the potent mglu receptor antagonist ly341495 identifies roles for both cloned and novel mglu...
TRANSCRIPT

Neuropharmacology 37 (1998) 1445–1458
The potent mGlu receptor antagonist LY341495 identifies roles forboth cloned and novel mGlu receptors in hippocampal synaptic
plasticity
S.M. Fitzjohn a,*, Z.A. Bortolotto a, M.J. Palmer a, A.J. Doherty a, P.L. Ornstein b,D.D. Schoepp b, A.E. Kingston c, D. Lodge c, G.L. Collingridge a
a Department of Anatomy, Uni6ersity of Bristol, Uni6ersity Walk, Bristol BS8 1TD, UKb Eli Lilly & Co., Lilly Corporate Centre, Indianapolis, IN, USA
c Lilly Research Centre, Eli Lilly & Co., Erl Wood Manor, Windlesham, Surrey GU20 6PH, UK
Accepted 31 July 1998
Abstract
Understanding the roles of metabotropic glutamate (mGlu) receptors has been severely hampered by the lack of potentantagonists. LY341495 (2S-2-amino-2-(1S,2S-2-carboxycyclopropyl-1-yl)-3-(xanth-9-yl)propanoic acid) has been shown to blockgroup II mGlu receptors in low nanomolar concentrations (Kingston, A.E., Ornstein, P.L., Wright, R.A., Johnson, B.G., Mayne,N.G., Burnett, J.P., Belagaje, R., Wu, S., Schoepp, D.D., 1998. LY341495 is a nanomolar potent and selective antagonist at groupII metabotropic glutamate receptors. Neuropharmacology 37, 1–12) but can be used in higher concentrations to block allhippocampal mGlu receptors, identified so far by molecular cloning (mGlu1–5, 7,8). Here we have further characterised the mGlureceptor antagonist activity of LY341495 and have used this compound to investigate roles of mGlu receptors in hippocampallong-term potentiation (LTP) and long-term depression (LTD). LY341495 competitively antagonised DHPG-stimulated PIhydrolysis in AV12-664 cells expressing either human mGlu1 or mGlu5 receptors with Ki-values of 7.0 and 7.6 mM, respectively.When tested against 10 mM L-glutamate-stimulated Ca2+ mobilisation in rat mGlu5 expressing CHO cells, it produced substantialor complete block at a concentration of 100 mM. In rat hippocampal slices, LY341495 eliminated 30 mM DHPG-stimulated PIhydrolysis and 100 mM (1S,3R)-ACPD-inhibition of forskolin-stimulated cAMP formation at concentrations of 100 and 0.03 mM,respectively. In area CA1, it antagonised DHPG-mediated potentiation of NMDA-induced depolarisations and DHPG-inducedlong-lasting depression of AMPA receptor-mediated synaptic transmission. LY341495 also blocked NMDA receptor-independentdepotentiation and setting of a molecular switch involved in the induction of LTP; effects which have previously been shown tobe blocked by the mGlu receptor antagonist (S)-MCPG. These effects may therefore be due to activation of cloned mGlureceptors. In contrast, LY341495 did not affect NMDA receptor-dependent homosynaptic LTD; an effect which may thereforebe independent of cloned mGlu receptors. Finally, LY341495 failed to antagonise NMDA receptor-dependent LTP and, in areaCA3, NMDA receptor-independent, mossy fibre LTP. Since in the same inputs these forms of LTP were blocked by (S)-MCPG,a novel type of mGlu receptor may be involved in their induction. © 1998 Elsevier Science Ltd. All rights reserved.
Keywords: Metabotropic glutamate (mGlu) receptor; 2S-2-Amino-2-(1S,2S-2-carboxycyclopropyl-1-yl)-3-(xanth-9-yl)propanoic acid (LY341495);Synaptic plasticity; Long-term potentiation (LTP); Long-term depression (LTD); Depotentiation
1. Introduction
L-Glutamate, the principal excitatory neurotransmit-ter in the brain, activates a family of G-protein linkedreceptors, termed metabotropic glutamate (mGlu) re-
ceptors, of which eight subtypes have been cloned(Nakanishi and Masu, 1994; Conn and Pin, 1997).However, identification of the roles of these receptorshas been hindered by the lack of potent antagonists(Watkins and Collingridge, 1994). This is typified in thehippocampus where pharmacological studies have in-vestigated the potential roles of mGlu receptors in theinduction of LTP and LTD (Bliss and Collingridge,
* Corresponding author. Tel.: +44 117 9287407; fax: +44 1179291687; e-mail: [email protected].
0028-3908/98/$ - see front matter © 1998 Elsevier Science Ltd. All rights reserved.
PII: S0028-3908(98)00145-2

S.M. Fitzjohn et al. / Neuropharmacology 37 (1998) 1445–14581446
1993; Bear and Abraham, 1996). Of the cloned mGlureceptor subtypes, group I (i.e. mGlu1 and mGlu5 re-ceptors) have been most often implicated in LTP forseveral reasons. Firstly, these receptors couple to PLC(Nakanishi and Masu, 1994; Conn and Pin, 1997) andcan thereby affect Ca2+ signalling and kinases thathave been implicated in the induction of LTP (Bliss andCollingridge, 1993). Secondly, antagonists that haveactions at group I mGlu receptors, in particular (S)-MCPG (Eaton et al., 1993), can block the induction ofLTP (Bashir et al., 1993a; Sergueeva et al., 1993; Aibaet al., 1994; Bortolotto et al., 1994; O’Connor et al.,1994; Richter-Levin et al., 1994; Little et al., 1995;Wang et al., 1995; Manahan-Vaughan, 1997; Breakwellet al., 1998). Thirdly, deficits in LTP have been de-scribed in mice lacking mGlu1 receptors (Aiba et al.,1994; Conquet et al., 1994) and mGlu5 receptors (Lu etal., 1997). However, these data are controversial, since(S)-MCPG does not invariably block the induction ofLTP (Chinestra et al., 1993; Bortolotto et al., 1994;Manzoni et al., 1994; Selig et al., 1995; Thomas andO’Dell, 1995; Wang et al., 1995; Martin and Morris,1997) and since deficits in LTP in area CA1 of themGlu1 receptor knockout were not observed in one ofthe two original reports (Conquet et al., 1994). Asimilar controversy surrounds studies on the roles ofmGlu receptors in LTD with both positive (Bashir etal., 1993b; Bashir and Collingridge, 1994; Bolshakovand Siegelbaum, 1994; Yang et al., 1994; Manahan-Vaughan, 1997; Oliet et al., 1997) and negative (Selig etal., 1995; Oliet et al., 1997) reports on the effectivenessof (S)-MCPG.
