apport des longues séquences lacustres à la connaissance des variations des climats et des...
TRANSCRIPT

Veget Hist Archaeobot (2005) 14:99–118DOI 10.1007/s00334-005-0065-z
O R I G I N A L A R T I C L E
Fr�d�ric Guiter · Val�rie Andrieu-Ponel ·Gunnar Digerfeldt · Maurice Reille ·Jacques-Louis de Beaulieu · Philippe Ponel
Vegetation history and lake-level changes from the Younger Dryasto the present in Eastern Pyrenees (France): pollen, plant macrofossilsand lithostratigraphy from Lake Racou (2000 m a.s.l.)Received: 24 May 2004 / Accepted: 25 January 2005 / Published online: 1 June 2005� Springer-Verlag 2005
Abstract A multi-proxy palaeoecological investigationincluding pollen, plant macrofossil, radiocarbon andsedimentological analyses, was performed on a smallmountain lake in the Eastern Pyrenees. This has allowedthe reconstruction of: (1) the vegetation history of the areabased on five pollen diagrams and eight AMS14C datesand (2) the past lake-level changes, based on plantmacrofossil, lithological and pollen analysis of twostratigraphical transects correlated by pollen analysis. Thepalaeolake may have appeared before the Younger Dryas;the lake-level was low and the vegetation dominated bycold steppic grasslands. The lake-level rose to its highestlevel during the Holocene in the Middle Atlantic (at ca.5060€45 b.p.). Postglacial forests (Quercetum mixtumand Abieto-Fagetum) developed progressively in thelower part of the valley, while dense Pinus uncinataforests rapidly invaded the surroundings of the mire andremained the dominant local vegetation until present. Theobserved lowering of the lake levels during the Late At-lantic and the Subboreal (from 5060 € B.P. to 3590€40b.p.) was related to the overgrowth of the mire. The firstobvious indications of anthropogenic disturbances of thevegetation are recorded at the Atlantic/Subboreal bound-ary as a reduction in the forest component, which hasaccelerated during the last two millennia.
Keywords Pollen analysis · Lake-level changes ·Lateglacial · Holocene · Pyrenees · Human impact
Introduction
The climate is one of the major factors influencing veg-etation dynamics and controlling the rate and mode ofdeposition and the composition of lake sediments. Thesesediments therefore provide important archives for re-constructing past climate changes and several methodshave been developed to extract climatic information fromfossil assemblages contained within them (Iversen 1958;Howe and Webb 1983; Guiot et al. 1989; Huntley 1990;Prentice et al. 1992; Moore 2002). While well-constrainedreconstructions based on fossil pollen data appear toprovide robust estimations of palaeotemperatures, calcu-lations of palaeoprecipitation and moisture-balance areless reliable, particularly for cool temperate regions wheremoisture availability is not a limiting factor for the veg-etation (Guiot et al. 1993; Almquist-Jacobson 1995).Other factors play a role in the evolution of past plantcommunities; migration lag and competition processescan buffer the effects of climate on vegetation dynamics.In areas that have a long history of human settlement, thevegetation composition has been modified by the actionsof man during the Holocene (Behre 1988). This is par-ticularly true in the Mediterranean region (Bottema et al.1990). A further drawback is that it is difficult to obtainnon-anthropogenic modern pollen analogues for recon-structing past climate changes (Davis et al. 2001). Pollendata therefore reflect the integrated effects of climate,biotic processes (Lischke et al. 2002) and anthropogenicdisturbances. We therefore require other independentclimate proxies.
Lakes are sensitive to climatically-induced changes inthe local water budget, and lake-level fluctuations providea good indicator of past climate changes (Digerfeldt 1972;Magny 1990; Harrison and Digerfeldt 1993). The level ofa closed lake is directly related to the equilibrium betweenprecipitation and evaporation over the lake surface and itscatchment (Street-Perrot and Harrison 1985; Harrison andDigerfeldt 1993). The presence of outlets can inducecomplex responses to climatic changes, by increasing or
F. Guiter ()) · V. Andrieu-Ponel · M. Reille · J.-L. de Beaulieu ·P. PonelIMEP, UMR 6116, Universit� Aix-Marseille III,Europ�le M�diterran�en de l’Arbois,B�timent Villemin, BP 80, 13545 Aix-en-Provence, Francee-mail: [email protected]
G. DigerfeldtDepartment of Quaternary Geology,Lund University,Tornav�gen 13, 22363 Lund, Sweden

decreasing outflow (Harrison and Digerfeldt 1993).However, major lake-level changes are not compensatedby changes in outflow, and small overflowing lakes maybe used to reconstruct palaeoclimatic changes (Winkler etal. 1986; Digerfeldt 1988; Guiot et al. 1993). In addition,modern closed lakes may have previously been over-flowing lakes in more humid periods (Street-Perrot andHarrison 1985).
A large number of lake level studies have been com-piled in the European Lake Status Data Bank (Yu andHarrison 1995; Yu 1996). This data set is particularlyfocused on North Europe (Digerfeldt 1972, 1976, 1988;Gaillard 1984; Almquist-Jacobson 1995; Vassiljev et al.1998; Noe-Nygaard et al. 2001), and the Alpine and Juramountains (Magny et al. 1988, 2001; Magny 1992, 2001;Lowe 1992; Magny and Ruffaldi 1995; Digerfeldt et al.1997). At present, however, there have been no lake-levelreconstructions from the French Pyrenees, despite thenumerous pollen studies in this area (Jalut 1973, 1988;Reille 1990a, b, 1993a, b; Andrieu 1991; Reille and Lowe1993; Andrieu et al. 1993; Reille and Andrieu 1994;Galop 1998). In this paper, we present a multidisciplinarystudy of an original Pyrenean overflowing mire called LeRacou, including lake-level and vegetation history re-constructions. The study includes the analyses of twotransects of eight marginal sediment cores, and combinespalaeobotanical (pollen, plant macrofossils) and sedi-mentological investigations.
Description of the study area
Physiography
The Racou mire is located at 2000 m a.s.l., on the right-hand bank of the upper TÞt river, 12 km north-west ofMont-Louis (Pyrenees Orientales) (Fig. 1). The surface ofthe lake is about 700 m2 and the maximal area of theintra-morainic former lake was approximately 17000 m2
(based on the area delimited by the current peaty shore)(Fig. 2). Today, the mire is about one meter deep and isdrained by a small outlet into the Angoustrine river(Figs. 1 and 2).
Vegetation and climate
The Racou mire is located close to the altitudinal treelineand at the boundary between subalpine forest and grass-lands. This ecotone is considered to be a highly dynamiczone and sensitive to climate changes (Kullman 1990;Hansen and di Castri 1992). The surrounding area isdominated by a dense Pinus uncinata forest mixed withBetula pubescens, characteristic of the “subalpine belt”(Gaussen 1925). Pinus uncinata, widespread in the sub-alpine eastern Pyrenean region, is associated with Arcto-staphylos uva-ursi and Juniperus communis (Gaussen1925; Gruber 1978). In places where the pine forest issparse or has been disturbed, there is an increased pres-ence of Betula pubescens, particularly on the edges of themire.
The modern climate of the area is characterised by arelatively warm and dry summer, and a cold winter. De-
Fig. 1 Location and geomor-phological context of LakeRacou. 1. Terminal moraine,morainic ridge. 2. Lake, mire
100

spite the presence of a dry summer, the climate is nottypically Mediterranean as the mean monthly precipita-tions are never lower than twice the mean monthly tem-perature (�C) (Bagnouls and Gaussen 1953). Freezingconditions occur during at least 7 months of the year.Finally, the maximum precipitation occurs in springwhilst summer and winter precipitation is low.
Geomorphological background
During the W�rm period, the TÞt glacier was the mostextensive of the eastern Pyrenees glacier systems. Severalsmall advances, the most extensive of which was 16 kmlong (Viers 1961, 1971; Taillefer 1983), left a complexseries of morainic ridges in the High TÞt valley (Fig. 1),above 1200 m altitude (Viers 1961). Compared to otherEuropean glaciers, the W�rmian expansion of the TÞtglacier was moderate. According to Taillefer (1983), thismay be a result of the small catchment area and the rel-ative drought induced by the location of glaciers insidethe massif. On the basis of geomorphological and pale-
oecological data, Andrieu et al. (1988, 2000) concludedthat the deglaciation in the Pyrenees occurred prior to38.4 kyr B.P. The subsequent ice retreat was irregular andcharacterised by successive phases of glacial advancesand episodes of deglaciation. Following the final retreatof the TÞt glacier, the valley was rapidly occupied bysmall intra-morainic lakes, e.g. Racou and La Borde(Fig. 1) (Reille 1990a).
Material and methods
Eight profiles were taken with a Russian corer (Jowsey 1966) alongtwo transects (A and B) in the overgrown part of the lake (Fig. 2).The ninth profile (C 20) was taken separately in the north-easternpart of the mire (Fig. 2). The transects ran from the mire margin tothe centre. The reference name of each core is composed of a lettercorresponding to the transect, and a number corresponding to thedistance (in meters) to the shore. Table 1 summarises the analysescarried out for each profile.
For pollen analysis, samples from gyttja and peat sediment weretreated using Faegri and Iversen’s method (1989). Samples from thebasal clay sediments were extracted using floatation on denseThoulet liquor (Goeury and Beaulieu 1979). A mean of at least 450pollen grains of terrestrial vascular plants were counted per slide.
Fig. 2 Location and lithology of the profiles sampled (profiles marked with an asterisk have been analysed both for pollen and plantmacrofossils)
Table 1 Summary of the anal-yses carried out on each Racouprofile
Reference core Lithology Pollen analysis Macrofossil analysis AMS14C dating
A 10p p p ;
A 23p p p ;
A 37p p ; ;
A 61p
Disturbed ; ;B 13
pDisturbed
p ;B 20
p p p ;B 40
p ; ; ;B 60
p ; ; ;C 20
p p p p
101

The counts are shown here as percentages of pollen sums thatexcluded spores and aquatic plant pollen. In all the diagrams, Pinusconsistently represents up to 60% of the pollen sum during thewhole Holocene. As a result, the fluctuations of other pollen curvesare reduced and the vegetation dynamics are masked. To avoid this,Pinus counts were excluded from the pollen sum. The zonation ofthe pollen diagrams is based on changes in percentages of the maintaxa.
Pinus stomata, well preserved in sediments (Trautmann 1953),were searched for systematically. In contrast to Pinus pollen, whichis wind-dispersed over large distances and often over-represented inpollen diagrams (Couteaux 1991), these stomata are residues ofneedles that are deposited locally, and indicate the local presence ofpine trees. This is particularly relevant for Pinus uncinata in highaltitude sites in the Pyrenees. Reille and Lowe (1993) have shownthat Pinus uncinata pollen frequencies may represent about 60 to75% of the modern pollen rain, even when the surface samples arelocated at some distance from any pine populations or above thetimberline. A high pollen frequency of Pinus is not a reliable pa-rameter and the local presence of pine can only be assumed wherePinus needles or stomata occur.
Samples for plant macrofossil analyses were washed through a0.4 mm sieve, the dominant seeds and fruits, and Chara oospores,are expressed in number per 50 cm3 of sediment. The sand contentof the basal sediment of the A 10, A 23 and A 37 cores wasdetermined by a simple granulometric analysis (washing andweighing). Information about the14C AMS measurements, (madeon terrestrial macrofossils from the C20 profile by the Lyons Ra-diocarbon Laboratory), is given in Table 2. Dates referred to in thetext are expressed in uncalibrated ages.
