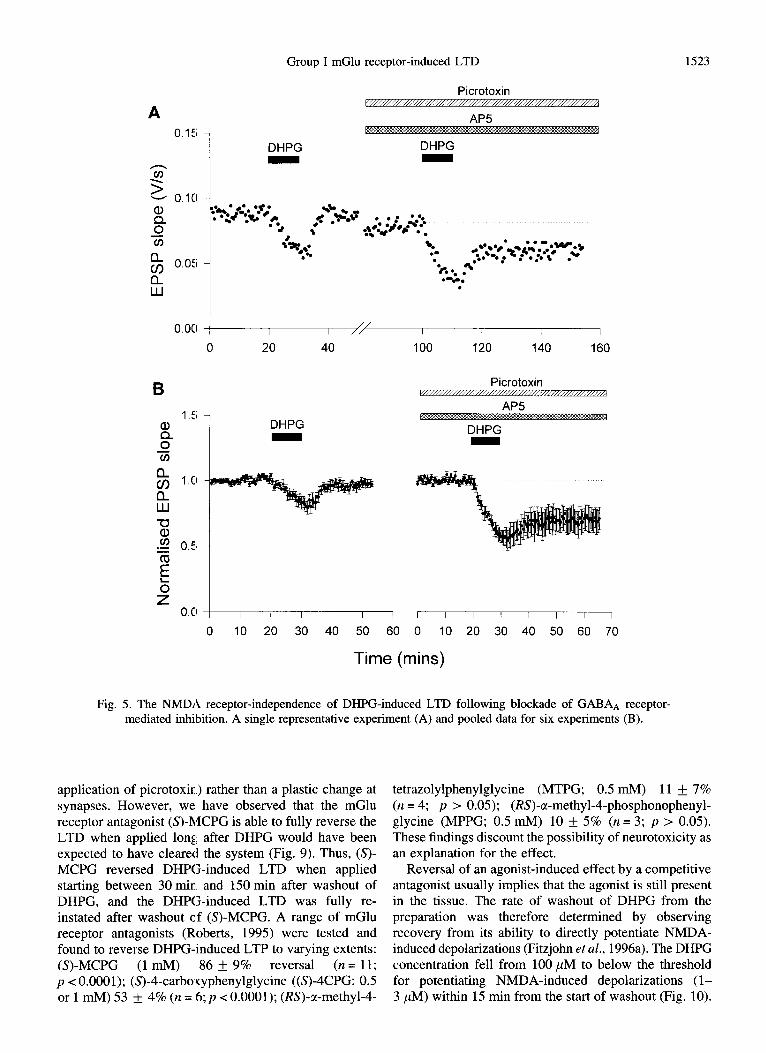
which picrotoxin had been added, LTD was readily induced in the presence of AP5 (Fig. 5). A summary of the likelihood of induction and level of DHPG-induced LTD in the presence or absence of Mg2+, picrotoxin and
AP5 is presented in Fig. 6.
Actions of other mGlu receptor agonists
To determine the specificity of DHPG at inducing LTD, a range of mGlu receptor agonists were compared in Mg2+-free medium. In contrast to DHPG, the highly potent group II agonist (2S,l’R,2’R,3’R)-2-(2’,3’-dicar- boxycyclopropyl)glycine (DCG-IV; 100 nM; Ishida et al., 1993) had no acute or long-term effects on synaptic transmission (Fig. 7). Similarly, the highly potent and
selective group II mGlu receptor agonist (lS,2&5R,6S)- 2-aminobicyclo[3.1 .O]hexane-2,6_dicarboxylate mono-
hydrate (LY354740; 1 FM; Schoepp et al., 1997) was also without any effect (data not shown). The group III agonist (S)-2-amino-4-phosphonobutanoic acid (L-AP4; 10 PM) also did not induce LTD (Fig. 7). A surprising
observation was that (lS,3R)-ACPD (30-100 ,uM), which activates group I mGlu receptors as well as other mGlu receptor subtypes (Pin and Duvoisin, 1995), was also unable to induce LTD (cf. Overstreet et al., 1997), even though it induced a similar initial depression to that obtained using DHPG (Fig. 7).
The threshold concentration of DHPG required to induce LTD was between 1 PM (no effect; IZ = 2) and
1522 M. J. Palmer et al.
A a, 1.5
E u, % 1.0 z 8 .cn 0.5 75 E 8 z 0.0
B 0.20
s 2 0.15
B _o 0.10 cn
% E? 0.05
0.00
Q) 1.5
E 7z
% 1.0
El
B .v, 0.5 z E i7 z 0.0
DHPG . 1 mM Mg*’ . Mg*‘-free
A Mg*‘-free + AP5
I I I I I I
0 10 20 30 40 50 60
Mg*‘-free 1+
AP5
- DHPG - DHPG
I I I I I I I I I
0 20 40 60 80 100 120 140 160
Mg*‘-free
AP5 1+ - DHPG - DHPG
I I I I I I I r I I I I I
0 10 20 30 40 50 60 0 10 20 30 40 50 60
Time (mins)
Fig. 4. The NMDA receptor-dependence of DHPG-induced LTD in Mg 2+-free medium. (A) A comparison of the pooled data for 19 experiments with the data obtained in “normal” and Mg 2+-free conditions (replotted from Fig. 1). (B) A single representative experiment and pooled data for six experiments in which LTD was completely
blocked by AP5. In all experiments AP5 was applied at a concentration of 100 PM.
3 FM (16 + 6%; n = 3). Collectively, these data suggest that selective activation of group I mGlu receptors is required to induce this form of LTD. Most agents that activate group I mGlu receptors have similar activity at mGlui and mGlus receptors (Pin and Duvoisin, 1995). However, (KS)-2-chloro-Shydroxyphenylglycine (CHPG) has been shown to activate mGlus but not mGlul receptors (Doherty et al., 1997). We therefore tested the effects of CHPG. In each case, 1 mM CHPG induced
LTD (19 f 5%, 12 = 6, p < 0.01; Fig. 8(B)). The level of LTD achieved was similar to that seen using 30 ,uM DHPG (24 + 4%, n = 12, p <O.OOl; Fig. 8(C)).
