intra-floodplain seed dispersal limitation and wetland community restoration
TRANSCRIPT

September 2014 ECOLOGICAL RESTORATION 32:3 • 249
Ecological Restoration Vol. 32, No. 3, 2014ISSN 1522-4740 E-ISSN 1543-4079©2014 by the Board of Regents of the University of Wisconsin System.
RESEARCH ARTICLE
Intra-Floodplain Seed Dispersal Limitation and Wetland Community Restoration
Scott H. Markwith, Garren Mezza, Stevee N. Kennard and Stephen G. Bousquin
ABSTRACTThe broadleaf marsh community in the post-restoration reaches of the Kissimmee River floodplain, Florida, US, has failed to achieve the expected dominance based on pre-disturbance conditions. We examined the hypothesis that seed dispersal is a limiting factor delaying the expansion of characteristic taxa of a dominant wetland plant community, broadleaf marsh, from remnant populations into a partially restored river floodplain. Hydrochory and anemochory seed traps were deployed in the Kissimmee River for two week periods during four sampling periods over the course of a year. Study plots were divided among an area influenced by backwater effects from a downstream water control structure and the substantially drier floodplain with more fragmented and degraded broadleaf marsh communities to the north. Seed trapping showed that seeds of characteristic broadleaf marsh taxa, e.g., bulltongue arrowhead (Sagittaria lancifolia), pickerelweed (Pontederia cordata), and common buttonbush (Cephalanthus occidentalis), were only rarely dispersed and in small volumes north of the confluence of Oak Creek. Seed limitation due to a partially restored physical environment may be one of the factors contributing to minimal recovery of wetland communities in the Kissimmee River floodplain. As with fragmented regional scale landscapes with substantial distances among patches and an inhospitable matrix ecosystem, our research indicates that seed limitation may also be an issue for wetland restoration at local scales in contiguous floodplain wetlands such as the Kissimmee River. Active revegetation practices may be necessary for accelerating expansion of plant communities in large-scale river/floodplain restoration projects where local seed sources are spatially and hydrologically constrained.
Keywords: anemochory, broadleaf marsh, hydrochory, Kissimmee River, macrophytes, passive restoration
By understanding the processes driving succession, scientists can
suggest ways to increase rates at which desired states, i.e. reference conditions, are reached through the process of restoration (Temperton et al. 2004). In many communities, the availability of propagules is a limiting factor in reaching those desired states (Robin-son and Handel 1993, Guariguata et al. 1995, Lamb et al. 1997, as cited in Young et al. 2001). Increasingly, the assumption that propagules will be readily available for colonization of physically restored areas is being questioned (Galatowitsch and van der Valk 1996, Zobel et al. 2006, Trow-bridge 2007). Restoration of physi-cal processes may be a prerequisite to successful ecological restoration, but
may not be sufficient (Suding et al. 2004, Trowbridge 2007). Research in floodplain wetlands in Europe and the U.S. indicates that dispersal limitation may be a problem in these ecosystems (Bakker and Berendse 1999, Donath et al. 2003, Trowbridge 2007). Nar-rowing or elimination of the dispersal niche (i.e. the set of environmental conditions that determine where a seed arrives) with hydrologic altera-tion in floodplains may be respon-sible for some limitation, but Young et al. (2005) suggest the niche may be extended with restoration.
Seeds of wetland plants can be effectively dispersed by water (hydro-chory), wind (anemochory), or ani-mals (zoochory). These dispersal processes are influential in ecologi-cal restoration, can be affected by flow regulation and climate change (Nilsson et al. 2010), and can give advanced warning of invasive or exotic species colonization (Neff and
Baldwin 2005). Wetland plant species can be dispersed by several different mechanisms, but hydrochory is typi-cally the primary vector in wetland systems (Neff and Baldwin 2005) and has a long scientific pedigree recog-nizing its population and commu-nity level importance (Ridley 1930, Skoglund 1989, Boedeltje et al. 2003, Merritt et al. 2010, Markwith and Leigh 2008, Monette and Markwith 2012). Anemochory is often over-looked due to hydrochory’s preemi-nence in wetlands, but anemochory plays a key role in dispersing small, lightweight, specifically adapted seeds (Neff and Baldwin 2005, Soons 2006). Both anemochory and zooch-ory can transport seeds over long dis-tances (Ozinga et al. 2004), although in lower densities than hydrochory, and unlike water can disperse seeds in all directions, including areas of the wetland that are not connected by surface water flows (Soons 2006).

250 • September 2014 ECOLOGICAL RESTORATION 32:3
The Kissimmee River is the main tributary to Lake Okeechobee, which historically fed the Everglades marsh system in southern Florida, US. The floodplain was historically dominated by the broadleaf marsh community, with hydroperiods > 200 days per year and wet season water depths > 1 m (Kushlan 1990). In the 1960s, the entire Kissimee River was canalized by the U.S. Army Corps of Engi-neers, reducing the river length by almost half. The flood control proj-ect included the 100 m wide C-38 canal and five water control structures, i.e. locks and dams, that maintained nearly constant water levels (Toth 2010). One of the largest river res-toration projects in the United States began with implementation of Phase I in 1999, which included restoration of flow to 39 km of original and recarved river channel, backfilling > 15 km of canal, some channel recarving, and removal of one water control structure (SFWMD 2012).
Prior to canalization of the Kissim-mee River, the predominant wetland plant communities were broadleaf marsh (dominated by bulltongue arrowhead [Sagittaria lancifolia], pickerelweed [Pontederia cordata], and sometimes including common buttonbush (Cephalanthus occiden-talis)) (Milleson et al. 1980, Toth et al. 1995, as cited in Toth 2010), wet prairie (dominated by maidencane [Panicum hemitomon] at the longer end of the hydroperiod gradient and various grasses, sedges, and forbs in shorter hydroperiod habitat), and wet-land shrub (dominated by common buttonbush and coastal plain willow [Salix caroliniana]). Shortly after canalization, and dewatering of the floodplain, these communities were largely replaced by upland vegetation and pasture. By 1996 (25 years after canalization), the three once-domi-nant wetland plant communities had substantially dropped in proportion of floodplain area, i.e. broadleaf marsh dropped from 46% to 7% of the flood-plain, wet prairie from 21% to 11%, and wetland shrub from 13% to 9%
(Bousquin 2005, Carnal and Bous-quin 2005). The South Florida Water Management District (SFWMD) pre-dicted these same floodplain wetland communities would recover to at least 80% of the floodplain area after water depth, extent, duration, and frequency of floodplain inundation were restored in response to canal backfilling and reconnection of remnant river chan-nels. Wetzel et al. (2001) found that especially for the broadleaf marsh and wetland shrub communities, but also within wet prairie, reestablishment of the former vegetation would require propagule dispersal from off-site sources because a viable seed bank was absent. van der Valk et al. (2009) concluded that propagule sources in remnant wetlands were abundant, and expansion of the dominant wetland species would proceed apace. A decade after Phase I was completed, recovery of the historically dominant broad-leaf marsh community in the river floodplain was yet to achieve the areal coverage predicted by the SFWMD (Carnal 2005, Spencer and Bousquin 2014).
