functional identity of the active sites of crustacean and viral thymidylate synthases
TRANSCRIPT

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Functional identity of the active sites of crustacean and viral thymidylate synthases
Aldo A. Arvizu-Flores a, Emmanuel Aispuro-Hernandez a, Karina D. Garcia-Orozco a,Alejandro Varela-Romero b, Elisa Valenzuela-Soto a, Enrique F. Velazquez-Contreras b,Arturo Rojo-Domínguez c, Gloria Yepiz-Plascencia a, Frank Maley d, Rogerio R. Sotelo-Mundo a,!a Centro de Investigación en Alimentación y Desarrollo (CIAD), A.C. Carretera a La Victoria Km 0.6, Hermosillo, Sonora, 83000, Mexicob Universidad de Sonora, Blvd. Rosales y Blvd. Luis Encinas, Hermosillo, Sonora, 83000, Mexicoc Universidad Autónoma Metropolitana, Unidad Cuajimalpa. Departamento de Ciencias Naturales, Pedro Antonio de los Santos 84, San. Miguel Chapultepec. Mexico D.F. 11850, Mexicod Wadsworth Center, New York State Department of Health, Albany, New York, 1220,1 USA
a b s t r a c ta r t i c l e i n f o
Article history:Received 15 May 2009Received in revised form 15 June 2009Accepted 16 June 2009Available online 25 June 2009
Keywords:Litopenaus vannameiShrimpPrawnThymidylate synthaseWhite spot syndrome virusAntifolate bindingDifferential inhibitionAntifolate
Thymidylate synthase (TS) catalyzes the synthesis of deoxythymidine monophosphate (dTMP), which is anessential precursor for DNA synthesis. The rationale underlying drug design is to identify compounds thatdifferentially inhibit a viral or parasite enzyme vs. the host homologue. We studied the TS of the white spotsyndrome virus (WSSV TS) and the corresponding TS from the host, the marine invertebrate shrimp Lito-penaeus vannamei. TS is the only de novo source of dTMP and is essential for host and viral DNA replication.To establish proof of principle, we cloned a full-length TS cDNA from the white shrimp L. vannamei (shrimpTS) that corresponds to a deduced sequence of 289 amino acids and over-expressed it to study inhibition ofboth shrimp and viral TSs. Steady-state kinetic parameters for both TSs are similar, and dissociation (Kd) orhalf maximal inhibitory concentration constants (IC50) did not show differential inhibition between thefolate analogues. Differences in their amino acid sequence are not re!ected in theoretical molecular modelsof both TSs, since both appear to have identical active sites. These results suggest that the eukaryotic TS activesite is very constrained into the functional residues involved in reductive methylation of 2"-deoxyuridine-5"-monophosphate (dUMP).
© 2009 Elsevier Inc. All rights reserved.
1. Introduction
Thymidylate synthase (TS, E.C. 2.1.1.45) catalyzes the synthesis of2"-deoxythymidine-5"-monophosphate (dTMP) from 2"-deoxyuri-dine-5"-monophosphate (dUMP) and 5,10-methylene-5,6,7,8-tetra-hydrofolate (MTHF) as cofactor. TS is the only de novo source of dTMPin many organisms from all kingdoms, including some viruses, and itsenzymatic inhibition becomes a hindrance for DNA replication(Carreras and Santi, 1995). TS is a strict dimer, since both active sitesare formed by residues of both subunits, and has a non-allostericligand-induced conformational change (Stroud and Finer-Moore,2003). Therefore, TS stands as an important model to understandthe structure–function relationships and as a paradigm for structure-directed drug design against proliferative diseases.
TS has been thoroughly studied in many species by means ofprotein crystallography, site directed mutagenesis, and kinetics(Finer-Moore et al., 2003; Stroud and Finer-Moore, 2003), although
it has not been examined much in invertebrates or non-modelorganisms. Since TS is present practically in all kingdoms, it wasanticipated that it would be present in crustaceans, although it has notbeen described before in them. The presence of TS in the white spotsyndrome virus genome, described as ORF54 (van Hulten et al., 2001)led to consideration of this gene as an antiviral target (Li et al., 2004).The white spot syndrome virus (WSSV) is a pathogen of cultivatedshrimp and an economic menace to aquaculture in developingcountries, so it is important to develop speci"c antivirals directedtowards the elimination of this virus. WSSV has a circular doublestranded DNA genome of approximately 300 kb, which encodes TS,among other gene products (Leu et al., 2009).
Differential targeting of enzymes as a drug target design approach iscommon in medicinal chemistry. Dihydrofolate reductase (DHFR)inhibitors that have antibacterial properties, such as trimethoprim, arestill used, while investigations on TS inhibitors against parasites arecurrently underway (Atreya et al., 2003). Even if the simple inhibition ofTS is not validated as a therapeutic approach, its inhibitors maypotentiate the action of other compounds targeted to thymidine kinaseor DNA polymerase, by reducing the deoxynucleotide pools thatcompete with these compounds, as shown in Scheme 1 (Panel A)(Prichard et al., 1993). Previous studies on WSSV TS show that it isexpressed and translated during viral infection (Li et al., 2004). Much of
Comparative Biochemistry and Physiology, Part C 150 (2009) 406–413
! Corresponding author. Aquatic Molecular Biology Laboratory, Centro de Investiga-ción en Alimentación y Desarrollo, A.C., Carretera a La Victoria Km 0.6, Hermosillo,Sonora, México 83000, Mexico, P.O. Box 1735. Tel.: +52 662 289 2400x352; fax: +52662 280 04 21.