Results
In the present study, we have used the pollen analyses to:
1. reconstruct the evolution of the vegetation in the val-ley, following the final TÞt glacier retreat,
2. correlate the profiles studied along transects A and B.
The simplified pollen diagrams of 5 profiles are repre-sented in Figs. 3–7. The A 37 core is the most detailedand continuous profile for the Racou site; and the de-scription of pollen zones is based principally on the A 37pollen diagram (Fig. 3), but also includes some features ofother diagrams. A synthesis of the vegetation history ofthe eastern Pyrenees (Reille and Lowe 1993) has allowedus to compare our results with other records, particularly
those of La Borde (Fig. 1) and Gourg N�gre, sites close tothe study area. Figure 8 summarises the biostratigraphicalfeatures used to define the Local Pollen AssemblageZones (LPAZ) and correlate the profiles. A diagramshowing the varying proportions of shore and lake taxa, ispresented in Fig. 9. This was not used in establishing thebiostratigraphy, but for the lake-level reconstructions.Figures 10 and 11 show results of plant macrofossilsanalyses. In the plant macrofossil diagram of the A 37core (Fig. 11), the dominant seeds and fruits, and Charaoospores, are shown.
Vegetation history
Lateglacial (Younger Dryas)
During the Younger Dryas, the vegetation around Racoumire was dominated by steppic plants as Artemisia,Ephedra, Chenopodiaceae, Poaceae (A 37 profile: LPAZ1–4, Fig. 3; C 20 profile: LPAZ 1–2, Fig. 7). This accordswith many other Pyrenean sites (Reille 1990a, 1993a, b;Andrieu et al. 1993; Reille and Andrieu 1995). TheYounger Dryas was a rapid climatic deterioration that hada marked impact on plant communities across Europe(Lotter 1999; O’Connell et al. 1999; Ammann et al. 2000;Magny et al. 2001; Litt et al. 2001), including southernmountain areas (Penalba 1989; Reille and Lowe 1993;Beaulieu et al. 1994).
The Lateglacial vegetation is characterised by a suc-cession from an early spread of Chenopodiaceae andEphedra (LPAZ 1–2), followed by a domination of Ar-temisia associated with Poaceae and Caryophyllaceae(LPAZ 3). This differs from the vegetation dynamicsrecorded at La Borde (Reille 1990a), also located in theTÞt valley (Reille 1990a; Fig. 1) and other Pyreneanpollen records (Reille and Lowe 1993). During LPAZ 2,the percentages of Ephedra and Chenopodiaceae arehigher than those recorded at Le Monge (High Pyrenees,Reille and Andrieu 1995), Ari�ge (Reille 1990b, 1993a),and the Balc�re lake profile (Reille 1990b; Fig. 1).
The maximum phase of Chenopodiaceae and Ephedraobserved at the base of the A 37 pollen diagram was
Table 2 Radiocarbon ages ob-tained on plant macrofossilsfrom the C 20 profile
Sample(cm)
Material dated Pollen events dated AMS14Cdatings
AMS14C datings(cal.)
40–50 Mosses First occurrences of Juglans 1215€45 a.d. 941–689135–140 Pinus needles Decline of Abies 2515€35 795–519 b.c
225–230 Pinus needles First continuous occurrencesof Fagus
3590€40 2105–1782 b.c
265–270 Pinus needles Forest optimum 5060€45 3964–3712 b.c
320–325 Pinus needles First continuous occurrencesof Tilia
7360€70 6393–6032 b.c
345–350 Pinus needles Corylus maximum 8725€55 7960–7759 b.c