Reversal of DHPG-induced LTD by (S)-MCPG
DHPG-induced LTD could, theoretically, be the consequence of a neurotoxic effect of group I mGlu receptor activation, facilitated by the conditions used to enhance neuronal excitability (i.e. no added Mg2+ or
Group I mGlu receptor-induced LTD 1523
Picrotoxin
DHPG
AP5
DHPG
0.00 / I 1 I I
0 20 40 100 120 140 160
Picrotoxin
DHPG
AP5
DHPG
% 1 .o
Ei u
g .- 0.5 ?? E z z
0.0 I I I I I I I I
0 10 20 30 40 50 60 0 10 20 30 40 50 60 70
Time (mins)
Fig. 5. The NMDA receptor-independence of DHPG-induced LTD following blockade of GABA* receptor- mediated inhibition. A single representative experiment (A) and pooled data for six experiments (B).
application of picrotoxin) rather than a plastic change at synapses. However, we have observed that the mGlu receptor antagonist (S)-MCPG is able to fully reverse the LTD when applied long after DHPG would have been expected to have cleared the system (Fig. 9). Thus, (s>- MCPG reversed DHPG-induced LTD when applied starting between 30 min and 150 min after washout of DHPG, and the DHPG-induced LTD was fully re- instated after washout cd (S)-MCPG. A range of mGlu receptor antagonists (Roberts, 1995) were tested and found to reverse DHPG-induced LTP to varying extents: (S)-MCPG (1 mM) 86 f 9% reversal (n = 11; p < 0.0001); (S)-4-carboxyphenylglycine ((S)4CPG; 0.5 or 1 mM) 53 f 4% (n = 6; p < 0.0001); (RS)-a-methyl4-
tetrazolylphenylglycine (MTPG; 0.5 mM) 11 f 7% (n = 4; p > 0.05); (KS)-a-methyl-4-phosphonophenyl- glycine (MPPG; 0.5 n&l) 10 f 5% (n = 3; p > 0.05). These findings discount the possibility of neurotoxicity as an explanation for the effect.
Reversal of an agonist-induced effect by a competitive antagonist usually implies that the agonist is still present in the tissue. The rate of washout of DHPG from the preparation was therefore determined by observing recovery from its ability to directly potentiate NMDA- induced depolarizations (Fitzjohn et al., 1996a). The DHPG concentration fell from 100 ,uM to below the threshold for potentiating NMDA-induced depolarizations (l- 3 PM) within 15 min from the start of washout (Fig. 10).
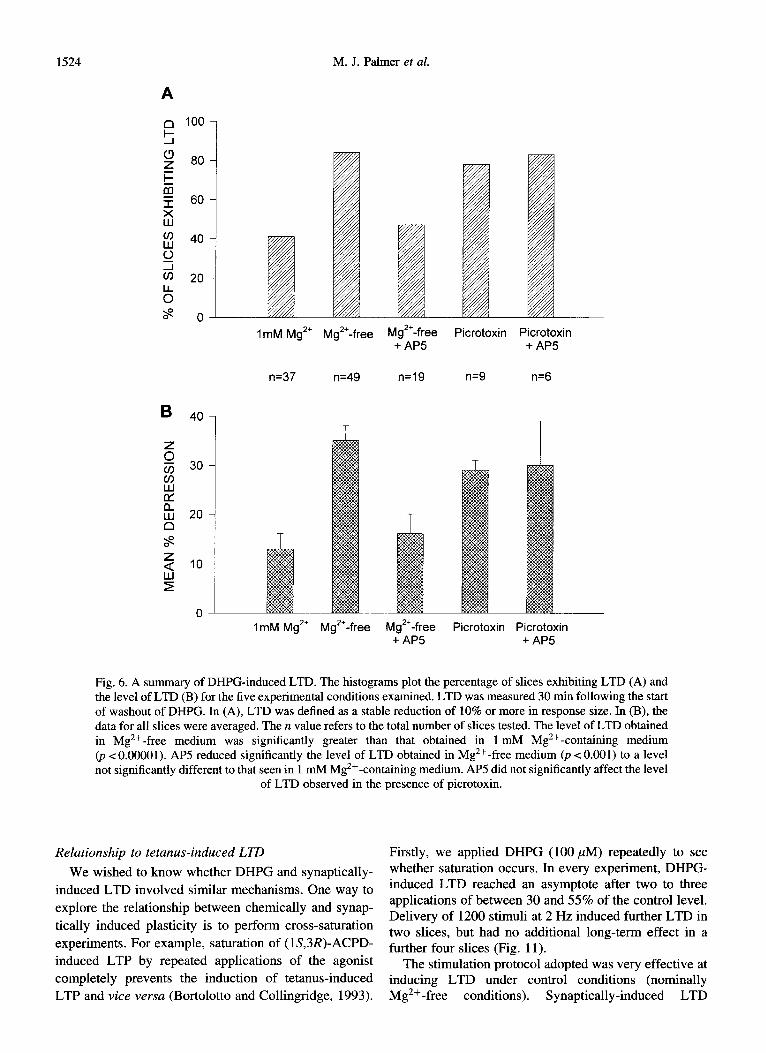
1524 M. J. Palmer et al.
ImM Mg2+ M$+-free M+gl;fee Picrotoxin P~$05Xin
n=37 n=49 n=19 n=9 n=6
ImM Mg” Mg”-free Mg*‘-free Picrotoxin Picrotoxin + AP5 + AP5
Fig. 6. A summary of DHPG-induced LTD. The histograms plot the percentage of slices exhibiting LTD (A) and the level of LTD (B) for the five experimental conditions examined. LTD was measured 30 min following the start of washout of DHPG. In (A), LTD was defined as a stable reduction of 10% or more in response size. In (B), the data for all slices were averaged. The n value refers to the total number of slices tested. The level of LTD obtained in Mg’+-free medium was significantly greater than that obtained in 1 mM Mg2+-containing medium @ < 0.00001). AP5 reduced significantly the level of LTD obtained in Mg 2+-free medium (p < 0.001) to a level not significantly different to that seen in 1 mM Mg2+ -containing medium. AP5 did not significantly affect the level
of LTD observed in tbe presence of picrotoxin.