Recently, LY341495 has been shown to inhibit allknown mGlu receptor subtypes (except for retina-spe-cific mGlu6 which was not tested) with IC50-valuesranging from 14 nM (mGlu3) to 22 mM (mGlu4)(Kingston et al., 1998). Therefore, LY341495 is the firstcompound that can be used to block the activation ofall the mGlu receptor subtypes in the brain that havebeen identified so far by cloning, and thereby determineto what extent the activation of one or more of thesemGlu receptors is essential for specific functions.
In the present study we have further characterisedLY341495 to determine the effective concentration re-quired to antagonise activation of all mGlu receptors,so far identified by molecular cloning. We have thenused 100 mM LY341495 to determine whether activa-tion of any of these mGlu receptors is necessary for theinduction of both NMDA receptor-dependent andNMDA receptor-independent forms of hippocampalLTP and LTD.
Some of these data have been presented as abstractsat the Forum of European Neuroscience, Berlin (Bor-tolotto et al., 1998; Fitzjohn et al., 1998).
2. Materials and methods
(S)-a-methyl-4-carboxyphenylglycine ((S)-MCPG),(RS)-3,5-dihydroxyphenylglycine (DHPG), N-methyl-D-aspartate (NMDA), (1S,3R)-1-aminocyclopentane-1,3-dicarboxylic acid ((1S,3R)-ACPD) and (D)-2-ami-no-5-phosphopentanoate (AP5) were obtained fromTocris Cookson (Bristol, UK). 2S-2-Amino-2-(1S,2S-2-carboxycyclopropyl-1-yl)-3-(xanth-9-yl)propanoic acid(LY341495) was synthesised as described previously(Kingston et al., 1998).
2.1. Experiments using recombinant receptors
Cell lines stably expressing human mGlu receptorswere derived and [3H]inositol monophosphate accumu-lation (PI turnover) was measured as described previ-ously (Kingston et al., 1995). Briefly, AV12-664 cellswere plated at 2.5×105 cells per well in Dulbecco’smodified Eagle’s medium (DMEM) supplemented with5% dialysed foetal calf serum, 1 mM sodium pyruvate,10 mM HEPES, 1% penicillin/streptomycin, 50 ug/mlG418 and 0.2 mg/ml hygromycin B, maintained at 37°Cin a humidified atmosphere of 5% CO2. After 24 h cellswere labelled with [3H]inositol for a further 20 h.Medium was then replaced with assay medium compris-ing DMEM supplemented with 10 mM HEPES, 10mM inositol and 10 mM lithium chloride. LY341495was added to cultures 20 min prior to DHPG, and cellsincubated for a further 60 min before terminating thereaction by replacing medium with acetone:methanol(1:1). [3H]inositol phosphates were separated by Sep-Pak Accell Plus QMA ion exchange chromatographyand the [3H]inositol monophosphate fraction elutedwith 0.1M triethylammonium bicarbonate buffer.
A CHO cell line stably expressing the rat mGlu5a
receptor was kindly provided by Professor S. Nakanishi(Kyoto) and L-glutamate-evoked Ca2+ release fromintracellular stores measured (Doherty et al., 1997).Briefly, cells were plated onto glass coverslips (22 mmdiameter, thickness 1, 1–2×105 cells per coverslip) inDMEM, (4.5g/l D-glucose) supplemented with 10% di-alysed foetal bovine serum (10000 Mr cut-off, Sigma), 2mM L-glutamine, 1% (w/v) L-proline, 1 mg/l penicillinand 100 U/l streptomycin. After 24–48 h, cells werewashed three times in Krebs buffer containing 4.5g/LD-glucose and 2 mM CaCl2 and loaded with 5 mM ofthe membrane permeable Ca2+ indicator Fluo3-AMfor 20 min. Cells were viewed on a Bio-Rad MRC600confocal microscope equipped with an argon ion laserusing a standard FITC filter set and perfused continu-ously with Krebs buffer at a rate of approximately 2ml/min. Kalman integrations of five individual imageswere obtained every 10 s. L-Glutamate (2 ml) was bathapplied and LY341495 was pre-incubated for 1 minprior to co-application with L-glutamate. The fluores-

S.M. Fitzjohn et al. / Neuropharmacology 37 (1998) 1445–1458 1447
cence of ten individual cells in each preparation wasmeasured using NIH-Image (public domain softwaredeveloped by the US National Institutes of Health andavailable on the Internet at http://rsb.info.nih.gov/nih-image/) and expressed relative to baseline. The mean peakfluorescence was taken as a measure of mGlu receptoractivation.
2.2. Neurochemical assays in hippocampal slices
For rat hippocampal PI hydrolysis studies withLY341495, cross-chopped (0.3 mm) tissue from 7-day-oldrats was incubated at 37°C for 60 min with [3H]myo-inos-itol, washed and pre-incubated for 20 min with buffercontaining 10 mM non-labeled myo-inositol, 10 mMlithium chloride, and LY341495 or water vehicle. DHPGor water vehicle (basal value) was added and tissue wasincubated for an additional 60 min. The [3H]Inositolmonophosphate was isolated by ion exchange columnchromatography as described above. For cAMP studies,cross-chopped (0.3 mm) hippocampal tissue from 45–50-day-old rats was pre-incubated at 37°C for 30 min, thenincubated for 15 min in buffer containing adenosinedeaminase (1 U/ml) in the presence of forskolin (30 mM),1S,3R-ACPD (100 mM), and various concentrations ofLY341495 or water vehicle (control). The incubation wasterminated by adding ice-cold 6 mM disodium EDTA,then immediately homogenizing the tissue. Levels ofcAMP in extracts of this homogenate were determinedusing a commercially available cAMP assay kit (Amer-sham TRK.432).
2.3. Electrophysiology
Experiments were performed on hippocampal slicesobtained from young (12–18 day old) or adult (6–10week old) rats, using extracellular (Bortolotto et al., 1994;Fitzjohn et al., 1996a) or grease-gap (Palmer et al., 1997)recording techniques. Slices were maintained in a mediumcomprising (mM) NaCl (124), KCl (3), NaHCO3 (26),NaH2PO4 (1.25), CaCl2 (2), MgSO4 (1), D-glucose (10),bubbled with O2/CO2: 95/5%. The Schaffer collateral-commissural pathway was stimulated at 30-s intervals (forexperiments using two inputs, stimuli were delivered every15 s to alternate inputs). Individual responses weredisplayed and the average field EPSP slopes of foursuccessive responses were plotted on-line using softwarewritten in-house (Anderson and Collingridge, 1997). TheLTD and depotentiation were induced by delivering 900shocks at 1 Hz, and LTP induced by delivering 100 shocksat 100 Hz at test intensity.
2.4. Data analysis
Data are presented as means9S.E.M. The n-valuesrefer to the number of slices from which a given exper-
iment was performed, each slice having been obtainedfrom a separate rat. Differences were analysed statisti-cally using ANOVA, followed by the appropriate Stu-dent’s t-test.
