snap25 in hippocampal ca1 region is involved in memory consolidation
TRANSCRIPT

SNAP-25 in hippocampal CA1 region is involvedin memory consolidation
Qiuling Hou,1,* Xiang Gao,2,* Xuehan Zhang,1 Lingwei Kong,2 Xinming Wang,1 Wei Bian,2 Yanyang Tu,2 Meilei Jin,3
Guoping Zhao,4 Baoming Li,1 Naihe Jing2 and Lei Yu51Laboratory of Higher Brain Functions, Institute of Neurobiology, Fudan University, Shanghai, China2Laboratory of Molecular Cell Biology, Laboratory of Stem Cell Biology, Institute of Biochemistry and Cell Biology and3Research Center of Biotechnology, Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences, Shanghai, China4Chinese National Human Genome Center at Shanghai, Pudong, Shanghai, China5Department of Cell Biology, Neurobiology and Anatomy, University of Cincinnati College of Medicine, 3125 Eden Avenue,Cincinnati, OH 45267-0521, USA
Keywords: antisense oligonucleotides, hippocampus, long-term potentiation, memory, rat, SNAP-25
Abstract
As a synaptosomal protein, SNAP-25 plays a role in a number of neuronal functions including axonal growth, dendrite formation,fusion of synaptic vesicles with membrane and the expression of long-term potentiation (LTP) in the hippocampus. Using alearning ⁄memory behavior screening, we identified SNAP-25 as one of the differentially expressed genes in the hippocampus uponbehavioral training. The inhibition of SNAP-25 with intracerebroventricular antisense oligonucleotide caused a deficit in long- but notshort-term memory for step-down inhibitory avoidance. Intra-CA1 infusion of the SNAP-25 antisense oligonucleotide impaired long-term contextual fear memory and spatial memory and interfered with the LTP of synaptic transmission in the CA1 region. Theinhibitory effect on LTP was not mediated by a pre-synaptic mechanism because paired pulse facilitation of synaptic transmissionwas not affected after administration of the antisense oligonucleotide. Together, the results suggest that SNAP-25 in the CA1 regionis involved in memory consolidation.
Introduction
Learning and memory are important biological functions for survival.Impairment of the learning ⁄memory capacity is often associated withneurological diseases and major insults to the brain, resulting in thedeterioration of cognitive functions and quality of life. Learn-ing ⁄memory is a complex set of processes involving the acquisition,consolidation and retrieval of information (Squire & Kandel, 1999).Extensive studies have demonstrated that the hippocampus is a criticalpart of the central nervous system for learning ⁄memory (Squire &Kandel, 1999) and its damage selectively impairs the ability to learnand remember (Morris et al., 1982; Sutherland et al., 1982; Seldenet al., 1991; Kim et al., 1993; Phillips & LeDoux, 1995; Logue et al.,1997; Sandin et al., 1997; Squire & Kandel, 1999). Because of theimportance of learning ⁄memory in human life, animal models havebeen developed to explore various aspects of learning ⁄memorybehavior and to unravel the underlying mechanisms at the molecular,cellular, system and organism levels (Bliss & Collingridge, 1993;Eichenbaum & Otto, 1993; Silva et al., 1998; Stevens, 1998; Elgersma& Silva, 1999; Squire & Kandel, 1999; Martin et al., 2000).
Using such animal models, a number of genes have been identifiedto be involved in learning ⁄memory, including genes for both structuralproteins and functionally important proteins such as protein kinases,
phosphatases and other cell signaling proteins (Elgersma & Silva,1999; Sanes & Lichtman, 1999; Squire & Kandel, 1999). Some ofthese genes have been shown to play a critical role using a variety ofexperimental manipulations, including the complete deletion of genefunction by homologous recombination-based gene targeting (Huanget al., 1995; Abel et al., 1997; Silva et al., 1998; Elgersma & Silva,1999) and partial inhibition of protein function in the relevant brainregions by antisense oligonucleotide delivery (Guzowski & McGaugh,1997; Ikegami & Inokuchi, 2000).SNAP-25 is a synaptosomal protein, initially identified as a brain-
enriched molecule in a brain ⁄ liver differential screening (Branks &Wilson, 1986), which is highly expressed by neurons in thehippocampus and other brain regions (Oyler et al., 1989, 1992;Geddes et al., 1990). As an integral component of the synaptic vesicle-docking ⁄ fusion core complex, SNAP-25 is required in a late post-docking step of Ca2+-dependent neurotransmitter exocytosis usingin vitro preparations (Banerjee et al., 1996; Mehta et al., 1996). WhenSNAP-25 function was disrupted by antibody microinjection, neuro-transmitter release was inhibited without affecting the synaptic vesiclenumber at the plasma membrane, supporting a critical role of SNAP-25 in membrane fusion but not in synaptic vesicle targeting to the pre-synaptic membrane (Low et al., 1999).More importantly, two lines of evidence suggest that hippocampal
SNAP-25 may be involved in learning and memory. Firstly, theselective inhibition of SNAP-25 expression by SNAP-25 antisenseoligonucleotides prevents neurite elongation (Osen-Sand et al., 1993).The axonal elongation and transformation of growth cones to synaptic
Correspondence: Dr Lei Yu, as above.E-mail: [email protected]
*Q.H. and X.G. contributed equally to this work.
Received 7 March 2004, revised 31 May 2004, accepted 1 July 2004
European Journal of Neuroscience, Vol. 20, pp. 1593–1603, 2004 ª Federation of European Neuroscience Societies
doi:10.1111/j.1460-9568.2004.03600.x

terminals contribute to the remodeling of nerve terminals, which mayserve as a morphological substrate for memory and learning. Secondly,elevated mRNA levels of SNAP-25 are associated with the expressionof long-term potentiation (LTP) in granule cells of the dentate gyrus(Roberts et al., 1998), correlating SNAP-25 mRNA up-regulation withthe expression of LTP in the hippocampus. LTP in the hippocampus isconsidered to be a form of synaptic plasticity that underlieslearning ⁄memory (Morris et al., 1986; Morris, 1989; Bliss &Collingridge, 1993; Martin et al., 2000). We report here that SNAP-25 is indeed involved in learning and memory, particularly in memoryconsolidation.
Materials and methods
RNA preparation and cDNA library differential screening
Adult male Sprague-Dawley rats (200–250 g, purchased fromShanghai Laboratory Animal Center, Shanghai Institutes for Biolo-gical Sciences, Chinese Academy of Sciences, Shanghai, China)were used in this work. Animals were randomly assigned into twogroups (n ¼ 5 or 6 for each group). The training group receivedstep-down inhibitory avoidance training and the control groupreceived sham training (with no electric shock). The rats wereanesthetized and decapitated2 6 h after training and hippocampaltissues were collected for RNA extraction using Oligotex reagent(Qiagen, Valencia, CA, USA). Poly(A)+ RNA (2 lg) from eachgroup was reverse transcribed with Superscript II reverse transcrip-tase (Gibco ⁄BRL, Rockville, MD, USA) and cDNAs from thecontrol and training groups were labeled with a-32P-dATP by randompriming. These radioactive probes were used to hybridize duplicatefilters of a rat brain cDNA library (8-week-old Sprague-Dawley malerat; Gibco ⁄BRL) and differentially expressed clones were isolatedand subjected to DNA sequence analysis.
Oligonucleotides
The following unmodified oligonucleotides were used:SNAP-25 antisense oligonucleotide(5¢-ATGTCTGCGTCCTCGGCCAT-3¢)and mis-sense oligonucleotide(5¢-ATCTCAGCGTGCTTCGCCTT-3¢)and scramble oligonucleotide(5¢-TAGCTTCGGCTCGCTCGCTA-3¢).The antisense oligonucleotide has been shown to be effective inreducing SNAP-25 gene expression (Osen-Sand et al., 1993). Thescramble oligonucleotide consisted of the same base composition asthe antisense oligonucleotide.
Northern blot analysis
Adult male Sprague-Dawley rats3 (200–250 g) were anesthetized anddecapitated 6 h after receiving step-down inhibitory avoidance trainingand hippocampal tissues were collected for RNA extraction. TotalRNA (40 lg) from rat hippocampus was subjected to electrophoresis ina 1.0% agarose gel and transferred onto an N+ nylon membrane. Thehybridization probes were 32P-labeled by random priming using a0.5-kb fragment of a rat SNAP-25 cDNA clone (0.1 kb 5¢ untranslatedregion and 0.4 kb protein-coding region) and a 1.8-kb cDNA fragmentof rat GAPDH. The RNA blots were hybridized at 65 �C in ahybridization solution containing 0.2 m Na3PO4 (pH 7.2), 1 mm
EDTA (pH 8.0), 1% bovine serum albumin, 7% sodium dodecyl
sulfate, 15% formamide and 100 lg ⁄mL denatured salmon spermDNA. After overnight hybridization, blots were washed twice in40 mm Na3PO4 (pH 7.2), 1 mm EDTA (pH 8.0) and 1% sodiumdodecyl sulfate for 1 h at 65 �C and exposed to X-ray films overnightwith an enhancer screen at )80 �C. The hybridized signal wasvisualized using SNAPscan 1212 (AGFA, Ridgefield Park, NJ, USA).
In situ hybridization
Rats were anesthetized and perfused 6 h after receiving step-downinhibitory avoidance training4 and the brains were fixed in phosphate-buffered saline (PBS) containing 4% paraformaldehyde for 4 h atroom temperature5 . The fixed tissues were washed in PBS and thenembedded in Paraplast�. Tissue blocks were sectioned at a thicknessof 10 lm. A plasmid containing the 0.5-kb rat SNAP-25 cDNA waslinearized with either BamHI or EcoRI and transcribed in vitro with T7or SP6 RNA polymerase using digoxigenin-labeled UTP to generatesense and antisense riboprobes. In situ hybridization was performed aspreviously described (Yang et al., 2000).
