approaches for determining phytoplankton nutrient limitation
TRANSCRIPT

Approaches for determining phytoplankton nutrient limitation
John Beardall 1, *, Erica Young 2 and Simon Roberts1
1 Department of Biological Sciences, Monash University, PO Box 18, Clayton, Victoria 3800,Australia
2 School of Biology and Biochemistry, Queen’s University of Belfast, Belfast BT9 7BL, Northern Ireland, U.K.
Key words: Phytoplankton, nutrient limitation, chlorophyll fluorescence, moleculardiagnosis.
ABSTRACT
Aquatic primary productivity is frequently limited by the availability of nutrients. The ability toidentify factors limiting algal growth is of considerable importance to our understanding of theecology of aquatic plants and to water management practices. Methods used to identify limitingresources in the past have included a) analysis of nutrient availability, b) elemental compositionand cell quotas for various nutrients, c) bio-assays monitoring growth of test species or of naturalpopulations following nutrient enrichment and d) measurements of various physiological param-eters, such as enhancement of respiration and dark carbon fixation rates and perturbation of photo-synthetic rate following re-supply of nutrients.
In this paper we briefly review the merits and methodological limitations of these approachesfor the assessment of the nutrient status of algal populations. We discuss how an understanding ofbiochemical and metabolic changes induced by nutrient limitation has led to the development ofrapid and simple tools to monitor the nutrient status of aquatic plants and algae. In particular, wedescribe the use of transient changes in chlorophyll a fluorescence as a potential tool for rapidassessment of algal nutrient status and the development of molecular probes specific to nutrientlimited cells, such as flavodoxin as a diagnostic tool for Fe-limitation.
Introduction
Aquatic primary productivity, especially in surface waters, is frequently limited bythe availability of nutrients. Principally this involves limitations in the supply ofnitrogen and phosphate (Schindler, 1977; Wynne and Berman, 1980; Birch et al.,1981; Lean and Pick, 1981). Although N and P were originally believed to be limit-ing in marine and freshwater ecosystems respectively (Hecky and Kilham, 1988; butsee Elser et al., 1990; Dodds et al., 1993), more recent investigations have shown
Aquat.sci.63 (2001) 44–691015-1621/01/010044-26 $ 1.50+0.20/0© Birkhäuser Verlag, Basel, 2001 Aquatic Sciences
* Corresponding author, e-mail: [email protected]

Phytoplankton nutrient limitation 45
that some marine systems such as the North Pacific Sub-tropical Gyre and regionsof the Mediterranean Sea are, in fact, P limited (Karl, 1999; Krom et al., 1991). In“high nutrient, low chlorophyll” (HNLC) areas such as the equatorial Pacific andSouthern Ocean, iron has been shown to be an important limiting resource (Martinand Fitzwater 1988; Behrenfeld et al., 1996; Timmermans et al., 1998; Boyd et al.,1999), and even Zn has been implied as a limiting nutrient in some situations (Bru-land, 1989; Morel et al., 1994). The identification of nutrient limitation of phyto-plankton growth and the identity of the limiting factor(s) in a water body is of con-siderable importance to our understanding of the ecology of aquatic systems and towater management practices. It enables managers to draw up appropriate nutrientloading budgets for catchments and respond to possible perturbations on an in-formed basis. To have reliable and rapid techniques to do this is of considerablesignificance.
A number of techniques have been employed in the past in order to determinefactors limiting growth and production of algae. In this paper, it is our intention toreview the merits and drawbacks of a number of these approaches, including somewhich have developed from an increasing understanding of algal metabolism undernutrient limitation. We will also review more recently developed molecular ap-proaches and briefly describe a new, rapid and specific assay technique, based uponnutrient induced transient changes in chlorophyll a fluorescence emission fromnutrient-deficient microalgae.
Background – defining nutrient limitation
In dealing with nutrient limitation, it is necessary to address precisely what is meantby limitation of productivity and algal growth. Many ‘bioassay’ or ‘enrichment’ ex-periments, used in the past to investigate the factors limiting algal growth, fail todifferentiate between Liebig limitation i.e. limitation of the extent of growth (maxi-mum yield or standing stock of algae) and the limitation of instantaneous growthrate (Fig. 1). Studies based on measurements of Liebig limitation do not necessarilyreflect limitation by any particular nutrient at the time of sampling and at the pre-vailing cell density and species composition of the population. Rather they indicatesimply that should growth continue, one particular nutrient will become the limitingfactor. This distinction has major implications for the way that we approach studiesof nutrient limitation in natural populations (see Cullen et al., 1992). In oligotrophicwaters, a low biomass of rapidly dividing phytoplankton can be maintained byheavy grazing pressure combined with efficient recycling of nutrients in the watercolumn. In that case, the growth rate of individual cells is not limited by nutrientavailability, but the maximum population size is determined by the overall nutrientconcentration. However, the questions of biomass accumulation versus rate proces-ses may be particularly relevant with the current requirement to model and predicteffects of global climate change and the role of the oceans in biological draw-downof anthropogenic CO2 (Raven and Falkowski, 1999).
46 Beardall et al.
Long term manipulation and growth of phytoplankton
Bioassays with ‘test organisms’
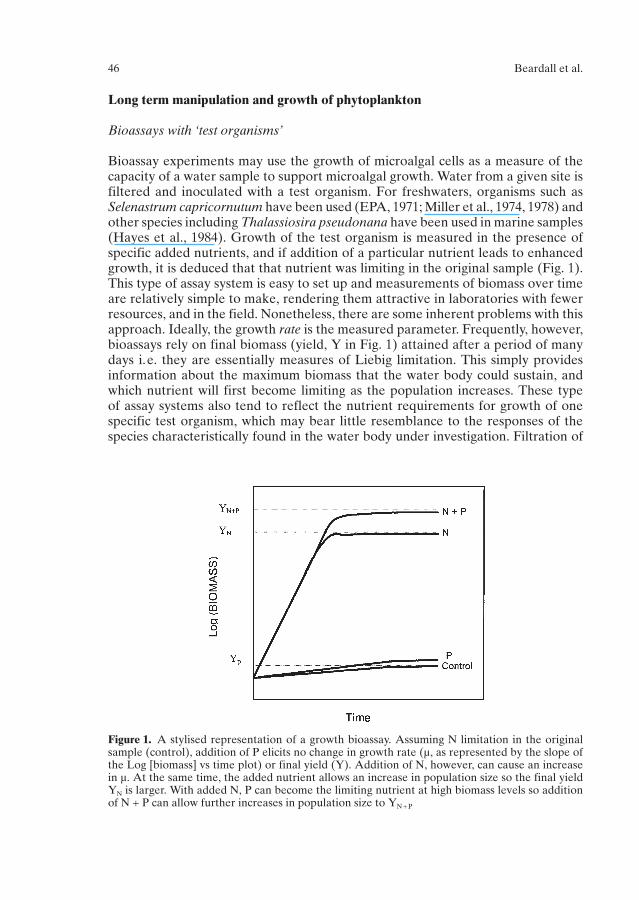
Bioassay experiments may use the growth of microalgal cells as a measure of thecapacity of a water sample to support microalgal growth. Water from a given site isfiltered and inoculated with a test organism. For freshwaters, organisms such asSelenastrum capricornutum have been used (EPA, 1971; Miller et al., 1974, 1978) andother species including Thalassiosira pseudonana have been used in marine samples(Hayes et al., 1984). Growth of the test organism is measured in the presence ofspecific added nutrients, and if addition of a particular nutrient leads to enhancedgrowth, it is deduced that that nutrient was limiting in the original sample (Fig. 1).This type of assay system is easy to set up and measurements of biomass over timeare relatively simple to make, rendering them attractive in laboratories with fewerresources, and in the field. Nonetheless, there are some inherent problems with thisapproach. Ideally, the growth rate is the measured parameter. Frequently, however,bioassays rely on final biomass (yield, Y in Fig. 1) attained after a period of manydays i.e. they are essentially measures of Liebig limitation. This simply providesinformation about the maximum biomass that the water body could sustain, andwhich nutrient will first become limiting as the population increases. These type of assay systems also tend to reflect the nutrient requirements for growth of onespecific test organism, which may bear little resemblance to the responses of the species characteristically found in the water body under investigation. Filtration of
Figure 1. A stylised representation of a growth bioassay. Assuming N limitation in the originalsample (control), addition of P elicits no change in growth rate (µ, as represented by the slope ofthe Log [biomass] vs time plot) or final yield (Y). Addition of N, however, can cause an increasein µ. At the same time, the added nutrient allows an increase in population size so the final yieldYN is larger. With added N, P can become the limiting nutrient at high biomass levels so additionof N + P can allow further increases in population size to YN + P

the water sample prior to inoculation with the test organism may also remove colloids and organic complexes that are frequently a source of nutrients, especiallyphosphorus (Wood and Oliver, 1995).
Enrichment experiments with natural populations
More commonly, enrichment experiments have been carried out with natural watersamples, relying upon the initial natural phytoplankton population as inoculum. A sample containing a natural population is “spiked” with various nutrients or witha nutrient mix lacking one particular nutrient. Carbon assimilation (Menzel andRyther 1961; Beardall et al., 1982) or growth (Cullen et al., 1992; Dodds et al., 1993)or the variable chlorophyll a fluorescence parameter Fv/Fm (Kolber et al. 1994; seebelow) is then determined over a period of time varying from several hours toseveral weeks or even longer (Healey, 1979; Hecky and Kilham, 1988). If produc-tivity (14C fixation or photosynthetic O2 evolution) or growth (measured as changesin biomass from chlorophyll concentration or cell numbers) is similar in sampleswithout a particular nutrient and in a control with all tested nutrient additions, it isusually deduced that the nutrient is not limiting. On the other hand, if a particularnutrient addition stimulates production and/or growth, that is indicative of thatnutrient being a limiting factor in the original water mass.
As with bioassays based on test organisms, experiments involving enrichment ofnatural phytoplankton populations are not without problems (see Esler et al., 1990).For instance, the response to bioassay enrichment experiments may be influencedby the species composition of the seeding/inoculum phytoplankton sample andaddition of one macronutrient may induce limitation of other nutrients. Forinstance, Menzel and Rhyther (1961) demonstrated a stimulatory effect of ironenrichment on 14C uptake by phytoplankton samples from the Sargasso Sea over24 h. However, over a longer period of 3 days, N and P additions were neededto produce a comparable stimulation. Chemical speciation is important and, espe-cially with trace metals, contamination and chelation effects can be significant (seeButler, 1998). Assessment of micronutrient deprivation on the basis of concentra-tions available in the water column requires trace-metal-clean methods for sam-pling water and relatively sophisticated analytical methods (Bruland, 1989; Twisset al., 2000). Chemical speciation of transition metals, and organic complexationof trace metals within the water column (particularly in freshwater environments,where the organic load, including humic substances, may be higher) complicatesthe relationship between measured concentrations and bioavailability for phyto-plankton uptake (see Anderson and Morel, 1982; Butler, 1998; McKay et al., unpublished). The complexity of trace metal chemistry has important implica-tions both for interpretation of micronutrient availability as well as for enrich-ment bioassays, where alleviation of micronutrient limitation has commonly beenachieved by addition of iron together with a chelating agent (usually EDTA) (e.g. Menzel and Rhyther, 1961; Peeters and Peperzak, 1990; Boyd et al., 1998). Furthermore, Muggli and Harrison (1996) showed the potential for inhibitory effects of EDTA on growth of phytoplankton. In enrichment bioassays, it can alsobe difficult to resolve trace metal limitation as the alleviation of one deficiency pre-
Phytoplankton nutrient limitation 47

