interactions des ondes et des particules avec la matière biologique 1 ère partie
TRANSCRIPT

Interactions des ondes et des particules avec la matière
biologique
1ère Partie

Matière
La matière biologique est formée par :
• Cellules : formées par plusieurs molécules (ADN,
Protéines, glucides, eau…) ;
• Tissu : ensemble de cellules ;
• Organe : ensemble formé de plusieurs tissus ;
• Système : ensemble formé de divers organes qui
collaborent ;
• Organisme : ensemble intégré des systèmes.

Dans une cellule, il y a environ 1012 molécules d’eau
et 109 macromolécules, divers organites et une seule
molécule d’ADN. Ces molécules qu’on appelle
biomolécules sont des composés chimiques que l’on
trouve dans les organismes vivants.


Molécule
Atome
Nuage électronique
Noyau
Protons Neutrons
3 Quarks2 up 1down
3 Quarks
1up 2down
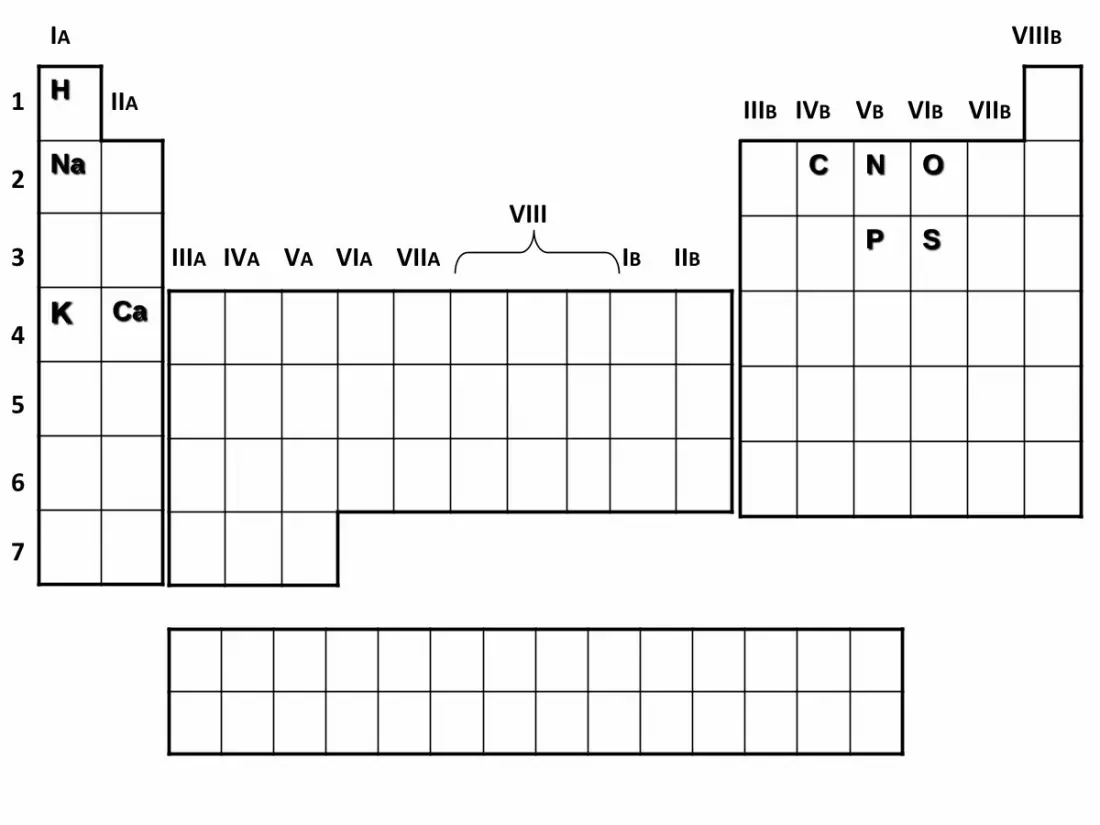
IA
H
Na
K Ca
C N O
P S
IIA
IIIA IVA VA VIA VIIA
VIII
IB IIB
IIIB IVB VB VIB VIIB
VIIIB
1
2
3
4
5
6
7

D'autres éléments existent, ce sont les oligoéléments
indispensables à la vie

IA
H
Na
Ca Fe Co Cu Zn
C N O
P S
I
IIA
IIIA IVA VA VIA VIIA
VIII
IB IIB
IIIB IVB VB VIB VIIB
VIIIB
1
2
3
4
5
6
7
le fer pour l'hémoglobine donc le transport de l'oxygène dans le sang
le zinc et un certain nombre de cations divalents qui maintiennent la structure de certaines enzymes (métalloprotéases
l'iode qui est un constituant essentiel de certaines hormones produites par la glande thyroïde

Un électron est donc d’autant plus lié que n est
plus petit (proche du noyau). Une relation plus ou
moins empirique donne la valeur de l’énergie de
liaison d’un électron pour un atome de numéro
atomique Z et pour une couche de nombre
quantique principal n :
En = - 13,6 . Z2 / n2
n=1
n=2
n=3

Transitions électroniques
• Il s’agit des différents déplacements
d’électrons entre les niveaux d’énergie
possibles, avec les phénomènes
d’absorption ou d’émission qui les
accompagnent.

E (eV) = E2- E1 = -13,6 [ 1 / n22 - 1 / n1
2 ]
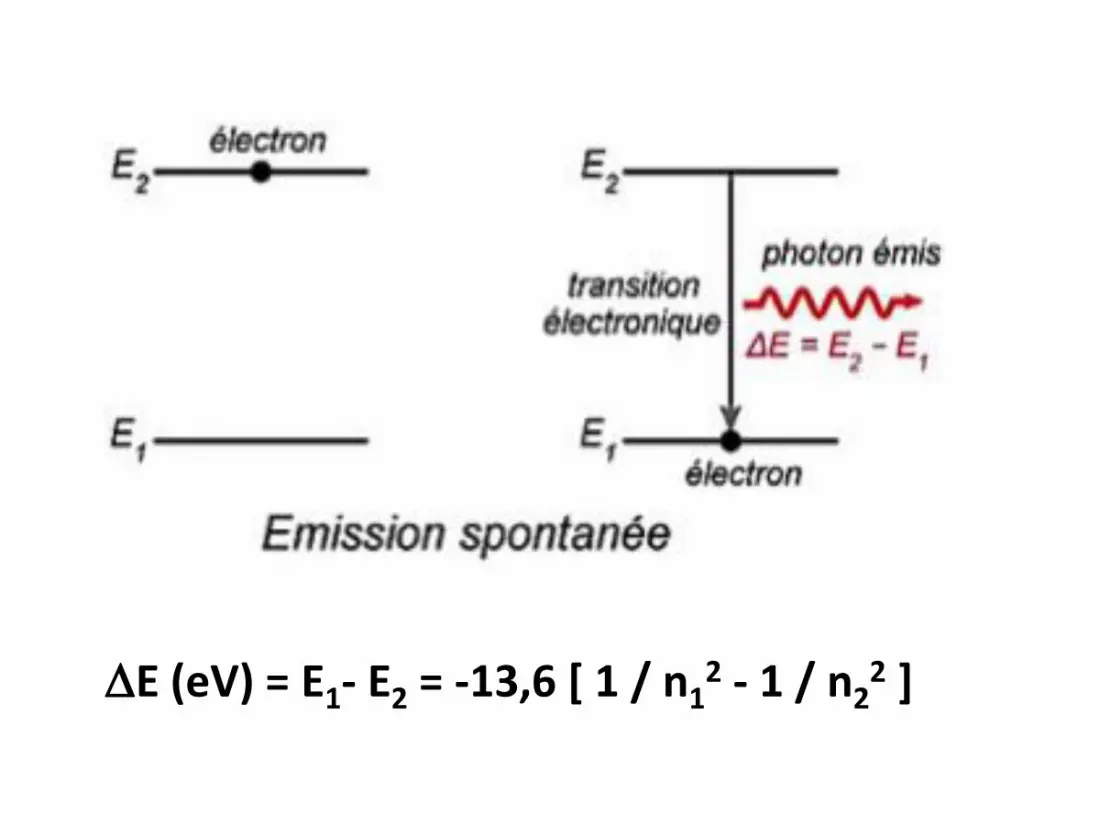
E (eV) = E1- E2 = -13,6 [ 1 / n12 - 1 / n2
2 ]

Les atomes n’existent que très rarement à l’état isolé.
Ils s’unissent généralement entre eux pour
former des molécules.
Les électrons des couches internes plus difficiles
à enlever que ceux des couches externes.
Processus chimiques courants impliquent la
redistribution des électrons des couches périphériques ou
électrons de valence.

Molécule
Les forces qui assurent la cohésion des atomes d’une
molécule et des ions d’un composé ionique solide sont
appelées liaisons chimiques. La plupart des propriétés
d’une substance dépendent de la nature des liaisons
chimiques. Les liaisons sont classées en fonction de
l’énergie qu’il faut mettre en œuvre pour les rompre, ce
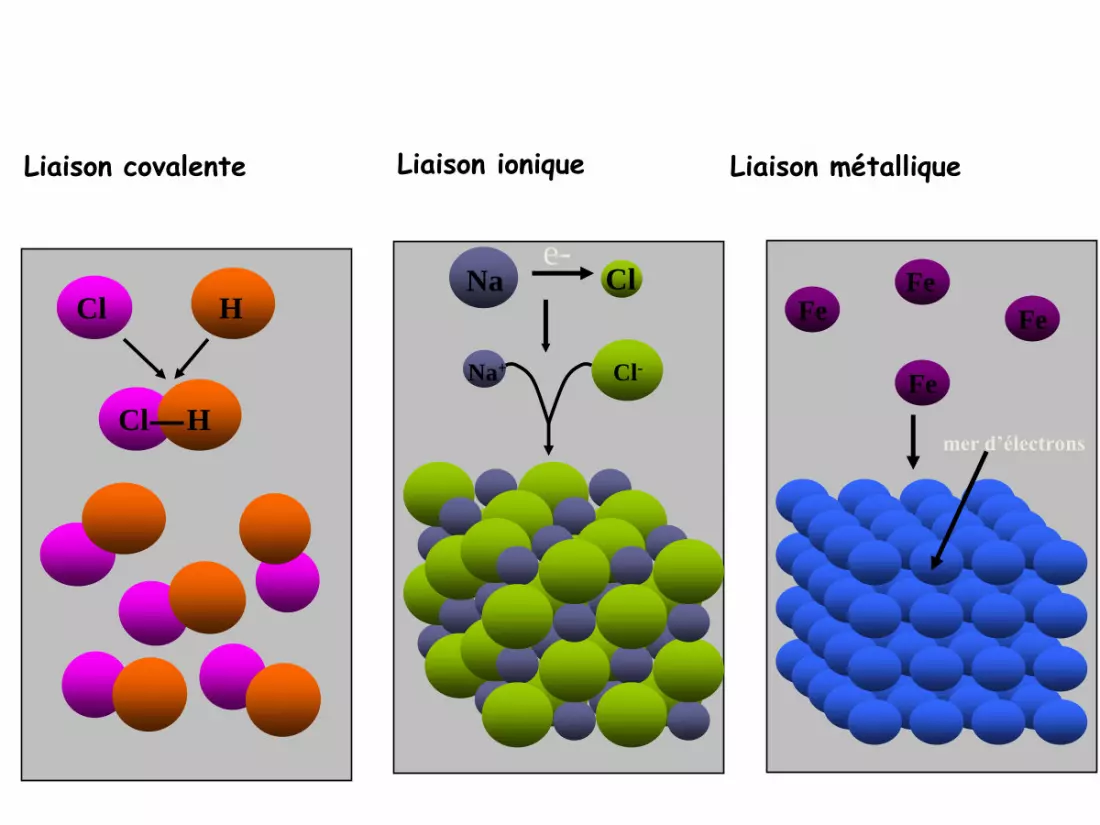
qui conduit à distinguer deux grands types de liaisons :