380–384 Wood Spread of Betula, fallof Juniperus
9735€55 9275–8964 b.c
396,5–384 RanunculusBatrachiumseeds
Base of the sequence 10360€55 10660–10009 b.c
102
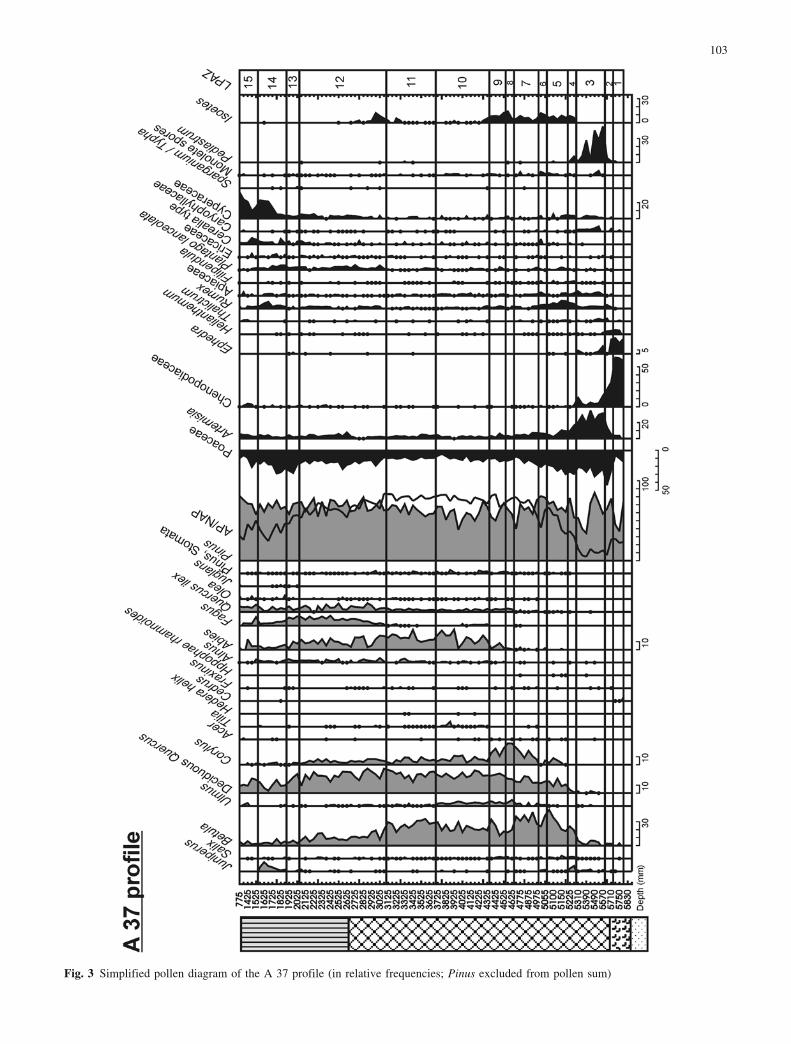
Fig. 3 Simplified pollen diagram of the A 37 profile (in relative frequencies; Pinus excluded from pollen sum)
103
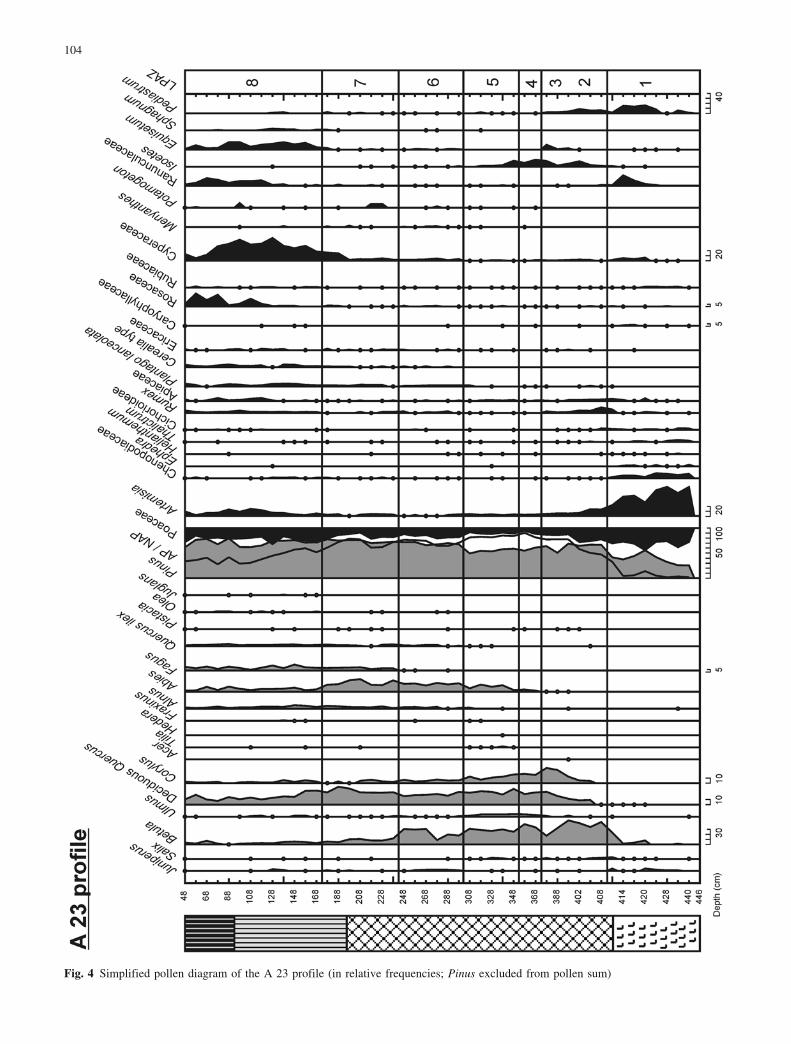
Fig. 4 Simplified pollen diagram of the A 23 profile (in relative frequencies; Pinus excluded from pollen sum)
104
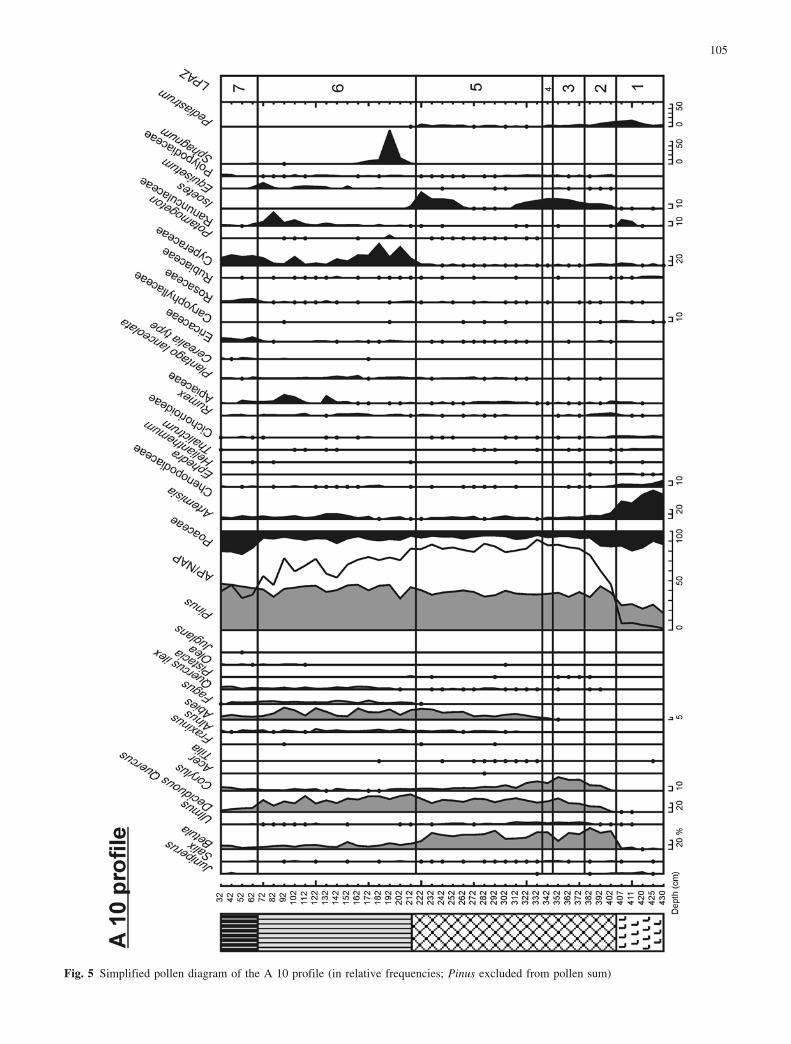
Fig. 5 Simplified pollen diagram of the A 10 profile (in relative frequencies; Pinus excluded from pollen sum)
105
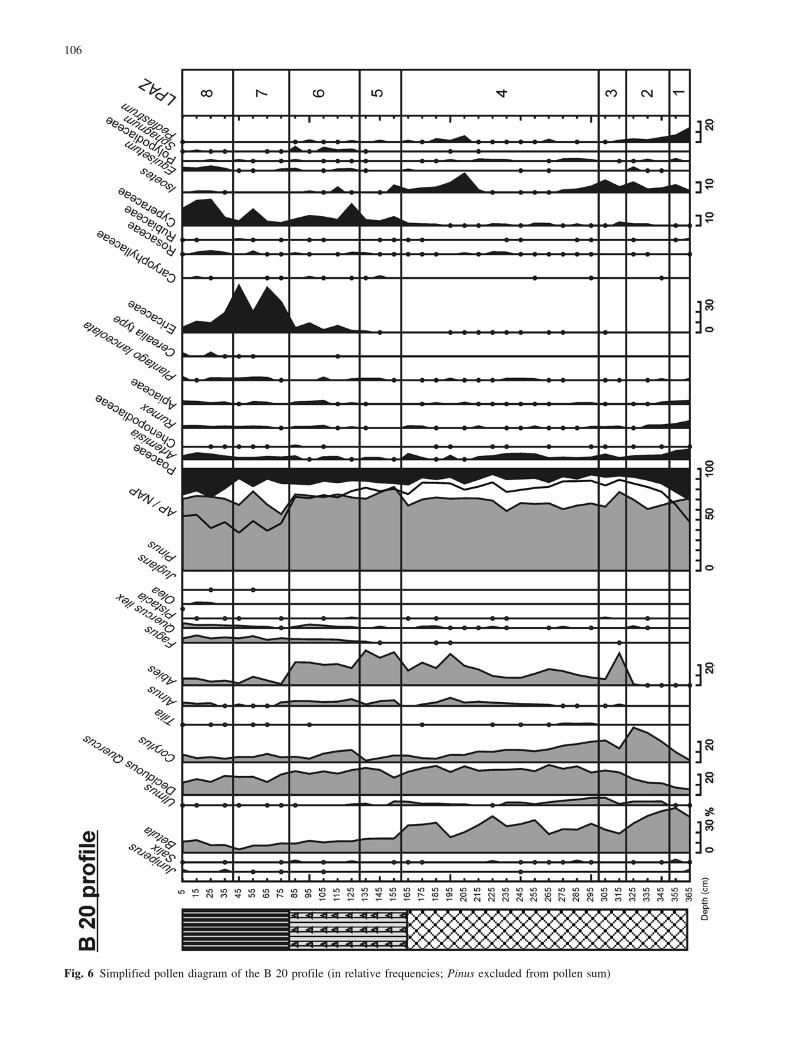
Fig. 6 Simplified pollen diagram of the B 20 profile (in relative frequencies; Pinus excluded from pollen sum)
106
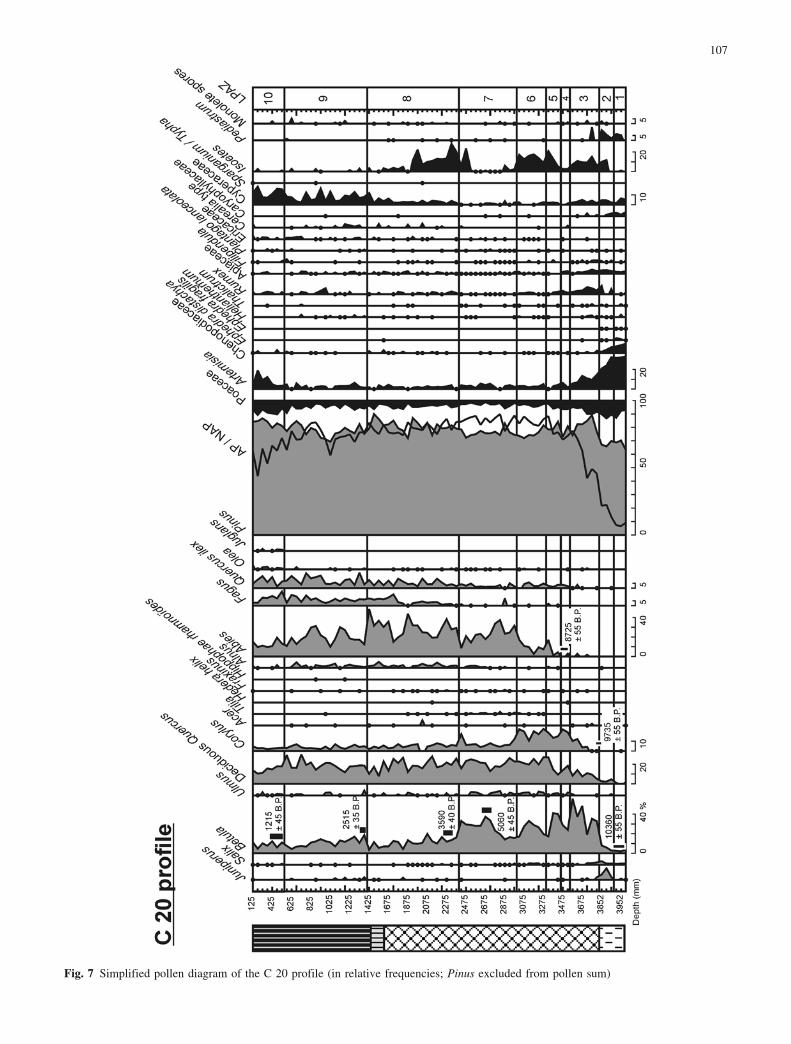
Fig. 7 Simplified pollen diagram of the C 20 profile (in relative frequencies; Pinus excluded from pollen sum)
107
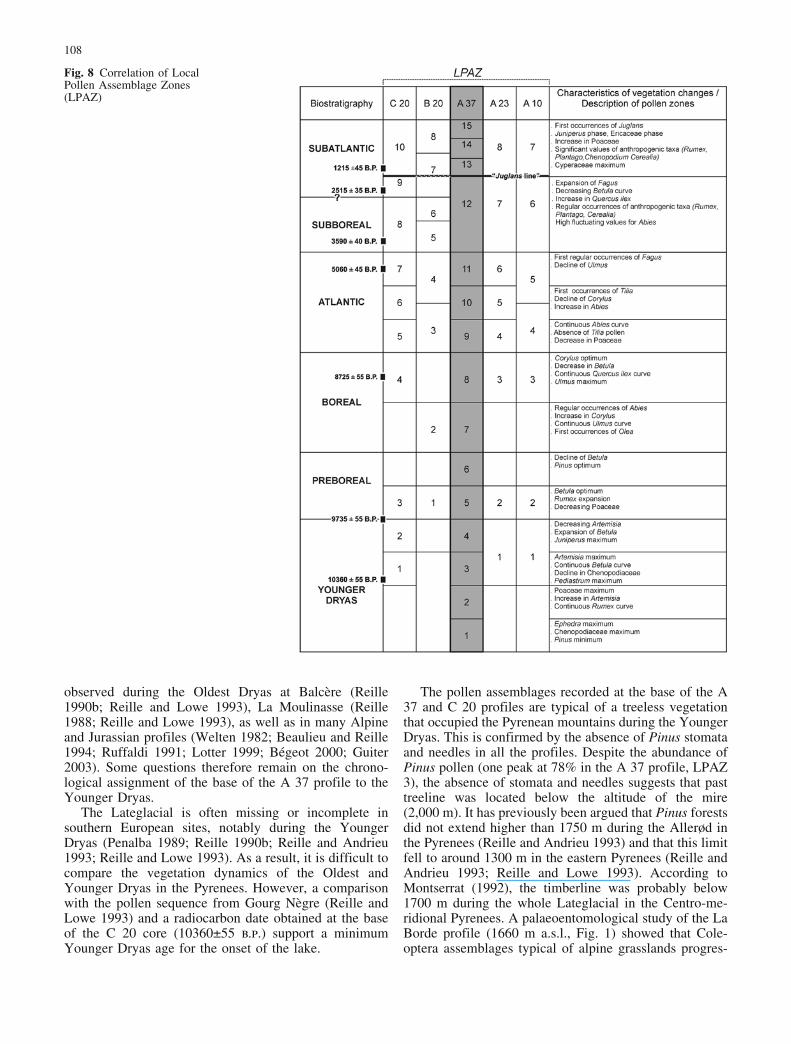
observed during the Oldest Dryas at Balc�re (Reille1990b; Reille and Lowe 1993), La Moulinasse (Reille1988; Reille and Lowe 1993), as well as in many Alpineand Jurassian profiles (Welten 1982; Beaulieu and Reille1994; Ruffaldi 1991; Lotter 1999; B�geot 2000; Guiter2003). Some questions therefore remain on the chrono-logical assignment of the base of the A 37 profile to theYounger Dryas.
The Lateglacial is often missing or incomplete insouthern European sites, notably during the YoungerDryas (Penalba 1989; Reille 1990b; Reille and Andrieu1993; Reille and Lowe 1993). As a result, it is difficult tocompare the vegetation dynamics of the Oldest andYounger Dryas in the Pyrenees. However, a comparisonwith the pollen sequence from Gourg N�gre (Reille andLowe 1993) and a radiocarbon date obtained at the baseof the C 20 core (10360€55 b.p.) support a minimumYounger Dryas age for the onset of the lake.
The pollen assemblages recorded at the base of the A37 and C 20 profiles are typical of a treeless vegetationthat occupied the Pyrenean mountains during the YoungerDryas. This is confirmed by the absence of Pinus stomataand needles in all the profiles. Despite the abundance ofPinus pollen (one peak at 78% in the A 37 profile, LPAZ3), the absence of stomata and needles suggests that pasttreeline was located below the altitude of the mire(2,000 m). It has previously been argued that Pinus forestsdid not extend higher than 1750 m during the Allerød inthe Pyrenees (Reille and Andrieu 1993) and that this limitfell to around 1300 m in the eastern Pyrenees (Reille andAndrieu 1993; Reille and Lowe 1993). According toMontserrat (1992), the timberline was probably below1700 m during the whole Lateglacial in the Centro-me-ridional Pyrenees. A palaeoentomological study of the LaBorde profile (1660 m a.s.l., Fig. 1) showed that Cole-optera assemblages typical of alpine grasslands progres-
Fig. 8 Correlation of LocalPollen Assemblage Zones(LPAZ)
108

sively replaced tree-dependent communities and becamedominant at the beginning of the Younger Dryas (Ponel etal. 1999). In summary, at this time the La Borde mire wassurrounded by steppic grasslands, possibly with a sparseshrubby cover (Juniperus, Ephedra) (Reille 1990a; Ponelet al. 1999).