Western blot analysis
Adult male Sprague-Dawley rats (200–250 g) were anesthetized anddecapitated6 either 6 h after step-down inhibitory avoidance training orat different time points (0, 1, 2.5, 6 or 24 h) after contextual fearconditioning and hippocampal tissues were collected for proteinisolation. The hippocampi (about 100 mg) of each rat were homo-genized ultrasonically in 1 mL of extraction buffer (150 mm NaCl,0.1% sodium dodecyl sulfate, 10 mm EDTA, 1% deoxycholic sodium,1% Triton X-100, 20 mm NaF, 0.25 mm phenylmethylsulfonylfluoride, 5 lg ⁄mL leupeptin and 50 mm Tris-HCl, pH 8.2). Thesupernatant fluids were collected after centrifugation at 15 000 r.p.m.for 10 min at 4 �C. After quantification, 100-lL aliquots wereprecipitated in 1 mL 70% ethanol with 0.13% KOAc, air-dried andresolved in 100 lL loading buffer. Samples (20 lg protein per lane)were subjected to preparative sodium dodecyl sulfate–polyacrylamidegel electrophoresis in a 10% gel and electrophoretically transferred tonitrocellulose membrane (Amersham Pharmacia, Piscataway, NJ,USA) using a Trans-Blot device (Bio-Rad, Hercules, CA, USA) at30 V constant voltages overnight at 4 �C. Membranes were soaked in1% bovine serum albumin and 5% normal goat serum in PBS for 1 hat room temperature to block non-specific binding, rinsed in PBS andincubated with an anti-SNAP-25 antibody (Affiniti, UK; diluted1 : 2000 in 1% bovine serum albumin, 5% normal goat serum in PBS)overnight at 4 �C. Membranes were washed twice for 15 min each inPBS with 0.01% Tween-20 and incubated for 1 h with a secondaryantibody, goat anti-rabbit IgG (1 : 10 000; Zymed, South SanFrancisco, CA, USA). After washing twice for 15 min each in PBSwith 0.01% Tween-20, membranes were incubated with an anti-rabbit-peroxidase ⁄ anti-peroxidase (PAP)7 antibody (1 : 20 000; Zymed) for1 h, washed once in PBS and the signal was detected by the ECLsystem (Amersham Pharmacia). A western blot of tubulin wasperformed in the same way, using a monoclonal anti-tubulin antibody(1 : 500; Sigma, St Louis, MO, USA) as the first antibody and a goatanti-mouse-horseradish peroxidase antibody (1 : 2000; Zymed) as thesecond antibody. Blots were quantified using gel work softwareversion 3.01 (UVP, Upland, CA, USA)8 . To quantify the results ofin situ hybridization, the average density of the SNAP-25 mRNAsignal in various hippocampal regions was calculated by themetamorph program (Universal Imaging Corporation, Downing-town, PA, USA).
1594 Q. Hou et al.
ª 2004 Federation of European Neuroscience Societies, European Journal of Neuroscience, 20, 1593–1603

Hippocampal neuron cell culture
Hippocampal cells were prepared from post-natal day 2 Sprague-Dawley rats as described previously (Brewer, 1997, 1999). The cellswere plated at 200 cells ⁄mm2 into poly-l-lysine-coated six-well plateswith neurobasal ⁄B27 supplemented with 10 ng ⁄mL basic fibroblastgrowth factor (bFGF)9 . The medium was replenished every 4 days andcells were cultured for 4 weeks for maturation. SNAP-25 antisenseand mis-sense oligonucleotides were used at 5 lg ⁄mL in the culturemedium. After 24 h the neuronal cells received fresh mediumcontaining the same concentration of oligonucleotides.
Intracerebroventricular and intrahippocampal administrationof oligonucleotides
Adult male Sprague-Dawley rats (200–250 g) were housed for a weekbefore surgical operation. For intracerebroventricular administration,rats were anesthetized with pentobarbital (40 mg ⁄ kg, i.p.) andmounted on a stereotaxic unit (SN-2; Narishige, Japan). An infusioncannula was stereotaxically implanted into the third ventricle (Paxinos& Watson, 1986). An Alzet 7-day osmotic mini-pump (ALZAPharmaceuticals, Palo Alto, CA, USA), which was filled with thedesired oligonucleotide (50 lg ⁄ lL) or saline solution, was embeddedunder the animal’s skin on the back and was connected to the infusioncannula with polyethylene (PE)10 tubing. The infusion rate of the mini-pump was 1 lL ⁄ h.
For intra-CA1 administration, the rat was cannulated, underpentobarbital anesthesia (40 mg ⁄ kg, i.p.), bilaterally in the CA1region (from Bregma 3.3 mm caudally, 1.8 mm laterally and fromskull surface 1.5 mm ventrally). After the animal recovered fromanesthesia, it was returned to the home cage for 1 week before anyexperimental manipulation. Saline or oligonucleotide solution wasinfused into the CA1 region via a Hamilton syringe inserted into theimplanted guide cannula. The tip of the syringe was placed 2.8–3.0 mm ventral to the skull surface. For each side of the hippocampus,a volume of 1.5 lL was administered in 3 min (0.5 lL ⁄min) followedby a 2-min waiting time before the injection cannula was taken out.Oligonucleotides were used at 1 nmol ⁄ lL.
Step-down inhibitory avoidance training
Adult male Sprague-Dawley rats (200–250 g) were trained for step-down inhibitory avoidance as described previously (Heise, 1984;Izquierdo et al., 1995, 1998) with some modifications. The trainingapparatus consisted of a wooden box [55 (L) · 40 (W) · 40 (H) cm]with an elevated platform [15 (L) · 15 (W) · 7.5 (H) cm] in onecorner of the box. During the training, a rat was first placed in theapparatus box for free exploration for 2 min and returned to the homecage. It was then placed on the elevated platform with its head towardthe corner. When the animal stepped down from the platform (the timeinterval from its placement on the platform to stepping down wasrecorded), it was allowed to explore the box for 30 s before beingreturned to the home cage. This procedure was repeated three times foreach animal with a 1-min interval between sessions. Next, the powersupply (45 V, AC) was switched on to electrify the metal grid on theapparatus floor and the animal was placed on the elevated platform asbefore. When the animal stepped down from the platform it received acontinuous electric foot shock. The rat would attempt to escape theshock and stepped up onto the elevated platform. If the animal stayedon the platform for 10 min, the training was considered completed andthe animal was returned to the home cage. If the animal stepped downagain within the 10-min period it would be shocked again. In our
experiments, most animals completed step-down inhibitory avoidancetraining after receiving one shock. Occasionally, an animal would stepdown from the platform after the first shock. Such animals were notincluded in subsequent studies. Control animals experienced the sameprocedure except that the apparatus floor was not electrified. To testthe memory for step-down inhibitory avoidance, an animal was placedon the platform for 5 min without electrifying the floor and itson-platform staying time was recorded.
Contextual and auditory fear conditioning
Adult male Sprague-Dawley rats (200–250 g) were trained in afreezing monitor system (San Diego Instruments, San Diego, CA,USA) consisting of a Plexiglas box [36 (L) · 23 (W) · 18 (H) cm]and a floor lined with stainless steel rods that can be electrified.Animals received training as described previously (Tang et al., 2001).Rats were placed into the apparatus box for 2.5 min and a tone signal(2200 Hz, 96 dB) was then given for 30 s [conditioned stimulus (CS)]with an electric shock (unconditioned stimulus; 1 mA) during the last1 s of the CS. After an interval of 30 s, a second CS–unconditionedstimulus paring was delivered. The animal was allowed to stay in thebox for an additional 30 s and its freezing behavior during this timeperiod was recorded as ‘immediate’.Contextual fear memory was tested 1 or 24 h post-training. Each
animal was placed in the box where it was trained for 3 min withoutthe presence of a CS or unconditioned stimulus and the duration of itsfreezing behavior was recorded. Auditory fear memory was testedimmediately after the contextual fear memory test; each animal wasplaced in a novel box for 3 min and was then exposed to the CS. Thecumulative duration of the animal’s freezing behavior was recorded asa measure of memory retention.
Morris water maze training
Adult male Sprague-Dawley rats (200–250 g) were trained in a Morriswater maze apparatus (San Diego Instruments). The water in the mazewas opaque so that the platform, once submerged, was not visible. Theposition of the platform was fixed at the south-east quadrant. For alltrials, video recordings were made for subsequent analysis.Six hours after intra-CA1 solution infusion (saline or oligonucle-
otide), the animals received training as described previously (Guzow-ski & McGaugh, 1997). Two training sessions were carried out, sixtrials per session, with a 1-h interval between the two sessions. Eachtrial started when a rat was placed at the junction between two adjacentquadrants (the east, north, west or south pole of the maze). The startingposition was randomly selected but counter-balanced among the fourpositions. If the animal found the submerged platform within 60 s, itwas allowed to stay there for 30 s and was then returned to a holdingcage for 30 s before the next trial started. If the animal did not find thesubmerged platform within 60 s, it was guided to the platform,allowed to stay there for 30 s and then returned to a holding cage for30 s before the next trial started.Memory retention was tested 48 h after training using a procedure
described previously (Guzowski & McGaugh, 1997). A total of threetrials were performed. The submerged platform was placed at the sameposition (the south-east quadrant) as in training and the startingposition was randomly selected between the north and west poles. Theanimal was placed in the water in the starting position and allowed tonavigate in the water for up to 60 s. Once the animal found thesubmerged platform, it was returned immediately to a holding cage.The inter-trial interval was 60 s.