● Liaisons fortes
(énergie supérieure à 100 kJ.mol-1)
→ Liaison covalente→ Liaison ionique→ Liaison métallique
● Liaisons faibles(énergie de 0,2 à 50 kJ. mol-1)
→ Liaison de VAN DER WALS → Liaison hydrogène

La liaison Chimique forte peut être achevée:
- Si deux atomes partagent 2 électrons(chacun contribue par un): liaison covalente.
- Si deux atomes partagent 2 électrons(un contribue par 2 et l’autre par 0): liaison covalente de coordinence.
-Par un transfert complet d’un ou de plusieurs électrons: liaison ionique.
- Si tous les atomes partagent des électrons entre eux: liaison métallique.
e-Na
Na+ Cl-
ClCl H
Cl H
Fe Fe
mer d’électrons
Fe
Fe
Fe
Fe
Liaison covalente Liaison ionique Liaison métallique
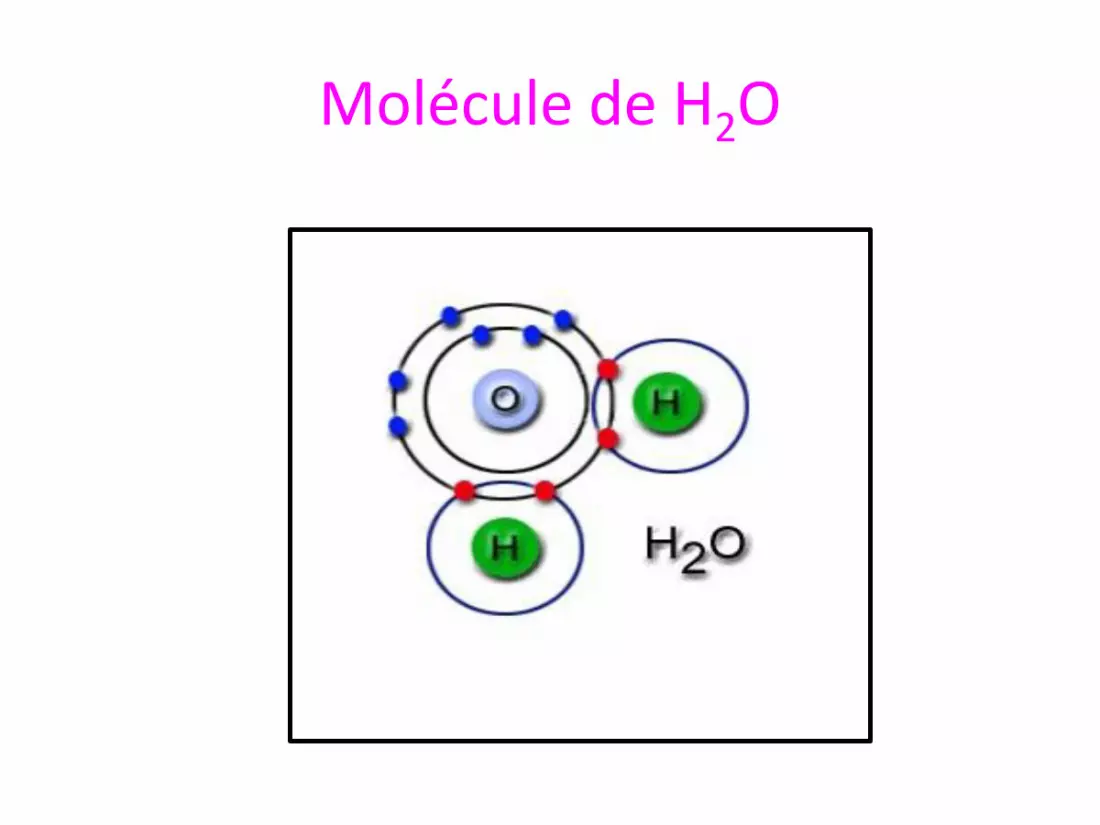
Molécule de H2O

H+ 0 électron
H+H3N +
H+
H3N H

Formation du chlorure de Sodium

Liaisons faibles
(énergie de 0,2 à 50 kJ. mol-1)

La liaison hydrogène
Il se forme chaque fois qu’un atome d’hydrogène est liée
à un atome très électronégatif (F, Cl, O,..)
La liaison est polarisée, l’atome d’hydrogène porte une
charge partielle positive + . Il peut donc y avoir une
interaction électronique entre cette charge positive et la
partie négative d’une autre molécule.
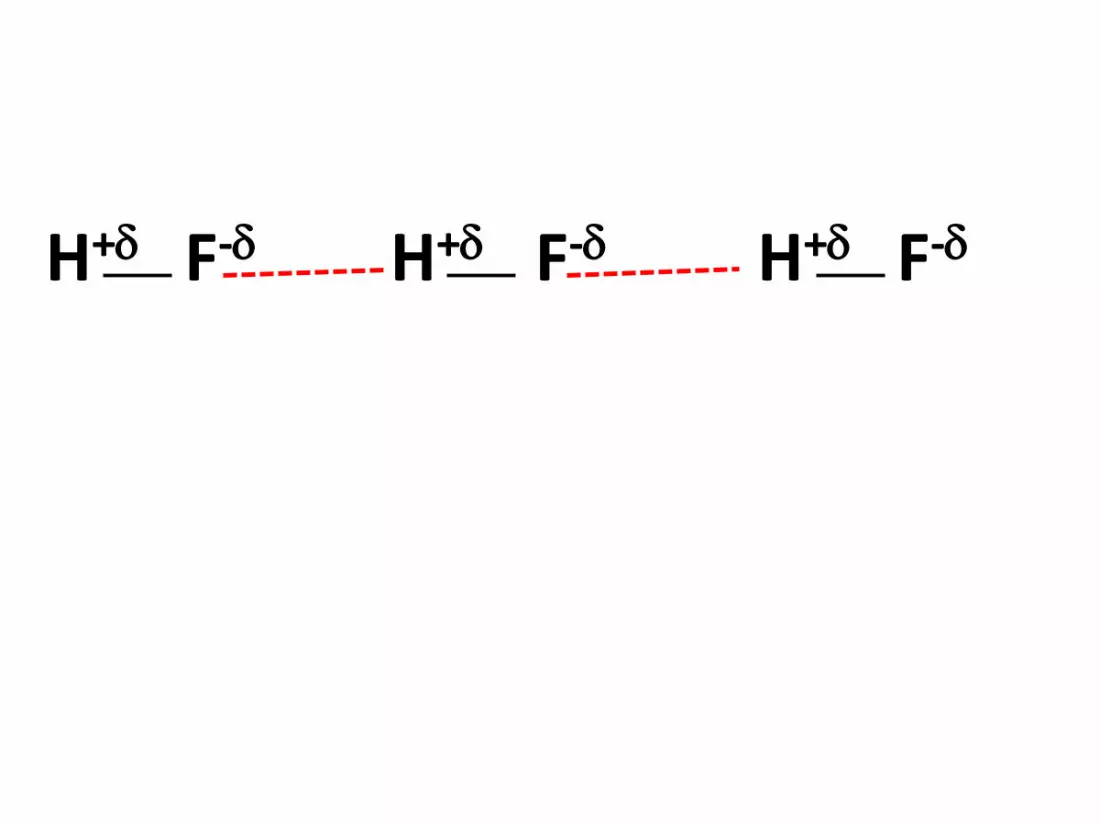
H+ F- H+ F- H+ F-



Liaison hydrogène:
Bien qu'énergétiquement plus faibles que les liaisons
covalentes, ces liaisons ont une importance
biologique. Leur effet cumulatif renforce la forme
tridimensionnelle des macromolécules et maintient
leur structure.
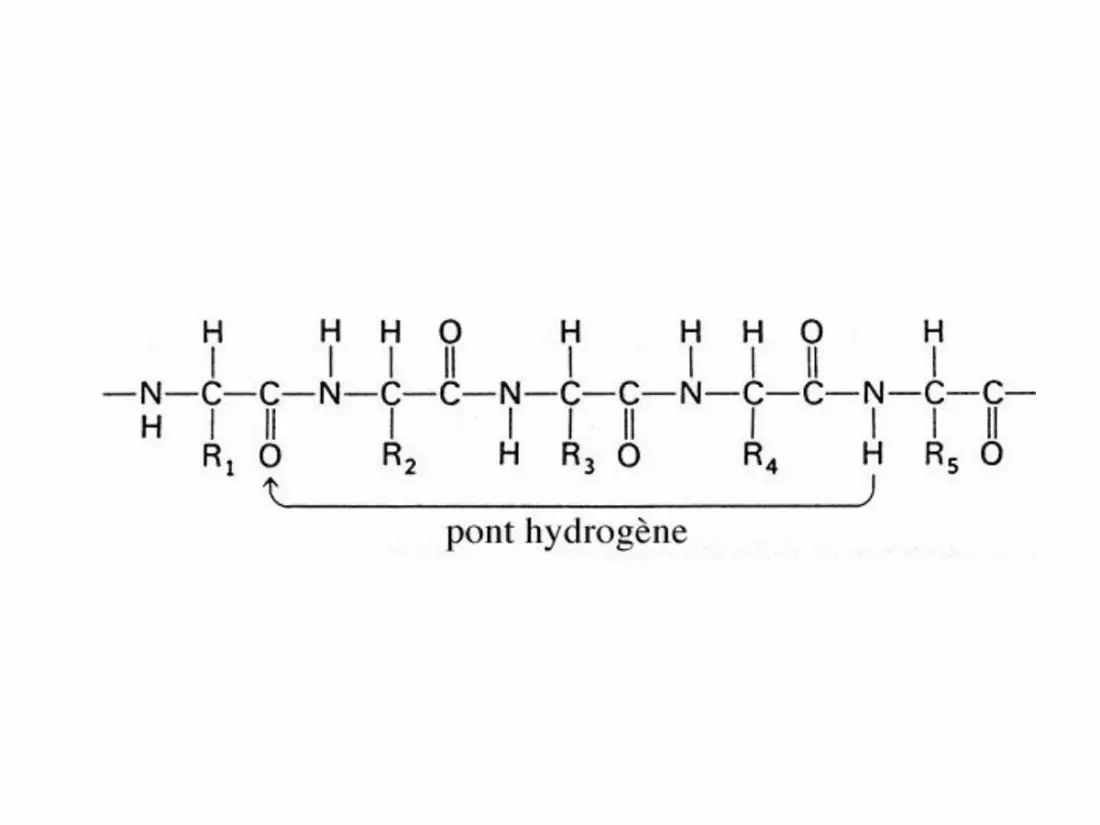

• Structure secondaire d'un polypeptide, montrant sa forme hélicoïdale et les ponts hydrogène (points
roses) qui en sont responsables (d'après
Miller, L., 1994.- Biology. Discovering Life. D. C. Heath
and Compagny).