This open vegetation would imply that the mire wassensitive to extra-regional pollen input (Heim 1970;Tauber 1977) and received pollen grains from a widesource area. The most striking indicator of this openlandscape is Cedrus pollen, commonly recorded in La-teglacial spectra throughout Southern Europe (Beaulieuand Reille 1973; Pons and Reille 1986; Reille 1990a;Reille and Lowe 1993) even though this taxon had beenabsent from Europe since the beginning of the Quaternary(Paganelli 1961; Ravazzi and Moscariello 1998). Cedruswas, however, present in Northern Africa during theYounger Dryas (Salamani 1991) and may have beendispersed by winds over the Mediterranean Sea as far asthe Pyrenees.
The end of the Lateglacial is characterised in the Ra-cou records by a marked increase in pollen and macro-fossils of Ranunculus-Batrachium type (Figs. 9 and 10). Asharp expansion of Ranunculus was observed in the La
Borde pollen record during the Oldest Dryas (Reille1990a). However, a small re-expansion of Ranunculus-Batrachium was also found in several eastern Pyreneansites in the latter part of the Younger Dryas (Reille andLowe 1993). This development of Ranunculus-Batrachi-um follows a maximum phase of Pediastrum (Fig. 9) thatmay indicate a significant increase in organic matter inthe basin. Similar phases have been recorded in Germansites and have been interpreted as lacustrine eutrophica-tion (Brauer et al. 1999; Kalis et al. 2003). According toJankovska and Komarek (2000), Pediastrum boryanum,predominant in Racou sediments, is abundant in smalldystrophic water bodies under cold conditions. However,some doubts remain as to whether eutrophication is theonly environmental factor that can cause an expansion ofPediastrum colonies (see Lake-level changes section be-low).
Finally, abundant fossil zygospores of Debarya glyp-tosperma were observed in the A 37 profile during the lateYounger Dryas (LPAZ 3 and 4). This member of theZygnemataceae, nowadays scarce (Transeau 1951), gen-erally occupies shallow water bodies on siliciferous soilsand indicates mesotrophic conditions (Ellis-Adam andvan Geel 1978). Collectively this evidence may indicate
Fig. 9 Pollen records from aquatic and shore vegetation (diagrams in relative frequencies). “H” indicates the period for which highestlake-levels have been reconstructed
109

changes in the lacustrine environment in response toclimatic improvement, beginning at the end of theYounger Dryas and continuing during the Early Holo-cene.
Early Holocene (Preboreal, Boreal)
Several European sites are marked by hiatuses during theEarly Holocene (Reille and Lowe 1993; Lowe and Wat-son 1993; Beaulieu et al. 1994). The Preboreal is, how-ever, documented in all the Racou pollen records (Fig. 8).The Lateglacial/Holocene boundary is dated between10360€55 b.p. and 9735€55 b.p. in the C 20 profile(Fig. 7). This corresponds to the chronological limit de-fined for eastern Pyrenees (ca. 10300 b.p. in Reille andLowe 1993).
An increase of Betula at the beginning of the Preborealis consistently found in all the Racou profiles but alsoregionally (Reille 1990a, b; Reille and Lowe 1993). Thiscoincides with an increase of tall grass/herb communities,notably Rumex, Filipendula, Plantago, and Apiaceae.Concurrent with the birch expansion, a decrease of totalN.A.P. and of Artemisia is recorded while Poaceae valuesremain relatively stable.
The start of continuous curves of Quercus, Corylusand Ulmus suggests a rapid change to postglacial forestdynamics as a result of warmer conditions. The earliestoccurrences of Pinus needles and stomata together withhigh pollen frequencies (A 37 profile: LPAZ 5, Figs. 3and 10) also indicate the presence of pine forests close tothe mire at the beginning of the Holocene. According to
Reille and Andrieu (1993), the Pinus timberline reachedthe altitude of 2200 m in the Pyrenean valleys at the onseton the Holocene. In the Racou profiles, the constant highpollen values of Pinus, together with occurrences ofneedles and stomata throughout the Holocene, show thata dense pine forest (most probably Pinus uncinata) in-vaded the subalpine area during the Preboreal and re-mained dominant until the present day, providing a rareexample of a forest ecosystem that has been stable duringthe last 10 millennia.
The extra-local vegetation is only weakly recorded inthe pollen profiles due to the domination of Pinus.However, there is some evidence for the continuing de-velopment of mixed-deciduous forests at lower altitudeduring the Boreal, for example the maximum of Corylusassociated with Quercus and Ulmus (A 37 profile: LPAZ7–8; C 20 profile: LPAZ 4). Corylus probably dominatedthe woodlands at medium altitudes and was replaced atlower altitude (<800 m) by Quercus and Ulmus (Reilleand Lowe 1993). In spite of this rapid development ofdeciduous woodlands, Pinus forests appear to have per-sisted around lake Racou, as well as the nearby sites of LaBorde (Reille 1990a) and Gourg N�gre (Reille and Lowe1993).
Regular occurrences of Abies are recorded during thelate Boreal in the A 10, A 37, A 23 and C 20 profiles. Thebeginning of the continuous curve of Abies recorded inthe C 20 profile is dated to about 8725€55 b.p. (Fig. 7).This age is in good agreement with radiocarbon datesobtained for this event in the Gourg N�gre and La Bordepollen diagrams (Reille 1990a; Reille and Lowe 1993).Collectively this suggests that Abies populations began to
Fig. 10 Batrachium seed records and sand granulometry (expressed in g per sample at constant volume) in the A 10, A 23 and A 37profiles at the boundary Lateglacial/Holocene
110

spread in the TÞt valley from their nearby glacial refugesand may have invaded the eastern Pyrenean forests (Reille1990b).
Another notable feature is the continuous curve ofQuercus ilex together with regular occurrences of Oleapollen (A 37 profile: LPAZ 6–8). In the C 20 record,evergreen oak pollen is continuously present from theBoreal period. The occurrence of these two taxa indicatesthe input of regional pollen from the vegetation devel-oping in the lowlands. This may indicate drier conditionsin the region (Reille 1990b; Reille and Lowe 1993).However, Quercus ilex values increase throughout theAtlantic biozone (A 37 profile, LPAZ 9–11, Fig. 3; C 20profile, LPAZ 5–7, Fig. 7), which is believed to have beenthe wettest period of the present interglacial. Furthermore,this drought would have occurred in the plains, whereclimatic conditions are more “Mediterranean”. Lake-levelreconstructions have helped to interpret this conflictingevidence (see Lake-level changes section below).
Middle Holocene (Atlantic)
The transition from the Boreal to the Atlantic biozone isdefined as the level where Corylus started to decrease andAbies and Quercetum mixtum pollen assemblages in-creased. The Atlantic has been described in many studiesas the climatic optimum of the Holocene (Kalis et al.2003). This period is characterised by a diversification offorests throughout the eastern Pyrenees (Reille and Lowe1993) and the Alps (Ammann 1989; Clerc et al. 1989;Lotter 1999; Ponel et al. 2001).
In the Racou sequence, the highest AP/NAP ratios arereached during the Atlantic (A 37 profile: LPAZ 9–11; C20 profile: LPAZ 5–7; Fig. 8), coinciding with a decreasein Corylus. A Quercus forest mixed with Fraxinus,Hedera, Acer and Tilia had probably developed in thelower TÞt valley, similar to the oak acidiphilous forests
currently occupying the Vallespir region (eastern FrenchPyrenees).
In the early Subboreal, Abies expanded and probablybecame the dominant conifer at medium altitudes in themountain belt. Its maximum is clearly recorded in boththe B 20 profile (LPAZ 5, Fig. 6) and A 23 profile (LPAZ7, Fig. 4). In the records from La Borde and Gourg N�gre(Reille 1990a; Reille and Lowe 1993), the spread of Abiesappears to cause a decrease in Pinus percentages. Thissuggests that the vegetation up to 2000 m altitude wasdominated by an Abies forest (Reille and Lowe 1993).The data obtained at the Racou site do not support thishypothesis. No evidence is seen in the plant macrofossils(pine needles are recorded throughout the Holocene) or inthe pollen diagrams of a decline of pine forests due tocompetition with fir. Unlike Corylus and Betula per-centages, Pinus values remain very high and relativelystable, suggesting that pine was the dominant tree speciesaround the mire throughout the Holocene. The southernexposure of the site might explain the continual presenceof Pinus, a heliophilous species, at Lake Racou whilstAbies, a sciaphilous species, was probably constrained byhumid conditions to the northern slopes of mountains.
The late Atlantic period is characterised by occur-rences of Fagus pollen that indicate the establishment ofthis tree in the valley between 3590€40 and 5060€45 b.p.(C 20 profile, Fig. 7). This corresponds with radiocarbondates obtained at Gourg N�gre (Reille and Lowe 1993)and coincides with the decline of Ulmus (A 37 profile:LPAZ 11; C 20 profile: LPAZ 7). The mid-Holocenedecline in elm forests has been observed elsewhere inEurope (Ammann 1988), resulting from plant disease(Peglar and Birks 1993) or a combination of inter-specificcompetition and human impact (Heitz-Weniger 1976;Ammann 1989; Lotter 1999). At Lake Racou, there is noclear evidence in the pollen diagrams of any anthro-pogenic disturbances, with the exception of the continu-ous curve of Plantago lanceolata in the A 37 profile(LPAZ 11, Fig. 3), A 23 profile (LPAZ 6, Fig. 4) and B 20
Fig. 11 Plant macrofossil record from A 37 profile
111

profile (LPAZ 4, Fig. 6). During this period, there arelarge fluctuations in Abies percentages in the A 37 and C20 profiles. This may indicate the beginning of distur-bances or clearings in the fir forests. Other indications arerequired, however, before these vegetation events can beattributed to human impact.
Late Holocene (Subboreal, Subatlantic)
The beginning of the Subboreal is characterised by acontinuous curve of Fagus together with a slight decreasein the A.P./N.A.P. ratio (A 37 profile: LPAZ 12; C 20profile: LPAZ 8/9; B 20 profile: LPAZ 5 and 6). Betulaundergoes the greatest decline whilst the most of the otherligneous taxa remain largely unchanged throughout thezone. The Abies record is marked by a succession of in-creases and decreases, notably in the C 20 diagram(LPAZ 8 and 9). The fir reaches maximum values in boththe C 20 record (LPAZ 8) and the B 20 record (LPAZ 5).The Subboreal is also characterised by several episodes ofexpansion of Rumex, Cerealia and Apiaceae together witha marked increase of Plantago lanceolata values thatclearly indicate successive disturbances of the vegetation.According to Tinner et al. (1999), such herbaceouscommunities recurrently spread after fires, to which Abiesis particularly sensitive. The Subboreal, which corre-sponds to the Late Neolithic, saw the first major humanexpansion in the Pyrenean Mountains (Reille and Andrieu1995; Galop 1998). This is characterised by the onset ofagro-pastoral activities and increasing disturbances offorest ecosystems. In the TÞt region, there is a sharp in-crease in the use of fires for forest clearing at about 4800b.p. (Galop 1998). Elsewhere in the region (Reille andLowe 1993), a marked decline of Abies is generally ob-served in pollen diagrams immediately following occur-rences of cereals.
