glycogen accumulations in differentiating mesonephric ducts and tubuli in male human embryos
TRANSCRIPT

Anat Embryol (1983) 168:445-453 Anatomy and Embryology �9 Springer-Verlag 1983
Glycogen Accumulations in Differentiating Mesonephric Ducts and Tubuli in Male Human Embryos
Lauri J. Pelliniemi 1, Pirkko Kellokumpu-Lehtinen 2, and Anita P. H o l l e r 3 Laboratory of Electron Microscopy 1, and Department of Anatomy 2, University of Turku, SF-20520 Turku 52, Finland, and Department of Anatomy 3 Harvard Medical School, Boston, MA. 02115, U.S.A.
Summary. Human mesonephric duct epithelial cells contained empty appearing regions in the infranuclear cytoplasm when prepared for trans- mission electron microscopy using glutaraldehyde and osmium fixation. The same regions stained positively with PAS in Epon sections for light microscopy suggesting that glycogen was present. Incubation with saliva abolished the reaction. For electron microscopy the glycogen stained very intensely if a mixture of osmium tetroxide and potassium ferro- cyanide was used instead of osmium alone. Glycogen accumulations were present between the ages of 5 to 10 weeks and absent at the age of 15 weeks. Reports by others indicate that glycogen may be present in different reactive forms in relation to its staining behaviour after various fixatives. The present results, and similar studies in other tissues, indicate that osmium tetroxide-potassium ferrocyanide fixative should be used routinely for preservation of embryos and fetuses and where indicated, for ultrastructural identification of glycogen and cytoplasmic filaments in clinical specimens.
Key words: Electron microscopy - Mesonephric duct - Fetus - Glycogen - Human development
Introduction
Differentiating cells in human blastocyst and in human embryonic and fetal genital systems contain accumulations of glycogen-like material (Hertig et al. 1958; Fukuda 1976; Niemi and Ikonen 1965; Pinkerton et al. 1961; Wartenberg et al. 1971). Similar accumulations have not been seen in animal gonads (Eddy 1974; Gondos and Conner 1973; Zamboni and Merchant 1973). On the other hand, large glycogen accumulations have been observed in the epithelial cells of mesonephric duct, which differentiates into epididy-
Offprint requests to: Lauri J. Pelliniemi, M.D., Laboratory of Electron Microscopy, University of Turku, Kiinamyllynkatu 10, SF-20520 Turku 52, Finland.

446 L.J. Pelliniemi et al.
mis in ram (Tiedemann 1971), but not in rat (Flickinger 1969) or man (De Martino and Zamboni 1966; Gould and Bernstein 1979; Holstein 1969).
We report here for the first time the presence of glycogen in human embryonic and fetal epithelial cells of the mesonephric duct. This material is not preserved by standard glutaraldehyde and osmium fixation, and there- fore may not have been visulaized in earlier studies.
Materials and Methods
The mesonephric ridges containing the mesonephric tubules, the mesonephric ducts, and the paramesonephric ducts were collected in the region adjacent to the gonad from 14 male human embryos and fetuses obtained from clinical interruptions of pregnancy. The ovulation ages were determined according to Streeter (1920) from tile measured foot lengths (FL) or crown- rump lengths (CR). The ages and corresponding lengths ranged from 5 weeks (CR 12 ram) to 15 weeks (FL 9.1 mm, CR 135 mm). The sex was identified by X chromatin staining and Y chromatin fluorescence before gonadal sex differentiation (Pelliniemi 1972) and by histologi- cal examination of gonads in older fetuses.
The ridges with the ducts were cut into 1 mm long pieces and immersed in a fixative containing 0.24 tool/1 glutardialdehyde (Glutaraldehyde EM, Leiras OY, Finland), 4.1 mmol/1 Ca C12, and 0.1 tool/1 sodium cacodylate-HC1 buffer (pH 7.4) for three hours at +4 ~ (Petlin- iemi 1975a). After washing overnight in 0.23 mol/1 sucrose, the tissue pieces were postfixed with 39 mmol/l osmium tetroxide in either 0.2 mol/l s-collidine-HC1 buffer (pH 7.4) or in 0A tool/1 s-collidine-HC1 buffer (pH 7.4) containing 36 mmol/1 potassium ferrocyanide (Kar- novsky 1971), both at 4 ~ for 2 h. The specimens were dehydrated in ethanol and embedded in Epon 812 (E. Merck, Darmstadt, FRG). Cross-sections of ducts were cut on a Sorvall MT 2 ultramicrotome with a diamond knife and stained with toluidine blue for light microscopy and with uranyl acetate (Watson 1958) and lead citrate (Venable and Coggeshall 1965) for electron microscopy. The specimens were examined with Philips 200 and JEOL JEM-100 C electron microscopes.