3. Results
3.1. Quantification of LY341495 as an antagonist athuman mGlu1 and mGlu5 receptors
The initial analysis of the actions of LY341495 atcloned human group I mGlu receptors suggested thatit can be used to block activation of these receptors,as well as group II receptors at which it has nano-molar potency (Kingston et al., 1998). However, it wasnecessary to first establish the potency of LY341495at mGlu1 and mGlu5 receptors. Thus, its ability toantagonise DHPG-stimulated PI hydrolysis in AV12-664cells permanently expressing human mGlu1a or mGlu5a
receptors was quantified using Schild analysis (Fig. 1).LY341495 had similar activity at both mGlu1 (Ki 7.091.2 mM; n=5) and mGlu5 (Ki 7.692.6 mM; n=5)receptors.
3.2. LY341495 antagonises the actions of L-glutamateat rat mGlu5 receptors
Since most LTP and LTD experiments are performedusing rat tissue it was important to determine theeffectiveness of LY341495 using a rat recombinantmGlu receptor subtype. We chose mGlu5 receptors inview of the high concentration of this subtype in areaCA1 of the rat hippocampus (Lujan et al., 1996). We alsoused L-glutamate as the agonist since the effectivenessof mGlu receptor antagonists may be agonist-dependent(Brabet et al., 1995) and L-glutamate is the most likelyendogenous activator of mGlu5 receptors. We usedCa2+ mobilisation from intracellular stores of indivi-dual cells as a functional assay (Bashir et al., 1993a).When tested against a concentration of L-glutamatewhich produced approximately an EC80 response (10mM), LY341495 produced dose-dependent antagonismover the range 10 to 100 mM (Fig. 2), with an estimatedIC50 of 14.694.2 mM (n=4; Hill slope, 1.0290.32).At the maximum concentration of LY341495 tested(100 mM) responses to 10 mM L-glutamate were oftenabolished (Fig. 2A) though in some cells a small (B10%)response remained. In a further two experiments, 10 mML-glutamate produced a maximal response and theantagonism by 100 mM LY341495 was correspondinglyless (responses depressed to 2592% of control),consistent with its competitive mode of action. Thesedata show that LY341495 is similarly effective againsthuman and rat clones and shows no obvious agonistpreference.

S.M. Fitzjohn et al. / Neuropharmacology 37 (1998) 1445–14581448
Fig. 1. LY341495 is a potent mGlu1 and mGlu5 receptor antagonist. (a) Parallel displacement of DHPG-induced inositol monophosphateaccumulation by LY341495 in mGlu5a receptor expressing cells. (b) Corresponding Schild plot. (c, d) Pooled data (n=5) showing competitiveantagonism of DHPG induced inositol monophosphate accumulation in mGlu5a (c) and mGlu1a (d) receptor expressing cells. The Schildcoefficients were 1.190.1 and 0.890.1 for mGlu5a and mGlu1a expressing cell lines, respectively. The symbols represent (�) control, (�) +3 mMLY341495, (") +10 mM LY341495, and (�)+30 mM LY341495.
3.3. LY341495 is an effecti6e mGlu receptor antagonistin hippocampal slices
Before LY341495 could be used to study synapticplasticity in the hippocampus it was necessary to es-tablish its effectiveness as an mGlu receptor antago-nist in the hippocampal slice preparation. LY341495fully antagonised (1S,3R)-ACPD-induced inhibition offorskolin-stimulated cAMP formation at a concentra-tion of 0.03 mM (Fig. 3a, b). When tested against 30mM DHPG-stimulated PI hydrolysis a concentration of100 mM was required for almost complete antagonism(Fig. 3c); Schild analysis yielded a Ki-value of 1.490.5mM (n=4; Fig. 3d; see also Chung et al., 1997; Huberet al., 1997). Basal forskolin-stimulated cAMP produc-tion and PI hydrolysis were unaffected by LY341495(data not shown).
Next, LY341495 was tested as an antagonist ofDHPG-induced potentiation of NMDA-induced depo-larisation (Fitzjohn et al., 1996), an effect believed to bemediated via mGlu5 receptors (Doherty et al., 1997).LY341495 reversibly antagonised the potentiation in-duced by DHPG (10 mM) in a dose-dependent manner(Fig. 3e, f), with an IC50-value of 1.4 mM. These datashow that LY341495 can be used effectively to antago-nise mGlu receptor-mediated events in hippocampalslices. However, although the most potent known an-tagonist at mGlu1 and mGlu5 receptors, with Ki-valuesof less than 10 mM, these experiments show that inorder to ensure complete antagonism of group I mGlureceptors it is necessary to use LY341495 at a relativelyhigh concentration (\30 mM). We used LY341495 at aconcentration of 100 mM to maximise the chances ofcomplete functional blockade.

S.M. Fitzjohn et al. / Neuropharmacology 37 (1998) 1445–1458 1449
Fig. 2. LY341495 antagonises L-glutamate-induced Ca2+ mobilisation in mGlu5 receptor expressing cells. (A) Change in fluorescence intensity ofa single CHO cell expressed as a percentage of baseline in response to 2 ml aliquots of 10 mM L-glutamate delivered in the presence or absenceof LY341495. The antagonist was pre-incubated for 1 min. Breaks in traces correspond to periods when the cells were not illuminated(approximately 15 min between applications). (B) Pooled data for four coverslips showing dose-dependent antagonism by LY341495 of asubmaximal response to 10 mM L-glutamate. (For each coverslip the mean peak fluorescence for ten cells was averaged).
3.4. LY341495 blocks the induction of long-lastingsynaptic depression induced by mGlu receptor acti6ation
To investigate its effects on synaptic plasticity we firsttested the ability of LY341495 to antagonise a form ofsynaptic plasticity induced by pharmacological activa-tion of mGlu receptors in hippocampal slices. It hasbeen shown that DHPG can induce a long-lastingsynaptic depression in the CA1 region of the hippocam-pus (Fitzjohn et al., 1996a; Palmer et al., 1997). In theadult hippocampus this effect is enhanced by treat-ments which increase neuronal excitability, such asMg2+-free medium or picrotoxin, (Palmer et al., 1997)but in slices from young animals such treatment is notnecessary to obtain substantial LTD (Fitzjohn et al.,1996a).
Using slices from 12–18-day-old rats, LY341495eliminated, in a reversible manner, the ability of DHPG
(50 mM) to both acutely depress synaptic transmissionand induce a long-term synaptic depression (Fig. 4).Thus, the peak depression obtained during the applica-tion of 50 mM DHPG and the depression measured 60min following washout of DHPG were 9791% ofcontrol (P\0.05) and 9594% (P\0.05) when DHPGwas applied in the presence of LY341495, compared to5999% (PB0.05) and 8494% (PB0.05) whenDHPG was re-applied 60 min following the start ofwashout of LY341495 (n=8). LY341495 had no effecton basal synaptic transmission, demonstrating thatnone of the cloned mGlu receptors are involved inmodulating basal transmission in the hippocampus.