E-mail address: [email protected] (R.R. Sotelo-Mundo).
1532-0456/$ – see front matter © 2009 Elsevier Inc. All rights reserved.doi:10.1016/j.cbpc.2009.06.008
Contents lists available at ScienceDirect
Comparative Biochemistry and Physiology, Part C
j ourna l homepage: www.e lsev ie r.com/ locate /cbpc

Author's personal copy
the research in this area is focused on the innate immunity against thevirus, including as an antiviral strategy the use of double stranded RNAto interfere with the expression of viral genes (RNAi) (Liu et al., 2009).
In order to assess the potential for WSSV TS inhibition as anantiviral strategy it is important to study nucleotide metabolism incrustaceans. Therefore, we cloned and over-expressed the Paci"cwhite shrimp (Litopenaeus vannamei) TS to address differentialinhibition studies against the white spot syndrome virus TS.
2. Materials and methods
2.1. Cloning and RACE
TS fromwhite shrimp L. vannamei (shrimp TS) was ampli"ed from acDNA library constructed with the Zap Express cDNA Synthesis System(Stratagene) (Romo-Figueroa et al., 2004). Degenerate primers TSFwandTSRv (see Fig. 1) were designed over conserved regions of aligned TSamino acid sequences obtained from GenBank. The 5" and 3" ends of theshrimp TS coding sequence were completed by RACE, combining thespeci"c primers TSFw1 and TSRv1 with universal primers located in thelibrary's vector. Complete cDNAsequencewas ampli"edusing theprimerset TSUTRFwand TSUTRRv from the cDNA library, thoroughly sequencedand deposited in GenBank under accession number FJ972199.
The cDNA ofWSSV TS was ampli"ed from a previously synthesizedcDNA library of WSSV-infected shrimp (Clavero-Salas et al., 2007)constructed with the SMART System (Clontech). The primers forWSSV TS were designed from the nucleotide sequences reported inGenBank (accession numbers AAK77723, AAL88992, and NP477589).
2.2. Recombinant expression and puri!cation
The amino acid sequence of both shrimp TS and WSSV TS wereoptimized by codon usage and obtained as synthetic genes (GeneArtAG). Both synthetic genes were cloned in pET11a (Novagen) into theNdeI and BamHI sites. Once veri"ed by DNA sequencing, the cloneswere used to transform E. coli BL21(DE3). The pET11a-shrimp TS clonewas grown in Luria Brothmedium at 37 °C in an orbital incubator untilthe A600nm reached 0.6; then, IPTG was added to a "nal concentrationof 0.5 mM and incubationwas continued for16 h. For the viral enzyme,the pET11a-wssvTS clone was grown in Terri"c Broth at 25 °C until theA600nm reached 0.6; then, IPTG was added to a "nal concentration of
0.2 mM and incubation was continued for 16 h. In both cases, the cellpellet containing the recombinant TS was obtained by centrifugationat 4500!g for 20 min at 4 °C and washed with 0.9% NaCl. For thepuri"cation of the recombinant TSs, the cell pellet obtained from 1 L ofculture was dissolved in 100 mL of 20 mM Tris–HCl, pH 7.5, plus 5 mMDTT and 0.5 mM PMSF, and then sonicated six times for 1 min in anice-bath. The cell debris was removed by centrifugation at 35,000!gfor 30 min at 4 °C. Nucleic acids were precipitated by adding 15 mL of5% streptomycin sulfate to each 100 mL of the supernatant fractionand removed by centrifugation after stirring for 10 min in an ice-bath.
TSs were isolated in the supernatant fraction between 65% and 80%saturation of ammonium sulfate. The precipitated protein was exten-sively dialyzed against 25 mMpotassium phosphate, pH 7.5, and loadedonto a Q-Sepharose column, on an AKTA chromatographer. The columnwas washed with 25 mM potassium phosphate, pH 7.5, plus 0.1 M NaCland eluted with a 50 mL linear gradient of 0.1 to 0.2 M NaCl in 25 mMpotassium phosphate, pH 7.5. The fractions containing pure TS werepooled and frozen at!80 ºC after precipitationwith ammoniumsulfate.The net yield was about 5 mg of pure TS from 1 L of culture. Since bothTSs were puri"ed without any af"nity tag, we validated the identity ofthe over-expressed proteins by mass spectrometry at the ProteomicsFacility of the Bio5 Institute, University of Arizona (Tucson, AZ, USA). AllMS/MS samples were analyzed using Sequest (ThermoFinnigan, SanJose, CA, USA; version 27, rev. 12) and X!Tandem (www.thegpm.org;version 2007.01.01.1). Scaffold (version Scaffold_2_03_01, ProteomeSoftware Inc., Portland, OR, USA) was used to validate MS/MS-basedpeptide and protein identi"cations.
