the effects of copulatory behavior on sperm transport and fertility in rats
TRANSCRIPT

The Effects of Copulatory Behavior on Sperm Transport and Fertility
in Rats
NORMAN T. ADLER AND JAMES PATRICK TONER, JR.
Department of Psychology University of Pennsylvania
Philadelphia, Pennsylvania 19104
INTRODUCTION
Judging from the pervasive use of contraception in our own species, one might guess that getting pregnant is a simple process. From the point of view of biological mechanism and function, this is far from the truth. While high premiums are paid to a system which reliably produces pregnancy under the right conditions, severe penalties are paid for pregnancy under unfavorable circumstances.
The adaptive balancing of easy pregnancy with prudent pregnancy has produced a complex set of controls which regulate both the opportunity to mate and the effects of mating. These controls exist at many levels of biological organization, from mo- lecular to social. This principle of integration at multiple levels of biological organi- zation mandates that students of reproductive behavior adopt the corresponding strategy of integrative, multidisciplinary investigation-precisely the approach that Jay Rosenblatt has developed so elegantly, interestingly, and influentially.
In sexually reproducing species, sperm and ova must meet for fertilization to occur. In mammals, the union of egg and sperm typically requires sperm to pass from the male into the female's reproductive tract where they must pass, successively, through her cervix, uterus, uterotubal junction, and a length of oviduct. Despite the importance of these processes for fertility (and infertility), many investigators find the present understanding of the process inc~mplete.'-~
One of the ways to further our understanding of mammalian fertility is to analyze the role of copulatory behavior in this process. It is now well documented that behavior influences sperm transport and fertility in species spanning a broad taxonomic range.G22 In recent years, we have investigated the behavioral control of transcervical and transuterine sperm transport in terms of its underlying mechanisms and its functional sign$cance. This work proceeds on three levels of biological organization: the organ- ismic level, the social level, and the physiological level.
Although this narrative is something of a case study, concentrating on the ways in which behavior controls pregnancy in the rat, we think the biopsychological prin- ciples uncovered are rather general, with potential relevance for understanding prob- lems of human fertility and infertility.
21

22 ANNALS NEW YORK ACADEMY OF SCIENCES
OUTLINE SEXUAL BEHAVIOR AND THE INDUCTION OF PREGNANCY
The copulatory pattern of rats consists of a series of 10 to 15 intromissions culminating in ejaculation, during which the pair remains immobile for a number of seconds. At ejaculation, the male deposits sperm and a coagulating “copulatory plug” in the vagina. This plug lodges tightly up against the cervix and holds the sperm in contact with the channels in the c e r ~ i x . ’ ~ , ~ ~
The sequence of intromissions and ejaculation constitutes an ejaculatory series. This series is followed by a pause in the mating (the postejaculatory internal or PEI). The composite pattern of ejaculatory series and PEI is repeated on the average of seven times for a given pair of rats in one mating session.
The stimulation derived from this mating sequence controls the onset of subsequent pregnancy. It does so by influencing the probability of becoming progestational (i.e., secreting progesterone) and by transporting sperm through the reproductive tract into the uterus.10~16*23”7 While both the hormonal basis of gestation and gamete transport are necessary components of pregnancy induction (and hence neither is sufficient), in the remainder of this paper we will concentrate on the latter aspect-sperm transport.
Although early workers thought that sperm were deposited directly in the uterus of the female rat at e j a c u l a t i ~ n ~ ~ * ~ * their interpretation was based on qualitative es- timates of sperm numbers in the uterus. Moreover, the earliest examination of the uterus made by these investigators was on the order of 1 min after ejaculation:* which at least leaves open the question of how many sperm were transported in the first minute following ejaculation.
More modem data, based on quantitative analysis of sperm transport over a more extended temporal range of the PEI, show transcervical sperm transport to be gradual.16 In fact, several reviewers believe that convincing evidence has not been found for direct intrauterine deposition of sperm in any specie^?^*'^
In the rat there are several physiological requirements for transcervical sperm transport to occur. First, the reproductive system of the female rat must be primed by pre-ejaculatory factors; second, the form and placement of the copulatory plug at ejaculation must be adequatez3; and third, the plug’s position must not be disrupted by exogenous stimulation during the first minutes after ejaculation.’6
All three of these physiological requirements for successful sperm transport in the female rat are influenced by copulatory behavior. In the remainder of this paper, we will discuss the reciprocal relationship between behavior and physiology in initiating sperm output, transport, and subsequent fertilization. In the next section, we will discuss the influence of the individual behavioral components on sperm transport.
BEHAVIORAL INITIATION OF SPERM TRANSPORT
Pre-ejaculatory Behavior: Female Effects
Females require “priming” by the male rat’s pre-ejaculatory intromissions if they are to transport optimal numbers of sperm following e j ac~ la t ion . ’~~~~ Without two or more intromissions, sperm transport and pregnancy do not occur.” Although we do

ADLER & TONER EFFECTS OF COPULATORY BEHAVIOR ON SPERM 23
not yet understand the precise mechanism by which the intromissions exert their effect, our tentative hypothesis is that the mechanism may be humoral, since the pre-ejac- ulatory intromissions may be stored for a considerable period of time.