Relationship to tetanus-induced LTD
We wished to know whether DHPG and synaptically-
induced LTD involved similar mechanisms. One way to
explore the relationship between chemically and synap-
tically induced plasticity is to perform cross-saturation
experiments. For example, saturation of (lS,3R)-ACPD-
induced LTP by repeated applications of the agonist
completely prevents the induction of tetanus-induced
LTP and vice versa (Bortolotto and Collingridge, 1993).
Firstly, we applied DHPG (100 PM) repeatedly to see
whether saturation occurs. In every experiment, DHPG- induced LTD reached an asymptote after two to three applications of between 30 and 55% of the control level. Delivery of 1200 stimuli at 2 Hz induced further LTD in two slices, but had no additional long-term effect in a further four slices (Fig. 11).
The stimulation protocol adopted was very effective at inducing LTD under control conditions (nominally Mg*+-free conditions). Synaptically-induced LTD
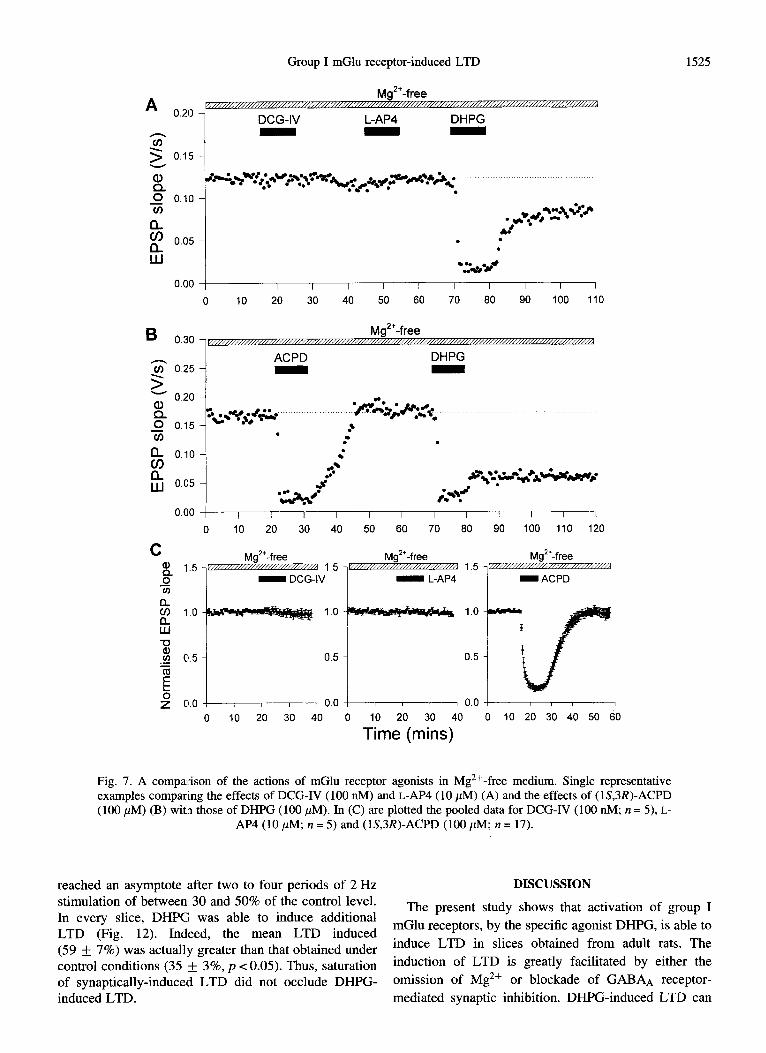
Group I mGlu receptor-induced LTD 1525
A Mg*‘-free
0.21 DCG-IV
z 2 0.15
I -
LAP4 DHPG
0.00 / I I I I I I I I I I I
0 IO 20 30 40 50 60 70 80 90 100 110
B 0.30 , Mg”-free
s 0.25 ACPD DHPG
5 ; 0.20
z 0.15 I - *#+y..-- ~
.r WI .
#T .
0.00 ) I I I I I I I I I I I I
0 lo 20 30 40 50 60 70 80 90 100 110 120
C $ 1.5
Mg*'-free Mg*'-free I 1.5
0 - DCG-IV 1
- L-AP4 TJ
ii 1.0 1.0
ii
$ ,u, 0.5 0.5 i+ E b 2 0.0 0.0
1.5 - Mg2'-free
-ACPD
0 10 20 30 40 0 10 20 30 40 0 10 20 30 40 50 60
Time (mins)
Fig. 7. A comparison of the actions of mGlu receptor agonists in Mg *+-free medium. Single representative examples comparing the effects of DCG-IV (100 nM) and L-AP4 (10 PM) (A) and the effects of (lS,3R)-ACPD (100 PM) (B) with those of DI-WG (100 PM). In (C) are plotted the pooled data for DCG-IV (100 nM; n = 5), L-
AI’4 (10 PM; it = 5) and (lS,3R)-ACPD (100 PM; n = 17).
reached an asymptote after two to four periods of 2 Hz
stimulation of between 30 and 50% of the control level.
In every slice, DHPG was able to induce additional LTD (Fig. 12). Indeed, the mean LTD induced
(59 f 7%) was actually greater than that obtained under control conditions (35 + 3%, p c 0.05). Thus, saturation of synaptically-induced LTD did not occlude DHPG-
induced LTD.