2.3. Steady-state kinetics and inhibition
TS activity was measured using the spectrophotometric assay forthe reductive methylation of dUMP by MTHF (Wahba et al., 1962),with modi"cations (Maley et al., 1995). A unit of TS activity is de"nedas the amount of enzyme that converts one micromole of dUMP todTMP per min at 30 °C under the conditions of the assay. Proteinconcentration was determined by the bicinchoninic acid (BCA)method (Pierce). For the determination of kinetic parameters, initialvelocities were obtained with 0.25 µM of shrimp or WSSV TS andcombinations of various concentrations of MTHF (0–150 µM) anddUMP (0–20 µM). Analysis of steady-state kinetics was done with theGraFit software.
Scheme 1. Panel A. DNA replication pathway for the WSSV and drug targeting enzymes. Panel B. Chemical structure of substrates and inhibitors for thymidylate synthase.
407A.A. Arvizu-Flores et al. / Comparative Biochemistry and Physiology, Part C 150 (2009) 406–413

Author's personal copy
The half maximal inhibitory concentration (IC50), which is theconcentration of a compound that leads to the half maximal enzymevelocity, was calculated for two well known TS inhibitors: PDDF (10-propargyl-5,8-dideazafolate) and U89 ((S)-2-(5(((1,2-dihydro-3-methyl-1-oxobenzo[f]quinazolin-9 yl)methyl)amino)1-oxo-2-isoin-dolinyl) glutaric acid) (Dev et al., 1994)(see Scheme 1, panel B). Theanalysis was done using 0.25 µM of either shrimp or WSSV TS, 20 µMand 150 µM of dUMP and MTHF, respectively.
2.4. Intrinsic "uorescence
Intrinsic tryptophan!uorescencewas used tomeasure the antifolatebinding af"nity to both TSs (Sharma and Kisliuk, 1975; Anderson et al.,1999; Felder et al., 2002; Lovelace et al., 2007; Arvizu-Flores et al., 2009).Fluorescence was monitored using a QM-2003 !uorometer (PhotonTechnology International, Inc.) with a Xenon lamp as light source.Samples were excited at 295 nm and emission datawere collected from300 to 500 nm with accumulation of two readings. Measurements ofintrinsic !uorescence were performed with 1 µM of shrimp TS orWSSVTS, in potassium phosphate buffer, pH 7.5, containing 10 µM of dUMP.Fluorescence titration data were collected in the absence of and in the
presence of a three-fold excessof the antifolates PDDForU89. Eachpointfor titrationwasprepared independentlyandbaselineemission fromthefree ligand was subtracted to obtain the "nal emission spectrum.Dissociation constants (Kd) for the antifolates were estimated byanalysis of quenching data with the modi"ed Stern–Volmer plots aftercorrecting for inner "lter effects (Samworth et al.,1988; Lakowicz,1999;Arvizu-Flores et al., 2009).
2.5. Molecular modeling
Molecular modeling was done using the MOE program version2007–09 (Chemical Computing Group) by aligning the deducedamino acid sequence of shrimp or viral TS to the human TScoordinates 1HVY (Phan et al., 2001), as a crystallographic structuretemplate. The TS homodimer from shrimp and WSSV were modeledincluding ligands in the active site cavity from the template structureof human TS. Electrostatic calculations were done using PDB2PQR(Dolinsky et al., 2004) and the Adaptive Poisson–Boltzmann Solversoftware (APBS) as implemented in Pymol (Baker et al., 2001). Thestructural model "gures were made with MOE and Pymol 1.0
Fig. 1. cDNA and deduced amino acid sequence of the TS from white shrimp Litopenaeus vannamei. Primer regions for DNA ampli"cation are indicated by arrows. A typical poly A+
signal and a poly A+ tail are indicated in the sequence. The peptide fragments sequenced are indicated in bold letters and underlined.
408 A.A. Arvizu-Flores et al. / Comparative Biochemistry and Physiology, Part C 150 (2009) 406–413

Author's personal copy
(DeLano, 2002). All the molecular modeling and graphics programswere run under Mac OS X.
2.6. Phylogeny
PAUP was used to elucidate the phylogenetic relationship ofshrimp TS (GenBank accession no. FJ972199, this work) with other TSsequences reported in GenBank. Nucleotide and predicted amino acidsequences were analyzed. Protein-coding gene sequences wereinitially identi"ed using BLAST searches (Altschul et al., 1990) againstGenBank data, and subsequently compared to WSSV TS sequence
(AAL88992). Amino acid sequences were deduced using the standardgenetic code.
Phylogenetic relationships were determined using the completeamino acid sequences of 52 TS, including the shrimp sequence(this work) and WSSV TSs, as well as other invertebrates, viruses,and related groups including vertebrates, protists, fungi, plants,bacteria, and bacteriophages. All sequences were aligned usingClustal W (Thompson et al., 1994) and construction of phylogenetichypotheses from the dataset was done using maximum parsimony(MP). We used amino acid sequences to obtain the most parsi-monious tree with tree bisection-reconnection branch-swappingheuristic searches in PAUP, in which all characters were equally
Fig. 2. Alignment and comparison with structural deductions of TS amino acid sequences from different species. Upper line: H stands for !-helix, E stands for "-strand,and=represents a "-hairpin. Lower line: ! represents an amino acid in contact with ligand, # represents a catalytic amino acid.
409A.A. Arvizu-Flores et al. / Comparative Biochemistry and Physiology, Part C 150 (2009) 406–413

Author's personal copy
weighted and starting trees were obtained by 1000 random step-wise additions.