Molécule d’ADN
Adénine – thymineCytosine - Guanine

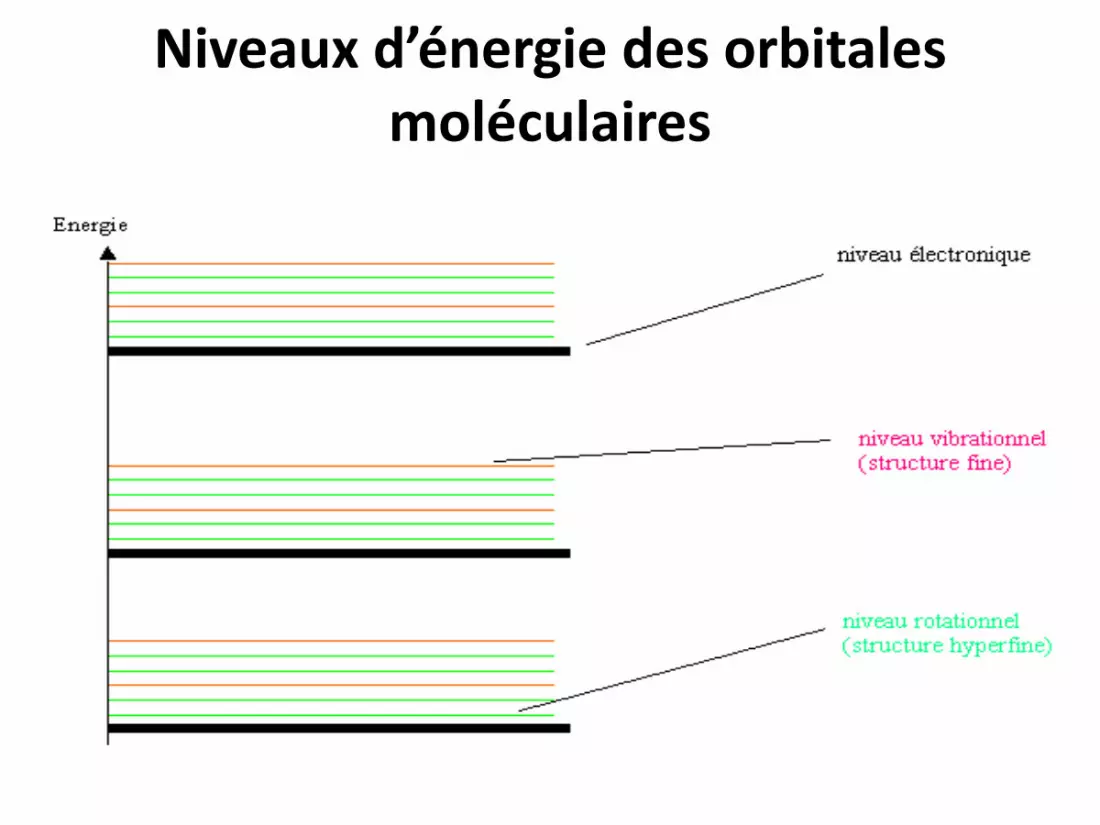
Niveaux d’énergie des orbitales moléculaires

Rayonnement
Le rayonnement est un transfert d'énergie sous
forme d'ondes ou de particules.



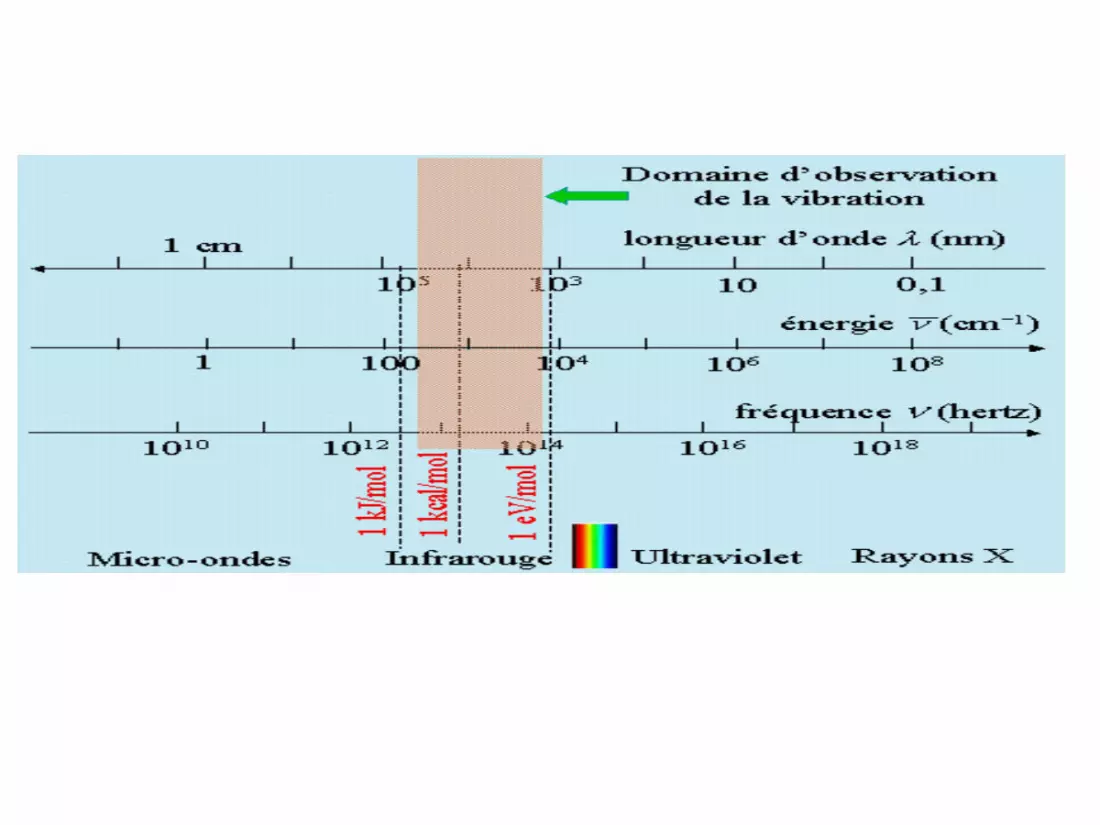
E = h ν = hc/
Les caractéristiques d’une onde électromagnétique :
Fréquence ν ( Hz) et période T(S) ν = 1 / T
Longueur d’onde m, µm, nm = c T = c / ν
Avec c vitesse de la lumière (célérité) c 3 108 m.s-1


Rayonnements de particules
• Rayonnement corpusculaire/particulaire:
Rayonnement formé essentiellement par
des particules neutres ou chargées.

Rayonnement de particules chargées :
Ces particules sont caractérisées par :
• Une charge
• Une masse au repos m0
• Une vitesse v
• Une masse cinétique, m= m0 / (1- v2/c2 )1/2
• Quantité de mouvement P= mV
• Une énergie totale E= mc2

L’énergie varie de 10keV à 3MeV.
Le spectre de β- est spectre continu avec une intensité maximale pour E = Emax/3
accompagné d’une émission γ si le radionucléide est dans un état excité.
Exemple : d’une réaction β- pour l’isotope radioactif cobalt 60(60Co) qui se transforme en nickel 60 (60Ni) stable :
6027Co 60
28Ni + e− + ν¯
Les rayonnements β-

Les rayonnements β+
Le rayonnement bêta plus est constitué d’un positon (particule de même masse que l’électron mais chargée positivement).
Par exemple, l’iode 122 est un radioactif bêta plus et se transforme en tellure 122. Notons que pour les deux types de désintégration bêta, le noyau garde le même nombre de nucléons (donc la même masse atomique).
eXX A
Z
A
Z
0
11

Les émetteurs sont des noyaux lourds, la plupart de
nombre atomique supérieur à 82. Si Y est produit à un
niveau excité, l’émission α s’accompagne de l’émission
d’un photon gamma (γ). Le spectre de ce rayonnement
est un spectre de raies dont l’énergie d’émission est
comprise entre 3 et 9 MeV
Les rayonnements alpha

Particules non chargées (neutron)
Interaction aléatoire
Ionisations indirectes par l’intermédiaire de particules
secondaires chargées mises en mouvement.

Relation énergie et longueur d’onde
• Chaque photon transporte un « quantum » d’énergie
proportionnel à la fréquence de l’onde.
• Relation de Planck :
• E = hv
• E = énergie en joules
• v= fréquence en hertz
• h = cte de Planck = 6,62 10-34 J.s = 4,136 10-15 eV s

= h/mV (dualité onde corpuscule)
les particules et la lumière sont des quanta
d'énergie dont le comportement est à la fois
corpusculaire et ondulatoire (dualité onde-
corpuscule).


Classification des rayonnements
rayonnements
non ionisants ionisants
électromagnétiquesh < 12.4 eV
> 0.1 m
électromagnétiquesh > 12.4 eV < 0.1 m
particules
non chargées chargées
légères lourdesUV - visible - IRmicro ondesondes radio photons X et neutrons + - p+
D II D I

Rayonnements ionisants
• Les rayonnements d’énergie suffisante sont capables
d’ioniser les atomes, c’est-à-dire d’arracher un électron à un
atome. De même, ils sont capables de couper des liaisons
intra-moléculaires (par exemple : ionisation de l’eau).
• Dans le cas des tissus biologiques, on considère que les
rayonnements d’énergie supérieure à 13,6 eV (énergie
d’ionisation de l’hydrogène ou de l’oxygène) sont ionisants.

Rayonnements non ionisants• Un rayonnement non-ionisant c’est rayonnement pour lequel l’énergie
électromagnétique transportée par chaque quantum est insuffisante
pour provoquer l’ionisation d'atomes ou de molécules.
• Ces radiations peuvent cependant avoir suffisamment d'énergie pour
provoquer le passage d'un électron sur un niveau d’énergie plus élevé.
Certains de ces rayonnements peuvent avoir des effets biologiques
(chapitre II).
• Parmi les rayonnements non-ionisants, on compte les rayonnements du
proche ultraviolet, la lumière visible, l'infrarouge, les micro-ondes.

Chapitre IIEffets des rayonnements non
ionisants sur la matière biologique

Pourquoi étudier l’action des rayonnements dans les
systèmes biologiques?
C’est indispensable pour maîtriser l’action des rayonnements
dans les systèmes biologiques, pour établir des normes quant
à l’utilisation des radiations et définir les risques
d’exposition.