Glycogen was demonstrated by the periodic acid-Schiff (PAS) reaction after removal of the plastic from 1 gm sections (Lane and Europa 1965). Control sections were incubated with saliva prior to the PAS reaction for 30 min at room temperature.
Results
The epithelial cells of the mesonephric duct contained large clear areas in the infranuclear cytoplasm (Fig. 1) when fixed with conventional glutaral- dehyde and osmium tetroxide. These areas were not empty, however, be- cause they stained positively with the PAS reaction (Fig. 2). If the sections were preincubated with saliva, the PAS reaction was abolished. When a mixture of osmium tetroxide and potassium ferrocyanide was substituted for osmium tetroxide alone, the material in the previously clear areas was positively stained.
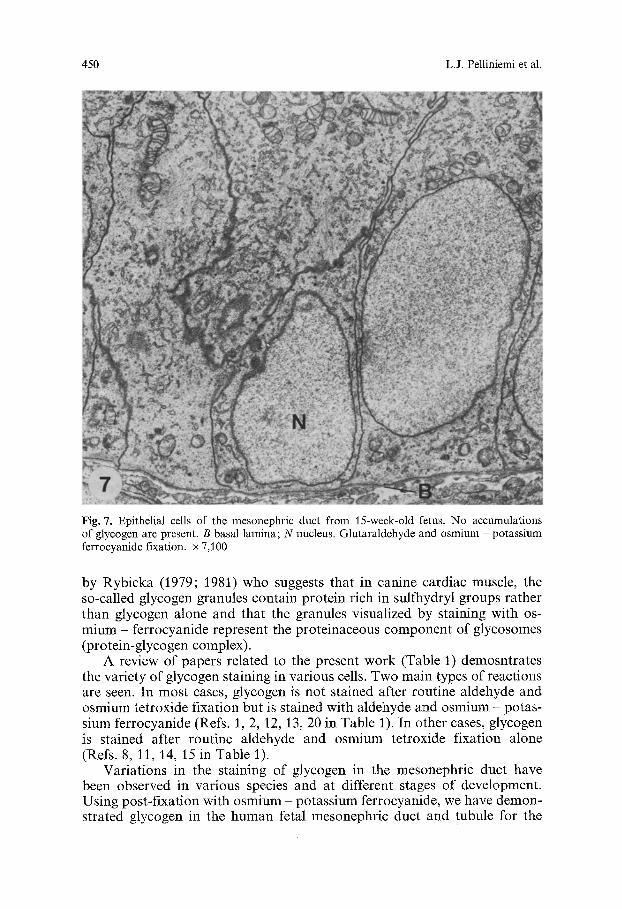
Electron microscopy showed that the basal areas of epithelial cells in osmium-fixed material were stained very weakly in comparison to the rest of the cytoplasm (Figs. 3 and 5). Osmium - potassium ferrocyanide fixation revealed accumulations of 30-50 nm glycogen particles with globular sub- units in the weakly staining areas (Figs. 4 and 6). These glycogen accumula- tions were most prominent between the ages of 5 to 10 weeks of prenatal development, and disappeared by the age of 15 weeks (Fig. 7).

Glycogen in Human Mesonephric Duct 447
Fig. 1. A survey light micrograph of mesonephric duct in cross section from an eight-week-old fetus. Glutaraldehyde and osmium fixation, toluidine blue staining, x 550
Fig. 2. Light micrograph of a mesonephric duct in cross-section stained with periodic acid - Schiff (PAS) reaction for the demonstration of glycogen. Large deposits of PAS-positive material (arrows) are present predominantly in the basal portions of the epithelial cells in the mesonephric duct. An eight-week-old fetus, glutaraldehyde and osmium fixation, x 550
Similar but smaller accumulations of glycogen particles were also ob- served, albeit less frequently, in the proximal mesonephric tubuli, the pri- mordia of the ductuli efferentes. The epithelial cells of the paramesonephric ducts, which do not degenerate until after the tenth week, contained relative- ly little glycogen.