cipitates another eg. alleviation of macronutrient limitation induces micronutrientdeficiency.
In addition, physical enclosure of a natural population can have deleteriouseffects upon physiological performance and result in changes in species compositionof the phytoplankton, and the population can be isolated from important nutrientsources such as sediments, particulate matter or recycling (Venrick et al., 1977; Hea-ley, 1979). Removal of zooplankton can also cause a relief from grazing pressure thatcan result in significant stimulation of growth even in unenriched control bottles(Cullen et al., 1992). If population growth is thus stimulated, the biomass mayincrease until the availability of some nutrient eventually becomes limiting (i.e. aLiebig limitation eventually sets in). Furthermore, if samples are incubated at ahigher photon flux than they might experience in situ, populations that were ori-ginally light-limited are exposed to conditions in which that limitation is alleviated,allowing a potential limitation by some nutrient to be expressed. Results obtainedfrom such experiments may bear little resemblance to the nutrient status of the ori-ginal plankton assemblage.
Furthermore, in enrichment bioassays based on short term 14C fixation or O2
exchange, addition of the limiting nutrient can actually cause short-term decreasesin the rate of photosynthesis (see Elrifi and Turpin, 1987) which are related to inter-actions between nutrient uptake and assimilation and other metabolic processes(see below).
An important development in the bio-assay approach to assessing nutrient limi-tation is the recent use of large-scale in situ enrichment experiments. These havebeen exemplified by the increasing number of Fe-enrichment trials, which haveidentified Fe-limitation in “high nutrient-low chlorophyll” (HNLC) regions of openocean. These Fe enrichment experiments will be mentioned below repeatedly in thecontext of specific parameters, but for a review of the general design of such expe-riments see Coale et al. (1998).
More immediate indices of nutrient status
Elemental ratios and macromolecular composition
Since the seminal work of Redfield (1958), the elemental composition of phyto-plankton, and even the composition of the water in which the organisms are grow-ing, has been used as a potential index of nutrient limitation. Phytoplanktoncells/populations on average assimilate C, N and P in the approximate ratio 106:16:1 (the Redfield ratio), so a water body with a N:P ratio of 30 is likely to lead toP limitation of algal growth whereas a ratio of 5 would imply N limited growth; inScenedesmus sp., P limitation switches to N limitation when the cellular N:P ratio isabout 30 (Rhee, 1974). Rees et al. (1995) measured the C:N assimilation ratio in afield study and used this to assess N limitation during the development of a springphytoplankton bloom. The use of elemental ratios of a water body can provideevidence for possible Liebig limitation of algal populations but are less useful forascertaining whether a given phytoplankton population is nutrient limited at a givenpoint in time. Furthermore, the reliance on Redfield ratios has been questioned by
48 Beardall et al.

Falkowski (2000) who pointed out that the ratio is far from constant and has beenembraced by aquatic ecologists as conveying more information than is warranted.This is further complicated by the difficulties in determining the availability ofnutrients for algal growth from a water sample, especially in turbid systems withhigh levels of particulates (White and Payne, 1980; White et al., 1985; Wood and Oli-ver, 1995). Methods for extracting phosphorus, for example, are complex and maynot reflect the availability of that element to phytoplankton (Bostrom et al., 1988);measurements of particle-associated exchangeable phosphorus show much bettercorrelation to growth bioassays than does total dissolved phosphorus (Oliver, 1993).The use of nutrient concentrations prevailing in a particular water body to deter-mine which nutrient is limiting also does not account for regeneration of nutrientswithin surface waters (which can vary significantly between water bodies), theautotrophic and heterotrophic community structure, and inorganic and organicnutrient inputs (i.e. variable f ratios) (Dugdale and Goering, 1967; Eppley andPeterson, 1979). In addition to concentrations within the water column, assessmentof the pool size and rate of turnover of a particular nutrient is needed to assess(potential) limitation (Harris, 1986). Sas (1989) has demonstrated the importanceof sediment in supporting phytoplankton growth, where there have been largereductions in phosphorus load in the water column.
The control of phytoplankton growth rate through other factors such as lightmay also complicate interpretation of elemental ratios for assessing nutrient status(see Goldman et al., 1979; Laws and Bannister, 1980; Tett et al., 1985), thoughgrowth at low light per se was not found to influence particulate C:N:P ratios in N-limited cultures of the chrysophyte Pavlova lutheri (Goldman, 1986) or C:N ratiosin the diatom Thalassiosira pseudonana (Thompson, 1999).
The elemental composition of algae reflects the macromolecular composition(protein, carbohydrate/lipid) composition of cells, which in turn reflects the in-tegration of the various processes involved in nutrient uptake and assimilation. Ele-mental ratios thereby provide an integrated measure of an algal cell’s more imme-diate nutrient history (Goldman et al., 1979; Healey, 1979; Fresnedo and Serra,1992). Thus nitrogen limitation usually results in diminished protein content andrelatively enhanced carbohydrate or lipid storage (Morris, 1981; Shifrin and Chis-holm, 1981; Ganf et al., 1986) while P limitation can also cause a shift in the pro-portions of protein, lipid and carbohydrate (Harris and Piccinin, 1983; Harrison etal., 1990; Theodorou et al., 1991; Reitan et al., 1994). Unfortunately, changes in ele-mental ratios of algal samples are not necessarily specific to the limiting nutrientand C:N ratios and N:P ratios can be influenced not only by N limitation but alsoby Si limitation or inorganic carbon availability (Table 1). In addition, measure-ments of elemental ratios or chemical composition of natural populations of phyto-plankton are prone to interference from debris and other microorganisms such asbacteria and microzooplankton that can be collected on the filters used to samplethe phytoplankton. One possible way to avoid interference from other organismsand debris in estimations of cell composition is the application of Fourier TransformInfrared (FTIR) spectroscopy. In combination with microscopy, this technique canbe used to determine macromolecular content of individual algal cells or colonies,thereby avoiding any signal from debris or other particles (see Beardall et al., 2001 –this issue). However, the changes in macromolecular composition of cells brought
Phytoplankton nutrient limitation 49
50 Beardall et al.
about by nutrient limitation are, as stated above, not sufficiently specific to allowFTIR techniques to identify which nutrient is limiting in a given situation.
Interactions between nutrient status and algal metabolism
Nutrient Uptake Kinetics
Nutrient limitation of algal growth results in changes to algal metabolism that canbe used as a measure of nutrient status. Given the extended time necessary for con-ducting bioassays and the additional problems, outlined above, with these and otherparameters such as elemental ratios, physiological analyses offer the possibility ofmore rapid estimates of nutrient status. The first step in assimilation of nutrientsinto organic matter is their uptake into algal cells and there have been numerousstudies investigating the effects of nutrient status on kinetics of nutrient uptake. Innatural waters the concentration of two of the major nutrients, nitrogen and phos-phorus, are often very low in comparison to measured values of half saturation constants (Km) for the inorganic ions (Harrison et al., 1996).
It is well established that algae respond to nutrient limitation by increasing up-take capacity and/or efficiency for the specific nutrient. For example, Gotham and
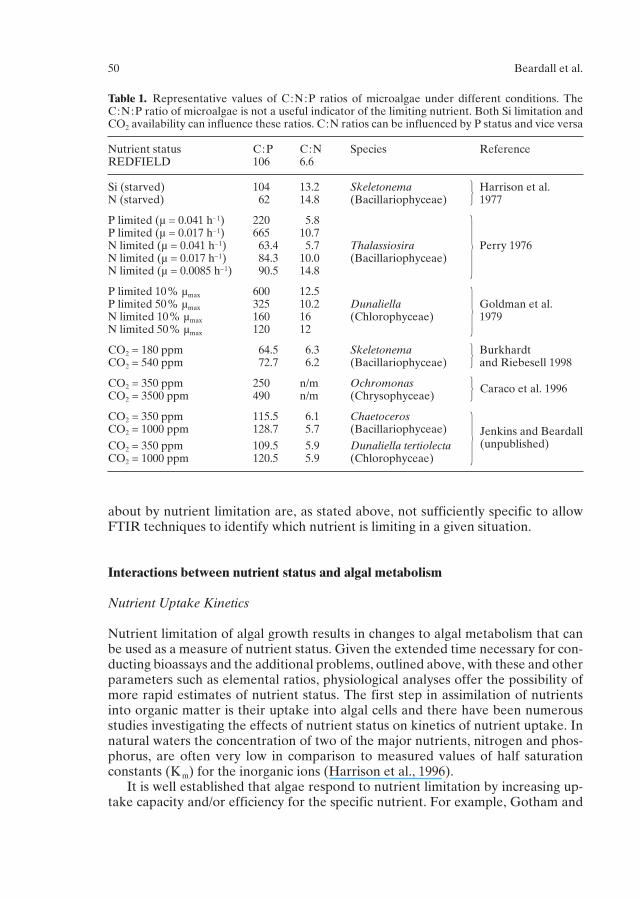
Table 1. Representative values of C:N:P ratios of microalgae under different conditions. TheC:N:P ratio of microalgae is not a useful indicator of the limiting nutrient. Both Si limitation andCO2 availability can influence these ratios. C:N ratios can be influenced by P status and vice versa
Nutrient status C:P C:N Species ReferenceREDFIELD 106 6.6
Si (starved) 104 13.2 Skeletonema Harrison et al. N (starved) 62 14.8 (Bacillariophyceae) 1977
P limited (µ = 0.041 h–1) 220 5.8P limited (µ = 0.017 h–1) 665 10.7N limited (µ = 0.041 h–1) 63.4 5.7 Thalassiosira Perry 1976N limited (µ = 0.017 h–1) 84.3 10.0 (Bacillariophyceae)N limited (µ = 0.0085 h–1) 90.5 14.8
P limited 10% µmax 600 12.5P limited 50% µmax 325 10.2 Dunaliella Goldman et al. N limited 10% µmax 160 16 (Chlorophyceae) 1979N limited 50% µmax 120 12
CO2 = 180 ppm 64.5 6.3 Skeletonema Burkhardt CO2 = 540 ppm 72.7 6.2 (Bacillariophyceae) and Riebesell 1998
CO2 = 350 ppm 250 n/m OchromonasCO2 = 3500 ppm 490 n/m (Chrysophyceae)
CO2 = 350 ppm 115.5 6.1 ChaetocerosCO2 = 1000 ppm 128.7 5.7 (Bacillariophyceae)
CO2 = 350 ppm 109.5 5.9 Dunaliella tertiolectaCO2 = 1000 ppm 120.5 5.9 (Chlorophyceae)
Caraco et al. 1996
Jenkins and Beardall(unpublished)�
��
���

Phytoplankton nutrient limitation 51
Rhee (1981) demonstrated that the maximal rate of phosphate uptake, for a rangeof freshwater cyanobacteria and microalgae, was enhanced as P-limited growth ratedecreased, and Graziano et al. (1996) reported that maximal uptake rates of Duna-liella tertiolecta under P-limitation were an order of magnitude greater than in P-replete cells. In most cases, increased uptake rate is associated with an increase inporter density (uptake sites) on the plasmalemma, rather than induction of isozym-es with different kinetic properties, so the affinity of uptake for P, as reflected inhalf-saturation constants, is unchanged (Graziano et al., 1996; Donald et al., 1997).The utilization of the inorganic nitrogen sources, nitrate, nitrite and ammonium, byphytoplankton in field conditions has been subject to intensive study, and the organ-ic N sources urea, amino acids and even purines and pyrimidines have been shownto support microalgal growth (Syrett, 1981). The kinetics of N uptake in relation tonitrogen deficiency is well characterised in a range of laboratory cultures of phyto-plankton species grown under both steady-state N limitation and N depletion (tostimulate a starvation condition) (reviewed by Collos and Slawyk, 1980; Goldmanand Glibert, 1983). N or P-deficient cells exhibit rapid uptake rates of the limitingnutrient immediately after it is re-supplied (Goldman and Glibert, 1983; Parslow etal., 1984b; Cochlan and Harrison, 1991). The maximal rate of uptake under theseconditions is significantly greater than the rate required to maintain the growth ofthe organism at its maximal rate (Parslow et al., 1984b) i.e. the cells exhibit “luxuryuptake”. This uncoupling between nutrient uptake and growth rates increases as Ndeficiency becomes more severe. Transient uptake rates vary with species, nitrogenhistory and nutrient status (Parslow et al., 1984a; Collos, 1980) although considera-ble species differences have been observed in the way N deficiency influencesnitrate or ammonium uptake (e.g. Dortch et al., 1982). Furthermore, the determi-nation of uptake rates can be complicated by a range of factors including incuba-tion conditions and time, non-stable substrates, recent presence of alternative Nsources (NO3
– vs NH4+) and different cellular N status (Flynn, 1998). It has been
established that the presence of ammonium can decrease nitrate uptake and/orassimilation capacity (e.g. Dortch et al., 1982, 1991) and, although the nature ofthis interaction is far from ubiquitous (e.g. Dortch, 1990), interactions betweenNO3
– and NH4+ sources for phytoplankton uptake have been observed in situ (e.g.
Harrison et al., 1996).
Secondary effects of uptake and incorporation
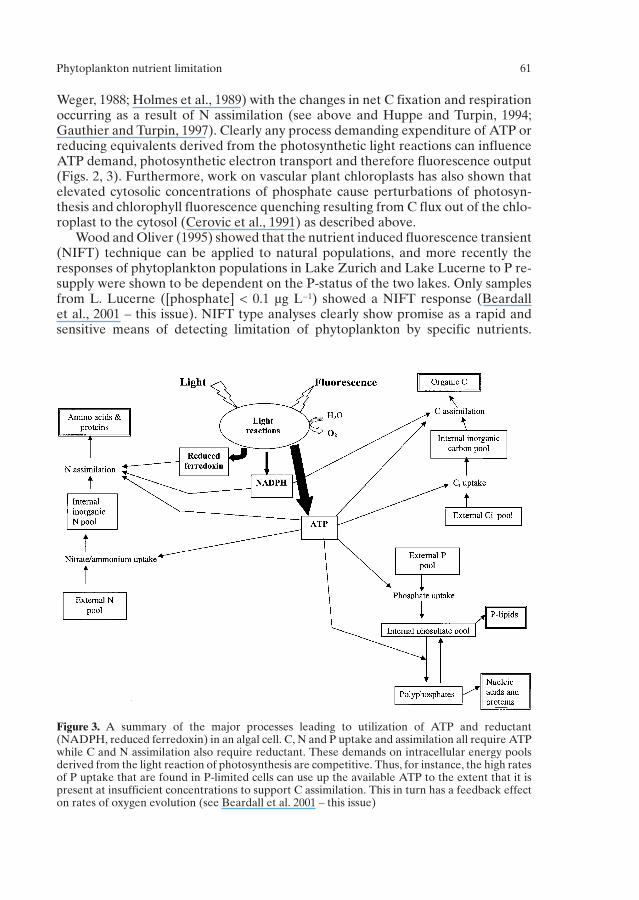
The uptake and assimilation of N and P both require the expenditure of energy asATP and/or reducing equivalents (Fig. 3). Many of the physiological parametersthat are used as indices of nutrient status are based on changes in cell function thatoccur during nutrient limitation and/or on the perturbations in energy metabolismthat occur when a limiting nutrient is re-supplied to cells. The ATP content of algalcells decreases during the onset of P or N deficiency and increases over a period of days following re-supply of the limiting nutrient (Healey, 1979). Re-supply of alimiting nutrient leads to ‘surge’ uptake of that nutrient (Parslow et al., 1984a; seeabove). Consequently ATP derived from the light reactions of photosynthesis canbe used for rapid uptake of the nutrient at the expense of C assimilation (Fig. 3). If

52 Beardall et al.
the ATP is derived from enhanced cyclic photophosphorylation, rather than linearelectron transport, photosynthetic O2 evolution will be diminished while nutrient isbeing taken up by the cells (Healey, 1979; Roberts, 1997; see also Beardall et al.,2001). The combination of enhanced nutrient uptake by nutrient limited cells, reduced O2 evolution and C assimilation rates and enhanced CO2 evolution rate following nutrient re-supply (Lean and Pick, 1981; Lean et al., 1982; Birch et al.,1986; Goldman and Dennett, 1986; Istvanovics et al., 1992) led Lean and Pick (1981)to use the ratio of C fixation (or O2 evolution) to nutrient uptake rate as an index oflimitation for natural populations of lake phytoplankton. Ratios above 100 weretaken as indicative of nutrient sufficiency while lower ratios suggested differentdegrees of limitation (Table 2). This index compared well to other parameters suchas turnover time for phosphate (Lean and Pick, 1981).
Similar observations on the suppression of photosynthetic C fixation were re-corded by Falkowski and Stone (1975) following NO3
– and NH4+ additions to N-
stressed phytoplankton cells, which they interpreted as a competition between Nassimilation and C fixation for ATP (Fig. 3). Such observations of short-term inhi-bition of C assimilation or O2 evolution confound interpretation of short-termenrichment experiments based on changes in 14C assimilation (see above).
Morris et al. (1971) first noted that N limited phytoplankton showed enhancedrates of carbon assimilation in the dark, and changes in net CO2 and O2 exchange.These phenomena have provided useful indices of N-limitation (Vincent, 1981;White et al., 1985; Elrifi and Turpin, 1987) but it was not until the elegant series ofstudies by Turpin and co-workers (see Huppe and Turpin, 1994 for a summary) thatthe biochemical basis of these observations was fully comprehensible. In brief, re-supply of NO3
– or NH4+ to N-stressed cells leads to a stimulation of inorganic N as-
similation and amino acid biosynthesis. To satisfy the demand for C– skeletons tosupport ammonium assimilation to glutamate, glycolysis and mitochondrial respi-ration are stimulated. Carbon to support this respiratory activity is diverted fromthe chloroplast, via the triose-phosphate shuttle, and is also supplemented by anap-lerotic carbon fixation via b-carboxylation. Since b-carboxylation is light indepen-dent, this is the basis for the observed enhancement of dark C fixation following N-addition. Export of triose sugars from the chloroplast depletes Calvin cycle C3 inter-mediates and inhibits recycling of the C acceptor RuBP. This results in a reduction
Table 2. Scale of P-limitation derived from the ratio of C assimilationto phosphate uptake capacity (mol mol–1) in natural populations ofphytoplankton from North American lakes (from Lean and Pick 1981).This scale of nutrient deficiency correlates with other estimates, basedon measurements such as P-turnover times in the populations
Nutrient status Molar ratio of C assimilation/ phosphate uptake capacity
Replete > 100Low deficiency 30–100Moderate deficiency 10–30Extreme nutrient stress < 10

Phytoplankton nutrient limitation 53
in Calvin cycle activity which affects the regeneration of electron acceptors (e.g.NADP+) that are necessary to maintain linear electron flow and photosynthetic O2
evolution. This dynamic also provides a rationale for the observed inhibition of O2
evolution following N re-supply to N-limited cells (see also Elrifi and Turpin, 1987).
Biochemical and Molecular approaches
A large body of research has characterized the physiological, biochemical andmolecular changes associated with acclimation of microalgal cells to nutrient stress.Although this work has predominantly concentrated upon a few key species, mani-pulated under laboratory culture conditions, it provides an essential theoreticalbackground to nutrient stress effects on algal metabolism, and has thus characte-rised changes which can be applied as diagnostic tools for assessing nutrient limita-tion in natural phytoplankton populations. The parameters discussed in this sectionhave in most cases been reported in combination with additional physiologicalassessment of algal photosynthetic competence, discussed in part above, though inrecent years measures of photochemical efficiency (see section on chlorophyll afluorescence below) have become more common.
The impact of macronutrient and Fe deprivation on cellular pigment ratios hasbeen reported in a number of phytoplankton taxa (e.g. Falkowski et al., 1989; Her-zig and Falkowski 1989; Cleveland and Perry, 1987; Sosik and Mitchell, 1991;Greene et al., 1992; Geider et al., 1993, 1998; van Leeuwe and Stefels, 1998). UnderN and P limitation, cellular chlorophyll content, and hence chl a :C ratio, decreases(e.g. Geider et al., 1998), though P limitation can provoke less rapid responses thanFe or N stress in some species (see La Roche et al., 1993). Geider et al. (1993)showed changes in chl c :chl a ratios in the diatom Phaeodactylum tricornutum underN, P and Fe deprivation, but found no significant changes in chl a :chl b ratio in N-limited cells of the chlorophyte Dunaliella tertiolecta (Geider et al., 1998), althoughin N-starved D. tertiolecta, chlorophyll b was preferentially lost over chlorophyll a(Young, 1999). Generally, N deprivation affects chlorophyll more strongly thancarotenoids, with elevated carotenoid to chlorophyll ratios a feature of N-stressedcells (e.g. Cleveland and Perry 1987; Falkowski et al., 1989; Herzig and Falkowski1989; Sosik and Mitchell, 1991). Heath et al. (1990) applied light absorption ratiosat 480 nm versus 665 nm to quantify the carotenoid:chlorophyll ratio as a potentialrapid index of phytoplankton nutritional status. They found absorption ratiosstrongly correlated with C:N ratio, independent of light and temperature effects, butthat these ratios varied between species. Although inter-specific differences wereless than between nutrient replete and nutrient stressed populations, field trials sug-gested that for mixed populations under mild N stress, this may not be as clear adiagnostic tool as desired. Geider et al. (1998) have more recently demonstratedcomplex interactions between the different carotenoid pools in D. tertiolecta in re-sponse to N and P stress. Furthermore, changes in pigment content, ratios and dis-tribution under N, P or Fe deprivation are related to changes in absorption crosssection areas of Photosystem II (PSII) and chlorophyll-specific absorption whichcan change during N, P or Fe limitation (Herzig and Falkowski, 1989; Greene et al.,1992; Geider et al., 1993).