Que se passe-t-il après l’impact d’un rayonnement sur une cellule?

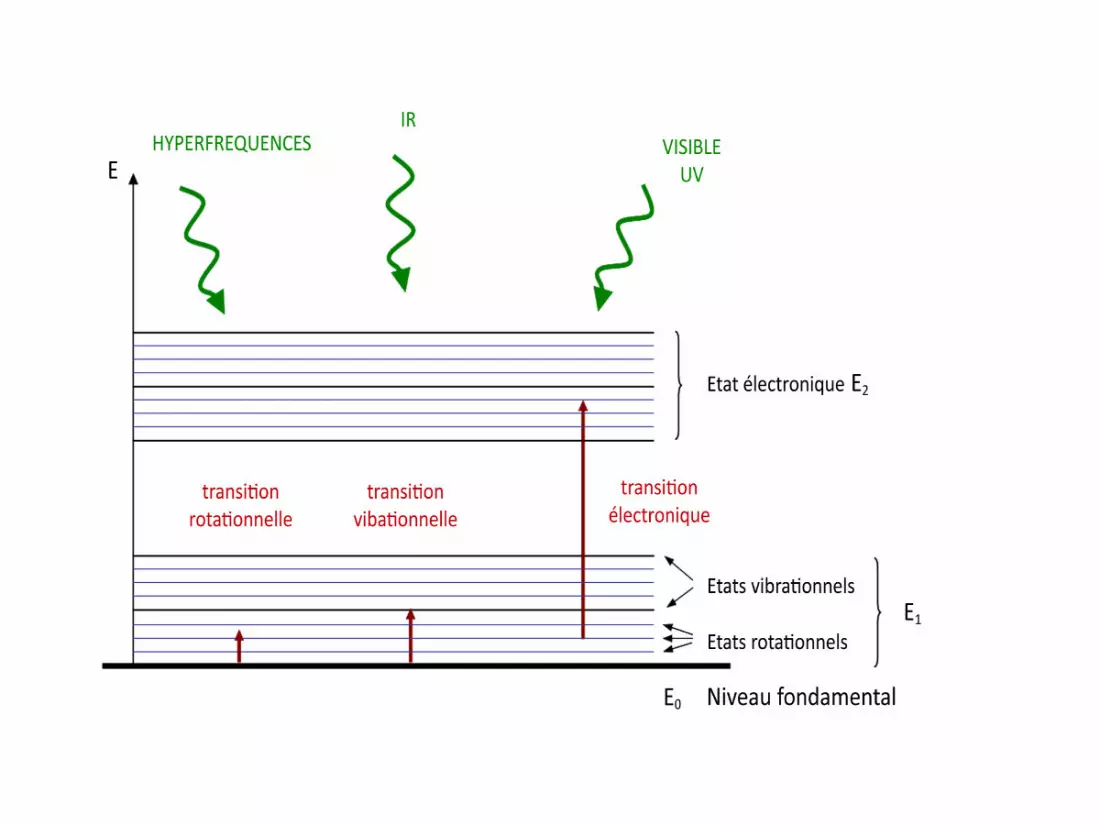

Dans l’ultraviolet et le Visible
L’énergie absorbée est suffisamment
importante pour permettre des transitions
énergétiques entre niveaux électroniques.
Au-delà d’un certain seuil énergétique,
l’absorption peut provoquer une dissociation
des molécules par rupture de liaison.


Seule les transitions électroniques
( UV et Visible) ont un intérêt photochimique
MBio M*Bio
h
Cette excitation implique une modification de
l’état énergétique moléculaire.
E M*Bio > E M Bio

Dans l’infrarouge proche :
• L’absorption du rayonnement est beaucoup moins énergétique que dans le visible ou les ultraviolets et les transitions d’énergie se font entre le niveau fondamental et les niveaux vibrationnels des molécules.
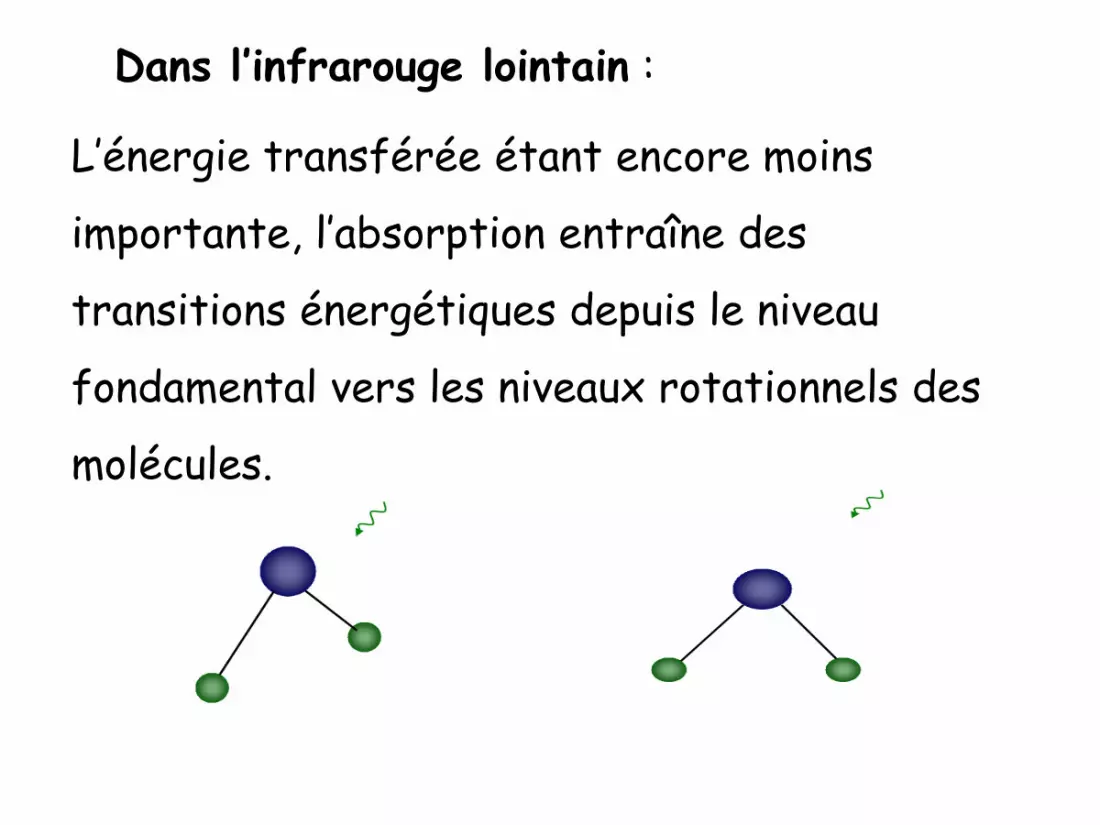
Dans l’infrarouge lointain :
L’énergie transférée étant encore moins
importante, l’absorption entraîne des
transitions énergétiques depuis le niveau
fondamental vers les niveaux rotationnels des
molécules.

Effets des rayonnements UV, Visible sur la matière biologique

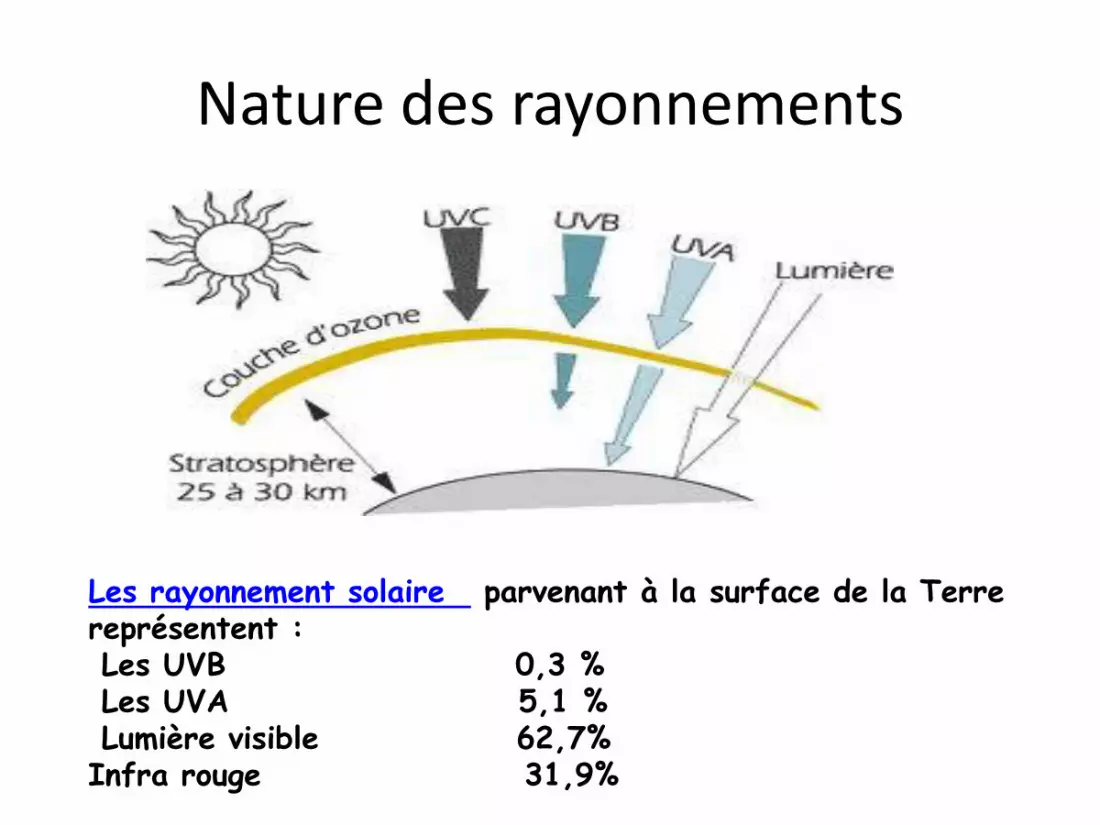
Nature des rayonnements
Les rayonnement solaire parvenant à la surface de la Terre représentent :Les UVB 0,3 %Les UVA 5,1 % Lumière visible 62,7% Infra rouge 31,9%

• La diminution de la couche d’ozone dans
certaines régions du globe fait que la
lumière solaire parvenant à la surface de la
terre a tendance à s’enrichir en
rayonnements UVB et UVC.