LY341495 had a similar effect on the acute andlong-lasting synaptic depression induced by DHPG inslices from adult animals perfused with Mg2+-freemedium, recorded using a grease-gap technique (Palmeret al., 1997). Pooled data are illustrated in Fig. 5a. The

S.M. Fitzjohn et al. / Neuropharmacology 37 (1998) 1445–14581450
Fig. 3. LY341495 blocks mGlu receptor activation in rat hippocampal slices. (a, b) Antagonism by LY341495 of 100 mM (1S,3R)-ACPDinhibition of 30 mM forskolin-stimulated cAMP formation in rat hippocampal slices (n=4 for both sets of experiments; *, PB0.05). (c)Antagonism by LY341495 of 30 mM DHPG-stimulated PI hydrolysis in rat hippocampal slices (n=3; *, PB0.05). (d) Competitive antagonismof DHPG-stimulated PI hydrolysis. Each point is the mean of four experiments. (e) Inhibition by 10 mM LY341495 of 10 mM DHPG-inducedpotentiation of 15 mM NMDA-induced depolarisations. Calibration bar, 0.1 mV, 10 min. NMDA was applied as a 2 ml aliquot at times indicatedby the black triangles. (f) The percent inhibition of 10 mM DHPG-induced potentiation of 15–20 mM NMDA-induced depolarisations is plottedversus LY341495 concentration. Each point is the mean of three separate experiments.
peak depression obtained during the application of 30mM DHPG and the depression measured 35 min fol-lowing washout of DHPG were 10091% of control(P\0.05) and 9893% (P\0.05) when DHPG wasapplied in the presence of LY341495 compared to6694% (PB0.05) and 7496% (PB0.05), whenDHPG was re-applied 40 min following the start ofwashout of LY341495 (n=5). An interesting propertyof DHPG-induced LTD is that it can be temporarily
reversed by application of mGlu receptor antagonists,such as (S)-MCPG (Palmer et al., 1997). This enablesthe time-course of the effect of an mGlu receptorantagonist to be determined directly. This is illustratedin Fig. 5b where substantial LTD was induced by threeapplications of DHPG. LY341495 (100 mM) rapidlyreversed the synaptic depression, with almost completereversal after 10 min. An application time of 20 minwas used for all subsequent experiments to ensure a

S.M. Fitzjohn et al. / Neuropharmacology 37 (1998) 1445–1458 1451
Fig. 4. LY341495 blocks DHPG-induced long-lasting synaptic depression. (a) Representative example to show reversible block of 50 mMDHPG-induced LTD. Traces are the average of four successive responses obtained at the times indicated by 1–4. Calibration bar, 0.5 mV, 10 ms.(b) Pooled data (n=8; in three of these experiments LY341495 was applied for an extra 10 min before and following the standard 40-minapplication). Data were normalised with respect to the 20-min periods preceding application of DHPG.
steady-state concentration of LY341495 in the slicepreparation.
3.5. LY341495 does not block NMDAreceptor-dependent homosynaptic LTD
The effects of LY341495 were then examined onNMDA receptor-dependent homosynaptic LTD inslices obtained from young rats (Dudek and Bear, 1992;Mulkey and Malenka, 1992). As illustrated in Fig. 6,homosynaptic LTD was prevented by the specificNMDA receptor antagonist AP5 (50 mM; n=3), butLY341495 (100 mM; n=5) was without effect. Therewas no significant difference between the amounts ofLTD observed (responses depressed to 6593% and7295% of control) 30 min after ceasing 1 Hz stimula-tion delivered in the presence or absence of LY341495,respectively; P\0.05; Fig. 6c).
3.6. LY341495 blocks NMDA receptor-independentdepotentiation
Next, the effects of LY341495 were examined onNMDA receptor-independent depotentiation of pre-es-tablished LTP (Bashir and Collingridge, 1994), in slicesobtained from young rats. In order to study pureNMDA receptor-independent depotentiation it wasnecessary to use AP5 to block NMDA receptor-depen-dent LTD. In contrast to the lack of effect on homosy-naptic LTD, LY341495 (100 mM) prevented theinduction of AP5-resistant depotentiation in a fullyreversible manner (Fig. 7). Thus, 1 Hz stimulationresulted in responses, measured 40 min later, of 10195% of control (AP5 plus LY341495, 1 Hz stimulationstarted 60 min following a tetanus), compared to 7595% of control (AP5 alone, 1 Hz stimulation started165–175 min following the same tetanus; n=5; PB
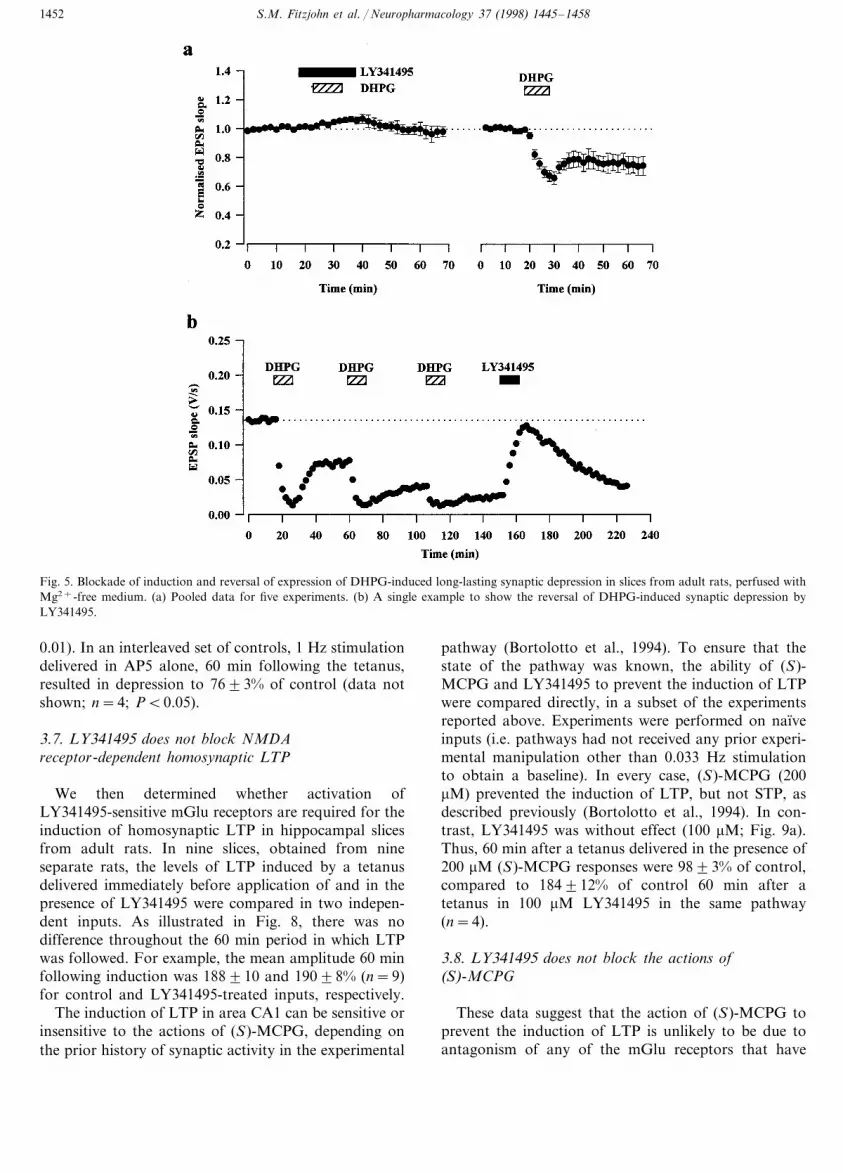
S.M. Fitzjohn et al. / Neuropharmacology 37 (1998) 1445–14581452
Fig. 5. Blockade of induction and reversal of expression of DHPG-induced long-lasting synaptic depression in slices from adult rats, perfused withMg2+-free medium. (a) Pooled data for five experiments. (b) A single example to show the reversal of DHPG-induced synaptic depression byLY341495.