DISCUSSION
The present study shows that activation of group I
mGlu receptors, by the specific agonist DHPG, is able to
induce LTD in slices obtained from adult rats. The
induction of LTD is greatly facilitated by either the
omission of Mg2+ or blockade of GABA* receptor-
mediated synaptic inhibition. DHPG-induced LTD can
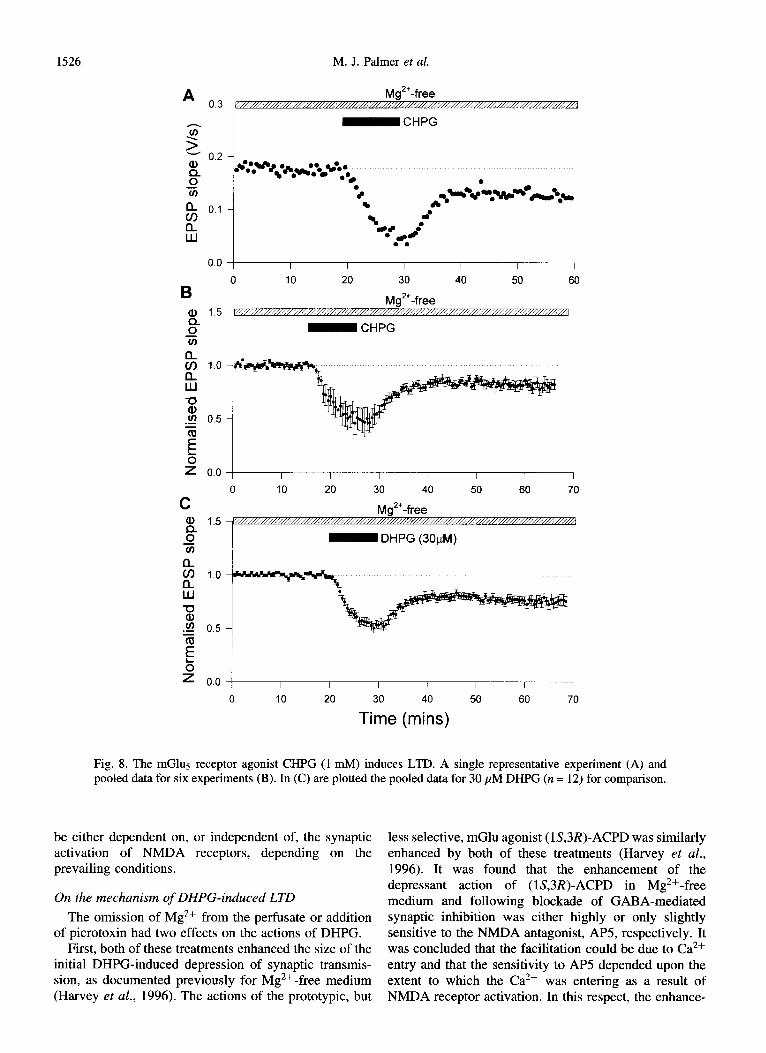
1526 M. J. Palmer et al.
A 0.3
-z-
2 0.2
4 Yz
a if 0.1
w
0.0
B
g 1.5
0 ii
$ 1.0
z
8 .g 0.5
3 E
b 2 0.0
C g 1.5
0 Yi
% 1.0
z
$ .cn 0.5 Yfl E
8
= 0.0
lL Mg”-free /N//////,,,,,,,,,A
- CHPG
I I I I I I I
0 10 20 30 40 50 60
Mg”-free
- CHPG
I I I I I I I I
0 10 20 30 40 50 60 70
Mg”-free
7-P - DHPG (3OkM)
I
0 I
10
I I I I I 1
20 30 40 50 60 70
Time (mins)
Fig. 8. The mGlu5 receptor agonist CHUG (1 mh4) induces LTD. A single representative experiment (A) and pooled data for six experiments (B). In (C) are plotted the pooled data for 30 PM DHPG (n = 12) for comparison.
be either dependent on, or independent of, the synaptic activation of NMDA receptors, depending on the prevailing conditions.
On the mechanism of DHPG-induced LTD
The omission of Mg ‘+ from the perfusate or addition of picrotoxin had two effects on the actions of DHPG.
First, both of these treatments enhanced the size of the initial DHPG-induced depression of synaptic transmis- sion, as documented previously for Mg2+-free medium (Harvey et al., 1996). The actions of the prototypic, but
less selective, mGlu agonist (l&3@-ACPD was similarly enhanced by both of these treatments (Harvey et al., 1996). It was found that the enhancement of the depressant action of (lS,3R)-ACPD in Mg2+-free medium and following blockade of GABA-mediated synaptic inhibition was either highly or only slightly sensitive to the NMDA antagonist, AP5, respectively. It was concluded that the facilitation could be due to Ca2+ entry and that the sensitivity to AP5 depended upon the extent to which the Ca2+ was entering as a result of NMDA receptor activation. In this respect, the enhance-

Group I mGlu receptor-induced LTD 1527
A 015
+z 2 Q 0 10
3 Q_ 005
i 0 00
B 020
-Z
2 0.15
g 0 0.10
Z
: 0.05
ti 0.00
C
g ,I.5
3
ii ,I.0
k
m $j 0.5
.- 7s E 6 0.0
z
Mg”-free
1 (S)-MCPG DHPG (S)-MCPG
. =A
. . ?D
I
0
I I I I
40 60 80 100
Mg*‘-free
MTPG MPPG (S)-MCPG
0 20 40 60 80 100 120 140 160 180 200 220
Mg”-free
DHPG (S)-MCPG
0 20 40 60 80 100
Time (mins)
Fig. 9. Reversal of DHPG-induced LTD by (S)-MCPG. (A) A single representative experiment showing the effects of (S)-MCPG (1 n&l) before and following the induction of LTD by DHPG. (B) A single experiment comparing the effects of four mGlu receptor antagonists (1 mM (S)4CPG, 0.5 mM MTPG, 0.5 mM MPPG and 1 mM (5’)- MCPG) on DHPG-induced LTD. (C) Pooled data for the five experiments where (S)-MCPG was applied for
15 min commencing 30 min following washout of DHPG.