Pre-ejaculatory Behavior: Male Effects
Although we have known for some time that pre-ejaculatory intromissions affect the female’s capacify to transport sperm, we have recently obtained evidence that the number of intromissions a male rat delivers afects his ejaculatoly performan~e.~’
To obtain these data, we allowed males and females different copulatory experiences by switching females during mating. Thirty-four virgin females were mated to sexually rested male rats. During 31 of these mating tests, the male was switched from a stimulus female to the experimental female in the course of his ejaculatory series. The switch was performed at random points in the series. Consequently, the experimental male and female experienced different numbers of pre-ejaculatory intromissions. Some females had as few as one pre-ejaculatory intromission and some males as few as three.
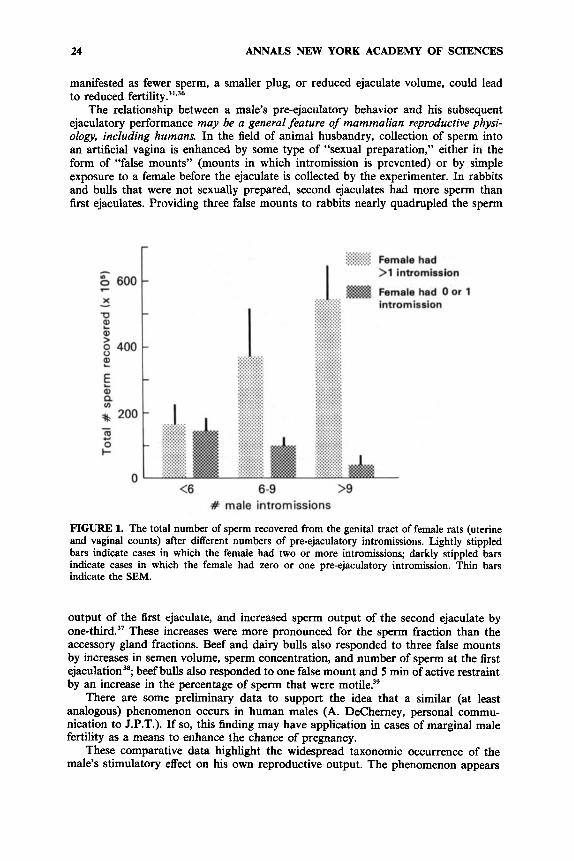
Reduced numbers of sperm were recovered when the female received fewer than two intromissions and/or when the male had fewer than six intromissions overall. Thus, the presence of sperm in the female rat’s genital tract depended upon both (a) the number of intromissions the female received and (b) the number the male delivered. These relationships are illustrated in FIGURE 1.
These data confirm our previous results showing that female rats require pre- ejaculatory intromissions to transport sperm into the ~ te rus . ’~~’~*” The present data also show that there is a clear-cut threshold in females: sperm recovery was always meager when she received fewer than two intromissions and was adequate if she received three or more intromissions.
In contrast to the effect on females, the role of the stimulation the male receives from the intromissions he delivers has not been previously documented. We found that few sperm were recovered when males had fewer than six intromissions, even when stimulation to the female was above her threshold (i.e., she received more than two intromissions). When males had six or more intromissions, many sperm were usually recovered. This transition from poor to good sperm recovery was not, however, as abrupt as that in the female.
These self-stimulatory effects of the male rat’s behavior on his own sperm output could have functional consequences. First, it is well known that the number of pre- ejaculatory intromissions among male rats is variable. Second, this variability in the number of intromissions is not a constant organismic variable between individual males but varies as a function of environmental and social contingencies. Experimen- tally, it is a simple matter to vary the number of intromissions by a variety of techniq~es.~’-~~ More to the functional orientation of the present discussion, there is an ecological relationship between social factors and reproductive function. Subdom- inant males have fewer than half the number of pre-ejaculatory intromissions that dominant males have.3’ In a related finding, subdominant male rats seem to have depressed fertility in the presence of the dominant conspecific (but normal fertility when mating in the absence of the dominant male36). We suggest that in a natural context subdominant males would take fewer intromissions to ejac~late:~ and thus would produce a suboptimal ejaculate. This reduced ejaculatory potency, whether
24 ANNALS NEW YORK ACADEMY OF SCIENCES
manifested as fewer sperm, a smaller plug, or reduced ejaculate volume, could lead to reduced f e r t i l i t~ .~ ’ ,~~
The relationship between a male’s pre-ejaculatory behavior and his subsequent ejaculatory performance may be a general feature of mammalian reproductive physi- ology, including humans. In the field of animal husbandry, collection of sperm into an artificial vagina is enhanced by some type of ‘‘sexual preparation,” either in the form of “false mounts” (mounts in which intromission is prevented) or by simple exposure to a female before the ejaculate is collected by the experimenter. In rabbits and bulls that were not sexually prepared, second ejaculates had more sperm than first ejaculates. Providing three false mounts to rabbits nearly quadrupled the sperm
r 600
X - - t E L
<6
........ ............... ............... ............... ....... Female had >1 intromission I
or 1
6-9 >9 # male intromissions
FIGURE 1. The total number of sperm recovered from the genital tract of female rats (uterine and vaginal counts) after different numbers of pre-ejaculatory intromissions. Lightly stippled bars indicate cases in which the female had two or more intromissions; darkly stippled bars indicate cases in which the female had zero or one pre-ejaculatory intromission. Thin bars indicate the SEM.