Due to minimal expansion of the broadleaf marsh community in the Kissimmee River floodplain post-res-toration, we designed this research to test the hypothesis that seed limita-tion due to the failure of hydrochory and anemochory to disperse seeds to potential sites for colonization plays a substantial role in hindering the spread of this community. Specific research questions included: 1) what was the pattern of seed dispersal of characteristic wetland community taxa by hydrochory and anemochory in the restored floodplain of the Kissim-mee River; and 2) was seed dispersal limitation restricting the expansion of broadleaf marsh taxa from remnants into restored wetland habitat? Few studies have directly examined the composition and spatial patterns of the dispersing seed pool in the context of floodplain wetland restoration. This information will help inform future Kissimmee River adaptive manage-ment as the project continues into
additional phases, and explain how seed dispersal may influence veg-etation restoration success, or delay success, in large floodplain wetlands.
Methods
Study AreaThe study area in the floodplain of the Kissimmee River was located east of the unincorporated community of Lorida, Florida, USA, in what is con-sidered the Central Florida region. The climate is subtropical, and has distinct wet and dry seasons. The wet season spans June through September, with consistent mean high and low temper-atures ~34 °C and ~22 °C, respectively, and mean wet season precipitation of 79.3 cm. The dry season spans Octo-ber through May, with mean high and low temperatures in January ranging from ~23 °C to ~8 °C, respectively, and mean dry season precipitation of 53.0 cm.
Sampling locations were positioned in Phase I of the Kissimmee River restoration, where the C-38 canal was backfilled and some channel recarv-ing was conducted between 1999 and 2001. The locations were placed proxi-mal to the confluence of Oak Creek (Figure 1). The confluence of Oak Creek represented an approximate break point between the floodplain that remained impacted hydrologi-cally by backwater from a downstream water control structure, and the north-ern section that was unaffected by the backwater (Spencer and Bousquin 2014). The sampling locations south of the confluence of Oak Creek were dominated by broadleaf marsh and wetland shrub communities. These locations supported the historically dominant taxa of those two com-munities, i.e. bulltongue arrowhead, pickerelweed, common buttonbush, and coastal plain willow, and frequent representation from other native taxa, such as dotted smartweed (Polygonum punctatum), fragrant flatsedge (Cyperus odoratus), marsh pennywort (Hydro-cotyle spp.), water spangles (Salvinia

September 2014 ECOLOGICAL RESTORATION 32:3 • 251
Figure 1. Vegetation map of Kissimmee River floodplain area of Phase I of restoration and sampling locations, FL, USA. Spatial data created by SFWMD from 2008 aerial imagery. Inset A: Area of interest of study in Kissimmee River restoration area. Inset B: Sampling protocol schematic for plot positioning within sampling locations in relation to broadleaf marsh, seed shadow, and non-seed shadow for broadleaf marsh plot types.
minima), and duckweed (Lemna spp.). Two exotic invasive taxa, Peruvian primrose-willow (Ludwigia peruvi-ana) and water hyacinth (Eichhornia crassipies), were also common. Sam-pling locations in the northern sec-tion of the floodplain were strongly dominated by wet prairie commu-nities and taxa historically typical of that community, i.e. maidencane and southern cutgrass (Leersia hexandra), with dotted smartweed also common. The remnant broadleaf marsh patches in the floodplain north of Oak Creek, identified with aerial photo interpre-tation and ground-truthing by the SFWMD, were generally depauper-ate of taxa historically dominant in those communities. Broadleaf marsh
wetland remnants were severely frag-mented by large swaths of floodplain converted to upland vegetation com-munities after canalization, and were primarily found in sections of flood-plain affected by backwater conditions from water control structures where hydroperiods remained suitable for broadleaf marsh.
Field Sampling and Data CollectionSampling locations were composed of two study plots within remnant broad-leaf marsh, two study plots in areas of wet prairie or wetland shrub directly within the remnant broadleaf marsh’s seed shadow (i.e. the hypothetical area within the dominant floodplain
flow-path directly ‘downstream’ from the broadleaf marsh remnant and potentially receiving dispersed seeds of broadleaf marsh taxa), and two study plots outside of the seed shadow of the broadleaf marsh remnant (i.e. non-seed shadow plots) (see Figure 1 Inset B for design schematic). Distances among plots within each location ranged from 50–100 m. Based on this sampling design we expected to find seeds of broadleaf marsh taxa in hydro-chory traps within broadleaf marsh remnants and seed shadow plots, but we did not expect to find seeds from broadleaf marsh taxa in the non-seed shadow hydrochory traps because the gradient controlled water flow pattern could not disperse seeds outside of the

252 • September 2014 ECOLOGICAL RESTORATION 32:3
seed shadow. This design, however, was not expected to show differences among sites with anemochory traps because that dispersal mechanism is more typically multi-directional.
There were eight sampling loca-tions (four north and four south of the confluence of Oak Creek, Figure 1) that contained six study plots each, for a total of 21 broadleaf marsh study plots, 13 seed shadow study plots, and 14 non-seed shadow study plots. We found that the entire area of sampling location 1 (L1), which was classified as having wetland shrub vegetation adja-cent to broadleaf marsh in 2008 by SFWMD, was dominated by broad-leaf marsh at the time of sampling. Variation in the 2008 classification maps and sampled vegetation com-munities in 2010/2011 was appar-ently due to the scale of mapping and some vegetation dynamic change at the broadleaf marsh/wetland shrub boundary between mapping and sampling periods. For analysis, all the study plots installed at L1 were classi-fied broadleaf marsh. Also, one of the expected seed shadow plots in L2 was dominated by broadleaf marsh taxa and was classified as broadleaf marsh for analysis purposes. To assess disper-sal of broadleaf marsh seeds into areas remote from broadleaf marsh rem-nants, we monitored seed dispersal in one remote wet prairie plot ( primar-ily maidencane) and one remote wet-land shrub plot ( primarily Peruvian primrose-willow).
Hydrochory and anemochory were sampled concurrently for a two week period approximately every three months (traps were deployed and collected, respectively, October 24 to November 7, 2010; January 16 to Jan-uary 30, 2011; April 16 to April 30, 2011; and July 10 to July 24, 2011). The timing of sampling events was scheduled with one event during the peak of the wet season ( July); one event following shortly after the peak tropi-cal cyclone season and accumulation of wet season rains, when floodplain water depths are typically substantial but beginning to recede (October/
November); and two events during the dry season that dominates 8 months of the year ( January and April). Ane-mochory sampling was possible year round; however, hydrochory, and any other sampling protocols based on the presence of water could only be conducted when flooded.
Each study plot consisted of one PVC stake to which one hydro-chory trap and one anemochory trap was affixed. Hydrochory traps were attached by rope to the permanently located PVC stakes. Floating hydro-chory seed trap design was based upon Middleton (1995) with minor variation. Traps were constructed from standard 2 gallon buckets with the bases cut out, nylon mesh fabric liners with square hole sizes of 500 µm to capture seeds (Industrial Netting part # NN1500), and flotation devices (sections of Styrofoam noodles: trap opening when floating, i.e. mouth of the trap = 215 cm2, and internal trap area, i.e. water surface area inside the trap = 2000 cm2, respectively).