ment of synaptic transmission in Mg2+-free medium is Second, omission of Mg2+ from the perfusate or largely, but not totally, antagonized by AP5 (Coan and addition of picrotoxin greatly facilitated the likelihood Collingridge, 1985), whilst the enhancement of synaptic and magnitude of DHPG-induced LTD. Again, the AP5 transmission following blockade of GABA* receptor- sensitivity of DHPG-induced LTD paralleled the AP5 mediated synaptic inhibition is only slightly sensitive to sensitivity of the initial DHPG-induced depression, AP5 (Herron et al., 1985). A similar explanation could suggesting a similar Ca2+-dependence. That is, under account for the varying AP5 sensitivity of the DHPG- conditions (e.g. Mg2+-free medium) where most Ca2+ induced initial depression of synaptic transmission entry is a consequence of NMDA receptor activation, observed in the present study. then AP5 can prevent the induction of LTD. However,

1528 M. J. Palmer et al.
z DHPG Y 0
I
10 min
Fig. 10. Rapid washout of the effects of DHPG. The trace is a DC recording showing depolarizations induced by 20 PM NMDA (applied for 2 min at times indicated by arrowheads). Note the rapid recovery from the potentiation
induced by 100 PM DHPG. (The potentiation was sustained if DHPG was not washed out.)
conditions can be reached (e.g. addition of picrotoxin) where sufficient Ca*+ enters without the need for NMDA receptor activation and hence AP5 has no effect. Consistent with this hypothesis, APS-insensitive LTD induced by (lS,3R)-ACPD has been described in the dentate gyrus of adult rats following blockade of GABA* receptor-mediated inhibition (O’Mara ef al., 1995).
Although the similarities in the sensitivity to Mg*+- free conditions, picrotoxin and AP5 of the DHPG- induced initial synaptic depression and the DHPG- induced LTD suggest a similar underlying mechanism, the two effects of DHPG could be separated. Thus, on occasion DHPG induced large synaptic depressions but no LTD in Mg*+-free medium (see Harvey et al., 1996). The simplest explanation is that there is an additional requirement for DHPG to induce LTD which, for unknown reasons, was not met in some slices. In this scenario, a single mechanism would enable DHPG to both depress synaptic transmission acutely (above that which results from postsynaptic depolarization; Davies et al., 1995; Gereau and Conn, 1995) and to induce LTD (provided the other condition(s) had been met). The possibility that this mechanism involves a Ca*+-depen- dent coupling between mGlu receptors and their signal- ling cascade (e.g. at the level of phospholipase C) has been discussed previously (Harvey et al., 1996).
We have not attempted to address the locus of expression of DHPG-induced LTD in adult tissue. However, in parallel experiments on slices obtained from young (13-14-day-old) rats we have found that DHPG induces LTD by a mechanism that appears to be presynaptic (Fitzjohn et al., 1996a, 1996b).
Pharmacology of DHPG-induced LTD
The finding that neither DCG-lV, LY354740 nor L- AP4 was able to induce LTD suggests that group I mGlu receptors are specifically involved in the generation of this form of LTD. Of course, these results do not preclude a role for group II or III mGlu receptors in LTD in other
pathways or in area CA1 under different experimental conditions. It also does not exclude an involvement of mGlu receptors, such as mGlu7, that will not be activated by these concentrations of agonists.
A surprising observation is that (lS,3R)-ACPD was not able to induce LTD, since this prototypic mGlu receptor agonist is of similar potency to DHPG at activating group I mGlu receptors (Schoepp et al., 1994). Furthermore, (l&3@-ACPD has been found to induce LTD in area CA1 under other experimental conditions (Overstreet et al., 1997; Bolshakov and Siegelbaum, 1994). Future experiments will address the reason for the difference between (lS,3R)-ACPD and DHPG in inducing LTD under the conditions of the present experiments. (l&3@- ACPD has also been found to induce LTP under certain conditions (e.g. Bortolotto and Coilingridge, 1993, 1995; Collins and Davies, 1993; Overstreet et al., 1997). The failure to observe LTP in the present experiments may be due to one or more factors, such as the removal of the CA3 region of the slice (Bortolotto and Collingridge, 1993).
The possibility that DHPG is acting via a mechanism independent of group I mGlu receptors seems unlikely, since it appears to be a specific agonist (Schoepp et al., 1994) and is effective at inducing LTD, in a dose- dependent manner, over the concentration range at which it activates mGlut and mGlus receptors. Furthermore, CHPG mimicked the effects of DHPG at a concentration that would suggest that both agents were having their effects via mGlus receptors. Finally, the potent mGlu receptor antagonist LY341495 (Ornstein et al., 1997) prevented the induction of LTD induced by DHPG (unpublished observations).
Reversal of DHPG-induced LTD by (S)-MCPG
The finding that (S)-MCPG reversed DHPG-induced LTD was unexpected. The possibility that DHPG washes out very slowly from the tissue and that the (S)-MCPG reversal of DI-IPG-induced LTD was due to competition
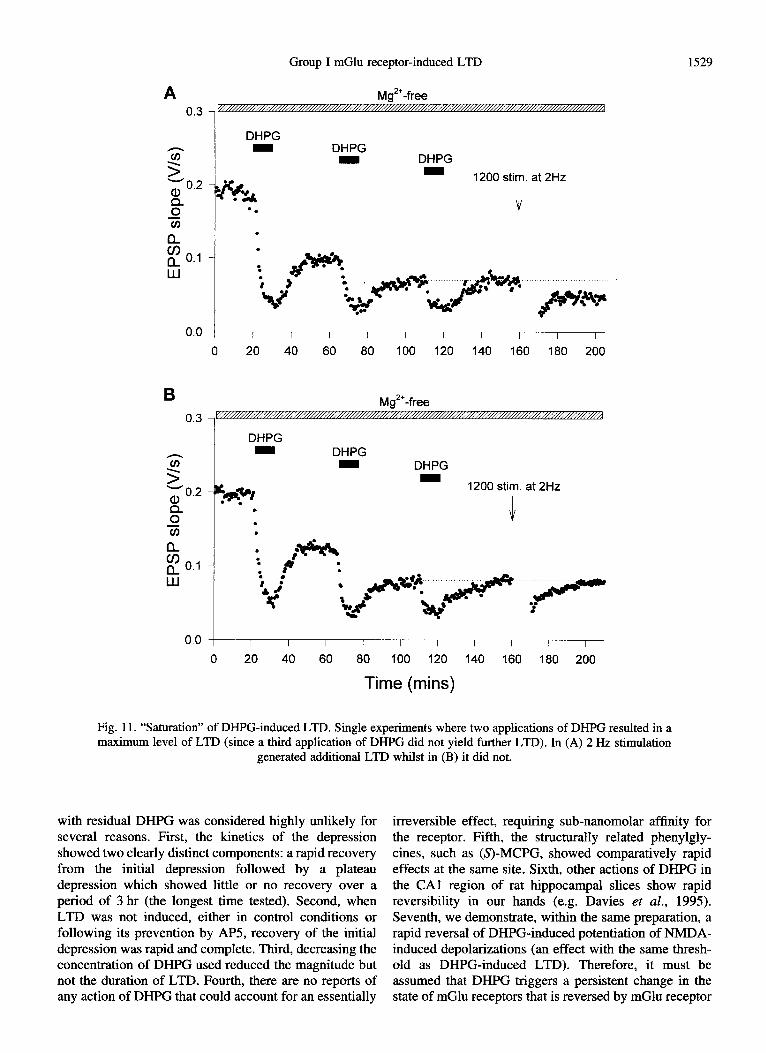
Group I mGlu receptor-induced LTD 1529
Mg*‘-free
DHPG DHPG
- -
1200 stim. at 2Hz
0.0 / I I I I I I I I I I
0 20 40 60 80 100 120 140 160 180 200
B Mg’+-free
0.3 ,
DHPG
1200 dim. at 2Hz
00 I I I I I I I I I I I
0 20 40 60 80 TOO 120 140 160 180 200
Time (mins)
Fig. 11. “Saturatio:n” of DHPG-induced LTD. Single experiments where two applications of DHPG resulted in a maximum level of LTD (since a third application of DHPG did not yield further LTD). In (A) 2 Hz stimulation
generated additional LTD whilst in (B) it did not.