Discussion
Accumulations of glycogen-like material are found in a great variety of epithelial tissues during prenatal development (cf. Table 1). After conven- tional aldehyde and osmium fixation, glycogen areas have an empty or a faint cotton-wool appearance. In these cases glycogen is not preserved with the fixative appropriately enough to bind subsequent heavy metal stains. A common feature in the differentiating cells is that glycogen gradual- ly disappears as the cells mature. However, in human epididymis, the basal cells contain glycogen in the adult period (Montagna and Hamilton 1952).
Ultrastructural and, in some cases, histochemical features are the criteria currently used to identify glycogen in tissue sections (Table 1). Biochemical studies and X-ray microanalysis indicate that osmium - ferrocyanide fixa- tives stain glycogen particles selectively (de Bruijn 1973), the binding takes place through the C2-C 3 dihydroxyl groups in glycogen (de Bruijn and den Breejen 1976), and Os-Fe complex is present in these particles (de Bruijn and van Buitenen 1980). The mechanism by which glycogen is stained in- volves chelation of the osmium - ferrocyanide complexes by donor atoms in the tissue macromolecules which, in turn, facilitates the reduction of osmium in sire to a more stable lower oxidation state and results in more osmium deposition than post-fixation with osmium tetroxide alone (White et al. 1979). A different view on the staining mechanism has been proposed
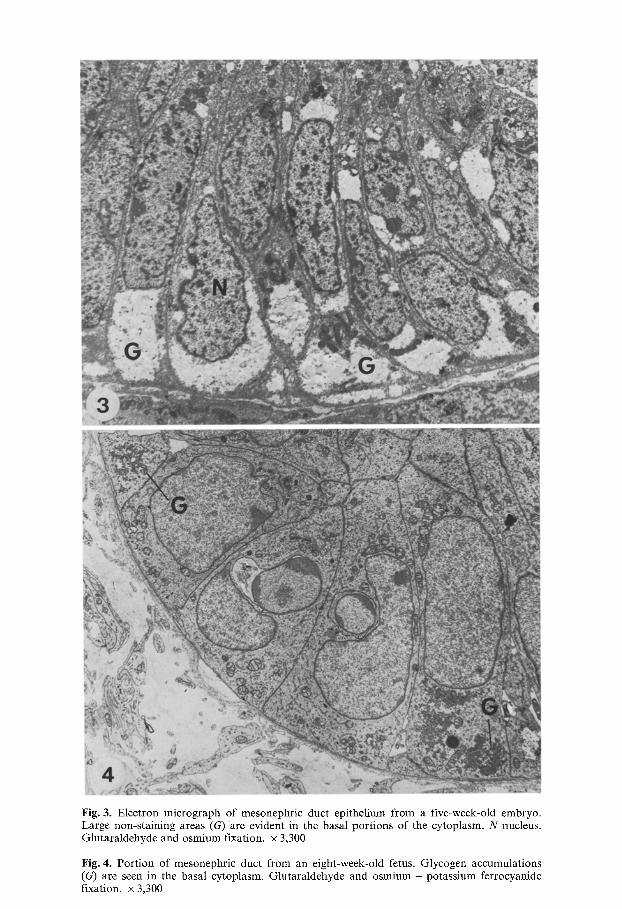
Fig. 3. Electron micrograph of mesonephric duct epithelium from a five-week-old embryo. Large non-staining areas (G) are evident in the basal portions of the cytoplasm. N nucleus. Glutaraldehyde and osmium fixation, x 3,300