output of the first ejaculate, and increased sperm output of the second ejaculate by ~ne-third.~’ These increases were more pronounced for the sperm fraction than the accessory gland fractions. Beef and dairy bulls also responded to three false mounts by increases in semen volume, sperm concentration, and number of sperm at the first ejaculation”; beef bulls also responded to one false mount and 5 min of active restraint by an increase in the percentage of sperm that were m~t i le . )~
There are some preliminary data to support the idea that a similar (at least analogous) phenomenon occurs in human males (A. DeCherney, personal commu- nication to J.P.T.). If so, this finding may have application in cases of marginal male fertility as a means to enhance the chance of pregnancy.
These comparative data highlight the widespread taxonomic occurrence of the male’s stimulatory effect on his own reproductive output. The phenomenon appears

ADLER & TONER EFFECTS OF COPULATORY BEHAVIOR ON SPERM 25
to be a general one as well from the point of view of behavioral dynamics for it illustrates what Cheng has called behavioral selfsfeedbaclC4O This term refers to an animal’s behavior influencing its own subsequent behavior orphysiology. Other examples include the self-licking in female rats which promotes mammary development4’; the nest-coos of female ring doves which stimulate normal follicular development”; and the vocalizations of young sparrows which are necessary for normal adult singingP3
Self-stimulation is often an efficient mechanism for an animal to regulate activities which cannot be coordinated by purely endogenous means. For example, egg-laying needs to be coordinated with the availability of an adequate nest and cannot depend solely on internal indices. Canaries check the environment directly, for tactile contact made with a completed nest allows egg-laying to occur.” In contrast, egg-laying in ring doves follows a certain amount of nest-building behavior, independent of the condition of the ne~t .4~ In both of these cases, the adequacy of the external environment for receiving the precious egg is assured by making sure that the external world has been prepared-either by checking it directly (as in the canary) or by using a “be- havioral token” like nest-building activity (as in the ring dove).
Ejaculatory Behavior
Once the female is primed by the precopulatory intromissions, she is ready to receive the sperm. Mere deposition of the sperm in the vagina is, however, not sufficient-the behavior of the pair at ejaculation influences fertility. More sperm are transported into the uterus when the pair of rats remains immobile at ejaculation with close pelvic contact for a second or more after the final ejaculatory thrust.’6 This beneficial effect of ejaculatory immobility might be mediated by better placement of the vaginal plug against the cervixI6 since (a) the better the plug conforms to the shape of the cervix, the more sperm are transported” and (b) the placement of the plug is a function of the duration of the pair’s ejaculatory immobility response. That is, pairs that remained immobile at ejaculation had plugs with tighter fits; and tighter- fitting plugs transport more sperm.’b’*
Postejaculatory Behavior
The PEI is a crucial period in the reproductive life of the female rat, because intromissions delivered too soon after an ejaculation are able to reduce sperm transport, sometimes by two orders of magnitude, thereby disrupting Postejaculatory intromissions may exert their inhibitory effects by mechanically dislodging the vaginal plug.
The most critical postejaculatory period for this disruption is the first 6 min following ejaculation.‘” This length of time corresponds to the male rars PEL We have obtained a fair amount of evidence that this important period of reproductive quiescence in the male is assured by an active inhibitory state.-’ We conclude that one function of this state of inhibition is to prevent the male from “prematurely” resuming mating. This in turn would permit the female sufficient time without cop- ulatory stimulation to complete sperm transport.

26 ANNALS NEW YORK ACADEMY OF SCIENCES
The periodic action of inhibition is not specific to the male rat’s sexual behavior. Although the point is so basic that it is at times overlooked, a major topographic aspect of behavior is its temporal organization. In this sequencing of responses, be- havioral inhibition is a potent and ubiquitous mechanism for “turning off’ behavior at the appropriate point.
Perhaps because of the pervasiveness of behavior inhibition, we should not have been surprised to find an analogous example of lowered sexual responsiveness during the female rat’s postejaculatory inter~a1.4~~~’ In the male, the postejaculatory inhibition provides a pause in the stream of copulatory behavior, helping to reconcile the conflict between (a) the male’s producing sufficient sperm (from prolonged copulation and multiple ejaculatory series) and (b) permitting the female rat’s reproductive system the quiescence to “process” his sperm from the immediately preceding ejaculation. In her copulatory behavior, the female rat also has a conflict.