SNAP-25 in area CA1 and memory consolidation 1595
ª 2004 Federation of European Neuroscience Societies, European Journal of Neuroscience, 20, 1593–1603
Immediately after the retention trials were completed, a three-trialvisible platform test was conducted. The platform was raised to abovethe water surface and covered with white gauze to be highly visible. Theplatform position was randomized among the four quadrants and thestarting position was fixed at the north or west pole. For each trialthe animal was first placed on the platform for 30 s and then placed in thewater in the starting position. The animal was allowed to navigate in thewater for up to 60 s. Once the animal found the visible platform, it wasimmediately returned to a holding cage. The inter-trial interval was 60 s.
Recording of long-term potentiation and oligonucleotideadministration
Adult male Sprague-Dawley rats (200–250 g) were used. The animalswere anesthetized with urethane carbamate (1.25 g ⁄ kg, i.p.) andmounted on a stereotaxic apparatus (SN-2; Narishige) with the incisorbar 3.3 mm below the interaural line. Body temperature wasmaintained at 36 ± 1.0 �C. After parting the skin and exposing theskull, two small holes were drilled on the skull to allow electrodepenetration. One hole was located at 4.9 mm posterior to Bregma and3.8 mm lateral to the mid-sagittal suture line for insertion of astimulating electrode and the other at 3.4 mm posterior to Bregma and2.5 mm lateral to the mid-sagittal suture line for insertion of aninfusion-recording electrode, based on the stereotaxic coordinates ofPaxinos & Watson (1986).
A stimulating electrode (concentric electrode, 200 lm diameter)was inserted into the Schaffer collateral pathway at a 15� angle to thefrontal plane and an infusion-recording electrode, constructed bygluing a Teflon-coated stainless steel wire (120 lm diameter) to aguide cannula (400 lm inside and 650 lm outside diameter), wasinserted vertically into the ipsilateral CA1 region. A stainless steelscrew (1.0 mm diameter) was implanted on the contralateral skull,serving as a reference electrode (implantation site 5 mm posterior toBregma and 1.5 mm lateral to mid-sagittal suture line). A test pulse(50 ls in duration) was delivered, once every 1.0 min, to the Schaffer
Contro
l
Traini
ng
SNAP-25
GAPDH
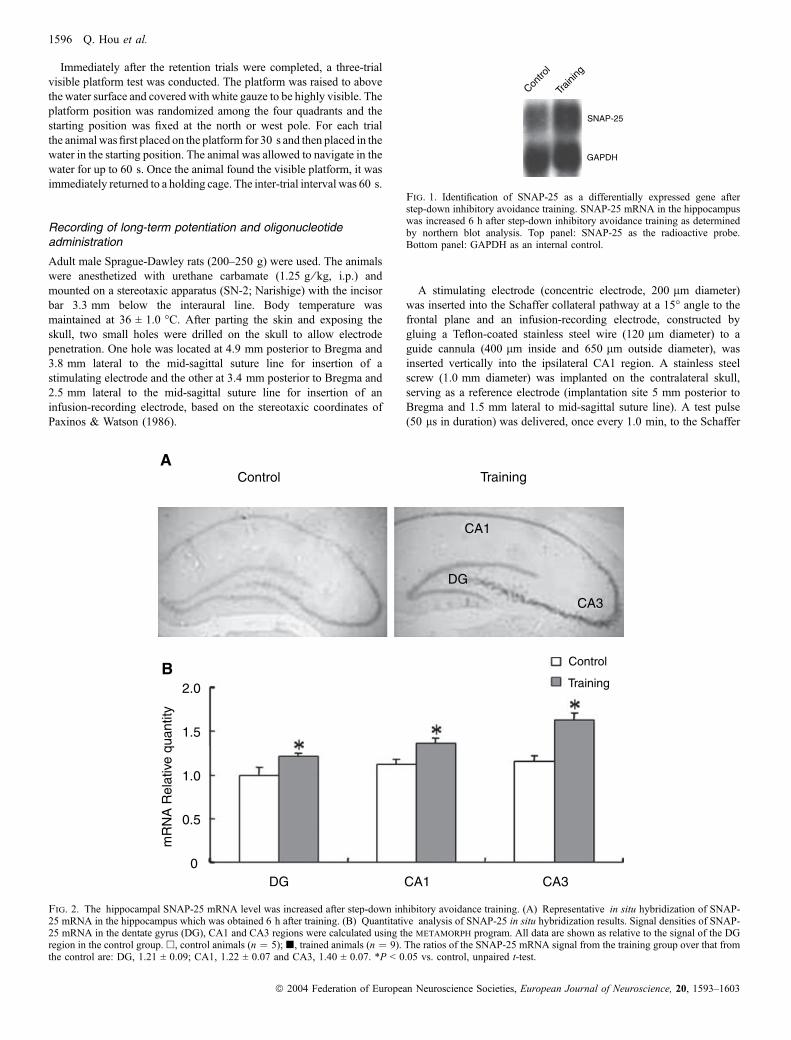
Fig. 1. Identification of SNAP-25 as a differentially expressed gene afterstep-down inhibitory avoidance training. SNAP-25 mRNA in the hippocampuswas increased 6 h after step-down inhibitory avoidance training as determinedby northern blot analysis. Top panel: SNAP-25 as the radioactive probe.Bottom panel: GAPDH as an internal control.
Control Training
Control
Training
DG
mR
NA
Rel
ativ
e qu
antit
y
CA1 CA3
2.0
1.5
0.5
1.0
0
CA1
DG
CA3
A
B
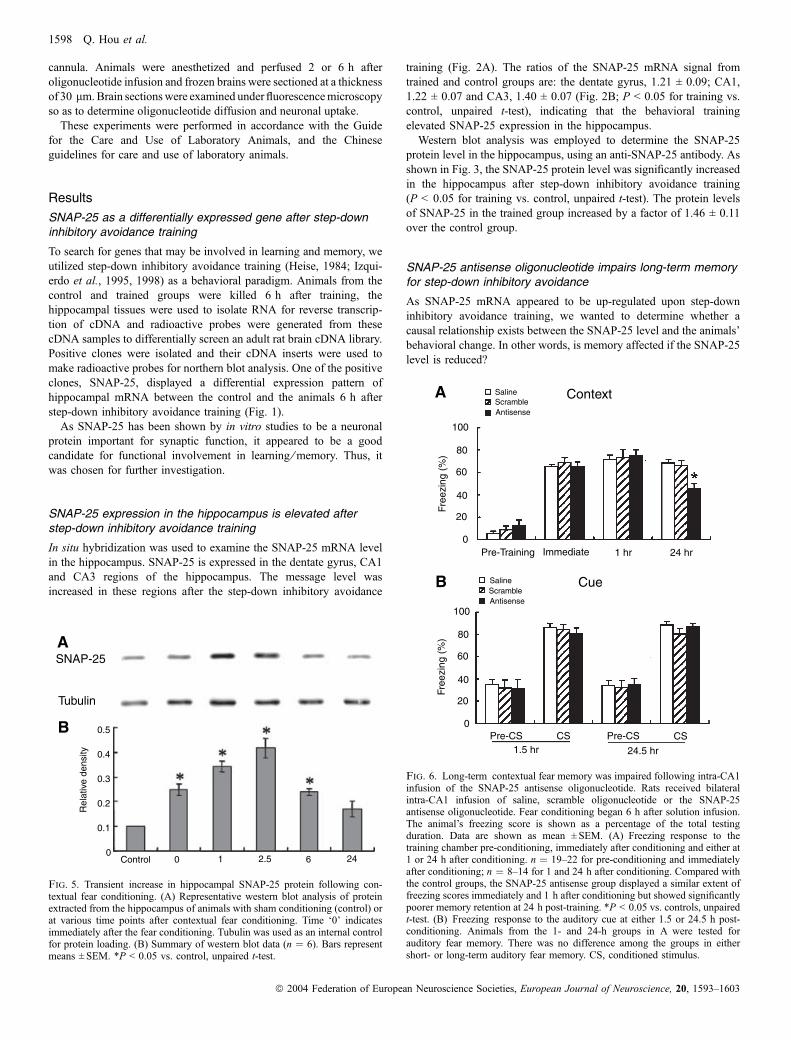
Fig. 2. The hippocampal SNAP-25 mRNA level was increased after step-down inhibitory avoidance training. (A) Representative in situ hybridization of SNAP-25 mRNA in the hippocampus which was obtained 6 h after training. (B) Quantitative analysis of SNAP-25 in situ hybridization results. Signal densities of SNAP-25 mRNA in the dentate gyrus (DG), CA1 and CA3 regions were calculated using the metamorph program. All data are shown as relative to the signal of the DGregion in the control group.h, control animals (n ¼ 5);j, trained animals (n ¼ 9). The ratios of the SNAP-25 mRNA signal from the training group over that fromthe control are: DG, 1.21 ± 0.09; CA1, 1.22 ± 0.07 and CA3, 1.40 ± 0.07. *P < 0.05 vs. control, unpaired t-test.
1596 Q. Hou et al.
ª 2004 Federation of European Neuroscience Societies, European Journal of Neuroscience, 20, 1593–1603

collateral pathway. The tip positions of the stimulating and infusion-recording electrodes were finely adjusted with two micromanipulatorsuntil an optimal field excitatory post-synaptic potential (fEPSP) wasobtained. A current intensity that elicited an fEPSP with two-thirds ofthe maximal response was used in the subsequent experiment.
The distance between the tip of the recording electrode and that ofthe guide cannula was 1.0 mm. The drug solution was deliveredthrough an injection needle (300 lm outside diameter) inserted intothe guide cannula. The injection needle extended 0.5–0.8 mm beyondthe tip of the guide cannula (0.5–0.2 mm from the tip of the recordingelectrode; approximately within or near to the layer of cell body in theCA1 region). Oligonucleotides (SNAP-25 antisense or scramble,0.165 nmol) or saline were infused at 1 lL in 4 min. The injectionneedle was left in place throughout the subsequent electrophysiolog-ical recording. During and after infusion, an fEPSP was evoked, onceevery 1 min, by delivering a single current pulse (50 ls in duration) tothe Schaffer collateral pathway.