Changes in Proteins
An important technique with diagnostic potential for assessing nutrient deprivationin phytoplankton is characterization of cell protein profiles. Single dimensional or2-dimensional polyacrylamide gel electrophoresis separates and resolves total pro-teins or polypeptides and the relative abundance and presence of particular proteinbands in response to nutrient deprivation can be identified. Several authors haveshown changes in ratios between relatively abundant chloroplast proteins such asthe large and small sub-units of Rubisco (LSU/SSU), the PSII reaction centre pro-tein D1, and light-harvesting chlorophyll-binding proteins (LHCI/II) under nutri-ent stress (e.g. Falkowski et al., 1989; Geider et al., 1993, 1998; La Roche et al., 1993;Graziano et al., 1996). Nitrogen deprivation in microalgae generally results in a pre-ferential repression of chloroplast protein synthesis (Plumley and Schmidt, 1989). Asuite of specific proteins have been identified in a range of species as being synthe-sised or lost in response to nutrient deficiency (e.g. La Roche et al., 1993; Li et al.,1998), yet relatively few have as yet been characterised to the extent of being use-ful as a priori markers for physiological limitation by nutrient stress.
A specific membrane protein (nrp1), identified in Emiliania huxleyi as regulatedby inorganic nitrogen limitation, was identified by Palenik and Koke (1995) using abiotinylated marker. In Synechococcus, novel membrane polypeptides have beenidentified under Fe, P and Mg limitation conditions (Scanlan et al., 1989) and a Syn-echococcus protein (PstS), induced under P-stress, showed homology with a peri-plasmic phosphate-binding protein of E. coli (Scanlan et al., 1993; Carr and Mann,1994). Graziano et al. (1996) reported that abundance of a unique stress proteinexpressed under P stress in D. tertiolecta correlated with rising C:P ratios and ele-vated Vmax for phosphate uptake induced under increasing phosphate limitation. LaRoche et al. (1993) provided further evidence that induction of specific proteinsindicated nutrient-limitation, but that the presence of these novel proteins is lessuseful for determining which nutrient is limiting in mixed assemblages. Diagnosingnutrient limitation on the basis of bands on a gel is rather tenuous. However theidentification and characterization of nutrient stress proteins in phytoplanktonpotentially enables highly specific immunological detection of nutrient stress mar-kers, as the successful identification of Fe-stress proteins in diatoms attests (see sec-tion below).
Dortch et al. (1985) reviewed a range of biochemical indices of nitrogen de-ficiency in natural phytoplankton populations, including deducing nitrogen suffi-ciency from high amino acid/protein ratios. They discussed variations in inorganicnitrogen storage, enzyme activity and protein/DNA ratios and their utility in under-standing nutrient deficiency in phytoplankton.
Specific enzyme markers for nutrient limitation
Alkaline Phosphatase
Extracellular alkaline phosphatase (AP) is expressed in a large range of phyto-plankton taxa in response to P-limitation. AP is highly stable in seawater, and thus
54 Beardall et al.

has potential as a indicator of inorganic P limitation, though it may reflect recent Pstatus, rather than the prevailing P status. Alkaline phosphatase catalyses the bre-akdown of exogenous organic-P compounds to yield utilizable inorganic P forms.The appearance of AP as an index of P-limitation was first proposed by Reichart etal. (1967) and extensively applied in freshwater systems by Berman (1970). Alkalinephosphatase activity (APA) has been detected in P-limited but not in nutrient-replete or N-limited phytoplankton (e.g. Sakshaug et al., 1984; Dyhrman and Pale-nik, 1997), and in freshwaters APA has been associated with orthophosphate deple-tion during spring phytoplankton blooms (Stevens and Parr, 1977). Significant APAhas been shown to correlate well with chlorophyll concentration during freshwaterphytoplankton blooms, and also correlated well with indices of P limitation (bioas-say, C:P ratios) and depletion of soluble reactive phosphorus (SRP) concentrationin the water column (Vrba et al., 1995; Rose and Axler, 1998). Rose and Axler(1998) showed a reduction in APA with P enrichment, whilst other nutrient addi-tions increased activity. APA is thus apparently a good index of phytoplankton Pstatus, though Rose and Axler (1998) recommended caution in interpreting APAdata in isolation as, when N-P co-limitation or secondary N limitation is present,APA activity may indicate P limitation without corresponding growth increasesoccurring under P enrichment. This re-iterates the strength of multiple assessmentmethods used simultaneously. Detection of significant APA may require relativelysevere P limitation in some species. Graziano et al. (1996) showed APA in severelyP-starved D. tertiolecta whilst APA was not detected in P-limited cells. In contrast,a distinctive protein marker of P-stress in D. tertiolecta was detected shortly afterthe onset of P-starvation (Graziano et al., 1996). APA detected in water samples caninclude a contribution from bacterial enzymes, though size-fractionation of watersamples can reduce this contamination. APA can also be released from cells and befound dissolved in water samples. The relationship between APA and the degree ofP stress varies amongst species (Graziano et al., 1996 and refs therein), and theremay be a diversity of P-limitation within mixed phytoplankton assemblages. Li etal. (1998) investigated the spatial distribution of APA in different plankton sizefractions and in aqueous fractions from the Red Sea. The majority of particulate-associated APA was in the picoplanktonic fraction, and there was significant corre-lation of APA with Synechococcus numbers, implying that APA responses to P-limitation may depend on species composition. González-Gil et al. (1998) recentlyreported a new in situ assay for APA in single cells, for detection by microscopy orin combination with flow cytometry and this may prove a useful advance in under-standing the nutrient status of individual species within a mixed population (see alsoZettler et al., 1996).
Enzymes of Nitrogen Assimilation
Fewer studies have looked at enzymes of nitrogen assimilation as indices of nutri-ent limitation, though changes in activities of these enzymes in relation to N-limita-tion, N-starvation and interactive effects of nitrogen source (NH4
+ vs NO3–) are well
understood (e.g. Goldman and Glibert, 1983). Nonetheless, activities of severalenzymes associated with N-assimilation, viz. nitrate reductase (NR), glutamine syn-
Phytoplankton nutrient limitation 55

thetase (GS) and NADPH-glutamate dehydrogenase (GDH) have been investi-gated as possible indicators of N-limitation (Dortch et al., 1979). Changes in GS andGDH activities have been associated with increasing N-limitation in (chemostat-grown) Chlorella stigmatophora (Everest et al., 1986). NR activity has been mea-sured in concert with 15N uptake, 14C fixation, pigment concentration and particu-late N, and related to the N concentration in the water column in marine systems(Blasco and Packard, 1974; Packard et al., 1978). Wynne et al. (1990) examined NRactivity together with nitrate and ammonium uptake capacity and cellular N:P ra-tios in freshwater phytoplankton, but suggested that NR activity was related tonutritional N status and nutrient pre-history rather than ambient conditions. Fur-thermore, conflicting responses to N-depletion have been observed in a range ofmarine microalgae with NR activity increasing (e.g. Kessler and Osterheld, 1970;Watt et al., 1992) or decreasing (Eppley et al., 1969, Berges et al., 1995) as depletionprogressed. Consequently, measurements of levels of enzymes of nitrogen assimila-tion are probably unlikely, in isolation, to provide a definitive tool for detecting N-limitation in phytoplankton populations.
Ferredoxin – Flavodoxin
Since the proposal that oceanic productivity can be limited by iron availability inhigh nutrient/low chlorophyll (HNLC) regions of the open ocean (Martin andFitzwater, 1988), there has been strong interest in assessing physiological Fe limita-tion of phytoplankton, with the redox catalysts ferredoxin and flavodoxin emergingas promising candidates for a specific assay. Ferredoxin (Fd) is a highly conservednon-haem iron-sulphur protein which plays a central role in energy metabolism ofcells by mediating electron transfer. A response common to prokaryotic organismsand eukaryotic phytoplankton subjected to iron-stress is the substitution of Fd withflavodoxin (Flv), a non-iron containing functional equivalent (La Roche et al.,1993). Although some prokaryotic and eukaryotic algae produce trace levels of Flvconsitutively (see Doucette et al., 1996), elevated expression of Flv in a range of dia-toms species tested is induced by iron-stress, leading La Roche et al. (1993) to sug-gest that Fd and Flv can be used as molecular probes of Fe-stress in the ocean. Fla-vodoxin induction is also found in members of the Prymnesiophyceae and Dino-phyceae (Erdner et al., 1999) and the cyanobacterium Nostoc (Hutber et al., 1977).The detection of Fd and Flv in phytoplankton has utilised predominantly immuno-logical techniques (La Roche et al., 1993, 1995, 1996; McKay et al., 1997, 1999; Tim-mermans et al., 1998), but also FPLC, and HPLC (Doucette et al., 1996 and refe-rences therein; Erdner and Anderson, 1999). In further characterising the broaderpotential for immunodetection of Fd/Flv expression to diagnose Fe-limitation inphytoplankton, McKay et al. (1999) detected Fd in a range of microalgae usingantibodies generated against Fd purified from the diatom Thalassiosira weissflogii.Abundant Fd was also detected by HPLC in Fe-replete cells of T. weissflogii butundetectable in Fe-limited cells (Doucette et al., 1996). McKay and co-workers(1999) showed that flavodoxin accumulation varied inversely with levels of ferre-doxin, which was not detectable in cells of the Fe-deficient diatom Phaeodactylumtricornutum. Erdner and co-workers proposed an Fd abundance ratio (Fd/(Fd +
56 Beardall et al.

Phytoplankton nutrient limitation 57
Flv)) as a quantitative index of Fe stress (Doucette et al., 1996; Erdner and Ander-son, 1999). Relative Fd and Flv can be related (McKay et al., 1999) to a frequentlyused parameter of physiological limitation, Fv/Fm (see below). However McKay etal. (1997) also showed that Flv can be expressed in the early stages of Fe-limitationand may be independent of growth rate and other indices of physiological limitationby Fe in both P. tricornutum and T. weissflogii.
Whilst most reports of Fd/Flv expression have concentrated on a few key diatomspecies in laboratory culture, the variable abundance of ferredoxin and flavodoxinin response to Fe stress has been demonstrated in field populations, in context within situ oceanic Fe-enrichment experiments (La Roche et al., 1995, 1996; Timmer-mans et al., 1998; Erdner and Anderson, 1999) and in freshwater systems (McKayet al., unpublished). La Roche et al. (1996) reported detection of flavodoxin inphytoplankton from HNLC regions but not from high iron regions of the sub-arcticPacific. Furthermore, relative flavodoxin abundance declined over 6 days followingFe enrichment despite large concurrent increases in chlorophyll, and Flv could bespecifically localised to chloroplasts of single diatom cells using immunofluores-cence microscopy (La Roche et al., 1996). In Fe-enrichment experiments, diatomsapparently respond most dramatically to Fe additions, which requires caution forinterpreting Flv and Fd data. Timmermans et al. (1998) showed an apparent in-crease in Flv abundance following minimal Fe enrichment in bottle experiments.However, in their experiments, there was a shift in species composition towards dia-toms, and the incomplete alleviation of Fe deficiency resulted in an increase in Flv,in the increasing diatom population, relative to a total protein marker. An impor-tant control for immunodetection of any specific protein marker is calibrationagainst total algal protein, or comparison with a non-specific immunomarker, typi-cally Rubisco LS (e.g. McKay et al., 1999).
Immunodetection of Fd and Flv provides a very sensitive tool for assessing Fedeficiency. However, limited antibody cross-reactivity allows restricted taxonomicrepresentation in reports of Flv expression under Fe stress (see McKay et al., 1999),which has also biased existing studies towards diatoms. Diatoms are major taxa ofecological relevance which are particularly iron-sensitive, however additional anti-sera would could allow Flv /Fd expression to be characterized in a wider range ofphytoplankters, and thereby broaden and strengthen Flv/Fd abundance as a dia-gnostic for Fe deficiency. Detection of Flv and Fd with HPLC (Doucette et al., 1998;Erdner and Anderson, 1999) avoids the phylogenic bias but is less sensitive thanimmunodetection, so requires a larger volume of cells, which could be difficult toachieve from HNLC areas of open ocean.
Other metal-containing enzymes can also potentially be used as markers fortrace metal deficiency. The role of trace metals in algal photosynthesis has beenrecently reviewed by Raven et al. (1999). Antisera generated against the Mn- andFe-containing isoforms of superoxide dismutase (Matta et al., 1992) could providean assay for Mn deficiency and an additional marker for Fe stress. The Cu-con-taining redox catalyst plastocyanin is similarly replaced with a non-Cu containingcompound in some picoplankton species that may become Cu deficient in fresh-waters where organic complexation may limit Cu bioavailability (Arudchandranand Bullerjahn, 1996).