Les molécules biologiques, molécules de
la matière vivante susceptibles d’être
affectées par les radiations RNI, sont
les molécules organiques non saturées
(protéines, les acides nucléiques..).
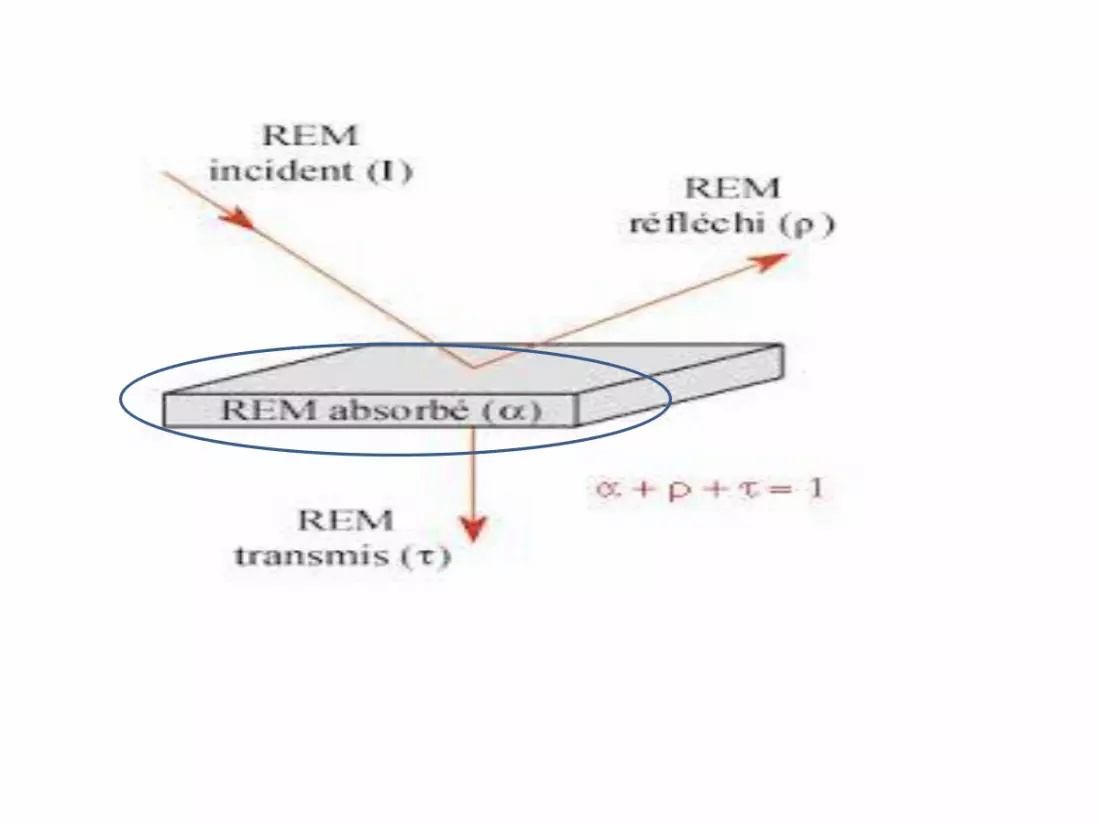

Après absorption d’un photon UV, l’énergie acquise
peut être dissipée par un certain nombre de
mécanismes :
• Transformation en chaleur
• Vibration des atomes constituant la molécule
• Transition électronique des molécules


Effets directs et indirects des rayonnements UV sur la matière
biologique

A poids égal, les acides nucléiques absorbent 10 à 20
fois plus de rayonnements que les protéines. La
molécule d’ADN peut alors être profondément
remaniée par une exposition aux rayonnements UV.

Caractériser et quantifier les agressions que subit l’ADN sous l’action des radiations (UV)
Excitation directe des molécules de l’ADN lui-même
Action sur les molécules produisant des espèces réactives qui attaquent l’ADN
Effets directs Effets indirects
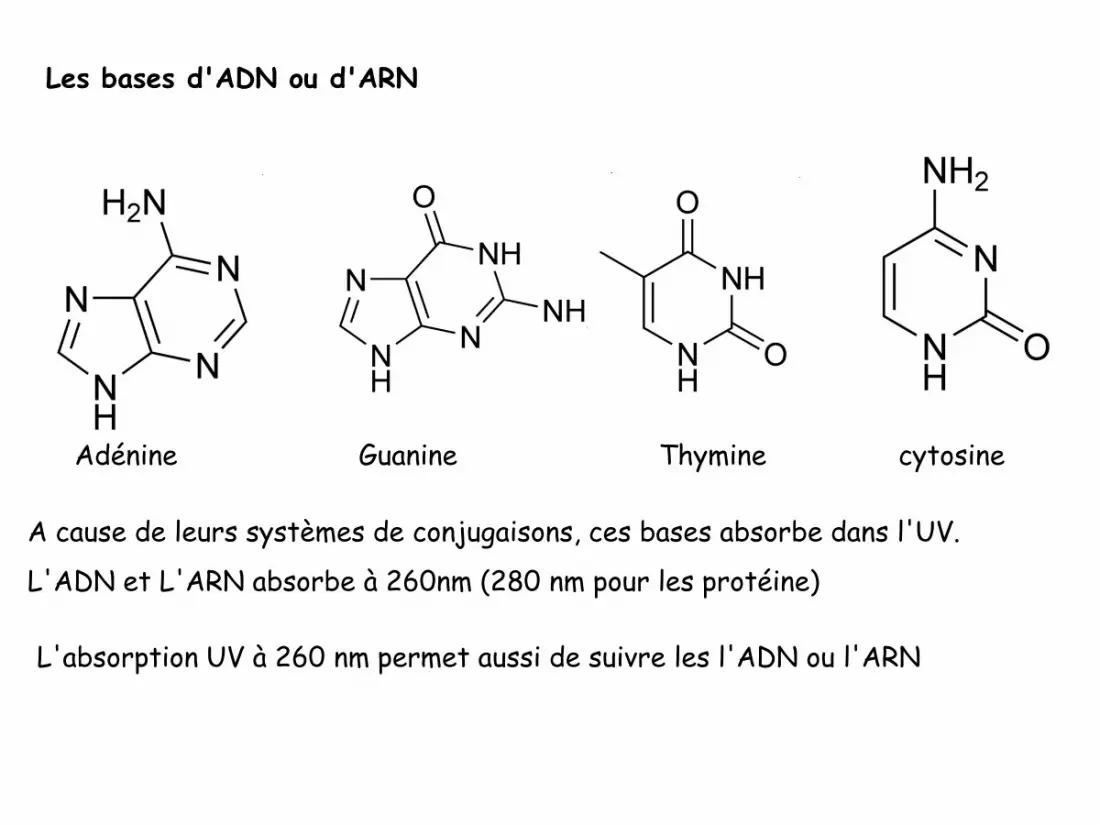
Les bases d'ADN ou d'ARN
Adénine Guanine Thymine cytosine
A cause de leurs systèmes de conjugaisons, ces bases absorbe dans l'UV.
L'ADN et L'ARN absorbe à 260nm (280 nm pour les protéine)
L'absorption UV à 260 nm permet aussi de suivre les l'ADN ou l'ARN

Effets directs des rayonnements UV sur ADN
• L’interaction des rayonnements UV avec
l’ADN conduisent à sa dénaturation (la
rupture des liaisons hydrogènes, donc
séparer les 2 brins d'ADN, la rupture
de liaisons phosphodiester).



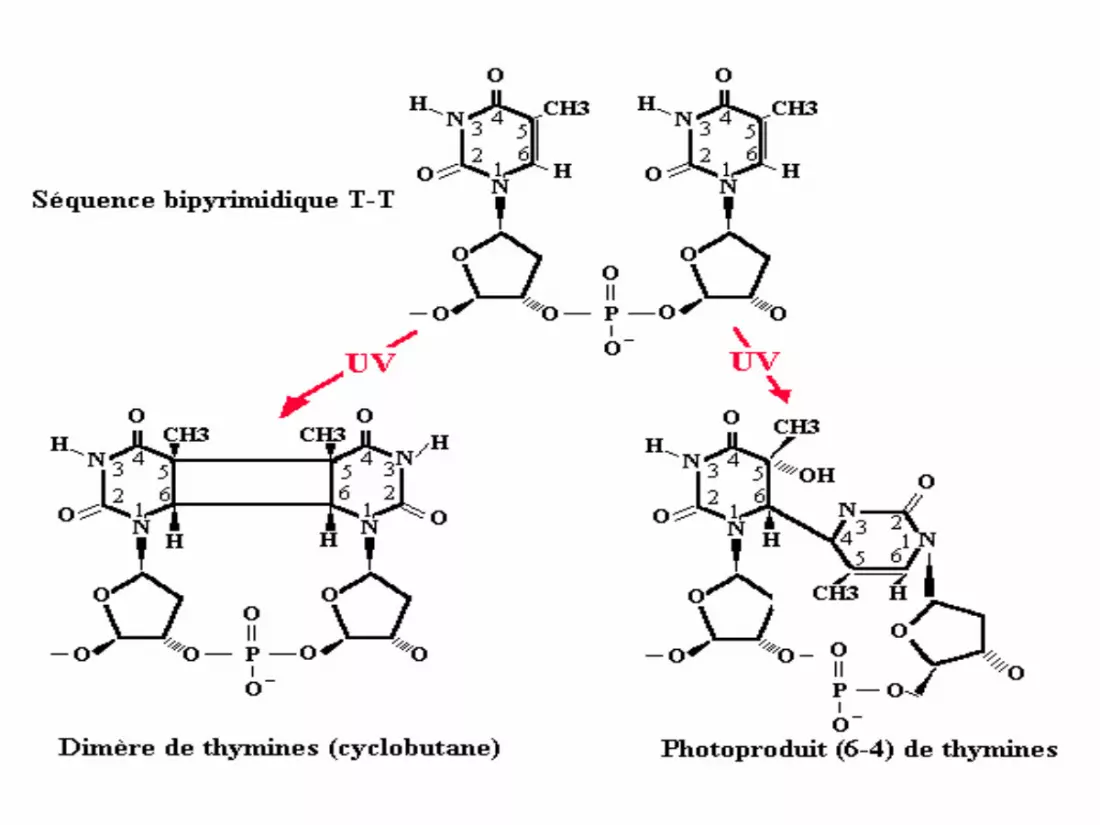
L'énergie absorbée induit le plus souvent la formation
d'une liaison covalente entre deux pyrimidines adjacentes
sur un même brin formant un dimère de pyrimidine (DP).
On peut avoir 2 liaisons covalentes entre les C5 et les C6
de deux pyrimidines adjacentes. Cette dimérisation peut
concerner les quatre types de séquences bipyrimidiques
(TT,TC,CT,CC).
Les CPD (les dimères cyclobutaniques de pyrimidines)

L'absorption de l'énergie des UV par l'ADN
entraîne la formation de dimères de thymines
adjacentes, ce qui provoque la distorsion de la
double hélice.


• Une double liaison covalente empêche l'ADN
polymérase d'effectuer la réplication de la
cellule. Généralement la mort cellulaire ou le
cancer s'en suivent lorsque non réparées par
les enzymes.

• La formation des CPD (dimères cyclobutaniques de
pyrimidines) dépend de la longueur d’onde du rayonnement
UV (UVB ou UVC), du matériel biologique étudié (ADN
cellulaire, plasmide, etc.), du contexte de séquence autour
du site bipyrimidique mais également du site bipyrimidique
lui‐même.