0.01). In an interleaved set of controls, 1 Hz stimulationdelivered in AP5 alone, 60 min following the tetanus,resulted in depression to 7693% of control (data notshown; n=4; PB0.05).
3.7. LY341495 does not block NMDAreceptor-dependent homosynaptic LTP
We then determined whether activation ofLY341495-sensitive mGlu receptors are required for theinduction of homosynaptic LTP in hippocampal slicesfrom adult rats. In nine slices, obtained from nineseparate rats, the levels of LTP induced by a tetanusdelivered immediately before application of and in thepresence of LY341495 were compared in two indepen-dent inputs. As illustrated in Fig. 8, there was nodifference throughout the 60 min period in which LTPwas followed. For example, the mean amplitude 60 minfollowing induction was 188910 and 19098% (n=9)for control and LY341495-treated inputs, respectively.
The induction of LTP in area CA1 can be sensitive orinsensitive to the actions of (S)-MCPG, depending onthe prior history of synaptic activity in the experimental
pathway (Bortolotto et al., 1994). To ensure that thestate of the pathway was known, the ability of (S)-MCPG and LY341495 to prevent the induction of LTPwere compared directly, in a subset of the experimentsreported above. Experiments were performed on naıveinputs (i.e. pathways had not received any prior experi-mental manipulation other than 0.033 Hz stimulationto obtain a baseline). In every case, (S)-MCPG (200mM) prevented the induction of LTP, but not STP, asdescribed previously (Bortolotto et al., 1994). In con-trast, LY341495 was without effect (100 mM; Fig. 9a).Thus, 60 min after a tetanus delivered in the presence of200 mM (S)-MCPG responses were 9893% of control,compared to 184912% of control 60 min after atetanus in 100 mM LY341495 in the same pathway(n=4).
3.8. LY341495 does not block the actions of(S)-MCPG
These data suggest that the action of (S)-MCPG toprevent the induction of LTP is unlikely to be due toantagonism of any of the mGlu receptors that have

S.M. Fitzjohn et al. / Neuropharmacology 37 (1998) 1445–1458 1453
Fig. 6. LY341495 does not affect homosynaptic LTD. (a) A representative example to show reversible block of homosynaptic LTD by AP5 (50mM) but not by LY341495 (100 mM). Calibration bar, 0.5 mV, 10 ms. In this and subsequent panels, the period of 1 Hz stimulation is shown bythe connected arrows and responses during this time are averages of 60 consecutive records. (b, c) Pooled data for five slices. In one input (b) 1Hz stimulation was applied in the presence of LY341495 (100 mM) and in the other input (c) 1 Hz stimulation was applied either before orfollowing washout of LY341495.
been cloned. An alternative mechanism has been sug-gested recently; that (S)-MCPG blocks the induction ofLTP via an agonist action at group II mGlu receptors(Breakwell et al., 1998). This conclusion was based onthe observation that a group II antagonist 2S,1%S,2%S-2-methyl-2-(2%-carboxycyclopropyl)-glycine (MCCG) pre-vented the inhibitory action of (S)-MCPG on theinduction of LTP. However, MCCG is a weak, partialagonist at group II mGlu receptors (Bushell et al.,1996). In contrast, LY341495 is a full antagonist withnanomolar potency at both mGlu2 and mGlu3 receptors(Kingston et al., 1998). We therefore tested this theorydirectly by determining the ability of (S)-MCPG toblock the induction of LTP in the presence ofLY341495. As illustrated in Fig. 9b, the ability of(S)-MCPG to block the induction of LTP was unaf-fected by the presence of LY341495 (n=4). Therefore,
agonist activity at group II mGlu receptors cannotexplain the ability of (S)-MCPG to block the inductionof LTP.
3.9. LY341495 blocks the setting of the molecularswitch
Appropriate synaptic activity converts a pathwayfrom an (S)-MCPG-sensitive state to an (S)-MCPG-in-sensitive state via activation of receptors which arethemselves (S)-MCPG-sensitive (Bortolotto et al.,1994). This led to the hypothesis that activation of(S)-MCPG-sensitive mGlu receptors sets a molecularswitch which negates the need for subsequent activationof mGlu receptors for the induction of further LTP. Wetherefore determined whether LY341495 could preventthe setting of the (S)-MCPG-sensitive molecular switch,

S.M. Fitzjohn et al. / Neuropharmacology 37 (1998) 1445–14581454
Fig. 7. LY341495 blocks depotentiation. (a,b) A single example (a) and pooled data from five slices (b) to show reversible blockade of NMDAreceptor-independent depotentiation (50 mM AP5) by LY341495 (100 mM). LTP (at arrow) was induced using a single tetanus (100 Hz, 1 s). Datawere normalised to the 20 min preceding the first period of 1 Hz stimulation.
by delivering a tetanus in its presence and then testingthe sensitivity of further LTP to (S)-MCPG. A tetanusdelivered immediately prior to application of LY341495set the molecular switch whilst a tetanus delivered to anindependent input in the presence of LY341495 did not,since further LTP was specifically blocked by (S)-MCPG in the latter input (Fig. 10). Thus, 60 min aftera tetanus in (S)-MCPG, following a prior tetanus inLY341495, responses were 10299% of control, com-pared to 140916% in the input where the first tetanuswas delivered immediately before application ofLY341495 (n=5; PB0.01).