with residual DHPG was considered highly unlikely for several reasons. First, the kinetics of the depression showed two clearly distinct components: a rapid recovery from the initial depression followed by a plateau depression which showed little or no recovery over a period of 3 hr (the longest time tested). Second, when LTD was not induced, either in control conditions or following its prevention by AP5, recovery of the initial depression was rapid and complete. Third, decreasing the concentration of DHPG used reduced the magnitude but not the duration of LTD. Fourth, there are no reports of any action of DHPG that could account for an essentially
irreversible effect, requiring sub-nanomolar affinity for the receptor. Fifth, the structumhy related phenylgly- tines, such as (S)-MCPG, showed comparatively rapid effects at the same site. Sixth, other actions of DHPG in the CA1 region of rat hippocampal slices show rapid reversibility in our hands (e.g. Davies et al., 1995). Seventh, we demonstrate, within the same preparation, a rapid reversal of DHPG-induced potentiation of NMDA- induced depolarizations (an effect with the same tbresh- old as DHPG-induced LTD). Therefore, it must be assumed that DHPG triggers a persistent change in the state of mGlu receptors that is reversed by mGlu receptor
1530 M. J. Palmer et al.
A Mg”-free 0.20
1200 at 2 Hz 1200 at 2 Hz
i 1200at2Hz
4
DHPG
0.00 / I I I I I I I I I I I
0 20 40 60 80 100 120 140 160 180 200 220
Mg”-free
1200 at 2 Hz
z 1200 at 2 Hz g DHPG 1.0
m
z u
2 G 0.5
E
8 Z
0.0 0 20 40 60 160 180 200 220 240 260 280 300
Time (mins)
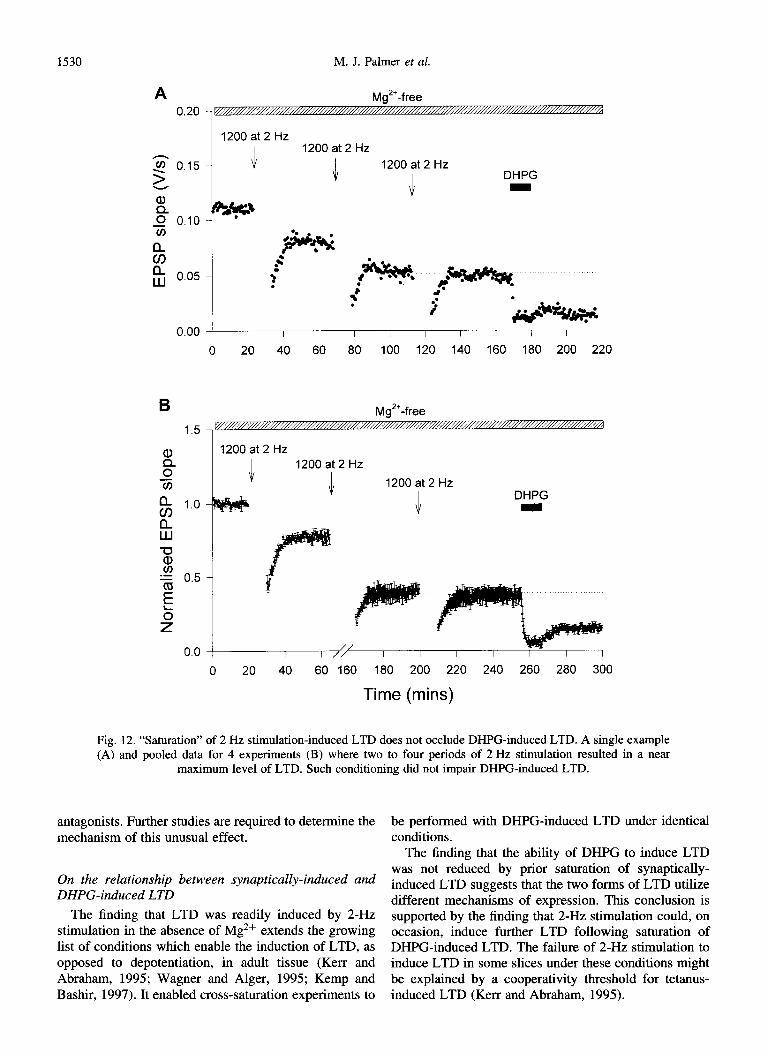
Fig. 12. “Saturation” of 2 Hz stimulation-induced LTD does not occlude DHPG-induced LTD. A single example (A) and pooled data for 4 experiments (B) where two to four periods of 2 Hz stimulation resulted in a near
maximum level of LTD. Such conditioning did not impair DHPG-induced LTD.
antagonists. Further studies are required to determine the be performed with DHPG-induced LTD under identical mechanism of this unusual effect. conditions.