Peruvian primrose-willow’s seed was the only relevant and dominant wetland taxa in the study whose seed or diaspore was smaller than 500 µm, thus the hydrochory traps allowed pas-sage of these seeds. Supplementary traps lined with muslin, deployed in July, were unfortunately asynchronous with the seed phenology of the species.
The anemochory traps were con-structed of a wind deflector to channel seeds into the traps, and a frame struc-ture holding a coconut fiber mat for seed trapping. The coconut fiber mat had 360 degree coverage, i.e. all direc-tions, and was attached 2 m above the ground surface to the same stake as the hydrochory trap (based upon Neff and Baldwin 2005).
After two weeks of deployment, traps were collected and transported back to Florida Atlantic University’s Biogeography Lab for seed extrac-tion, processing, and identification. Seeds too small to be identified with the naked eye were examined for diagnostic traits under a Leica S6 D microscope and a digital image
was captured with a Leica DFC295 3.1 megapixel digital camera. Seeds were visually sorted and identified using Martin and Barkley (2000), and a comprehensive seed key for wetland taxa developed using herbar-ium specimen, online resources, and direct field identification of maternal specimen.
The buoyancy period and germina-tion rate of the characteristic broad-leaf marsh taxa, bulltongue arrowhead, pickerelweed, and common button-bush, were examined using mature dia-spores and seed collected fresh from maternal sources in the river flood-plain. For buoyancy experiments, 50 seeds of bulltongue arrowhead and common buttonbush, and 25 seeds and 25 diaspores of pickerelweed, were placed in partitioned 4.2 L containers of water. The number of seeds/dia-spores sunk per taxa was counted each day until all seeds/diaspores had sunk; sunken seeds/diaspores were removed each day after counting. Water was added twice per week to maintain depth, and complete water changes were conducted monthly. Reporting for buoyancy period is the number of days required for 50% of the seeds to sink (FD50) (e.g., Mossman, 2009). Germination experiments were con-ducted in the greenhouse on the Boca Raton campus of Florida Atlantic Uni-versity. Seeds were stored from collec-tion to experiment start date in dry and dark conditions in envelopes. Half of the collected seeds were placed in 4 °C for cold stratification for 8 weeks, while the other half were stored at room tem-perature. Seeds of the three character-istic taxa were attached to constantly moist filters in petri dishes that were placed standing on end in a container of shallow water, ~3 cm deep. Forty seeds of each species were tested and split evenly among two petri dishes per species. The number of seeds that germinated per species was counted each day until all seeds had germinated or three months had passed. Water was added twice per week to maintain depth, and complete water changes were conducted monthly.

September 2014 ECOLOGICAL RESTORATION 32:3 • 253
Water depth (m) and flow veloc-ity (cm/s) were measured upstream from the PVC stake at each plot when the traps were deployed. Veloc-ity was measured using a SonTek/YSI Flowtracker Handheld-ADV Acoustic Doppler Velocity meter. Flow readings were taken just below the surface at 2.5 cm depth. Wind speed (m/s) and direction were recorded using a porta-ble anemometer (Kestrel 4500 Pocket Weather Tracker). Standing aquatic vegetation adjacent to the seed traps was sampled every 3 months within a 2 m × 0.5 m sample frame to iden-tify individual taxa, record their per-cent cover, and to characterize species richness.
Statistical and Data AnalysisTo characterize the seed pool, species richness, Shannon-Weiner diversity index, and total number of seeds at each trap were calculated for both hydrochory and anemochory sam-ples. Hydrochory traps that were physically destroyed (only 6 traps total were destroyed over the entire period) or where the dispersal vector was absent, i.e. no water in the study plot (100 traps total had no water over the entire period) were excluded from statistical analyses. In addition, where no water was present in a study plot, water depth was recorded as zero; however, water velocity was recorded as “no data” because velocity could not be measured. This method was chosen to differentiate water with no velocity from the inability to measure velocity due to no water. Anemochory traps that were damaged or destroyed were also excluded from analysis (10 traps over the entire period). Vari-ables were normalized using typical transformations where necessary.
Repeated measures Analysis of Variance (ANOVA) was used to test for significant differences between subjects grouped by study plot type or between subjects for plots north and south of Oak Creek. Within subjects tests were conducted based on sampling period and interactions with study plot type and relationship
to Oak Creek. These tests were con-ducted on seed pool variables for both the hydrochory and anemochory data (i.e. the per trap total number of seeds, species richness, and Shannon-Weiner diversity index), the number of seeds/trap for the most common species (i.e. species with a mean number of seeds/trap > 5), dispersal vector parameters (i.e. the per plot water depth, water velocity, wind direction, and mean wind speed), and standing vegetation parameters (i.e. the per plot percent cover and species richness). Repeated measures ANOVA was performed using STATA release 9 (STATA Corp., College Station, TX).
Results
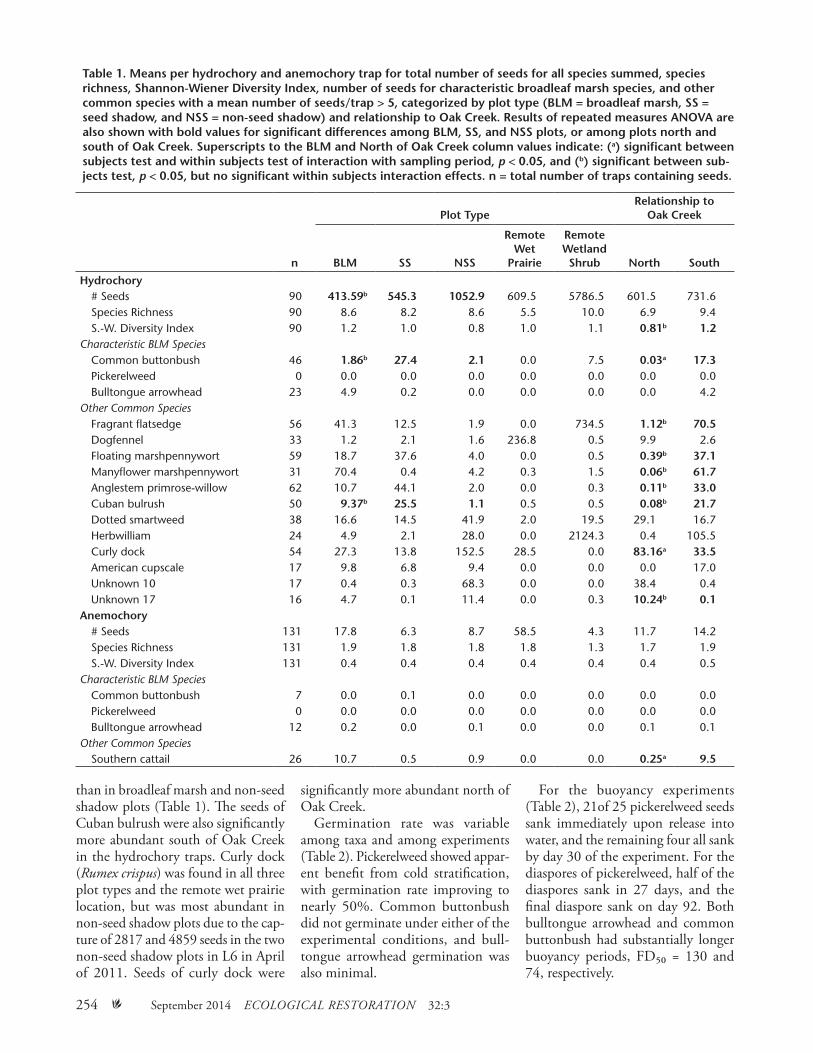