The copulating female also faces a strategic conflict: her survival depends upon both her mating and mothering. Davis and Hall” have determined that approximately 50% of all pregnancies in populations of wild rats result from postpartum mating. (Shortly after giving birth to one litter, the female rat becomes receptive and can copulate, thus initiating a second litter.) Since female rats will spend approximately 80% of their time caring for their l i t t e r~ ,5~-~~ this presents a problem: time taken to mate is time taken away from mothering, and vice versa. To investigate how the female handles this potential conflict, we studied the occurrence and temporal organization of maternal and copulatory behavior in postpartum female We found that a strategic compromise exists in which the female exhibits both mating and mothering, alternating between the two in the manner that McFarland5’ has called “behavioral time-sharing.” While time-sharing is not as elementary a mechanism as that controlling the male’s PEI, it does allow for the orderly alternation of two behavior patterns, with the dominant pattern allowing the other to appear only in its wake.
In the case of postpartum mating in the rat, copulatory behavior seems to be the dominant behavior-since the female virtually always completed an ejaculatory series before returning to the next. (Only 2 of the 32 series were interrupted; the preponderance of maternal behavior occurred during the postejaculatory intervals.) While “sexual motivation” remains high during the ejaculatory series (preventing the expression of maternal behavior), it declines after ejaculation, during the PEI.
Thus, it seems that in both sexes of this species, the postejaculatory interval is a “sexually protected” period. Both the male and the female entered an altered (and reduced) state of copulatory readiness during the critical time when sperm must travel from the vagina into the uterus.
SOCIAL FACTORS IN THE CONTROL OF SPERM TRANSPORT
Sperm Transport: Inhibition
In the previous sections we have described the effects of copulation on sperm transport, concentrating on the organismic level (a single copulating pair, for a single ejaculation). As we pointed out earlier, though, behavior has consequences on other levels (both higher, social levels and lower, physiological levels). In this section, we would like to explore some of the consequences of sperm transport on the social level.

ADLER & TONER EFFECTS OF COPULATORY BEHAVIOR ON SPERM 21
Of course, sexual behavior is, by definition, social since it involves at least two animals-the male and the female. The social dimension in our case study, however, applies to mating with several males (and females) that are normally present in the immediate social environment of the copulating pair.
To begin this analysis we would like to reconsider the postejaculatory phase of the mating encounter. Although the ejaculating male’s own PEI may reflect an active physiological process that protects his own sperm from disruption by his too-quick resumption of mating, there is still the possibility that another male could interfere with the transport of the first male’s sperm. We have already seen that one male can “cancel” the sperm transport of a male that has just ejaculated, substituting his own sperm instead.’*,“
This inhibitory effect of one male’s copulatory behavior on the reproductive success of another is an example of what has been broadly called “sperm omp petition."^^ One molar definition of sperm competition is: the set of behavioral and physiological mechanisms by which the probability of one male’s sperm fertilizing the available ova is increased at the expense of another.
Sperm Transport: Protection
Like other dyadic relationships in biology (e.g., prey -predator relations), sperm competition between males involves pressures from both sides of the relationship. Thus, while the copulatory disruption of sperm transport is an adaptive mechanism by which one male might cancel the effects of a previous male’s mating, the “disad- vantaged” male might be expected to generate a set of responses (secondary adap- tations) to prevent his sperm from being displaced, or at least to mitigate the effect of displacement. Though the mechanism is not yet clear, there is evidence that this might be occurring in rats: Following ejaculation by a dominant male, the female waits twice as long before resuming mating than if the ejaculation was delivered by a male of lower social tatu us.^' Thus, one way in which a male (at least a dominant male) can protect his gametic investment is to modify the behavior of the female, so as to protect the sperm he has just ejaculated from being dislodged by a second (subdominant) male.
In addition to protecting his sperm by modifying the behavior of the female, there is a second way in which a dominant male can ensure his sperm’s success against that of a genetic adversary: by reducing the gametic output of the subordinate male, as discussed in the previous section.
A third, related strategy by which a male might protect his investment of sperm is to increase the amount or quality of copulatory contact with a given female. Lanier et ~ 1 . ~ ’ have shown that, in a competitive mating situation, male rats increase the proportion of pups that they sire when they deliver a greater number of ejaculatory series, relative to another male. They concluded that the proportion of sperm from each male is important in determining reproductive advantage.