It is therefore clear that the vegetation surroundingLake Racou was disturbed by human activities during theSubboreal: the significant spreading of Rumex, Plantago,Apiaceae and Poaceae indicate pastoral activities close tothe site (semi-nomadic livestock) and/or forest fires. Thefluctuations in Abies percentages and the regular occur-rences of cereals probably indicate continued anthro-pogenic activity, which also helped the establishment ofFagus in the area, where it replaced Abies (Jalut 1974;Reille 1990a, b; Reille and Lowe 1993; Reille and An-drieu 1994). The increasing curve of Quercus ilex (C 20profile, LPAZ 8; A 37 profile, LPAZ 12) is further evi-dence of the disturbance of forests in the lowlands, whereman had been present from approximately 7800 b.p.(Planchais 1985, 1987; Guenet 1995).
The palynostratigraphical boundary between the Sub-boreal and the Subatlantic is difficult to define. Tradi-tionally, the first occurrence of Juglans, which delimitsthe “Juglans line” (Beug 1975), is thought to indicate thebeginning of the Roman settlements in the region at about2200 b.p. This was used to define the limit Subboreal/Subatlantic by Reille and Lowe (1993). However, the
chronological boundary between the Subboreal and Sub-atlantic is commonly placed at about 2700 b.p. (the onsetof the Iron age). This would correspond approximately tothe transition between LPAZ 8 and LPAZ 9 in the C 20record (Fig. 7). The first occurrence of Juglans, verifiedin the C 20 record by the complete examination of du-plicate slides, is dated to 1215€45 b.p. (a.d. 689 to 941).Previous studies have shown that the first Juglans recordis, in fact, asynchronous throughout the Pyrenees (Plan-chais 1985; Galop 1998), and cannot be therefore used asa reliable chronostratigraphical marker. At Racou, how-ever, the occurrence of Juglans appears to be synchronousin all profiles and can be used as a good biostratigraphicalmarker for within-site core correlation.
The regular occurrences of Juglans coincide withsignificant values of Olea and with a number of clearindicators of intensified human disturbance: (1) a de-crease of Abies and deciduous forests; (2) a re-expansionof open-land vegetation (Poaceae, Artemisia), (3) an in-crease of nitrophilous taxa (Plantago lanceolata) andanthropogenic herb communities (Rumex, Filipendula,Cerealia, Secale).
The Fagus optimum comes after the “Juglans line”and coincides with the decline of Abies (A 37 profile:LPAZ 13; C 20 profile: LPAZ 9-10). This was previouslynoted in pollen sequences from La Restanque (Reille andAndrieu 1994) and Biscaye (Reille and Andrieu 1995). Atfirst, anthropogenic disturbances favoured Fagus expan-sion, but then resulted in a general reduction of the forestincluding both Abies and Fagus (C 20 profile: LPAZ 9,Fig. 7). With the exception of Pinus, all tree communitieswere strongly affected by human activities. This can beseen in the strong decrease of the A.P./N.A.P. ratio andthe record of Juniperus/Ericaceae shrublands in the LateSubatlantic (A 37 profile, LPAZ 13). The major expan-sion of Ericaceae recorded in the B 20 profile (LPAZ 7,Fig. 6) was not observed in the other diagrams. As the B20 profile is located at the edge of the peat-bog (Fig. 2)and pollen of Ericaceae is only dispersed over short dis-tances, this Ericaceae peak indicates a highly localisedclearing, possibly on the peat-bog itself. The slight de-cline of Pinus and Betula curves supports this, indicatingthat the forest surrounding the mire was affected (LPAZ7, Fig. 6).
The continuous curves of cereals and high values forPlantago lanceolata type indicate that agro-pastoral ac-tivities have intensified in the TÞt valley during the lasttwo millennia. This period of maximum human impacthas been observed in all Pyrenean sites (Jalut 1974; Reille1990a, b; Reille and Lowe 1993; Reille and Andrieu1994, 1995; Galop 1998).
Lake-level changes
Younger Dryas
During the Younger Dryas, a distinct lake-level loweringis recorded by an increase in coarse minerogenic matter in
112

the sediment and the deposition of sandy clay gyttja in theA 23 profile and sandy gravel in the A 10 profile(Fig. 10). The drop in level may have culminated duringthe later part of the zone. In the diagrams from transect A,the coarse minerogenic matter in the sediment shows adistinct decrease towards the end of Younger Dryas, in-dicating a rise in lake level (Fig. 10).
In the macrofossil diagrams, the Younger Dryas ischaracterised by high frequencies of Ranunculus-Batra-chium seeds, which disappear at the boundary to thePreboreal (Fig. 10). Lateglacial occurrences of Ranun-culus-Batrachium have been previously observed at sev-eral sites in the region (Reille and Lowe 1993; Reille andAndrieu 1994). Whilst the seeds cannot be identified tospecies level, most of the Lateglacial finds may originatefrom Ranunculus confervoides, which seems to be theonly European species found in artic-alpine lakes (Cook1966).
The occurrences of Ranunculus-Batrachium in the A10, A 23 and A 37 profiles during the Younger Dryas andtheir subsequent disappearance during the Preboreal mayresult from climate changes. However, the shallow waterconditions caused by the lake-level lowering during theYounger Dryas may have played a role. Ranunculus-Ba-trachium is a floating-leaved plant that normally grows inshallow water within the shore region. This can explainalso the simultaneous rich occurrence of Chara oosporesin the A 37 profile.
In the pollen diagrams, the Younger Dryas is charac-terised by high values of Pediastrum. The percentagedecreases sharply around the boundary to the Holocene.In the A 10 and A 23 profiles, it is also present during thePreboreal and the Boreal in relatively low and decreasingvalues (Fig. 9). The occurrence of Pediastrum is influ-enced by several environmental factors, complicating theinterpretation. The decrease cannot be explained by cli-mate change, as this would cause an equivalent increase atthe Holocene boundary. It also seems unlikely that anysignificant change of the trophic conditions in the lakeoccurred. However, since most Pediastrum species com-monly occur as periphytic or benthic forms (attached tolake vegetation or bottom sediment, respectively), thistaxon is most frequently found in the shore region of lakes(Cronberg 1982). The occurrences of Pediastrum maytherefore have been influenced by the recorded lake-levellowering during the Younger Dryas and the succeedingrise during the beginning of the Holocene. The floating-leaved Ranunculus cf. confervoides, which was commonin the shore region resulting from the low lake-levelsduring the Younger Dryas, would have provided a fa-vourable habitat for epiphytic forms of Pediastrum. At thesame time, the shallow water favoured the occurrence ofbenthic forms. On this basis, the disappearance of Ra-nunculus cf. confervoides at the transition to the Holocenemay help explain the coincident decrease of Pediastrum.The subsequent decrease during the Preboreal and theBoreal may be connected to the continued rise in lake-level, which increased the depth of water in the shoreregion.
The recorded lake-level lowering in Lake Racou isconsistent with both biostratigraphical and lithostrati-graphical evidence from other sites in the region, showinga cold and dry climate during the Younger Dryas (Reilleand Lowe 1993; Beaulieu et al. 1994).
Preboreal, Boreal and Atlantic
The lake-level rise, which appears to have started towardsthe end of the Younger Dryas, continued during thePreboreal and the Boreal. Pediastrum values decrease inmost diagrams (Fig. 9). However, despite the rise in lake-level, the lake must have remained relatively shallow,indicated by the high frequency of Isoetes in the pollendiagrams during both the Preboreal and the Boreal(Fig. 9). The occurrence of Isoetes is determined pri-marily by the light conditions in the bottom water, andaccordingly it grows principally within the lake margin.Based on the continued decrease and temporary disap-pearance of Isoetes, the rise in lake-level appears to haveculminated during the middle part of the Atlantic. At thistime, the water was too deep and the light conditions inthe bottom water too poor for the occurrence of Isoetes.
Subboreal and Subatlantic
In both the A 10 and B 20 profiles, the lake was over-grown by a sedge fen at the boundary between the At-lantic and the Subboreal. At A 23, this took place duringthe Subboreal and at A 37 during the earlier part of theSubatlantic. This would be partly responsible for the in-creasing sediment accumulation and resulting decrease inwater depth. In the A 10, A 37 and B 20 profiles, this isshown by the reappearance of Isoetes (Fig. 9). In A 37, aspread of Potamogeton was observed prior to the finalovergrowth.
However, the fact that the overgrowth became so ex-tensive and proceeded so rapidly suggests that this wasaccelerated by a simultaneous lowering of lake-level. AtB 20, the layer of sedge peat containing Pinus stumpsindicates a drier period with a lowered watertable duringthe Subboreal, during which time Pinus was able tospread and occupy the marginal part of transect B.
During the Upper Holocene (i.e. Subatlantic), thelowering appears to have been succeeded by a rise inlake-level. The marginal fen in transect B became againwetter and Pinus was restricted to areas closer to theshore. Pinus stumps are not found in the peat formedduring this period. The formation of quagmire (floatingpeat) observed in the A 61 profile may also be indicativeof this rise (Fig. 2). During the rise in lake-level, the peatoverlying the detritus gyttja was probably lifted by in-ternal gas pressure. Such quagmire formation is commonin flooded peatlands.
113

Discussion
Chronology of deglaciation
The oldest pollen record (profile A 37) begins during theYounger Dryas (LPAZ 1–4; Fig. 3) and no pollen wasfound below in the basal clay from 6 to 5.88 m. The siteof La Borde (Fig. 1) (Reille 1990a) located at1660 m a.s.l. in the lower TÞt valley, provided an oldercontinuous pollen record from the beginning of the La-teglacial Interstadial. This asynchronism between theonsets of the two lakes suggests that the glacier had re-treated from La Borde during the Oldest Dryas, whiledead-ice or permanent snow cover occupied the high TÞtvalley above 2000 m a.s.l. until the middle of the YoungerDryas. An alternative hypothesis is the existence of poortaphonomic conditions at Lake Racou during the La-teglacial interstadial, which prevented the deposition orpreservation of pollen at this time.
Palaeoclimatic context
Younger Dryas
During the Younger Dryas, the pollen data together withthe low lake-levels suggest that the climate was bothcooler and dryer than present. The general decrease intemperature is supported by a number of European studies(Berglund et al. 1984; Allen et al. 1996; Coope et al.1998; Lemdahl 2000; Wick 2000; Magny et al. 2001), andhas been observed in fossil insect records from Pyreneansites. At La Restanque (Ponel et al. 2002) and La Borde(Ponel et al. 1999), the discovery of Helophorus glacialisremains in Younger Dryas sediments suggests that meanJuly temperatures did not exceeded 10�C.
However, some data from Northern Europe suggestthat the Younger Dryas was a cold period with increasedlake-levels, as for example in Denmark (Noe-Nygaardand Heiberg 2001). The climate may have been colderand more oceanic than the more continental Bølling/Al-lerød Interstadial (Coope et al. 1998). Studied in greaterdetail, the records from Jura and Swiss mountains show apalaeohydrological tripartition (Magny and Ruffaldi1995; Magny 2001), with a low mid-Younger Dryas lake-level intercalated between two transgressive episodes.Similar fluctuations in lake-level have also been recordedat Le Locle, Jura (Magny et al. 2001). This tripartition hasalso been observed from alpine pollen records (Lotter etal. 1992; Richard and B�geot 2000; Guiter 2003), where itis characterised by:
– A first episode with an increase of pioneer herbs and adecrease of Pinus,
– A second period characterised by an expansion of Pi-nus and a transitory decline of heliophilous grasses,
– A third period characterised by an expansion of steppicplants together with an increase in Betula.