A total of 63,743 seeds among 58 taxa were trapped by the hydrochory traps over the four sampling periods, and 2,445 seeds among 35 taxa were trapped by the anemochory traps. Common buttonbush was the most abundant characteristic broadleaf marsh species in hydrochory traps (Table 1). Common buttonbush had significantly greater seed density in seed shadow plots and south of the confluence of Oak Creek than in other plot types, but was also present in broadleaf marsh, non-seed shadow, and the remote wetland shrub plots. Only 3 seeds of common buttonbush were found in hydrochory traps in the locations north of Oak Creek, in L6 non-seed shadow and L7 broadleaf marsh plots. Its greatest abundance was in plots where established individ-uals were found in the standing vegeta-tion quadrats (1619 seeds), compared to plots not supporting established individuals of common buttonbush (102 seeds). Common buttonbush was rare in anemochory traps. Bull-tongue arrowhead was largely confined to broadleaf marsh habitat (~5 seeds/hydrochory trap), occasionally in seed shadow plots, but seeds of this species were very rarely found north of Oak Creek (only 3 seeds in location L5, the rest in L1, L2, and L3) (Table 1). At plots where bulltongue arrowhead was
measured in the standing vegetation, 300 seeds of the species were trapped, while those plots without bulltongue arrowhead in the standing vegetation resulted in 119 trapped seeds. How-ever, two traps in the L1 location, where all plots were classified as broad-leaf marsh, accounted for 105 of the 119 seeds in traps without representa-tion of bulltongue arrowhead in the adjacent standing vegetation quadrats. Bulltongue arrowhead was present but rare in anemochory traps. Bulltongue arrowhead was also never found in the remote wet prairie or remote wet-land shrub plots in either hydrochory or anemochory traps. Pickerelweed diaspores and seed were not found in any of the hydrochory or anemochory traps in any sampling period (Table 1). Pickerelweed was observed flowering and fruiting in the vicinity of some traps, but none were identified from the trap samples.
Other taxa common in the hydro-chory seed pool were variably abun-dant among the sampling locations (Table 1). Fragrant flatsedge, many-flower marshpennywort (Hydrocotyle umbellata), and dotted smartweed are all taxa typical of both broad-leaf marsh and wet prairie commu-nities. Both fragrant flatsedge and manyflower marshpennywort were significantly more abundant in the seed pool south of Oak Creek and in broadleaf marsh plots, but not signifi-cantly, while dotted smartweed was more abundant north of Oak Creek and in non-seed shadow plots, but not significantly. Fragrant flatsedge also showed great abundance at the remote wetland shrub location, as did herbwilliam (Ptilimnium capil-laceum), whereas the upland species dogfennel (Eupatorium capillifolium) maintained a presence in the standing vegetation and seed pool in drier areas of the floodplain, e.g., the remote wet prairie location.
Two invasive exotic taxa were among the most common taxa in the hydrochory seed pool. Cuban bulrush (Oxycaryum cubense) was significantly more abundant in seed shadow plots
254 • September 2014 ECOLOGICAL RESTORATION 32:3
Table 1. Means per hydrochory and anemochory trap for total number of seeds for all species summed, species richness, Shannon-Wiener Diversity Index, number of seeds for characteristic broadleaf marsh species, and other common species with a mean number of seeds/trap > 5, categorized by plot type (BLM = broadleaf marsh, SS = seed shadow, and NSS = non-seed shadow) and relationship to Oak Creek. Results of repeated measures ANOVA are also shown with bold values for significant differences among BLM, SS, and NSS plots, or among plots north and south of Oak Creek. Superscripts to the BLM and North of Oak Creek column values indicate: (a) significant between subjects test and within subjects test of interaction with sampling period, p < 0.05, and (b) significant between sub-jects test, p < 0.05, but no significant within subjects interaction effects. n = total number of traps containing seeds.
Plot TypeRelationship to
Oak Creek
n BLM SS NSS
Remote Wet
Prairie
Remote Wetland
Shrub North South
Hydrochory # Seeds 90 413.59b 545.3 1052.9 609.5 5786.5 601.5 731.6 Species Richness 90 8.6 8.2 8.6 5.5 10.0 6.9 9.4 S.-W. Diversity Index 90 1.2 1.0 0.8 1.0 1.1 0.81b 1.2Characteristic BLM Species Common buttonbush 46 1.86b 27.4 2.1 0.0 7.5 0.03a 17.3 Pickerelweed 0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 Bulltongue arrowhead 23 4.9 0.2 0.0 0.0 0.0 0.0 4.2Other Common Species Fragrant flatsedge 56 41.3 12.5 1.9 0.0 734.5 1.12b 70.5 Dogfennel 33 1.2 2.1 1.6 236.8 0.5 9.9 2.6 Floating marshpennywort 59 18.7 37.6 4.0 0.0 0.5 0.39b 37.1 Manyflower marshpennywort 31 70.4 0.4 4.2 0.3 1.5 0.06b 61.7 Anglestem primrose-willow 62 10.7 44.1 2.0 0.0 0.3 0.11b 33.0 Cuban bulrush 50 9.37b 25.5 1.1 0.5 0.5 0.08b 21.7 Dotted smartweed 38 16.6 14.5 41.9 2.0 19.5 29.1 16.7 Herbwilliam 24 4.9 2.1 28.0 0.0 2124.3 0.4 105.5 Curly dock 54 27.3 13.8 152.5 28.5 0.0 83.16a 33.5 American cupscale 17 9.8 6.8 9.4 0.0 0.0 0.0 17.0 Unknown 10 17 0.4 0.3 68.3 0.0 0.0 38.4 0.4 Unknown 17 16 4.7 0.1 11.4 0.0 0.3 10.24b 0.1Anemochory # Seeds 131 17.8 6.3 8.7 58.5 4.3 11.7 14.2 Species Richness 131 1.9 1.8 1.8 1.8 1.3 1.7 1.9 S.-W. Diversity Index 131 0.4 0.4 0.4 0.4 0.4 0.4 0.5Characteristic BLM Species Common buttonbush 7 0.0 0.1 0.0 0.0 0.0 0.0 0.0 Pickerelweed 0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 Bulltongue arrowhead 12 0.2 0.0 0.1 0.0 0.0 0.1 0.1Other Common Species Southern cattail 26 10.7 0.5 0.9 0.0 0.0 0.25a 9.5
than in broadleaf marsh and non-seed shadow plots (Table 1). The seeds of Cuban bulrush were also significantly more abundant south of Oak Creek in the hydrochory traps. Curly dock (Rumex crispus) was found in all three plot types and the remote wet prairie location, but was most abundant in non-seed shadow plots due to the cap-ture of 2817 and 4859 seeds in the two non-seed shadow plots in L6 in April of 2011. Seeds of curly dock were
significantly more abundant north of Oak Creek.