Les photoproduits (6‐4) (pyrimidines (6‐4) et pyrimidone(6‐4 PP))
• Les 6‐4 PP sont quantitativement les secondes lésions les
plus fréquentes après une irradiation UVB ou UVC. Les
6‐4 PP résultent de la formation d’un pont stable entre
les positions C6 et C4 de deux pyrimidines adjacentes.
Ces appellations faisant référence à la nature des liaisons
formées pour créer le dommage


Absorption indirecte par (UVA et des UVB faible énergie)
• Les UVA (320 nm à 400 nm),
représentent la partie la plus importante
des UV, peuvent également induire des
dommages sur l’ADN de façon indirecte.

• En effet, les UVA ne sont que très
faiblement absorbés par les bases de
l’ADN, mais ils peuvent exciter certains
composés endogènes jouant un rôle de
photo- sensibilisateurs (chromophores).

Action sur les molécules produisant des espèces réactives qui attaquent l’ADN
Effets indirects

S0 état fondamental
S1 état excité
T1 état intermédiaire
E

Cet état intermédiaire joue un rôle clé dans l’induction des
dommages liés aux UVA. Il peut soit :
• Réagir Directement avec d’autres molécules, comme les
bases de l’ADN, (photosensibilisation de type I).
• Transférer son énergie aux molécules oxygène
(photosensibilisation de type II), menant ainsi à la formation
d’espèces réactives de l’oxygène (reactive oxygen species,
ROS) comme l’oxygène singulet (1O2) ou l’anion superoxyde
(O2•- ).
Remarque:
ERO (espèces réactives oxygénées) = ROS (reactive oxygen species)

Ces espèces réactives oxygénées entrainent la
Formation de lésions oxydatives.
Elles correspondent à des oxydations de bases
provoquées par des agents super-oxyde
(°O2-, H2O2 et °OH).

La formation des radicaux libres rend les molécules (ou les
atomes) instables. Ces molécules essayent alors sans cesse de
capter ou céder un électron à une autre molécule de leur
entourage, propageant ainsi le phénomène, en transformant à
leur tour les autres molécules en radicaux libres. Lorsqu’elle
se produit dans l’organisme, cette réaction en chaîne est
communément appelée stress oxydant.

• Les principales cibles sont les lipides, constituants essentiels
des membranes externes et internes de la cellule.
Une fois oxydées, les membranes cellulaires deviennent
rigides et ne sont plus capables de s'adapter aux variations de
leur environnement.
• Cependant, l’ADN est également touché par ces radicaux
libres qui lèsent le matériel génétique.

Conclusion
• L’accumulation de ROS dans la cellule
peut provoquer des lésions directes sur
tous les composants cellulaires :
peroxydation des lipides , oxydation
des protéines et altérations des acides
nucléiques.

Sur l’ADN, cinq principaux types de lésions peuvent être induits par les ROS
• la modification de bases,
• la création de sites abasiques,
• la formation d’adduits lipidiques,
• les pontages ADN‐ protéines,
• les cassures de l’hélice d’ADN (simple ou double‐brin).
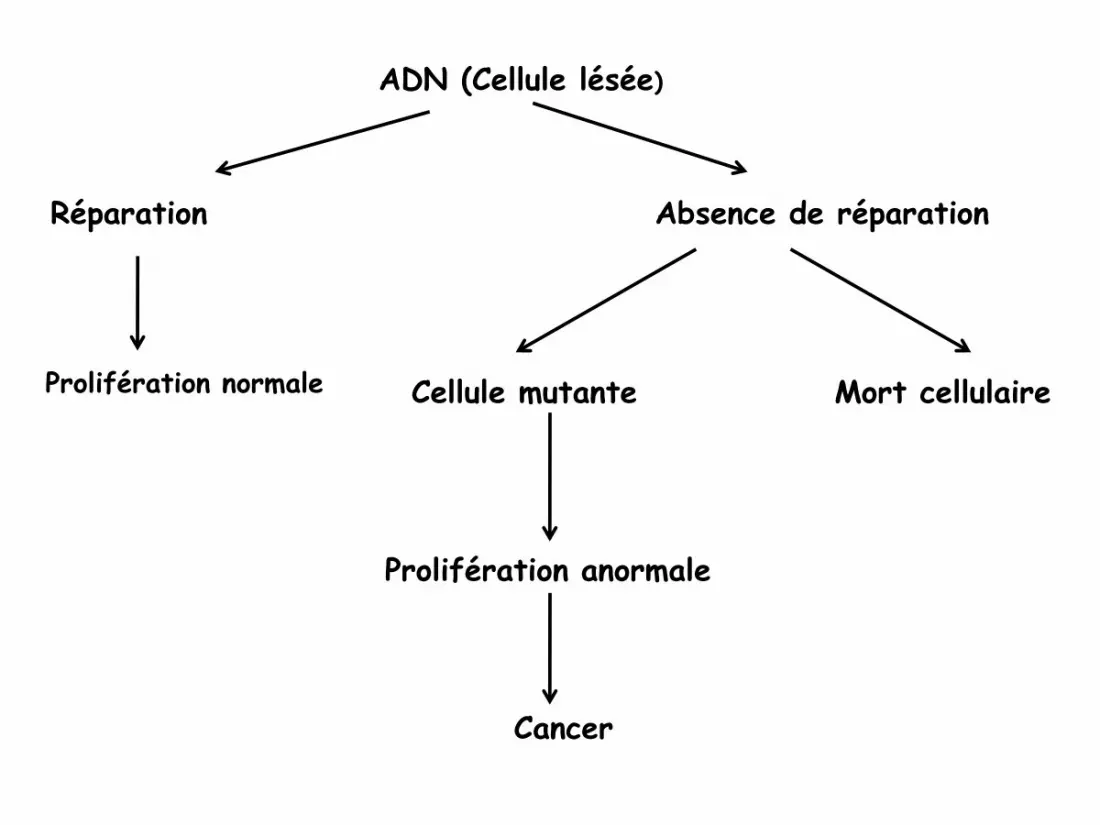
ADN (Cellule lésée)
Réparation Absence de réparation
Cellule mutante Mort cellulaire
Prolifération anormale
Cancer
Prolifération normale

Effets sur les yeux
Les yeux sont particulièrement sensibles aux
rayons UV. Même une exposition de quelques
secondes peut provoquer des affections
temporaires connues sous le nom de
photokératite et conjonctivite. La
photokératite constitue une affection
douloureuse causée par l'inflammation de la
cornée.

Spectrométrie UV- Visible
Analyse qualitative
• Les groupes d’atomes qui absorbent sont
appelés des groupes chromophores et ceux
qui n’absorbent pas mais qui provoquent
seulement des modifications de
l’absorption par un chromophore sont dits
auxochromes.

• Quand un chromophore est soumis à des influences
électroniques, la bande d’absorption peut se déplacer
vers les grandes longueurs d’onde, c’est l’effet
batochrome, ou vers les faibles longueurs d’onde, c’est
l’effet hypsochrome.
• Si l’absorption lumineuse est augmentée, on dit qu’il y a
un effet hyperchrome. Si elle est diminuée, il y a un
effet hypochrome.

Chromophores max max
Chromophores conjugués
>C=C-C=C< (linéaire) 220 30000
>C=C-C=O (dans un cycle) 244 10000
Acides nucléiques
Adénosine 260 15000
Guanosine 255 14000
Thymidine 265 10000
Cytidine 270 9000
ADN double brin 260
ADN simple brin 260
ARN 260
Protéines
Liaison peptidique 190 4-8000
Pont disulfure 250 300
Phénylalanine 257 200
Tyrosine 275 1400
Tryptophane 280 5600
Longueur d’onde de quelques groupes chromophores
E

Le spectrophotomètre UV-Visible
C'est un des appareil les plus utilisé en laboratoire

L'absorption: la loi de Beer Lambert
.C.L ε(λ)-eIo
It

La Densité optique (D.O) ou l'aborsbance A est
.C.L ε(λ))It
Ilog()log( 0 TDOA
L'intensité transmise décroit de façon exponentiel en
fonction de la largeur de la cuve et de la concentration.
C'est la loi de Beer-Lambert
ε est l'absorptivité molaire (aussi appelé coefficient d'extinction
molaire), exprimée en L·mol−1·cm−1. Elle dépend de la longueur d'onde, la
nature chimique de l'entité et la température.
L est la longueur du trajet optique dans la solution traversée, elle
correspond à l'épaisseur de la cuvette utilisée (en cm).
C est la concentration molaire de la solution (en mol.L−1).

Application
• Soit une solution de carotène dans le
chromophore à 5mg/l . Calculer le coefficient
d’absorption molaire, sachant que l= 1 cm et la
T= 7,5% M= 536g/mol

Solution
• A = DO = l C
• DO = log 100/7,5 = 1,124
• exprimée en L·mol−1·cm−1.• C (g/l) C (mol/l)
• C (concentration molaire) = 5. 10-3/ 536
• = A/l.C = 1,124 / 1 . 9,3
• = 1,2. 10+5 (mol/l) -1cm-1

Application 2
• La transmission d’une solution est de
20% dans une cuve de 1cm. Quelle serait
la transmission de cette solution dans
une cuve de 5cm.

• DO1 = log I0/ I1 = log 1/ T1 = l1 C
• DO2 = log I0/ I2 = log 1/ T2 = l2 C
• ( log 1/ T1 ) / (log 1/ T2) = 1 / 5
• (log 1/ T2) = ( log 1/ T1 ) . 5 = 3,28
• 1/ T2 = 103,28 T2 = 1/ 103,28

L'absorption de la lumière est dépendante de:
• la longueur d'onde (transition électronique)
• de la nature chromophore
• la concentration (ou densité), C
• la longueur du chemin optique parcouru dx

Exemple d’un spectre UV
ADNProtéine (sérum albumine)

-A260 est le maximum d'absorption de l'ADN. Dans certaines
limites, cette absorbance est proportionnelle à la
concentration en ADN.
- l'ADN absorbe environ 2 fois moins à 280 nm qu'à 260 nm.
- A280 est le maximum d'absorption des protéines. Dans
certaines limites, cette absorbance est proportionnelle à la
concentration en protéine(s).

Quand on réalise un extrait cellulaire d'ADN, il est
accompagné par des protéines dont la présence peut
être indésirable.
La valeur du rapport A260/A280 fournit une bonne
indication sur la pureté de l'échantillon. En effet, la
présence de protéines fait augmenter A280.
Si le rapport, mesuré par spectroscopie est:
1.8 < A260/A280 < 2 ADN extrait est pur
A260/A280 < 1.8 ADN contaminé par des protéines
A260/A280 > 2 Une présence importante d’ARN

Le rapport A260/A280 pour l'échantillon d'ADN étudié,
déterminé d'après les spectres ci-dessus est égal à
1,4/0,8 soit 1,75. L'ADN est faiblement contaminé par
des protéines.

