morphine induced inhibition of the activities of accessory reproductive ducts in male rats
TRANSCRIPT

2008 Kyung Hee University Press 67
Oriental Pharmacy and Experimental Medicine 2008 8(1), 67-72
www.opem.org
OPEM
Morphine induced inhibition of the activities of accessory reproductive ducts
in male rats
Ramesh L Londonkar1, Sharangouda
2 and Saraswati B Patil
2,*
1Department of Biotechnology, Gulbarga University, Gulbarga-585106, Karnataka, India;
2Department of
Zoology, Gulbarga University, Gulbarga-585106, Karnataka, India
SUMMARY
Adult male albino rats were treated with 0.5 mg and 0.75 mg morphine/100 g body weightintraperitoneally for 30 days. All the animals were autopsied on 31st day. Epididymis and vasdeferens were dissected out, weighed and processed for histological and biochemical studies.Morphine has caused a reduction in the weight of epididymis and vas deferens in both the dosesof drug treated groups. The total cholesterol content is increased while protein, DNA and RNAcontents and epididymal sperm counts are decreased. The acid phosphatase content is decreased10.12 ± 0.11 in caput, 9.26 ± 0.30 in cauda of epididymis and in vas deferens 8.14 ± 0.15 in 0.5 mgtreated groups and in 0.75 mg treated rats shows 9.52 ± 0.27 in caput, 9.14 ± 0.18 in cauda ofepididymis and in vas deferens 7.84 ± 0.11is decreased, whereas alkaline phosphatase isincreased. The surface epithelial cell height of these ducts is reduced and secretory activity isinhibited with the disruption of epithelial cell projections. The gravimetric and histometricchanges of epididymis and vas deferens may be due to non-availability of androgens in morphinetreated rats.
Key words: Morphine; Epididymis; Vas deferens; Rat
INTRODUCTION
Morphine, a narcotic analgesic opiate has been
discovered by Sertuner in 1806. It binds to opioid
receptors in Central Nervous System (CNS) and
exerts its chief pharmacological actions on the
central nervous system (Russell et al., 1989). Morphine
administered to female rats during parturition
inhibits the uterine contractibility, disturbs the
normal maternal behavior in lactating rats (Russell
and Spears, 1984). Like other CNS influencing
drugs morphine is also known to inhibit the release
of follicular stimulating hormone (FSH) and luteinizing
hormone (LH) from the pituitary, acting through
hypothalamus blocking the neural stimulus to the
gonadotrophin releasing hormone (Blake, 1974;
Blake, 1978; Anderson et al., 1982). According to Jaffe
and Martin (1985), opioids act on the hypothalamus
and inhibit the release of gonadotrophin releasing
hormone (GnRH) and corticotrophin releasing factor
(CRF), thus decreasing the circulating concentrations
of LH, FSH, adrenocorticotrophic hormone (ACTH)
and β-endorphin.
Epididymis is an important site at which the
spermatozoa undergo progressive morphological
and physiological changes. It provides a favourable
milieu for acquiring the motility, fertilizing ability,
storage and survival of spermatozoa (Kasturi et al.,
1995). The vas deferens in addition to its role in
*Correspondence: Saraswati B Patil, Department ofZoology, Gulbarga University, Gulbarga-585106,Karnataka, India. E-mail: [email protected]
DOI 10.3742/OPEM.2008.8.1.067

2008 Oriental Pharmacy and Experimental Medicine 8(1), 67-72
68 Ramesh L Londonkar et al.
transport of spermatozoa is also involved in
maturation and survival of spermatozoa (Orgebin-
Crist, 1969; Orgebin-Crist and Danzo, 1975).
Therefore, the epididymis and vas deferens
instead of remaining mere passage ducts have
an important role in male reproduction and
fertility. Any drug influencing the CNS activities
can also modify the function of gonads and in
turn accessory organs. As the development and
function of epididymis and vas deferens
depends directly on gonadal activities, it is of
interest to study the effect of morphine on the
accessory reproductive organs.