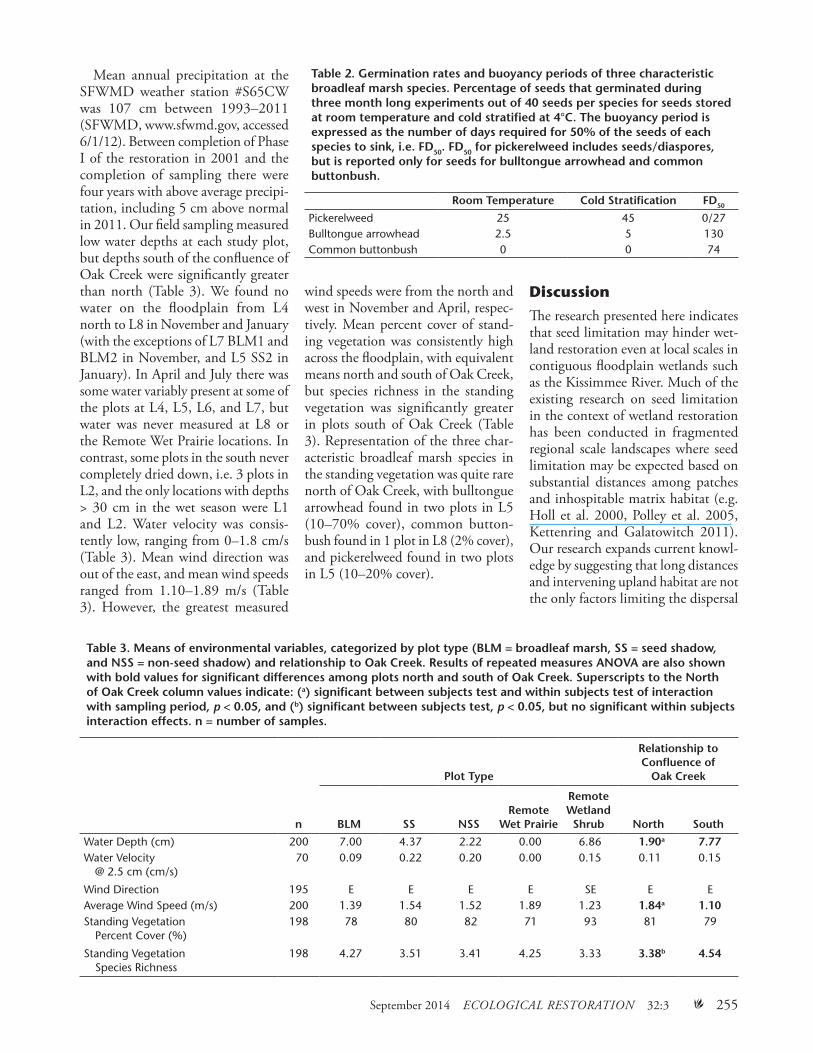
Germination rate was variable among taxa and among experiments (Table 2). Pickerelweed showed appar-ent benefit from cold stratification, with germination rate improving to nearly 50%. Common buttonbush did not germinate under either of the experimental conditions, and bull-tongue arrowhead germination was also minimal.
For the buoyancy experiments (Table 2), 21of 25 pickerelweed seeds sank immediately upon release into water, and the remaining four all sank by day 30 of the experiment. For the diaspores of pickerelweed, half of the diaspores sank in 27 days, and the final diaspore sank on day 92. Both bulltongue arrowhead and common buttonbush had substantially longer buoyancy periods, FD50 = 130 and 74, respectively.
September 2014 ECOLOGICAL RESTORATION 32:3 • 255
Table 2. Germination rates and buoyancy periods of three characteristic broadleaf marsh species. Percentage of seeds that germinated during three month long experiments out of 40 seeds per species for seeds stored at room temperature and cold stratified at 4°C. The buoyancy period is expressed as the number of days required for 50% of the seeds of each species to sink, i.e. FD50. FD50 for pickerelweed includes seeds/diaspores, but is reported only for seeds for bulltongue arrowhead and common buttonbush.
Room Temperature Cold Stratification FD50
Pickerelweed 25 45 0/27Bulltongue arrowhead 2.5 5 130Common buttonbush 0 0 74
Table 3. Means of environmental variables, categorized by plot type (BLM = broadleaf marsh, SS = seed shadow, and NSS = non-seed shadow) and relationship to Oak Creek. Results of repeated measures ANOVA are also shown with bold values for significant differences among plots north and south of Oak Creek. Superscripts to the North of Oak Creek column values indicate: (a) significant between subjects test and within subjects test of interaction with sampling period, p < 0.05, and (b) significant between subjects test, p < 0.05, but no significant within subjects interaction effects. n = number of samples.
Plot Type
Relationship to Confluence of
Oak Creek
n BLM SS NSS
Remote Wet Prairie
Remote Wetland
Shrub North South
Water Depth (cm) 200 7.00 4.37 2.22 0.00 6.86 1.90a 7.77Water Velocity @ 2.5 cm (cm/s)
70 0.09 0.22 0.20 0.00 0.15 0.11 0.15
Wind Direction 195 E E E E SE E EAverage Wind Speed (m/s) 200 1.39 1.54 1.52 1.89 1.23 1.84a 1.10Standing Vegetation Percent Cover (%)
198 78 80 82 71 93 81 79
Standing Vegetation Species Richness
198 4.27 3.51 3.41 4.25 3.33 3.38b 4.54
Mean annual precipitation at the SFWMD weather station #S65CW was 107 cm between 1993–2011 (SFWMD, www.sfwmd.gov, accessed 6/1/12). Between completion of Phase I of the restoration in 2001 and the completion of sampling there were four years with above average precipi-tation, including 5 cm above normal in 2011. Our field sampling measured low water depths at each study plot, but depths south of the confluence of Oak Creek were significantly greater than north (Table 3). We found no water on the floodplain from L4 north to L8 in November and January (with the exceptions of L7 BLM1 and BLM2 in November, and L5 SS2 in January). In April and July there was some water variably present at some of the plots at L4, L5, L6, and L7, but water was never measured at L8 or the Remote Wet Prairie locations. In contrast, some plots in the south never completely dried down, i.e. 3 plots in L2, and the only locations with depths > 30 cm in the wet season were L1 and L2. Water velocity was consis-tently low, ranging from 0–1.8 cm/s (Table 3). Mean wind direction was out of the east, and mean wind speeds ranged from 1.10–1.89 m/s (Table 3). However, the greatest measured
wind speeds were from the north and west in November and April, respec-tively. Mean percent cover of stand-ing vegetation was consistently high across the floodplain, with equivalent means north and south of Oak Creek, but species richness in the standing vegetation was significantly greater in plots south of Oak Creek (Table 3). Representation of the three char-acteristic broadleaf marsh species in the standing vegetation was quite rare north of Oak Creek, with bulltongue arrowhead found in two plots in L5 (10–70% cover), common button-bush found in 1 plot in L8 (2% cover), and pickerelweed found in two plots in L5 (10–20% cover).