3.10. LY341495 does not block NMDAreceptor-independent, mossy fibre LTP
Finally, we examined the ability of LY341495 toblock the induction of mossy fibre LTP (Fig. 11). Ineach experiment, LY341495 failed to prevent the induc-tion of AP5-resistant LTP (responses 60 min aftertetanus 16098% of control; n=7). In five of theseslices, the sensitivity to (S)-MCPG (200 mM) was firstlydetermined; in each case (S)-MCPG prevented the in-duction of mossy fibre LTP as described previously
(Bashir et al., 1993a). Single examples of mossy fibre-in-duced LTP, in the presence of 100 mM LY341495, areshown in Fig. 11a, b and pooled data for all fiveexperiments comparing (S)-MCPG and LY341495 ispresented in Fig. 11c.
4. Discussion
In the present study we have utilised a new com-pound, LY341495 (Kingston et al., 1998), which is themost potent mGlu receptor antagonist at all knownmGlu receptor subtypes which are expressed in thehippocampus (mGlu1-5,7,8), to investigate the roles ofmGlu receptors in various forms of hippocampalsynaptic plasticity. LY341495 (100 mM) did not affectbasal synaptic transmission or the induction of LTP,demonstrating no activity at AMPA or NMDA recep-tors, consistent with its lack of effect on AMPA,NMDA and kainate receptors in ligand binding experi-ments (Kingston et al., 1998). Its ability to antagonisemGlu5 receptors is of particular interest for studies ofsynaptic plasticity in area CA1, where mGlu5 receptorsare the predominant postsynaptic subtype (Lujan et al.,

S.M. Fitzjohn et al. / Neuropharmacology 37 (1998) 1445–1458 1455
1996), particularly since (S)-MCPG is a weak antago-nist at mGlu5 receptors when they are activated byL-glutamate (Brabet et al., 1995). Indeed, LY341495was a highly effective antagonist at blocking DHPG-in-duced potentiation of NMDA responses, an effectlikely to be mediated via activation of mGlu5 receptors(Doherty et al., 1997). Fortunately, for physiologicalstudies, LY341495 is an effective antagonist when usedagainst rat mGlu receptors stimulated by L-glutamate.However, the pharmacological analysis shows that, irre-spective of the agonist employed, it is necessary to usea high concentration of LY341495 to ensure completefunctional blockade of group I mGlu receptors by thiscompetitive antagonist. We therefore used LY341495 ata concentration of 100 mM for all the studies onsynaptic plasticity.
The different forms of synaptic plasticity studiedcould be divided into three categories based on their
Fig. 9. LY341495 neither mimics nor blocks the actions of (S)-MCPG. (a) Lack of effect of LY341495 (100 mM) on (S)-MCPG (200mM)-sensitive LTP (n=4). (b) LY341495 does not prevent (S)-MCPG from inhibiting the induction of LTP (n=4). Note that inboth (a) and (b), the first tetanus to input 2 (lower panels) wasdelivered immediately prior to application of LY341495.
Fig. 8. LY341495 does not affect LTP in area CA1. Pooled data fornine two-input experiments. In input 1, the tetanus was appliedimmediately prior to the application of 100 mM LY341495 whilst ininput 2 the tetanus was applied following 20-min incubation, immedi-ately prior to washout of LY341495. In the lower trace the two inputshave been aligned with respect to the tetanus and superimposed.
sensitivity to (S)-MCPG and LY341495. The first cate-gory is plasticity which is sensitive to both antagonists.Both depotentiation of pre-established LTP and settingof the molecular switch are sensitive to LY341495 and,as has been shown previously, to (S)-MCPG (Bor-tolotto et al., 1994; Bashir and Collingridge, 1994).These results are consistent with a role for one or moreof the cloned subtypes of mGlu receptor which aresensitive to both antagonists (such as mGlu1, mGlu2 ormGlu5). The finding that LY341495 prevented the set-ting of the molecular switch but not the induction ofLTP differentiates it from (S)-MCPG which blocksboth aspects of synaptic plasticity, and suggests thatswitch setting and the induction of LTP per se in naıveinputs involve two different subtypes of (S)-MCPG-sensitive mGlu receptors.

S.M. Fitzjohn et al. / Neuropharmacology 37 (1998) 1445–14581456
The second category is plasticity which is affected byneither antagonist. Of the LY341495-insensitive formsof plasticity described here, NMDA receptor-dependenthomosynaptic LTD has been shown to be independentof (S)-MCPG-sensitive mGlu receptors (Selig et al.,1995; Oliet et al., 1997), except in extremely younganimals or when the divalent cation concentration ismodified (Bolshakov and Siegelbaum, 1994; Oliet et al.,1997). This plasticity could involve one or more mGlureceptor subtypes which are insensitive to both antago-nists or, more simply, be independent of mGlu receptoractivation.
The final category is plasticity which is sensitive to(S)-MCPG but not LY341495 and applies to both
Fig. 11. LY341495 does not block mossy fibre (MF) LTP. (a) Singleexample to show lack of effect of LY341495 (100 mM) on AP5 (50mM)-resistant, mossy fibre LTP. (b) A single example showing acomparison of the effects of (S)-MCPG (200 mM) and LY341495 onmossy fibre LTP. (c) Pooled data for five experiments, performed asin (b) including the data for the associational (Ass) input.
Fig. 10. LY341495 (100 mM) blocks the setting of the molecularswitch. (a, b) A single example (a) and pooled data for five slices (b).The stimulus intensity was reduced in both inputs at the timesindicated by the inverted triangles. Note that in both (a) and (b), thefirst tetanus to input 2 (lower panels) was delivered immediately priorto application of LY341495. In both inputs the second tetanus wasdelivered in the presence of (S)-MCPG. In (a), a third tetanus wasapplied to input 1 to verify that the failure to induce LTP in responseto the second tetanus was due to the action of (S)-MCPG rather thanthe failure of the input to exhibit further LTP. A similar result wasobtained in all three slices so tested.
NMDA receptor-dependent and NMDA receptor-inde-pendent LTP. Since LY341495 was used at a concentra-tion over 10-fold the Ki-value for antagonism of bothmGlu1 and mGlu5 receptors it is unlikely that either ofthe two known PLC-coupled mGlu receptors are neces-sary for the induction of LTP in these pathways. Thissuggests that the deficits in LTP observed in areas CA1(Aiba et al., 1994) and CA3 (Conquet et al., 1994) inmGlu1, and in area CA1 in mGlu5 (Lu et al., 1997),receptor knockouts are unrelated to acute loss of mGlureceptor but rather may be a background effect or dueto compensation for the loss of receptor throughoutdevelopment. However, the finding that LTP in areaCA1 is normal in a different mGlu1 receptor knockout(Conquet et al., 1994) is consistent with the presentfindings.

S.M. Fitzjohn et al. / Neuropharmacology 37 (1998) 1445–1458 1457
The finding that homosynaptic LTP and LTD do notrequire activation of LY341495-sensitive mGlu recep-tors for their induction does not exclude the possibilitythat such receptors could, under certain circumstances,be involved in the modulation of the induction of eitheror both of these processes, for example via facilitationof NMDA receptor activation. It is possible thereforethat under appropriate conditions LY341495 couldhave some effect on homosynaptic LTP or LTD. Fur-ther experiments are required to address this issue.