MATERIALS AND METHODS
Healthy male albino rats of Wistar strain from
inbred colony, weighing 180 - 200 g, of 80 - 90 days
old were maintained at room temperature of 20 ±
28oC with lighting schedule of 12 h light and 12 h
darkness. They were maintained in individual
cages and divided in groups each containing six
animals and fed with balanced diet as described by
CFTRI (Central Food and Technological Research
Institute) Mysore, Karnataka, India and water ad
libitum.
The animals were divided into following groups
Group-I: Received 0.2 ml saline/100 g body weight
i.p. for 30 days and served as control.
Group-II: Received 0.5 mg morphine/100 g body
weight i.p. for 30 days in 0.2 ml saline.
Group-III: Received 0.75 mg morphine/100g body
weight i.p. for 30 days in 0.2 ml saline.
All the animals were sacrificed by cervical dislocation
after 24 h of the last injection. The epididymis and
vas deferens were dissected out immediately and
separated from adherent tissue, weighed up to the
nearest mg on electronic balance. Organs from one
side of each animal were fixed in Bouin’s fluid for
histological studies. They were embedded in paraffin,
sectioned at 5 µ, stained with Ehrlich hematoxylin
and Eosin. The micrometric measurements like
diameter of epididymis, its epithelial cell height
and lumen diameter of vas deferens were made
from randomly chosen 20 sections appearing
round at cross sections from each group using
ocular and stage micrometers. The sperm count
from cauda epididymis was done by using
haemocytometer (Kempinas and Lamano- Carvalho,
1987). Organs from other side were used for
biochemical estimations.
The total cholesterol (Peters and Vanslyke, 1946),
protein (Lowry et al., 1951), nucleic acids (Glick,
1985) and phosphatases (Toussaky and Shorr, 1953)
of epididymis and vas deferens were estimated
and “Student’s t-test” was employed for statistical
analysis.
RESULTS
Changes in the epididymis
Gravimetric changes (Table 1)
The weight of caput and cauda epididymis is
decreased significantly (P < 0.05) with 0.5 mg of
morphine treatment and highly significantly (P <
0.001) with 0.75 mg of morphine treatment when
Table 1. Effect of morphine on gravimetric and histometric changes of epididymis
GroupsWeight of epididymis
(mg/100 g body weight)Diameter (µm) Epithelial cell height (µm) Caudal
sperm count(million/ml)Caput Cauda Caput Cauda Caput Cauda
Control 395.84 ± 2.05 289.68 ± 2.12 2,645.24 ± 25.50 2324.45 ± 22.26 31.31 ± 1.40 32.45 ± 0.79 58.25 ± 4.34
0.5 mg 380.27 ± 3.01 277.52 ± 2.36 2,486.35 ± 22.54 2,238.81 ± 25.320 26.20 ± 21*0 29.83 ± 0.56 49.43 ± 5.00
0.75 mg 352.40 ± 6.10 263.45 ± 3.20 2,291.73 ± 29.80 2,214.27 ± 18.090 23.15 ± 1.38 27.75 ± 0.50* 46.72 ± 3.25**
Dose: 0.5 mg and 0.75 mg/100 g body weight. Duration: 30 days. Minimum 6 animals were maintained in eachgroup. Each datum represents the mean ± S.E. *P < 0.05, **P < 0.01.
Morphine induced inhibition of the activities of accessory reproductive ducts in male rats 69
2008 Oriental Pharmacy and Experimental Medicine 8(1), 67-72
compared to saline treated controls.
Biochemical changes (Table 3)
Protein
There is significant (P < 0.005) decrease in the protein
content of caput and cauda epididymis due to
0.5 mg morphine treatment, but it is highly
significant (P < 0.001) due to 0.75 mg morphine
treatment.
Cholesterol
The total Cholesterol content of caput and cauda
epididymis is increased significantly (P < 0.05) with
0.5 mg morphine treatment and highly significantly
(P < 0.01) with 0.75 mg morphine treatment when
compared to control group.
Nucleic acids
The DNA and RNA content of caput and cauda
epididymis is reduced almost significantly (P <
0.005 with 0.05) with 0.5 mg morphine treatment
and highly significantly (P < 0.001) with 0.75 mg
morphine administration.