Le dosage d'une solution d'ADN par spectroscopie à 260nm indique
une absorbance (DO) de 0,78. Quelle est la quantité d'ADN
contenue dans 15µl de cette solution?
Rappel : ADN : 1DO -> 50ng/µl
ADN dénaturé : 1DO -> 37ng/µl
Exercice 2
Solution 2
DO : 1 => 50ng/µl0.78 => X
X = 0.78 * 50 = 39ng/µl
39ng/µl * 15 = 585ng

Méthodes d’analyse
• La substance à doser possède un pic d'absorption caractéristique

Vérifier la nature du spectreOn repère l'axe des abscisses et on vérifie le domaine dans lequel on travail.
Déterminer la position du pic d'absorbance.On repère sur l'axe des abscisses la longueur d'onde pour laquelle A est maximum.
Déterminer la couleur absorbéeOn utiliser ses connaissances ou on lit l'étoile des couleurs complémentaires, si elle est donnée, pour savoir quelle est la couleur correspondante.
Conclure sur la couleur du milieu étudiéOn repère sur l'étoile la couleur opposée à celle trouvée ci- dessus: il s'agit de la couleur du milieu étudié.On peut donc, uniquement à partir du spectre, retrouver la couleur du milieu étudié.

Exercice 1
On donne le spectre de deux espèces M et P. Déterminer les
coordonnées du maximum d’absorption de chacune d’elles. Quelle est
la couleur de l’espèce M en solution ? Peut-on doser l’espèce M dans
une solution contenant P ? Et inversement ?

Graphiquement, λmax(M) = 425 nm et λmax(P) = 630 nm.
La solution de M absorbe dans le violet, elle est jaune vert (couleur
complémentaire).
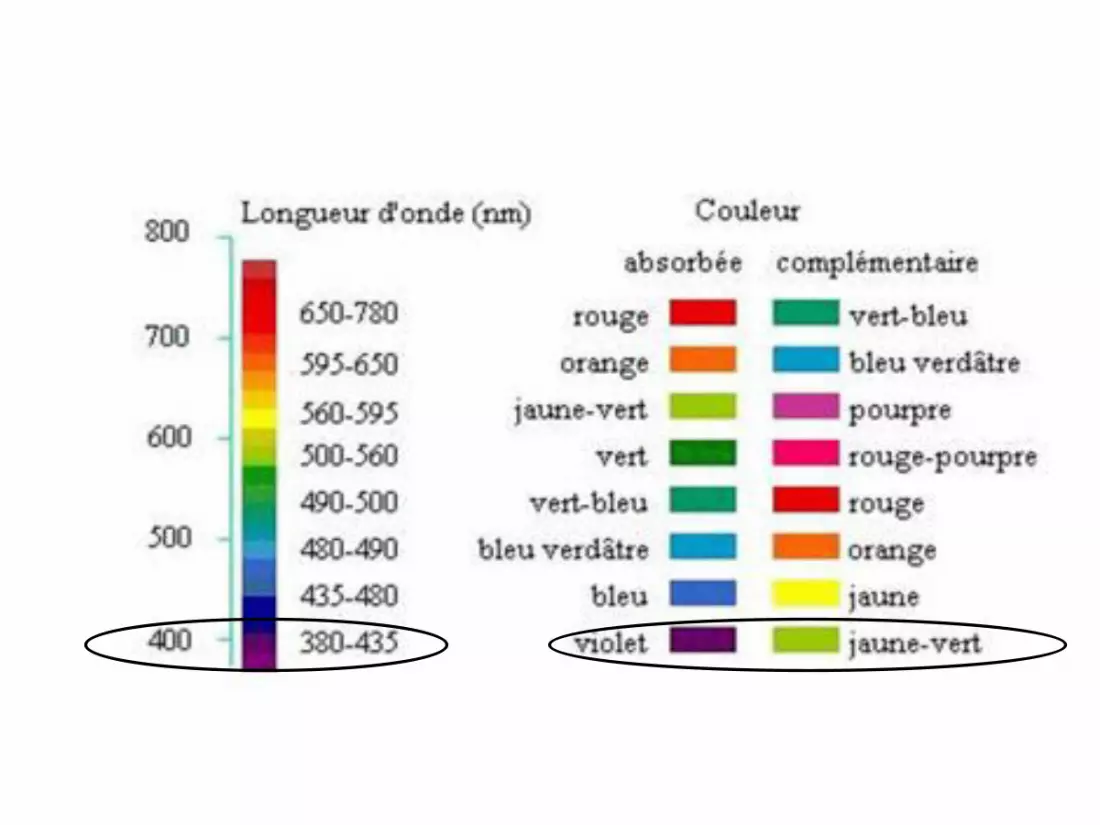

Si on se place à 425 nm, A = A(M) + A(P) car l’espèce P
absorbe aussi à 425 nm. On ne peut doser M à 425 nm en
présence de P.
À 630 nm, seule P absorbe, on peut réaliser une étude
quantitative pour P.
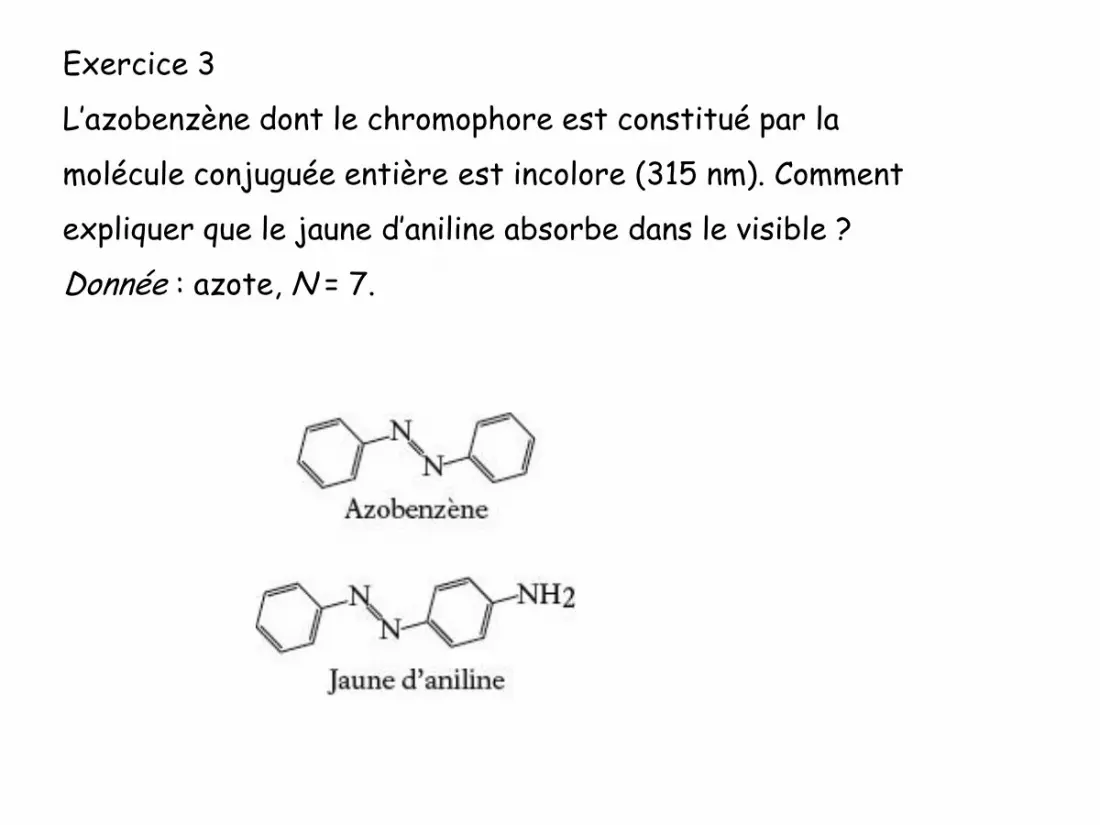
Exercice 3
L’azobenzène dont le chromophore est constitué par la
molécule conjuguée entière est incolore (315 nm). Comment
expliquer que le jaune d’aniline absorbe dans le visible ?
Donnée : azote, N = 7.

Le jaune d’aniline possède le même chromophore
mais avec un substituant en plus, le groupe amino -
NH2. L’azote N de structure (K)2(L)5 engendre
trois liaisons. Il possède donc un doublet non liant
qui participe à la conjugaison. Ainsi, on observe un
déplacement du maximum d’absorption vers des
longueurs d’onde plus élevées (effet bathochrome)
qui se trouve dans le visible. La molécule est donc
colorée.

2) La substance à doser ne possède pas de pic d'absorption caractéristique; il faut alors effectuer une réaction colorée; on fait un dosage indirect. Il y a deux méthodes

Méthode par comparaison avec un étalon unique:
Elle consiste à mesurer dans les mêmes conditions
l'absorbance Ad de la solution à doser et l'absorbance
Aet d'une solution "étalon" ou "standard" de
concentration connue Cet, puis calculer la
concentration de la solution à doser Cd.
Cd = Ad * Cet / Aet

Méthode avec une gamme d'étalonnage :
Elle consiste à préparer une gamme de dilutions
d'une solution étalon "mère", à mesurer
l'absorbance de chacune de ces solutions étalons
"filles", puis à tracer la courbe d'étalonnage
A = f(c)

On utilise trois types de solution :- une solution de concentration connue d'une protéine= référence ou standard par rapport à la protéine à doser.
-une solution de la protéine à doserdont on veut déterminer la concentration = échantillon à doser (essai)
-une solution de réactifqui développe une coloration en réagissant avec des acides aminés spécifiques de ces protéines.
. Principe d'une gamme étalon et d'une droite d’étalonnage


On laisse réagir dans les conditions et le temps nécessaires puis on mesure l'absorbance de tous les tubes.
A partir des tubes de la gamme étalon on trace une courbe d’étalonnage : absorbance = f (quantité protéine par tube) ou f (concentration en protéines par tube)

Cette proportionnalité permet de déterminer la quantité de protéine contenue dans un volume de prise d'essai de l'échantillon à doser.
sans oublier les facteurs de dilution

Effets des rayonnements IR sur la biomolécule
• L’absorption par les tissus biologiques des
photons infrarouges modifie l’état de
vibration ou de rotation moléculaire.

En raison de leur faible énergie, les photons infrarouges
ne peuvent pas produire d’ionisation, pas plus que de
réaction photochimique. L’irradiation infrarouge est
globalement perçue comme une augmentation de la
chaleur. Effets uniquement thermiques

Effets délétères
• Seul l’IR proche parvient jusqu’à la rétine (lésion rétinienne par effet
thermique par focalisation sur la rétine en manipulant des lasers
émettant dans l’infrarouge)
• Les effets cutanés peuvent exister et être responsables de brûlures.
Heureusement, la sensation d’excès de chaleur ou de douleur est un
signal suffisant pour éviter les lésions. Les mécanismes de compensation
sont bien connus, comme la vasodilatation et la sudation.
• Les effets provoqués par IR moyen et lointain : atteinte de la
thermorégulation, « coup de chaleur »


Analyse par fluorescence

• Certains composés organiques non saturés
(composés chromophores : protéines, acides
nucléiques) où il y’a perte d’énergie, une partie
de celle-ci est émise sous forme d’une radiation
lumineuse pour permettre le retour de la
molécule à son niveau d’énergie initiale
(phénomène de fluorescence). C’est ainsi que
plusieurs composés organiques irradiés par une
lumière UV fluorescent dans le domaine visible.