Fig. 4. Port ion of mesonephric duct from an eight-week-old fetus. Glycogen accumulations (G) are seen in the basal cytoplasm. Glutaraldehyde and osmium - potassium ferrocyanide fixation, x 3,300
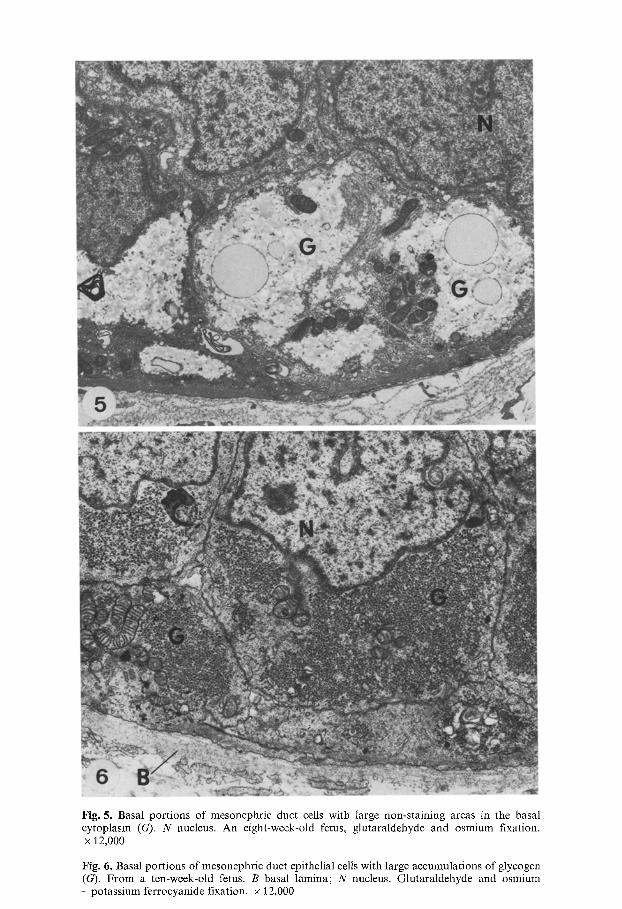
Fig. 5. Basal portions of mesonephric duct cells with large non-staining areas in the basal cytoplasm (G). N nucleus. An eight-week-old fetus, glutaraldehyde and osmium fixation. x 12,000
Fig. 6. Basal portions of mesonephric duct epithelial cells with large accumulations of glycogen (G). From a ten-week-old fetus. B basal lamina; N nucleus. Glutaraldehyde and osmium - potassium ferrocyanide fixation. • 12,000
450 L.J. Pelliniemi et al.
Fig. 7. Epithelial cells of the mesonephric duct from 15-week-old fetus. No accumulations of glycogen are present. B basal lamina; N nucleus. Glutaraldehyde and osmium potassium ferrocyanide fixation, x 7,100
by Rybicka (1979; 1981) who suggests that in canine cardiac muscle, the so-called glycogen granules contain protein rich in sulfhydryl groups rather than glycogen alone and that the granules visualized by staining with os- mium - ferrocyanide represent the proteinaceous component of glycosomes (protein-glycogen complex).
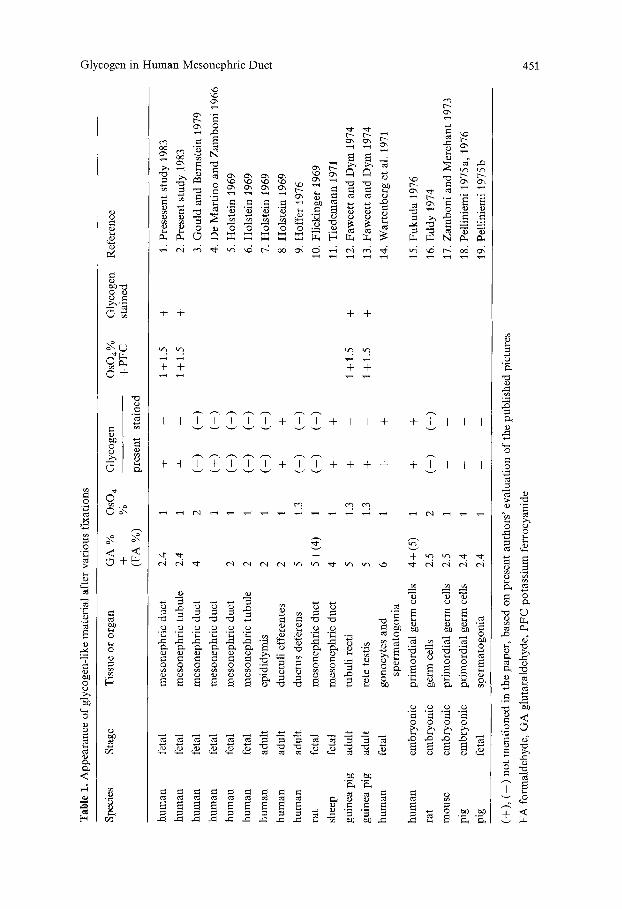
A review of papers related to the present work (Table 1) demosntrates the variety of glycogen staining in various cells. Two main types of reactions are seen. In most cases, glycogen is not stained after routine aldehyde and osmium tetroxide fixation but is stained with aldehyde and osmium - potas- sium ferrocyanide (Refs. 1, 2, 12, 13, 20 in Table 1). In other cases, glycogen is stained after routine aldehyde and osmium tetroxide fixation alone (Refs. 8, 11, 14, 15 in Table 1).