In determining the final proportion of sperm that a female receives from several males, the total number of ejaculations is surely important. We have recently discovered that there is another major factor operating, viz., the particular ejaculatory series which is (are) del i~ered.~~
In studying the fertility of successive ejaculatory series, we were interested in determining the basic, physiological parameters of reproduction in this species which is characterized by multiple ejaculatory series. There is another, more ecological reason
28 ANNALS NEW YORK ACADEMY OF SCIENCES
for pursuing this research, though. The concepts of sperm competition and mating strategy play a central theoretical role in behavioral ecology and sociobiology. Often, these theoretical considerations are pursued with great formal clarity. In addition to this theoretical rigor, however, basic empirical data are needed to adequately evaluate competing theoretical hypotheses about the biopsychology and behavioral ecology of reproductive processes. So although it is important to consider the effects of multiple ejaculations in a social setting with several ma1es,2°860 it would also seem important to work out the basic pattern of fertility supported by each ejaculation in order to satisfactorily elaborate the evolutionary consequences of complex mating patterns.
Thirteen male and 276 female Sprague-Dawley CD rats (Charles River) were used in this experiment. Two groups of males were randomly selected from our pool of sexually experienced animals: an older group (n=9), 223 to 402 days old at the beginning of the study, and a younger group (n=4), 103 to 172 days old.
After 2 weeks of rest, a given male was mated successively to 6 females (in the case of the older males) or 7 females (in the case of the younger males). Within a minute after each ejaculation the female was removed and replaced with another estrous female. Females mated to the younger males were given five preliminary intromissions by a “stimulus” male before being mated to the experimental male. (These priming intromissions increased the chance that any reduction in fertility would not be due to a lack of progesterone secretion and would instead depend on reduced numbers of sperm.) One group of females was examined for numbers of sperm; another group (mated to the same males) was allowed to become pregnant, and the number of embryos was counted. The results are illustrated in FIGURES 2 and 3.
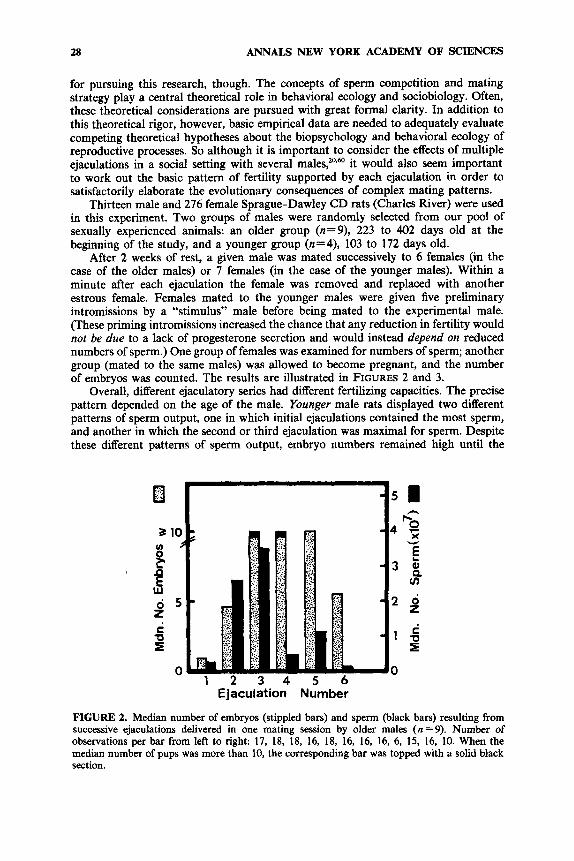
Overall, different ejaculatory series had different fertilizing capacities. The precise pattern depended on the age of the male. Younger male rats displayed two different patterns of sperm output, one in which initial ejaculations contained the most sperm, and another in which the second or third ejaculation was maximal for sperm. Despite these different patterns of sperm output, embryo numbers remained high until the
$ W
0 z r 0 E
0 z c
Ejaculation Number
FIGURE 2. Median number of embryos (stippled bars) and sperm (black bars) resulting from successive ejaculations delivered in one mating session by older males (n =9). Number of observations per bar from left to right: 17, 18, 18, 16, 18, 16, 16, 16, 6, 15, 16, 10. When the median number of pups was more than 10, the corresponding bar was topped with a solid black section.
ADLER & TONER EFFECTS OF COPULATORY BEHAVIOR ON SPERM 29
5 1
' P 0
Ejaculation Number
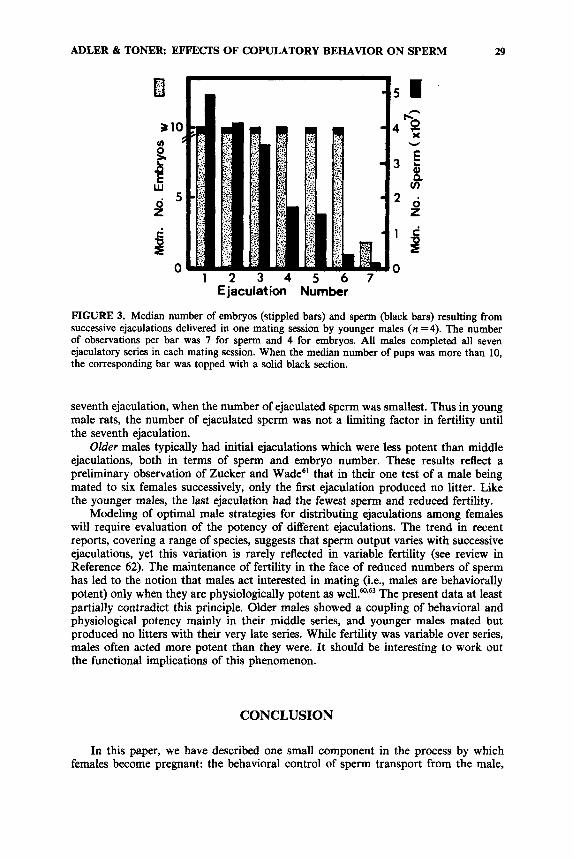
FIGURE 3. Median number of embryos (stippled bars) and sperm (black bars) resulting from successive ejaculations delivered in one mating session by younger males (n =4). The number of observations per bar was 7 for sperm and 4 for embryos. All males completed all seven ejaculatory series in each mating session. When the median number of pups was more than 10, the corresponding bar was topped with a solid black section.