This tripartition can be observed in the record from pro-file A 37 (LPAZ 1–4). However, the lake-level changesdo not support this hypothesis, as only a low lake-level(with a succeeding rise at the transition to the Holocene)has been reconstructed at Racou. It is possible that there isa hiatus in the sediments that correspond to the beginningof the Younger Dryas.
Early and Mid Holocene
In a number of European records, the beginning of theHolocene is characterised by an abrupt fall in lake-level(Ammann 1975; Magny 2001; Hormes et al. 2001; Kaliset al. 2003). The hypothesis of an Early Holocene dryperiod (Guiot and Pons 1986; Reille and Lowe 1993) is,however, in contradiction to the rise in lake-level recon-structed at Racou during the Preboreal and Boreal. Inaddition, the pollen records at Racou do not show an earlyHolocene aridification; significant values of Rumex, Fil-ipendula and Apiaceae during the Preboreal are indicativeof relative wet conditions.
Analysis of fossil insect remains at a nearby site, LaRestanque (1620 m a.s.l., eastern Pyrenees, France), haveshown a sharp increase in Drusus discolor at the start ofthe Holocene (Ponel et al. 2002). The abundance of thisTrichopteran indicates that there was a substantial inputof running water into the mire; this agrees well with a risein lake-level at Lake Racou.
Palaeohydrological investigations from several Euro-pean regions have shown discordant patterns throughoutthe Holocene, with discrepancies observed between proxydata from southern and central Europe (Magny et al.2002). Unfortunately, there is no existing work on lake-level changes in the Pyrenees that can be used to assessthe Racou records. However, comparison with recordsfrom the Mediterranean region shows a coherent patternwith a first phase dominated by relatively wet conditionsuntil the Mid Holocene, followed by a progressive cli-matic aridification during the second part of the Holo-cene. In addition, recent palaeohydrological investiga-tions in Spain (Reed et al. 2001) have also shown a humidperiod in the mid-Holocene (at about 6000–5830 b.p.)followed by a general desiccation such as recorded atLake Racou (Fig. 9).
Late Holocene
Using ratios between deciduous and sclerophytic plants inMediterranean pollen sequences, Jalut et al. (2000) haveidentified six episodes of drought throughout the Holo-cene with the last two phases occurring during the Sub-atlantic. This is inconsistent with the Racou records,where the Late Holocene is characterised by an over-growth of the mire coupled with a lowering of the water-table, followed by a rise in lake-level that probably cor-responds to the beginning of the Subatlantic. Moreover,the ratios between sclerophytics and deciduous vegetation
114

do not have a reliable climatic significance (Magny et al.2002), and the results from Lake Racou appear to supportthis. Furthermore, several studies in North-West Europehave shown a change to cooler and wetter conditions,together with a change in the species composition of peat-forming Sphagna at the Subboreal/Subatlantic transition(van Geel and Renssen 1998; van Geel and Berglund2000). Therefore, the increase of humidity in the lateHolocene at Lake Racou and in the Eastern Pyreneesappears to be consistent with a more general increase inWestern Europe.
Human activities in the region
The history of human activities in Pyrenees is relativelywell-documented from archaeological data. During theMagdalenian period, the Pyrenean Mountains were oc-cupied by nomadic populations as shown by regular oc-currences of lithic industry found all over the chain(Schmider 1978). At that time, anthropogenic pressurewas restricted to hunting activities and vegetation re-mained slightly disturbed. This is highlighted by very lowcharcoal deposition in the archaeological sites dated tothis period (Solari and Vernet 1992; Heinz et al. 2004).The Racou pollen records do not indicate any significantimpact on the vegetation until the mid Holocene (At-lantic).
The three oldest archaeological sites near Lake Racou(about 60 km away) are located in the eastern Pyreneesforeland and are dated to the Early Neolithic:
The stratigraphical sequence at Montou evidenced thatdrastic environmental changes occurred between the LateNeolithic and the Late Bronze Age (Heinz et al. 2004).This period was characterised by spreading of ther-mophilous/mesophilous Mediterranean plant formationscorrelated with high charcoal accumulations and vegeta-tion changes.
The study of Cova de l’Esp�rit cave indicated thatearly notable effects of man on vegetation cover weredated to the Middle Neolithic and corresponded to thedevelopment of agriculture and animal husbandry in thePyrenean foreland (Solari and Vernet 1992).
Anthropogenic facies have been described in theB�lesta cave (Brochier and Claustre 1994); in this area,extensive pastoral activities led to the opening of theforest during the Neolithic. Anthracological and archae-ological investigations evidenced that increased humanoccupation during the early Bronze Age (Late Neolithic)resulted in vast deforested areas (Brochier et al. 1998).
The nature, extent and chronology of human activitiesin our study area at that time remain poorly documented.However, in the close vicinity of Lake Racou, Neolithicvestiges of animal husbandry have been observed fromthe lower part of the TÞt valley to the highest altitudes(Galop 1998), suggesting that pastoral activities werewidespread in the region. All the Racou pollen records arecharacterised during the Late Holocene by phases ofRumex, Plantago, Poaceae and Apiaceae, indicating that
pastoral activities occurred nearby. This confirms alsothat the intensification of human pressure started duringthe Middle/Late Neolithic.
Lastly, several archaeological studies have clearlydemonstrated that Pyrenean high-altitude vegetation wasdeeply affected by deforestation during the Middle andModern Ages (Rendu 1987; Davasse et al. 1997; Riera etal. 2004), but human occupation of the TÞt valley itselfduring the mediaeval period remains poorly known.Moreover, the Racou pollen records are not sufficientlydetailed for the last millennium and, generally the vege-tation dynamics during this period remain poorly docu-mented in spite of the large number of palaeoecologicalstudies carried out in the Pyrenees.
Acknowledgements We would like to thank C. PeÇalba for usefulcomments and suggestions. S. Brewer is thanked for constructivecomments and improving the English
References
Allen, J.R.M., Huntley, B., Watts, W.A. (1996). The vegetation andclimate of northwest Iberia over the last 14000 year. Journal ofQuaternary Science, 11, 125–147
Almquist-Jacobson, H. (1995). Lake-level fluctuations at Ljustj�r-nen, central Sweden and their implications for the Holoceneclimate of Scandinavia. Palaeogeography, Palaeoclimatology,Palaeoecology, 118, 269–290
Ammann, B. (1975). Vegetationskundliche und pollenanalytischeUntersuchungen aus dem Heidenweg im Bielersee. Beitr�geGeobotanische Landesaufnahme der Schweiz, 56, 76
Ammann, B. (1988). Palynological evidence of prehistoric an-htropogenic forest changes on the Swiss Plateau. In: Birks,H.J.B., Kaland P.E., Moe, D. (eds) The cultural landscape-past,present and future. Cambridge University Press, Cambridge, pp289–302
Ammann, B. (1989). Late-Quaternary Palynology at Lobigensee.Regional vegetation history and local lake development. Dis-sertationes Botanicae, 137, 1–157
Ammann, B., Birks, H.J.B., Brooks, S.J., Eicher, U., von Grafen-stein, U., Hofmann, W., Lemdahl, G., Schwander, J., Tobolski,K., Wick, L. (2000). Quantification of biotic responses to rapidclimatic changes around the Younger Dryas. A synthesis.Palaeogeography, Palaeoclimatology, Palaeoecology, 159,313–347
Andrieu, V. (1991). Dynamique du pal�oenvironnement de lavall�e montagnarde de la Garonne (Pyrenees centrales, France)de la fin des temps glaciaires l’actuel. Thesis, Toulouse, 330p
Andrieu, V., Eicher, U., Reille, M. (1988). Chronologie de lad�glaciation des Pyrenees franÅaises. Dynamique de s�dimen-tation et contenu pollinique des pal�olacs: application l’in-terpr�tation du retrait glaciaire. Bulletin de l’AFEQ, 213, 55–67
Andrieu, V., Eicher, U., Reille, M. (1993). The end of the lastPleniglacial in the Pyrenees (France): pollen analytical, isotopicand radiometric data. Comptes Rendus Academic ScienceParis, S�rie II., 316, 245–250
Andrieu, V., de Beaulieu, J.-L., Ponel, P., Guiter, F. (2000). En-vironnement glaciaire et v�g�tations dans les PyreneesfranÅaises entre 25 ka cal. BP et 15 ka cal. BP. Rapport duProgramme National d’Etude de la Dynamique du Climat(PNEDC) Novembre 2000, pp 1–6
Bagnouls, F., Gaussen, H. (1953). Saison s�che et indicex�rothermique. Documents Cartographiques de la ProductionV�g�tale des Services G�n�raux, Toulouse, 8, 47
de Beaulieu, J.L., Reille, M. (1973). L’interpr�tation des spectrespolliniques de p�riodes froides : propos du C�dre au Qua-
115

ternaire en Europe. IX Congr�s international INQUA,Christchurch, 1973, pp 198–199
de Beaulieu, J.L., Reille, M. (1994). Pal�oenvironnement tardi-glaciaire et holoc�ne des lacs de Pell�autier et Siguret (Hautes-Alpes, France). Histoire de la v�g�tation d’apr�s les analysespolliniques. Ecologia Mediterranea, 9, 19–34
de Beaulieu, J.L., Andrieu V., Ponel, P., Reille, M. (1994). TheWeichselian Late-glacial in Southwestern Europe. Journal ofQuaternary Science, 9, 101–107
B�geot, C. (2000). Histoire de la v�g�tation et du climat au cours duTardiglaciaire et du d�but de l’Holoc�ne sur le massif jurassiencentral partir de l’analyse pollinique et des macrorestesv�g�taux. Thesis, Franche-Comt� University, 199 p
Behre, K.-E. (1988). The role of man in European vegetation his-tory. In: Huntley, B., Webb, T. III (eds) Vegetation history.Kluwer, Dordrecht, pp 633–672
Berglund, B.E., Lemdahl, G., Liedberg-Jçnsson, T. (1984). Bioticresponse to climatic changes during the time span 13,000–10,000 B.P. A case study from SW Sweden. In: Mçrner, N.-A.,Karlen, W. (eds). Climatic changes on a Early to millennialbasis, Reidel Publishing Company, Dodrecht, pp 25–36