The LTP recording was started 6 h after solution injection. Threetrains of tetanus stimulation were delivered to the Schaffer collateralpathway. Each train of tetanus consisted of 20 pulses of 50-ls durationeach with a 5-ms interpulse interval (i.e. at a frequency of 200 Hz) andthe intertetanus interval was 30 s.
The effect of SNAP-25 antisense on paired-pulse facilitation (PPF)was examined before and after LTP was induced. PPF was measuredusing various interpulse intervals (150, 180, 200 and 250 ms). As PPFis mediated by a pre-synaptic mechanism, a change in PPF following agiven treatment would indicate whether a pre-synaptic mechanismunderlies the action of that treatment.
Fluorescence microscopy
The SNAP-25 antisense oligonucleotide was fluorescently labeled at the5¢ end and was administered to the CA1 region of pentobarbitalanesthetized rat hippocampus (1 nmol ⁄ lL, 1.5 lL) via an implanted
A
B
C
Pre-training
1 hr post-training
24 hr post-training
Saline Scramble Antisense
On-
plat
form
tim
e (s
) 600
500
400
300
200
100
0
Contro
l
Miss
ense
Antise
nse
SNAP-25
Tubulin
Control
Control
Missense
TrainingMissense
Antisense
TrainingAntisense
Nor
mal
ized
inte
nsity
1.2
1.0
0.8
0.6
0.4
0.2
0
1.2
1.0
0.8
0.6
0.4
0.2
0
Rel
ativ
e de
nsity
Training
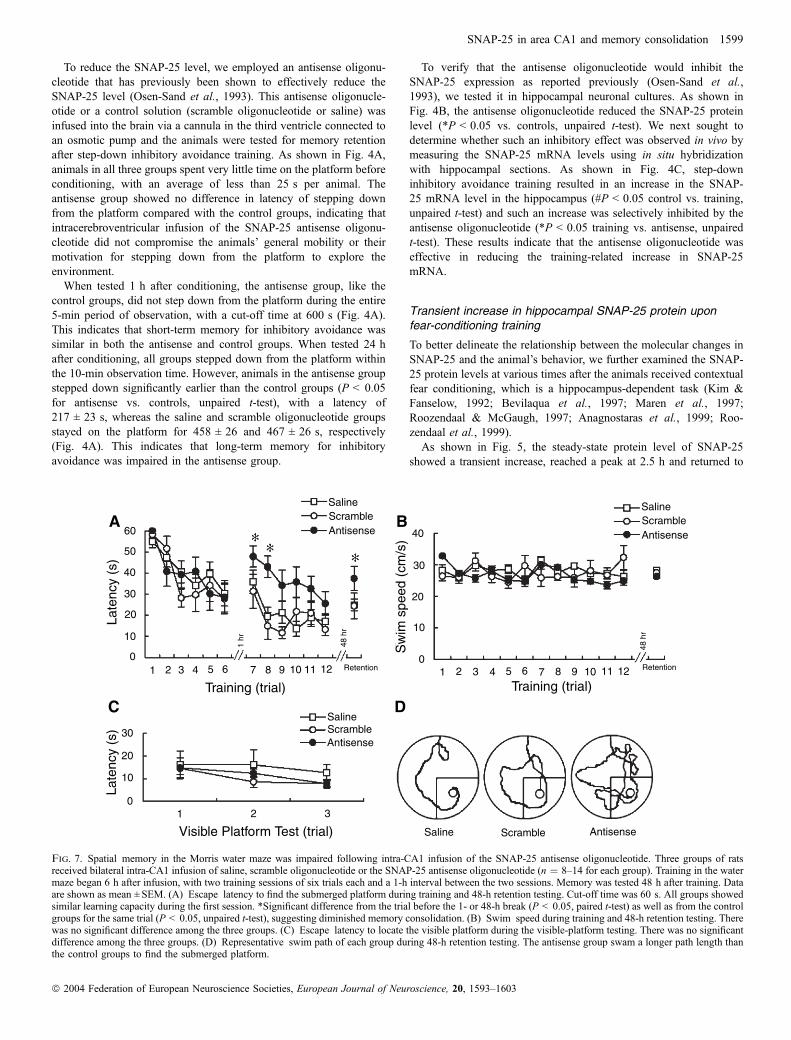
Fig. 4. The SNAP-25 antisense oligonucleotide impaired long-term memoryfor step-down inhibitory avoidance. (A) Animals were given an intracerebro-ventricular infusion of saline, scramble oligonucleotide or the SNAP-25antisense oligonucleotide into the third ventricle by 7-day osmotic pump. Fourdays after the infusion started, animals received step-down inhibitory avoidancetraining. Memory was tested either at 1 or 24 h after training. When tested 24 hafter conditioning, all groups stepped down from the platform within the 10-min observation time. However, animals in the antisense group stepped downsignificantly earlier than the control groups (*P < 0.05 for antisense vs.controls, unpaired t-test). Data are shown as mean ± SEM, n ¼ 10 for eachgroup. (B) The SNAP-25 antisense oligonucleotide was effective in reducingexpression of SNAP-25 protein in cultured hippocampal neurons. The SNAP-25 antisense oligonucleotide was as described previously (Osen-Sand et al.,1993). Proteins were extracted for western blot analysis. Data are shown asmean ± SEM, n ¼ 5 for each group. *P < 0.05 vs. controls, unpaired t-test.(C) The SNAP-25 antisense oligonucleotide was effective in reducing SNAP-25 mRNA level in the hippocampus. Hippocampal paraffin sections wereprepared for in situ hybridization analysis. Data are shown as mean ± SEM,n ¼ 3 for each group. #P < 0.05 control vs. training and *P < 0.05 training vs.antisense, unpaired t-test.
Control Training
SNAP-25
Tubulin
Control Training
Rel
ativ
e in
tens
ity
2.0
1.5
1.0
0.5
0
A
B
Fig. 3. The hippocampal SNAP-25 protein level was increased afterstep-down inhibitory avoidance training. (A) Western blot analysis of SNAP-25 protein in the hippocampus obtained 6 h after training, using an anti-SNAP-25 antibody. Tubulin was used as an internal control to normalize the SNAP-25protein level. (B) Relative protein level of SNAP-25 from the training group tothat of the control group is 1.46 ± 0.11 (mean ± SEM, n ¼ 5). *P < 0.05 vs.control, unpaired t-test.
SNAP-25 in area CA1 and memory consolidation 1597
ª 2004 Federation of European Neuroscience Societies, European Journal of Neuroscience, 20, 1593–1603
cannula.11 Animals were anesthetized and perfused 2 or 6 h afteroligonucleotide infusion and frozen brains were sectioned at a thicknessof 30 lm.Brain sectionswere examined under fluorescencemicroscopyso as to determine oligonucleotide diffusion and neuronal uptake.These experiments were performed in accordance with the Guide
for the Care and Use of Laboratory Animals, and the Chineseguidelines for care and use of laboratory animals.12
Results
SNAP-25 as a differentially expressed gene after step-downinhibitory avoidance training
To search for genes that may be involved in learning and memory, weutilized step-down inhibitory avoidance training (Heise, 1984; Izqui-erdo et al., 1995, 1998) as a behavioral paradigm. Animals from thecontrol and trained groups were killed 6 h after training, thehippocampal tissues were used to isolate RNA for reverse transcrip-tion of cDNA and radioactive probes were generated from thesecDNA samples to differentially screen an adult rat brain cDNA library.Positive clones were isolated and their cDNA inserts were used tomake radioactive probes for northern blot analysis. One of the positiveclones, SNAP-25, displayed a differential expression pattern ofhippocampal mRNA between the control and the animals 6 h afterstep-down inhibitory avoidance training (Fig. 1).As SNAP-25 has been shown by in vitro studies to be a neuronal
protein important for synaptic function, it appeared to be a goodcandidate for functional involvement in learning ⁄memory. Thus, itwas chosen for further investigation.
SNAP-25 expression in the hippocampus is elevated afterstep-down inhibitory avoidance training
In situ hybridization was used to examine the SNAP-25 mRNA levelin the hippocampus. SNAP-25 is expressed in the dentate gyrus, CA1and CA3 regions of the hippocampus. The message level wasincreased in these regions after the step-down inhibitory avoidance
training (Fig. 2A). The ratios of the SNAP-25 mRNA signal fromtrained and control groups are: the dentate gyrus, 1.21 ± 0.09; CA1,1.22 ± 0.07 and CA3, 1.40 ± 0.07 (Fig. 2B; P < 0.05 for training vs.control, unpaired t-test), indicating that the behavioral trainingelevated SNAP-25 expression in the hippocampus.Western blot analysis was employed to determine the SNAP-25
protein level in the hippocampus, using an anti-SNAP-25 antibody. Asshown in Fig. 3, the SNAP-25 protein level was significantly increasedin the hippocampus after step-down inhibitory avoidance training(P < 0.05 for training vs. control, unpaired t-test). The protein levelsof SNAP-25 in the trained group increased by a factor of 1.46 ± 0.11over the control group.
SNAP-25 antisense oligonucleotide impairs long-term memoryfor step-down inhibitory avoidance
As SNAP-25 mRNA appeared to be up-regulated upon step-downinhibitory avoidance training, we wanted to determine whether acausal relationship exists between the SNAP-25 level and the animals’behavioral change. In other words, is memory affected if the SNAP-25level is reduced?