On the relationship between synaptically-induced and DHPG-induced LTD
The finding that LTD was readily induced by ~-HZ stimulation in the absence of Mg2+ extends the growing list of conditions which enable the induction of LTD, as opposed to depotentiation, in adult tissue (Kerr and Abraham, 1995; Wagner and Alger, 1995; Kemp and Bashir, 1997). It enabled cross-saturation experiments to
The finding that the ability of DHPG to induce LTD was not reduced by prior saturation of synaptically- induced LTD suggests that the two forms of LTD utilize different mechanisms of expression. This conclusion is supported by the finding that ~-HZ stimulation could, on occasion, induce further LTD following saturation of DHPG-induced LTD. The failure of ~-HZ stimulation to induce LTD in some slices under these conditions might be explained by a cooperativity threshold for tetanus- induced LTD (Kerr and Abraham, 1995).

Group I mGlu receptor-induced LTD 1531
The relative importance of mGlu receptors and NMDA receptors in synaptically-evoked LTD in area CA1 of the hippocampus is highly variable from preparation to preparation (see Introduction). Recent evidence suggests that two independent forms of synaptically-induced LTD, one induced by mGlu and the other by NMDA receptor activation, can coexist in area CA1 (Oliet et aZ., 1997). This raises the possibility that a form of synaptically- induced LTD may be found which could occlude with DHPG-induced LTD. The findings that AP5 can prevent DHPG-induced LTD and (lS,3R)-ACPD-induced LTD (Overstreet et al., 1997) means, however, that mGlu and NMDA receptor-dependent forms of LTD should not necessarily be viewed as fully independent processes.
CONCLUDING REMARKS
We have shown that a form of LTD can be readily induced in area CA1 of slices obtained from adult animals by the application of mGlu receptor agonists. A preliminary pharmacological characterization of this effect suggests the involvement of mGlu5 receptors. We have also shown striking differences in the NMDA receptor dependence of this form of LTD, depending on the experimental conditions used, and suggest that this simply reflects the way in which Ca2+ is elevated in the neurones that undergo ILTD.
Acknowledgements-Supported by the MRC, the European Economic Community (BMH4CT960228) and The Wellcome Trust. Some of these experiments were performed at the Department of Pharmacology, The Medical School, University of Birmingham, Birmingham B15 2TT, U.K.
REFERENCES
Anderson W. W. and Collingridge G. L. (1997) A data acquisition program for on-line analysis of long-term potentiation and long-term depression. Society for Neuro- science Abstracts 264: 19.
Barrionuevo G., Schottler F. and Lynch G. (1980) The effects of repetitive low frequency stimulation on control and “potentiated” synaptic responses in the hippocampus. Life Sciences 27: 2385-239 1.
Bashir Z. I., Jane D. E., Sunter D. C., Watkins J. C. and Collingridge G. L. (1993) Metabotropic glutamate receptors contribute to the induction of long-term depression in the CA1 region of the hippocampus. European Journal of Pharmacology 239: 265-266.
Bashir Z. I. and Collingridge G. L. (1994) An investigation of depotentiation of long-term potentiation in the CA1 region of the hippocampus. Experimental Brain Research 100: 437- 443.
Bear M. F. and Abraham W. C. (1996) Long-term depression in hippocampus. Annual Review of Neuroscience 19: 437-462.
Blake J. F., Brown M. W. #and Collingridge G. L. (1988) CNQX blocks acidic amino acid induced depolarisations and synaptic components mediated by non-NMDA receptors in rat hippocampal slices. Neuroscience 89: 182-186.
Bliss T. and Collingridge G. L. (1993) A synaptic model of
memory: long-term potentiation in the hippocampus. Nature
361: 31-39. Bolshakov V. Y. and Siegelbaum S. A. (1994) Postsynaptic
induction and presynaptic expression of hippocampal long- term depression. Science 264: 1148-1152.
Bortolotto Z. A. and Collingridge G. L. (1993) Characterisation of LTP induced by the activation of glutamate metabotropic receptors in area CA1 of the hippocampus. Neuropharma-
cology 32: l-9. Bortolotto Z. A. and Collingridge G. L. (1995) On the
mechanism of long-term potentiation induced by (lS,3R)- l-aminocyclopentane-1,3-dicarboxylic acid (ACPD) in rat hippocampal slices. Neurophannacology 34: 1003-1014.
Coan E. J. and Collingridge G. L. (1985) Magnesium ions block an N-methyl-D-aspartate receptor-mediated component of synaptic transmission in rat hippocampus. Neuroscience Letters 53: 21-26.
Collins D. R. and Davies S. N. (1993) Co-administration of (lS,3R)-ACPD and arachidonic acid potentiates synaptic transmission in rat hippocampal slices. European Journal of Pharmacology 240: 325-326.
Davies C. H., Clarke V. R. J., Jane D. E. and Collingridge G. L. (1995) Pharmacology of postsynaptic metabotropic gluta- mate receptors in rat hippocampal CA1 pyramidal neurones. British Journal of Pharmacology 116: 1859-1869.
Doherty A. J., Palmer M. J., Henley J. M., Collingridge G. L. and Jane D. E. (1997) (RS)-2-Chloro-5_hydroxyphenylgly- tine (CHPG) activates mGlu5, but not mGlu1, receptors expressed in CHO cells and potentiates NMDA responses in the hippocampus. Neuropharmacology 36: 265-267.
Dudek S. M. and Bear M. F. (1992) Homosynaptic long-term depression in area CA1 of the hippocampus and the effects of NMDA receptor blockade. Proceedings of the National Academy of Sciences U.S.A. 89: 43634367.