58 Beardall et al.
Nucleic Acids
RNA/DNA ratios and DNA/Carbon ratios have been briefly examined to a limitedextent for their potential to estimate amount of living material in a natural phyto-plankton assemblage (e.g. Dortch et al., 1985). This technique has a major limita-tion in application to a mixed population, in that the pools of C and of nucleic acidscan be contributed inequitably from different taxa, so changing ratios could repre-sent changing species composition rather than a response to nutrient stress. How-ever nucleic acid ratios could potentially be used as an adjunct to estimating growthfrom algal yield in bioassay style experiments. The use of PCR technology mayenable detection of specific mRNA species associated with nutrient-regulatedgenes (e.g. see Leonhardt and Straus, 1992).
A problem common to assessment of nutrient deficiency using elemental analy-ses and enzyme activities and other physiological/biochemical markers is interfer-ence or uncertainty associated with presence of non-photoautotrophic organisms(microzooplankton, bacteria, viruses) and in phylogenetic variation. This source ofambiguity is not associated with use of methods specific to photosynthetic functioni.e. measuring productivity or chlorophyll (spectrophotometric or fluorescence), orgrowth characteristics with chlorophyll as a biomass determinant, or to the target-ing of specific algal taxa made possible with some immunological and molecularapproaches (see above).
Chlorophyll a Fluorescence
Light energy absorbed by chlorophyll in the light-harvesting antennae of the chlo-roplast thylakoid membranes has 3 major fates (Fig. 2). Most of the absorbedenergy is re-emitted as heat, some can be used to do photochemical work in drivingATP synthesis and NADP+ reduction, and a proportion is re-emitted as fluores-cence. These pathways compete for the light energy absorbed. Thus the proportionof energy used to do photochemical work is inversely related to the amount of fluo-rescence emission from chlorophyll a. The capacity for photochemical work is influ-enced by the nutrient status of cells and any damage to the photochemical appara-tus caused by photoinhibition. The instantaneous capacity for photochemical workalso depends upon reactions in the chloroplast, which utilize photosynthetically-derived ATP and reductant. Changes in rates of photochemical processes are thusreflected in changes in fluorescence output. Fluorescence therefore provides anextremely sensitive tool for examining energy metabolism in photosynthetic cellsand the interactions between processes such as C assimilation and nutrient assimi-lation that compete for electrons derived from the H2O-splitting reactions in PSII.
Chlorophyll a fluorescence can be readily measured by a range of fluorometers,provides a non-invasive assessment of the efficiency of photochemical conversionby PSII, and has become an increasingly powerful tool for investigations of nutrientlimitation in phytoplankton. The fluorescence parameter most commonly used isthe variable fluorescence expressed as Fv/Fm:
Fv/Fm = (Fm – F0)/Fm

Phytoplankton nutrient limitation 59
where Fm is the maximum fluorescence from a sample (i.e. when capacity forchemical work is saturated) and Fv is the difference between Fm and the minimumfluorescence F0, obtained after a period of dark adaptation, when the capacity forchemical work is maximal.
For details of fluorescence and its use in measuring photochemical efficiency thereader is referred to Maxwell and Johnson (2000). Although many early studiesobtained Fm values by blocking electron transport with the inhibitor DCMU (e.g.Geider et al., 1993), many investigations now use either Fast Repetition Rate (FRR;Kolber et al., 1988) or pulse amplitude modulated (PAM) fluorometers (Schreiber,1994). In these systems, Fm is obtained following an intense light flash which rapidlysaturates the photochemical apparatus.
Measurements of variable fluorescence
Variable fluorescence is extensively used to monitor effects of environmental stresson algal photosynthesis and applications include phytoplankton nutrient deficiency.The photosynthetic light harvesting and energy transduction apparatus is biochem-ically rich in N and Fe, and P plays a crucial role in cellular metabolism (as a com-ponent of membranes) and in energy transduction (as adenine nucleotide phospha-tes). Thus deficiency in these nutrients result in major impairment of photochemi-cal efficiency, Fv/Fm. Decreases in Fv/Fm have been observed during N-, P- or Fe-star-vation in a range of algal taxa (e.g. Kolber et al., 1988; Herzig and Falkowski, 1989;Sosik and Mitchell, 1991; Geider et al., 1993, 1998; La Roche et al., 1993; Greene etal., 1992; Berges et al., 1996; McKay et al., 1997; Young, 1999). Changes in variable
Figure 2. The fate of light energy absorbed by chlorophyll in algal cells. Light absorption causesexcitation of a chlorophyll molecule when an electron in the chlorophyll molecule is lifted to a highenergy level. As this electron returns to the low energy ground state, it releases energy, which canbe given off as heat or as fluorescence or can be used to power chemical work such as the assimi-latory processes shown. Clearly, these demands on the excitation energy are competitive and chan-ges in assimilation rates will be reflected in opposing changes in fluorescence or heat output

fluorescence in response to Si limitation and re-supply have also been observed indiatoms (Lippemeier et al., 1999). Fv/Fm recovers following re-supply of the limitingnutrient (Geider et al., 1993, La Roche et al., 1993; Young, 1999) and can be used asa basis for bioassays on natural populations of phytoplankton (e.g. Boyd et al., 1996).Following in situ Fe enrichment, Behrenfeld et al. (1996) reported increases in Fv/Fm
from ~0.25 to ~0.55 over 24 h which persisted over the 8 days in a HNLC region ofthe equatorial Pacific. Similar data have been reported following Fe enrichment inother HNLC regions of the equatorial Pacific (Kolber et al., 1994), the sub-arcticPacific (Boyd et al., 1996, 1998) and most recently, the Southern Ocean (Boyd et al.,1999). Thus measurements of Fv/Fm following re-supply of nutrients are an impor-tant index of nutrient status although, if carried out in samples isolated in contain-ers, they could be subject to similar problems to those outlined above for standardbioassays.
Nutrient induced fluorescence transients (NIFT)
Measurable changes in variable chlorophyll a fluorescence in response to nutrientlimitation or re-supply requires several days or hours respectively. Recent observa-tions of more rapid changes in chlorophyll a fluorescence emission, occurring overseveral minutes, in response to supply of a limiting nutrient offer the possibility ofalmost instantaneous assessment of phytoplankton nutrient status. These studieshave shown that addition of the limiting nutrient to N-limited or P-limited culturesof microalgae will induce characteristic changes in in vivo fluorescence output(Wood and Oliver, 1995; Beardall et al., 1996; Gauthier and Turpin, 1997; Roberts,1997; Young, 1999; Beardall et al., 2001). The qualitative nature of the response isdependent on the nutrient that is limiting and which nutrient form is re-supplied.Thus NH4
+ re-supply to N-limited cells elicits a small rise, followed by a significantdrop in fluorescence output, whereas micromolar NO3
– additions to the same cellscauses a significant rise in fluorescence which then returns to the initial value over~10 min (Young, 1999). PO4
3– addition to P-limited cells causes a drop of up to 40%in fluorescence emission which recovers to the initial level after several minutes(Beardall et al., 1996). Addition of a nutrient to nutrient replete cells or a non-limit-ing nutrient generally elicits no response, although Wood and Oliver (1995) re-ported some non-specific response to NH4
+ in P-limited cells of Microcystis aerugi-nosa. The duration, magnitude and rate of change in fluorescence emission aredependent on the concentration of nutrient added and the nutrient concentrationseliciting half-maximal responses are similar to those for uptake of N and P from thegrowth medium (Beardall et al., 1996; Roberts, 1997; Young, 1999). Furthermore, inthe case of phosphate re-supply, fluorescence transients persist until the externalphosphate is depleted, suggesting that the fluorescence transients are associated, atleast in part, with uptake processes (Beardall et al., 1996).
Observed changes in fluorescence output are mirrored by the well-documenteddecrease in O2 evolution following re-supply of limiting nutrients (Holmes et al.,1989; Beardall et al., 1996; Gauthier and Turpin, 1997; Young, 1999; Beardall et al.,2001). Turpin and co-workers associated quenching of chlorophyll fluorescenceoutput following NH4
+ re-supply to N-limited Scenedesmus minutum (Turpin and
60 Beardall et al.
Phytoplankton nutrient limitation 61
Weger, 1988; Holmes et al., 1989) with the changes in net C fixation and respirationoccurring as a result of N assimilation (see above and Huppe and Turpin, 1994;Gauthier and Turpin, 1997). Clearly any process demanding expenditure of ATP orreducing equivalents derived from the photosynthetic light reactions can influenceATP demand, photosynthetic electron transport and therefore fluorescence output(Figs. 2, 3). Furthermore, work on vascular plant chloroplasts has also shown thatelevated cytosolic concentrations of phosphate cause perturbations of photosyn-thesis and chlorophyll fluorescence quenching resulting from C flux out of the chlo-roplast to the cytosol (Cerovic et al., 1991) as described above.
Wood and Oliver (1995) showed that the nutrient induced fluorescence transient(NIFT) technique can be applied to natural populations, and more recently theresponses of phytoplankton populations in Lake Zurich and Lake Lucerne to P re-supply were shown to be dependent on the P-status of the two lakes. Only samplesfrom L. Lucerne ([phosphate] < 0.1 µg L–1) showed a NIFT response (Beardallet al., 2001 – this issue). NIFT type analyses clearly show promise as a rapid andsensitive means of detecting limitation of phytoplankton by specific nutrients.
Figure 3. A summary of the major processes leading to utilization of ATP and reductant(NADPH, reduced ferredoxin) in an algal cell. C, N and P uptake and assimilation all require ATPwhile C and N assimilation also require reductant. These demands on intracellular energy poolsderived from the light reaction of photosynthesis are competitive. Thus, for instance, the high ratesof P uptake that are found in P-limited cells can use up the available ATP to the extent that it ispresent at insufficient concentrations to support C assimilation. This in turn has a feedback effecton rates of oxygen evolution (see Beardall et al. 2001 – this issue)