Beug, H.-J. (1975). Man as a factor in the vegetational history ofthe Balkan Peninsula. Proceedings of the 1st InternationalSymposium on Balkan Flora and Vegetation, Varna 1973, pp72–77
Brauer, A., Endres, C., Gunter, C., Litt, T., Stebich, M., Negen-dank, F.W.J. (1999). High-resolution sediment and vegetationresponses to Younger Dryas climate change in varved lakesediments from Meerfelder Maar, Germany. Quaternary Sci-ence Reviews, 18, 321–326
Brochier, E., Claustre, F. (1994). Le parcage des bovines et leprobl�me des liti�res du N�olithique final l’ffge du Bronzedans la Grotte de B�lesta. 24�me congr�s pr�historique deFrance: Habitats, Economies et Soci�t� du Nord-Ouestm�diterran�en, Carcassonne 26–30 septembre 1994, pp 27–36
Brochier, E., Claustre, F., Heinz, C.(1999). Environmental impactof Neolithic and Bronze Age farming in the eastern Pyreneesforelands, based on multidisciplinary investigations at LaCaune de B�lesta (B�lesta Cave), near Perpignan, France.Vegetation History and Archaeobotany, 7, 1–9
Bottema, S., Entjes-Nieborg, G., van Zeist, W. (1990). Man’s rolein the shaping of the Eastern Mediterranean Landscape. Pro-ceedings of the INQUA/BAI symposium on the impact of an-cient man on the landscape of the eastern Mediterranean regionand the near east, Groningen/Netherlands, 6–9 March 1989.Balkema, Rotterdam
Clerc, J., Magny, M., Mouthon, J. (1989). Histoire du milieu la-custre du Bas-Dauphin�: Le Grand Lemps. Etude paly-nologique des remplissages tardiglaciaires et holoc�nes, et miseen �vidence de fluctuations lacustres l’aide d’analyses s�di-mentologiques et malacologiques. Revue de Pal�obiologie, 8,1–19
Cook, C.D.K. (1966). A monographic study of Ranunculus sub-genus Batrachium (DC.) A. Gray. Mitteilungen der Botanis-chen Staatssammlung M�nchen, 6, 47–237
Coope, G.R., Lemdahl, G., Lowe, J.J., Walking, A. (1998). Tem-perature gradients in northern Europe during the last glacial-Holocene transition (14-9 ka14C yr B.P.) interpreted from Co-leoptera assemblages. Journal of Quaternary Science, 13, 419–433
Couteaux, M. (1991). La limite sup�rieure de la forÞt et sa valeur deseuil. Actes du colloque de Perpignan, 139–159
Cronberg, G. (1982). Phytoplankton changes in Lake Trummeninduced by restoration. Folia Limnologica Scandinavia, 18, 1–145
Davasse, B., Galop, D., Rendu, C. (1997). Paysages du N�olithique nos jours dans les Pyr�n�es de l’Est d’apr�s l’�cologie his-torique et l’arch�ologie pastorale. La dynamique des paysagesprotohistoriques, antiques, m�di�vaux et modernes. 17�mes
rencontres internationales d’Arch�ologie et d’Histoire d’An-tibes, pp 577–599
Davis, B., Stevenson, T., Juggins, S., Brewer, S. (2001). Short-termclimate events in the Mediterranean area during the Holocene:some preliminary results using pollen based reconstructions.Terra Nostra, 2, 24–29
Digerfeldt, G. (1972). The Post-glacial development of LakeTrummen. Regional vegetation history, water-level changesand palaeolimnology. Folia limnologica scandinavia, 16, 1–96
Digerfeldt, G. (1976). A Pre-Boreal water-level change in LakeLyngsjç, central Holland. Geologiska Fçreingens I StockholmFçrhandlingar, 97, 167–173
Digerfeldt, G. (1988). Reconstruction and regional correlation ofHolocene lake-level fluctuations in Lake Bysjçn, South Swe-den. Boreas, 17, 165–182
Digerfeldt, G., de Beaulieu, J.L., Guiot, J., Mouthon, J. (1997).Reconstruction and paleoclimatic interpretation of Holocenelake-level changes in Lac de St L�ger, Haute Provence,southeast France. Palaeogeography, Palaeoclimatology,Palaeoecology, 136, 231–258
Ellis-Adam, A.C., van Geel, B. (1978). Fossil zygospores of De-barya glyptosperma (de Bary) Wittr. (Zygnemataceae) in Ho-locene sandy soils. Acta Botanica, 27, 389–396
Faegri, K., Iversen, J. (1989). Faegri, K., Kaland, P.E., Krzywinski,K.. Textbook of Pollen analysis, 4th edn. Wiley and Sons,Chichester
Gaillard, M.J. (1984). A palaeohydrological study of Krage-holmssjçn (Scania, South Sweden): regional vegetation historyand water-level changes. Lundqua report, 25, 1–40
Galop, D. (1998). La forÞt, l’homme et le troupeau dans les Pyr-�n�es. Thesis, Toulouse
Gaussen, H. (1925). Le pin crochets dans les Pyr�n�es. Bulletinde la societe d’histoire naturelle de Toulouse, 3, 150–157
van Geel, B., Berglund, B.E. (2000). A causal link between a cli-matic deterioration around 850 cal. B.C. and a subsequent risein human population density in N-W Europe? Terra Nostra2000: 5th ELPD Workshop, Pallanza, 7, 126–129
van Geel, B., Renssen, H. (1998). Abrupt climate change around2,650 B.P. in North-West Europe: evidence for climatic tele-connection and a tentative explanation. In: Water, environmentand society in times of climatic change, Kluwer, Dordrecht, pp21–41
Goeury, C., de Beaulieu, J.L. (1979). A propos de la concentrationdu pollen l’aide de la liqueur de Thoulet dans les s�dimentsmin�raux. Pollen et Spores, 21, 239–252
Gruber, M. (1978). La v�g�tation des Pyr�n�es ari�geoises etcatalanes occidentales. Thesis, Aix-Marseille III, 305p
Guenet, P., (1995). Analyse pollinique du sondage du petit Caste-lou. Temps et espace dans le bassin de l’Aude du N�olithique l’Age du Fer (Guilaine J. dir.). CNRS, SDA et Conseil G�n�ralde l’Aude, Centre d’Anthropologie, Toulouse, 1995 , pp 334–341
Guiot, J., Pons, A. (1986). Une m�thode de reconstruction quanti-tative du climat partir de chroniques pollenanalytiques. Leclimat de la France depuis 15,000 ans. Comptes Rendus del’Acad�mie des Sciences, Paris, S�rie II, 302, 911–916
Guiot, J., Pons, A., de Beaulieu, J.L., Reille, M. (1989). A 140,000year climatic reconstruction from two European records. Na-ture, 338, 309–313
Guiot, J., Harrison, S.P., Prentice, C. (1993). Reconstruction ofHolocene precipitation patterns in Europe using pollen andlake-level data. Quaternary Research, 40, 139–149
Guiter, F. (2003). Contribution pollen-analytique l’histoire de lav�g�tation au cours des derniers 100000 ans dans la r�giond’Evian (Haute Savoie, France): implications pour l’histoire dudernier glacier du Rh�ne. Thesis, Aix-Marseille III, 271 p
Hansen, A., di Castri, F. (eds) (1992). Landscape boundaries:Consequences for biotic diversity and ecological flows.Springer, New York, 452 p
Harrison, S.P., Digerfeldt, G. (1993). European lakes as palaeo-hydrological and palaeoclimatic indicators. Quaternary Sciencereviews, 12, 233–248
116

Heim, J. (1970). Les relations entre les spectres polliniques r�centset la v�g�tation actuelle en Europe occidentale. Thesis, LouvainUniversity, 181 p
Heinz, C., Figueiral, I., Terral, J.-F., Claustre, F. (2004). Holocenevegetation changes in the northwestern Mediterranean: newpalaeoecological data from charcoal analysis and quantitativeeco-anatomy. The Holocene, 14, 621–627
Heitz-Weniger, A. (1976). Zum Problem des mittelholoz�nen Ul-menabfalls im Gebiet des Z�richsees (Schweiz). Bauhinia, 5,215–219
Hormes, A., M�ller, B.U., Schl�chter, C. (2001). The Alps withlittle ice: evidence for right Holocene phases of reduced glacierextent in the Central Swiss Alps. The Holocene, 11, 255–265
Howe, S., Webb, T. III (1983). Calibrating pollen data in climaticterms: improving the methods. Quaternary Science Reviews, 2,17–51
Huntley, B. (1990). European vegetation history: paleovegetationmaps from pollen data—13 000 year B.P. to present. Journal ofQuaternary Science, 5, 103–122
Iversen, J. (1958). The bearings of glacial and interglacial epochson the formation and extinction of plant taxa. In: Hedberg, O.(ed) Systematics of today. Acta Universitatis Upsaliensis(1958), pp 210–105
Jalut, G. (1973). Evolution du climat et de la v�g�tation de l’ex-tr�mit� orientale des Pyr�n�es au Tardiglaciaire et au Post-glaciaire, d’apr�s l’analyse pollinique. Comptes Rendus del’Acad�mie des Sciences, Paris, 276, 2653–2656
Jalut, G. (1974). Evolution de la v�g�tation et variations clima-tiques durant les quinze derniers mill�naires dans l’extr�mit�orientale des Pyr�n�es. Thesis, Toulouse University, 176 p
Jalut, G. (1988). Les principales �tapes de l’histoire de la forÞtpyr�n�enne franÅaise depuis 15 000 ans. Monografias del In-stituto Pirenaico de Ecologia, Jaca, 4, 609–615
Jalut, G., Amat, A.E., Bonnet L., Gauquelin, T., Fontugne, M.(2000). Holocene climatic changes in the Western Mediter-ranean, from south-east France to south-east Spain. Palaeo-geography, Palaeoclimatology, Palaeoecology, 160, 255–290
Jankovska, V., Komarek, J. (2000). Indicative value of Pediastrumand other coccal green algae in palaeoecology. Folia Geob-otanica, 35, 59–82
Jowsey, P.C. (1966). An improved peat sampler. New Phytologist,65, 245–248
Kalis, A.J., Merkt, J., Wunderlich, J. (2003). Environmentalchanges during the Holocene climatic optimum in central Eu-rope –human impact and natural causes. Quaternary ScienceReviews, 22, 33–79
Kullman, L. (1990). Dynamics of altitudinal tree-limits in Sweden:a review. Norsk Geographisk Tidsskrift, 44, 103–116
Lemdahl, G. (2000). Lateglacial and Early Holocene insect as-semblages from sites at different altitudes in the Swiss Alps—implications on climate and environment. Palaeogeography,Palaeoclimatology, Palaeoecology, 159, 293–312
Lischke, H., Lotter, A.F., Fischlin, A. (2002). Untangling a Holo-cene pollen record with forest model simulations and inde-pendent climate data. Ecological Modelling, 150, 1–21
Litt, T., Brauer, A., Goslar, T., Merkt, J., Balaga, K., M�ller, H.,Ralska-Jasiewiczowa, M., Stebich, M., Negendank, J.F.W.(2001). Correlation and synchronisation of Lateglacial conti-nental sequences in northern central Europe based on annuallylaminated lacustrine sediments. Quaternary Science Reviews,20, 1233–1249
Lotter, A.F. (1999). Late-glacial and Holocene vegetation historyand dynamics as shown by pollen and plant macrofossil anal-ysis in annually laminated sediments from Soppensee, centralSwitzerland. Vegetation History and Archaeobotany, 8, 165–184