3. Results
3.1. Shrimp TS cDNA
A 260 bp fragment corresponding to shrimp TS was ampli"edfrom a cDNA library using degenerate primers, and the predictedamino acid sequence has up to 66% identity with vertebrate TSs(Ciesla et al., 1995; Du et al., 2006). Using RACE, we completed acDNA of 1123 bp that includes 5" and 3" UTR and corresponds to adeduced protein of 289 amino acids (Fig. 1). A typical polyadenyla-tion signal (Clavero-Salas et al., 2007) and a poly A+ tail were foundat 6 and 29 nucleotides downstream of the termination codon,respectively. The deduced amino acid sequence of shrimp TS wascompared to related TS sequences and an alignment of thesesequences is presented in Fig. 2. As indicated in previous reports,the TS amino acid sequence is highly conserved during evolution(Carreras and Santi, 1995), with a value of 63% sequence identitybetween shrimp and human, and 53% with E. coli, and 62% identitybetween shrimp and WSSV TS (Fig. 2).
Critical invariant residues were identi"ed in the deduced aminoacid sequence, such as the catalytic cysteine C171 (ecC1461), a lysineresidue important for folate binding K53 (ecK48) (Arvizu-Flores et al.,2008), arginine residues important for nucleotide binding R151'(ecR126')2(Strop et al., 1997) and R191 (ecR166) (Sotelo-Mundo et al.,2006), pyrimidine speci"city N202 (ecN177) (Hardy and Nalivaika,1992), among others.
3.2. Phylogenetic analysis
Phylogenetic relationships between the TSs of the white shrimpand WSSV were investigated using their amino acid sequences.Several amino acid sequences aligned were parsimony informative.Amino acid maximum parsimony tree (length=4743 steps, consis-tency index=0.557, retention index=0.507) produced trees withstrong statistical bootstrap support (100%) and included shrimp andviral TSs in the same clade, with high homoplasy. A strict consensus ofthe two shortest trees (with uninformative characters excluded)
1 ec stands for E. coli amino acid sequence numbering.2 ' stands for the sequence of the opposite monomer, this is that R127' is an arginine
residue located in the opposite active site.
Fig. 3. The strict consensus tree for TSs from the white shrimp and WSSV using amino acid sequences. The GenBank accession numbers are appended for each sequence betweenparentheses. 1Symbiotic of Acyrthosiphon pisum, 2symbiotic of Schizaphis graminum, and 3symbiotic of Baizongia pistaciae.
410 A.A. Arvizu-Flores et al. / Comparative Biochemistry and Physiology, Part C 150 (2009) 406–413
Author's personal copy
obtained from themaximum parsimony analysis of 254 aligned aminoacids for shrimp and WSSV TSs and the out group species and relatedspecies show that shrimp TS had a closest evolutionary relationship tothe TS from invertebrates and vertebrates (Fig. 3).
3.3. Kinetics of shrimp TS and WSSV TS
Michaelis–Menten kinetic parameters and inhibition analysis arecritical to investigate whether viral TS could be selectively inhibitedand could thus serve as an antiviral target. The values for KM dUMP andKM MTHF were found to be very similar between the shrimp TS andWSSV TS and other species (Table 1). Interestingly, the MTHF KM isabout one order of magnitude larger than the constant for thenucleotide, and the values previously reported for WSSV TS were verysimilar (Li et al., 2004). After recalculation of these constants, usingthe spectroscopic assay (Wahba and Friedkin, 1962; Maley et al.,1995), instead of the difference absorption method, the same trend inthe constant values was obtained. kcat values for shrimp andWSSV TSsare in the range of the Cryptosporidium hominis turnover number,which is considered one of the fastest TS (Atreya et al., 2003).
To address the differential inhibition of WSSV TS compared to theshrimp enzyme, the IC50 values were obtained using semi-log plots ofinhibitor concentration vs. initial velocity (Fig. 4). The data wasadjusted to a dose–responsemodel with non-linear "t, obtaining goodregression values (r2N0.97). Both enzymes were more sensitive toU89 compared to PDDF, but between shrimp and viral, there was nodifferential inhibition (Table 2).