62 Beardall et al.
However, current work in our, and other, laboratories has yet to thoroughly eva-luate the applicability of the technique to natural populations, especially at low cellnumbers.
Current Challenges and Ongoing Questions
Several issues continue to provide methodological problems and challenges forinterpreting assays of nutrient limitation in phytoplankton:
1. There are problems extrapolating from laboratory studies to an understandingof nutrient limitation on larger spatial scales. This is illustrated in an extremecase by extrapolations from measurements involving the small volumes asso-ciated with flow cytometry to ecosystem or population levels.
2. We need to address nutrient patchiness (see Turpin and Harrison, 1979) and hete-rogeneity in phytoplankton assemblages, species composition and nutrientswithin a single sample – flow cytometry offers a way of examining individualspecies/cells, but can only deal with very limited volumes.
3. How do we extract important data from a mixed sample? How differently dodifferent algal groups respond to nutrient enrichment? How can we define limi-tation on a population scale if, for example, diatoms are limited by silicate avail-ability, yet other groups are potentially limited by N or P?
4. Changes in species composition can occur with enrichment so that bioassayexperiments provide information about limitation of total primary productivitypotential and the capacity to increase primary productivity by alleviating differ-ent nutrient deficiencies rather than identifying factors limiting growth rates ofthe initial population. The growth rate of a phytoplankton population may onlybe marginally limited but enrichment might cause changes in species composi-tion as competition issues come into play.
Despite several decades of interest in assessing nutrient limitation in phytoplank-ton, there remain problems that are common to many of the methods discussed inthis review. However each of the methods is characterised by particular advantagesand limitations. The best approach is, therefore, not to rely exclusively on one tech-nique but to employ a suite of techniques and parameters to identify nutrient limi-tation in phytoplankton, which greatly strengthens interpretation of results. Thecontinued refinement of existing methods and identification of useful new indicesassessing nutrient limitation of phytoplankton growth requires ongoing input fromphysiologists to oceanographers and limnologists alike.
ACKNOWLEDGEMENTS
This contribution was presented as a keynote lecture at the 7th International GAP Workshop, heldon 9–17 Sept. 1999 in Zürich, Switzerland, and supported by the Swiss National Science Founda-tion (SNF), the Swiss Academies of Natural and Technical Sciences (SANW and SATW), theSwiss Society of Hydrology and Limnology (SGHL), by EAWAG, Zürich Water Supply and theUniversity of Zürich, as well as by Hoffmann-La Roche, Lonza, Novartis, Canberra Packard S.A,Millipore AG and Faust Laborbedarf AG. John Beardall is grateful for the support of the Aus-tralian Research Council for his laboratory’s studies on algal physiology. Erica Young acknowl-

edges support from a Monash Graduate Scholarship, Dr. John Berges for access to an extensivereference collection and Dr Michael McKay for access to unpublished material. Dr. Don Ander-son was kind enough to provide early access to papers at proof stage.Note added in proof: Hameed et al. have recently published an interesting comparison of methodsfor determining nutrient limitation in phytoplankton (Hameed, H.A., S. Kilinc, S. McGowan andB. Moss, 1999. Physiological tests and bioassays: aids or superfluities to the diagnosis of phyto-plankton nutrient limitation? A comparative study in the Broads and the Meres of England. Eur.J. Phycol. 34: 253–269.
REFERENCES
Anderson, M.A. and F.M.M. Morel, 1982. The influence of aqueous iron chemistry on the uptakeof iron by the coastal diatom Thalassiosira weissflogii. Limnol. Oceanogr. 27: 789–813.
Arundchandran, A. and G.S. Bullerjahn, 1996. Expression of the petE gene encoding plastocyaninin the photosynthetic prokaryote, Prochlorothrix hollandica. Biochem. Biophys. Res. Comm.226: 626–630.
Beardall, J., P. Foster, D. Voltolina and G. Savidge, 1982. Observations on the surface watercharacteristics in the western Irish Sea: July 1977. Est. Coast. Shelf Sci. 14: 589–598.
Beardall, J., E. Young and S. Roberts, 1996. Interactions between photosynthesis and the acquisi-tion of N and P in Dunaliella. In: Photosynthesis: from Light to Biosphere, vol. V (ed. P.Mathis), pp. 435–438. Dordrecht: Kluwer Academic Publishers.
Beardall, J., T. Berman, P. Heraud, M. Kadiri, B. Light, G. Patterson, S. Roberts, B. Sulzberger, E.Sahan, U. Uehlinger and B. Wood, 2001. A comparison of methods for detection of phosphatelimitation in microalgae. Aquatic Sciences 63: 107–121.
Behrenfeld, M.J., A.J. Bale, Z.S. Kolber, J. Aiken and P.G. Falkowski, 1996. Confirmation of ironlimitation of phytoplankton photosynthesis in the equatorial Pacific Ocean. Nature 383:508–511.
Berges, J.A., W.P. Cochlan and P.J. Harrison, 1995. Laboratory and field response of algal nitratereductase to diel periodicity in irradiance, nitrate exhaustion and the presence of ammonium.Mar. Ecol. Prog. Ser. 124: 259–269.
Berges, J.A., D.O. Charlebois, D.C. Mauzerall and P.G. Falkowski, 1996. Differential effects ofnitrogen limitation on photosynthetic efficiency of photosystems I and II in microalgae. PlantPhysiol. 110: 689–696.
Berman, T., 1970. Alkaline phosphatase and phosphorus availability in Lake Kinneret. Limnol.Oceanogr. 15: 633–674.
Birch, P.B., D.M. Gordon and A.J. McComb, 1981. Nitrogen and phosphorus nutrition of Clado-phora in the Peel-Harvey estuarine system, Western Australia. Bot. Mar. 24: 381–387.
Birch, D.G., I.R. Elrifi and D.H. Turpin, 1986. Nitrate and ammonium induced photosyntheticsuppression in N-limited Selensatrum minutum. Plant Physiol. 82: 708–712.
Blasco, D. and T.T. Packard, 1974. Nitrate reductase measurements in upwelling regions: I. Signi-ficance of the distribution off Baja California and Northwest Africa. Tethys 6: 239–246.
Bostrom, B., G. Persson and B. Broberg, 1988. Bioavailability of different phosphorus forms infreshwater systems. Hydrobiologia 170: 133–55.
Boyd, P.W., D.L. Muggli, D.E. Varela, R.H. Goldblatt, R. Chretien, K.J. Orians and P.J. Harrison,1996. In vitro iron enrichment experiments in the NE subarctic Pacific. Mar. Ecol. Prog. Ser.136: 179–193.
Boyd, P., J.A. Berges and P.J. Harrison, 1998. In vitro enrichment experiments at iron-rich and -poor sites in the NE subarctic Pacific. J. Exp. Mar. Biol. Ecol. 227: 133–151.
Boyd, P.W., J. La Roche, M., Gall, R. Frew, and R.M.L. McKay, 1999. The role of iron, light andsilicate in controlling algal biomass in Sub-Antarctic waters SE of New Zealand. J. Geophys.Res. – Oceans. 104: 13395–13408.
Bruland, K.W., 1989. Complexation of zinc by natural organic ligands in the central North Pacific.Limnol. Oceanogr. 34: 269–285.
Burkhardt, S. and U. Riebesell, 1998. CO2 availability affects elemental composition (C:N:P) ofthe marine diatom Skeletonema costatum. Mar. Ecol. Prog. Ser. 155: 67–76.
Phytoplankton nutrient limitation 63

Butler, A., 1998. Acquisition and utilization of transition metal ions by marine organisms. Science281: 207–210.
Caraco, N.F., J.J. Cole and R.A. Miller, 1966. Carbon dioxide and stoichiometry of phytoplankton.Suppl. EOS, Trans. Am Geophys Union Vol 76, No. 3.
Carr, N.G. and N.H. Mann, 1994. Oceanic cyanobacterial picoplankton. In: The Molecular Biologyof Cyanobacteria. (ed. D.A. Bryant), pp. 27–48. Kluwer, Dordrecht.
Cerovic, Z.G., Z.Vucinic and D.A. Walker, 1991. Photosynthetic oxygen evolution and chlorophyllfluorescence in intact isolated chloroplasts on a solid support: the influence of orthophosphate.Planta 184: 248–53.
Cleveland, J.S. and M.J. Perry, 1987. Quantum yield, relative specific absorption and fluorescencein nitrogen-limited Chaetoceros gracilis. Mar. Biol. 94: 489–497.
Coale, K.H., K.S. Johnson, S.E. Fitzwater, S.P.G. Blain, T.P. Stanton and T.L. Coley, 1998. IronEx-I, an in situ iron-enrichment experiment: Experimental design, implementation and results.Deep-Sea Res. II 45: 919–945.
Cochlan, W.P. and P.J. Harrison, 1991. Uptake of nitrate, ammonium, and urea by nitrogen-starvedcultures of Micromonas pusilla (Prasinophyceae): Transient responses. J. Phycol. 27: 673–679.
Collos, Y. and G. Slawyk, 1980. Nitrogen Uptake and Assimilation by Marine Phytoplankton. In:Primary Productivity in the Sea (ed. P.G. Falkowski), pp. 195–211. Plenum Press New York.
Collos, Y., 1980. Transient situations in nitrate assimilation by marine diatoms. 1. Changes in upta-ke parameters during nitrogen starvation. Limnol. Oceanogr. 25: 1075–1081.
Cullen, J.J., X. Yang and H.L. MacIntyre, 1992. Nutrient limitation of marine photosynthesis. In:Primary Productivity and Biogeochemical Cycles in the Sea (ed. P.G. Falkowski and A.D.Woodhead), pp. 69–88. Plenum Press New York.
Dodds, W.K., E.A. Strausss and R. Lehmann, 1993. Nutrient dilution and removal bioassays toestimate phytoplankton response to nutrient control. Arch. Hydrobiol. 128: 467–481.
Donald, K.M., D.J. Scanlan, N.G. Carr, N.M. Mann and I. Joint, 1997. Comparative phosphorusnutrition of the marine cyanobacterium Synechococcus WH7803 and the marine diatom Tha-lassiosira weissflogii. J. Plankton Res. 19: 1793–1813.
Dortch, Q., 1990. The interaction between ammonium and nitrate uptake in phytoplankton Mar.Ecol. Prog. Ser. 61: 183–201.
Dortch, Q., S.I. Ahmed and T.T. Packard, 1979. Nitrate reductase and glutamate dehydrogenaseactivities in Skeletonema costatum as measures of nitrogen assimilation rates. J. Plank. Res. 1:169–185.
Dortch, Q., J.R. Clayton, S.S. Thoreson, S.L. Bressler and S.I. Ahmed, 1982. Response of marinephytoplankton to nitrogen deficiency: decreased nitrate uptake vs enhanced ammonium up-take. Mar. Biol. 70: 13–19.
Dortch, Q., J.R. Clayton, S.S. Thoresen, J.S. Cleveland, S.L. Bressler and S.I. Ahmed, 1985. Nitro-gen storage and use of biochemical indicies to assess nitrogen deficiency and growth rate innatural phytoplankton populations. J. Mar. Res. 43: 437–464.
Dortch, Q., P. A. Thompson and P. J. Harrison, 1991. Short-term interaction between nitrate andammonium uptake in Thalassiosira pseudonana: effect of preconditioning nitrogen source andgrowth rate. Mar. Biol. 110: 183–193.
Doucette, G.J., D.L. Erdner, M.L. Peleato, J.J. Hartman and D.M. Anderson, 1996. Quantitativeanalysis of iron-stress related proteins in Thalassiosira weissflogii: measurement of flavodoxinand ferredoxin using HPLC. Mar. Ecol. Prog. Ser. 130: 269–276.
Dugdale, R.C. and J.J. Goering, 1967. Uptake of new and regenerated forms of nitrogen in primaryproductivity. Limnol. Oceanogr. 12: 196–206.
Dyhrman, S.T. and B.P. Palenik, 1997. The identification and purification of a cell-surface alkalinephosphatase from the dinoflagellate Prorocentrum minimum (Dinophycaeae). J. Phycol. 33:602–612.
Elrifi, I.R. and D.H. Turpin, 1987. Short-term physiological indicators of N deficiency in phyto-plankton: a unifying model. Mar. Biol. 96: 425–432.
Environmental Protection Agency, USA, National Eutrophication Research Program. 1971 Algalassay program: bottle test.
Eppley, R.W. and B.J. Peterson, 1979. Particulate organic matter flux and planktonic new produc-tion in the deep ocean. Nature 282: 677–680.
64 Beardall et al.