Phosphatases
The acid phosphatase is decreased significantly
(P < 0.05) in caput and non-significantly in cauda
epididymis due to 0.5 mg morphine administration.
But highly significant reduction (P < 0.01) in
caput and significant (P < 0.005) reduction in
cauda epididymis is observed with 0.75 mg
morphine treatment. There is a significant increase
(P < 0.05) in alkaline phosphatase activity of caput
and non-significant increase in cauda epididymis
with 0.5 mg morphine treatment. Highly significant
(P < 0.01) increase in both caput and cauda
epididymis is observed after the administration of
0.75 mg morphine.
Histometric changes (Table 1)
The epithelial cells of saline treated control group
are tall, healthy, columnar and filled with secretary
material. But those of morphine treated cells are
smaller, disrupted and are with less secretary
material. The diameter of caput epididymis is
reduced almost significantly (P < 0.05) with 0.75 mg
morphine treatment.
Sperm morphology and number
The cauda epididymal sperms of normal rat shows
sickle shaped head and straight tailpiece. But in
morphine treated rats the sperms are abnormal as
their head region reduced and the tail is wrinkled
or coiled. A significant reduction (P < 0.05) in the
sperm count of cauda epididymis with 0.5 mg
morphine and highly significant (P < 0.01) reduction
with 0.75 mg morphine is observed.
Changes in vas deferens
Gravimetric changes (Table 2)
A non-significant decrease with 0.5 mg morphine
and significant decrease (P < 0.01) with 0.75 mg
morphine is observed in the weight of vas deferens.
Biochemical changes (Table 3)
Protein
There is non-significant decrease in the protein
content of vas deferens due to 0.5 mg morphine
treatment is observed but it is almost significant
(P < 0.05) due to 0.75 mg of morphine treatment.
Table 2. Effect of morphine on gravimetric and histometric changes of vas deferens
GroupsWeight of Vas Deferens(mg/100 g body weight)
Diameter (µm)Epithelial cell height
(µm)Diameter of lumen
(µm)
Control 83.19 ± 1.8 278.00 ± 22.13 28.00 ± 1.28 225.32 ± 2.61
0.5 mg 076.85 ± 1.99 251.62 ± 26.00 27.16 ± 1.24 236.20 ± 4.25
0.75 mg 0052.31 ± 1.60**0200.28 ± 15.10**
021.38 ± 1.05** 243.21 ± 3.55**
Dose: 0.5 mg and 0.75 mg/100 g body weight. Duration: 30 days. Minimum 6 animals were maintained in eachgroup. Each datum represents the mean ± S.E. *P < 0.05, **P < 0.01.
2008 Oriental Pharmacy and Experimental Medicine 8(1), 67-72
70 Ramesh L Londonkar et al.
Cholesterol
Though the total cholesterol content of vas
deferens is increased with both the doses, it is
significant (P < 0.01) only with 0.75 mg morphine
treatment.
Nucleic acids
The DNA and RNA content of vas deferens is
reduced in both the treated groups, but it is
significant (P < 0.05) only with 0.75 mg morphine
treatment.
Phosphatases
Acid phosphatase activity of vas deferens is
significantly (P < 0.01) reduced but the alkaline
phosphatase activity is significantly increased (P <
0.01) with 0.75 mg morphine treatment. Similar
changes are observed with 0.5 mg morphine
administration.
Histometric changes (Table 2)
Morphine administration has caused the reduction
in the diameter of vas deferens and its epithelial
cell eight, thereby increasing the lumen diameter.