3.4. Molecular modeling of shrimp TS
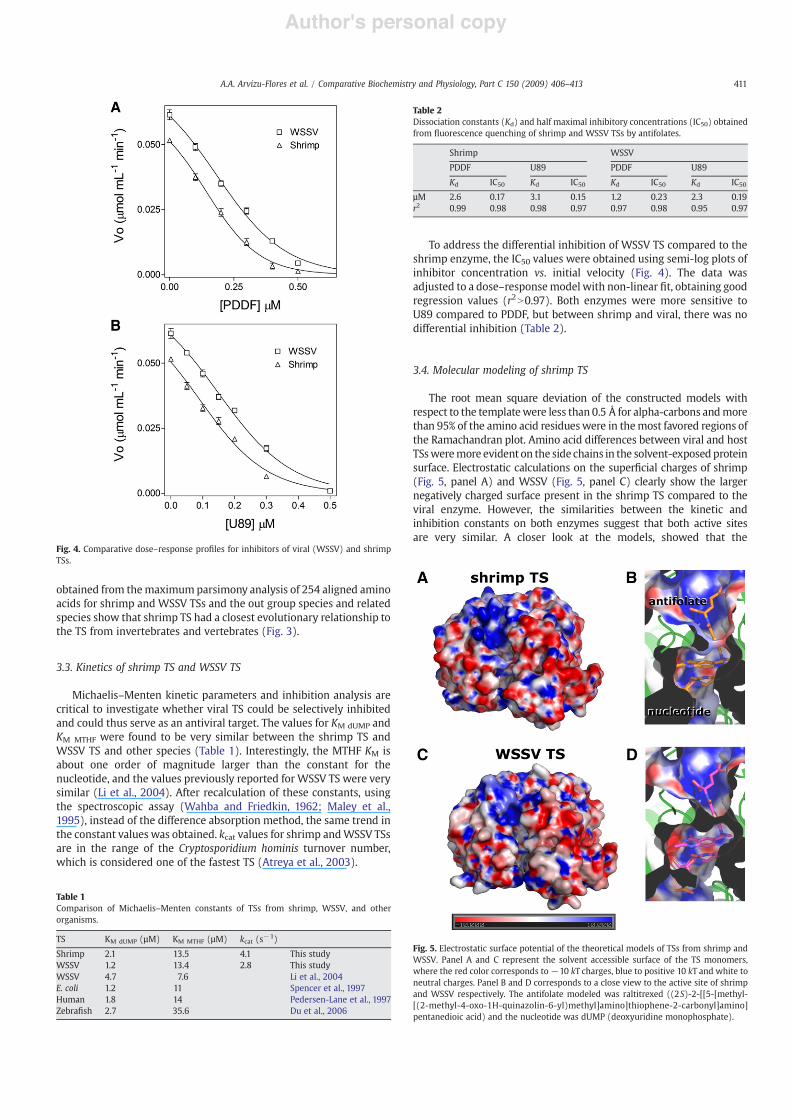
The root mean square deviation of the constructed models withrespect to the templatewere less than 0.5 Å for alpha-carbons andmorethan 95% of the amino acid residueswere in themost favored regions ofthe Ramachandran plot. Amino acid differences between viral and hostTSsweremore evident on the side chains in the solvent-exposed proteinsurface. Electrostatic calculations on the super"cial charges of shrimp(Fig. 5, panel A) and WSSV (Fig. 5, panel C) clearly show the largernegatively charged surface present in the shrimp TS compared to theviral enzyme. However, the similarities between the kinetic andinhibition constants on both enzymes suggest that both active sitesare very similar. A closer look at the models, showed that the
Fig. 4. Comparative dose–response pro"les for inhibitors of viral (WSSV) and shrimpTSs.
Table 1Comparison of Michaelis–Menten constants of TSs from shrimp, WSSV, and otherorganisms.
TS KM dUMP (µM) KM MTHF (µM) kcat (s!1)
Shrimp 2.1 13.5 4.1 This studyWSSV 1.2 13.4 2.8 This studyWSSV 4.7 7.6 Li et al., 2004E. coli 1.2 11 Spencer et al., 1997Human 1.8 14 Pedersen-Lane et al., 1997Zebra"sh 2.7 35.6 Du et al., 2006
Table 2Dissociation constants (Kd) and half maximal inhibitory concentrations (IC50) obtainedfrom !uorescence quenching of shrimp and WSSV TSs by antifolates.
Shrimp WSSV
PDDF U89 PDDF U89
Kd IC50 Kd IC50 Kd IC50 Kd IC50
µM 2.6 0.17 3.1 0.15 1.2 0.23 2.3 0.19r2 0.99 0.98 0.98 0.97 0.97 0.98 0.95 0.97
Fig. 5. Electrostatic surface potential of the theoretical models of TSs from shrimp andWSSV. Panel A and C represent the solvent accessible surface of the TS monomers,where the red color corresponds to!10 kT charges, blue to positive 10 kT and white toneutral charges. Panel B and D corresponds to a close view to the active site of shrimpand WSSV respectively. The antifolate modeled was raltitrexed ((2S)-2-[[5-[methyl-[(2-methyl-4-oxo-1H-quinazolin-6-yl)methyl]amino]thiophene-2-carbonyl]amino]pentanedioic acid) and the nucleotide was dUMP (deoxyuridine monophosphate).
411A.A. Arvizu-Flores et al. / Comparative Biochemistry and Physiology, Part C 150 (2009) 406–413

Author's personal copy
conformation and the sign of charges at the shrimp active site (Fig. 5,panel B) and WSSV (Fig. 5, panel D) are strongly conserved.
3.5. Binding of antifolates to TS
Calculation of dissociation constants (Kd) is important during thescreening of ligands as potential inhibitors or leads in drug design. Weapplied Stern–Volmer plots (Fig. 6) to calculate Kd by !uorescencequenching of the TS from shrimp and WSSV by PDDF and U89(Scheme 1, panel B). Some marginal differences between the Kd werefound, which are within one order of magnitude (Table 2). Such smalldifferences do not warrant any further study since a signi"cantcompetitive inhibition of the endogenous host enzyme may occur.
4. Discussion
Comparative analysis between species may aid in understandingthe evolution of protein structure because regions of conserved aminoacid sequences re!ect structural constraints on the molecule.Differential inhibition using TS as a target has been long postulatedin the literature (Maley et al., 1979), but lack of shrimp cell linesprecluded the direct study of antiviral compounds. Therefore, abiochemical approach to the problem was used to study thedifferential inhibition of recombinant shrimp and WSSV TSs.