Variations in the staining of glycogen in the mesonephric duct have been observed in various species and at different stages of development. Using post-fixation with osmium - potassium ferrocyanide, we have demon- strated glycogen in the human fetal mesonephric duct and tubule for the
Tab
le 1
. A
ppea
ranc
e of
gly
coge
n-li
ke m
ater
ial
afte
r va
riou
s fi
xati
ons
Spe
cies
S
tage
T
issu
e or
org
an
GA
%
OsO
4 G
lyco
gen
OS
O4%
+
%
+ P
FC
(F
A %
) pr
esen
t st
aine
d
Gly
coge
n st
aine
d R
efer
ence
�9
%
hu
man
fe
tal
mes
onep
hric
duc
t 2.
4 1
+ -
1 +
1.5
hu
man
fe
tal
mes
onep
hric
tub
ule
2.4
1 +
- 1
+ 1.
5
hu
man
fe
tal
mes
onep
hric
duc
t 4
2 (-
) ( -
)
hu
man
fe
tal
mes
onep
hric
duc
t 1
(+)
( -
)
hu
man
fe
tal
mes
onep
hric
duc
t 2
1 (-
) (-
)
hu
man
fe
tal
mes
onep
hric
tub
ule
2 1
(-)
( -
)
hu
man
ad
ult
epid
idym
is
2 1
(-)
(-)
hu
man
ad
ult
duct
uli
effe
rent
es
2 1
+ +
hu
man
ad
ult
duct
us d
efer
ens
5 1.
3 (-
) (-
)
rat
feta
l m
eson
ephr
ic d
uct
5 +
(4)
1 (-
) (-
)
shee
p fe
tal
mes
onep
hric
duc
t 4
1 +
+
guin
ea p
ig
adul
t tu
buli
rec
ti
5 1.
3 +
- 1
+ 1.
5
guin
ea p
ig
adul
t re
te t
esti
s 5
1.3
+ -
1 +
1.5
hu
man
fe
tal
gono
cyte
s an
d 6
1 +
+ sp
erm
atog
onia
hu
man
em
bryo
nic
prim
ordi
al g
erm
cel
ls
4 +
(5)
1 +
+
rat
embr
yoni
c ge
rm c
ells
2.
5 2
(-)
(-)
mou
se
embr
yoni
c pr
imor
dial
ger
m c
ells
2.
5 1
- -
pig
embr
yoni
c pr
imor
dial
ger
m c
ells
2.
4 1
- -
pig
feta
l sp
erm
atog
onia
2.
4 1
- -
+ + + +
1. P
rese
sent
stu
dy J
983
2. P
rese
nt s
tudy
198
3 =
3. G
ou
ld a
nd B
erns
tein
197
9 o
4. D
e M
arti
no
and
Zam
bo
ni
1966
5. H
olst
ein
1969
:r
6. H
olst
ein
1969
7. H
olst
ein
1969
8. H
olst
ein
1969
9. H
offe
r 19
76
10.
Fli
ckin
ger
1969
11.
Tie
dem
ann
197
1
12.