Table 3. Biochemical changes in epididymis and vas deferens
Contents GroupsEpididymis
Vas deferensCaput Cauda
Protein (mg/100 mg)
Control 016.46 ± 0.22 015.80 ± 0.17 14.86 ± 0.12
0.5 mg 0 15.78 ± 0.16*013.82 ± 0.21* 13.60 ± 0.15*
0.75 mg 0 15.23 ± 0.09**0 12.56 ± 0.21** 12.82 ± 0.60*
Cholesterol (µg/100 mg)
Control 146.45 ± 2.21 100.05 ± 2.12 78.64 ± 2.21
0.5 mg 158.89 ± 3.13 0 124.56 ± 2.64** 82.51 ± 3.30
0.75 mg 178.58 ± 5.11*0 149.11 ± 20.20** 89.26 ± 1.00**
DNA (µg/100 mg)
Control 186.10 ± 2.91 162.30 ± 3.21 153.61 ± 3.60
0.5 mg 173.61 ± 4.00* 150.00 ± 3.11* 148.36 ± 8.42
0.75 mg 165.00 ± 2.21** 146.22 ± 3.20** 140.00 ± 3.82*
RNA (µg/100 mg)
Control 868.30 ± 7.52 791.31 ± 5.42 772.43 ± 4.01
0.5 mg 0835.01 ± 9.50* 758.60 ± 4.23* 759.62 ± 4.27
0.75 mg 0 824.10 ± 4.72** 735.02 ± 2.31** 753.32 ± 3.50*
Acid Phosphatase (µg/100 g)
Control 011.48 ± 0.43 10.42 ± 0.39 0 8.60 ± 0.13
0.5 mg 010.12 ± 0.11 9.26 ± 0.30 8.14 ± 0.15
0.75 mg 0 9.52 ± 0.27 9.14 ± 0.18* 7.84 ± 0.11**
Alkaline Phosphatase (µg/100 g)
Control 0 5.75 ± 0.10 5.33 ± 0.09 4.23 ± 0.19
0.5 mg 0 6.26 ± 0.12* 5.67 ± 0.14 4.51 ± 0.28
0.75 mg 0 6.64 ± 0.09** 6.21 ± 0.16** 4.96 ± 0.74*
Dose: 0.5 mg and 0.75 mg/100 g body weight. Duration: 30 days. Minimum 6 animals were maintained in eachgroup. Each datum represents the mean ± S.E. *P < 0.05, **P < 0.01.
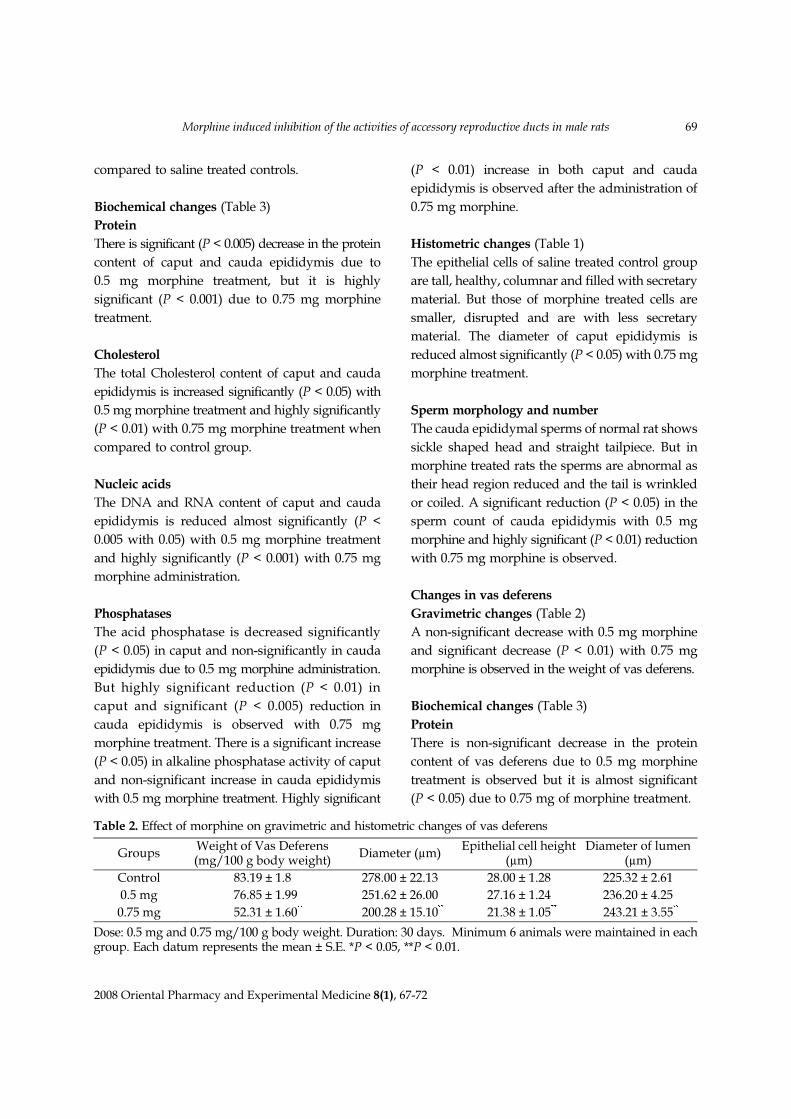
Fig. 1. Cross section of the cauda epididymis ofnormal adult rat showing tall healthy and secretaryepithelial cells. The surface villi are projectingtowards the lumen. The nuclei of the cells givepseudostratified appearance and spermatozoa arespermatozoa are clearly seen in the lumen. H & EStaining (400×). AC, Apical Cell; BC, Basal Cell; HC,Halo Cell; L, Lumen; N, Nucleus; PC, Principal Cell;SP, Spermatozoa; V, Villi.