The phylogenetic analysis of TS amino acid sequences showed thatWSSV TS is grouped out from the branch of shrimp TS (Fig. 3). This"nding is consistent with a previous report on the characterization ofWSSVTS,where the authors suggested that theviral enzymehad a closerrelationship to protozoa than to prokaryotes (Li et al., 2004). Based on aconsensus tree (Fig. 3), WSSV TS is apart from the TS-region frombifunctional DHFR-TS of kinetoplastids (Leishmania sp., Trypanosomasp.), herpesviruses, vertebrates, insects, and nematodes. Shrimp TS wasfoundmore related to a cnidaria (Nemastostella vectensis) and to a wasp(Nasonia vitripennis), but in this analysis crustaceans are strongly under-represented, since this is the "rst report for a marine invertebrate TS.
The phylogeny of double stranded viruses is more complex, andseveral non-occluded rod-shaped invertebrate viruses were leftoutside of the Baculoviridae family. Among them were the Heliothiszea virus 1 (HzV-1), the Orcytes rhinoceros virus (OrV), and the WSSV.OrV has been postulated as a Nudiviridae (Wang et al., 2007) and theWSSV in the Nimaviridae family (Genus Whispovirus) (Leu et al.,2009). Nonetheless, all those viral TSs share the high conservation ofthe catalytic and active site residues.
Many invariant and conserved residues in TS have a function incatalysis and substrate binding. An example of these constraints is thetwo fully conserved threonine (T51 and T52) residues before K53
(ecK48), which contacts MTHF via a water molecule (Fig. 2) (Maleyet al., 1982). These two Thr residues form a double "-turn thatpositions the interacting Lys residue in its optimal position at theactive site. The conservative mutation of K48Q in E. coli TS decreasedkcat and increased KM by several orders of magnitude respective to thewild type enzyme (LaPat-Polasko et al., 1990; Arvizu-Flores et al.,2008) without affecting the formation of the "-turns.
The presence of the invariant arginines in shrimp and WSSV TSs(ecR21, ecR126', ecR127', and ecR166) in the phosphate binding pocketand other residues, such as ecY209 or ecN229, may explain such similarvalues in the Michaelis–Menten kinetic constants for species asdivergent as human, zebra"sh, or E. coli. The similarity between thesteady-state kinetic constant and the equilibriumdissociation constantssuggests that the folded proteins have the same structure and functionalactive sites. Amino acid identity is not necessarily a good indicator forstructural or functional identity; since a 100-fold differential inhibitioncould be obtained using folyl-polyglutamates when comparing the T2bacteriophage with the host E. coli TS (Maley et al., 1979).
It must be considered that PDDF and U89 are competitive–activesite inhibitors (see Scheme 1, panel B), and, in fact, there aredifferences between shrimp and viral TS sequences (Fig. 5, panel Aand C). For example, the N-terminal !-helices in shrimp TS(MRHDEYQYLDLIRQIM) and in WSSVTS (MEGEHQYLNLVREIL) aredifferent and this region is structurally located within the TS dimerinterface. Therefore, non-competitive and acompetitive inhibitors arealternatives to use TS as an antiviral target. This means that an ideallead compoundwould speci"cally bind to the TS dimer interface of theviral TS andwould have low af"nity for the shrimp enzyme. The dimerinterface does not belong to the active site cavity but ligand binding atthis sitemay prevent the conformational change required for catalysis.That would be an example of acompetitive inhibition. Other studiessuch as molecular docking would require the three-dimensionalcrystal structure of both proteins. The biochemical approach usingrecombinant proteins and biophysical techniques to measure inhibi-tion and binding, combined with molecular modeling and dockingmay be the way forward in systems where cell lines are not available;such studies are required to address emerging pathogens such as thewhite spot syndrome virus. Aside of this, understanding the phylogenyand possible gene transfer between novel viruses is important as othercrustaceans or insects may be affected by emerging diseases.
Acknowledgments
This work was supported by the Consejo Nacional de Ciencia yTecnología (CONACYT) México D.F. grant 48991-Z to R. Sotelo-Mundo.A. Arvizu-Flores received a graduate scholarship from the CONACYT.Author A. Arvizu-Flores designed and performed experiments, ana-lyzed data and wrote the paper; E. Aispuro-Hernandez and K. Garcia-Orozco designed and performed experiments, and reviewed the paper;A. Varela-Romero analyzed data and reviewed the paper; E. Valenzuela-Soto, A. Rojo-Dominguez and E. Velazquez-Contreras discussed anddesigned experiments, contributed techniques and reviewed the paper,F. Maley and G. Yepiz-Plascencia designed experiments, wrote andreviewed the paper, R. Sotelo-Mundo designed experiments, analyzeddata, wrote and reviewed the paper. We thank Dr. Maria Islas-Osunaand Alonso Lopez-Zavala, M.Sc. for reviewing and suggesting improve-ments to the manuscript, and Monica Villegas, Salvador Carrasco andMonica Resendiz for technical support.
References
Altschul, S.F., Gish, W., Miller, W., Myers, E.W., Lipman, D.J., 1990. Basic local alignmentsearch tool. J. Mol. Biol. 215, 403–410.