Since the (S)-MCPG-sensitive receptor involved inthe induction of LTP in naıve inputs is insensitive toLY341495, all cloned mGlu receptors can be excludedas the mediators of this central plastic event. As (S)-MCPG-sensitive LTP can be induced by activation ofmGlu receptors using (1S,3R)-ACPD (Bashir et al.,1993a,b), but not DHPG (Bortolotto and Collingridge,1995), it is most likely to involve a novel mGlu receptorsubtype rather than some different class of receptor.One such candidate which stimulates PI turnover andhas the appropriate pharmacology has recently beendescribed in liver (Copani et al., 1997); it is activated by(1S,3R)-ACPD, but not by DHPG, and is antagonisedby (S)-MCPG. It will be interesting to see whether thisreceptor, or a related family member, is expressed in thehippocampus.
In conclusion, homosynaptic LTP and LTD are read-ily induced in the presence of LY341495, a potentantagonist at all cloned mGlu receptor subtypes thatare expressed in the hippocampus. Thus, although themGlu receptor subtypes that have been identified bycloning are involved in some aspects of plasticity, theyare not essential for the dominant forms of synapticplasticity in the hippocampus.
Acknowledgements
This project was supported by the MRC, BBSRCand The European Economic Community (BioMedBMH4-CT96-0228 and Biotech PL96-0049). We thankBryan G. Johnson and Craig R. Salhoff for technicalassistance.
References
Aiba, A., Chen, C., Herrup, K., Rosenmund, C., Stevens, C.F.,Tonegawa, S., 1994. Reduced hippocampal long-term potentia-tion and context-specific deficit in associative learning in mGluR1mutant mice. Cell 79, 365–375.
Anderson, W.W., Collingridge, G.L., 1997. A data acquisition pro-gram for on-line analysis of long-term potentiation and long-termdepression, Soc. Neurosci. Abstr. 264.19.
Bashir, Z.I., Collingridge, G.L., 1994. An investigation of depotentia-tion of long-term potentiation in the CA1 region of the hippocam-pus. Exp. Brain Res. 100, 437–443.
Bashir, Z.I., Bortolotto, Z.A., Davies, C.H., Berretta, N., Irving,A.J., Seal, A.J., Henley, J.M., Jane, D.E., Watkins, J.C.,Collingridge, G.L., 1993a. Induction of LTP in the hippocampusneeds synaptic activation of glutamate metabotropic receptors.Nature 363, 347–350.
Bashir, Z.I., Jane, D.E., Sunter, D.C., Watkins, J.C., Collingridge,G.L., 1993b. Metabotropic glutamate receptors contribute to theinduction of long-term depression in the CA1 region of thehippocampus. Eur. J. Pharmacol. 239, 265–266.
Bear, M.F., Abraham, W.C., 1996. Long-term depression in thehippocampus. Annu. Rev. Neurosci. 19, 437–462.
Bliss, T.V.P., Collingridge, G.L., 1993. A synaptic model of memory:long-term potentiation in the hippocampus. Nature 361, 31–39.
Bolshakov, V.Y., Siegelbaum, S.A., 1994. Postsynaptic induction andpresynaptic expression of hippocampal long-term depression. Sci-ence 264, 1148–1152.
Bortolotto, Z.A., Bashir, Z.I., Davies, C.H., Collingridge, G.L., 1994.A molecular switch activated by metabotropic glutamate recep-tors regulates induction of long-term potentiation. Nature 368,740–743.
Bortolotto, Z.A., Collingridge, G.L., 1995. On the mechanism oflong-term potentiation induced by (1S,3R)-1-aminocyclopentane-1,3-dicarboxylic acid (ACPD) in rat hippocampal slices. Neu-ropharmacology 34, 1003–1014.
Bortolotto, Z.A., Ornstein, P.L., Schoepp, D.D., Kingston, A.E.,Lodge, D., Collingridge, G.L., 1998. Actions of LY341495, apotent mGlu receptor antagonist, on hippocampal LTP. Eur. J.Neurosci. 10, 224.
Brabet, I., Mary, S., Bockaert, J., Pin, J.-P., 1995. Phenylglycinederivatives discriminate between mGluR1- and mGluR5-mediatedresponses. Neuropharmacology 34, 895–903.
Breakwell, N.A., Rowan, M.J., Anwyl, R., 1998. MCPG blocksinduction of LTP in CA1 of rat hippocampus via agonist actionat an mGluR group II receptor. J. Neurophysiol. 79, 1270–1276.
Bushell, T.J., Jane, D.E., Tse, H.-W., Watkins, J.C., Garthwaite, J.,Collingridge, G.L., 1996. Pharmacological antagonism of theactions of group II and III mGluR agonists in the lateral per-forant path of rat hippocampal slices. Br. J. Pharmacol. 117,1457–1462.
Chinestra, P., Aniksztejn, L., Diabira, D., Ben-Ari, Y., 1993. (RS)-al-pha-methyl-4-carboxyphenylglycine neither prevents induction ofLTP nor antagonizes metabotropic glutamate receptors in CA1hippocampal neurons. J. Neurophysiol. 70, 2684–2689.
Chung, D.S., Traynelis, S.F., Murphy, T.J., Conn, P.J., 1997. 4–Methylhomoibotenic acid activates a novel metabotropic gluta-mate receptor coupled to phosphoinositide hydrolysis. J.Pharmacol. Exp. Ther. 283, 742–749.
Conn, P.J., Pin, J.-P., 1997. Pharmacology and fuctions of metabo-tropic glutamate receptors. Annu. Rev. Pharmacol. Toxicol. 37,205–237.
Conquet, F., Bashir, Z.I., Davies, C.H., Daniel, H., Ferraguti, F.,Bordi, F., Franz-Bacon, K., Reggiani, A., Matarese, V., Conde,F., Collingridge, G.L., Crepel, F., 1994. Motor deficit and impair-ment of synaptic plasticity in mice lacking mGluR1. Nature 372,237–243.
Copani, S.F., Bruno, V., Knopfel, T., Meltzger, G., Nicoletti, F.,1997. Metabotropic glutamate receptor agonists stimulatepolyphosphoinositide hydrolysis in primary cultures of rat hepa-tocytes. Eur. J. Pharmacol. 338, R1–R2.
Doherty, A.J., Palmer, M.J., Henley, J.M., Collingridge, G.L., Jane,D.E., 1997. (RS)-2-Chloro-5-hydroxyphenylglycine (CHPG) acti-vates mGlu5, but not mGlu1, receptors expressed in CHO cellsand potentiates NMDA responses in the hippocampus. Neu-ropharmacology 36, 265–267.
Dudek, S.M., Bear, M.F., 1992. Homosynaptic long-term depressionin area CA1 of hippocampus and effects of N-methyl-D-aspartatereceptor blockade. Proc. Natl. Acad. Sci. USA 89, 4363–4367.