Fitzjohn S. M., Irving A. J., Palmer M. J., Harvey J., Lodge D. and Collingridge G. L. (1996a) Activation of group I mGluRs potentiates NMDA responses in rat hippocampal slices. Neuroscience Letters 203: 211-213.
Fitzjohn, S. M., Lodge, D. and Collingridge, G. L. (1996b) Evidence for a presynaptic locus of expression of LTD induced by group I mGluRs in the CA1 region of the immature rat hippocampus. Society for Neuroscience Abstracts 686.3.
Fujii S., Saito K., Miyakawa H., Ito K. and Kato H. (1991) Reversal of long-term potentiation (depotentiation) induced by tetanus stimulation of the input to CA1 neurons of guinea pig hippocampal slices. Brain Research 555: 112-122.
Gereau R. W. and Conn P. J. (1995) Roles of specific metabotropic glutamate receptor subtypes in regulation of hippocampal CA1 pyramidal cell excitability. Journal of Neurophysiology 74: 122-129.
Harvey J. and Collingridge G. L. (1993) Signal transduction pathways involved in the acute potentiation of NMDA responses by lS,3R-ACPD in rat hippocampal slices. British Journal of Pharmacology 109: 1085-1090.
Harvey J., Palmer M. J., Irving A. J., Clarke V. R. J. and Collingridge G. L. (1996) NMDA receptor dependence of mGlu-mediated depression of synaptic transmission in the CA1 region of the rat hippocampus. British Journal of Pharmacology 119: 1239-1247.
Herron C. E., Williamson R. and Collingridge G. L. (1985) A selective N-methyl-D-aspartate antagonist depresses epilepti-

1532 M. J. Palmer et al.
form activity in rat hippocampal slices. Neuroscience Letters 61: 255-260.
Ito I., Kohda A., Tanabe S., Hirose E., Hayashi N., Mitsunaga
Irving A. J., Palmer M. J., Harvey J., Seabrook G. R. and
S. and Sugiyama H. (1992) 3,5_Dihydroxyphenylglycine: a
Collingridge G. L. (1996) Long-term depression of synaptic transmission in the adult rat hippocampus by coactivation of
potent agonist of metabotropic glutamate receptors.
NMDA and group I metabotropic glutamate receptors. Journ& of Physiology (London) 495P: 49-50P.
Neuroreport 3: 1013-1016.
Ishida M., Saitoh T., Shimamoto K., Ohfune Y. and Shinozaki H. (1993) A novel metabotropic glutamate receptor agonist: marked depression of monosynaptic excitation in the new- born rat isolated spinal cord. British Journal of Phurma- cology 109: 1169-l 177.
Kemp N. and Bashir Z. I. (1997) NMDA receptor-dependent and-independent long-term depression in the CA1 region of the adult rat hippocampus in vitro. Neurophamtacology 36: 397-399.
Kerr D. S. and Abraham W. C. (1995) Cooperative interactions among afferents govern the induction of homosynaptic long- term depression in the hippocampus. Proceedings of the National Academy of Sciences U.S.A. 92: 11637-l 1641.
Manahan-Vaughan D. and Reymann K. G. (1995) lS,3R- ACPD dose-dependently induces a slow-onset potentiation in the rat hippocampal CA1 region in vivo. Neurophamta- cozogy 34: 1103-l 105.
Mulkey R. M. and Malenka R. C. (1992) Mechanisms underlying induction of homosynaptic long-term depression in area CA1 of the hippocampus. Neuron 9: 967-975.
Oliet S. H. R., Malenka R. C. and Nicoll R. A. (1997) Two distinct forms of long-term depression coexist in CA1 hippocampal pyramidal cells. Neuron 18: 969-982.
O’Mara S. M., Rowan M. J. and Anwyl R. (1995) Metabotropic glutamate receptor-induced homosynaptic long-term depres- sion and depotentiation in the dentate gyrus of the rat hippocampus in vitro. Neurophatmacology 34: 983-989.
Ornstein P. L., Kingston A., Bleisch T. J., Arnold M. B., Wright R. A., Johnson B. G. and Schoepp D. D. (1997) Pharmaco- logical characterization of LY341495, a potent antagonist of metabotropic glutamate receptors. Society for Neuroscience Abstracts 788: 10.
Overstreet L. S., Pastemak J. F., Colley P. A., Slater N. T. and Trommer B. L. (1997) Metabotropic glutamate receptor mediated long-term depression in developing hippocampus. Neuropharmacology 36: 83 l-844.
Pin J.-P. and Duvoisin R. (1995) The metabotropic glutamate receptors: structure and functions. Neuropharmacology 34: l-26.
Roberts P. J. (1995) Pharmacological tools for the investigation of metabotropic glutamate receptors (mGluRs): phenygly- tine derivatives and other selective antagonists-an update. Neuropharmacology 34: 813-819.
Schoepp D. D., Goldsworthy J., Johnson B. G., Salhoff C. R. and Baker S. R. (1994) 3,5_Dihydroxyphenylglycine is a highly selective agonist for phosphoinositide-linked metabo- tropic glutamate receptors in the rat hippocampus. Journal of Neurochemistry 63: 769-772.
Schoepp D. D., Johnson B. G., Wright R. A., Salhoff C. R., Mayne N. G., Wu S., Cockerham S. L., Paul Burnett L., Belegaje R., Bleakman D. and Monn J. A. (1997) LY354740 is a potent and highly selective group II metabotropic glutamate receptor agonist in cells expressing human glutamate receptors. Neurophamuzcology 36: 1-l 1.
Selig D. K., Lee H. K., Bear M. F. and Malenka R. C. (1995) Re-examination of the effects of MCPG on hippocampal LTP, LTD, and depotentiation. Journal of Neurophysiology 74: 1075-1082.
Staubli U. and Lynch G. (1990) Stable depression of potentiated synaptic responses in the hippocampus with l- 5 Hz stimulation. Bruin Research 513: 113-l 18.
Wagner J. J. and Alger B. E. (1995) GABAergic and developmental influences on homosynaptic LTD and depo- tentiation in rat hippocampus. Journal of Neuroscience 15: 1577-1586.