Discussion
The research presented here indicates that seed limitation may hinder wet-land restoration even at local scales in contiguous floodplain wetlands such as the Kissimmee River. Much of the existing research on seed limitation in the context of wetland restoration has been conducted in fragmented regional scale landscapes where seed limitation may be expected based on substantial distances among patches and inhospitable matrix habitat (e.g. Holl et al. 2000, Polley et al. 2005, Kettenring and Galatowitch 2011). Our research expands current knowl-edge by suggesting that long distances and intervening upland habitat are not the only factors limiting the dispersal

256 • September 2014 ECOLOGICAL RESTORATION 32:3
ability of wetland plants into restored habitat. Seed limitation may also occur within a single, contiguous wetland system, and in such cases restoration of abiotic factors may not be sufficient to spur passive restoration from all remnant plant communities.
Seeds of characteristic broadleaf marsh taxa were more numerous in broadleaf marsh and seed shadow plots than non-seed shadow, which was expected based on transport and subsequent deposition within the gradient driven flow-path. However, similar to Donath et al. (2003), who found distances only in the 10s of meters were spanned between remnant and restored populations within 10 years, the assumption of hydrology as mechanism for long-distance dispersal within the floodplain was not sup-ported. The vast majority of broadleaf marsh seeds were trapped adjacent to potential maternal sources, and thus likely did not move substantial dis-tances. The pattern was predominant despite bulltongue arrowhead bear-ing seeds that are light weight and have a flat winged shape that may be conducive to anemochory, and that the buoyancy periods for bulltongue arrowhead and common button-bush were also sufficient to facilitate long distance hydrochory. Emergent standing vegetation was quite dense through most of the wetland, thus pre-senting a high friction environment where hydrochory may be slowed or completely stopped when seeds adhere to emergent surfaces (Schneider and Sharitz 1988). Open water in the Kis-simmee River floodplain conducive to long distance transport is not spatially extensive, especially north of the back-water effect, except in extreme flood conditions following tropical storms and hurricanes, e.g., Hurricane Isaac in 2012. Thus, the dispersal niche for many species has not been fully expanded with canal backfilling, rela-tively short hydroperiods, and shallow depths common in most years. Resto-ration may depend in part on the yet to be implemented Kissimmee Chain of Lakes Headwaters Revitalization
regulation schedule in the river head-waters, that will attempt to mimic historic hydroperiods (Bousquin et al. 2005, Williams et al. 2005).
Limited potential for long-distance dispersal away from maternal sources is particularly problematic at the Kis-simmee River given patterns in the standing vegetation, where the domi-nant broadleaf marsh taxa were only rarely found north of Oak Creek, i.e. healthy and expansive broadleaf marsh remnants are downstream and distant from the areas in greatest need of colo-nization. The historically characteristic taxa of the broadleaf marsh commu-nity were only rarely dispersing by hydrochory and anemochory and in quite low densities to locations in the northern end of the Phase I flood-plain. Many areas classified as broad-leaf marsh remnants by SFWMD in the upper reaches of Phase I support negligible populations of the charac-teristic broadleaf marsh taxa and may have inappropriate hydroperiods to support broadleaf marsh, even though they are hydrologically more similar to broadleaf marsh than the surrounding shorter hydroperiod floodplain. These severely depleted remnants may not serve as sufficient local source popu-lations for broadleaf marsh expan-sion via either sexual or vegetative reproduction.
The majority of the northern end of the Phase I floodplain was cov-ered by maidencane-dominated wet prairie, which differs from broadleaf marsh in the dominant characteris-tic species but has largely overlapping hydroperiod tolerances (Lowe 1983, Lowe 1986, Kushlan 1990, Richard-son 2000). Toth’s (2010) examination of niche-assembly in the Kissimmee River floodplain rejected as oversim-plified the single filter hypothesis that hydrology is the primary driver of broadleaf marsh restoration, and found variation in community trajec-tories that were not explained well by restoration of hydrologic regime. Even with a completely restored hydrologi-cal regime implemented at a relatively late stage, community restoration may
be forestalled by the substantial influ-ence of priority effects on wet prai-rie and broadleaf marsh trajectories. Community assembly models have found that historical contingencies such as invasion order and preemp-tion by early colonists are important determinants of community compo-sition in experimental communities (Platt and Weiss 1985, Robinson and Edgemon 1988, Drake et al. 1996), while other studies have shown that open colonization sites may exist and diversity is recruitment limited (Tilman 1997), an apparent problem at the Kissimmee River. The estab-lished communities may not change in a deterministic manner even if the dispersal niche can be expanded and more propagules of broadleaf marsh species arrive in wet prairie wetlands.
Pickerelweed was not trapped during sampling, seeds sunk imme-diately upon release into water, the diaspores had a relatively short buoy-ancy period, and structures suggesting wind dispersal are absent. Although hydrochory and anemochory are the dominant dispersal mechanisms in wetlands (Neff and Baldwin 2005), zoochory can also act as an impor-tant dispersal mechanism (Barrat-Segretain, 1996, Mueller and van der Valk 2002, Vittoz and Engler, 2007). Mossman (2009) indicated that ducks may eat diaspores of pickerelweed, but this hypothesis was not tested at the Kissimmee River. Neither common buttonbush nor bulltongue arrow-head have fleshy pericarps attractive to animals and indicative of poten-tial ingestion, however, there remains potential for adherence to water fowl and wading bird’s legs or feathers in water or mud. Mist netting or use of net guns to capture birds and examine their feces and for seeds attached to legs and feathers would help fill this gap in our knowledge.
Not only was the density of seed rain on many parts of the floodplain negligible for broadleaf marsh species, but the effectiveness of that dispersal was questionable given potentially inappropriate habitat for germination

September 2014 ECOLOGICAL RESTORATION 32:3 • 257
and low germination rates once the seeds arrived. The combination of the probability of deposition and prob-abilities of germination and estab-lishment are all factors affecting the overall probability of an individual colonizing a location. The germina-tion rate was greatest with pickerel-weed, and increased with cold stratifi-cation. Whigham and Simpson (1982) found rather high germination rates, > 50%, for pickerelweed after cold stratification > 10 weeks and increas-ing with longer stratification. Getty and Dumroese (2009) reported high germination rates when seeds were submerged in water. The germina-tion rates with bulltongue arrowhead and common buttonbush were much reduced. Similarly, Mossman (2009) measured a relatively low 29% germi-nation rate for bulltongue arrowhead seeds, while Collon and Velasquez (1989) found up to 100% germina-tion at 25°C. DuBerry (1963) and Bonner (1974) reported a germination rate of 78% to 84% for common but-tonbush. In all studies, germination rate was variable with treatments, but greater than those reported herein. The probabilities of establishment remain unknown.
Conclusions
Passive restoration is promoted as an efficient and relatively cost effective methodology based on the reversal of degradation and expectation that native species will naturally recolonize the target location. Expecting plant species to disperse to a site and recreate a reference system within the monitor-ing window is a deterministic view of community assembly (Trowbridge 2007). The appeal of this equilibrial succession philosophy, or “self-design” (Mitsch et al. 1998), for resource man-agers is that it supports the allocation of limited financial resources nearly exclusively to land acquisition and physical restoration. However, if biotic factors, e.g., seed limitation, influ-ence the restoration trajectory, abiotic restoration may be insufficient and
active management may be necessary to establish the desired communities (Seabloom and van der Valk 2003).