seventh ejaculation, when the number of ejaculated sperm was smallest. Thus in young male rats, the number of ejaculated sperm was not a limiting factor in fertility until the seventh ejaculation.
Older males typically had initial ejaculations which were less potent than middle ejaculations, both in terms of sperm and embryo number. These results reflect a preliminary observation of Zucker and Wade6' that in their one test of a male being mated to six females successively, only the first ejaculation produced no litter. Like the younger males, the last ejaculation had the fewest sperm and reduced fertility.
Modeling of optimal male strategies for distributing ejaculations among females will require evaluation of the potency of different ejaculations. The trend in recent reports, covering a range of species, suggests that sperm output varies with successive ejaculations, yet this variation is rarely reflected in variable fertility (see review in Reference 62). The maintenance of fertility in the face of reduced numbers of sperm has led to the notion that males act interested in mating (i.e., males are behaviorally potent) only when they are physiologically potent as The present data at least partially contradict this principle. Older males showed a coupling of behavioral and physiological potency mainly in their middle series, and younger males mated but produced no litters with their very late series. While fertility was variable over series, males often acted more potent than they were. It should be interesting to work out the functional implications of this phenomenon.
CONCLUSION
In this paper, we have described one small component in the process by which females become pregnant: the behavioral control of sperm transport from the male,

30 ANNALS NEW YORK ACADEMY OF SCIENCES
through the reproductive tract of the female, to the point where sperm and egg unite. We have concentrated on the behavioral (and social) levels of organization and have tried to show how physiological parameters underlying successful reproduction are regulated by higher levels. This process, in one species, represents a kind of microcosm reflecting the behavioral- physiological integration that occurs across a wide spectrum of behavior patterns and species and therefore has general applicability in bio- psychological thought. In developing this research program, much of our thinking has been illuminated by our contacts, social and intellectual, with Jay Rosenblatt over many years. We hope we have many more years of instruction from him.
REFERENCES
1.
2. 3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13. 14.
15.
16.
17.
18.
19.
20.
KELLY, J. V. 1962. Myometrial participation in human sperm transport: a dilemma. Fert.
Fox, C. t B. Fox. 1967. Uterine suction during orgasm. Br. Med. J. 1: 300-301. MOGHISSI, K. S. 1969. Sperm migration in the human female genital tract. J. Reprod.
Med. 3: 73. HAFEZ, E. S. E. 1973. Transport of spermatozoa in the female reproductive tract. Am. J.
Obstet. Gynecol. 115: 703. SETTLAGE, D. S. F., M. MOTOSHIMA BE D. TREDWAY. 1973. Sperm transport from the
external cervical 0s to the fallopian tubes in women: a time and quantitation study. Fertil. Steril. 24. 655.
MASTERS, W. H. t W. E. JOHNSON. 1966. Human Sexual Response. Little, Brown t Co. Boston, Mass.
DEWSBURY, D. A. 1967. A quantitative description of the behaviour of rats during cop- ulation. Behaviour 2 9 154- 178.
DEWSBURY, D. A. 198 1. On the function of the multiple-intromission, multiple-ejaculation copulatory patterns of rodents. Bull. Psychon. SOC. 18: 221-223.
ADLER, N. T. 1968. Effects of the male’s copulatory behavior in the initiation of pregnancy in the female rat. Anat. Rec. 160: 304.
ADLER, N. T, 1969. Effects of the male’s copulatory behavior on successful pregnancy of the female rat. J. Cornp. Physiol. Psychol. 69: 613-622.
ADLER, N. T. 1983. The neuroethology of reproduction. I n Advances in Vertebrate Neu- roethology. J. Ewert, R. Capranica & D. Ingle, U s . : 1033- 1061. Plenum Press. London, England.
ADLER, N. T. t S. R. ZOLOTH. 1970. Copulatory behavior can inhibit pregnancy in female rats. Science 168: 1480- 1482.