Faw
cett
and
Dy
m 1
974
13.
Faw
cett
and
Dym
197
4
14.
War
tenb
erg
et a
l. 1
971
15.
Fu
ku
da
1976
16.
Edd
y 19
74
17.
Zam
bo
ni
and
Mer
chan
t 19
73
18.
Pel
lini
emi
1975
a, 1
976
19.
Pel
lini
emi
1975
b
(+),
(--
) no
t m
enti
oned
in
the
pape
r, b
ased
on
pres
ent
auth
ors'
eva
luat
ion
of t
he p
ubli
shed
pic
ture
s
FA
for
mal
dehy
de,
GA
glu
tara
ldeh
yde,
PF
C p
otas
sium
fer
rocy
anid
e

452 L.J. Pelliniemi et al.
first time. The non-staining areas shown in the human mesonephric duct by de Mart]no and Zamboni (1966, Figs. I and 2) also suggest the presence of glycogen but the authors do not mention it in the text. Using only osmium tetroxide as a post-fixative, glycogen is not observed in the mesonephric duct of 1,2- and 16-week-old fetuses (Holstein 1969; Gould and Bernstein 1979) or in the fetal rat epididymis (Flickinger 1969). By contrast, a compre- hensive analysis of the mesonephric duct in the ram reveals glycogen accu- mulations similar to those in the present study (Tiedemann 1971). However, the glycogen deposits in the ram mesonephric duct differ from those in the human in that they require only aldehyde fixation and post-fixation with osmium rather than aldehyde and osmium - potassium ferrocyanide fixation as the glycogen in the human mesonephic duct.
The differences in the staining behaviour of glycogen in various cells suggest that glycogen is present as chemically different forms or complexes in various species and cells. On the other hand, the ambiguities in the inter- pretations of the reaction mechanism of the osmium potassium ferrocyanide fixative as described above hamper the selective application of this useful tool. Regardless of the exact mechanism of staining, it is clear that osmium - potassium ferrocyanide provides excellent preservation and contrast of glycogen complexes which in many cases otherwise remain unstained. Addi- tional contrast is also given to glycocalyx-related carbohydrate-containing structures, and cytoplasmic filaments (David-Ferreira and David-Ferreira 1980). Ferrocyanide-reduced osmium is therefore recommended as a routine fixative for the preservation of embryos and fetuses and where indicated, for ultrastrctural identification of glycogen and cytoplasmic filaments in clinical specimens.
Acknowledgements. The authors wish to express their appreciation to Dr. Don Fawcett for his constructive criticism of the manuscript and to Sirpa From, Georgia Hsiao, Mauno Lehti- m/iki, Marja Mattila and Ulla Mfintyl~i for excellent technical and secretarial assistance.
References
Bruijn WC de (1973) Glycogen, its chemistry and morphologic appearance in the electron microscope. ~1. A modified OsO 4 fixative which selectively contrasts glycogen. J Ultrastruct Res 42: 29 50
Bruijn WC de, Breejen P den (1976) Glycogen, its chemistry and morphological appearance in the electron microscope. III. identification of the tissue ligands involved in the glycogen contrast staining reaction with the osminm(IV)-iron(II) complex. Histochem J 8:121-~42
Bruijn WC de, Buitenen JMH van (1980) X-ray microanalysis of aldehyde-fixed glycogen contrast-stained by OsVl.Fd I and OsV1.Ru TM complexes. J Histochem Cytochem 28:]242-1250
David-Ferreira KL, David-Ferreira JF (1980) Association between intermediate sized filaments and mitochondria in rat Leydig cells. Cell Biol Int Rep 4:655-662
Eddy EM (]974) Fine structural observations on the form and distribution of nuage in germ cells of the rat. Anat Rec 178:73]-757
Fawcett DW, Dym M (1974) A glycogen-rich segment of the tubuli recti and proximal portion of the rete testis in the guinea-pig. J Reprod Fertil 38:40]-409