SNAP-25
Tubulin
Rel
ativ
e de
nsity
Control 0 1 2.5 6 24
0.5
0.4
0.3
0.2
0.1
0
A
B
Fig. 5. Transient increase in hippocampal SNAP-25 protein following con-textual fear conditioning. (A) Representative western blot analysis of proteinextracted from the hippocampus of animals with sham conditioning (control) orat various time points after contextual fear conditioning. Time ‘0’ indicatesimmediately after the fear conditioning. Tubulin was used as an internal controlfor protein loading. (B) Summary of western blot data (n ¼ 6). Bars representmeans ± SEM. *P < 0.05 vs. control, unpaired t-test.
Context
Cue
Saline
Saline
Scramble
Scramble
Antisense
Antisense
Pre-Training Immediate 1 hr 24 hr
Free
zing
(%
)Fr
eezi
ng (
%)
100
80
60
40
20
0
100
80
60
40
20
0Pre-CS Pre-CS CS CS
1.5 hr 24.5 hr
A
B
Fig. 6. Long-term contextual fear memory was impaired following intra-CA1infusion of the SNAP-25 antisense oligonucleotide. Rats received bilateralintra-CA1 infusion of saline, scramble oligonucleotide or the SNAP-25antisense oligonucleotide. Fear conditioning began 6 h after solution infusion.The animal’s freezing score is shown as a percentage of the total testingduration. Data are shown as mean ± SEM. (A) Freezing response to thetraining chamber pre-conditioning, immediately after conditioning and either at1 or 24 h after conditioning. n ¼ 19–22 for pre-conditioning and immediatelyafter conditioning; n ¼ 8–14 for 1 and 24 h after conditioning. Compared withthe control groups, the SNAP-25 antisense group displayed a similar extent offreezing scores immediately and 1 h after conditioning but showed significantlypoorer memory retention at 24 h post-training. *P < 0.05 vs. controls, unpairedt-test. (B) Freezing response to the auditory cue at either 1.5 or 24.5 h post-conditioning. Animals from the 1- and 24-h groups in A were tested forauditory fear memory. There was no difference among the groups in eithershort- or long-term auditory fear memory. CS, conditioned stimulus.
1598 Q. Hou et al.
ª 2004 Federation of European Neuroscience Societies, European Journal of Neuroscience, 20, 1593–1603
To reduce the SNAP-25 level, we employed an antisense oligonu-cleotide that has previously been shown to effectively reduce theSNAP-25 level (Osen-Sand et al., 1993). This antisense oligonucle-otide or a control solution (scramble oligonucleotide or saline) wasinfused into the brain via a cannula in the third ventricle connected toan osmotic pump and the animals were tested for memory retentionafter step-down inhibitory avoidance training. As shown in Fig. 4A,animals in all three groups spent very little time on the platform beforeconditioning, with an average of less than 25 s per animal. Theantisense group showed no difference in latency of stepping downfrom the platform compared with the control groups, indicating thatintracerebroventricular infusion of the SNAP-25 antisense oligonu-cleotide did not compromise the animals’ general mobility or theirmotivation for stepping down from the platform to explore theenvironment.
When tested 1 h after conditioning, the antisense group, like thecontrol groups, did not step down from the platform during the entire5-min period of observation, with a cut-off time at 600 s (Fig. 4A).This indicates that short-term memory for inhibitory avoidance wassimilar in both the antisense and control groups. When tested 24 hafter conditioning, all groups stepped down from the platform withinthe 10-min observation time. However, animals in the antisense groupstepped down significantly earlier than the control groups (P < 0.05for antisense vs. controls, unpaired t-test), with a latency of217 ± 23 s, whereas the saline and scramble oligonucleotide groupsstayed on the platform for 458 ± 26 and 467 ± 26 s, respectively(Fig. 4A). This indicates that long-term memory for inhibitoryavoidance was impaired in the antisense group.
To verify that the antisense oligonucleotide would inhibit theSNAP-25 expression as reported previously (Osen-Sand et al.,1993), we tested it in hippocampal neuronal cultures. As shown inFig. 4B, the antisense oligonucleotide reduced the SNAP-25 proteinlevel (*P < 0.05 vs. controls, unpaired t-test). We next sought todetermine whether such an inhibitory effect was observed in vivo bymeasuring the SNAP-25 mRNA levels using in situ hybridizationwith hippocampal sections. As shown in Fig. 4C, step-downinhibitory avoidance training resulted in an increase in the SNAP-25 mRNA level in the hippocampus (#P < 0.05 control vs. training,unpaired t-test) and such an increase was selectively inhibited by theantisense oligonucleotide (*P < 0.05 training vs. antisense, unpairedt-test). These results indicate that the antisense oligonucleotide waseffective in reducing the training-related increase in SNAP-25mRNA.
Transient increase in hippocampal SNAP-25 protein uponfear-conditioning training
To better delineate the relationship between the molecular changes inSNAP-25 and the animal’s behavior, we further examined the SNAP-25 protein levels at various times after the animals received contextualfear conditioning, which is a hippocampus-dependent task (Kim &Fanselow, 1992; Bevilaqua et al., 1997; Maren et al., 1997;Roozendaal & McGaugh, 1997; Anagnostaras et al., 1999; Roo-zendaal et al., 1999).As shown in Fig. 5, the steady-state protein level of SNAP-25
showed a transient increase, reached a peak at 2.5 h and returned to
Saline
AntisenseScramble
Saline
AntisenseScramble
Saline
Saline
Antisense
Antisense
Scramble
Scramble
Training (trial)
Visible Platform Test (trial)
Late
ncy
(s)
Sw
im s
peed
(cm
/s)
Late
ncy
(s)
Training (trial)
1 hr
48 h
r
RetentionRetention
48 h
r
60
50
40
40
30
30
30
20
20
20
1010
10
0 0
0
1 1
1
2 2
2
3 3
3
4 45 56 67 78 89 910 1011 1112 12
A B
DC
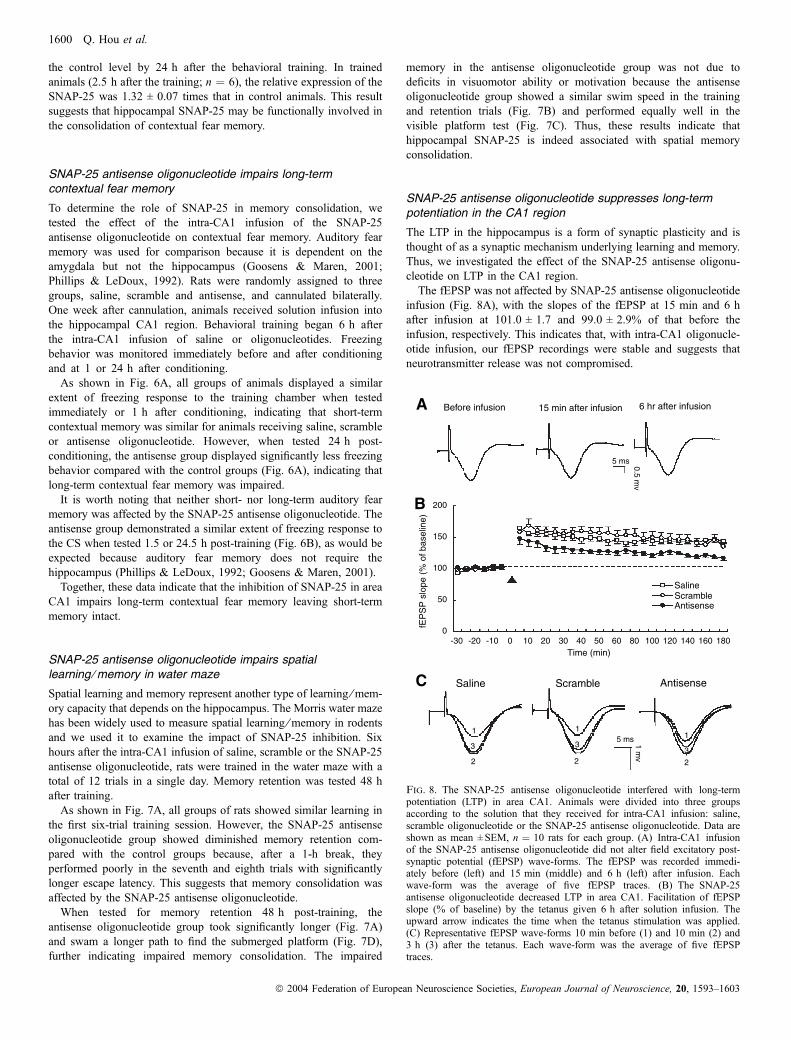
Fig. 7. Spatial memory in the Morris water maze was impaired following intra-CA1 infusion of the SNAP-25 antisense oligonucleotide. Three groups of ratsreceived bilateral intra-CA1 infusion of saline, scramble oligonucleotide or the SNAP-25 antisense oligonucleotide (n ¼ 8–14 for each group). Training in the watermaze began 6 h after infusion, with two training sessions of six trials each and a 1-h interval between the two sessions. Memory was tested 48 h after training. Dataare shown as mean ± SEM. (A) Escape latency to find the submerged platform during training and 48-h retention testing. Cut-off time was 60 s. All groups showedsimilar learning capacity during the first session. *Significant difference from the trial before the 1- or 48-h break (P < 0.05, paired t-test) as well as from the controlgroups for the same trial (P < 0.05, unpaired t-test), suggesting diminished memory consolidation. (B) Swim speed during training and 48-h retention testing. Therewas no significant difference among the three groups. (C) Escape latency to locate the visible platform during the visible-platform testing. There was no significantdifference among the three groups. (D) Representative swim path of each group during 48-h retention testing. The antisense group swam a longer path length thanthe control groups to find the submerged platform.
SNAP-25 in area CA1 and memory consolidation 1599
ª 2004 Federation of European Neuroscience Societies, European Journal of Neuroscience, 20, 1593–1603
the control level by 24 h after the behavioral training. In trainedanimals (2.5 h after the training; n ¼ 6), the relative expression of theSNAP-25 was 1.32 ± 0.07 times that13 in control animals. This resultsuggests that hippocampal SNAP-25 may be functionally involved inthe consolidation of contextual fear memory.