Large scale projects like the Kissim-mee River restoration, e.g., 39 km of river restoration, require great expen-ditures for land purchases and con-struction for physical restoration, e.g., > US$600 million (SFWMD 2009). These massively complex human and natural systems lend themselves to passive restoration where planning is envisioned and implemented in broad strokes, even though inordinate impacts may be caused by ecologi-cal processes working on some of the smallest biological units, i.e. the seed scale. Our research supports the asser-tion that active restoration by manual seed dispersal or planting seedlings may be necessary to overcome seed limitation and accelerate broadleaf marsh expansion and approximation of reference conditions (Kettenring and Galatowitsch 2011). However, several fundamental questions could be considered by scientists, manag-ers, and stakeholders prior to adaptive management, including a) is broad-leaf marsh a critical vegetation state that must be restored to historical reference proportions for appropri-ate ecosystem functioning, or b) can our expectations and timelines be suf-ficiently suspended to observe how natural processes produce outcomes over the longterm that may or may not ever match our preconceptions of the restored ecosystem?
AcknowledgementsThis research was funded by the South Florida Water Management District under P.O. 4500053356. The authors would like to thank the following individuals for field assistance: Dale Gawlik, Tyler Beck, Rich Botta, Dean Monette; Riverwoods Field Lab for providing field accommodations; and anonymous reviewers for the assistance strengthening the manuscript.
ReferencesBakker, J.P. and F. Berendse. 1999. Con-
straints in the restoration of ecological diversity in grassland and heathland
communities. Trends in Ecology and Evolution 14:63–68.
Barrat-Segretain M.H. 1996. Strategies of reproduction, dispersion and com-petition in river plants: A review. Vegetation 123:13–37.
Boedeltje, G., J.P. Bakker, R.M. Bekker, J.M. Van Groenendael and M. Soes-bergen. 2003. Plant dispersal in a low-land stream in relation to occurrence and three specific life-history traits of the species in the species pool. Journal of Ecology 91:855–866.
Bonner, F.T. 1974. Cephalanthus occi-dentalis L., common buttonbush. In C.S. Schopmeyere, tech. coord. Seeds of woody plants in the United States. Washington, D.C.: United States Department of Agriculture Forest Service Agriculture Handbook 450.
Bousquin, S. 2005. Chapter 8: Chan-nelized Kissimmee River Floodplain Vegetation. In S.G. Bousquin, D.H. Anderson, G.E. Williams and D J. Colangelo (eds), Kissimmee River Res-toration Studies, Establishing a baseline: Pre-restoration studies of the channel-ized Kissimmee River, vol. I. SFWMD Technical Publication ERA 432.
Bousquin, S., D.H. Anderson, D.J. Col-angelo and G.E. Williams. 2005. Chapter I: Introduction to baseline studies of the channelized Kissimmee River. In S.G. Bousquin, D.H. Ander-son, G.E. Williams and D.J. Colan-gelo (eds). Kissimmee River Restora-tion Studies, Establishing a baseline: Pre-restoration studies of the channel-ized Kissimmee River, vol. I. SFWMD Technical Publication ERA 432.
Carnal, L. 2005. Expectation 13: Areal coverage of broadleaf marsh. In S.G. Bousquin, D.H. Anderson, G.E. Wil-liams and D.J. Colangelo (eds), Kis-simmee River Restoration Studies, Defining success: Expectations for resto-ration of the Kissimmee River, vol. II. SFWMD Technical Publication ERA 433.
Carnal, L. and S. G. Bousquin. 2005. Chapter 10, Areal coverage of flood-plain plant communities in Pool C of the channelized Kissimmee River. In S. G. Bousquin, D. H. Ander-son, G. E. Williams and D. J. Col-angelo (eds), Kissimmee River Resto-ration Studies, Establishing a baseline: Pre-restoration studies of the channel-ized Kissimmee River, vol. I. SFWMD Technical Publication ERA 432.
Collon, E.G. and J. Velasquez. 1989. Dis-persion, germination and growth of

258 • September 2014 ECOLOGICAL RESTORATION 32:3
seedlings of Sagittaria lancifolia. Folia Geobotanica 24:37–49.
Donath, T.W., N. Holzel and A. Otte. 2003. The impact of site conditions and seed dispersal on restoration suc-cess in alluvial meadows. Applied Vegetation Science 6:13–22.
Drake, J.A., G.R. Huxel and C.L. Hewitt. 1996. Microcosms as models for gen-erating and testing community theory. Ecology 77:670–677.
DuBerry, A.P., Jr. 1963. Germina-tion of bottomland tree seed while immersed in water. Journal of Forestry 61:225–226.
Galatowitsch, S.M. and A.G. van der Valk. 1996. The vegetation of restored and natural prairie wetlands. Ecological Applications 6:102–112.
Getty L.A. and R.K. Dumroese. 2009. Optimum storage and germination conditions for seeds of pickerelweed (Pontederia cordata L.) from Florida. Native Plants Journal 10:4–12.
Guariguata, M.R., R. Rhemgans and F. Montagnini. 1995. Early woody invasion under tree plantations in Costa Rica: Implications for forest regeneration. Restoration Ecology 3:252–250.
Holl, K.D., M.E. Loik, E.H. V. Lin and I.A. Samuels. 2000. Tropical mon-tane forest restoration in Costa Rica: Overcoming barriers to dispersal and establishment. Restoration Ecology 8:339–349.
Kettenring, K.M. and S.M. Galatow-itsch. 2011. Seed rain of restored and natural prairie wetlands. Wetlands 31:283–294.
Kushlan, J.A. 1990. Freshwater marshes. Pages 324–363 in R.L. Myers and J.J. Ewel (eds), Ecosystems of Florida. Orlando, FL: University of Central Florida Press.
Lamb, D., J. Parrotta, R. Keenan and N. Tucker. 1997. Rejoining habitat fragments: Restoring degraded rain-forest lands. Pages 366–385 in W.F. Laurance and R.O. Bierregaard (eds), Tropical Forest Remnants Ecology, Man-agement and Conservation of Frag-mented Communities. Chicago, IL: University of Chicago Press.
Lowe, E.F. 1983. Distribution and struc-ture of floodplain plant communi-ties in the Upper Basin of the St. Johns River, Florida. Technical Publi-cation SJ83–8, St. Johns River Water Management District.
Lowe, E.F. 1986. The relationship between hydrology and vegetational pattern
within the floodplain marsh of a sub-tropical, Florida lake. Florida Scientist 49:213–233.
Markwith, S.H and D.S. Leigh. 2008. Subaqueous hydrochory: Open-channel hydraulic modeling of non-buoyant seed movement. Freshwater Biology 53:2274–2286.
Martin, A.C. and W.D. Barkley. 2000. Seed Identification Manual. Caldwell, NJ: The Blackburn Press.
Merritt, D.M., C. Nilsson and R. Jans-son. 2010. Consequences of propa-gule dispersal and river fragmentation for riparian plant community diversity and interannual turnover. Ecological Monographs 80:609–626.
Middleton, B. 1995. Sampling devices for the measurement of seed rain and hydrochory in rivers. Bulletin of the Torrey Botanical Club 122:152–155.