Flickinger CJ (1969) Fine structure of the wolffian duct and cytodifferentiation of the epididy- mis in fetal rats. Z Zellforsch 96:344-360

Glycogen in Human Mesonephric Duct 453
Fukuda T (1976) Ultrastructure of primordial germ cells in human embryo. Virchows Archiv Cell Pathot 20:85-89
Gondos B, Conner LA (1973) Ultrastructure of developing germ cells in the fetal rabbit testis. Am J Anat 136:23~42
Gould SF, Bernstein MH (1979) Fine structure of fetal human testis and epididymis. Arch Androl 2:93-99
Hertig AT, Adams EC, MeKay DG, Rock J, Mulligan WJ, Menkin MF (1958) A thirteen-day human ovum studied histochemically. Am J Obstet Gynecol 76:1025-J040
Hoffer AP (1976) The ultrastructure of the ductus deferens in man. Biol Reprod 14:425-443 Holstein A-F (1969) Morphologische Studien am Nebenhoden des Menschen. Zwanglose Ab-
handlungen aus dem Gebiet der normalen und pathologischen Anatomie. Heft 20. Georg Thieme, Stuttgart
Karnovsky MJ (1971) Use of ferrocyanide-reduced osmium tetroxide in electron microscopy. J Cell Biol 284:146 Abstract
Lane BP, Europa DL (1965) Differential staining of ultrathin sections of Epon-embedded tissues for light microscopy. J Histochem Cytochem 13:579-582
Martino C de, Zamboni L (1966) A morphologic study of the mesonephros of the human embryo. J Ultrastruct Res 16: 399-472
Montagna W, Hamilton JB (1952) Histological studies of human testes. II. The distribution of glycogen and other HIO4-Schiff reactive substances. Anat Rec 112:237-244
Niemi M, Ikonen M (1965) Primordial germ cells in foetal and postnatal human testis. Ann Med Exp Fenn 43:23-28
Pelliniemi LJ (1972) Sex determination of human embryos by quinacrine fluoromicroscopy. Scand J Clin Lab Invest 29 (Suppl) 122:55
Pelliniemi LJ (1975a) Ultrastructure of gonadal ridge in male and female pig embryos. Anat Embryol 147 : 19-34
Pelliniemi LJ (1975b) Ultrastructure of the early ovary and testis in pig embryos. Am J Anat 144:89 112
Pelliniemi LJ (1976) Ultrastructure of the indifferent gonad in male and female pig embryos. Tissue Cell 8:163-174
Pinkerton JHM, McKay DG, Adams EC, Hertig AT (1961) Development of the human ovary - A study using histochemical technics. Obstet Gynecol 18:152-181
Rybicka K (1979) Glycosomes (protein-glycogen complex) in the canine heart. Ultrastructure, histochemistry and changes induced by acidic treatment. Virchows Arch Cell Pathol 30: 335-347
Rybicka K (1981) Simultaenous demonstration of glycogen and protein in glycosomes of cardiac tissue. J Histochem Cytochem 29:4-8
Streeter GL (1920) Weight, sitting height, head size, foot length and menstrual age of the human embryo. Carnegie Inst Wash Publ 55, Contrib to Embryol 11:143-159 + 6 charts
Tiedemann K (1971) Die Ultrastruktur des Epithels des Wolffschen Ganges und des Ductus deferens beim Schafembryo. Z Zellforsch 113:230-248
Venable JH, Coggeshall R (1965) A simplified lead citrate stain for use in electron microscopy. J Cell Biol 25:407-408
Wartenberg H, Holstein A-F, Vossmeyer J (1971) Zur Cytologie der pr~inatalen Gonaden- Entwicklung beim Menschen. II. Elektronenmikroskopische Untersuchungen tiber die Cy- togenese von Gonocyten und fetalen Spermatogonien im Hoden. Z Anat Entwickl-Gesch 134:165-185
Watson ML (1958) Staining of tissue sections for electron microscopy with heavy metals. J Biophys Biochem Cytol 4:475-478 +plates 226-229
White DL, Mazurkiewicz JE, Barrnett RJ (1979) A chemical mechanism for tissue staining by osmium tetroxide-ferrocyanide mixtures. J Histochem Cytochem 27:1084-1091
Zamboni L, Merchant H (1973) The fine morphology of mouse primordial germ cells in extragonadal locations. Am J Anat 137:29%335
Accepted November 7, 1983