SNAP-25 antisense oligonucleotide impairs long-termcontextual fear memory
To determine the role of SNAP-25 in memory consolidation, wetested the effect of the intra-CA1 infusion of the SNAP-25antisense oligonucleotide on contextual fear memory. Auditory fearmemory was used for comparison because it is dependent on theamygdala but not the hippocampus (Goosens & Maren, 2001;Phillips & LeDoux, 1992). Rats were randomly assigned to threegroups, saline, scramble and antisense, and cannulated bilaterally.One week after cannulation, animals received solution infusion intothe hippocampal CA1 region. Behavioral training began 6 h afterthe intra-CA1 infusion of saline or oligonucleotides. Freezingbehavior was monitored immediately before and after conditioningand at 1 or 24 h after conditioning.As shown in Fig. 6A, all groups of animals displayed a similar
extent of freezing response to the training chamber when testedimmediately or 1 h after conditioning, indicating that short-termcontextual memory was similar for animals receiving saline, scrambleor antisense oligonucleotide. However, when tested 24 h post-conditioning, the antisense group displayed significantly less freezingbehavior compared with the control groups (Fig. 6A), indicating thatlong-term contextual fear memory was impaired.It is worth noting that neither short- nor long-term auditory fear
memory was affected by the SNAP-25 antisense oligonucleotide. Theantisense group demonstrated a similar extent of freezing response tothe CS when tested 1.5 or 24.5 h post-training (Fig. 6B), as would beexpected because auditory fear memory does not require thehippocampus (Phillips & LeDoux, 1992; Goosens & Maren, 2001).Together, these data indicate that the inhibition of SNAP-25 in area
CA1 impairs long-term contextual fear memory leaving short-termmemory intact.
SNAP-25 antisense oligonucleotide impairs spatiallearning ⁄memory in water maze
Spatial learning and memory represent another type of learning ⁄mem-ory capacity that depends on the hippocampus. The Morris water mazehas been widely used to measure spatial learning ⁄memory in rodentsand we used it to examine the impact of SNAP-25 inhibition. Sixhours after the intra-CA1 infusion of saline, scramble or the SNAP-25antisense oligonucleotide, rats were trained in the water maze with atotal of 12 trials in a single day. Memory retention was tested 48 hafter training.As shown in Fig. 7A, all groups of rats showed similar learning in
the first six-trial training session. However, the SNAP-25 antisenseoligonucleotide group showed diminished memory retention com-pared with the control groups because, after a 1-h break, theyperformed poorly in the seventh and eighth trials with significantlylonger escape latency. This suggests that memory consolidation wasaffected by the SNAP-25 antisense oligonucleotide.When tested for memory retention 48 h post-training, the
antisense oligonucleotide group took significantly longer (Fig. 7A)and swam a longer path to find the submerged platform (Fig. 7D),further indicating impaired memory consolidation. The impaired
memory in the antisense oligonucleotide group was not due todeficits in visuomotor ability or motivation because the antisenseoligonucleotide group showed a similar swim speed in the trainingand retention trials (Fig. 7B) and performed equally well in thevisible platform test (Fig. 7C). Thus, these results indicate thathippocampal SNAP-25 is indeed associated with spatial memoryconsolidation.
SNAP-25 antisense oligonucleotide suppresses long-termpotentiation in the CA1 region
The LTP in the hippocampus is a form of synaptic plasticity and isthought of as a synaptic mechanism underlying learning and memory.Thus, we investigated the effect of the SNAP-25 antisense oligonu-cleotide on LTP in the CA1 region.The fEPSP was not affected by SNAP-25 antisense oligonucleotide
infusion (Fig. 8A), with the slopes of the fEPSP at 15 min and 6 hafter infusion at 101.0 ± 1.7 and 99.0 ± 2.9% of that before theinfusion, respectively. This indicates that, with intra-CA1 oligonucle-otide infusion, our fEPSP recordings were stable and suggests thatneurotransmitter release was not compromised.
Before infusion 15 min after infusion 6 hr after infusion
5 ms
5 ms
0.5 mv
1 mv
Saline
Saline
Scramble
Scramble
Time (min)
Antisense
Antisense
fEP
SP
slo
pe (
% o
f bas
elin
e)
200
150
100
50
0-30 -20 -10 0 10 20 30 40 50 60 80 100 120 140 160 180
11
3 32 2
1
3
2
A
C
B
Fig. 8. The SNAP-25 antisense oligonucleotide interfered with long-termpotentiation (LTP) in area CA1. Animals were divided into three groupsaccording to the solution that they received for intra-CA1 infusion: saline,scramble oligonucleotide or the SNAP-25 antisense oligonucleotide. Data areshown as mean ± SEM, n ¼ 10 rats for each group. (A) Intra-CA1 infusionof the SNAP-25 antisense oligonucleotide did not alter field excitatory post-synaptic potential (fEPSP) wave-forms. The fEPSP was recorded immedi-ately before (left) and 15 min (middle) and 6 h (left) after infusion. Eachwave-form was the average of five fEPSP traces. (B) The SNAP-25antisense oligonucleotide decreased LTP in area CA1. Facilitation of fEPSPslope (% of baseline) by the tetanus given 6 h after solution infusion. Theupward arrow indicates the time when the tetanus stimulation was applied.(C) Representative fEPSP wave-forms 10 min before (1) and 10 min (2) and3 h (3) after the tetanus. Each wave-form was the average of five fEPSPtraces.
1600 Q. Hou et al.
ª 2004 Federation of European Neuroscience Societies, European Journal of Neuroscience, 20, 1593–1603
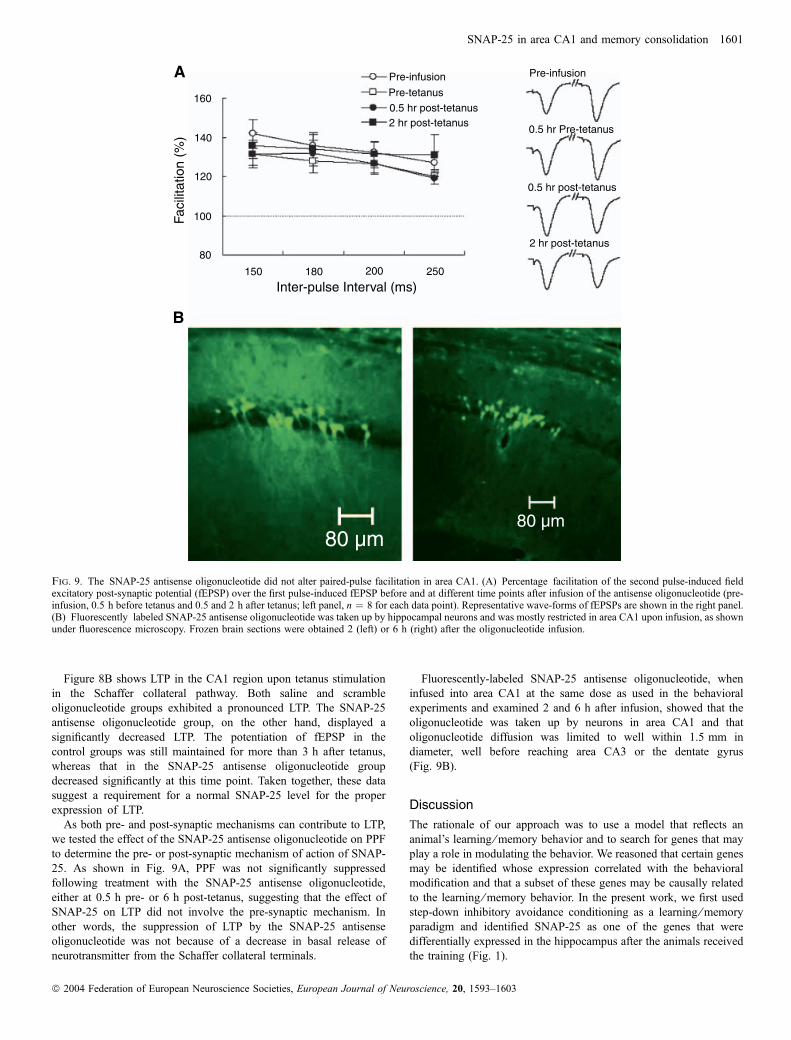
Figure 8B shows LTP in the CA1 region upon tetanus stimulationin the Schaffer collateral pathway. Both saline and scrambleoligonucleotide groups exhibited a pronounced LTP. The SNAP-25antisense oligonucleotide group, on the other hand, displayed asignificantly decreased LTP. The potentiation of fEPSP in thecontrol groups was still maintained for more than 3 h after tetanus,whereas that in the SNAP-25 antisense oligonucleotide groupdecreased significantly at this time point. Taken together, these datasuggest a requirement for a normal SNAP-25 level for the properexpression of LTP.
As both pre- and post-synaptic mechanisms can contribute to LTP,we tested the effect of the SNAP-25 antisense oligonucleotide on PPFto determine the pre- or post-synaptic mechanism of action of SNAP-25. As shown in Fig. 9A, PPF was not significantly suppressedfollowing treatment with the SNAP-25 antisense oligonucleotide,either at 0.5 h pre- or 6 h post-tetanus, suggesting that the effect ofSNAP-25 on LTP did not involve the pre-synaptic mechanism. Inother words, the suppression of LTP by the SNAP-25 antisenseoligonucleotide was not because of a decrease in basal release ofneurotransmitter from the Schaffer collateral terminals.
Fluorescently-labeled SNAP-25 antisense oligonucleotide, wheninfused into area CA1 at the same dose as used in the behavioralexperiments and examined 2 and 6 h after infusion, showed that theoligonucleotide was taken up by neurons in area CA1 and thatoligonucleotide diffusion was limited to well within 1.5 mm indiameter, well before reaching area CA3 or the dentate gyrus(Fig. 9B).