Milleson, J.M., R.L. Goodrick and J.A. van Arman. 1980. Plant communities of the Kissimmee River valley. Tech-nical Publication #80–7, South Flor-ida Water Management District, West Palm Beach, Florida.
Mitsch, W.J., X. Wu, R.W. Nairn, P.E. Weihe, N. Wang, R. Deal and C.E. Boucher. 1998. Creating and restoring wetlands. Bioscience 48:1019–1030.
Monette, D. and S.H. Markwith. 2012. Hydrochory in the Florida Everglades: Temporal and spatial variation in seed dispersal phenology, hydrology, and restoration of wetland structure. Ecological Restoration 30:180–191.
Mossman, R. 2009. Seed dispersal and reproduction patterns among Ever-glades plants. FIU Electronic Theses and Dissertations. Paper 135. digitalcommons.fiu.edu/etd/135.
Mueller, M.H. and A.G. van der Valk. 2002. The potential role of ducks in wetland seed dispersal. Wetlands 22:170–178.
Neff, K.P. and A.H. Baldwin. 2005. Seed dispersal into wetlands: Tech-niques and results for a restored tidal freshwater marsh. Wetlands 25:392–404.
Nilsson, C., R.L. Brown, R. Jansson and D.M. Merritt. 2010. The role of hydrochory in structuring riparian and wetland vegetation. Biological Reviews 85:837–58.
Ozinga, W.A., R.M. Bekker, J. Scham-inee and J.M. Van Groenendael. 2004. Dispersal potential in plant communities depends on environ-mental conditions. Journal of Ecology 92:767–777.
Platt, W.J. and I.M. Weiss. 1985. An experimental study of competition among fugitive prairie plants. Ecology 66:708–720.
Polley, H.W., J.D. Derner and B.J. Wilsey. 2005. Patterns of plant species diver-sity in remnant and restored tall-grass prairie. Restoration Ecology 13:480–487.
Richardson, C.J. 2000. Freshwater wet-lands. Pages 488–499 in (eds) M.G. Barbour and W.D. Billings, North American Terrestrial Vegetation, 2nd ed. Cambridge, UK: University Press.
Ridley H.N. 1930. The Dispersal of Plants Throughout the World. Kent, UK: L. Reeve & Co., Ltd.
Robinson, G.R. and S.N. Handel. 1993. Forest restoration on a closed land-fill: Rapid addition of new species by bird dispersal. Conservation Biology 7:271–278.
Robinson, J.V. and M.A. Edgemon. 1988. An experimental evalua-tion of the effects of invasion his-tory on community structure. Ecology 69:1410–1417.
Seabloom, E.W. and A.G. van der Valk. 2003. Plant diversity, composition, and invasion of restored and natural prairie pothole wetlands: implications for restoration. Wetlands 23:1–12.
SFWMD. 2009. South Florida Envi-ronmental Report: Vol. I. The South Florida Environment. my.sfwmd.gov.
SFWMD. 2012. Restoring the Kissimmee River. ww.sfwmd.gov/portal/page/portal/ xweb protecting and restoring/kissimmee river. Accessed: February 2, 2012.
Schneider, R.L. and R.R. Sharitz. 1988. Hydrochory and regeneration in a bald cypress-water tupelo swamp forest. Ecology 69:1055–1063.
Skoglund, S.J. 1989. Seed dispersal agents in two regularly flooded river sites. Canadian Journal of Botany 68:754–760.
Soons, M.B. 2006. Wind dispersal in freshwater wetlands: Knowledge for conservation and restoration. Applied Vegetation Science 9:271–78.
Spencer, L.J. and S.G. Bousquin. 2014. Interim responses of wetland vegeta-tion to phase I of the Kissimmee River restoration project: Comparisons of vegetation maps from five periods in the river’s history. Restoration Ecology 22:397–408.
Suding, K.N., K.L. Gross and G.R. Houseman. 2004. Alternative states and positive feedbacks in restoration

September 2014 ECOLOGICAL RESTORATION 32:3 • 259
ecology. Trends in Ecology and Evolution 19:46–53.
Temperton, V., R. Hobbs, T. Nuttle and S. Halle. 2004. Assembly Rules and Restoration Ecology: Bridging the Gap Between Theory and Practice. Washington, DC: Island Press.
Tilman, D. 1997. Community invisibility, recruitment limitation, and grassland biodiversity. Ecology 78:8–92.
Toth, L.A. 2010. Restoration response to relict broadleaf marsh to increased water depths. Wetlands 30:263–274.
Toth, L.A., D.A. Arrington, M.A. Brady and D.A. Musziak. 1995. Conceptual evaluation of factors potentially affect-ing restoration of habitat structure within the channelized Kissimmee River ecosystem. Restoration Ecology 3:160–180.
Trowbridge, W.B. 2007. The role of sto-chasticity and priority effects in floodplain restoration. Ecological Applications 17:1312–1324.
van der Valk, A.G., L.A. Toth, E.B. Gibney, D.H. Mason and P.R. Wetzel. 2009. Potential propagule sources for reestablishing vegetation on the flood-plain of the Kissimmee River, Florida, USA. Wetlands 29:976–987.
Vittoz, P. and R. Engler. 2007. Seed dis-persal distances: A typology based on dispersal modes and plant traits. Botanica Helvetica 117:109–124.
Wetzel, P.R., A.G. van der Valk and L.A. Toth. 2001. Restoration of wetland vegetation on the Kissimmee River floodplain: potential role of seed banks. Wetlands 21:189–198.
Whigham, D.F. and R.L. Simpson. 1982. Germination and dormancy studies of Pontederia cordata L. Bulletin of the Torrey Botanical Club 109:524–528.
Williams, G.E., J.W. Koebel, D.H. Ander-son, S.G. Bousquin, D.J. Colangelo, J.L. Glenn, B.L. Jones, C. Carlson, L. Carnal and J. Jorge. 2005. Chapter 11: Kissimmee River restoration and upper basin initiatives. South Flor-ida Environmental Report, vol. I. The South Florida environment—Water year 2004. SFWMD and FLDEP Technical Publication ERA 432.
Young, T.P., J.M. Chase and R.T. Hud-dleston. 2001. Community succes-sion and assembly: Comparing, con-trasting and combining paradigms in the context of ecological restoration. Ecological Restoration 19:5–18.
Young, T.P., D.A. Petersen and J.J. Clary. 2005. The ecology of restoration:
Historical links, emerging issues and unexplored realms. Ecology Letters 8:662–673.
Zobel, M., M. Opik, M. Moora and M. Partel. 2006. Biodiversity and ecosystem functioning: It is time for dispersal experiments. Journal of Vegetation Science 17:543–547.
Scott H. Markwith (corresponding author), Department of Geosciences, Flor-ida Atlantic University, 777 Glades Road, Boca Raton, FL 33431, [email protected]. Garren Mezza, Department of Geosci-ences, Florida Atlantic University, Boca Raton, FL. Stevee N. Kennard, Department of Geosci-ences, Florida Atlantic University, Boca Raton, FL. Stephen G. Bousquin, South Florida Water Management District, West Palm Beach, FL.
Pontederia cordata. USDA-NRCS PLANTS Database / USDA NRCS. Wetland flora: Field office illustrated guide to plant species. USDA Natural Resources Conservation Service.