Morphine induced inhibition of the activities of accessory reproductive ducts in male rats 71
2008 Oriental Pharmacy and Experimental Medicine 8(1), 67-72
These changes are significant only with 0.75 mg
morphine administration.
DISCUSSION
The male accessory reproductive organs play an
important role in the sperm maturation, motility
and formation of semen (Orgebin-Crist, 1969;
Hamilton, 1975; Martin and Sloan 1977). Spermatozoas
formed in the seminiferous tubules are transported
from the testis into epididymis and remain in the
duct system for varying periods of time before
being ejaculated (Gaddum and Glover, 1965).
During this period they acquire motility and
fertilizing capacity in the epididymis (Gaddum
and Glover, 1965).
Opioids by acting through the hypothalamus
inhibit the release of GnRH and CRF, thus decreasing
the circulating concentrations of LH, FSH and
ACTH (Reisine and Pasternak, 1996). The epididymis
and the accessory glands of reproduction depend
on testicular androgens (Price and William Ashman,
1961). Testosterone being an important factor for
the maintenance of accessory sex organs (Ojeda
and Urbanski, 1994). In turn the synthesis and
release of androgens depends on the availability of
pituitary gonadotrophins like FSH and LH/ICHS
(Connel and Eik-Nes, 1868; Johnson and Ewing,
1971; Hanson et al., 1973).
Morphine being a µ-opioid receptor against
inhibits the release of pituitary gonadotrophin
(Barraclough and Sawyer, 1955; Bruni et al., 1977),
which is essential for the androgen synthesis.
Therefore, reduction in the weight of epididymis
and vas deferens is observed in the present study.
As the androgen regulate the synthesis of DNA,
RNA and protein (Brooks, 1981; Duggan and
North, 1983; Martin, 1984). The decrease in the
DNA, RNA and protein contents after morphine
treatment supports the reduced androgen productivity
by the testis. Significant increase in the cholesterol
content of these ducts indicates the hampered
conversion of androgen from cholesterol, which is
dependent on the availability of pituitary
gonadotrophins.
Decrease in the acid phosphatase activity and
increase in the alkaline phosphatase activity after
morphine treatment may be due to decline in the
endogenous androgen production (Kasturi et al.,
1995). It is also one of the factors for the suppression of
spermatogenesis, which is evidenced by the
lowered sperm count in the cauda epididymis after
morphine treatment.
REFERENCES
Anderson K, Fuxe K, Eneroth P, Agnathi LF. (1982)
Involvement of cholinergic nicotine like receptors
as modulators of amine turnover in various
terminal systems and prolactin, LH, FSH and ICSH
secretion in castrated male rats. Acta Physiol. Scand.
116, 41-50.
Barraclough CA, Sawyer CH. (1955) Inhibition of the
release of pituitary the epididymis of the rabbit. J.
Exp. Zool. 63, 319-329.
Blake CA. (1974) Localization of inhibitory actions in
ovulation blocking drugs on release of Luteinizing
hormone in ovariectomized rats. J. Endocrinol. 95,
999-1005.