Eppley, R.W., J.L. Coatsworth and L. Solorzano, 1969. Studies of nitrate reductase in marinephytoplankton Limnol. Oceanogr. 14: 194–205.
Erdner, D.L. and D.M. Anderson, 1999. Ferredoxin and flavodoxin as biochemical indica-tors of iron limitation during open-ocean iron enrichment. Limnol. Oceanogr. 44: 1609–1615.
Erdner, D.L., N.M. Price, G.J. Doucette, M.L. Pelato and D.L. Anderson, 1999. Characterizationof ferredoxin and flavodoxin as markers of iron limitation in marinbe phytoplankton. Mar.Ecol. Prog. Ser. 184: 43–53.
Esler, J.J., E.R. Marzolf and C.R. Goldman, 1990. Phosphorus and nitrogen limitation of phyto-plankton growth in freshwaters of North America: A review and critique of experimentalenrichments. Can. J. Fish. Aquat. Sci. 47: 1468–1477.
Everest, S.A., C.R. Hipkin and P.J. Syrett, 1986. Enzyme activities in some marine phytoplanktersand the effect of nitrogen limitation on nitrogen and carbon metabolism in Chlorella stigma-tophora. Mar. Biol. 90: 165–172.
Falkowski, P.G., 2000. Rationalizing elemental ratios in unicellular algae. J. Phycol. 36: 3–6.Falkowski, P.G. and D.P. Stone, 1975. Nitrate uptake in marine phytoplankton: Energy sources and
the interaction with carbon fixation. Mar. Biol. 32: 77–84.Falkowski, P.G., A. Sukenik and R. Herzig, 1989. Nitrogen limitation in Isochrysis galbana (Hapto-
phyceae). II. Relative abundance of chloroplast proteins. J. Phycol. 25: 471–478.Flynn, K.J., 1998. Estimation of kinetic parameters for the transport of nitrate and ammonium into
marine phytoplankton. Mar. Ecol. Prog. Ser. 169: 13–28.Fresnedo, O. and J.L. Serra, 1992. Effects of nitrogen starvation on the biochemistry of Phormi-
dium laminosum (Cyanophyceae) J. Phycol 28: 183–193.Ganf, G.S., S.J.L. Stone and R.L. Oliver, 1986. Use of protein to carbohydrate ratios to analyse for
nutrient deficiency in phytoplankton. Aust. J. Mar. Freshw. Res. 37: 183–97.Gauthier, D.A. and D.H. Turpin, 1997. Interactions between inorganic phosphate (Pi) assimilation,
photosynthesis and respiration in the Pi-limited green alga Selenastrum minutum. Plant CellEnvir. 20: 12–24.
Geider, R.J., J. La Roche, R.M. Greene and M. Olaizola, 1993. Response of the photosyntheticapparatus of Phaeodactylum tricornutum (Bacillariophyceae) to nitrate, phosphate, or ironstarvation. J. Phycol. 29: 755–766.
Geider, R.J., H.L. MacIntyre, L.M. Graziano and R.M.L. McKay, 1998. Responses of the photo-synthetic apparatus of Dunaliella tertiolecta (Chlorophyceae) to nitrogen and phosphorus limi-tation. Eur. J. Phycol. 33: 315–332.
Goldman, J.C., 1986. On phytoplankton growth rates and particulate carbon:nitrogen:phosphorusratios at low light. Limnol. Oceanogr. 31: 1358–63.
Goldman, J.C. and M.R. Dennett, 1986. Dark CO2 uptake by the diatom Chaetoceros simplex inresponse to nitrogen pulsing. Mar. Biol. 90: 493–500.
Goldman, J.C. and P.M. Glibert, 1983. Kinetics of inorganic nitrogen uptake by phytoplankton. In:Nitrogen in the Marine Environment. (Carpenter, E.J. and Capone, D.G. Eds.), AcademicPress. pp. 233–274.
Goldman, J.C., J.J. McCarthy and D.G. Peavey, 1979. Growth rate influence on the chemical com-position of phytoplankton in oceanic waters. Nature. 279: 2110–215.
González-Gil, S., B.A. Keafer, R.V.M. Jovine, A. Aguilera, S. Lu and D.M. Anderson, 1998.Detection and quantification of alkaline phosphatase in single cells of phosphorus-starvedmarine phytoplankton. Mar. Ecol. Prog. Ser. 164: 21–35.
Gotham, I.J. and G. Rhee, 1981. Comparative kinetic studies of phosphate-limited growth andphosphate uptake in phytoplankton in continuous culture. J. Phycol. 17: 257–65.
Graziano, L.M., J. La Roche and R.J. Geider, 1996. Physiological response to phosphorus limita-tion in batch and steady-state cultures of Dunaliella tertiolecta (Chlorophyta): a unique stressprotein as an indicator of phosphate deficiency. J. Phycol. 32: 825–838.
Greene, R.M., R.J. Geider, Z. Kolber and P.G. Falkowski, 1992. Iron-induced changes in light har-vesting and photochemical energy conversion processes in eukaryotic marine algae. Plant Phy-siol. 100: 565–575.
Harris, G.P., 1986. Phytoplankton Ecology: Structure, Function and Fluctuation. Chapman andHall, London. 384 pp.
Phytoplankton nutrient limitation 65

Harris, G.P. and B.B. Piccinin, 1983. Phosphorus limitation and carbon metabolism in a unicellu-lar alga: Interaction between growth rate and the measurement of net and gross photosynthe-sis. J. Phycol. 19: 185–92.
Harrison, P.J., H.L. Conway, R.W. Holmes and C.O. Davis, 1977. Marine diatoms grown in che-mostats under silicate or ammonium limitation. III Cellular composition and morphology ofChaetoceros debilis, Skeletonema costatum and Thalassiosira gravida. Mar. Biol. 43: 19–31.
Harrison, G.I., L.R. Harris and B.D. Irwin, 1996. The kinetics of nitrogen utilization in the oceanicmixed layer: Nitrate and ammonium interactions at nanomolar concentrations. Limnol.Oceanogr. 41: 16–32.
Harrison, P.J., P.A. Thompson and G.S. Calderwood, 1990. Effects of nutrient and light limitationon the biochemical composition of phytoplankton. J. Appl. Phycol. 2: 45–5.
Hayes, P.K., T.M. Whitaker and G.E. Fogg, 1984. The distribution and nutrient status of phyto-plankton in the Southern Ocean between 20° and 70°W. Polar Biol. 3: 153.
Healey, P.F., 1979 Short-term responses of nutrient-deficient algae to nutrient addition. J. Phycol.15: 289–299.
Heath, M.R., K. Richardson and T. Kiørboe, 1990. Optical assessment of phytoplankton nutrientdepletion. J. Plank. Res. 12: 381–396.
Hecky, R.E. and P. Kilham, 1998. Nutrient limitation of phytoplankton in freshwater and marineenvironments: a review of recent evidence on the effects of enrichment. Limnol. Oceanogr. 33:796–822.
Herzig, R. and P.G. Falkowski, 1989. Nitrogen limitation in Isochrysis galbana (Haptophyceae).I. Photosynthetic energy conversion and growth efficiencies. J. Phycol. 25: 462–471.
Holmes, J.J., H.G. Weger and D.H. Turpin, 1989. Chlorophyll a fluorescence predicts total photo-synthetic electron flow to CO2 or NO3
–/NO2– under transient conditions. Plant Physiol. 91:
331–337.Huppe, H.C. and D.H. Turpin, 1994. Integration of carbon and nitrogen metabolism in plant and
algal cells. Ann. Rev. Plant Physiol. Plant Mol. Biol. 45: 577–607.Hutber, G.N., K.G. Hutson and L.J. Rogers, 1977. Effect of iron deficiency on levels of two ferre-
doxins and flavodoxin in a cyanobacterium. FEMS Microbiol. Lett. 1: 193–196.Istanovics, V., K. Petterson, D. Pierson and R. Bell, 1992. Evaluation of phosphorus deficiency
indicators for summer phytoplankton in Lake Erken. Limnol. Oceanogr. 35: 1258–1266.Karl, D.M., 1999. A sea of change: Biogeochemical variability in the North Pacific subtropical
gyre. Ecosystems 2: 181–214.Kessler, E. and H. Osterheld, 1970. Nitrification and induction of nitrate reductase in nitrogen-
deficient algae. Nature 228: 287–288.Kolber, Z., J. Zehr and P. Falkowski, 1988. Effects of growth irradiance and nitrogen limitation on
photosynthetic energy conversion in photosystem II. Plant Physiol. 88: 923–929.Kolber, Z.S., R.T. Barber, K.H. Coale, S.E. Fitzwater, R.M. Greene, K.S. Johnson, S. Lindley and
P.G. Falkowski, 1994. Iron limitation of phytoplankton photosynthesis in the equatorial PacificOcean. Nature 371: 145–149.
Krom, M.D., N. Kress and S. Brenner, 1991. Phosphorus limitation of primary productivity in theeastern Mediterranean Sea. Limnol. Oceanogr. 36: 424–432.
La Roche, J., R.J. Geider, L.M. Graziano, H. Murray and K. Lewis, 1993. Induction of specificproteins in eukaryotic algae grown under iron-, phosphorus-, or nitrogen-deficient conditions.J. Phycol. 29: 767–777.
La Roche, J., H. Murray, M. Orellana and J. Newton, 1995. Flavodoxin expression as an indicatorof iron limitation in marine diatoms. J. Phycol. 31: 520–530.
La Roche, J., P.W. Boyd, R.M.L. McKay and R.J. Geider, 1996. Flavodoxin as an in situ marker foriron stress in phytoplankton. Nature 382: 802–805.
Laws, E.A. and T. T. Bannister, 1980. Nutrient- and light-limited growth of Thalassiosira fluviatilisin continuous culture, with implications for phytoplankton growth in the ocean. Limnol.Oceanogr. 25: 457–473.
Lean, D.R.S. and F.R. Pick, 1981. Photosynthetic response of lake plankton to nutrient enrich-ment: A test for nutrient limitation. Limnol. Oceanogr. 26: 1001–1019.
Lean, D.R.S., T.P. Murphy and F.R. Pick, 1982. Photosynthetic response of lake plankton to com-bined nitrogen enrichment. J. Phycol. 18: 509–521.
66 Beardall et al.