Lotter, A.F., Eicher, U., Siegenthaler, U., Birks, H.J.B. (1992).Late-glacial climatic oscillations as recorded in Swiss lakesediments. Journal of Quaternary Science, 7, 187–204
Lowe, J.J. (1992). Pollen stratigraphy and radiocarbon dating ofLateglacial and early Holocene lake sediments from thenorthern Apeninnes, Italy. Boreas, 21, 193–208
Lowe, J.J., Watson, C. (1993). Lateglacial and early Holocenepollen stratigraphy of the northern Apennines, Italy. QuaternaryScience Reviews, 12, 727–738
Magny, M. (1990). Une approche pal�oclimatique de l’Holoc�ne:les fluctuations des lacs du Jura et des Alpes du NordfranÅaises. Thesis, BesanÅon
Magny, M. (1992). Holocene lake-level fluctuations in Jura and thenorthern subalpine ranges, France: regional pattern and climaticimplications. Boreas, 21, 319–334
Magny, M. (2001). Palaeohydrological changes as reflected bylake-level fluctuations in the Swiss plateau, the Jura mountainsand the northern French Pre-Alps during the Last Glacial-Ho-locene: a regional synthesis. Global and Planetary Change, 30,85–101
Magny, M. Ruffaldi, P. (1995). Younger Dryas and early Holocenelake-level fluctuations in the Jura mountains, France. Boreas,24, 155–172
Magny, M., Richard, H., Evin, J. (1988). Nouvelle contribution al’histoire holoc�ne des lacs du Jura franÅais : recherches s�di-mentologiques et palynologiques sur les lacs de Chalain, deClairvaux et de l’Abbaye. Revue de Pal�obiologie, 7, 11–23
Magny, M., Guiot, J., Schoellammer, P. (2001). Quantitative re-construction of Younger Dryas to Mid-Holocene Paleoclimatesat Le Locle, Swiss Jura, using Pollen and lake-level data.Quaternary Research, 57, 170–180
Magny, M., Miramont, C., Sivan, O. (2002). Assessment of theimpact of climate and anthropogenic factors on HoloceneMediterranean vegetation in Europe on the basis of palaeohy-drological records. Palaeogeography, Palaeoclimatology,Palaeoecology, 186, 47–59
Montserrat, J.M. (1992). Evolucion Glaciar y Postglaciar del Climay la Vegetacion en la Vertiente Sur del Pirineo; Estudio Paly-nologico. CSIC, Zaragoza, Spain
Moore, P.D. (2002). Climate record spruced up. Nature, 417, 133–134
Noe-Nygaard, N., Heiberg, E.O. (2001). Lake-level changes in theLate Weichselian Lake Tovelde, Mon, Denmark: induced bychanges in climate and base level. Palaeogeography, Palaeo-climatology, Palaeoecology, 174, 351–382
O’Connell, M., Huang, C.C., Eicher, U. (1999). Multidisciplinaryinvestigations, including stable-isotope studies, of thick Late-glacial sediments from Tory Hill, Co. Limerick, western Ire-land. Palaeogeography, Palaeoclimatology, Palaeoecology,147, 169–208
Paganelli, A. (1961). Giornale Botanico Italiano, 69, 109–117Peglar, S.M., Birks, H.J.B. (1993). The mid-Holocene Ulmus fall at
Diss Mere, South-East England- disease or human impact.Vegetation History and Archaeobotany, 2, 1–13
Penalba, M.C. (1989). Dynamique de v�g�tation tardiglaciaire etholoc�ne du centre-nord de l’Espagne d’apr�s l’analyse polli-nique. Unpublished thesis, Aix-Marseille III
Planchais, N. (1985). Analyse pollinique du remplissage Holoc�nede la lagune de Canet (plaine du Roussillon, d�partement desPyr�n�es Orientales). Ecologia Mediterranea, 11, 117–127
Planchais, N. (1987). Impact de l’Homme lors du remplissage del’estuaire du Lez (Palavas, H�rault) mis en �vidence par l’an-alyse pollinique. Pollen et Spores, 29, 73–88
Ponel, P., Coope, G.R., Andrieu-Ponel, V., Reille, M. (1999).Coleopteran evidence for a mosaic of environments at highaltitude in the eastern Pyrenees, France, during the climatictransition between the Allerød and Younger Dryas. Journal ofQuaternary Science, 14, 169–174
Ponel, P., Andrieu-Ponel, V., Parchoux, F., Juhasz, I., de Beaulieu,J.-L. (2001). Late-glacial and Holocene high-altitude environ-mental changes in Vall�e des Merveilles (Alpes-Maritimes,France): insect evidence. Journal of Quaternary Science, 16,795–812
Ponel, P., Andrieu-Ponel, V., Reille, M., Guiter, F. (2002). Aninsect record of the last 12 ky in Pyrenees, France: comparisonwith pollen data. Quaternary Sciences from Land to Sea: inhonor of J.T. Andrews, Colorado Convention Center, DenverAnnual Meeting October 2002, pp 241–1
117

Pons, A., Reille, M. (1986). Nouvelles recherches pollenanalytiques Padul (Granada): la fin du dernier glaciaire et l’Holoc�ne.Proceedings of the Symposium on Climatic Fluctuations duringthe Quaternary in the Western Mediterranean Regions, Madrid,1986, 405–419
Prentice, I.C., Cramer, W., Harrison, S.P., Leemans, R., Monserud,R.A., Solomon, A.M. (1992). A global biome model based onplant physiology and dominance, soil properties and climate.Journal of Biogeography, 19, 117–134
Ravazzi, C., Moscariello, A. (1998). Sedimentation, palaeoenvi-ronmental evolution and time duration of earliest Pleistoceneclimatic cycles in the 24–5 6 m FM core interval (Leffe Basin,Northern Basin). Mededelingen Nederlands Instituut voorToegepaste Geowetenschappen TNO, 60, 467–489
Reed, J.M., Stevenson, A.C., Juggins, S. (2001). A multi-proxyrecord of Holocene climatic change in southwestern Spain: Thelaguna di Medina, Cadiz. The Holocene, 11, 707–720
Reille, M. (1988). Nouvelles analyses polliniques de la tourbi�re dela Moulinasse: versant nord-oriental des Pyr�n�es (France).LBHP unpublished report, 21 p
Reille, M. (1990a). La tourbi�re de la Borde (Pyrenees Orientales,France): un site cl� pour l’�tude du Tardiglaciaire sud-eu-rop�en. Comptes Rendus de l’Acad�mie des Sciences, Paris,310, 823–829
Reille, M. (1990b). Recherches pollenanalytiques dans l’extr�mit�orientale des Pyr�n�es: donn�es nouvelles, de la fin du Gla-ciaire l’Actuel. Ecologia Mediterranea, 16, 317–357
Reille, M. (1993a). New pollenanalytical researches at Freych-in�de, Ari�ge, Pyrenees, France. Dissertationes Botanicae, 196,377–386
Reille, M. (1993b). L’interface Tardiglaciaire-Holoc�ne dans unsite du littoral atlantique sud-europ�en: Le Moura (PyreneesAtlantiques, France). Comptes Rendus de l’Acad�mie desSciences, Paris, 316, 463–468
Reille, M., Andrieu, V. (1993). Variations de la limite sup�rieuredes forÞts dans les Pyr�n�es (France) pendant le Tardiglaciaire.Comptes Rendus de l’Acad�mie des Sciences, Paris, 316, 547–551
Reille, M., Andrieu, V. (1994). Vegetation history and human ac-tion in Ariege (Pyrenees, France). Dissertationes Botanicae,234, 413–422
Reille, M., Andrieu, V. (1995). The late Pleistocene and Holocenein the Lourdes basin, Western Pyrenees, France: new pollenanalytical and chronological data. Vegetation History and Ar-chaeobotany, 4, 1–21
Reille, M., Lowe, J.J. (1993). A re-evaluation of the vegetationhistory of the eastern Pyrenees (France) from the end of theLast Glacial to the present. Quaternary Science Reviews, 12,47–77
Rendu, C. (1987). Quelques jalons pour une histoire des forÞts enCerdagne: le massif d’Oss�ja entre 1030 et 1430. Etudesroussillonnaises offertes Pierre Ponsich, Perpignan, Le Pub-licateur, pp245–251
Richard, H., B�geot, C. (2000). Le Tardiglaciaire du massif jur-assien: bilan et perspectives de recherches. Quaternaire, 11,145–154
Riera, S., Wansard, G., Juli, R. (2004). 2000-year environmentalhistory of a karstic lake in the Mediterranean Pre-Pyrenees: theEstanya lakes (Spain). Catena, 55, 293–324
Ruffaldi, P. (1991). Premi�re contribution l’�tude de la v�g�tationtardiglaciaire et holoc�ne du Bugey: l’exemple de la tourbi�rede Cerin (Ain, France). Revue de Pal�obiologie, 10, 137–149
Salamani, M. (1991). Premi�res donn�es palynologiques sur l’his-toire holoc�ne du massif de l’Akfadou (Grande Kabylie, Al-g�rie). Ecologia Mediterranea, 17, 145–159
Solari, M.-E., Vernet, J.-L. (1992). Late glacial and Holocenevegetation of the Corbi�res based on charcoal analysis at theCova de l’Esp�rit (Salses, Pyr�n�es Orientales, France). Reviewof Palaeobotany and Palynology, 71, 111–120
Street-Perrot, F.A., Harrison, S.P. (1985). Lake-levels and climatereconstruction. In: Hecht, A.D. (ed) Paleoclimate Analysis andModelling, Wiley, New York, pp 291–340
Taillefer, G. (1983). Les oscillations des glaciers fini- et post-gla-ciaires des Pyr�n�es Orientales. Actes du colloque “Oscillationsfini- et post-glaciaires des glaciers” Balkema, Rotterdam, pp231–244
Tauber, H. (1977). Investigations of aerial pollen transport in af-forested area. Dansk Botanisk Arkkiv, 32 p
Tinner, W., Hubschmid, P., Wehrli, M., Ammann, B., Conedera,M. (1999). Long-term forest fire ecology and dynamics insouthern Switzerland. Journal of Ecology, 87, 273–289
Transeau, E.N. (1951). The Zygnemataceae. Columbus GraduateSchool Monographs. Contributions in Botany, 1
Trautmann, W. (1953). Zur Unterscheidung fossiler Spaltçffnungender mitteleurop�ischen Coniferen. Flora, 140, 523–533
Vassiljev, J., Harrison, S.P., Guiot, J. (1998). Simulating the Ho-locene Lake-level Record of Lake Bysjçn, Southern Sweden.Quaternary Research, 49, 62–71
Viers, G. (1961). Le glaciaire du massif du Carlit (Pyr�n�es Ori-entales). Revue de G�ographie des Pyr�n�es du Sud Ouest, 32,5–33
Viers G. (1971). L’englacement quaternaire des Pyr�n�es orientaleset ses probl�mes climatiques. Colloque interdisciplinaire sur lesmilieux naturels supra-forestiers des montagnes du bassin oc-cidental de la M�diterran�e, Perpignan, pp 57–64
Welten, M. (1982). Vegetationsgeschichtliche Untersuchungen inder westlichen Schweizer Alpen: Bern-Wallis. Denkschriftender Schweizerischen Naturforschenden Gesellchaft, 95, 1–104
Wick, L. (2000). Vegetational response to climatic changesrecorded in Swiss Late Glacial lake sediments. Palaeogeogra-phy, Palaeoclimatology, Palaeoecology, 159, 231–250
Winkler, M.G., Swain, A.M., Kutzbach, J.E. (1986). Middle Ho-locene dry period in the northern Midwestern United States:lake levels and pollen stratigraphy. Quaternary Research, 25,235–250
Yu, G. (1996). Lake-level records and palaeoclimates of NorthernEurasia. Lundqua, 37, 1–17
Yu, G., Harrison, S.P. (1995). Lake status records from Europe:data base documentation. World data Center—A for Palaeo-climatology, NOAA Paleoclimatology Program, Paleoclima-tology Publications series report 3, 451 p
118