Anderson, A.C., O'Neil, R.H., DeLano, W.L., Stroud, R.M., 1999. The structural mechanismfor half-the-sites reactivity in an enzyme, thymidylate synthase, involves a relay ofchanges between subunits. Biochemistry 38, 13829–13836.
Fig. 6. Modi"ed Stern–Volmer plots for the quenching of shrimp and WSSV TSs byantifolates. F0/#F vs. 1/[antifolate] plots for the titration of shrimp TS with PDDF (!)and U89 (") and wssvTS with PDDF (#) and U89 ($) show linearity, typical for protein!uorescence quenching by binding.
412 A.A. Arvizu-Flores et al. / Comparative Biochemistry and Physiology, Part C 150 (2009) 406–413

Author's personal copy
Arvizu-Flores, A.A., Sugich-Miranda, R., Arreola, R., Garcia-Orozco, K.D., Velazquez-Contreras, E.F., Montfort, W.R., Maley, F., Sotelo-Mundo, R.R., 2008. Role of aninvariant lysine residue in folate binding on Escherichia coli thymidylate synthase:calorimetric and crystallographic analysis of the K48Q mutant. Int. J. Biochem. CellBiol. 40, 2206–2217.
Arvizu-Flores, A.A., Velazquez-Contreras, E.F., Machi, L., Arreola, R., Maley, F., Sotelo-Mundo, R.R., 2009. Spectroscopic analysis of folate binding to thymidylate synthaseactive site. Spectroscopy Lett. 42, 142–146.
Atreya, C.E., Johnson, E.F., Irwin, J.J., Dow, A., Massimine, K.M., Coppens, I., Stempliuk, V.,Beverley, S., Joiner, K.A., Shoichet, B.K., Anderson, K.S., 2003. A molecular dockingstrategy identi"es Eosin B as a non-active site inhibitor of protozoal bifunctionalthymidylate synthase-dihydrofolate reductase. J. Biol. Chem. 278, 14092–14100.
Baker, N.A., Sept, D., Joseph, S., Holst, M.J., McCammon, J.A., 2001. Electrostatics ofnanosystems: application tomicrotubules and theribosome. Proc.Natl.Acad. Sci. U. S. A.98, 10037–10041.
Carreras, C.W., Santi, D.V., 1995. The catalytic mechanism and structure of thymidylatesynthase. Annu. Rev. Biochem. 64, 721–762.
Ciesla, J., Weiner, K., Weiner, R., Reston, J., Maley, G., Maley, F., 1995. Isolation andexpression of rat thymidylate synthase cDNA: Phylogenetic comparison withhuman and mouse thymidylate synthases. Biochim. Biophys. Acta 1261, 233–242.
Clavero-Salas, A., Sotelo-Mundo, R.R., Gollas-Galvan, T., Hernandez-Lopez, J., Peregrino-Uriarte, A.B., Muhlia-Almazan, A., Yepiz-Plascencia, G., 2007. Transcriptome analysisof gills from the white shrimp Litopenaeus vannamei infected with White SpotSyndrome Virus. Fish Shellf. Immunol. 23, 459–472.
DeLano, W.L., 2002. The PyMOL Molecular Graphics System. DeLano Scienti"c, SanCarlos, CA, USA.
Dev, I.K., Dallas, W.S., Ferone, R., Hanlon, M., McKee, D.D., Yates, B.B., 1994. Mode ofbinding of folate analogs to thymidylate synthase. Evidence for two asymmetric butinteractive substrate binding sites. J. Biol. Chem. 269, 1873–1882.
Dolinsky, T.J., Nielsen, J.E., McCammon, J.A., Baker, N.A., 2004. PDB2PQR: An automatedpipeline for the setup of Poisson–Boltzmann electrostatics calculations. NucleicAcids Res. 32, W665–667.
Du, C., Niu, R., Chu, E., Zhang, P., Lin, X., 2006. Sequence analysis and functional study ofthymidylate synthase from zebra"sh, Danio rerio. J. Biochem. 139, 913–920.
Felder, T., Dunlap, R.B., Dix, D., Spencer, T., 2002. Differences in natural ligand and!uoropyrimidine binding to human thymidylate synthase identi"ed by transient-statespectroscopic and continuous variationmethods. Biochim. Biophys. Acta 1597,149–156.
Finer-Moore, J.S., Santi, D.V., Stroud, R.M., 2003. Lessons and conclusions from dissectingthemechanismof a bisubstrate enzyme: thymidylate synthasemutagenesis, function,and structure. Biochemistry 42, 248–256.
Hardy, L.W., Nalivaika, E.,1992. Asn177 in Escherichia coli thymidylate synthase is amajordeterminant of pyrimidine speci"city. Proc. Natl. Acad. Sci. U. S. A. 89, 9725–9729.
Lakowicz, J.R., 1999. Principles of Fluorescence Spectroscopy. Kluwer Academic/PlenumPublishers, New York.
LaPat-Polasko, L., Maley, G.F., Maley, F., 1990. Properties of bacteriophage T4thymidylate synthase following mutagenic changes in the active site and folatebinding region. Biochemistry 29, 9561–9572.