Blake CA. (1978) Paradoxical effects of drugs acting
on the central nervous system on the preovulatory
release of pituitary Luteinizing hormone in
proestrous rats. J. Endocrinol. 79, 319-326.
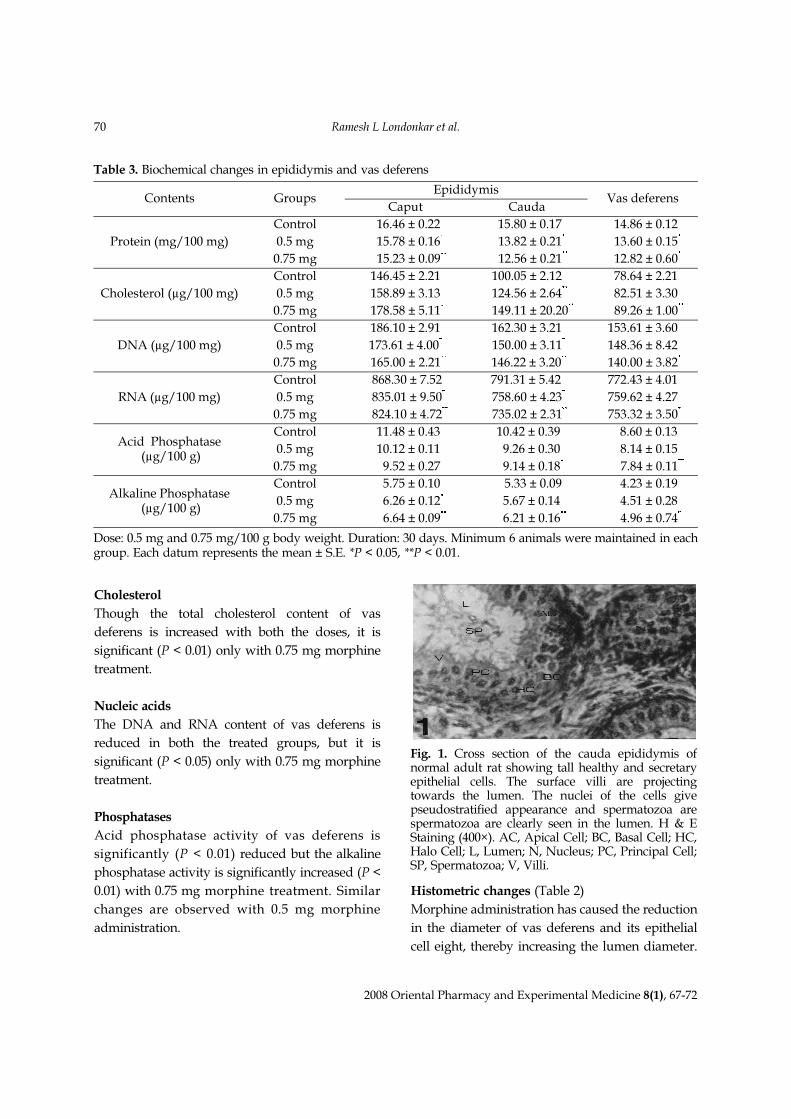
Fig. 2. Cross section of the cauda epididymis of adultrat treated with 0.75 mg morphine/100 g bodyweight showing epithelial cells with reduced heightand secretion. Surface villi are absent. The nuclei areshowing degenerative changes, spermatozoa are notdistinctly seen. H & E Staining (400×).

2008 Oriental Pharmacy and Experimental Medicine 8(1), 67-72
72 Ramesh L Londonkar et al.
Brooks DE. (1981) Effects of androgen on protein
synthesis and secretion in various regions of the rat
epididymis is analysed by two dimensional gel
electrophoresis. Mol. Cell. Endocrinol. 29, 255-261.
Bruni JF, Vanvut D, Marshall S, Meites J. (1977) Effect
of naloxone, morphine and metheonine enkepaline
on serum prolaction, Luteinising hormone, follicle
stimulating hormone, thyroid stimulating hormone
and growth hormone. Life Sci. 21, 461-466.
Connel GM, Eik-Nes KB. (1968) Testosterone
production by rabbit testis slices. Steroids 12, 507-521.