Discussion
The rationale of our approach was to use a model that reflects ananimal’s learning ⁄memory behavior and to search for genes that mayplay a role in modulating the behavior. We reasoned that certain genesmay be identified whose expression correlated with the behavioralmodification and that a subset of these genes may be causally relatedto the learning ⁄memory behavior. In the present work, we first usedstep-down inhibitory avoidance conditioning as a learning ⁄memoryparadigm and identified SNAP-25 as one of the genes that weredifferentially expressed in the hippocampus after the animals receivedthe training (Fig. 1).
Pre-infusion Pre-infusion
Pre-tetanus0.5 hr post-tetanus
0.5 hr post-tetanus
0.5 hr Pre-tetanus2 hr post-tetanus
2 hr post-tetanus
Inter-pulse Interval (ms)
80 µm80 µm
Faci
litat
ion
(%)
160
140
120
100
80
150 180 200 250
A
B
Fig. 9. The SNAP-25 antisense oligonucleotide did not alter paired-pulse facilitation in area CA1. (A) Percentage facilitation of the second pulse-induced fieldexcitatory post-synaptic potential (fEPSP) over the first pulse-induced fEPSP before and at different time points after infusion of the antisense oligonucleotide (pre-infusion, 0.5 h before tetanus and 0.5 and 2 h after tetanus; left panel, n ¼ 8 for each data point). Representative wave-forms of fEPSPs are shown in the right panel.(B) Fluorescently labeled SNAP-25 antisense oligonucleotide was taken up by hippocampal neurons and was mostly restricted in area CA1 upon infusion, as shownunder fluorescence microscopy. Frozen brain sections were obtained 2 (left) or 6 h (right) after the oligonucleotide infusion.
SNAP-25 in area CA1 and memory consolidation 1601
ª 2004 Federation of European Neuroscience Societies, European Journal of Neuroscience, 20, 1593–1603

SNAP-25 is a synaptosomal protein (Geddes et al., 1990; Oyleret al., 1989, 1992). It is involved in a number of neuronalfunctions, including exocytosis-based neurotransmitter release(Banerjee et al., 1996; Mehta et al., 1996) and axonal ⁄ dendriticgrowth (Osen-Sand et al., 1993; Grosse et al., 1999). SNAP-25mRNA up-regulation is also associated with LTP expression inthe hippocampus (Roberts et al., 1998). The involvement ofSNAP-25 in regulating axonal ⁄ dendritic growth and hippocam-pal LTP points to a possible role of SNAP-25 in learning andmemory.Indeed, our results showed this to be the case. In animals
receiving step-down inhibitory avoidance training, SNAP-25 wasup-regulated in the hippocampus both at the mRNA (Fig. 1) andprotein level (Fig. 3). In situ hybridization demonstrated anenhancement of the neuronal SNAP-25 message upon training inthe dentate gyrus, CA1 and CA3 regions (Fig. 2). Functionalperturbation experiments showed that animals receiving intracere-broventricular infusion of the SNAP-25 antisense oligonucleotideexhibited impaired long-term memory for step-down avoidance withintact short-term memory (Fig. 4A). As antisense oligonucleotidesare known to inhibit protein functions by retarding mRNAtranslation and selective mRNA degradation (Crooke, 2000) andthe oligonucleotide that we employed has been shown to reduceSNAP-25 expression (Osen-Sand et al., 1993), our results suggest aclose relationship of the altered SNAP-25 level with learning and ⁄ ormemory consolidation.Interestingly, contextual fear conditioning triggered a transient
increase in SNAP-25 protein levels (Fig. 5) which peaked at 2.5 hafter conditioning and subsided to the control level by 24 h. Thetiming of this elevated expression is consistent with the time windowfor memory consolidation (McGaugh, 2000), suggesting a causalrelationship of the SNAP-25 expression level with memory consol-idation. Indeed, intra-CA1 infusion of the SNAP-25 antisenseoligonucleotide produced a deficit in long-term contextual fearmemory without affecting short-term memory (Fig. 6A). In corrobor-ation with this evidence, we also observed an impaired spatial memoryfor the water maze (Fig. 7).Hippocampal LTP is thought to be a form of synaptic plasticity
that underlies learning ⁄memory (Morris et al., 1986; Morris, 1989;Bliss & Collingridge, 1993; Martin et al., 2000). We found that LTPwas significantly suppressed by intra-CA1 infusion of the SNAP-25antisense oligonucleotide (Fig. 8). Although extensive work hasidentified the role of SNAP-25 in pre-synaptic neurotransmitterrelease, our results clearly indicate that the impact of SNAP-25 onLTP is not mediated pre-synaptically. Specifically, PPF was notaffected by the SNAP-25 antisense oligonucleotide (Fig. 9A).Furthermore, the fluorescently labeled oligonucleotide was mostlyretained in area CA1 upon infusion (Fig. 9B) without noticeablediffusion outside the CA1 region. Thus, neuronal cell bodies in areaCA3 are unlikely to be affected by the SNAP-25 antisenseoligonucleotide.Increasing evidence suggests that post-synaptic SNAP-25 is
involved in the mechanism underlying trafficking of N-methyl-d-aspartate receptors to the cell surface (Lan et al., 2001a,b). Down-regulation of post-synaptic SNAP-25 in area CA1 would reducedelivery of N-methyl-d-aspartate receptors to the post-synapticmembrane and thus interferes with synaptic plasticity, like LTP,resulting in a deficit in memory consolidation.It should be pointed out that oligonucleotides have been known
to cause a non-specific toxicity effect which may influenceexperimental results. We are very aware of this potential concernand have used two or three controls in each experiment. In most
cases, we used saline, scramble oligonucleotide and a mis-senseoligonucleotide as controls. The saline injection controls for volumeinjection effects on the brain structure. The scramble oligonucleotideserves as a control for sequence non-specific toxicity of theoligonucleotide. The mis-sense oligonucleotide, because it resemblesthe sequence of the antisense oligonucleotide with only a few basesdifference, serves as a control for any serendipitous interaction withother genes by the antisense oligonucleotide. While these controlsdo not completely eliminate the potential for artifacts, together theydo represent a reasonable set of controls to show that theresults that we observed are indeed specific for the antisenseoligonucleotide.In summary, we identified SNAP-25 as a differentially expressed
gene in area CA1 upon behavioral training. SNAP-25 expression istransiently elevated after contextual fear conditioning, coincidingwith the time window for memory consolidation. The inhibition ofSNAP-25 leads to impaired long-term contextual fear memory andspatial memory as well as decreased LTP. Together, our resultssuggest that SNAP-25 in the CA1 region is involved in memoryconsolidation.
Acknowledgements
This work was supported in part by the Life Science Special Fund of theChinese Academy of Sciences for Human Genome Research (KJ95T-06 andKSCX1-Y02 to M.J., N.J. and B.L.), National Natural Science Foundation ofChina (grant nos 39930090 and 90208011 to N.J. and 39970244 and 30225023to B.L.), National Key Basic Research and Development Program of China(G1999054000 to N.J. and B.L. and 2002CB713802 to N.J.), National High-Tech Research and Development Program of China (2002AA223081 to B.L.)and National Institutes of Health (USA) grants (DA09444, DA11891 andDA1347 to L.Y.). We thank Drs M. E. Bardgett, J. T. Meij, J. A. Strong and M.Xu for helpful comments.
Abbreviations
CS, conditioned stimulus; fEPSP, field excitatory post-synaptic potential; LTP,long-term potentiation; PBS, phosphate-buffered saline; PPF, paired-pulsefacilitation.
References
Abel, T., Nguyen, P.V., Barad, M., Deuel, T.A., Kandel, E.R. & Bourtch-ouladze, R. (1997) Genetic demonstration of a role for PKA in the late phaseof LTP and in hippocampus-based long-term memory. Cell, 88, 615–626.
Anagnostaras, S.G., Maren, S. & Fanselow, M.S. (1999) Temporally gradedretrograde amnesia of contextual fear after hippocampal damage in rats:within-subjects examination. J. Neurosci., 19, 1106–1114.
Banerjee, A., Kowalchyk, J.A., DasGupta, B.R. & Martin, T.F.J. (1996) SNAP-25 is required for a late postdocking step in Ca2+-dependent exocytosis.J. Biol. Chem., 271, 20 227–20 230.
Bevilaqua, L., Ardenghi, P., Schroder, N., Bromberg, E., Schmitz, P.K.,Schaeffer, E., Quevedo, J., Bianchin, M., Walz, R., Medina, J.H. &Izquierdo, I. (1997) Drugs acting upon the cyclic adenosinemonophosphate ⁄ protein kinase A signalling pathway modulate memoryconsolidation when given late after training into rat hippocampus but notamygdala. Behav. Pharmacol., 8, 331–338.
Bliss, T.V. & Collingridge, G.L. (1993) A synaptic model of memory: long-term potentiation in the hippocampus. Nature, 361, 31–39.
Branks, P.L. & Wilson, M.C. (1986) Patterns of gene expression in the murinebrain revealed by in situ hybridization of brain-specific mRNAs. Mol. BrainRes., 1, 1–16.
Brewer, G.J. (1997) Isolation and culture of adult rat hippocampal neurons.J. Neurosci. Meth., 71, 143–155.
Brewer, G.J. (1999) Regeneration and proliferation of embryonic and adult rathippocampal neurons in culture. Exp. Neurol., 159, 237–247.
Crooke, S.T. (2000) Progress in antisense technology: the end of the beginning.Meth. Enzymol., 313, 3–45.
1602 Q. Hou et al.
ª 2004 Federation of European Neuroscience Societies, European Journal of Neuroscience, 20, 1593–1603

Eichenbaum, H. & Otto, T. (1993) LTP and memory: can we enhance theconnection? Trends Neurosci., 16, 163–164.