S.M. Fitzjohn et al. / Neuropharmacology 37 (1998) 1445–14581458
Eaton, S.A., Jane, D.E., Jones, P.L.S.J., Porter, R.H.P., Pook, P.C.-K., Sunter, D.C., Udvarhelyi, P.M., Roberts, P.J., Salt, T.E.,Watkins, J.C., 1993. Competitive antagonism at metabotropicglutamate receptors by (S)-4-carboxyphenylglycine and (RS)-a-methyl-4-carboxyphenylglycine. Eur. J. Pharmacol. Mol. Pharma-col. 244, 195–197.
Fitzjohn, S.M., Irving, A.J., Palmer, M.J., Harvey, J., Lodge, D.,Collingridge, G.L., 1996. Activation of group I mGluRs potenti-ates NMDA responses in rat hippocampal slices. Neurosci. Lett.203, 211–213.
Fitzjohn, S.M., Lodge, D., Collingridge, G.L., 1996a. Long-termdepression of synaptic transmission induced by group I metabo-tropic glutamate receptor activation in the CA1 region of the rathippocampus in vitro. J. Physiol. 493, 49P.
Fitzjohn, S.M., Ornstein, P.L., Schoepp, D.D., Kingston, A.E.,Lodge, D., Collingridge, G.L., 1998. Actions of the potent mGlureceptor antagonist LY341495 on hippocampal LTD and depo-tentiation. Eur. J. Neurosci. 10, 128.
Huber, K.M., Sawtell, N.B., Bear, M.F., 1997. Effects of novelmetabotropic glutamate receptor antagonists on glutamate stimu-lated phosphoinositide turnover and synaptic transmission in ratvisual cortex, Soc. Neurosci. Abstr. 41.1.
Kingston, A.E., Burnett, J.P., Mayne, N.G., Lodge, D., 1995. Phar-macological analysis of 4-carboxyphenylglycine derivatives: com-parison of effects on mGluR1a and mGluR5a subtypes.Neuropharmacology 34, 887–894.
Kingston, A.E., Ornstein, P.L., Wright, R.A., Johnson, B.G., Mayne,N.G., Burnett, J.P., Belagaje, R., Wu, S., Schoepp, D.D., 1998.LY341495 is a nanomolar potent and selective antagonist atgroup II metabotropic glutamate receptors. Neuropharmacology37, 1–12.
Little, Z., Grover, L. M., Teyler, T.J., 1995. Metabotropic glutamatereceptor antagonist, (R,S)-a-methyl-4-carboxyphenylglycine,blocks two distinct forms of long-term potentiation in the CA1 ofrat hippocampus. Neurosci. Lett. 201, 73–76.
Lu, Y.M., Jia, Z.P., Janus, C., Henderson, J.T., Gerlai, R., Wojtow-icz, J.M., Roder, J.C., 1997. Mice lacking metabotropic glutamatereceptor 5 show impaired learning and reduced CA1 long-termpotentiation (LTP) but normal CA3 LTP. J. Neurosci. 17, 5196–5205.
Lujan, R., Nusser, Z., Roberts, J.D.B., Shigemoto, R., Somogyi, P.,1996. Perisynaptic location of metabotropic glutamate receptorsmGluR1 and mGluR5 on dendrites and dendritic spines in the rathippocampus. 8, 1488–1500.
Manahan-Vaughan, D., 1997. Group 1 and 2 metabotropic glutamatereceptors play differential roles in hippocampal long-term depres-sion and long-term potentiation in freely moving rats. J. Neurosci.17, 3303–3311.
Manzoni, O.J., Weisskopf, M.G., Nicoll, R.A., 1994. MCPG antago-
nises metabotropic glutamate receptors but not long-term potenti-ation in the hippocampus. Eur. J. Neurosci. 6, 1050–1054.
Martin, S.J., Morris, R.G.M., 1997. (R,S)-a-methyl-4-car-boxyphenylglycine (MCPG) fails to block long-term potentiationunder urethane anaesthesia in vivo. Neuropharmacology 36,1339–1354.
Mulkey, R.M., Malenka, R.C., 1992. Mechanisms underlying induc-tion of homosynaptic long-term depression in area CA1 of thehippocampus. Neuron 9, 967–975.
Nakanishi, S., Masu, M., 1994. Molecular diversity and functions ofglutamate receptors. Nature 368, 740–743.
O’Connor, J.J., Rowan, M.J., Anwyl, R., 1994. Long-lasting en-hancement of NMDA receptor-mediated synaptic transmission bymetabotropic glutamate receptor activation. Nature 367, 557–559.
Oliet, S.H.R., Malenka, R.C., Nicoll, R.A., 1997. Two distinct formof long-term depression coexist in CA1 hippocampal pyramidalcells. Neuron 18, 969–982.
Palmer, M.J., Irving, A.J., Seabrook, G.R., Jane, D.E., Collingridge,G.L., 1997. The group I mGlu receptor agonist DHPG induces anovel form of LTD in the CA1 region of the hippocampus.Neuropharmacology 36, 1517–1532.
Richter-Levin, G., Errington, M.L., Maegawa, H., Bliss, T.V.P.,1994. Activation of metabotropic glutamate receptors is necessaryfor long-term potentiation in the dentate gyrus and for spatiallearning. Neuropharmacology 33, 853–857.
Selig, D.K., Lee, H.K., Bear, M.F., Malenka, R.C., 1995. Re-exami-nation of the effects of MCPG on hippocampal LTP, LTD, anddepotentiation. J. Neurophysiol. 74, 1075–1082.
Sergueeva, O.A., Fedorov, N.B., Reymann, K.G., 1993. An antago-nist of glutamate metabotropic receptors, (RS)-a-methyl-4-car-boxyphenylglycine, prevents the LTP-related increase inpostsynaptic AMPA sensitivity in hippocampal slices. Neurophar-macology 32, 933–935.
Thomas, M.J., O’Dell, T. J., 1995. The molecular switch hypothesisfails to explain the inconsistent effects of the metabotropic gluta-mate receptor antagonist MCPG on long-term potentiation. BrainRes. 695, 45–52.
Wang, Y., Rowan, M.J., Anwyl, R., 1995. (RS)-a-methyl-4-car-boxyphenylglycine inhibits long-term potentiation only followingthe application of low frequency stimulation in the rat dentategyrus in vitro. Neurosci. Lett. 197, 207–210.
Watkins, J.C., Collingridge, G.L., 1994. Phenylglycine derivatives asantagonists of metabotropic glutamate receptors. Trends Pharma-col. Sci. 15, 333–342.
Yang, X.-D., Connor, J.A., Faber, D.S., 1994. Weak excitation andsimultaneous inhibition induce long-term depression in hippocam-pal CA1 neurons. J. Neurophysiol. 71, 1586–1590.
.