DEVINE, M. C. 1975. Copulatory plugs in snakes: enforced chastity. Science 187: 844-845. DEVINE, M. C. 1977. Copulatory plugs, restricted mating opportunities, and reproductive
competition among male garter snakes. Behaviour 267: 154- 178. MARTAN, J. t B. A. SHEPHERD. 1976. The role of the copulatory plug in reproduction
of the guinea pig. J. Exp. Zool. 196: 79-84. MATTHEWS, M. t N. T. ADLER. 1977. Facilitative and inhibitory influences of reproductive
behavior on sperm transport in rats. J. Comp. Physiol. Psychol. 91: 727-741. MATTHEWS, M. L N. T. ADLER. 1978. Systematic interrelationship of mating, vaginal
plug position, and sperm transport in the rat. Physiol. Behav. 20 303-309. MATTHEWS, M. t N. T. ADLER. 1979. Relative efficiency of sperm transport and number
of sperm ejaculated in the female rat. Biol. Reprod. 2 0 540-544. Ross, P. & D. CREWS. 1977. Influence of the seminal plug on mating behaviour in the
garter snake. Nature 267: 344-345. DEWSBURY, D. A. BE T. G. HARTUNG. 1980. Copulatory behavior and differential repro-
duction of laboratory rats in a two-male, one-female competitive situation. Anim. Behav.
Steril. 13: 84.
28: 95-102.

ADLER & TONER: EFFECIS OF COPULATORY BEHAVIOR ON SPERM 31
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
42.
43.
44.
DEWSBURY, D. A. & D. J. BAUMGARDNER. 1981. Studies of sperm competition in two species of muroid rodents. Behav. Ecol. Sociobiol. 9 121-133.
ALLEN, T. 0. & N. T. ADLER. 1982. The neuroendocrine consequences of sexual behavior in mammals. In Reproduction. N. T. Adler & D. Pfaff, Eds. Plenum Press. New York, N.Y.
BLANDAU, R. 3. 1945. On the factors involved in sperm transport through the cervix uteri of the albino rat. Am. J. Anat. 77: 253-272.
WILSON, J. R., N. ADLER & B. LEBOEUF. 1965. The effects of intromission frequency on successful pregnancy in the female rat. Proc. Natl. Acad. Sci. USA 53: 1392-1395.
ADLER, N. T., J. A. RESKO & R. W. GOY. 1970. The effect of copulatory behavior on hormonal change in the female rat prior to implantation. Physiol. Behav. 5 1003- 1007.
CHESTER, R. V. & I. ZUCKER. 1970. Influence of male copulatory behavior on sperm transport, pregnancy, and pseudopregnancy in female rats. Physiol. Behav. 5: 35-43.
TERKEL, J. & C. H. SAWYER. 1978. Male copulatory behavior triggers nightly prolactin surges resulting in successful pregnancy in rats. Horm. Behav. 11: 304-309.
HARTMAN, C. G. & J. BALL. 1931. On the almost instantaneous transport of spermatozoa through the cervix and uterus in the rat. Proc. SOC. Exp. Biol. Med. 28: 312.
FREUND, M. 1973. Mechanisms and problems of sperm transport. In The Regulation of Mammalian Reproduction. S. J. Segal, R. Crozier, P. A. Cofman & P. G. Condiffe, Eds. C. C. Thomas. Springfield, Ill.
OVERSTREET, J. W., G. W. COOPER & D. F. KATZ. 1978. Sperm transport in the repro- ductive tract of the female rabbit. 2. The sustained phase of transport. Biol. Reprod. 1 9 115- 132.
TONER, J. P. & N. T. ADLER. 1986. The pre-ejaculatory behavior of male and female rats affects the number of sperm in the vagina and uterus. Physiol. Behav. 36 363-367.
BERMANT, G. 1964. Effects of single and multiple enforced intercopulatory intervals on the sexual behavior of male rats. J. Comp. Physiol. Psychol. 57: 398-403.
LARRSON, K. 1959. The effect of restraint upon copulatory behaviour in the rat. h i m . Behav. 7: 23-25.
SILBERBERG, A. & N. T. ADLER. 1974. Modulation of the copulatory sequence of the male rat by a schedule of reinforcement. Science 185 374-375.
MCCLINTOCK, M. K., J. J. ANISKO & N. T. ADLER. 1982. Group mating among Norway rats. 2. The social dynamics of copulation: competition, cooperation, and mate choice. Anim. Behav. 30 410-425.
COSTANZO, D. J. & R. K. ORNDOFF. 1984. Reproductive success of dominant versus subdominant male rats in a competitive mating situation. Abstract presented at Confer- ence on Reproductive Behavior, Pittsburgh, Pa.
MACMILLAN, K. L. & H. D. HAFS. 1967. Semen output of rabbits ejaculated after varying sexual preparation. Proc. Soc. Exp. Biol. Med. 125 1278-1281.