Leonhardt, K. and N.A. Straus, 1992. An iron stress operon involved in photosynthetic electrontransport in the marine cyanobacterium Synechococcus sp. PCC-7002. J. Gen. Microbiol. 138:1613–1621.
Li, H., J.W. Veldhuis and A.F. Post, 1998. Alkaline phosphatase activities among planktonic com-munities in the northern Red Sea. Mar. Ecol. Prog. Ser. 173: 107–115.
Lippemeier, S., P. Hartig and F. Colijn, 1999. Direct impact of silicate on the photosynthetic per-formance of the diatom Thalassiosira weissflogii assessed by on- and off-line PAM fluores-cence measurements. J. Plank. Res. 21: 269–283.
Martin, J.H. and S.E. Fitzwater, 1988. Iron deficiency limits phytoplankton growth in the north-east pacific subartic. Nature 331: 341–343.
Matta, J.L., N.S. Govind and R.K. Trench, 1992. Polyclonal antibodies agains iron-superoxide dis-mutase from Escherichia coli B cross-react with superoxide dismutases from Symbiodiniummicroadriaticum (Dinophyceae). J Phycol. 28: 343–346.
Maxwell, K. and G. Johnson, 2000. Chlorophyll fluorescence – a practical guide. J. Exp. Bot. 51:659–668.
McKay, R.M.L., R.J. Geider and J. La Roche, 1997. Physiological and biochemical response of thephotosynthetic apparatus of two marine diatoms to Fe stress. Plant Physiol. 114: 615–622.
McKay, R.M.L., J. La Roche, A.F. Yakunin, D.G. Durnford and R.J. Geider, 1999. Accumulationof ferredoxin and flavodoxin in a marine diatom in response to Fe. J. Phycol. 35: 510–519.
Menzel, D.W. and J.H. Ryther, 1961. Nutrients limiting the production of phytoplankton in theSargasso Sea, with special reference to iron. Deep Sea Res. 7: 276–281.
Miller, W.E., T.E. Maloney and E.J.C. Greene, 1974. Algal productivity in 49 lake waters, as deter-mined by algal assays. Water Res. 8: 667–79.
Miller, W.E., E.J.C. Greene and T. Shiroyama, 1978. The Selenastrum capricornutum Printz algalassay: bottle test. Experimental design, application and data interpretation protocol. EPA(USA)-60019-78-018. 124 p.
Morel, F.M.M., J.R. Reinfelder, S.B. Roberts, C.P. Chamberlain, J.G. Lee and D. Yee, 1994. Zincand carbon co-limitation of marine phytoplankton. Nature 369: 740–742.
Morris, I., 1981. Photosynthetic products, physiological state, and phytoplankton growth. In: Platt,T. (ed.) Physiological Bases of Phytoplankton Ecology. Can. Bull. Fish. Aquat. Sci. 210:83–102.
Morris, I., C.M. Yentsch and C.S. Yentsch, 1971. The physiological state with respect to nitrogenof phytoplankton from low-nutrient subtropical water as measured by the effect of ammoni-um ion on dark carbon fixation. Limnol. Oceanogr. 16: 859–68.
Muggli, D.L. and P.J. Harrison, 1996. EDTA suppresses the growth of oceanic phytoplankton fromthe Northeast Subarctic Pacific. J. Exp. Mar. Biol. Ecol. 205: 221–227.
Oliver, R.L., 1993. Phosphorus dynamics and bioavailability in turbid waters. Land and WaterResources Research and Development Corporation, Occasional Paper No. 03/93, Canberra,Australia.
Packard, T.T., R.C. Dugdale, J.J. Goering and R.T. Barber, 1978. Nitrate reductase activity in thesubsurface waters of the Peru Current. J. Mar. Res. 36: 59–76.
Palenik, B. and J.A. Koke, 1995. Characterization of a nitrogen-regulated protein identified by cellsurface biotinylation of a marine phytoplankton. App. Environ. Microbiol. 61: 3311–3315.
Parslow, J.S., P.J. Harrison and P.A. Thompson, 1984a. Development of rapid ammonium uptakeduring starvation of batch and chemostat cultures of the marine diatom Thalassiosira pseudo-nana. Mar. Biol. 83: 43–50.
Parslow, J.S., P.J. Harrison and P.A. Thompson, 1984b. Saturated uptake kinetics: Transient re-sponse of the marine diatom Thalassiosira pseudonana to ammonium, nitrate, silicate or phos-phate starvation. Mar. Biol. 83: 51–59.
Perry, M.J., 1976. Phosphate utilization by an oceanic diatom in phosphorus-limited chemostat cul-tures and in the oligotrophic waters of the central North Pacific. Limnol Oceanogr. 21: 88–107.
Peeters, J.C.H. and L. Peperzak, 1990. Nutrient limitation in the North Sea: A bioassay approach.Neth. J. Sea Res. 26: 61–73.
Plumley, F.G. and G.W. Schmidt, 1989. Nitrogen-dependent regulation of photosynthetic geneexpression. Proc. Nat. Acad. Sci. USA. 86: 2678–2682.
Raven, J.A. and P.G. Falkowski, 1999. Oceanic sinks for atmospheric CO2. Plant Cell Envir. 22:741–755.
Phytoplankton nutrient limitation 67

Raven, J.A., M.C.W. Evans and R.E. Korb, 1999. The role of trace metals in photosynthetic elec-tron transport in O2-evolving organisms. Photo. Res. 60: 111–149.
Rees, A.P., N.J.P. Owens, M.R. Heath, D.H. Plummer and R.S. Bellerby, 1995. Seasonal nitrogenassimilation and carbon fixation in a fjordic sea loch. J. Plank. Res. 17: 1307–1324.
Redfield, A.C., 1958. The biological control of chemical factors in the environment. Am. Sci. 46:205–21.
Reichardt, W., J. Overbeck and L. Steubing, 1967. Free dissolved enzymes in lake waters. Nature216: 1345–47.
Reitan, K.I., J.R. Rainuzzo and Y. Olsen, 1994. Effect of nutrient limitation on fatty acid and lipidcontent of marine microalgae. J. Phycol. 30: 972–79.
Rhee, G.-Y., 1974. Phosphate uptake under nitrate limitation by Scenedesmus sp. and its ecologicalimplications. J. Phycol. 10: 470–475.
Roberts, S.C., 1997. Physiological effects of phosphorus limitation on photosynthesis in two greenalgae. PhD thesis, Monash University, Melbourne, Australia. 116 pp.
Rose, C. and R.P. Axler, 1998. Uses of alkaline phosphatase activity in evaluating phytoplanktoncommunity phosphorus deficiency. Hydrobiologia 361: 145–156.
Sakshaug, E., E. Granéli, M. Elbrächter and H. Kayser, 1984. Chemical composition and alkalinephosphatase activity of nutrient-saturated and P-deficient cells of four marine dinoflagellates.J. Exp. Mar. Biol. Ecol. 77: 241–254.
Sas, H., 1989. Lake restoration by reduction of nutrient loading: expectations, experiences, extra-polations. Academia Verlag Richarz, St Augustin, p. 497.
Scanlan, D.J., N.H. Mann and N.G. Carr, 1989. Effect of iron and other nutrient limitations on thepattern of outer membrane proteins in the cyanobacterium Synechococcus PCC7942. Arch.Microbiol. 152: 224–228.
Scanlan, D.J., N.H. Mann and N.G. Carr, 1993. The response of the picoplanktonic marine cyano-bacterium Synechococcus species WH7803 to phosphate starvation involves a protein homo-logous to the periplasmic phosphate-binding protein of Escherichia coli. Mol. Biol. 10:181–191.
Schindler, D.W., 1977. Evolution of phosphorus limitation in lakes. Science 195: 260–262.Schreiber, U., 1994. New emitter-detector-cuvette assembly for measuring modulated chlorophyll
fluorescence of highly diluted suspensions in conjunction with the standard PAM fluorometer.Z. Naturfors. 49c: 646–656.
Shifrin, N.S. and S.W. Chisholm, 1981. Phytoplankton lipids: interspecific differences and effectsof nitrate, silicate and light-dark cycles. J. Phycol. 17: 374–384.
Sosik, H.M. and B.G. Mitchell, 1991. Absorption, fluorescence, and quantum yield for growth innitrogen-limited Dunaliella tertiolecta. Limnol. Oceanogr. 36: 910–921.
Stevens, R.J. and M.P. Parr, 1977. The significance of alkaline phosphatase activity in LoughNeagh. Freshw. Biol. 7: 351–355.
Syrett, P.J., 1981. Nitrogen metabolism of microalgae. In: Physiological Bases of PhytoplanktonEcology. (ed. T. Platt). Can. Bull. Fish. Aquat. Sci. 210: 182–210.
Tett, P., S.I. Heaney and M.R. Droop, 1985. The Redfield ratio and phytoplankton growth rate. J.Mar. Biol. Ass. U.K. 65: 487–504.
Theodorou, M.E., I.R. Elrifi, D.H. Turpin and W.C. Plaxton, 1991. Effects of phosphorus limita-tion on respiratory metabolism in the green alga Selenastrum minutum. Plant Physiol. 95:1089–1095.
Thompson, P., 1999. The response of growth and biochemical composition to variations in day-length, temperature, and irradiance in the marine diatom Thalassiosira pseudonana (Bacilla-riophyceae). J. Phycol. 35: 1215–1223.
Timmermans, K.R., M.A. van Leeuwe, J.T.M. de Jong, R.M.L. McKay, R.F. Nolting, H.J. Witte, J.van Ooyen, M.J.W. Swagerman, H. Kloosterhuis and H.J.W. de Baar, 1998. Iron stress in thePacific region of the Southern Ocean: evidence from enrichment bioassays. Mar. Ecol. Prog.Ser. 166: 27–41.
Turpin, D.H. and H.G. Weger, 1988. Steady-state chlorophyll a fluorescence transients duringammonium assimilation by the N-limited green alga Selenastrum minutum. Plant Physiol. 88:97–101.
Turpin, D.H. and P.J. Harrison, 1979. Limiting nutrient patchiness and its role in phytoplanktonecology. J. Exp. Mar. Biol. Ecol. 39: 151–166.
68 Beardall et al.

Twiss, M.R., J.-C. Auclair and N.M. Charlton, 2000. An investigation into iron-stimulated phyto-plankton productivity in epipelagic Lake Erie during thermal stratification using trace metalclean techniques. Can. J. Fish. Aquat. Sci. 57: 86–95.
van Leeuwe, M.A. and J. Stefels, 1998. Effects of iron and light stress on the biochemical compo-sition of Antarctic Phaeocystis sp. (Prymnesiophyceae). II. Pigment composition. J. Phycol. 34:496–503.
Venrick, E.L., J.R. Beers and J.F. Heinbokel, 1977. Possible consequences of containing micro-plankton for physiological rate measurements. J. Exp. Mar. Biol. Ecol 26: 55–76.
Vincent, W.F., 1981. Rapid physiological assays for nutrient demand by the plankton. II Phos-phorus. J. Plank. Res. 3: 699–710.
Vrba, J., V. Vyhná, J. Hejzlar and J. Nedoma, 1995. Comparison of phosphorus deficiency indicesduring a spring phytoplankton bloom in a eutrophic reservoir. Freshw. Biol. 33: 73–81.
Watt, D.A., A.M. Amory and C.F. Cresswell, 1992. Effect of nitrogen supply on the kinetics andregulation of nitrate assimilation in Chlamydomonas reinhardtii Dangeard. J. Exp. Bot. 43:605–615.
Wynne, D. and T. Berman, 1980. Hot water extractable phosphorus – an indicator of nutritionalstatus of Peridinium cinctum (Dinophyceae) from Lake Kinneret (Israel). J. Phycol. 16: 40–4.
Wynne, D. and T. Berman, 1990. The influence of environmental factors on nitrate reductaseactivity in freshwater phytoplankton. I. Field studies. Hydrobiologia 194: 235–245.
White, E. and G. Payne, 1980. Distribution and biological availability of reactive high molecularweight phosphorus in natural waters in New Zealand. Can. J. Fish. Aquat. Sci. 37: 664–669.
White, E., K. Law, G. Payne and S. Pickmere, 1985. Nutrient demand and availability amongplanktonic communities – an attempt to assess nutrient limitation to plant growth in 12 centralvolcanic plateau lakes. N. Z. J. Mar. Freshw. Res. 19: 49–62.
Wood, M.D. and R.L. Oliver, 1995. Fluorescence transients in response to nutrient enrichment ofnitrogen- and phosphorus-limited Microcystis aeruginosa cultures and natural phytoplanktonpopulations: a measure of nutrient limitation. Aust. J. Plant Physiol. 22: 331–340.
Young, E.B., 1999. Interactions between photosynthetic carbon and nitrogen acquisition in themarine microalga Dunaliella tertiolecta Butcher. PhD Thesis. Monash University, Melbourne,Australia. 228 pp.
Zettler, E.R., R.J. Olson, B.J. Binder, S.W. Chisholm, S.E. Fitzwater and R.M. Gordon, 1996. Iron-enrichment bottle experiments in the equatorial Pacific: response of individual phytoplanktoncells. Deep-Sea Res. II 43, 4–6.
Received 29 August 2000;revised manuscript accepted 25 September 2000.
Phytoplankton nutrient limitation 69
To access this journal online:http://www.birkhauser.ch
All in-text references underlined in blue are linked to publications on ResearchGate, letting you access and read them immediately.