Il existe deux types de fluorescence.
• la fluorescence dite « naturelle », ou auto-fluorescence, émise spontanément par la cellule. On peut citer par exemple la chlorophylle des cellules végétales.
• la fluorescence conférée par un fluorochrome ou fluorophore, substance chimique qui émet de la lumière si elle est excitée à une certaine longueur d’onde.

• Ces fluorochromes sont des substances composées de plusieurs noyaux aromatiquesconjugués ou encore des molécules planes et cycliques qui possèdent une ou plusieurs liaisons π.

Fluorophoreλ excitation
(nm)
λ émission
(nm)
Hoechst 33342 343 483
DAPI 345 455
Hoechst 33258 345 478
SYTOX Blue 431 480
Chromomycin A3 445 575
Mithramycin 445 575
YOYO-1 491 509
Sur le tableau ci-dessous nous donnons quelques fluophores pour marquer l’acide nucléique.


• Retour de l’électron du niveau excité S1 au niveau électronique fondamental est réalisé avec émission d'un photon Fluorescent. Ce retour est très rapide de l'ordre de 10-6s et se produit, pour la majeure partie, à une longueur d'onde plus grande que la longueur d'onde absorbée.
• E photon d'excitation > E photon émis
• λ excitation < λ émission



Spectre d’émission :
• Pour obtenir un spectre d'émission, on fixe une λ d'excitation (choisie
grâce à la littérature ou vers un maximum d'absorption). On balaye les λ
d'émission en mesurant l'intensité de fluorescence. On obtient un
spectre d'émission.
Spectre d'excitation :
• L’excitation est caractérisée par une longueur d’onde λexc, spécifique
du composé étudié.
Pour obtenir un spectre d'excitation, on fixe une λ d'émission (choisie
grâce à la littérature ou vers un maximum d'émission). Ensuite on balaye
les λ d'excitation en mesurant l'intensité de fluorescence. On obtient
un spectre d'excitation.

un spectre d'excitation et un spectre d'émission pour le
fluorochrome "TexasRed", un marqueur de fluorescence excitable
dans le visible utilisé en cytométrie de flux.

• le spectre de fluorescence (situé aux longueurs
d’onde plus élevées) est le reflet approximatif
du spectre d’absorption (situé aux longueurs
d’onde plus basses).
• La différence d’énergie ou de longueur d’onde
(Eex - Eem) est appelée déplacement de
stockes.

La fluorimétrie et l’analyse quantitative
On définit alors le rendement quantique de
fluorescence, qui est constant, comme le nombre de
photons émis à une longueur d'onde sur le nombre de
photons absorbés par la molécule.
F = IF / IA

• Nous connaissons le nombre de photons absorbés qui est proportionnel à IA. Le nombre de photons émis est alors de :
IF = F.IA
IF = F (I0 - It) avec It = I0e-A
• Soit
• IF = F I0 (1 - e- l C )
Dans le cas d'une solution diluée, un développement limité au premier ordre donne.
IF = F I0 . l C

Fluorimétrie et analyse qualitative
La fluorescence induite Les colorants utilisés, ne deviennent fluorescents que lorsqu’ils sont en contact avec la molécule étudiée. On utilise souvent des microscopes optiques pour visualiser les acides nucléiques (ADN, ARN). On distingue 2 types de colorants :
*Les colorants se fixant spécifiquement sur les paires de bases (Hoechst et DAPI spécifiques des bases A-T).
*Les intercalants : iodure de propidium et bromure d’éthidium (se fixent entre 2 brins d’ADN peu importe la séquence nucléotidique).

Ces colorants permettent de distinguer:
• l’hétérochromatine (ADN très condensé donc fluorescence très forte),
• l’euchromatine (ADN peu condensé donc fluorescence plus faible).
• L’ADN n’est pas réparti de manière homogène dans le noyau donc on a une coloration hétérogène avec ces colorants.

• L’immunofluorescence indirecte
On utilise des anticorps fluorescents pour marquer les
molécules à étudier et les visualiser en microscopie
optique, cette technique permet d’éviter les
modifications génétiques de la cellule. On peut rendre
artificiellement les anticorps fluorescents en les
couplant chimiquement à des fluorochromes.

La protéine joue le rôle de l’antigène. Sur
cette protéine il y a plusieurs épitopes (zones
que les anticorps reconnaissent). Pour plus de
précision, on va utiliser des anticorps
monoclonaux (mélange d’anticorps qui ne
reconnaissent qu’un seul épitope).

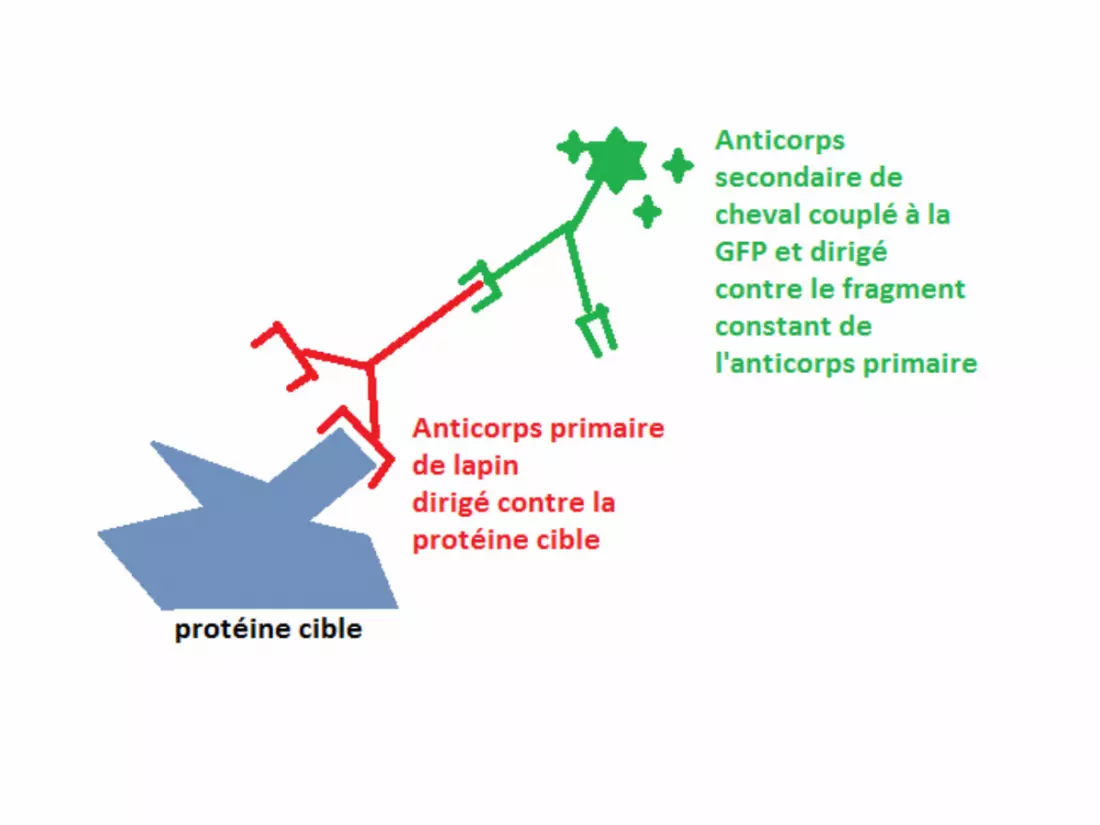
Pour la technique d’immunofluorescence indirecte,
on a besoin de 2 types d’anticorps :
• Les anticorps primaires, dirigés contre la
protéine d’intérêt.
• Les anticorps secondaires, dirigés contre
l’anticorps primaire.

Comment obtient-on ces anticorps ?
• On injecte à un animal l’antigène (la protéine d’intérêt).
L’animal va créer des anticorps pour se défendre et on ira les
récupérer.
• Les anticorps secondaires, dirigés contre l’anticorps primaire
(ils reconnaissent la partie constante des anticorps primaires)
et couplés à des fluorochromes, ils permettent de localiser
les anticorps primaires et donc les protéines d’intérêt.

Structure de la GFP.
GFP (Green Fluorescent Protein) est une protéine
naturellement fluorescente (Issue d'une méduse). Cette
protéine a la forme d’un tonneau avec au milieu 3 acides
aminés responsables de la fluorescence : le chromophore.

Solution
• on prélève 10 ml de ces ampoules que l’onintroduit dans une fiole jaugée de 100ml, on complete au trait de jauge à l’aide de l’eau et on homogéinise
• 10 ml de ces ampoules 2 ampoules
Exercice 3

10 ml des ampoules
Dilution
CAVA = CBVB
CA = CBVB/VA
VA= 10mlVB= 100mlCB = ?

• La densité optique de cette solution préparéeest égale à 0,45 à 380 nm dans une cuve de 1cm.
• CB ? • A = DO = l CB
• CB= 0,45/ 450 = 10-3g/100ml = 10-5g/ml
• CA = CBVB/VA = 10-5 100/10 = 10-4 g/ml
• Dans une ampoule on a 5ml10-4 g 1mlX 5ml
• La quantité du principe actif dans une ampoule estde 5 10-4 g

1) Donner la composition de la solution à blanc
• Dilution
On calcule la concentration de l’acide salicylique de concentration 100mg/ml.
CAVA = CBVB
100. 10 = CB100
CB= 10mg/ml DO= 0,35
Exercice 4
Solution

2) Déterminer le volume x de réactif ferrique à ajouter à la solution à doser.
• 100ml 0,25ml de Rf
200ml X
X= 0,5 ml

• 3) Calculer la concentration d’acide
salicylique dans la solution à doser

• CAS= 10mg/ml DO( AS) = 0,35DO(s)
• DO( AS) = l1 CAS
• DO(s) = l2 C(s)
• DO( AS)/ DO(s) = CAS/ C(s)
• C(s) = CAS . DO(s) / DO( AS)
• C(s) = 10 . 0,22/ 0,35 = 6,28mg/ml

C(s) = 6,28mg/ml dans 200ml
Pour calculer la concentration dans 20ml
on doit utiliser.
CAVA = CBVB
CA = CBVB/ VA= 200. 6,28/ 20
= 62, 28mg/ml

Exercice 5
ex= 410 nm ém =540nm
130nm
Shift
Excitation Emission

3)
F= IF /(I0 - It)
• IF= 92 A partir du spectre• I0= 2476 donnée• It = ?
• A = -log It / I0 = O,15 • It / I0 = O,71
F= IF/ I0 /(I0 - It)/ I0 = 92/ 2476 / (1 – 0,71)F= 0,128

4) T= It / I0 = O,71
• A = l C
• = 0,15 / 1cm 1.10-6M
= 15. 104M-1cm-1
= f(T)
A = -log(T)
= -log(T)/l.C