Duggan AW, North RA. (1983) Electrophysiology of
opioids. Pharmacol. Rev. 35, 219-282.
Gaddum P, Glover TD. (1965) Some relations of
rabbit’s spermatozoa to ligation of the epididymis.
J. Reprod. Fertil. 9, 119-126.
Glick D. (1958) Methods of biochemical analysis. Vol.
VI, pp. 10-17.
Hamilton DW. (1975) Structure and function of
epithelium lining and ductuli efferentes, ductus
epididymis and ducts deferens in the rat. In: Greep,
R.O., Astood E.B. (Eds), Handbook of physiology,
Section-7, Vol. 1, pp. 259-302.
Hanson V, Rensech E, Trygstud O, Torgeseno O,
Ritzen EM, French FS. (1973) FSH stimulation of
testicular androgen binding protein nature. New
Biol. 246, 56-58.
Johnson BJ, Ewing LL. (1971) Follicle stimulating
hormone and the regulation of testosterone
secretion in rabbit testis. Science 173, 635-640.
Kasturi MM, Manvannan B, Nazeer Ahmed, Parveen
DS, Pathan KM. (1995) Changes in epididymal
structural and function of albino rat treated with
Azadirachta indica leaves. Indian J. Exp. Biol. 33, 725-
729.
Kempinas WG, Lamano-Carvlho TL. (1987) A method
for estimation of concentration of spermatozoa in
the rat cauda epididymis. Laboratory Animals. pp.
154-156.
Lowry OH, Rosenbrough NJ, Farr NL, Randoll RJ.
(1951) Protein measurements with folic phenol
reagent. J. Biol. Chem. 193, 265-275.
Martin W R. (1984) Pharmacology of Opioids. Pharmacol.
Rev. 35, 283-323.
Martin WR, Sloan JW. (1977) Neuro pharmacology
and neurochemistry of subjective effects, analgesia,
tolerance and dependence produced by narcotic
analgesics. In: Handbook of experimental
pharmacology Vol. 45/I, Drug Addiction I, Morphine,
Sedative/Hypotonic and Alcohol Dependence
(Martin W.R., ed.). Springer Verlag, Berlin, 43-158.
Ojeda SR, Urbanski HF. (1994) Puberty in rat. In: the
of physiology reproductive. Second edition, Edted
by knobil and Neil J.D., Raven Press, New York.
Vol. II, 363-406.
Orgebin-Crist MC. (1969) Studies on the function of
the epidermis. Biol. Reprod. Suppl 1, 155-175.
Orgebin-Crist MC, Danzo BJ. (1975) Endocrine control
of the development and maintainance of fertilizing
ability in the epididymis. In: Handbook of physiology,
Section-7. (Eds. D.W. Hamilton and Greep. O). Am.
physiol. Soc. Washington D.C. 5, 319-338.
Peters JP, Vanslyke DD. (1946) Quantitative Clinical
Chemistry. Vol. I, Williams and Wilkins, Baltimore.
Price D, William Ashman HG. (1961) In: Young
William C. (Ed.), Sex and internal secretions, Vol. I,
Williams and Wilkins, Baltimore. pp. 366-448.
Reisine T, Pasternak. (1996) Opioid analgesic and
antagonistic. In: The pharmacological basis of
therapeutics (Eds. Hardman J.G., Limbird).
Motinoff PB, Ruddon RW, Gilman AG. (Mc grew
Hill. international edition New York) 9th edition.
521-556.
Russel JA, Spears N. (1984) Morphine inhibits
suckling induced oxytocin secretion in conscious
lactating rats, but also disrupts maternal behaviour.
J. Physiol. 346, 133-138.
Russel JA, Godsen RG, Humphreys EM, Cutting R,
Fitzsimons N, Johnson VS, Liddle S, Scott S,
Stirland JA. (1989) Interrruption of parturition in
rats by morphine: A result of inhibition of oxytocin
secretion. J. Endocrinol. 121, 521-536.
Taussaky HH, Sorr E. (1953) Microcolourometric method
for the determination of inorganic phosphorus. J.
Biol. Chem. 202, 675-682.