Leu, J.H., Yang, F., Zhang,X., Xu,X., Kou,G.H., Lo, C.F., 2009.Whispovirus. In:VanEtten, J.L. (Ed.),Lesser Known Large dsDNAViruses. Springer-Verlag, Berlin, Heidelberg , pp. 197–227.
Li, Q., Pan, D., Zhang, J.H., Yang, F., 2004. Identi"cation of the thymidylate synthasewithin the genome of white spot syndrome virus. J. Gen. Virol. 85, 2035–2044.
Liu, H., Söderhäll, K., Jiravanichpaisal, P., 2009. Antiviral immunity in crustaceans. FishShell. Immunol. doi:10.1016/j.fsi.2009.02.009.
Lovelace, L.L., Gibson, L.M., Lebioda, L., 2007. Cooperative inhibition of humanthymidylate synthase by mixtures of active site binding and allosteric inhibitors.Biochemistry 46, 2823–2830.
Maley, F., Pedersen-Lane, J., Changchien, L., 1995. Complete restoration of activity toinactive mutants of Escherichia coli thymidylate synthase: evidence that E. colithymidylate synthase is a half-the-sites activity enzyme. Biochemistry 34,1469–1474.
Maley, G.F., Maley, F., Baugh, C.M., 1979. Differential inhibition of host and viralthymidylate synthetases by folylpolyglutamates. J. Biol. Chem. 254, 7485–7487.
Maley, G.F., Maley, F., Baugh, C.M., 1982. Studies on identifying the folylpolyglutamatebinding sites of Lactobacillus casei thymidylate synthetase. Arch. Biochem. Biophys.216, 551–558.
Pedersen-Lane, J., Maley, G.F., Chu, E., Maley, F., 1997. High-level expression of humanthymidylate synthase. Protein Expr. Purif. 10, 256–262.
Phan, J., Koli, S., Minor, W., Dunlap, R.B., Berger, S.H., Lebioda, L., 2001. Humanthymidylate synthase is in the closed conformation when complexed with dUMPand raltitrexed, an antifolate drug. Biochemistry 40, 1897–1902.
Prichard, M.N., Prichard, L.E., Shipman Jr., C., 1993. Inhibitors of thymidylate synthaseand dihydrofolate reductase potentiate the antiviral effect of acyclovir. AntiviralRes. 20, 249–259.
Romo-Figueroa, M.G., Vargas-Requena, C., Sotelo-Mundo, R.R., Vargas-Albores, F.,Higuera-Ciapara, I., Soderhall, K., Yepiz-Plascencia, G., 2004. Molecular cloning ofa beta-glucan pattern-recognition lipoprotein from the white shrimp Penaeus(Litopenaeus) vannamei: correlations between the deduced amino acid sequenceand the native protein structure. Dev. Comp. Immunol. 28, 713–726.
Samworth, C.M., Degli Esposti, M., Lenaz, G.,1988. Quenching of the intrinsic tryptophan!uorescence of mitochondrial ubiquinol–cytochrome-c reductase by the binding ofubiquinone. Eur. J. Biochem. 171, 81–86.
Sharma, R.K., Kisliuk, R.L., 1975. Quenching of thymidylate synthetase !uorescence bysubstrate analogs. Biochem. Biophys. Res. Commun. 64, 648–655.
Sotelo-Mundo, R.R., Changchien, L., Maley, F., Montfort, W.R., 2006. Crystal structures ofthymidylate synthase mutant R166Q: structural basis for the nearly complete lossof catalytic activity. J. Biochem. Mol. Toxicol. 20, 88–92.
Spencer, H.T., Villafranca, J.E., Appleman, J.R., 1997. Kinetic scheme for thymidylatesynthase from Escherichia coli: determination from measurements of ligandbinding, primary and secondary isotope effects, and pre-steady-state catalysis.Biochemistry 36, 4212–4222.
Strop, P., Changchien, L., Maley, F., Montfort,W.R.,1997. Crystal structures of amarginallyactive thymidylate synthase mutant, Arg 126->Glu. Protein Sci. 6, 2504–2511.
Stroud, R.M., Finer-Moore, J.S., 2003. Conformational dynamics along an enzymaticreaction pathway: thymidylate synthase, “the movie”. Biochemistry 42, 239–247.
Thompson, J.D., Higgins, D.G., Gibson, T.J., 1994. CLUSTALW: improving the sensitivity ofprogressive multiple sequence alignment through sequence weighting, position-speci"c gap penalties and weight matrix choice. Nucleic Acids Res. 22, 4673–4680.
van Hulten, M.C., Witteveldt, J., Peters, S., Kloosterboer, N., Tarchini, R., Fiers, M.,Sandbrink, H., Lankhorst, R.K., Vlak, J.M., 2001. The white spot syndrome virus DNAgenome sequence. Virology 286, 7–22.
Wahba, A.J., Friedkin, M., 1962. Direct spectrophotometric evidence for oxidation oftetrahydrofolate during enzymatic synthesis of thymidylate. J. Biol. Chem. 237,3794–3801.
Wang, Y., van Oers, M.M., Crawford, A.M., Vlak, J.M., Jehle, J.A., 2007. Genomic analysis ofOryctes rhinoceros virus reveals genetic relatedness to Heliothis zea virus 1. Arch.Virol. 152, 519–531.
413A.A. Arvizu-Flores et al. / Comparative Biochemistry and Physiology, Part C 150 (2009) 406–413