ALMQUIST, J. 0. 1973. Effects of sexual preparation on sperm output, semen characteristics and sexual activity of beef bulls with a comparison to dairy bulls. J. Anim. Sci. 36
FOSTER, J., J. 0. ALMQUIST & R. C. MARTIG. 1970. Reproductive capacity of beef bulls. IV. Changes in sexual behavior and semen characteristics among successive ejaculations. J. Anim. Sci. 30 245-252.
CHENG, M.-F. 1983. Behavioural “self-feedback” control of endocrine states. In Hormones and Behaviour in Higher Vertebrates. J. Balthazart, E. Prove & R. Gilles, Eds. Springer- Verlag. West Berlin, FRG.
ROTH, L. & J. S. ROSENBLATT. 1968. Self-licking and mammary development during pregnancy in the rat. J. Endocrinol. 42: 363-378.
COHEN, J. & M.-F. CHENG. 1979. Role of vocalization in the reproductive cycle of ring doves (Streptopelia risoriu): effects of hypoglossal nerve section on the reproductive behavior and physiology of the female. Horm. Behav. 13 113- 127.
MARLER, P. 1970. A comparative approach to vocal learning: song development in white- crowned sparrows. J. Comp. Physiol. Psychol. 71: 1-25.
HINDE, R. A. & R. P. WARREN. 1959. The effect of nest building on later reproductive behaviour in domesticated canaries. Anim. Behav. 7: 35-41.
331-336.

32 ANNALS NEW YORK ACADEMY OF SCIENCES
45.
46.
47.
48.
49.
50.
51.
52.
53.
54.
55.
56.
57.
58.
59.
60. 61.
62.
63.
CHENG, M.-F. & J. BALTHAZART. 1982. The role of nest-building activity in gonadotrophin secretions and the reproductive success of ring doves (Streptopelia risoria). J. Comp. Physiol. Psychol. 96: 307-324.
KURTZ, R. & N. T. ADLER. 1973. Electrophysiological correlates of sexual behavior in the male rat. J. Comp. Physiol. Psychol. 84: 225-239.
ADLER, N. T. 1974. The behavioral control of reproductive physiology. In Reproductive Behavior. M. Montagna & W. A. Sadler, Eds. Plenum Press. New York, N.Y.
ANISKO, J. J., S. F. SUER, M. K. MCCLINTOCK & N. T. ADLER. 1978. Relation between 22 kHz ultrasonic signals and sociosexual behavior in the rat. J. Comp. Physiol. Psychol.
GILBERT, A. N., R. J. PELCHAT & N. T. ADLER. 1980. Postpartum copulatory and maternal behaviour in Norway rats under seminatural conditions. Anim. Behav. 28 989-995.
GILBERT, A. N., R. J. PELCHAT L N. T. ADLER. 1984. Sexual and maternal behaviour at the postpartum oestrus: the role of experience in time-sharing. Anim. Behav. 32:
DAVIS, H. N., & 0. HALL. 1951. The seasonal reproductive conditions of female Norway (Brown) in Baltimore Maryland. Physiol. Zool. 24: 9-20.
GROTA, R. L. & R. ADER. 1969. Continuous recording of maternal behavior in Rattus norvegicus. Anim. Behav. 17: 722-729.
ADER, R. & R. J. GROTA. 1970. Rhythmicity in the maternal behavior of Rattus norvegicus. Anim. Behav. 1 8 144-150.
LEON, M., P. G. CROSKERRY & G. K. SMITH. 1978. Thermal control of mother-young contact in rats. Physiol. Behav. 21: 793-811.
MCFARLAND, D. 1974. Time-sharing as a behavioral phenomenon. Adv. Study Behav. 5 201 -225.
PARKER, G. A. 1970. Sperm competition and its evolutionary consequences in the insects. Biol. Rev. 45 525-547.
MCCLINTOCK, M. K., J. P. TONER, J. J. ANISKO & N. T. ADLER. 1982. Group mating among Norway rats. 11. The social dynamics of copulation: competition, cooperation, and mate choice. Anim. Behav. 30 410-425.
LANIER, D. L., D. Q. BTEP & D. A. DEWSBURY. 1979. Role of prolonged copulatory behavior in facilitating reproductive success in a competitive mating situation in labo- ratory rats. J. Comp. Physiol. Psychol. 93: 781-792.
TONER, J. P. & N. T. ADLER. 1985. Potency of rat ejaculations varies with their order and with male age. Physiol. Behav. 35: 113-115.
DEWSBURY, D. A. 1982. Ejaculate cost and male choice. Am. Nat. 119 601-610. ZUCKER, I. & G. WADE. 1968. Sexual preferences of male rats. J. Comp. Physiol. Psychol. 66 816-819.
DEWSBURY, D. A. & D. K. SAWREY. 1985. Male capacity as related to sperm production, pregnancy initiation, and sperm competition in deer mice (Peromyscus maniculatus). Behav. Jkol. Sociobiol. 1 6 37-47.
HALLIDAY, T. & A. HOUSTON. 1978. The newt as an honest salesman. Anim. Behav. 26: 1273-1281.
92: 821-829.
1045-1053.