Elgersma, Y. & Silva, A.J. (1999) Molecular mechanisms of synaptic plasticityand memory. Curr. Opin. Neurobiol., 9, 209–213.
Geddes, J.W., Hess, E.J., Hart, R.A., Kesslak, J.P., Cotman, C.W. & Wilson,M.C. (1990) Lesions of hippocampal circuitry define synaptosomal-associated protein-25 (SNAP-25) as a novel presynaptic marker. Neu-roscience, 38, 515–525.
Goosens, K.A. & Maren, S. (2001) Contextual and auditory fear conditioningare mediated by the lateral, basal, and central amygdaloid nuclei in rats.Learn. Mem., 8, 148–155.
Grosse, G., Grosse, J., Tapp, R., Kuchinke, J., Gorsleben, M., Fetter, I.,Hohne-Zell, B., Gratzl, M. & Bergmann, M. (1999) SNAP-25 requirementfor dendritic growth of hippocampal neurons. J. Neurosci. Res., 56, 539–546.
Guzowski, J.F. & McGaugh, J.L. (1997) Antisense oligodeoxynucleotide-mediated disruption of hippocampal cAMP response element binding proteinlevels impairs consolidation of memory for water maze training. Proc. NatlAcad. Sci. U.S.A., 94, 2693–2698.
Heise, G.A. (1984) Behavioral methods for measuring effects of drugs onlearning and memory in animals. Med. Res. Rev., 4, 535–558.
Huang, Y.Y., Kandel, E.R., Varshavsky, L., Brandon, E.P., Qi, M., Idzerda,R.L., McKnight, G.S. & Bourtchouladze, R. (1995) A genetic test of theeffects of mutations in PKA on mossy fiber LTP and its relation to spatial andcontextual learning. Cell, 83, 1211–1222.
Ikegami, S. & Inokuchi, K. (2000) Antisense DNA against calcineurinfacilitates memory in contextual fear conditioning by lowering the thresholdfor hippocampal long-term potentiation induction. Neuroscience, 98, 637–646.
Izquierdo, I., Fin, C., Schmitz, P.K., Da Silva, R.C., Jerusalinsky, D., Quillfeldt,J.A., Ferreira, M.B., Medina, J.H. & Bazan, N.G. (1995) Memoryenhancement by intrahippocampal, intraamygdala, or intraentorhinal infu-sion of platelet-activating factor measured in an inhibitory avoidance task.Proc. Natl Acad. Sci. USA, 92, 5047–5051.
Izquierdo, I., Barros, D.M., Mello e Souza, T., de Souza, M.M., Izquierdo, L.A.& Medina, J.H. (1998) Mechanisms for memory types differ. Nature, 393,635–636.14
Kim, J.J. & Fanselow, M.S. (1992) Modality-specific retrograde amnesia offear. Science, 256, 675–677.
Kim, J.J., Rison, R.A. & Fanselow, M.S. (1993) Effects of amygdala,hippocampus, and periaqueductal gray lesions on short- and long-termcontextual fear. Behav. Neurosci., 107, 1093–1098.
Lan, J.Y., Skeberdis, V.A., Jover, T., Grooms, S.Y., Lin, Y., Araneda, R.C.,Zheng, X., Bennett, M.V. & Zukin, R.S. (2001a) Protein kinase C modulatesNMDA receptor trafficking and gating. Nat. Neurosci., 4, 382–390.
Lan, J.Y., Skeberdis, V.A., Jover, T., Zheng, X., Bennett, M.V. & Zukin, R.S.(2001b) Activation of metabotropic glutamate receptor 1 accelerates NMDAreceptor trafficking. J. Neurosci., 21, 6058–6068.
Logue, S.F., Paylor, R. & Wehner, J.M. (1997) Hippocampal lesions causelearning deficits in inbred mice in the Morris water maze and conditioned-fear task. Behav. Neurosci., 111, 104–113.
Low, P., Norlin, T., Risinger, C., Larhammar, D., Pieribone, V.A., Shupliakov,O. & Brodin, L. (1999) Inhibition of neurotransmitter release in the lampreyreticulospinal synapse by antibody-mediated disruption of SNAP-25function. Eur. J. Cell Biol., 78, 787–793.
Maren, S., Aharonov, G. & Fanselow, M.S. (1997) Neurotoxic lesions of thedorsal hippocampus and Pavlovian fear conditioning in rats. Behav. BrainRes., 88, 261–274.
Martin, S.J., Grimwood, P.D. & Morris, R.G. (2000) Synaptic plasticity andmemory: an evaluation of the hypothesis. Annu. Rev. Neurosci., 23, 649–711.
McGaugh, J.L. (2000) Memory — a century of consolidation. Science, 287,248–251.
Mehta, P.P., Battenberg, E. & Wilson, M.C. (1996) SNAP-25 and synapto-tagmin involvement in the final Ca(2+)-dependent triggering of neurotrans-mitter exocytosis. Proc. Natl Acad. Sci. U.S.A., 93, 10 471–10 476.
Morris, R.G. (1989) Synaptic plasticity and learning: selective impairment oflearning rats and blockade of long-term potentiation in vivo by the N-methyl-D-aspartate receptor antagonist AP5. J. Neurosci., 9, 3040–3057.
Morris, R.G., Anderson, E., Lynch, G.S. & Baudry, M. (1986) Selectiveimpairment of learning and blockade of long-term potentiation by anN-methyl-D-aspartate receptor antagonist, AP5. Nature, 319, 774–776.
Morris, R.G., Garrud, P., Rawlins, J.N. & O’Keefe, J. (1982) Place navigationimpaired in rats with hippocampal lesions. Nature, 297, 681–683.
Osen-Sand, A., Catsicas, M., Staple, J.K., Jones, K.A., Ayala, G., Knowles, J.,Grenningloh, G. & Catsicas, S. (1993) Inhibition of axonal growth by SNAP-25 antisense oligonucleotides in vitro and in vivo. Nature, 364, 445–448.
Oyler, G.A., Higgins, G.A., Hart, R.A., Battenberg, E., Billingsley, M., Bloom,F.E. & Wilson, M.C. (1989) The identification of a novel synaptosomal-associated protein, SNAP-25, differentially expressed by neuronal sub-populations. J. Cell Biol., 109, 3039–3052.
Oyler, G.A., Polli, J.W., Higgins, G.A., Wilson, M.C. & Billingsley, M.L.(1992) Distribution and expression of SNAP-25 immunoreactivity in ratbrain, rat PC-12 cells and human SMS-KCNR neuroblastoma cells. BrainRes. Dev. Brain Res., 65, 133–146.
Paxinos, G. & Watson, C. (1986) The Rat Brain in Stereotaxic Coordinates.Academic Press, San Diego, CA.
Phillips, R.G. & LeDoux, J.E. (1992) Differential contribution of amygdala andhippocampus to cued and contextual fear conditioning. Behav. Neurosci.,106, 274–285.
Phillips, R.G. & LeDoux, J.E. (1995) Lesions of the fornix but not theentorhinal or perirhinal cortex interfere with contextual fear conditioning.J. Neurosci., 15, 5308–5315.
Roberts, L.A., Morris, B.J. & O’Shaughnessy, C.T. (1998) Involvement of twoisoforms of SNAP-25 in the expression of long-term potentiation in the rathippocampus. Neuroreport, 9, 33–36.
Roozendaal, B. & McGaugh, J.L. (1997) Basolateral amygdala lesions blockthe memory-enhancing effect of glucocorticoid administration in the dorsalhippocampus of rats. Eur. J. Neurosci., 9, 76–83.
Roozendaal, B., Nguyen, B.T., Power, A.E. & McGaugh, J.L. (1999)Basolateral amygdala noradrenergic influence enables enhancement ofmemory consolidation induced by hippocampal glucocorticoid receptoractivation. Proc. Natl Acad. Sci. USA, 96, 11 642–11 647.
Sandin, J., Georgieva, J., Schott, P.A., Ogren, S.O. & Terenius, L. (1997)Nociceptin ⁄ orphanin FQ microinjected into hippocampus impairs spatiallearning in rats. Eur. J. Neurosci., 9, 194–197.
Sanes, J.R. & Lichtman, J.W. (1999) Can molecules explain long-termpotentiation? Nat. Neurosci., 2, 597–604.
Selden, N.R., Everitt, B.J., Jarrard, L.E. & Robbins, T.W. (1991) Complemen-tary roles for the amygdala and hippocampus in aversive conditioning toexplicit and contextual cues. Neuroscience, 42, 335–350.
Silva, A.J., Kogan, J.H., Frankland, P.W. & Kida, S. (1998) CREB andmemory. Annu. Rev. Neurosci., 21, 127–148.
Squire, L.R. & Kandel, E.R. (1999) Memory: From Mind to Molecules.Scientific American Library, New York.
Stevens, C.F. (1998) A million dollar question: does LTP ¼ memory? Neuron,20, 1–2.
Sutherland, R.J., Kolb, B. & Whishaw, I.Q. (1982) Spatial mapping: definitivedisruption by hippocampal or medial frontal cortical damage in the rat.Neurosci. Lett., 31, 271–276.
Tang, Y.P., Wang, H., Feng, R., Kyin, M. & Tsien, J.Z. (2001) Differentialeffects of enrichment on learning and memory function in NR2B transgenicmice. Neuropharmacology, 41, 779–790.
Yang, J., Bian, W., Gao, X., Chen, L. & Jing, N. (2000) Nestin expressionduring mouse eye and lens development. Mech. Dev., 94, 287–291.
SNAP-25 in area CA1 and memory consolidation 1603
ª 2004 Federation of European Neuroscience Societies, European Journal of Neuroscience, 20, 1593–1603